
Submitted:
03 July 2023
Posted:
04 July 2023
You are already at the latest version
Abstract
Inotodiol, a lanostane-type triterpenoid, and many phytochemicals from Chaga mushroom have been investigated for various allergic diseases. However, antiaging and anti-inflammatory activities of inotodiol under different types of oxidative stress and impact of inotodiol on collagen and hyaluronan synthesis are not sufficiently studied. Lanostane triterpenoids-rich concentrate, which contained 10% inotodiol as major (the inotodiol concentrate), was prepared from Chaga and compared with pure inotodiol in terms of anti-inflammatory activities on a human keratinocyte cell line, HaCaT cells, under various stimulation such as stimulation with ultraviolet (UV) B or tumor necrosis factor (TNF)-α. In stimulation with TNF-α, interleukin (IL)-1β, IL-6, and IL-8 genes were significantly repressed by 0.44~4.0 μg/mL of pure inotodiol. UVB irradiation induced overexpressions of pro-inflammatory cytokines, but those genes were significantly suppressed by pure inotodiol or inotodiol concentrate. Moreover, pure inotodiol/inotodiol concentrate could also modulate synthesis of collagen and hyaluronic acid by controlling COL1A2 and HAS2/3 expression, which implies a crucial role of pure inotodiol/inotodiol concentrate in prevention of skin aging. These results enlighten anti-inflammatory and anti-aging effect of pure inotodiol/inotodiol concentrate, and it is highly conceivable that pure inotodiol and inotodiol concentrate could be promising natural bioactive substances to be incorporated in therapeutic and beautifying applications.
Keywords:
lanostane teriterpenoid
; inotodiol
; keratinocyte
; anti-aging
; anti-inflammation
1. Introduction
Pro-inflammatory cytokines, one of the inflammatory mediators, produced by multiple types of cells such as macrophages, mast cells, lymphocytes, and keratinocytes contribute to the initiation of various diseases including vascular disease, lipid metabolism, cancer, and aging-related diseases [1,2]. Keratinocytes, well known for synthesis of keratin, produce proinflammatory cytokines, such as TNF-α, IL-1β, when they detect antigen and play a crucial role in immune response in the skin [3]. Pro-inflammatory cytokines are also induced by UV radiation for skin defense responses [4]. Even though these cytokines are important for modulation of the immune response for protection of the host but excessive production of pro-inflammatory cytokines may cause chronic inflammation and negative consequences [5].
Inflammaging, which was coined in 2,000 by Prof. Claudio Franceschi, is a concept that suggests that low grade and persistent proinflammatory status, which is characterized by high levels of proinflammatory markers might lead long term aging progression and tissue damage [4,6,7,8]. If inflammaing occurs in the skin, it can cause cancer or skin diseases related to aging [9]. It is not only aging that causes inflammaing but also external environment, such as ultraviolet (UV) radiation or smoking can be reasons for inflammaging [10].
Inotodiol, a lanostane-type triterpenoid, from Chaga mushroom has been used as a traditional medicine for diabetes, cardiovascular disorders, and tuberculosis [11,12]. Numerous studies have investigated inotodiol’s biological activities and it has been proven to have antitumor, anti-inflammatory, antiallergic, antiviral, antiaging, and antioxidant properties [13,14,15,16]. Recently, there is a movement to identify novel, safe, and costly effective molecules that can be used for treatment of various diseases including inflammatory diseases. So far, it has been reported that inotodiol exerts anti-inflammatory activity without showing detrimental effect in in vivo and in vitro studies [17]. Also, inotodiol shows antiaging effect under oxidative stress by inhibiting MAPK-NOX5 and NF-κB signaling pathways [16]. However, antiaging and anti-inflammatory activities of inotodiol under different types of oxidative stress and impact of inotodiol on collagen and hyaluronan synthesis are not sufficiently studied. We therefore complemented our previous data in terms of antiaging and anti-inflammatory effect of inotodiol.
The aims of this study were 1) to evaluate the impact of inotodiol on pro-inflammatory cytokine production by keratinocytes stimulated with UV or TNF-α and 2) to assess how inotodiol affects hyaluronic acid synthesis in HaCaT cells under the presence of TNF-α or not. Here, we demonstrated for the first time that inotodiol is capable of reducing inflammation induced by UVB and TNF-α and upregulates hyaluronan and collagen synthesis related genes in HaCaT cells that imply potential role of inotodiol in mitigating or delaying inflammaging.
2. Results
2.1. Evaluation of cytotoxic effect of inotodiol and inotodiol concentrate
The MTT assay was used to evaluate the cytotoxicity of pure inotodiol and inotodiol concentrate using HaCaT cell line and cytotoxicity was determined as per ISO EN 10993-5 criteria. Cell viability under 70% of the negative control considered to be toxic [18]. Cells were treated with various concentrations of inotodiol/inotodiol concentrate (0~20 μg/ml) for 24 h. Inotodiol were non-toxic (viability was >70%) other than at the highest concentration of 20 μg/ml. Inotodiol concentrate was non-toxic at all tested concentration (viability was >70%). At a lower concentration (~5 μg/ml), inotodiol shows more than 90% cell viability but it gradually decreased with the increase in concentrations of inotodiol (Figure 1). In inotodiol concentrate, no concentration dependent decrease in terms of cell viability was observed (Figure 1B). According to the results for cell viability, 5 μg/ml of inotodiol and 20 μg/ml inotodiol concentrate were chosen as the highest concentration for cell culture.
2.2. Impact of inotodiol and inotodiol concentrate on pro-inflammatory cytokine expression after treatment with TNF-α in HaCaT cells
To examine the effect of inotodiol/inotodiol concentrate on the pro-inflammatory cytokine mRNA expression level after treatment with TNF-α in HaCaT cells, the mRNA expression of IL-1β, IL-6, and IL-8 were evaluated by real-time qPCR. The mRNA expression level of IL-1β, IL-6, and IL-8 increased by treatment with TNF-α (Figure 2).
Addition of inotodiol /inotodiol concentrate, however, significantly reduced mRNA expression level of these cytokines especially at higher concentration. Addition of inotodiol under TNF-α presence reduced mRNA expression level of these cytokines in a dose dependent manner (Figure 3). Cells treated with inotodiol concentrate showed decreased mRNA expression of the cytokines at the lowest tested concentration (2.5 μg/ml).
2.3. Impact of inotodiol and inotodiol concentrate on pro-inflammatory cytokine expression after treatment with UV in HaCaT cells
Inflammation is one of the important mediators of photoaging and UV radiation promotes release of inflammatory mediators such as IL-1 and IL-6 through induction of pro-inflammatory genes from keratinocytes [19]. To investigate the ability of inotodiol/inotodiol concentrate to reduce inflammation level, mRNA expression level of two pro-inflammatory cytokines, IL-6 and TNF-α were assessed. By addition of inotodiol/inotodiol concentrate, mRNA expression level of IL-6 and TNF-α decreased (Figure 4). Especially, above 2.2 μg/ml inotodiol signify reduced these cytokine expressions. In cells treated with inotodiol concentrate, IL-6 mRNA expression level at the highest concentration (20 μg/ml) returned almost equivalent to the basal level.
2.4. Impact of inotodiol/inotodiol concentrate on hyaluronan synthesis and collagen
To investigate if inotodiol/inotodiol concentrate affects collagen synthesis, HaCaT cells were treated with inotodiol/inotodiol concentrate and mRNA expression level of COL1A2 was determined by qRT-PCR. Treatment with inotodiol/inotodiol concentrate increased COL1A2 expression in HaCaT cells. In addition, to examine the role of inotodiol/inotodiol concentrate in hyaluronic acid expression, HAS2 and HAS3 mRNA expression level were also assessed by qRT-PCR. Hyaluronic acid (HA) synthesis is regulated by an enzyme called hyaluronic acid synthases (HAS) [20] and three isoforms of HAS, HAS1, HAS2, and HAS3, have been reported so far [1,21]. As shown in Figure 5, inotodiol/inotodiol concentrate increased HAS2 and HAS3 expression.
Hyaluronidase (HYAL) has HA degrading activity through cleavage at the β-(1,4)-linkage [22]. To investing the influence of inotodiol on expression of HYAL 3 under stimulation of TNF-α (20 ng/ml), HaCaT cells were incubated with various concentrations of inotodiol under TNF-α stimulation for XX h. By addition of TNF-α to HaCaT cells without inotodiol, HYAL-3 mRNA expression increased (Figure 6). Treatment with inotodiol, however, significantly reduced HYAL-3 mRNA expression even with low concentration (0.55 μg/ml).
3. Discussion
Multiple stresses or stressors such as oxidative, chemical/physical, or antigenic stress lead to constant stimulation of immune response that can be characterized as inflamm-aging [6]. In the skin, tissue damage or other skin diseases can be triggered by inflamm-aging [23]. Inotodiol has been studied for its various biological activities so far and antioxidant and anti-inflammatory function are its well-known activities [24,25,26,27,28,29]. UV irradiation causes acute inflammation, which usually causes oxidative stress [30]. UV irradiation induces accumulation of ROS and it results in activation of NF-κB that can up-regulate expression of genes that involved in inflammation [31]. In this study, we investigated the functions of inotodiol/inotodiol concentrate in terms of modulation of UV or TNF-α induced inflammation for the first time. Previously it has been reported that inotodiol has anti-oxidative and anti-inflammatory activities in vivo and in vitro [32,33,34]. Moreover, recent study shows that inotodiol has anti-aging effects by protecting human dermal fibroblast through blocking MAPK-NOX5 and NF-κB activation and suppression of aging gene expressions [16]. However, it is still not elucidated if inotodiol affects cytokine expression after stimulation with UV and TNF-α and how inotodiol impacts on hyaluronic acid production after TNF-α stimulation. Here, we demonstrated that inotodiol reduced UV- and TNF-α-induced pro-inflammatory cytokine expressions and upregulated Hyaluronan synthesis by increasing HAS2 and HAS3 expression, which results in augmentation of HA synthesis. In addition, inotodiol prevented degradation of HA by down-regulation of hyaluronidase-3 expression under TNF-α stimulation. These data additionally support potent anti-inflammatory and anti-aging activities of inotodiol in skin cells that imply a potential use of inotodiol in cosmetic industry.
Intrinsic aging and extrinsic aging trigger skin aging through a complex biological process [35]. Ultraviolet (UV) radiation is one of the external factors that causes extrinsic aging, also called photoaging. When it comes to anti-aging in skin, hyaluronic acid (HA, hyaluronan) plays crucial role by binding water that results in keeping skin hydrated.
In HaCaT cells that exposed to UV or TNF-α, expression of pro inflammatory cytokine such as IL-1β, IL-6, and IL-8 increased, which verifies that inflammation plays crucial roles in terms of skin aging (fig. XX). TNF-α has been known for a key mediator of immune response and inflammation in skin through activation of NF-κB signaling [36]. Keratinocytes, epidermal Langerhans cells (LC) and macrophages secrete TNF-α and this TNF-α acts on these cells via autocrine manner and induces other pro-inflammatory cytokines [37].
To assess the ability of inotodiol/inotodiol concentrate to modulate inflammation, pro-inflammatory cytokine expressions were measured in HaCaT cells treated with or without inotodiol/inotodiol concentrate under UV or TNF-α stimulation. Inotodiol/inotodiol concentrate protected cells from increase of pro-inflammatory cytokines against UV or TNF-α stimulation and these results indicate a positive control of inotodiol/inotodiol concentrate in terms of inflamm-aging. Regarding control of IL-1β expression by treatment with inotodiol/inotodiol concentrate, expression of IL-1β decreased even with low concentrate of inotodiol/inotodiol concentrate. It has been reported that pro-inflammatory cytokines including IL-1β promote collagen degradation and down-regulate production of collagen [38]. In our study, we demonstrated that addition of inotodiol/inotodiol concentrate to HaCaT cells increased COL1A2 which means inotodiol/inotodiol concentrate positively modulate HaCaT cells in terms of collagen synthesis. Moreover, inotodiol up-regulated HAS2 and HAS3 and it indicates the ability of inotodiol/inotodiol concentrate to increase hyaluronic acid expression which is important to protect skin from senescence [35]. These findings suggest that inotodiol/inotodiol concentrate effectively control skin aging via suppression of pro-inflammatory cytokines and augment the ability of skin to maintain collagen and hyaluronic acid synthesis.
4. Materials and Methods
4.1. Materials
Dulbecco’s Modified Eagle’s medium (DMEM), Dulbecco’s Phosphate Buffered Saline (DPBS), Fetal Bovine serum (FBS) and Penicillin-Streptomycin solutions (P/S) were purchased from Sigma-Aldrich (St. Louis, MO, USA). 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) was purchased from Thermo Scientific (Massachusetts, USA) and both AccuPower cyclescript RT premix and AccuPower Taq PCR Premix, primer were purchased from Bioneer (Daejeon, South Korea).
4.2. Preparation of inotodiol concentrate and high purity inotodiol
Dried Chaga mushroom was purchased from Jungwoodang (Seoul, Korea). Chaga mushroom powder (200 g) was treated with 2 L of 70% ethanol and stirred at 50°C for 4 h. After centrifugation (12,000 x g, 10 min) at room temperature, the supernatant was obtained and concentrated using vacuum rotary evaporator (Deahan Science Co., Daejeon, Korea) to final volume of 200 mL. The concentrate was kept at 4°C for 15 h and the precipitant was separated by centrifugation. The precipitant obtained this step was designated as inotodiol concentrate, which had 10 % inotodiol approximately. To obtain high purity inotodiol, inotodiol concentrate was further purified using recycling HPLC system (MPLC, YMCKOREA, Seongnam, Korea) equipped with reverse-phase chromatography (ODS-AQ, 50x500 mm). Finally, 95% purity inotodiol was produced and used for the in vitro assays.
4.3. Cell culture
HaCaT cell, a human epithelial keratinocyte cell line was kindly provided by Dr. Jongil Park from Chungnam University medical school. HaCaT cells were cultured in Dulbecco’s Modified Eagle’s Medium supplemented with 10% Fetal bovine serum and 1X penicillin-streptomycin at 37°C under 5% CO2.
4.4. Cell viability assay
A standard MTT assay was performed to determine cytotoxicity. HaCaT cells were seeded at a density of 1X105 cells/well and incubated for 24 hours. Cells were treated with various concentrations (0.6 ~ 20 μg/ml) of pure inotodiol and inotodiol concentrate in a 96-well plate. After treatment with pure inotodiol and inotodiol concentrate for 24 h under 5% CO2, the cells were treated with 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT) solution (XXXmg/ml) for 3 hours, and the resultant formazan crystals were dissolved in 100 μL of Dimethyl sulfoxide (DMSO). The absorbance was measured by microreader at 570 nm. The results were expressed as percentage of treated cell number relative to the untreated control cells.
4.5. UVB irradiation and TNF-α treatment
HaCaT cells were seeded (0.1X105 cells/ml) in 6-well plates in DMEM medium with 10% FBS for 24 hours. After stabilization, cells were pretreated with pure-inotodiol and inotodiol concentrate for 18 hours. Cells were washed with PBS before irradiation and after washing, 1 mL of PBS was added to each well. For UVB irradiation, PBS was removed and DMEM without serum was added to each cell and cells were incubated for 18 hours. Rhe total energy of irradiation was 10 mJ/cm2 (wavelength 312 nm, 10 sec). For treatment with TNF-α, HaCaT cells were treated with 20 μg/ml TNF-α for 6 h at 37°C.
4.6. Quantitative polymerase chain reaction (qPCR)
Total RNA was isolated from HaCaT cells using TRIzol (Life Technologies, Carlsbad, CA, USA) according to the manufacturer’s protocol. cDNA was synthesized from 1 µg RNA by reverse transcriptional reaction. The qPCR amplification was performed with SYBR-Green/ROX qPCR Master Mix (2X) (BioFact Co. Daejeon, Korea). The PCR primers of IL-6, TNF-α, IL-1β, iNOS, PLTP, APOE, ABCA1, and GAPDH were obtained from Bioneer company (Yuseong-gu, Daejeon, Korea). Primer sequences are in Table 1.
4.7. Statistical analyses
At least three independent experiments were performed in triplicate and the results were presented as means ± standard deviation (SD). Statistical comparison between experimental groups and controls was performed using a Student’s t-tests. P values less than 0.05 were considered significant.
5. Conclusions
In conclusion, out results demonstrated that pure inotodiol and lanostane triterpenoids-rich inotodiol concentrate exert anti-inflammatory activities under various stimulation such as stimulation with UV or TNF-α, which might due to prevent NF-κB signaling activation but it should be further elucidated. Pure inotodiol/inotodiol concentrate could also modulate synthesis of collagen and hyaluronic acid by control COL1A2 and HAS2/3 expression, which implies a crucial role of inotodil/inotodiol concentrate in prevention of skin aging. These results enlighten anti-inflammatory and anti-aging effect of inotodiol/inotodiol concentrate and it is highly conceivable inotodiol/inotodiol concentrate as a promising natural bioactive substance to be incorporated in therapeutic and beautifying applications.
6. Patents
Not applicable.
Supplementary Materials
None.
Author Contributions
Conceptualization, JT Park and J Park; methodology, J park; formal analysis, and investigation, J Park, TMN Nguyen, and HA Park; resources, CK Lee; writing—original draft preparation, J Park; writing—review and editing, JT Park; supervision, JT Park; project administration and funding acquisition, CK Lee. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This work was supported by the Technology Commercialization support program, through the Korea Innovation Foundation funded by the Ministry of Science and ICT (project No. 1711177773).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
None.
Acknowledgments
None.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dhandapani, S.; Wang, R.; Hwang, K.C.; Kim, H.; Kim, Y.-J. Enhanced skin anti-inflammatory and moisturizing action of gold nanoparticles produced utilizing Diospyros kaki fruit extracts. Arab. J. Chem. 2023, 16. [Google Scholar] [CrossRef]
- Liu, C.; Chu, D.; Kalantar-Zadeh, K.; George, J.; Young, H.A.; Liu, G. Cytokines: From Clinical Significance to Quantification. Adv. Sci. 2021, 8, e2004433. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.; Rodrigues, P.; Pintado, M.; Tavaria, F. A systematic review of natural products for skin applications: Targeting inflammation, wound healing, and photo-aging. Phytomedicine 2023, 115, 154824. [Google Scholar] [CrossRef] [PubMed]
- Alves, G.d.A.D.; de Souza, R.O.; Rogez, H.L.G.; Masaki, H.; Fonseca, M.J.V. Cecropia obtusa extract and chlorogenic acid exhibit anti aging effect in human fibroblasts and keratinocytes cells exposed to UV radiation. PLOS ONE 2019, 14, e0216501. [Google Scholar] [CrossRef] [PubMed]
- Valeri, M.; Raffatellu, M. Cytokines IL-17 and IL-22 in the host response to infection. Pathog. Dis. 2016, 74. [Google Scholar] [CrossRef] [PubMed]
- Fulop, T.; Larbi, A.; Pawelec, G.; Khalil, A.; Cohen, A.A.; Hirokawa, K.; Witkowski, J.M.; Franceschi, C. Immunology of Aging: the Birth of Inflammaging. Clin. Rev. Allergy Immunol. 2021, 64, 109–122. [Google Scholar] [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef]
- Koppula, S., M. Akther, and M.E. Haque, Potential Nutrients from Natural and Synthetic Sources Targeting Inflammaging-A Review of Literature, Clinical Data and Patents. 2021. 13(11).
- Pająk, J.; Nowicka, D.; Szepietowski, J.C. Inflammaging and Immunosenescence as Part of Skin Aging—A Narrative Review. Int. J. Mol. Sci. 2023, 24, 7784. [Google Scholar] [CrossRef]
- Yap, W.N. Tocotrienol-rich fraction attenuates UV-induced inflammaging: A bench to bedside study. J. Cosmet. Dermatol. 2017, 17, 555–565. [Google Scholar] [CrossRef]
- Park, Y.-M.; Won, J.-H.; Kim, Y.-H.; Choi, J.-W.; Park, H.-J.; Lee, K.-T. In vivo and in vitro anti-inflammatory and anti-nociceptive effects of the methanol extract of Inonotus obliquus. J. Ethnopharmacol. 2005, 101, 120–128. [Google Scholar] [CrossRef]
- Kim, H.G., et al., Ethanol extract of Inonotus obliquus inhibits lipopolysaccharide-induced inflammation in RAW 264.7 macrophage cells. J Med Food, 2007. 10(1): p. 80-9.
- Chung, J.; Choi, M.; Park, S.; Kang, J.; Chung, E.H.; Park, J.; Kim, Y.M. Inotodiol suppresses allergic inflammation in allergic rhinitis mice. Int. Forum Allergy Rhinol. 2023, 13, 1603–1614. [Google Scholar] [CrossRef]
- Nguyen, P.C.; Nguyen, M.T.T.; Truong, B.T.; Kim, D.-R.; Shin, S.; Kim, J.-E.; Park, K.-B.; Park, J.-H.; Tran, P.L.; Ban, S.-Y.; et al. Isolation, Physicochemical Characterization, and Biological Properties of Inotodiol, the Potent Pharmaceutical Oxysterol from Chaga Mushroom. Antioxidants 2023, 12, 447. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, W.; Chen, C.; Zhang, C.; Duan, J.; Yao, H.; Wei, Q.; Meng, A.; Shi, J. Inotodiol protects PC12 cells against injury induced by oxygen and glucose deprivation/restoration through inhibiting oxidative stress and apoptosis. J. Appl. Biomed. 2018, 16, 126–132. [Google Scholar] [CrossRef]
- Lee, S.H.; Won, G.-W.; Choi, S.-H.; Kim, M.-Y.; Oh, C.-H.; Park, J.-T. Antiaging effect of inotodiol on oxidative stress in human dermal fibroblasts. BioMedicine 2022, 153, 113311. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.M.N.; Ban, S.-Y.; Park, K.-B.; Lee, C.-K.; Lee, S.-W.; Lee, Y.-J.; Baek, S.-M.; Park, J.-K.; Nguyen, M.T.T.; Kim, J.; et al. Evaluation of Toxicity and Efficacy of Inotodiol as an Anti-Inflammatory Agent Using Animal Model. Molecules 2022, 27, 4704. [Google Scholar] [CrossRef]
- Podgórski, R.; Wojasiński, M.; Ciach, T. Nanofibrous materials affect the reaction of cytotoxicity assays. Sci. Rep. 2022, 12, 1–12. [Google Scholar] [CrossRef]
- Bosch, R.; Philips, N.; Suárez-Pérez, J.A.; Juarranz, A.; Devmurari, A.; Chalensouk-Khaosaat, J.; González, S. Mechanisms of Photoaging and Cutaneous Photocarcinogenesis, and Photoprotective Strategies with Phytochemicals. Antioxidants 2015, 4, 248–268. [Google Scholar] [CrossRef]
- Sze, J.H.; Brownlie, J.C.; Love, C.A. Biotechnological production of hyaluronic acid: a mini review. 3 Biotech 2016, 6, 67. [Google Scholar] [CrossRef] [PubMed]
- Usui, T.; Amano, S.; Oshika, T.; Suzuki, K.; Miyata, K.; Araie, M.; Heldin, P.; Yamashita, H. Expression regulation of hyaluronan synthase in corneal endothelial cells. Investig. Ophthalmol. Vis. Sci. 2000, 41. [Google Scholar]
- Krupkova, O.; Greutert, H.; Boos, N.; Lemcke, J.; Liebscher, T.; Wuertz-Kozak, K. Expression and activity of hyaluronidases HYAL-1, HYAL-2 and HYAL-3 in the human intervertebral disc. Eur. Spine J. 2019, 29, 605–615. [Google Scholar] [CrossRef]
- Trentini, M., et al., Apple Derived Exosomes Improve Collagen Type I Production and Decrease MMPs during Aging of the Skin through Downregulation of the NF-κB Pathway as Mode of Action. 2022. 11(24).
- Nguyen, T.M.N.; Le, H.S.; Le, B.V.; Kim, Y.H.; Hwang, I. Anti-allergic effect of inotodiol, a lanostane triterpenoid from Chaga mushroom, via selective inhibition of mast cell function. Int. Immunopharmacol. 2020, 81, 106244. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Gao, D.; Cho, C.W.; Hwang, I.; Kim, H.M.; Kang, J.S. A Novel Bioanalytical Method for Determination of Inotodiol Isolated from Inonotus Obliquus and Its Application to Pharmacokinetic Study. Plants 2021, 10, 1631. [Google Scholar] [CrossRef] [PubMed]
- Nomura, M.; Takahashi, T.; Uesugi, A.; Tanaka, R.; Kobayashi, S. Inotodiol, a lanostane triterpenoid, from Inonotus obliquus inhibits cell proliferation through caspase-3-dependent apoptosis. Anticancer Res. 2008, 28. [Google Scholar]
- Duru, K.C., et al., The pharmacological potential and possible molecular mechanisms of action of Inonotus obliquus from preclinical studies. Phytother Res, 2019. 33(8): p. 1966-1980.
- Glamočlija, J.; Ćirić, A.; Nikolić, M.; Fernandes, .; Barros, L.; Calhelha, R.C.; Ferreira, I.C.; Soković, M.; van Griensven, L.J. Chemical characterization and biological activity of Chaga (Inonotus obliquus), a medicinal “mushroom”. J. Ethnopharmacol. 2015, 162, 323–332. [CrossRef]
- Zhang, X.; Bao, C.; Zhang, J. Inotodiol suppresses proliferation of breast cancer in rat model of type 2 diabetes mellitus via downregulation of β-catenin signaling. BioMedicine 2018, 99, 142–150. [Google Scholar] [CrossRef]
- Hu, L.; Nomura, S.; Sato, Y.; Takagi, K.; Ishii, T.; Honma, Y.; Watanabe, K.; Mizukami, Y.; Muto, J. Anti-inflammatory effects of differential molecular weight Hyaluronic acids on UVB-induced calprotectin-mediated keratinocyte inflammation. J. Dermatol. Sci. 2022, 107, 24–31. [Google Scholar] [CrossRef]
- Subedi, L., et al., Resveratrol-Enriched Rice Attenuates UVB-ROS-Induced Skin Aging via Downregulation of Inflammatory Cascades. 2017. 2017: p. 8379539.
- Cui, Y.; Kim, D.-S.; Park, K.-C. Antioxidant effect of Inonotus obliquus. J. Ethnopharmacol. 2005, 96, 79–85. [Google Scholar] [CrossRef]
- Lee, I.-K.; Kim, Y.-S.; Jang, Y.-W.; Jung, J.-Y.; Yun, B.-S. New antioxidant polyphenols from the medicinal mushroom Inonotus obliquus. Bioorganic Med. Chem. Lett. 2007, 17, 6678–6681. [Google Scholar] [CrossRef]
- Nguyen, T.M.N.; Ban, S.-Y.; Park, K.-B.; Lee, C.-K.; Lee, S.-W.; Lee, Y.-J.; Baek, S.-M.; Park, J.-K.; Nguyen, M.T.T.; Kim, J.; et al. Evaluation of Toxicity and Efficacy of Inotodiol as an Anti-Inflammatory Agent Using Animal Model. Molecules 2022, 27, 4704. [Google Scholar] [CrossRef]
- nová, R. and V. Pavlík, Hyaluronan: A key player or just a bystander in skin photoaging? 2022. 31(4): p. 442-458.
- Jeon, Y.J., et al., Rhododendrin ameliorates skin inflammation through inhibition of NF-κB, MAPK, and PI3K/Akt signaling. Eur J Pharmacol, 2013. 714(1-3): p. 7-14.
- Kristensen, M.; Chu, C.Q.; Eedy, D.J.; Feldmann, M.; Brennan, F.M.; Breathnach, S.M. Localization of tumour necrosis factor-alpha (TNF-α) and its receptors in normal and psoriatic skin: epidermal cells express the 55-kD but not the 75-kD TNF receptor. Clin. Exp. Immunol. 1993, 94, 354–362. [Google Scholar] [CrossRef]
- Qin, Z.; Okubo, T.; Voorhees, J.J.; Fisher, G.J.; Quan, T. Elevated cysteine-rich protein 61 (CCN1) promotes skin aging via upregulation of IL-1β in chronically sun-exposed human skin. AGE 2013, 36, 353–364. [Google Scholar] [CrossRef]
Figure 1.
Effect of pure inotodiol and inotodiol concentrate on the cell viability in HaCaT cells. Cytotoxicity (MTT assay) of pure-inotodiol (A) and inotodiol concentrate (B).
Figure 1.
Effect of pure inotodiol and inotodiol concentrate on the cell viability in HaCaT cells. Cytotoxicity (MTT assay) of pure-inotodiol (A) and inotodiol concentrate (B).
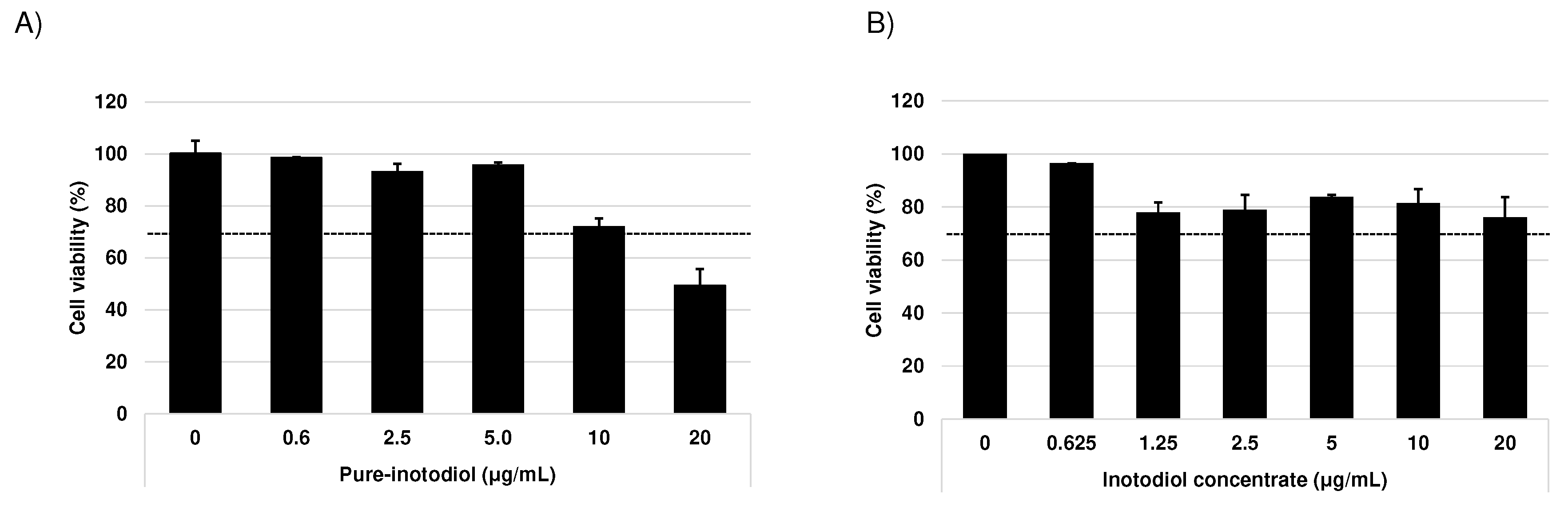
Figure 2.
Anti-inflammatory effects of pure-inotodiol (A and C) and inotodiol concentrate (B and D) on HaCaT cells treated with TNF-α (20 ng/mL). Statistical significance was denoted as * for p < 0.05 and ** for p < 0.01.
Figure 2.
Anti-inflammatory effects of pure-inotodiol (A and C) and inotodiol concentrate (B and D) on HaCaT cells treated with TNF-α (20 ng/mL). Statistical significance was denoted as * for p < 0.05 and ** for p < 0.01.
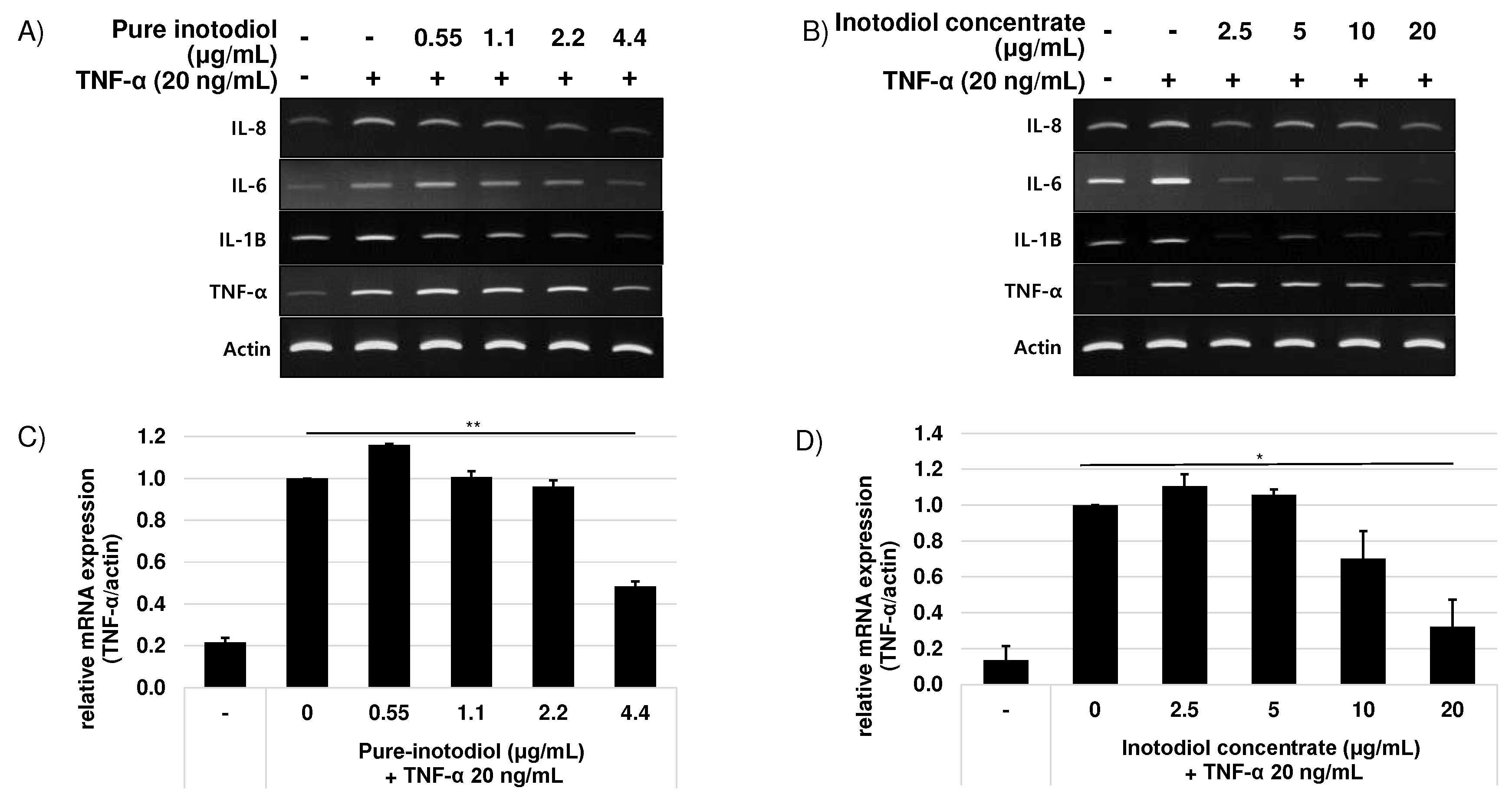
Figure 3.
Effects of pure inotodiol (A, C, and E) and the inotodiol concentrate (B, D, and F) on expression of proinflammatory cytokines of HaCaT cells treated with TNF-α (20 ng/mL). Statistical significance was denoted as * for p < 0.05 and ** for p < 0.01.
Figure 3.
Effects of pure inotodiol (A, C, and E) and the inotodiol concentrate (B, D, and F) on expression of proinflammatory cytokines of HaCaT cells treated with TNF-α (20 ng/mL). Statistical significance was denoted as * for p < 0.05 and ** for p < 0.01.
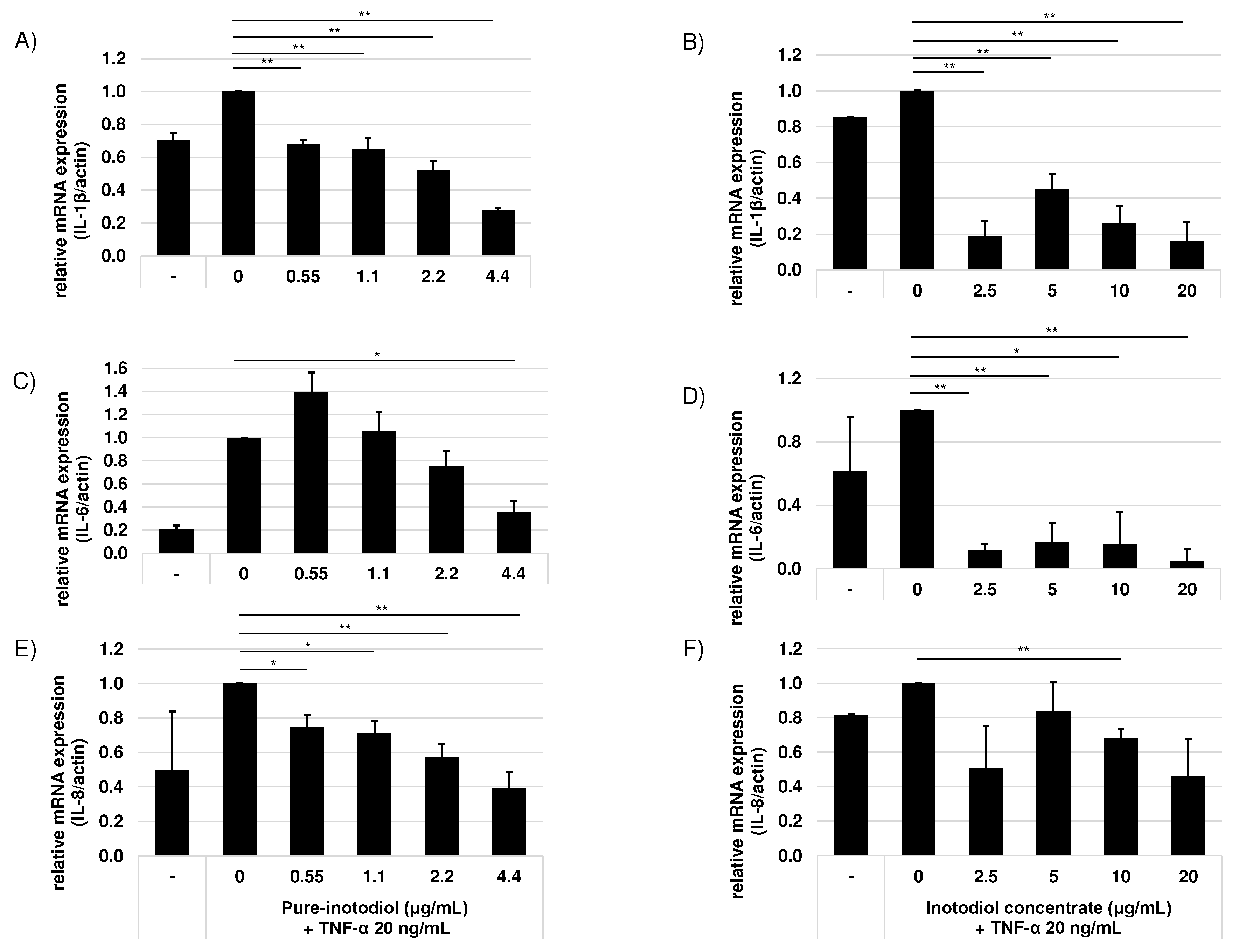
Figure 4.
Effects of pure inotodiol (A, C, and E) and the inotodiol concentrate (B, D, and F) on expression of pro-inflammatory cytokines of HaCaT cells induced by UVB exposure (10 mJ/cm2). Statistical significance was denoted as * for p < 0.05 and ** for p < 0.01.
Figure 4.
Effects of pure inotodiol (A, C, and E) and the inotodiol concentrate (B, D, and F) on expression of pro-inflammatory cytokines of HaCaT cells induced by UVB exposure (10 mJ/cm2). Statistical significance was denoted as * for p < 0.05 and ** for p < 0.01.
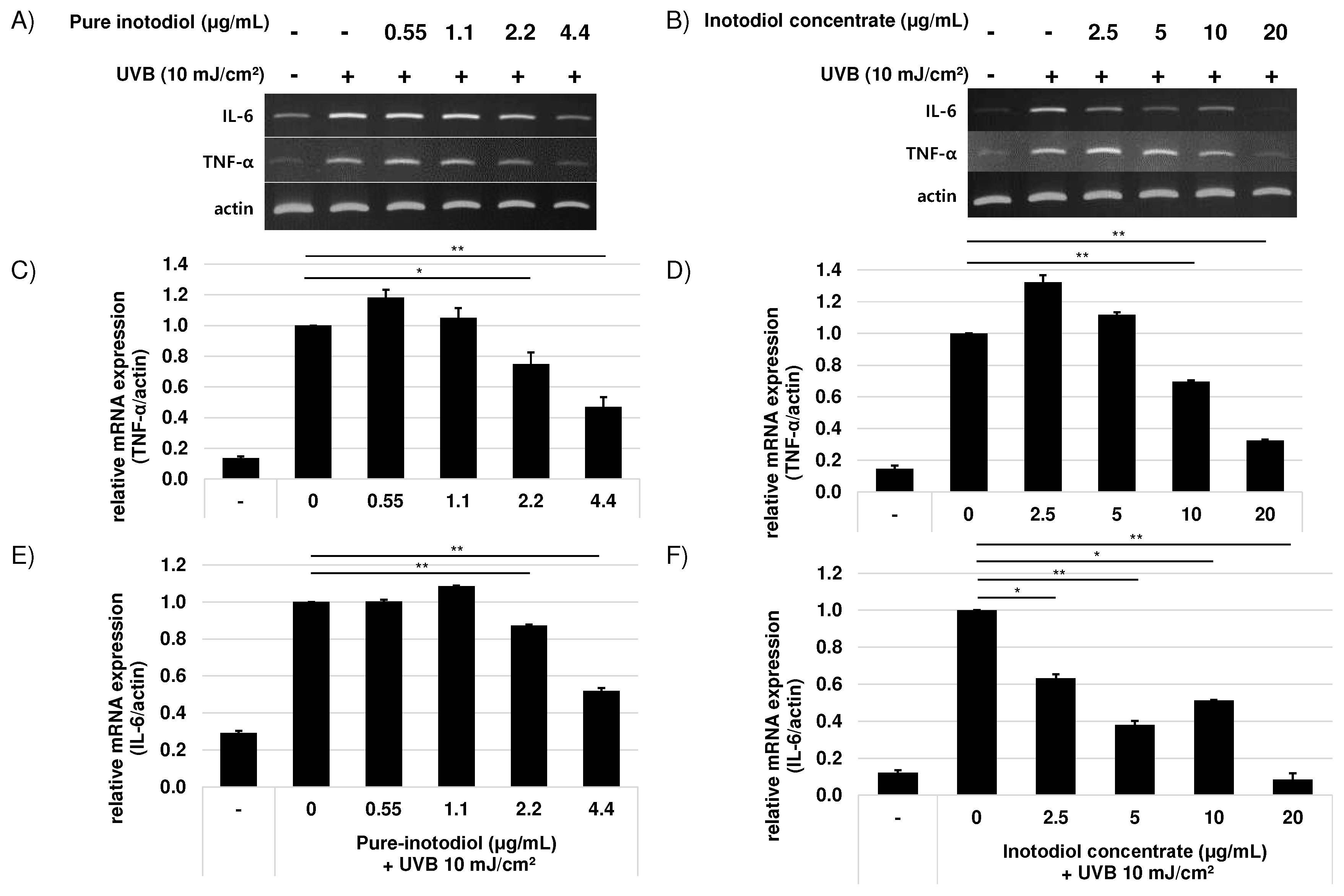
Figure 5.
Effects of pure inotodiol (A, C, E, and G) and the inotodiol concentrate (B, D, F, and H) on expression of genes involved in collagen (COL1α2) and hyaluronan (HAS-2 and HAS-3) synthesis in HaCaT cell. Statistical significance was denoted as * for p < 0.05 and ** for p < 0.01.
Figure 5.
Effects of pure inotodiol (A, C, E, and G) and the inotodiol concentrate (B, D, F, and H) on expression of genes involved in collagen (COL1α2) and hyaluronan (HAS-2 and HAS-3) synthesis in HaCaT cell. Statistical significance was denoted as * for p < 0.05 and ** for p < 0.01.
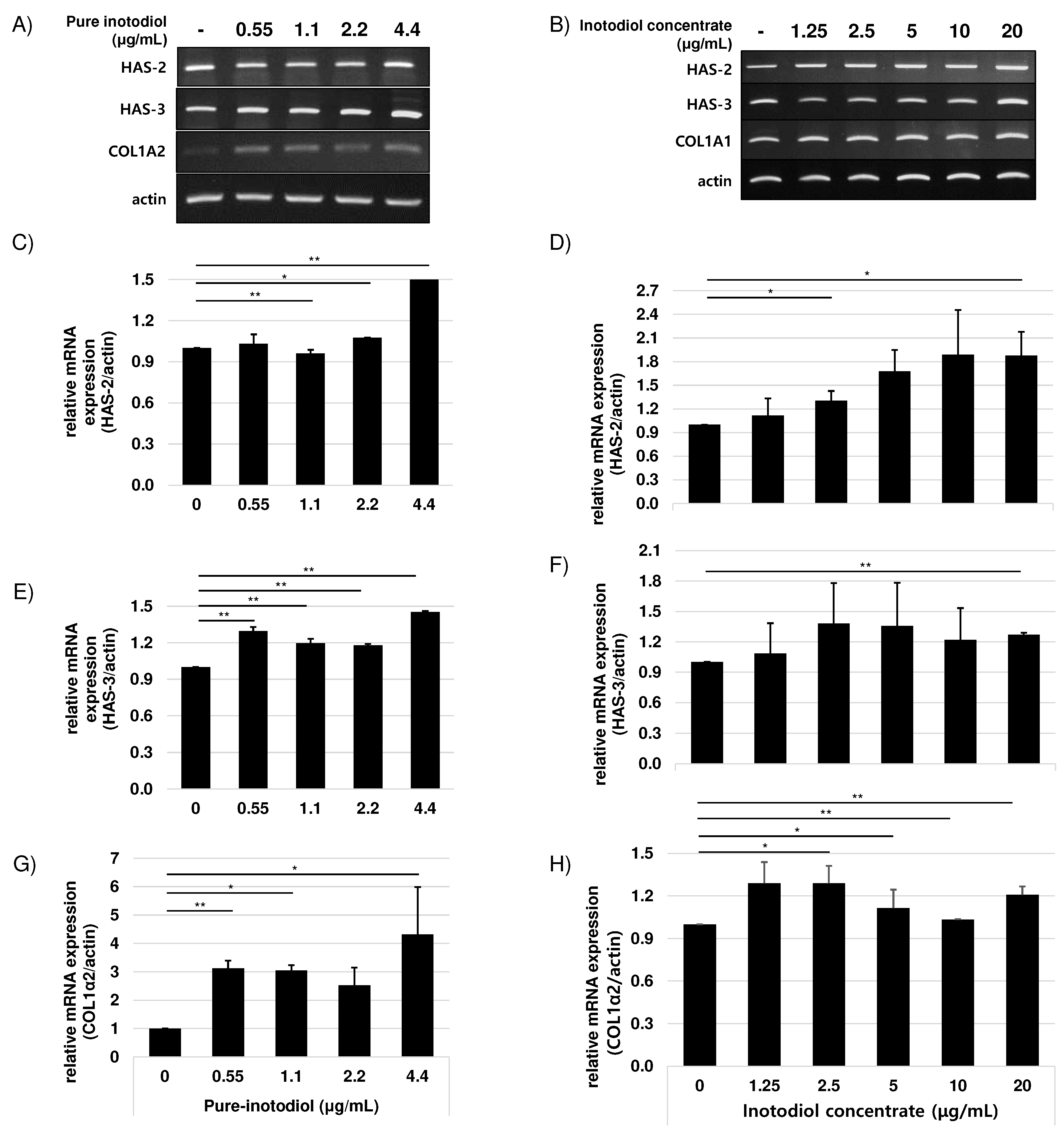
Figure 6.
Effects of pure inotodiol on expression of genes involved in hyaluronan (HYAL-3) degradation in HaCaT. Pro-inflammatory TNF-α was used for inducing inflammation to HaCaT cells. Statistical significance was denoted as * for p < 0.05 and ** for p < 0.01.
Figure 6.
Effects of pure inotodiol on expression of genes involved in hyaluronan (HYAL-3) degradation in HaCaT. Pro-inflammatory TNF-α was used for inducing inflammation to HaCaT cells. Statistical significance was denoted as * for p < 0.05 and ** for p < 0.01.
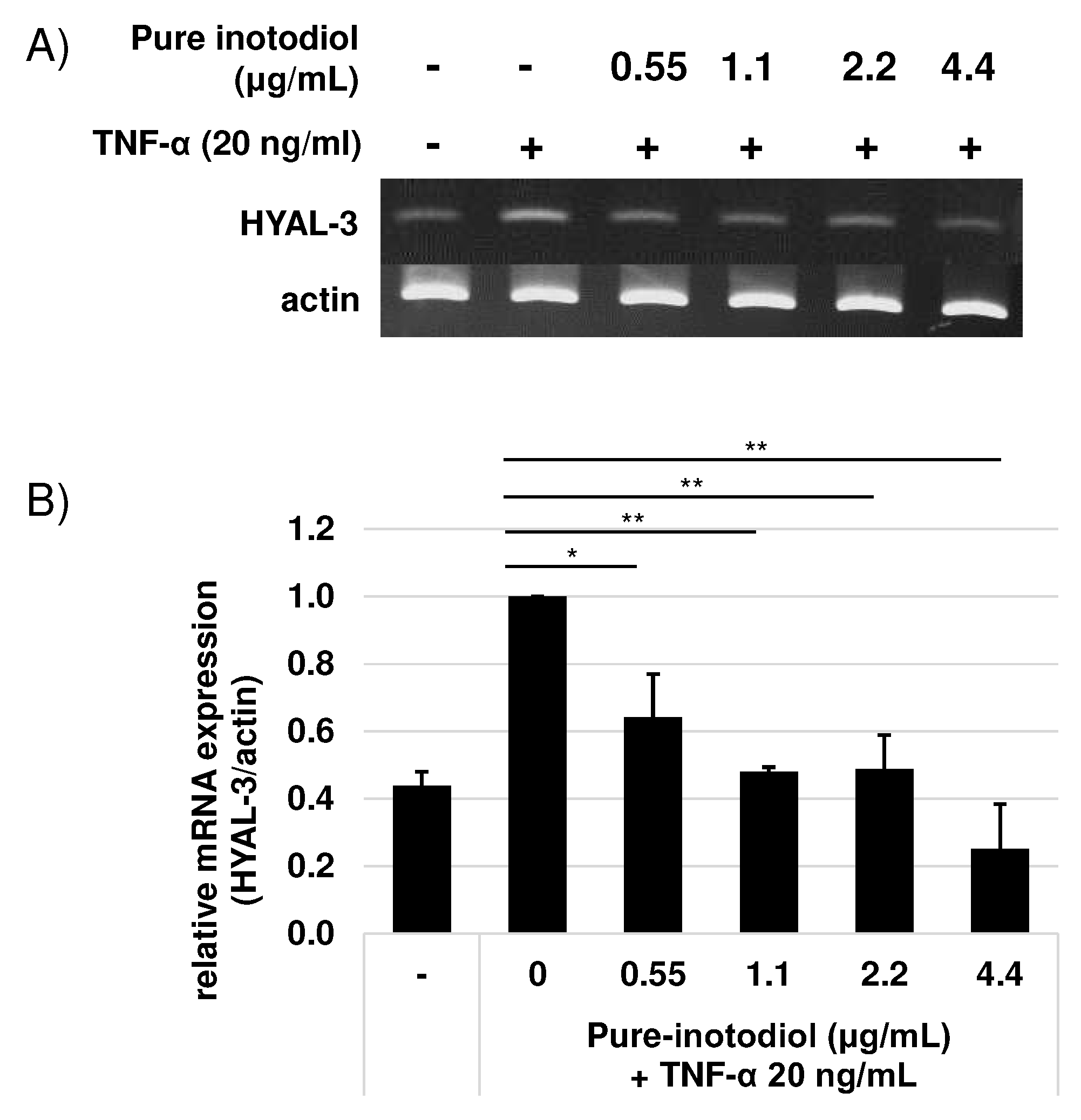

Table 1.
Genes and PCR primers used in this study.
| Gene | Forward | Reverse |
|---|---|---|
| GAPDH | AAGTGGATATTGTTGCCATC | ACTGTGGTCATGAGTCCTTC |
| IL-6 | ATGAACTCCTTCTCCACAAGC | GTTTTCTGCCAGTGCCTCTTTG |
| IL-8 | TCTGTGTGAAGGTGCAGTT | AGCCCTCTTCAAAAACTTCT |
| IL-1β | AAACAGATGAAGTGCTCCTTCCAGG | TGGAGAACACCACTTGTTGCTCCA |
| TNF-α | GAGCTGAGAGATAACCAGCTGGTG | CAGATAGATGGGCTCATACCAGGG |
| HAS-2 | TGGGTGTGTTCAGTGCAT | GCATTGTACAGCCATTCTCG |
| HAS-3 | CCCAGCCAGATTTGTTGATG | AGTGGTCACGGGTTTCTTCC |
| COL1A1 | AGCCAGCAGATCGAGAACAT | TCTTGTCCTTGGGGTTCTTG |
| COL1A2 | TCAAGGTTTCCAAGGACCTG | GTGTCCCCTAATGCCTTTGA |
| HYAL-3 | CAGTCCATTGGTGTGAGTG | CACAGGTGTAGAAAGGCTTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



