
Submitted:
30 June 2023
Posted:
03 July 2023
You are already at the latest version
Abstract
The octopus keel is a trait that has been hypothesized to be connected with burrowing in octopuses, but has never been explored in any detail. This research aimed to test for anWe investigated the association between these two traits phylogenies using two approaches independent sets of methods. First, we examined phylogenetic correlation between presence of a keel and known burrowing behavior in incirrate octopuses.by using a phylogenetic approach, benthic octopuses as a group were examined for known burrowing and the presence of a keel to determine whether an evolutionary relationship exists between these traits. Second, burrowing and non-burrowing captive Muusoctopus leioderma were evaluated for keel prominence to determine whether the keel is lost more rapidly in non-burrowing individuals. Pagel’s test for the coevolution of binary characteristics showed the model of best fit for the resulting phylogenetic tree to be one of evolutionary interdependence, and that non-burrowing Muusoctopus leioderma lost their keels over time, while burrowing individuals maintained their keels. Together, these results indicate the keel may be a trait associated with burrowing in octopuses.
Keywords:
octopus
; burrowing
; keel
; leioderma
; sediment
; phylogeny
; evolution
1. Introduction
Marine sediments are the most expansive and pervasive environments on Earth [1]. These seemingly barren underwater habitats make up roughly 70% of Earth’s surface, yet their depths and temperatures make them the least accessible environments possible for humans [1]. These habitats host a diverse range of marine life, and are largely dominated by burrowing organisms. Little is known about the behavior and ecology of these marine infauna, as the difficulty of examining these habitats has stymied their research [1].
Octopuses are a diverse group of invertebrates found in most marine environments. Charismatic, highly intelligent, and displaying a wide range of novel adaptations for invertebrates, this group plays an important role, both in ecological systems, and in facilitating public engagement and interest in science [2]. Octopuses act as both generalist predators and prey, playing crucial roles in trophic webs and ecological health [3,4,5]. Cephalopods, including octopuses, are also expanding in range and population [5,6,7,8] suggesting that their ecological significance will grow, and sparks questions about octopus’ resilience and adaptation.
The smooth-skinned octopus (Muusoctopus leioderma) is a deep-water species and most commonly found between 450-650 meters [9] throughout the northern Pacific ocean from California up to the Sea of Okhotsk off Siberia, and until recently had never been recorded at depths shallower than 70 meters [10,11]. However, a population of M. leioderma has recently been found to reside in Burrows Bay, WA, USA, at only 15 m depth and accessibly by SCUBA[12]. This exciting discovery allows for collection and study of live and minimally disturbed benthic octopuses via SCUBA that would normally be inaccessible.
This species actively modifies its environment by burrowing, an adaptation allowing organisms to move through and compact soft substrata to form semi-permanent structures [13]. Burrowing organisms have an unusual level of influence on their environment by directly altering marine sediment biochemistry and structure [14] and indirectly by impacting co-occurring bacteria, microalgae, macrofauna, seagrasses, and other secondary consumers affected by sediment nutrition [14].
Muusoctopus leioderma’s most notable defining characteristic is the presence of a keel, a flap of skin that protrudes from the lateral margin of the mantle [12]. This trait, shared with some other species of octopuses, and has been hypothesized to be linked to burrowing in several benthic species [15]. The keel is essentially just an expandable flap of skin, and lacks rigidity, and is therefore unlikely to be used directly for excavation. However, the keel’s apparent connection with known burrowing octopus species makes it an intriguing point of study as a possible burrowing trait for benthic octopuses and may lend clues to the burrowing strategy of M. leioderma and other octopuses.
2. Materials and Methods
2.1 Octopus Collection
Octopuses were collected from Burrows Bay, Skagit County, WA (48°28′12″ N, 122°40′53″ W) by SCUBA. Once found, they were captured and returned to Rosario Beach Marine Laboratory (RBML) in re-sealable plastic storage bags filled with saltwater. Upon arrival at RBML, octopuses were sexed, weighed, and assigned a name. Lengths were also obtained in the laboratory using the same laser dot camera method used to measure lengths during BRI recordings in situ (see below). These data, along with species, date caught, depth, the tank number the octopus were to be placed in, and any notes were recorded. Then octopuses were immediately transferred to a 61 cm x 33 cm x 41 cm acrylic aquarium, or “mud tank”, filled approximately halfway with sediment from Burrows Bay for their 24-hour laboratory acclimation period. The time that octopuses begin this acclimation period, along with the date, sex, name, and tank were recorded in Google Sheets.
2.2 Octopus Holding
Aquaria filled with sediment collected from Burrow Bay or “mud tanks” were connected to RBML’s seawater system to provide fresh saltwater. Tanks were modified for a flow-through system, with an inflow and passive outflow (Figure 1). Tanks outflow lines ran to a single plastic tub with a sump pump connected to a float switch, allowing all tanks to have short lines and consistent outflow to a single drain. Laboratory lighting conditions were controlled and matched with natural lighting conditions in Burrows Bay using custom lights. These lights were diffused green LEDs, controlled by a Raspberry Pi Pico microcontroller, and powered by a DC wall socket. Each mud tank was equipped with a light array programmed to brighten and dim closely following a diurnal cycle approximating what occurs naturally. Peak brightness hours and ramp down periods were calculated using “sunriset” function from the maptools R package [16] using July 31, 2022, a date roughly halfway through our data collection period, as a set date, and Burrows Bay, Skagit County, Washington, USA as our set location. There were also red light sources that were constantly illuminated and allowed nighttime photography of octopuses when the green lights were off without disturbing them. Muusoctopus leioderma’s eyes are non-reactive (no discernable pupillary response) to red light (pers obs).
Mud for the tanks was collected directly from Burrows Bay either by SCUBA or using a box core sampler or Van Veen grab samplers. Octopuses were held for at least 12 days, then periodically released by divers while new individuals were collected.
2.3 Muusoctopus leioderma Keel Prominence
After their initial acclimation period, captured individuals were periodically evaluated for keel prominence by taking images of the octopus’s keel with an iPhone 12 mini. Target keel days were days 2, 6, and 12 in captivity. However, keels could not be photographed when the octopus was in its burrow, so occasionally octopuses would have to be photographed later, as soon as the octopus was visible. Images were normally taken either in the early morning, or late at night. Octopuses were kept in treatment for a target of 3 keel collection dates in the mud tanks, although time restraints forced treatment to end early in some cases. After completing their treatment in the mud tanks, individuals were transferred to non-light controlled plastic containers, or “Octocondos,” connected to RBML’s saltwater system to await return and release to Burrows Bay via SCUBA, and allowing new octopuses to begin treatment. In some cases, the octopuses never emerged from their burrow after their initial burrowing event. These octopuses were dug out of their burrows after 10-20 days. Occasionally, such as when octopuses needed to be dug out from their burrows, images for keel evaluations were collected after octopuses were transferred to the Octocondos, and this was noted in the data. Later, individuals were categorized into two groups: burrowers, or octopuses that burrowed below the sediment layer, and non-burrowers, or octopuses that did not burrow in captivity.
Keels were evaluated qualitatively based on 5 levels of prominence, with a 0 or “Missing” being a completely absent keel, and a 4 or “Very Prominent” being the strongest level of prominence using a key created for this purpose (Appendix A). The key provided descriptions of distinctions between each of the levels of prominence, as well as multiple images highlighting those differences. Keels were evaluated independently by 4 observers, and the consistency of the evaluations was examined. These were blind evaluations, meaning evaluators did not know if images were of burrowing or non-burrowing octopuses, and they lacked access to other evaluators’ results.
Each observer had their own private sheet to perform evaluations independently from other observers. These results were averaged to get keel prominence values for each keel measurement of all individual octopuses. Individual octopuses were split into two groups based on burrowing status as burrowers and non-burrowers. Keel evaluations were categorized as either mud tank or Octocondo evaluations. Individuals that had one or fewer keel measurements were excluded because keel loss could not be determined. Octopuses’ keel prominence was compared with how many days it had been held in captivity, which was calculated using the dates of octopus collections and keel evaluations.
To test consistency between evaluators, we calculated an intraclass correlation coefficient (ICC) as an index of interrater reliability. ICC was run as a two way consistency model using the irr package in R [17].
Statistically significant differences in keel prominence between burrowing and non-burrowing octopuses were analyzed using a Linear Mixed Effect (LME) model from the nlme package in R [18]. The LME model also accounted for two different keel measurement scenarios: mud tanks or Octocondos. This model was analyzed using an ANOVA test from the car R package [19].
2.4 Octopoda Keel and Phylogenetics
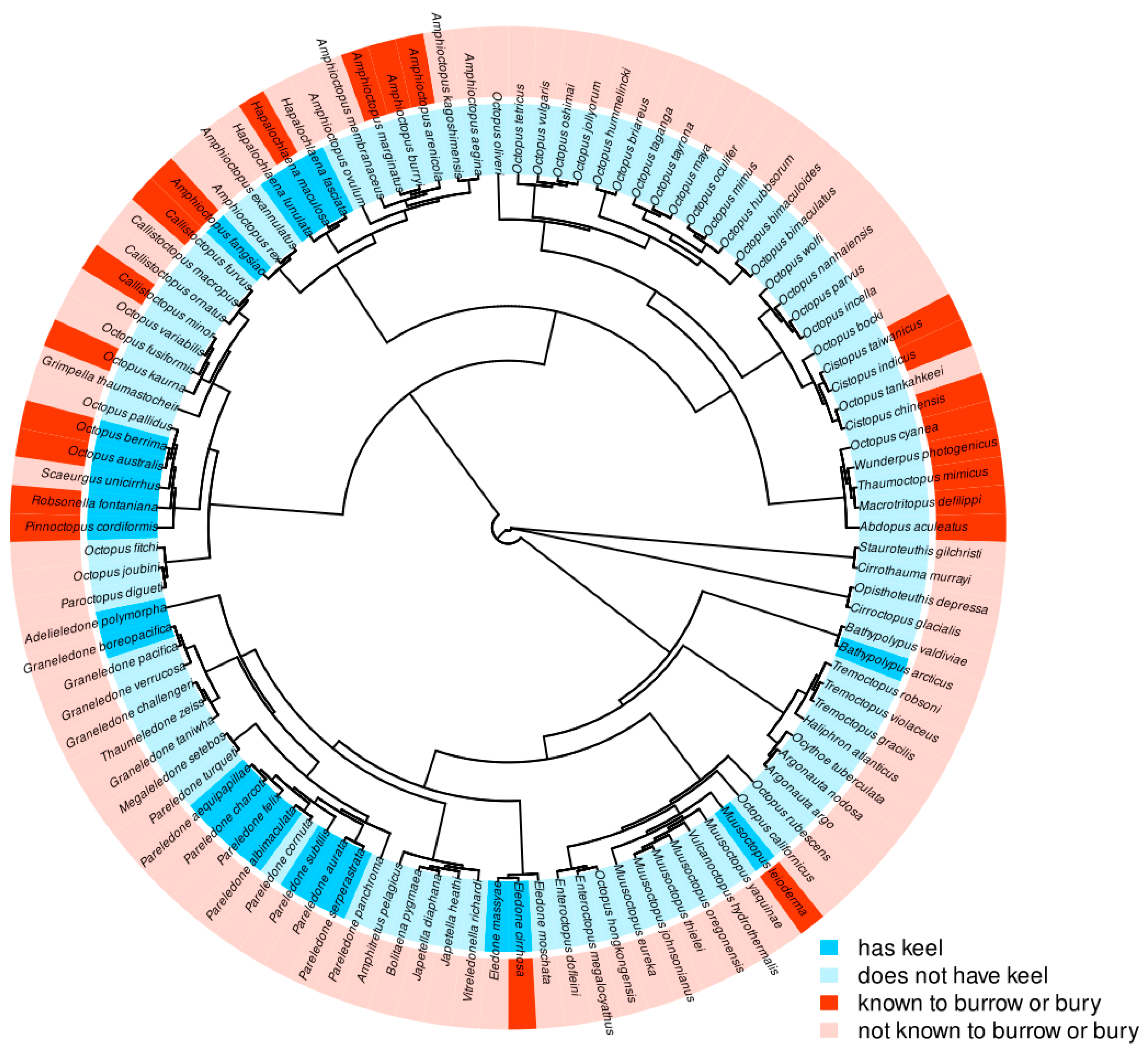
Phylogenetic analysis of the occurrence of keels across benthic octopuses in the order Octopoda in relation to burrowing was accomplished using visual identification of keels as “Present” or “Absent” by examining images from literature, species descriptions and reaching out to species-specific experts. These species were classified as “known to bury or burrow” or “not known to bury or burrow.” The octopus species selected for analysis were all benthic species with known genetic sequencing of the cytochrome oxidase subunit 1 (COI), cytochrome oxidase subunit 3 (COIII), and/or 16S genes obtained from GenBank. These genes were chosen because they provided the best coverage of the greatest number of species, and each of these species had sequences of one or more of these genes available, sufficient for an alignment. Species lacking genetic sequencing, or that lacked data on both keels and burrowing behavior, were mostly deep-water species with very little research focus, and these were omitted from the analysis. The final tree included 110 total species. Octopuses in which only keel presence was unknown, the keel was considered missing. Octopuses in which only burrowing behavior was unknown were considered “not known to bury or burrow”. Evolutionary correlation between mantle keel and burrowing or burying behavior were tested using Pagel's test in R.
Genetic sequences were obtained from GenBank using the read.GenBank function from the ape R package using accession numbers [20] for benthic octopus species with known sequencing of COI, COIII, and 16S, as these 3 genes provided the best genetic coverage of benthic octopuses. Sequences were aligned using the AlignSeqs function from the DECIPHER package [21]. This allowed for the creation of a NEXUS file that could then be used in creating the final tree. For each of the three genes, a model test was conducted with the phangorn package for each codon position independently, so the dataset could be partitioned into each of the three codon positions [22].
The resulting files were combined into a single nexus file and a MrBayes command block generated using a custom BASH script. This file was then used for the final analysis and production of the multi-gene tree using MrBayes [23].
To test for a statistically significant evolutionary relationship between keel presence/absence and known burrowing behavior, the multi-gene tree was analyzed using Pagel’s test. Pagel’s test examines the independent evolution of 2 binary characters by comparing a ratio of likelihoods of two models [24]. Pagel’s test was run using the phytools R package [25].
3. Results
3.1. Keel Regression in Non-Burrowing Muusoctopus leioderma
Keel evaluations of captive octopuses showed a clear effect of burrowing on keel prominence. Keel evaluations were conducted blindly and independently by four observers, and results were consistent (ICC = 0.913, p-value < 0.001, 95%-Confidence Interval: 0.85 < ICC < 0.953). Average keel prominence values of the linear mixed effects model with an ANOVA analysis showed a statistically significant interaction between days and burrowing status (Figure 2, Χ2 = 4.6648, df = 1, p-value = 0.03079). There was also a significant effect of day on keel prominence (Figure 2, Χ2 = 4.3657, df = 1, p-value = 0.03667). Non-burrowing octopuses lost their keel more rapidly than burrowing octopuses . In addition, the analysis found that whether the image was taken in the mud tank or in the Octocondo after removal from the mud tank did not have a significant effect on these results (linear mixed-effect model, Χ2 = 0.8436, p-value = 0.35837).
3.2. Keel Evolution in Burrowing Octopoda
The phylogenetic tree when analyzed using Pagel’s test found the model of best fit to be one of interdependence between the two tested traits, meaning the tree may demonstrate evidence of an interdependent evolutionary relationship between the presence of a keel and known burrowing behavior in benthic octopus species (Figure 3, Pagel’s test, AIC = 188.4336, likelihood ratio = 20.5514, p-value = 0.000388571).
4. Discussion
In burrowing captive Muusoctopus leioderma, we saw a significant interaction between the effect of octopus burrower/non-burrower status and number of days in the mud tank on keel prominence, meaning that the condition of an octopus being a burrower or non-burrower impacted how keel prominence changed over time. Specifically, we saw a stronger negative relationship between keel prominence and time in captivity for non-burrowers than for burrowers (Figure 2). This suggests that the keel is better maintained when octopuses continue burrowing in captivity. The mechanism behind this is still unclear, as the precise function of the keel is still unknown. It is possible that the keel is a “use it or lose it” trait, and that the keel disappears when the octopus does not continue the behavior. The keel loss could also be a result of stress from captivity, and that burrowing individuals were less prone to stress in their tanks, despite the minimal disturbance and light-controlled nature of the experiment.
Then, we broadened focus to benthic octopuses as a whole to explore the possibility of an evolutionary relationship between known burrowing behavior and keel presence. Pagel’s test is a statistical test that compares models of evolutionary relationships between binary traits, and this test revealed that the model of best fit for the phylogenetic tree was one of interdependence, suggesting the two traits co-evolved. While keels have been anecdotal linked to burrowing in octopuses [15], this is the first study to provide phylogenetic evidence of a link between the two traits.
It was originally speculated that the keel may act as a kind of rudder, allowing the octopus to orient and guide itself into and/or through the sediment [15]. Octopus keels are essentially a flap of skin with very little rigidity or structure, so it is less likely that it functions directly as an excavating appendage to displace sediment. However, the extra skin is the only external textural feature in this species, as M. leioderma lacks any papillae or other prominent skin texture. This unusually smooth and streamlined profile is ideal for burrowing, as it may help to reduce resistance from marine sediment. The keel is the only thing that disrupts this low resistance body shape, but the notion that this skin is being used to displace sediment is unlikely.
Instead, the keel may be involved in improving the efficiency of burrowing. In general, there are advantages for burrowing organisms with smaller body sizes. Smaller animals are faster and expend less energy in making their burrows compared to larger organisms [26,27]. However, there are also advantages to constructing a larger burrow. For example, a larger burrow would allow for improved water flow and could reduce the risk of anoxia in such a confined environment. The width of a burrow is usually proportional to the size of the organism [28], but larger individuals burrow more slowly [29] and expend more energy than smaller individuals [26,27]. A keel may help to offset these issues while allowing the octopus to build a larger burrow.
To accomplish this, the keel may act as an extra reservoir of skin that allows the octopus to expand its mantle during the burrowing process. Octopuses expand and retract their mantles as part of the normal respiration process, and octopus skin is very flexible, normally retracting into a highly textured surface of papillae or folds [30]. Muusoctopus leioderma’s skin texture is concentrated at the lateral margin of the mantle, but the mantle still expands and retracts during respiration. It’s possible that the extra skin allows the octopus to expand its body size while burrowing beyond what would be possible otherwise, allowing for a disproportionately large burrow, and overall less resistive skin for burrowing. Additionally, mantle expansion may also be a part of this octopus’s overall burrowing strategy. The razor clam (Ensis directus) utilizes a strategy of bodily expansion and retraction in its burrowing, and it is possible that M. leioderma could be burrowing in a similar way [31].
The elastic nature of octopus skin may also explain the loss of keels in captive non-burrowers. Individuals no longer expanding their mantles during burrowing could show reduced keels as the extra skin is pulled back into the mantle from disuse. There is currently no research on the impacts of inactivity on octopus skin, but inactivity in humans results in reduced skin elasticity, muscle mass, tone, and alteration of skin and muscle fiber composition [32].
Alternatively, the keel may just be an adaptation connected with M. leioderma’s smooth skin to improve sediment resistance. Next steps would involve trying to get M. leioderma to burrow in a transparent medium in the laboratory to document the keel below the sediment, and to investigate interspecific variation in mantle plasticity and skin texture to explore connections between octopus skin and burrowing behavior.
5. Conclusions
Burrowing is a life-strategy closely linked with marine sediment and deep-sea environments, and this connection makes burrowing an ideal target for understanding these inaccessible and understudied ecosystems. The evolution of phenotypes that facilitate this behavior varies based on both organism and environment. This study found evidence that the keel may be a burrowing associated traits in octopuses using two independent sets of methods. Phylogenetic analysis of known burrowing and keel presence in benthic octopuses provided a model of best fit of evolutionary interdependence between keels and known burrowing. Meanwhile, individual non-burrowing Muusoctopus leioderma were shown to lose their keel faster compared with burrowing individuals. Both of these results are what we’d expect to see if this were a burrowing-associated trait, and together indicate this is a phenotype linked with burrowing in octopuses.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Octopus holding and laboratory setup; Figure S2: Keel prominence values vs time in captivity for burrowing and non-burrowing individuals; Figure S3: Multi-species phylogenetic tree showing keel presence and known burrowing.
Author Contributions
Conceptualization, C.M.S. and K.L.O.; methodology, C.M.S. and K.L.O.; software, C.M.S. and K.L.O.; validation, C.M.S., K.C., C.H., G.E. and K.L.O.; formal analysis, C.M.S., K.C., C.H., G.E.; investigation, C.M.S.; resources, K.L.O.; writing—original draft preparation, C.M.S.; writing—review and editing, C.M.S. and K.L.O; visualization, C.M.S. and K.L.O.; supervision, C.M.S.; project administration, C.M.S. and K.L.O.; funding acquisition, C.M.S and K.L.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Walla Walla University, as well as by a scholarship provided by the American Academy of Underwater Sciences.
Data Availability Statement
The code for Muusoctopus leioderma keel evaluations presented in this study are openly available in Zenodo at https://doi.org/10.5281/zenodo.8021840. The code for octopus keel phylogenetic evaluations and accession numbers used in this study are openly available in Zenodo at https://doi.org/10.5281/zenodo.8021847. Keel evaluation images used in this study are openly available in Zenodo at https://doi.org/10.5281/zenodo.8040502.
Acknowledgments
Thank you to Walla Walla University and the American Academy of Underwater Sciences for their financial support of this research. Thanks to our field support: Dr. Alan Verde, Dr. Jim Nestler, Dr. Cecilia Brothers, Macky Tapley, Emily Stone, Nathan Iwakowshi, Jesse Humbert, Miranda Wredburg, Julia Beaudoin, Ricky Wright, and Ashley Morrow. Thank you for the personal communication from Drs. James Wood, Carlos Vazquez, Michael Vecchione, Jurgen Guerrero, Louise Allcock, Steve O’ Shea, and Christian Ibanez.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A: Keel Evaluation Key
Very Prominent (VP): Keel is highly distinctive and easily seen as a flap of skin protruding strongly from the lateral mantle. Keel appears as a ring of skin protruding strongly from the mantle when observed dorsally (from above) and the area of the skin extends beyond a simple skin line or ridge. Keel would likely “flap” if octopus were in motion.
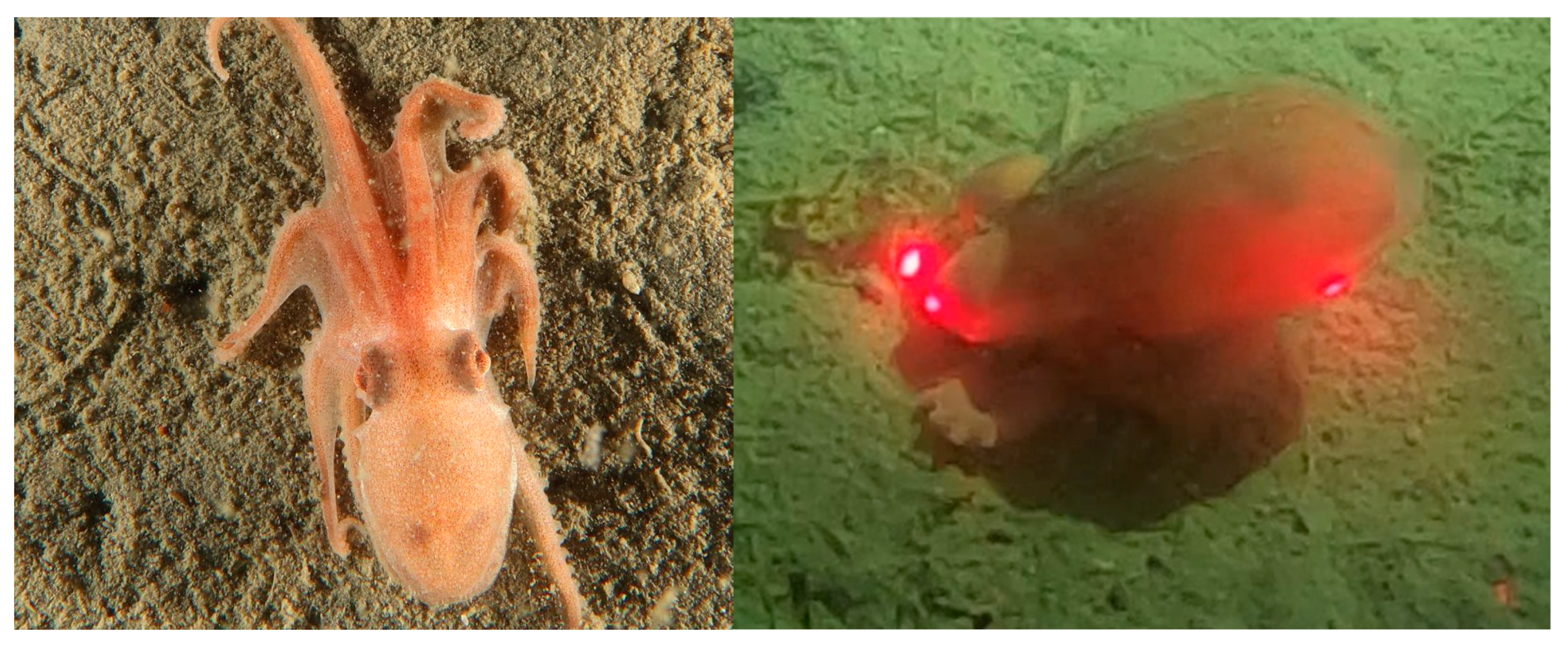
Prominent (P): Keel is distinctive and easily seen as a small skin fold protruding from the lateral mantle. Keel appears as a thick line protruding strongly from the mantle when observed dorsally. Keel is more robust than a simple skin ridge, but would likely not “flap” if octopus were in motion.

Apparent (A): Keel is visible as a ridge of skin protruding from the lateral mantle. Keel appears as a narrow line protruding from the mantle when observed dorsally. Keel is apparent as a skin ridge, but does not demonstrate excess flappy skin.

Weak (W): Keel is faintly visible as a line of skin, but does not protrude from the mantle. Keel is not readily observable from a dorsal view, and can only be seen upon close examination of the lateral mantle.
Missing (M): No distinctive lateral mantle ridge line is visible. Keel is completely missing from the octopus and no line can not be readily seen from any view.
References
- Dorgan, K.M.; Arwade, S.R.; Jumars, P.A. Burrowing in Marine Muds by Crack Propagation: Kinematics and Forces. Journal of Experimental Biology 2007, 210, 4198–4212. [Google Scholar] [CrossRef] [PubMed]
- Misra, M.; Miller, M.L. Marine Conservation Tourism and the Giant Pacific Octopus: A SWOT Analysis of Two Public Engagement Programs and the Viability of a Hybrid Program at the Seattle Aquarium, Washington, USA. Regional Studies in Marine Science 2022, 52, 102231. [Google Scholar] [CrossRef]
- Piatkowski, U.; Pierce, G.; Morais da Cunha, M. Impact of Cephalopods on the Food Chain and Their Interaction with the Environment and Fisheries: An Overview. Fisheries Research 2001, 52, 5–10. [Google Scholar] [CrossRef]
- Xavier, J.C.; Allcock, A.L.; Cherel, Y.; Lipinski, M.R.; Pierce, G.J.; Rodhouse, P.G.K.; Rosa, R.; Shea, E.K.; Strugnell, J.M.; Vidal, E.A.G.; et al. Future Challenges in Cephalopod Research. Journal of the Marine Biological Association of the United Kingdom 2015, 95, 999–1015. [Google Scholar] [CrossRef]
- ICES Working Group on Cephalopod Fisheries and Life History (WGCEPH). 2020. [CrossRef]
- Ramos, J.E.; Pecl, G.T.; Semmens, J.M.; Strugnell, J.M.; León, R.I.; Moltschaniwskyj, N.A. Reproductive Capacity of a Marine Species (Octopus Tetricus) within a Recent Range Extension Area. Marine and Freshwater Research 2015, 66, 999–1008. [Google Scholar] [CrossRef]
- Domínguez-Contreras, J.F.; Ceballos-Vázquez, B.P.; Hochberg, F.G.; Arellano-Martínez, M. A New Record in a Well-Established Population of Octopus Hubbsorum (Cephalopoda: Octopodidae) Expands Its Known Geographic Distribution Range and Maximum Size. American Malacological Bulletin 2013, 31, 95–99. [Google Scholar] [CrossRef]
- Doubleday, Z.A.; Prowse, T.A.; Arkhipkin, A.; Pierce, G.J.; Semmens, J.; Steer, M.; Leporati, S.C.; Lourenço, S.; Quetglas, A.; Sauer, W.; et al. Global Proliferation of Cephalopods. Current Biology 2016, 26, R406–R407. [Google Scholar] [CrossRef]
- Conners, M.E.; Conrath, C.; Aydin, K. Assessment of the Octopus Stock Complex in the Bering Sea and Aleutian Islands; Bering Sea and Aleutian Islands stock assessment and fisheries evaluation; North Paci fi c Fisheries Management Council: Anchorage, AK, 2014. [Google Scholar]
- Hochberg, F.G. Class Cephalopoda. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and the Western Santa Barbara Channel Vol 8; Scott, P.V., Blake, J.A., Eds.; Santa Barbara Museum of Natural History: Santa Barbara, California, 1998; Vol. 8, pp. 175–235. [Google Scholar]
- Conners, M.E.; Aydin, K.; Conrath, C. Assessment of the Octopus Stock Complex in the Bering Sea and Aleutian Islands. 2016, 56.
- Kore, L.G.; Johnson, B.M.; Hayden, M.; Winters, M.; Onthank, K.L. 4 The Odd Octopus: Morphological and Molecular Identification, Burrowing 5 Behavior, and Temperature Preference of a Deep-Water Octopus Found in a 6 Shallow-Water Bay. 35.
- Dorgan, K. The Biomechanics of Burrowing and Boring. The Journal of experimental biology 2015, 218, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Pillay, D.; Branch, G. Bioengineering Effects of Burrowing Thalassinidean Shrimps on Marine Soft-Bottom Ecosystems. In Oceanography and Marine Biology; Gordon, J., Ed.; Oceanography and Marine Biology - An Annual Review; CRC Press, 2011; Vol. 20115434 ISBN 978-1-4398-5364-1.
- Norman, M.; Reid, A. Guide to Squid, Cuttlefish and Octopuses of Australasia. In Guide to squid, cuttlefish and octopuses of Australasia; CSIRO publishing, 2000; pp. 17, 33, 35, 62, 77.
- Bivand, R.; Lewin-Koh, N. Maptools: Tools for Handling Spatial Objects; 2022.
- Gamer, M.; Lemon, J. Singh <puspendra.pusp22@gmail.com>, I.F.P. Irr: Various Coefficients of Interrater Reliability and Agreement, 2019. [Google Scholar]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer: New York, 2000. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Third.; Sage: Thousand Oaks CA, 2019. [Google Scholar]
- Paradis, E.; Schliep, K. Ape 5.0: An Environment for Modern Phylogenetics and Evolutionary Analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.S. Using DECIPHER v2. 0 to Analyze Big Biological Sequence Data in R. R Journal 2016, 8. [Google Scholar] [CrossRef]
- Schliep; Klaus; Potts; J, A.; Morrison; A, D.; Grimm; W, G. Intertwining Phylogenetic Trees and Networks. Methods in Ecology and Evolution 2017, 8, 1212–1220. [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Revell, L.J. Exercise 9: Testing for an Evolutionary Correlation between Binary Characters Using Pagel’s (1994) Method. Available online: https://www.phytools.org/Cordoba2017/ex/9/Pagel94-method.html (accessed on 17 February 2022).
- Revell, L.J. Phytools: An R Package for Phylogenetic Comparative Biology (and Other Things). Methods in Ecology and Evolution 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Lovegrove, B.G. The Cost of Burrowing by the Social Mole Rats (Bathyergidae) Cryptomys Damarensis and Heterocephalus Glaber: The Role of Soil Moisture. Physiological Zoology 1989, 62, 449–469. [Google Scholar] [CrossRef]
- Wu, N.; Alton, L.; Clemente, C.; Kearney, M.; White, C. Morphology and Burrowing Energetics of Semi-Fossorial Skinks (Liopholis Spp.). Journal of Experimental Biology 2015, 218, 2416–2426. [Google Scholar] [CrossRef]
- Schlacher, T.A.; Lucrezi, S.; Peterson, C.H.; Connolly, R.M.; Olds, A.D.; Althaus, F.; Hyndes, G.A.; Maslo, B.; Gilby, B.L.; Leon, J.X.; et al. Estimating Animal Populations and Body Sizes from Burrows: Marine Ecologists Have Their Heads Buried in the Sand. Journal of Sea Research 2016, 112, 55–64. [Google Scholar] [CrossRef]
- de la Huz, R.; Lastra, M.; López, J. The Influence of Sediment Grain Size on Burrowing, Growth and Metabolism of Donax Trunculus L. (Bivalvia: Donacidae). Journal of Sea Research 2002, 47, 85–95. [Google Scholar] [CrossRef]
- Madan, J.; Wells, M. Cutaneous Respiration in Octopus Vulgaris. Journal of experimental biology 1996, 199, 2477–2483. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Tao, J. Modeling Clam-Inspired Burrowing in Dry Sand Using Cavity Expansion Theory and DEM. Acta Geotech. 2020, 15, 2305–2326. [Google Scholar] [CrossRef]
- Demangel, R.; Treffel, L.; Py, G.; Brioche, T.; Pagano, A.F.; Bareille, M.-P.; Beck, A.; Pessemesse, L.; Candau, R.; Gharib, C.; et al. Early Structural and Functional Signature of 3-Day Human Skeletal Muscle Disuse Using the Dry Immersion Model. The Journal of Physiology 2017, 595, 4301–4315. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Octopus holding and laboratory setup, with water system, lighting, and all 5 experimental mud tanks shown.
Figure 1.
Octopus holding and laboratory setup, with water system, lighting, and all 5 experimental mud tanks shown.

Figure 2.
Keel prominence values vs time in captivity for burrowing and non-burrowing individuals. Individual shapes connected by thin lines show keel prominence for individual octopuses during captivity, with blue showing burrowing individuals, and red showing non-burrowers. Thick lines show results of LME. Results show a significant interaction between burrowing and day, with non-burrowing octopuses losing their keels faster than non-burrowers (ANOVA, days:burrow, Χ2 = 4.6648, df = 1, p-value = 0.03079).
Figure 2.
Keel prominence values vs time in captivity for burrowing and non-burrowing individuals. Individual shapes connected by thin lines show keel prominence for individual octopuses during captivity, with blue showing burrowing individuals, and red showing non-burrowers. Thick lines show results of LME. Results show a significant interaction between burrowing and day, with non-burrowing octopuses losing their keels faster than non-burrowers (ANOVA, days:burrow, Χ2 = 4.6648, df = 1, p-value = 0.03079).
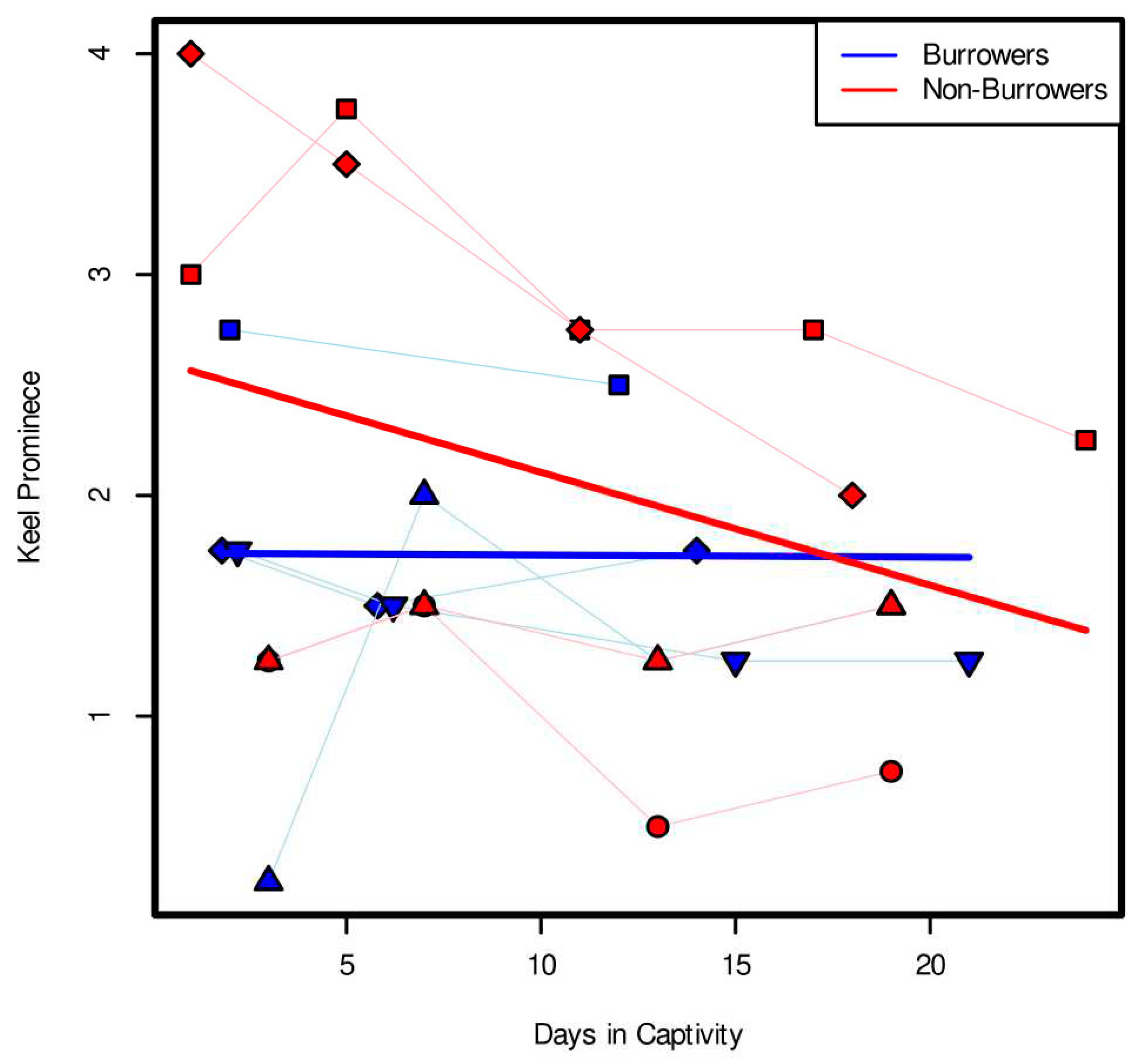
Figure 3.
Multi-species phylogenetic tree showing results of Pagel’s binary character correlation test between keel presence and known burrowing/burying behavior. ARD substitution model used between dependent and independent model rate matrices of 4 possibilities: burrowing, non-burrowing, keel, no keel. Results show interdependence to be the model of best fit (AIC=188.4336, likelihood-ratio = 20.5514, p-value = 0.000388571).
Figure 3.
Multi-species phylogenetic tree showing results of Pagel’s binary character correlation test between keel presence and known burrowing/burying behavior. ARD substitution model used between dependent and independent model rate matrices of 4 possibilities: burrowing, non-burrowing, keel, no keel. Results show interdependence to be the model of best fit (AIC=188.4336, likelihood-ratio = 20.5514, p-value = 0.000388571).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



