
Submitted:
29 June 2023
Posted:
03 July 2023
You are already at the latest version
Abstract
Type-2 diabetes mellitus is recognized as a serious public health concern with a considerable impact on human life, long-term health expenditures, and substantial health losses.
In this context, the use of dietary polyphenols to prevent and manage Type 2 diabetes mellitus is widely documented. These dietary compounds exert their beneficial effects by several actions, including the protection of pancreatic islet β-cell, the antioxidant capacities of these molecules, their effects on insulin secretion and actions, the regulation of intestinal microbiota, and their contribution to ameliorating diabetic complications, particularly those of vascular origin.
In the present review, we intend to highlight these multifaceted actions and the molecular mechanisms by which these plant-derived secondary metabolites exert their beneficial effects on type-2 diabetes patients.
Keywords:
type-2 diabetes mellitus
; polyphenols
; oxidative stress
; antioxidant capacity
1. Introduction
Diabetes mellitus is a heterogeneous group of chronic metabolic disorders characterized by hyperglycemia resulting from defects of insulin action, insulin secretion, or both [1].
This metabolic disease is a global health issue, which has been increasing from time to time and it is now considered as one of the most important disorders worldwide. According to International Diabetes Federation, 10.5% of adults of the world population are currently living with diabetes and this alarming indicator is predicted to rise to 11.3 %, (643 million people) by 2030 and to 12,2 % (783 million) by 2045 [2].
Noteworthy, a considerable proportion of the world's burden of diabetes is caused by type 2 diabetes mellitus (T2DM). In this regard, T2DM is recognized as a serious public health concern with a considerable impact on human life and health expenditures [3].
The onset and progression of T2DM are determined by a complex pathophysiological basis where oxidative stress is a crucial contributor not only involved in the disease development but also to diabetes complications, particularly those associated with both microvascular (retinopathy, nephropathy, and neuropathy) and macrovascular complications (ischemic heart disease, peripheral vascular disease, and cerebrovascular disease [4].
Acute or chronic hyperglycemia upregulates ROS production in the mitochondrial electron transfer chain. This excessive production of superoxide an inorganic radical anion mediates the downregulation of GADPH levels, which in turn activates the major pro-oxidative pathways involved in the pathogenesis of diabetes complications, such as the activation of protein kinase C, the polyol and hexosamine pathways, the formation of advanced glycation end products production (AGEs), as well as the increased expression of the receptor for AGEs [5,6,7].
On the other hand, antioxidant mechanisms are diminished in diabetic patients, which may further augment oxidative stress [8,9,10].
During the last few years, compelling shreds of evidence have shed light on the usefulness of dietary antioxidants as an alternative option in the treatment of T2DM, considering both the adverse effects conferred by conventional pharmacological treatments as well as the enormous economic burden that lifelong treatments place on patients [11].
In this regard, dietary polyphenols have emerged as an option to manage T2DM. These compounds are one of the most abundant secondary plant metabolites, which are grouped into four major families, flavonoids, ligands, stilbenes, and phenolic acids, and are widely found in fruits, vegetables, nuts, cereals, and in many beverages such as tea, coffee, and red wines. A growing body of experimental and clinical evidence supports the protective role of these compounds on several human diseases through their antioxidant activity and diverse molecular mechanisms [12,13,14,15,16].
This review is aimed to highlight the roles of this large and heterogeneous family of secondary metabolites of plants containing phenol rings, on pancreatic islet β-cell functioning and promotion of insulin production and signaling, protection against micro-and microvascular complications, protection against the progression of T2DM-associated obesity, management of dyslipidemia and gut microbiome dysbiosis. In addition, the capacity of polyphenols to reduce both the formation of advanced glycation products and their pathologic consequences is also addressed.
2. β-cell dysfunction and death
Currently, both clinical and experimental data support that during the development of T2DM, there is not only a progressive deterioration in β-cell functioning but also a marked reduction of the β-cell mass in the pancreatic islets of Langerhans [17,18,19].
Many factors such as the glucotoxicity associated with the hyperglycemic state, the oxidative and endoplasmic reticulum stresses, as well as the lipotoxicity due to chronic exposure to saturated free fatty acids (FFA), are crucial elements in decreased beta-cell functioning and, eventually in β cell death through apoptosis [17,20,21].
Hyperglycemia is a crucial factor in the onset of oxidative stress in T2DM and it even correlates with the progression of disease [22]. Additionally, β-cells are very susceptible to oxidative damage, because of their low antioxidant capacity [23,24], and consequently, oxidative stress is a very important contributor to the impairment of beta-cell functioning [21,25,26]. Furthermore, oxidative stress mediates the permeabilization of mitochondrial membranes, and consequently the release of cytochrome C and thus β-cell death by apoptosis [27].
Based on their antioxidant activities polyphenols are major regulators of oxidative stress and consequently the improvement of mitochondrial functions. Cocoa catechins can also improve insulin secretion by increasing the expression of some genes involved in mitochondrial respiration [28].
Resveratrol is known for its remarkable activities in improving pancreatic β-cell function mainly by its effect on Sirtuin-1, a master regulator for β-cell function [29]. Cinnamic acid derivatives can improve the insulin-secreting capacity of beta-cells, by raising the levels of intracellular calcium [30].
Noteworthy, compelling pieces of evidence support that the hyperglycemia-associated overexpression of human amylin, also known as islet amyloid polypeptide (IAPP), can form aggregates to favor amylin fibril formation, and these fibrils evoke the activation of caspases cascade and thus leading to β-cell death by apoptosis [31,32].
3. Insulin resistance
Insulin receptor (IR) is a tyrosine kinase receptor, which is autophosphorylated upon insulin binding and it is expressed in all tissues. The major responders to IR engagement by insulin are the liver, skeletal muscle, and adipose tissue [35].
Upon insulin binding complex signaling is activated including several substrates such as insulin or insulin-like growth factor (IGF)-1, insulin receptor (IR), insulin receptor substrate (IRS)-1, and phosphatidylinositol-3 kinase (PI3-K)/Akt or ERK kinases. The phosphorylation of IRS1 can recruit PI3K rendering Akt phosphorylated, which in turn can regulate crucial events such as the translocation of glucose transporter GLUT4 to the cell surface, promoting glycogen synthesis through inhibition of GSK3 activity, the induction of protein synthesis via activation of mTOR and the inhibition of FoxO transcription factors [36,37].
The inactivation of Akt and activation of FoxO1, through the suppression of IRS1 and IRS2 in different organs following hyperinsulinemia, over-nutrition, and inflammation, represent crucial mechanisms for insulin resistance in humans [38,39].
Compelling shreds of evidence support that oxidative stress is an important contributor to insulin resistance in T2DM [40], and that the production of mitochondrial H2O2 [41,42], and the overactivation of NAPDPH oxidase, via angiotensin II/AT1 receptor can mediate skeletal muscle insulin resistance [43]. Furthermore, oxidative stress has been shown to impair insulin signaling through the reduction of the activity of tyrosine phosphatase which in turn, raises the basal level of tyrosine phosphorylation in both the insulin receptor and downstream proteins involved in the signaling cascade [44,45].
The cJun-N-terminal kinases (JNKs) is major signal transducer driving the physiological response to several cellular stressors, including oxidative stress. Epigallocatechin gallate, the major green tea catechin can protect both the insulin receptor and IRS proteins from phosphorylation by JNKs, a crucial event in the onset of insulin resistance [46], as well as by reducing the expression of the negative regulator of insulin receptor PTP1B [47].
Resveratrol, which is one of the main polyphenolic compounds of red wines, peanuts and apples, is a potent activator of sirtuin 1 (SIRT1), which is a potent intracellular inhibitor of oxidative stress, and thus attenuates insulin resistance and improves insulin signaling in the skeletal muscle cells [48,49].
4. Obesity
Obesity is the major driving factor of T2DM and it is characterized by chronic low-grade inflammation with permanently increased oxidative stress [50,51]. The onset of a chronic condition of oxidative stress in obesity is supported by different mechanisms implicated in the homeostasis of adipose tissue, which contributes to the development of pathological systemic consequences [52]. On one hand, those associated with increased reactive oxygen species (ROS) production such as the adipocytes-associated endoplasmic reticulum stress, a sustained increase of NOx activities, as well as the high level of post-prandial-associated ROS generation, and on the other, the altered antioxidant defenses observed in obese patients [53,54,55,56].
In addition to the antioxidant properties of polyphenols, they exert several beneficial effects on obesity far beyond their antioxidant capacity [57], such as the attenuation of obesity-linked inflammation [58], the beneficial regulation of several key obesity pathways such as the modulation of food intake [59], the inhibition of pancreatic lipase [60]; decreasing lipogenesis by inhibiting both fatty acid synthase activity and the activation of the adenosine monophosphate (AMP)-activated protein kinase (AMPK) [61,62], and by increasing thermogenesis and mitochondrial biogenesis [63].
Finally, some polyphenols have been reported to mediate the suppression of the conversion of preadipocytes into adipocytes, which can store an excessive lipid load. This polyphenols-mediated suppression of adipocytes differentiation occurs by the regulation of crucial factors such as C/EBPα, PPARγ1, PPARγ2, and GLUT-4 in mature adipocytes [61,64,65].
5. Dysbiosis
Human gut microbiota is considered a complex microbial ecosystem composed of different microorganisms, including bacteria, archaea, viruses, fungi, and protists, which are involved in the regulation of many physiological processes and numerous diseases [66].
Firmicutes and Bacteroidetes are the main phyla that composed the adult gut flora, regulating the homeostatic production of microbiota-induced metabolites such as butyrate, which have anti-inflammatory and antioxidative properties, and the production of lipopolysaccharide (LPS), which can promote systemic inflammation and insulin resistance through induction of metabolic endotoxemia [67,68].
Growing data raised from both clinical and experimental evidence shows that T2DM patients have an altered gut microbiota, where the Bacterioidetes/Firmicutes ratio of the intestinal flora of diabetic patients significantly differs from non-T2DM adults [69,70].
A crucial consequence of the quantitative change in gut microbiota composition in T2DM patients is the impairment of the expression of gut-microbiota-related metabolites, which have crucial consequences in the metabolic regulation of glucose homeostasis, and insulin sensitivity [70].
Short-chain fatty acids (SCFAs) are considered one of the main microbial metabolites, that have crucial effects on the expression of glucagon-like peptide-1 (GLP-1) and GLP-2 via stimulating G-protein-coupled receptors (GPR), thus contributing to improve glucose homeostasis and amplification of insulin sensitivity [71].
Under this dysbiosis condition that affects T2DM patients, structural changes in the intestinal epithelium barrier allow LPS translocation into the bloodstream, resulting in increased plasmatic levels of LPS, which in consequence, activates Toll-like receptor-4 (TLR4) leading to the production of pro-inflammatory mediators, and sustaining low-grade systemic inflammation [72].
This condition known as metabolic endotoxemia induces a significant decrease in bacterial populations which are crucial producers of beneficial gut-derived metabolites such as SCFA, thus supporting the impairment of glucose metabolism and insulin resistance [73,74].
In addition, different studies have demonstrated that specific gut microbiota dysbiosis in mice model of T2DM, induces GLP-1 resistance and consequently, the impairment of GLP1-induced insulin secretion which is crucial in the acquisition of the insulin resistance condition in diabetic individuals [75].
At present, polyphenols have emerged as novel compounds that could interact with microbiota and exert strong regulatory effects on intestinal bacteria, with subsequent regulation of gut microbiota and its derivate metabolites [76]. These interactions between polyphenols and gut microbiota can positively affect crucial metabolic markers of T2DM, improving systemic inflammation and insulin sensitivity [77,78].
Growing evidence reveals that distinct types of polyphenolic compounds, such as genistein, curcumin, and grifolic acid can increase GLP-1 secretion from L-cells via different mechanisms [79,80,81,82]. Besides their effect to directly stimulate GLP-1 secretion, some polyphenols, particularly luteolin, apigenin, and resveratrol may also naturally suppress DPP-IV activity, which potentially increases the half-life of GLP-1, thus stimulating glucose-dependent insulin secretion and regulating glycemia [83].
Different studies demonstrate that different doses of oral intake of polyphenols including catechins, and (-)-epigallocatechin-3-gallate (EGCG), can also favor the increase of different microbial populations of SCFA-producing agents in fecal samples of human patients, thus improving the insulin sensitivity and glucose homeostasis of individuals [84,85].
In addition, other phenolic compounds including chlorogenic and ferulic acid can also act as antidiabetic agents, through significant upregulating of the expression of GLUT4 and PPAR-γ, thus favoring the uptake of 2-deoxyglucose (2DG) in time- and dose-dependent manner, and improving the pathogenesis of T2DM progression [86,87,88].
Another feature of diabetogenic and overfeeding individuals is the increased expression of branched-chain amino acids (BCAA) and BCAA-related metabolites such as the branched-chain keto acids (BCKA) from the gut microbiome [89], which are well established as among the strongest biomarkers of obesity, and insulin resistance, in consequence, potentiating the development of T2DM [90,91]. Conversely, experimental results have demonstrated that lowering BCAA and BCKA levels is associated with improved insulin sensitivity and reduced fat accumulation in mouse models [92].
Emerging studies have suggested that polyphenol administration may accelerate the catabolism of BCAA, inducing a lowering of circulating BCAA levels, thus improving glucose homeostasis and insulin sensitivity [93].
Additionally, some evidence also supports that intestinal catabolites of polyphenolic compounds by the action of the gut microbiota could act as a strong antiglycative agent [94,95]. In this sense, dietary polyphenolic intake may have a significant positive impact on the generation of glycation products and diabetes-related complications [96,97].
Taken together, those findings suggest that a polyphenols-enriched diet can strongly modulate the dysbiotic changes induced by hyperglycemia, improving the regulation of metabolites that mediates glucose homeostasis and insulin sensitivity in T2DM patients.
6. Vascular complications
Vascular complications in T2DM are those long-term complications that affect the blood vessels network, and are responsible for most of the morbidity, and required hospitalization in these patients. The vascular complications of diabetes are classified as either microvascular (retinopathy, nephropathy, and neuropathy) or macrovascular, which includes coronary artery, peripheral, and cerebral vascular diseases [98,99,100]. At present, a large body of compelling pieces of evidence support that oxidative stress has a key role in the pathogenesis of vascular complications in diabetes [101,102,103].
As a major regulator of vascular homeostasis, the vascular endothelial cells play crucial roles by controlling vascular tone through a balance between vasodilation and vasoconstriction, fibrinolysis, platelet adhesion and aggregation, leukocyte activation, adhesion, and transmigration, smooth muscle cell proliferation, and modulating the growth of blood vessels [104,105].
The onset of an imbalanced vasodilation and vasoconstriction, elevated reactive oxygen species (ROS), and proinflammatory factors, as well as a reduced nitric oxide (NO) bioavailability, are crucial elements in the onset of the systemic disorder known as endothelial dysfunction [106]. NO is produced in the endothelium by the endothelial nitric oxide synthase, a Ca2+-calmodulin-dependent enzyme that can convert the L-arginine to NO plus citrulline. By activation of soluble guanylyl cyclase and modulation of cation channels, nitric oxide promotes VSMC relaxation and thus regulates vascular tone. Additionally, nitric oxide is a crucial mediator in controlling platelet activation and aggregation [107].
When ROS bioavailability overtakes the antioxidant defenses due to the onset of oxidative stress, superoxide (O2−) rapidly inactivates NO and forms peroxynitrite (ONOO−), It is known that peroxynitrite inactivates prostacyclin synthase (PGIS) and thus favoring the deterioration of vascular health due to the vasodilatory, growth-inhibiting, antithrombotic, and antiadhesive effects of prostacyclin (PGI2). Additionally, peroxynitrite increases the release of prostaglandin H2 (PGH2) and thromboxane A2, which are potent vasoconstrictors, prothrombotic, growth- and adhesion-promoting agents [108,109,110].
A growing body of data supports the beneficial roles of polyphenols to protect against endothelial dysfunction induced by oxidative stimuli [111,112,113]. Of note, some polyphenols, as reported for resveratrol and its derivatives show dual protecting activities, either by the expression of Nox4, a ROS-generating enzyme highly expressed in the endothelium, and by enhancing the expression of two crucial members of the antioxidant defense of the vascular wall, such as glutathione peroxidase 1 (GPx1) and superoxide dismutase 1 (SOD1) [114]. Moreover, polyphenols seem to have peroxynitrite-scavenging activity [115].
Furthermore, different reports have demonstrated that some polyphenols such as resveratrol and others derived from strawberry and grapes skin and seeds, can promote the phosphorylation of eNOS at Ser1177 by PI3K/Akt pathway, which is essential for NO production [116,117,118]. In addition, resveratrol is reported to increase both endothelial eNOS mRNA and protein levels [119,120,121]. This effect seems to be associated with the effects of resveratrol on SIRT1 and FOXO factors [122].
7. Polyphenols and advanced glycation.
Advanced glycation is one of the major pathways involved in the onset and progression of T2DM complications, particularly those associated with the cardiovascular system [123]. Since the pioneering works of the Vlassara group [124,125], a huge and compelling body of evidence has demonstrated the paramount importance of advanced glycation end-products in diabetes complications, due to the hyperglycemic condition [126,127].
The formation of advanced glycation end-products (AGEs) involves the reaction of reducing sugars, such as glucose, with the terminal amino groups of proteins, nucleic acids, or phospholipids to initially form unstable Schiff bases, which evolve towards the formation of more stable compounds called Amadori products, which by a series of complex reaction yield the AGEs. Degradation of both Schiff bases and Amadori products rise to highly reactive short-chain carbonyl compounds, called α-dicarbonyls [128].
These highly reactive compounds can also be formed by hexose autoxidation, as well as by-products of either the glycolytic or polyol pathways and from lipid oxidation. Dicarbolyls can then react non-enzymatically with lysine or arginine residues to produce AGEs [129,130].
The AGEs exert their deleterious effects, either directly by cross-linking of proteins, thus disrupting protein functioning and turn-over [131,132], or indirectly by binding to a signaling receptor for AGE-modified proteins, known as the receptor of advanced glycation end-products (RAGE) [133,134].
Noteworthy, oxidative stress is an important contributor to the formation of endogenous AGEs, by leading to the increased formation of endogenous reactive aldehydes as glyoxal, methylglyoxal (MG), and thus favoring the formation of AGEs [135]. Additionally, when AGEs activate RAGE, NADPH oxidase is activated and thus increases ROS levels [136].
At present, compelling evidence derived from experimental and clinical data studies support the role of different polyphenols as very active inhibitors of the deleterious effects of AGEs, through several mechanisms [137,139].
By their antioxidant activities, polyphenols are potent antiglycation compounds and antiglycation activity strongly correlates with the free radical scavenging activity and antiglycation activity [94], as reported catechins, proanthocyanidins, anthocyanin, stilbenoids, and flavonols [139,140]. Additionally, polyphenols have other properties which are essential to reduce the formation of AGEs, such as the chelation of transition metal, as reported for chlorogenic and caffeic acids [141,142].
The capacity of trapping dicarbonyl compounds is another crucial activity reported for some polyphenols considering that dicarbonyls are one of the main precursors of AGEs [129], epigallocatechin-3-gallate, resveratrol, catechin, and epicatechin as well as different procyanidins can efficiently trap both glyoxal and methylglyoxal [137,143,144].
8. Lipid metabolism
T2DM has been widely associated with an increased risk for atherosclerotic cardiovascular disease which is closely related to raised plasmatic LDL levels with important oxidative changes, [150] which support diabetic hyperlipidemia and accelerated atherosclerosis, increasing the risk of macrovascular complication and cardiovascular morbidity. Noteworthy, LDL is a highly sensitive molecule to hyperglycemia-induced hyperglycemia damage and modification, making them highly pathogenic and atherogenic [151,152].
Under hyperglycemic conditions, transition metals in the presence of oxygen catalyze the autoxidation of glucose or lipid peroxidation [153].
In addition, excess ROS formation in T2DM patients fuels vascular inflammation, and mediates oxidized low-density lipoprotein (OxLDL) formation, which is considered a hallmark feature of atherosclerotic development due to the crucial induction of atherosclerotic plaque progression and destabilization in T2DM patients [154,155,156].
Besides the different pathways that conflux in activate NADPH oxidase and subsequent ROS production in T2DM patients, the increased expression of ox-LDL also stimulates NADPH oxidase, thus contributing to increment ROS formation and oxidative stress in T2DM patients [157].
In addition, hyperglycemia-mediated mitochondrial ROS production can also promote the NF-kB-mediated entry of monocytes in atherosclerotic lesions, fueling the inflammation and progression of unstable plaques, and increasing the risk of macrovascular complication in T2DM patients [158], thus, sustaining a vicious cycle that perpetuating ROS production and oxLDL formation, contributes to the progression of atherosclerosis unstable plaques on DM patients.
In last years, polyphenols have been postulated to lower lipids through different mechanisms that imply beneficial effects on cardiovascular diseases of T2DM patients [159]. Based on their antioxidant effects, different studies have shown that many polyphenols including resveratrol, apigenin as well as synthetic polyphenol-like molecules can inhibit NAPDH oxidase activity, thus decreasing vascular oxidation and atherogenesis in nondiabetic apolipoprotein (apo) E–deficient mice [160], as well as improve hyperlipidemia and atherosclerosis in diabetic individuals [161].
Resveratrol based on its antioxidant activities can impact lipid metabolism and is considered an important protective compound against LDL oxidation and atherosclerosis progression [162,163]. In this sense, the free radical scavenging activity of resveratrol has been investigated, revealing that this polyphenol compound can interact with free radicals to form relatively stable free radicals and non-radicals, resulting in inhibition of lipid peroxidation by Fenton reaction products [162,164], which may decrease the progression of accelerated atherosclerosis through inhibition of oxidation in T2DM patients [165,166].
More recently, it was demonstrated that resveratrol can upregulate endothelial nitric oxide synthase (eNOS) expression by increasing cAMP levels, and decreasing ox-LDL-induced oxidative stress in human endothelial cells, leading to a significant improvement of endothelial dysfunction and atherosclerosis in mice [167]. Similar results have been demonstrated for quercetin, an important flavonoid, which have been demonstrate protective effects in diabetic individuals through significantly reversed dyslipidemia and hepatic steatosis in diabetic mice, including lowered liver cholesterol and triglycerides contents [168,169].
Taken together, these findings suggest that dietary polyphenols may be crucial in the regulation of dysregulated lipid metabolism through the modulation of antioxidative mechanisms in T2DM patients.
9. Conclusion remarks and future directions
A compelling body of evidence suggests that dietary polyphenols may represent an important alternative to the management of DM as well as in many of its complications (See Figure 1). Most of the pieces of evidence derived from animals and in vitro studies support these issues, and only for some polyphenols, solid clinical shreds of evidence have been provided. However, much more research is needed on some topics that may be crucial to explain the current controversial results in some clinical studies. In this regard, a full understanding of the metabolisms and bioavailability, the assessment of dietary intake by measuring urine or blood polyphenol metabolites, duration of exposure, delivery systems that guarantee high stability, as well as more efforts to understand the structure-activity relationship of polyphenols, are crucial elements to be considered in the design and execution of more double-blinded clinical trials.
References
- Tomic, D.; Shaw, J.E.; Magliano, D.J. The burden and risks of emerging complications of diabetes mellitus. Nat. Rev. Endocrinol. 2022, 18, 525–539. [Google Scholar] [CrossRef]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF diabetes atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Hashim, M.J.; King, J.K.; Govender, R.D.; Mustafa, H.; Al Kaabi, J. Epidemiology of type 2 diabetes – global burden of disease and forecasted trends. J. Epidemiol. Glob. Health. 2019, 10, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of type 2 diabetes mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Yan, LJ. Pathogenesis of chronic hyperglycemia: from reductive stress to oxidative stress. J. Diabetes Res. 2014, 2014, 137919. [Google Scholar] [CrossRef] [PubMed]
- Black, H.S. A synopsis of the associations of oxidative stress, ROS, and antioxidants with diabetes mellitus. Antioxidants 2022, 11, 2003. [Google Scholar] [CrossRef] [PubMed]
- Rains, J.L.; Jain, S.K. Oxidative stress, insulin signaling, and diabetes. Free Radic. Biol. Med. 2011, 50, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B. Diabetes, oxidative stress, and antioxidants: A Review. J. Biochem. Mol. Toxicol. 2003, 17, 24–38. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Pasupuleti, V.R.; Arigela, C.S.; Gan, S.H.; Salam, S.K.N.; Krishnan, K.T.; Rahman, N.A.; Jeffree, M.S. A Review on Oxidative Stress, Diabetic Complications, and the Roles of Honey Polyphenols. Oxid. Med. Cell. Longev. 2020, 2020, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Andrés-Lacueva, C. Polyphenols and Health: Current State and Progress. J. Agric. Food Chem. 2012, 60, 8773–8775. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights Into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef]
- Cerf, M.E. Beta Cell Dysfunction and Insulin Resistance. Front. Endocrinol. 2013, 4. [Google Scholar] [CrossRef]
- White, M.G.; Shaw, J.A.M.; Taylor, R. Type 2 Diabetes: The Pathologic Basis of Reversible β-Cell Dysfunction. Diabetes Care 2016, 39, 2080–2088. [Google Scholar] [CrossRef]
- Ferrannini, E.; Mari, A. β-Cell Function in Type 2 Diabetes. Metabolism 2014, 63, 1217–1227. [Google Scholar] [CrossRef]
- Porte, D.; Kahn, S.E. Beta-Cell Dysfunction and Failure in Type 2 Diabetes: Potential Mechanisms. Diabetes 2001, 50, S160. [Google Scholar] [CrossRef]
- Cerf, M.E. Beta Cell Physiological Dynamics and Dysfunctional Transitions in Response to Islet Inflammation in Obesity and Diabetes. Metabolites 2020, 10, 452. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, J.S.; Sehrawat, A.; Mishra, J.; Sidhu, I.S.; Navik, U.; Khullar, N.; Kumar, S.; Bhatti, G.K.; Reddy, P.H. Oxidative Stress in the Pathophysiology of Type 2 Diabetes and Related Complications: Current Therapeutics Strategies and Future Perspectives. Free Radic. Biol. Med. 2022, 184, 114–134. [Google Scholar] [CrossRef] [PubMed]
- Drews, G.; Krippeit-Drews, P.; Düfer, M. Oxidative Stress and Beta-Cell Dysfunction. Pflugers Arch. 2010, 460, 703–718. [Google Scholar] [CrossRef]
- Dludla, P.V.; Mabhida, S.E.; Ziqubu, K.; Nkambule, B.B.; Mazibuko-Mbeje, S.E.; Hanser, S.; Basson, A.K.; Pheiffer, C.; Kengne, A.P. Pancreatic β-Cell Dysfunction in Type 2 Diabetes: Implications of Inflammation and Oxidative Stress. World J. Diabetes 2023, 14, 130–146. [Google Scholar] [CrossRef]
- Dinić, S.; Arambašić Jovanović, J.; Uskoković, A.; Mihailović, M.; Grdović, N.; Tolić, A.; Rajić, J.; Đorđević, M.; Vidaković, M. Oxidative Stress-Mediated Beta Cell Death and Dysfunction as a Target for Diabetes Management. Front. Endocrinol. 2022, 13, 1006376. [Google Scholar] [CrossRef]
- Eguchi, N.; Vaziri, N.D.; Dafoe, D.C.; Ichii, H. The Role of Oxidative Stress in Pancreatic β Cell Dysfunction in Diabetes. Int. J. Mol. Sci. 2021, 22, 1509. [Google Scholar] [CrossRef]
- Ma, Z.A. The Role of Peroxidation of Mitochondrial Membrane Phospholipids in Pancreatic β -Cell Failure. Curr. Diabetes Rev. 2012, 8, 69–75. [Google Scholar] [CrossRef]
- Rowley, T.J.; Bitner, B.F.; Ray, J.D.; Lathen, D.R.; Smithson, A.T.; Dallon, B.W.; Plowman, C.J.; Bikman, B.T.; Hansen, J.M.; Dorenkott, M.R.; et al. Monomeric Cocoa Catechins Enhance β-Cell Function by Increasing Mitochondrial Respiration. J. Nutr. Biochem. 2017, 49, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Vetterli, L.; Brun, T.; Giovannoni, L.; Bosco, D.; Maechler, P. Resveratrol Potentiates Glucose-Stimulated Insulin Secretion in INS-1E β-Cells and Human Islets through a SIRT1-Dependent Mechanism. J. Biol. Chem. 2011, 286, 6049–6060. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Moonsan, P.; Yibchok-anun, S. Insulin-Releasing Properties of a Series of Cinnamic Acid Derivatives in Vitro and in Vivo. J. Agric. Food Chem. 2008, 56, 7838–7844. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, J.; Dragunow, M.; Cooper, G.J.S. Fibrillogenic Amylin Evokes Islet β-Cell Apoptosis through Linked Activation of a Caspase Cascade and JNK1. J. Biol. Chem. 2003, 278, 52810–52819. [Google Scholar] [CrossRef] [PubMed]
- Kanatsuka, A.; Kou, S.; Makino, H. IAPP/Amylin and β-Cell Failure: Implication of the Risk Factors of Type 2 Diabetes. Diabetol. Int. 2018, 9, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, I.; Poppitt, S. Unfolding Novel Mechanisms of Polyphenol Flavonoids for Better Glycaemic Control: Targeting Pancreatic Islet Amyloid Polypeptide (IAPP). Nutrients 2017, 9, 788. [Google Scholar] [CrossRef] [PubMed]
- Mahboob, A.; Senevirathne, D.K.L.; Paul, P.; Nabi, F.; Khan, R.H.; Chaari, A. An Investigation into the Potential Action of Polyphenols against Human Islet Amyloid Polypeptide Aggregation in Type 2 Diabetes. Int. J. Biol. Macromol. 2023, 225, 318–350. [Google Scholar] [CrossRef]
- Boucher, J.; Kleinridders, A.; Kahn, C.R. Insulin Receptor Signaling in Normal and Insulin-Resistant States. Cold Spring Harb. perspect. biol. 2014, 6, a009191. [Google Scholar] [CrossRef]
- Saltiel, A.R. Insulin Signaling in Health and Disease. J. Clin. Investig. 2021, 131, e142241. [Google Scholar] [CrossRef]
- Siddle, K. Signalling by Insulin and IGF Receptors: Supporting Acts and New Players. J. Mol. Endocrinol. 2011, 47, R1–R10. [Google Scholar] [CrossRef]
- Taylor, R. Insulin Resistance and Type 2 Diabetes. Diabetes 2012, 61, 778–779. [Google Scholar] [CrossRef]
- Wondmkun, Y.T. Obesity, Insulin Resistance, and Type 2 Diabetes: Associations and Therapeutic Implications. Diabetes Metab. Syndr. Obes. 2020, 13, 3611–3616. [Google Scholar] [CrossRef]
- Hurrle, S.; Hsu, W.H. The Etiology of Oxidative Stress in Insulin Resistance. Biomed. J. 2017, 40, 257–262. [Google Scholar] [CrossRef]
- Anderson, E.J.; Lustig, M.E.; Boyle, K.E.; Woodlief, T.L.; Kane, D.A.; Lin, C.-T.; Price, J.W.; Kang, L.; Rabinovitch, P.S.; Szeto, H.H.; et al. Mitochondrial H2O2 Emission and Cellular Redox State Link Excess Fat Intake to Insulin Resistance in Both Rodents and Humans. J. Clin. Invest. 2009, 119, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, E.J.; Diamond-Stanic, M.K.; Marchionne, E.M. Oxidative Stress and the Etiology of Insulin Resistance and Type 2 Diabetes. Free Radic. Biol. Med. 2011, 51, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Sowers, J.R.; Nistala, R.; Gong, H.; Uptergrove, G.M.-E.; Clark, S.E.; Morris, E.M.; Szary, N.; Manrique, C.; Stump, C.S. Angiotensin II-Induced NADPH Oxidase Activation Impairs Insulin Signaling in Skeletal Muscle Cells. J. Biol. Chem. 2006, 281, 35137–35146. [Google Scholar] [CrossRef]
- Petersen, M.C.; Shulman, G.I. Mechanisms of Insulin Action and Insulin Resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef]
- Masenga, S.K.; Kabwe, L.S.; Chakulya, M.; Kirabo, A. Mechanisms of Oxidative Stress in Metabolic Syndrome. Int. J. Mol. Sci. 2023, 24, 7898. [Google Scholar] [CrossRef] [PubMed]
- Solinas, G.; Becattini, B. JNK at the Crossroad of Obesity, Insulin Resistance, and Cell Stress Response. Mol. Metab. 2017, 6, 174–184. [Google Scholar] [CrossRef]
- Mi, Y.; Zhang, W.; Tian, H.; Li, R.; Huang, S.; Li, X.; Qi, G.; Liu, X. EGCG Evokes Nrf2 Nuclear Translocation and Dampens PTP1B Expression to Ameliorate Metabolic Misalignment under Insulin Resistance Condition. Food Funct. 2018, 9, 1510–1523. [Google Scholar] [CrossRef]
- Vlavcheski, F.; Den Hartogh, D.J.; Giacca, A.; Tsiani, E. Amelioration of High-Insulin-Induced Skeletal Muscle Cell Insulin Resistance by Resveratrol Is Linked to Activation of AMPK and Restoration of GLUT4 Translocation. Nutrients 2020, 12, 914. [Google Scholar] [CrossRef]
- Zin, C.A.J.C.M.; Mohamed, W.M.I.W.; Khan, N.A.K.; Ishak, W.R.W. Effects of Fruit and Vegetable Polyphenols on the Glycemic Control and Metabolic Parameters in Type 2 Diabetes Mellitus: A Review. Prev. Nutr. Food Sci. 2022, 27, 257–264. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased Oxidative Stress in Obesity and Its Impact on Metabolic Syndrome. J. Clin. Invest. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Keaney, J.F.; Larson, M.G.; Vasan, R.S.; Wilson, P.W.F.; Lipinska, I.; Corey, D.; Massaro, J.M.; Sutherland, P.; Vita, J.A.; Benjamin, E.J. Obesity and Systemic Oxidative Stress: Clinical Correlates of Oxidative Stress in The Framingham Study. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Skalicky, J.; Muzakova, V.; Kandar, R.; Meloun, M.; Rousar, T.; Palicka, V. Evaluation of Oxidative Stress and Inflammation in Obese Adults with Metabolic Syndrome. Clin. Chem. Lab. Med. 2008, 46. [Google Scholar] [CrossRef]
- Chrysohoou, C.; Panagiotakos, D.B.; Pitsavos, C.; Skoumas, I.; Papademetriou, L.; Economou, M.; Stefanadis, C. The Implication of Obesity on Total Antioxidant Capacity in Apparently Healthy Men and Women: The ATTICA Study. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Savini, I.; Catani, M.; Evangelista, D.; Gasperi, V.; Avigliano, L. Obesity-Associated Oxidative Stress: Strategies Finalized to Improve Redox State. Int. J. Mol. Sci. 2013, 14, 10497–10538. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative Stress in Obesity: A Critical Component in Human Diseases. Int. J. Mol. Sci. 2014, 16, 378–400. [Google Scholar] [CrossRef]
- Aloo, S.-O.; Ofosu, F.K.; Kim, N.-H.; Kilonzi, S.M.; Oh, D.-H. Insights on Dietary Polyphenols as Agents against Metabolic Disorders: Obesity as a Target Disease. Antioxidants 2023, 12, 416. [Google Scholar] [CrossRef]
- Zamani-Garmsiri, F.; Emamgholipour, S.; Rahmani Fard, S.; Ghasempour, G.; Jahangard Ahvazi, R.; Meshkani, R. Polyphenols: Potential Anti-inflammatory Agents for Treatment of Metabolic Disorders. Phytother. Res. 2022, 36, 415–432. [Google Scholar] [CrossRef]
- Badshah, H.; Ullah, I.; Kim, S.E.; Kim, T.; Lee, H.Y.; Kim, M.O. Anthocyanins Attenuate Body Weight Gain via Modulating Neuropeptide Y and GABAB1 Receptor in Rats Hypothalamus. Neuropeptides 2013, 47, 347–353. [Google Scholar] [CrossRef]
- Buchholz, T.; Melzig, M.F. Polyphenolic Compounds as Pancreatic Lipase Inhibitors. Planta Med. 2015, 81, 771–783. [Google Scholar] [CrossRef]
- Kim, N.-H.; Jegal, J.; Kim, Y.N.; Heo, J.-D.; Rho, J.-R.; Yang, M.H.; Jeong, E.J. Chokeberry Extract and Its Active Polyphenols Suppress Adipogenesis in 3T3-L1 Adipocytes and Modulates Fat Accumulation and Insulin Resistance in Diet-Induced Obese Mice. Nutrients 2018, 10, 1734. [Google Scholar] [CrossRef]
- Rocha, A.; Bolin, A.P.; Cardoso, C.A.L.; Otton, R. Green Tea Extract Activates AMPK and Ameliorates White Adipose Tissue Metabolic Dysfunction Induced by Obesity. Eur. J. Nutr. 2016, 55, 2231–2244. [Google Scholar] [CrossRef]
- Lee, M.-S.; Shin, Y.; Jung, S.; Kim, Y. Effects of Epigallocatechin-3-Gallate on Thermogenesis and Mitochondrial Biogenesis in Brown Adipose Tissues of Diet-Induced Obese Mice. Food Nutr. Res. 2017, 61, 1325307. [Google Scholar] [CrossRef]
- Valli, V.; Heilmann, K.; Danesi, F.; Bordoni, A.; Gerhäuser, C. Modulation of Adipocyte Differentiation and Proadipogenic Gene Expression by Sulforaphane, Genistein, and Docosahexaenoic Acid as a First Step to Counteract Obesity. Oxid. Med. Cell. Longev. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Carpéné, C.; Pejenaute, H.; Del Moral, R.; Boulet, N.; Hijona, E.; Andrade, F.; Villanueva-Millán, M.J.; Aguirre, L.; Arbones-Mainar, J.M. The Dietary Antioxidant Piceatannol Inhibits Adipogenesis of Human Adipose Mesenchymal Stem Cells and Limits Glucose Transport and Lipogenic Activities in Adipocytes. Int. J. Mol. Sci. 2018, 19, 2081. [Google Scholar] [CrossRef] [PubMed]
- Matijašić, M.; Meštrović, T.; Čipčić Paljetak, H.; Perić, M.; Barešić, A.; Verbanac, D. Gut Microbiota beyond Bacteria—Mycobiome, Virome, Archaeome, and Eukaryotic Parasites in IBD. Int. J. Mol. Sci. 2020, 21, 2668. [Google Scholar] [CrossRef]
- Hills, R.; Pontefract, B.; Mishcon, H.; Black, C.; Sutton, S.; Theberge, C. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients 2019, 11, 1613. [Google Scholar] [CrossRef] [PubMed]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Reddy, D.N. Role of the Normal Gut Microbiota. World J. Gastroenterol. 2015, 21, 8787. [Google Scholar] [CrossRef]
- Larsen, N.; Vogensen, F.K.; Van Den Berg, F.W.J.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut Microbiota in Human Adults with Type 2 Diabetes Differs from Non-Diabetic Adults. PLOS ONE 2010, 5, e9085. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R. Microbiota and Diabetes: An Evolving Relationship. Gut 2014, 63, 1513–1521. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-Chain Fatty Acids in Control of Body Weight and Insulin Sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in Gut Microbiota Control Metabolic Endotoxemia-Induced Inflammation in High-Fat Diet–Induced Obesity and Diabetes in Mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Tanase, D.M.; Gosav, E.M.; Neculae, E.; Costea, C.F.; Ciocoiu, M.; Hurjui, L.L.; Tarniceriu, C.C.; Maranduca, M.A.; Lacatusu, C.M.; Floria, M.; et al. Role of Gut Microbiota on Onset and Progression of Microvascular Complications of Type 2 Diabetes (T2DM). Nutrients 2020, 12, 3719. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic Endotoxemia Initiates Obesity and Insulin Resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [PubMed]
- Grasset, E.; Puel, A.; Charpentier, J.; Collet, X.; Christensen, J.E.; Tercé, F.; Burcelin, R. A Specific Gut Microbiota Dysbiosis of Type 2 Diabetic Mice Induces GLP-1 Resistance through an Enteric NO-Dependent and Gut-Brain Axis Mechanism. Cell Metab. 2017, 25, 1075–1090.e5. [Google Scholar] [CrossRef] [PubMed]
- Koudoufio, M.; Desjardins, Y.; Feldman, F.; Spahis, S.; Delvin, E.; Levy, E. Insight into Polyphenol and Gut Microbiota Crosstalk: Are Their Metabolites the Key to Understand Protective Effects against Metabolic Disorders? Antioxidants 2020, 9, 982. [Google Scholar] [CrossRef]
- Anhê, F.F.; Roy, D.; Pilon, G.; Dudonné, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A Polyphenol-Rich Cranberry Extract Protects from Diet-Induced Obesity, Insulin Resistance and Intestinal Inflammation in Association with Increased Akkermansia Spp. Population in the Gut Microbiota of Mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef]
- Chen, K.; Gao, Z.; Ding, Q.; Tang, C.; Zhang, H.; Zhai, T.; Xie, W.; Jin, Z.; Zhao, L.; Liu, W. Effect of Natural Polyphenols in Chinese Herbal Medicine on Obesity and Diabetes: Interactions among Gut Microbiota, Metabolism, and Immunity. Front. Nutr. 2022, 9, 962720. [Google Scholar] [CrossRef]
- Hara, T.; Hirasawa, A.; Sun, Q.; Sadakane, K.; Itsubo, C.; Iga, T.; Adachi, T.; Koshimizu, T.; Hashimoto, T.; Asakawa, Y.; et al. Novel Selective Ligands for Free Fatty Acid Receptors GPR120 and GPR40. Naunyn-Schmiedeberg's Arch. Pharmacol. 2009, 380, 247–255. [Google Scholar] [CrossRef]
- Takikawa, M.; Kurimoto, Y.; Tsuda, T. Curcumin Stimulates Glucagon-like Peptide-1 Secretion in GLUTag Cells via Ca2+/Calmodulin-Dependent Kinase II Activation. Biochem. Biophys. Res. Commun. 2013, 435, 165–170. [Google Scholar] [CrossRef]
- Kato, M.; Nishikawa, S.; Ikehata, A.; Dochi, K.; Tani, T.; Takahashi, T.; Imaizumi, A.; Tsuda, T. Curcumin Improves Glucose Tolerance via Stimulation of Glucagon-like Peptide-1 Secretion. Mol. Nutr. Food Res. 2017, 61, 1600471. [Google Scholar] [CrossRef] [PubMed]
- Rehman, K.; Ali, M.B.; Akash, M.S.H. Genistein Enhances the Secretion of Glucagon-like Peptide-1 (GLP-1) via Downregulation of Inflammatory Responses. Biomed. Pharmacother. 2019, 112, 108670. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Johnson, M.H.; Lila, M.A.; Yousef, G.; De Mejia, E.G. Berry and Citrus Phenolic Compounds Inhibit Dipeptidyl Peptidase IV: Implications in Diabetes Management. Evid.-Based Complementary Altern. Med. 2013, 2013, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tzounis, X.; Vulevic, J.; Kuhnle, G.G.C.; George, T.; Leonczak, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P.E. Flavanol Monomer-Induced Changes to the Human Faecal Microflora. Br. J. Nutr. 2008, 99, 782–792. [Google Scholar] [CrossRef]
- Unno, T.; Sakuma, M.; Mitsuhashi, S. Effect of Dietary Supplementation of (-)-Epigallocatechin Gallate on Gut Microbiota and Biomarkers of Colonic Fermentation in Rats. J. Nutr. Sci. Vitaminol. 2014, 60, 213–219. [Google Scholar] [CrossRef]
- Prabhakar, P.K.; Doble, M. Synergistic Effect of Phytochemicals in Combination with Hypoglycemic Drugs on Glucose Uptake in Myotubes. Phytomedicine 2009, 16, 1119–1126. [Google Scholar] [CrossRef]
- Prabhakar, P.K.; Doble, M. Interaction of Phytochemicals with Hypoglycemic Drugs on Glucose Uptake in L6 Myotubes. Phytomedicine 2011, 18, 285–291. [Google Scholar] [CrossRef]
- Upadhyay, S.; Dixit, M. Role of Polyphenols and Other Phytochemicals on Molecular Signaling. Oxid. Med. Cell. Longev. 2015, 2015, 1–15. [Google Scholar] [CrossRef]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human Gut Microbes Impact Host Serum Metabolome and Insulin Sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martínez-González, M.A.; Salas-Salvadó, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-Analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef]
- Bragg, F.; Kartsonaki, C.; Guo, Y.; Holmes, M.; Du, H.; Yu, C.; Pei, P.; Yang, L.; Jin, D.; Chen, Y.; et al. Circulating Metabolites and the Development of Type 2 Diabetes in Chinese Adults. Diabetes Care 2022, 45, 477–480. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Lapworth, A.L.; An, J.; Wang, L.; McGarrah, R.W.; Stevens, R.D.; Ilkayeva, O.; George, T.; Muehlbauer, M.J.; Bain, J.R.; et al. Branched-Chain Amino Acid Restriction in Zucker-Fatty Rats Improves Muscle Insulin Sensitivity by Enhancing Efficiency of Fatty Acid Oxidation and Acyl-Glycine Export. Mol. Metab. 2016, 5, 538–551. [Google Scholar] [CrossRef] [PubMed]
- Bartova, S.; Madrid-Gambin, F.; Fernández, L.; Carayol, J.; Meugnier, E.; Segrestin, B.; Delage, P.; Vionnet, N.; Boizot, A.; Laville, M.; et al. Grape Polyphenols Decrease Circulating Branched Chain Amino Acids in Overfed Adults. Front. Nutr. 2022, 9, 998044. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.S.; Cuerrier, A.; Lamont, E.; Haddad, P.S.; Arnason, J.T.; Bennett, S.A.L.; Johns, T. Investigating Wild Berries as a Dietary Approach to Reducing the Formation of Advanced Glycation Endproducts: Chemical Correlates of In Vitro Antiglycation Activity. Plant Foods Hum. Nutr. 2014, 69, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Ramkissoon, J.; Mahomoodally, M.; Ahmed, N.; Subratty, A. Antioxidant and Anti–Glycation Activities Correlates with Phenolic Composition of Tropical Medicinal Herbs. Asian Pac. J. Trop. Med. 2013, 6, 561–569. [Google Scholar] [CrossRef]
- Coelho, O.G.L.; Ribeiro, P.V.M.; Alfenas, R.D.C.G. Can Grape Polyphenols Affect Glycation Markers? A Systematic Review. Crit. Rev. Food Sci. Nutr. 2023, 63, 1208–1218. [Google Scholar] [CrossRef]
- Larrosa, M.; González-Sarrías, A.; Yáñez-Gascón, M.J.; Selma, M.V.; Azorín-Ortuño, M.; Toti, S.; Tomás-Barberán, F.; Dolara, P.; Espín, J.C. Anti-Inflammatory Properties of a Pomegranate Extract and Its Metabolite Urolithin-A in a Colitis Rat Model and the Effect of Colon Inflammation on Phenolic Metabolism. J. Nutr. Biochem. 2010, 21, 717–725. [Google Scholar] [CrossRef]
- Nesto, RW. Correlation between cardiovascular disease and diabetes mellitus: current concepts. Am J Med. 2004, 116, 11–22. [Google Scholar] [CrossRef]
- Shi, Y.; Vanhoutte, P.M. Macro- and microvascular endothelial dysfunction in diabetes. J. Diabetes 2017, 9, 434–449. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Liu, S.; Gao, M.; Wang, W.; Chen, K.; Huang, L.; Liu, Y. Diabetic Vascular Diseases: Molecular Mechanisms and Therapeutic Strategies. Signal Transduct. Target. Ther. 2023, 8, 152. [Google Scholar] [CrossRef]
- Giugliano, D.; Ceriello, A.; Paolisso, G. Oxidative Stress and Diabetic Vascular Complications. Diabetes Care 1996, 19, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Rojas, A.; Figueroa, H.; Re, L.; Morales, M.A. Oxidative Stress at the Vascular Wall. Mechanistic and Pharmacological Aspects. Arch. Med. Res. 2006, 37, 436–448. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Mann, G.E. Vascular NAD(P)H Oxidase Activation in Diabetes: A Double-Edged Sword in Redox Signalling. Cardiovasc. Res. 2009, 82, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Deanfield, J.E.; Halcox, J.P.; Rabelink, T.J. Endothelial Function and Dysfunction: Testing and Clinical Relevance. Circulation 2007, 115, 1285–1295. [Google Scholar] [CrossRef] [PubMed]
- Krüger-Genge, A.; Blocki, A.; Franke, R.-P.; Jung, F. Vascular Endothelial Cell Biology: An Update. Int. J. Mol. Sci. 2019, 20, 4411. [Google Scholar] [CrossRef]
- Boulanger, C. M. Endothelium. Arterioscler. Thromb. Vasc. Biol. 2016, 36, e26–e31. [Google Scholar] [CrossRef]
- Tousoulis, D.; Kampoli, A.-M.; Tentolouris, C.; Papageorgiou, N.; Stefanadis, C. The Role of Nitric Oxide on Endothelial Function. Curr. Vasc. Pharmacol. 2012, 10, 4–18. [Google Scholar] [CrossRef]
- Zou, M.-H.; Cohen, R.A.; Ullrich, V. Peroxynitrite and Vascular Endothelial Dysfunction in Diabetes Mellitus. Endothelium 2004, 11, 89–97. [Google Scholar] [CrossRef]
- Szabo, C. Role of Nitrosative Stress in the Pathogenesis of Diabetic Vascular Dysfunction. Br. J. Pharmacol. 2009, 156, 713–727. [Google Scholar] [CrossRef]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef]
- Martins, T.F.; Palomino, O.M.; Álvarez-Cilleros, D.; Martín, M.A.; Ramos, S.; Goya, L. Cocoa Flavanols Protect Human Endothelial Cells from Oxidative Stress. Plant Foods Hum. Nutr. 2020, 75, 161–168. [Google Scholar] [CrossRef]
- Zhou, H.; Fu, B.; Xu, B.; Mi, X.; Li, G.; Ma, C.; Xie, J.; Li, J.; Wang, Z. Rosmarinic Acid Alleviates the Endothelial Dysfunction Induced by Hydrogen Peroxide in Rat Aortic Rings via Activation of AMPK. Oxid. Med. Cell Longev. 2017, 2017, 7091904. [Google Scholar] [CrossRef]
- Serraino, I.; Dugo, L.; Dugo, P.; Mondello, L.; Mazzon, E.; Dugo, G.; Caputi, A.P.; Cuzzocrea, S. Protective Effects of Cyanidin-3-O-Glucoside from Blackberry Extract against Peroxynitrite-Induced Endothelial Dysfunction and Vascular Failure. Life Sci. 2003, 73, 1097–1114. [Google Scholar] [CrossRef] [PubMed]
- Spanier, G.; Xu, H.; Xia, N.; Tobias, S.; Deng, S.; Wojnowski, L.; Forstermann, U.; Li, H. Resveratrol Reduces Endothelial Oxidative Stress by Modulating the Gene Expression of Superoxide Dismutase 1 (SOD1), Glutathione Peroxidase 1 (GPx1) and NADPH Oxidase Subunit (Nox4). J. Physiol. Pharmacol. 2009, 60 (Suppl. S4), 111–116. [Google Scholar] [PubMed]
- Alvarez, S.; Zaobornyj, T.; Actis-Goretta, L.; Fraga, C.G.; Boveris, A. Polyphenols and Red Wine as Peroxynitrite Scavengers: A Chemiluminescent Assay. Ann. N. Y. Acad. Sci. 2002, 957, 271–273. [Google Scholar] [CrossRef]
- Liang, X.-X.; Wang, R.-Y.; Guo, Y.-Z.; Cheng, Z.; Lv, D.-Y.; Luo, M.-H.; He, A.; Luo, S.-X.; Xia, Y. Phosphorylation of Akt at Thr308 Regulates P-eNOS Ser1177 during Physiological Conditions. FEBS Open Bio 2021, 11, 1953–1964. [Google Scholar] [CrossRef] [PubMed]
- Madeira, S.V.F.; Auger, C.; Anselm, E.; Chataigneau, M.; Chataigneau, T.; Soares de Moura, R.; Schini-Kerth, V.B. eNOS Activation Induced by a Polyphenol-Rich Grape Skin Extract in Porcine Coronary Arteries. J. Vasc. Res. 2009, 46, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Edirisinghe, I.; Burton-Freeman, B.; Tissa Kappagoda, C. Mechanism of the Endothelium-Dependent Relaxation Evoked by a Grape Seed Extract. Clin. Sci. 2008, 114, 331–337. [Google Scholar] [CrossRef]
- Wallerath, T.; Poleo, D.; Li, H.; Förstermann, U. Red Wine Increases the Expression of Human Endothelial Nitric Oxide Synthase. J. Am. Coll. Cardiol. 2003, 41, 471–478. [Google Scholar] [CrossRef]
- Wallerath, T.; Li, H.; Gödtel-Ambrust, U.; Schwarz, P.M.; Förstermann, U. A Blend of Polyphenolic Compounds Explains the Stimulatory Effect of Red Wine on Human Endothelial NO Synthase. Nitric Oxide 2005, 12, 97–104. [Google Scholar] [CrossRef]
- Wallerath, T.; Deckert, G.; Ternes, T.; Anderson, H.; Li, H.; Witte, K.; Förstermann, U. Resveratrol, a Polyphenolic Phytoalexin Present in Red Wine, Enhances Expression and Activity of Endothelial Nitric Oxide Synthase. Circulation 2002, 106, 1652–1658. [Google Scholar] [CrossRef]
- Xia, N.; Strand, S.; Schlufter, F.; Siuda, D.; Reifenberg, G.; Kleinert, H.; Förstermann, U.; Li, H. Role of SIRT1 and FOXO Factors in eNOS Transcriptional Activation by Resveratrol. Nitric Oxide 2013, 32, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Goldin, A.; Beckman, J.A.; Schmidt, A.M.; Creager, M.A. Advanced Glycation End Products: Sparking the Development of Diabetic Vascular Injury. Circulation 2006, 114, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M.; Vlassara, H.; Cerami, A. Nonenzymatic Glycosylation and the Pathogenesis of Diabetic Complications. Ann. Intern. Med. 1984, 101, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Cerami, A.; Vlassara, H.; Brownlee, M. Role of Advanced Glycosylation Products in Complications of Diabetes. Diabetes Care 1988, 11 Suppl 1, 73–79. [Google Scholar]
- Khalid, M.; Petroianu, G.; Adem, A. Advanced Glycation End Products and Diabetes Mellitus: Mechanisms and Perspectives. Biomolecules 2022, 12, 542. [Google Scholar] [CrossRef]
- Peppa, M.; Uribarri, J.; Vlassara, H. Glucose, Advanced Glycation End Products, and Diabetes Complications: What Is New and What Works. Clin. Diabetes 2003, 21, 186–187. [Google Scholar] [CrossRef]
- Twarda-Clapa, A.; Olczak, A.; Białkowska, A.M.; Koziołkiewicz, M. Advanced Glycation End-Products (AGEs): Formation, Chemistry, Classification, Receptors, and Diseases Related to AGEs. Cells 2022, 11, 1312. [Google Scholar] [CrossRef]
- Brings, S.; Fleming, T.; Freichel, M.; Muckenthaler, M.U.; Herzig, S.; Nawroth, P.P. Dicarbonyls and Advanced Glycation End-Products in the Development of Diabetic Complications and Targets for Intervention. Int. J. Mol. Sci. 2017, 18, 984. [Google Scholar] [CrossRef]
- Uribarri, J.; del Castillo, M.D.; de la Maza, M.P.; Filip, R.; Gugliucci, A.; Luevano-Contreras, C.; Macías-Cervantes, M.H.; Markowicz Bastos, D.H.; Medrano, A.; Menini, T.; et al. Dietary Advanced Glycation End Products and Their Role in Health and Disease. Adv. Nutr. 2015, 6, 461–473. [Google Scholar] [CrossRef]
- Gautieri, A.; Passini, F.S.; Silván, U.; Guizar-Sicairos, M.; Carimati, G.; Volpi, P.; Moretti, M.; Schoenhuber, H.; Redaelli, A.; Berli, M.; et al. Advanced Glycation End-Products: Mechanics of Aged Collagen from Molecule to Tissue. Matrix Biol. 2017, 59, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Rojas, A.; Añazco, C.; González, I.; Araya, P. Extracellular Matrix Glycation and Receptor for Advanced Glycation End-Products Activation: A Missing Piece in the Puzzle of the Association between Diabetes and Cancer. Carcinogenesis 2018, 39, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Rojas, A.; Delgado-López, F.; González, I.; Pérez-Castro, R.; Romero, J.; Rojas, I. The Receptor for Advanced Glycation End-Products: A Complex Signaling Scenario for a Promiscuous Receptor. Cell Signal. 2013, 25, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Hudson, B.I.; Lippman, M.E. Targeting RAGE Signaling in Inflammatory Disease. Annu. Rev. Med. 2018, 69, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’nikova, T.I.; Porozov, Y.B.; Terentiev, A.A. Oxidative Stress and Advanced Lipoxidation and Glycation End Products (ALEs and AGEs) in Aging and Age-Related Diseases. Oxid. Med. Cell. 2019, 2019, 1–14. [Google Scholar] [CrossRef]
- Wautier, M.P.; Chappey, O.; Corda, S.; Stern, D.M.; Schmidt, A.M.; Wautier, J.L. Activation of NADPH Oxidase by AGE Links Oxidant Stress to Altered Gene Expression via RAGE. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E685–694. [Google Scholar] [CrossRef]
- Yeh, W.-J.; Hsia, S.-M.; Lee, W.-H.; Wu, C.-H. Polyphenols with Antiglycation Activity and Mechanisms of Action: A Review of Recent Findings. J. Food Drug Anal. 2017, 25, 84–92. [Google Scholar] [CrossRef]
- Khanam, A.; Ahmad, S.; Husain, A.; Rehman, S.; Farooqui, A.; Yusuf, M.A. Glycation and Antioxidants: Hand in the Glove of Antiglycation and Natural Antioxidants. Curr. Protein Pept. Sci. 2020, 21, 899–915. [Google Scholar] [CrossRef]
- González, I.; Morales, M.A.; Rojas, A. Polyphenols and AGEs/RAGE Axis. Trends and Challenges. Food Res. Int. 2020, 129, 108843. [Google Scholar] [CrossRef]
- Sun, C.; Zhao, C.; Guven, E.C.; Paoli, P.; Simal-Gandara, J.; Ramkumar, K.M.; Wang, S.; Buleu, F.; Pah, A.; Turi, V.; et al. Dietary Polyphenols as Antidiabetic Agents: Advances and Opportunities. Food Front. 2020, 1, 18–44. [Google Scholar] [CrossRef]
- Gugliucci, A.; Bastos, D.H.M.; Schulze, J.; Souza, M.F.F. Caffeic and Chlorogenic Acids in Ilex Paraguariensis Extracts Are the Main Inhibitors of AGE Generation by Methylglyoxal in Model Proteins. Fitoterapia 2009, 80, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Genaro-Mattos, T.C.; Maurício, Â.Q.; Rettori, D.; Alonso, A.; Hermes-Lima, M. Antioxidant Activity of Caffeic Acid against Iron-Induced Free Radical Generation—A Chemical Approach. PLOS ONE 2015, 10, e0129963. [Google Scholar] [CrossRef]
- Bhuiyan, M.N.I.; Mitsuhashi, S.; Sigetomi, K.; Ubukata, M. Quercetin Inhibits Advanced Glycation End Product Formation via Chelating Metal Ions, Trapping Methylglyoxal, and Trapping Reactive Oxygen Species. Biosci. Biotechnol. Biochem. 2017, 81, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yagiz, Y.; Buran, T.J.; Nunes, C.D.N.; Gu, L. Phytochemicals from Berries and Grapes Inhibited the Formation of Advanced Glycation End-products by Scavenging Reactive Carbonyls. Food Res. Int. 2011, 44, 2666–2673. [Google Scholar] [CrossRef]
- Thornalley, P.J. The Glyoxalase System: New Developments towards Functional Characterization of a Metabolic Pathway Fundamental to Biological Life. Biochem. J. 1990, 269, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Frandsen, J.; Narayanasamy, P. Flavonoid Enhances the Glyoxalase Pathway in Cerebellar Neurons to Retain Cellular Functions. Sci. Rep. 2017, 7, 5126. [Google Scholar] [CrossRef] [PubMed]
- Buttari, B.; Profumo, E.; Facchiano, F.; Ozturk, E.I.; Segoni, L.; Saso, L.; Riganò, R. Resveratrol Prevents Dendritic Cell Maturation in Response to Advanced Glycation End Products. Oxid. Med. Cell. Longev. 2013, 2013, 1–12. [Google Scholar] [CrossRef]
- Kanlaya, R.; Thongboonkerd, V. Molecular Mechanisms of Epigallocatechin-3-Gallate for Prevention of Chronic Kidney Disease and Renal Fibrosis: Preclinical Evidence. Curr. Dev. Nutr. 2019, 3, nzz101. [Google Scholar] [CrossRef]
- Dong, L.; Li, Y.; Chen, Q.; Liu, Y.; Wu, Z.; Pan, D.; Yan, N.; Liu, L. Cereal Polyphenols Inhibition Mechanisms on Advanced Glycation End Products and Regulation on Type 2 Diabetes. Crit. Rev. Food Sci. Nutr. 2023, 1–19. [Google Scholar] [CrossRef]
- Jonas, R.A.; Crabtree, T.R.; Jennings, R.S.; Marques, H.; Katz, R.J.; Chang, H.-J.; Stuijfzand, W.J.; van Rosendael, A.R.; Choi, J.H.; Doh, J.-H.; et al. Diabetes, Atherosclerosis, and Stenosis by AI. Diabetes Care 2023, 46, 416–424. [Google Scholar] [CrossRef]
- Yuan, T.; Yang, T.; Chen, H.; Fu, D.; Hu, Y.; Wang, J.; Yuan, Q.; Yu, H.; Xu, W.; Xie, X. New Insights into Oxidative Stress and Inflammation during Diabetes Mellitus-Accelerated Atherosclerosis. Redox Biol. 2019, 20, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Poznyak, A.; Grechko, A.V.; Poggio, P.; Myasoedova, V.A.; Alfieri, V.; Orekhov, A.N. The Diabetes Mellitus-Atherosclerosis Connection: The Role of Lipid and Glucose Metabolism and Chronic Inflammation. Int. J. Mol. Sci. 2020, 21, 1835. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Q.; Zhang, F.; Zhu, W.; Wu, J.; Liang, M. Copper in Diabetes Mellitus: A Meta-Analysis and Systematic Review of Plasma and Serum Studies. Biol. Trace Elem. Res. 2017, 177, 53–63. [Google Scholar] [CrossRef]
- Banerjee, J.; Mishra, N.; Damle, G.; Dhas, Y. Beyond LDL-c: The Importance of Serum Oxidized LDL in Predicting Risk for Type 2 Diabetes in the Middle-Aged Asian Indians. Diabetes Metab. Syndr. 2019, 13, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.S.; Brownlee, M. Molecular and Cellular Mechanisms of Cardiovascular Disorders in Diabetes. Circ. Res. 2016, 118, 1808–1829. [Google Scholar] [CrossRef]
- Islam, M.A.; Amin, M.N.; Siddiqui, S.A.; Hossain, M.P.; Sultana, F.; Kabir, M.R. Trans Fatty Acids and Lipid Profile: A Serious Risk Factor to Cardiovascular Disease, Cancer and Diabetes. Diabetes Metab. Syndr. 2019, 13, 1643–1647. [Google Scholar] [CrossRef]
- Heinloth, A.; Heermeier, K.; Raff, U.; Wanner, C.; Galle, J. Stimulation of NADPH Oxidase by Oxidized Low-Density Lipoprotein Induces Proliferation of Human Vascular Endothelial Cells. J. Am. Soc. Nephrol. 2000, 11, 1819–1825. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, G.Z.; Rabinovitch, P.S.; Tabas, I. Macrophage Mitochondrial Oxidative Stress Promotes Atherosclerosis and Nuclear Factor-κB-Mediated Inflammation in Macrophages. Circ. Res. 2014, 114, 421–433. [Google Scholar] [CrossRef]
- Feldman, F.; Koudoufio, M.; Desjardins, Y.; Spahis, S.; Delvin, E.; Levy, E. Efficacy of Polyphenols in the Management of Dyslipidemia: A Focus on Clinical Studies. Nutrients 2021, 13, 672. [Google Scholar] [CrossRef]
- Cayatte, A.J.; Rupin, A.; Oliver-Krasinski, J.; Maitland, K.; Sansilvestri-Morel, P.; Boussard, M.F.; Wierzbicki, M.; Verbeuren, T.J.; Cohen, R.A. S17834, a New Inhibitor of Cell Adhesion and Atherosclerosis That Targets Nadph Oxidase. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1577–1584. [Google Scholar] [CrossRef]
- Zang, M.; Xu, S.; Maitland-Toolan, K.A.; Zuccollo, A.; Hou, X.; Jiang, B.; Wierzbicki, M.; Verbeuren, T.J.; Cohen, R.A. Polyphenols Stimulate AMP-Activated Protein Kinase, Lower Lipids, and Inhibit Accelerated Atherosclerosis in Diabetic LDL Receptor-Deficient Mice. Diabetes 2006, 55, 2180–2191. [Google Scholar] [CrossRef] [PubMed]
- Frémont, L.; Belguendouz, L.; Delpal, S. Antioxidant Activity of Resveratrol and Alcohol-Free Wine Polyphenols Related to LDL Oxidation and Polyunsaturated Fatty Acids. Life Sci. 1999, 64, 2511–2521. [Google Scholar] [CrossRef] [PubMed]
- Aviram, M.; Fuhrman, B. Wine Flavonoids Protect against LDL Oxidation and Atherosclerosis. Ann. N. Y. Acad. Sci. 2002, 957, 146–161. [Google Scholar] [CrossRef]
- Berrougui, H.; Grenier, G.; Loued, S.; Drouin, G.; Khalil, A. A New Insight into Resveratrol as an Atheroprotective Compound: Inhibition of Lipid Peroxidation and Enhancement of Cholesterol Efflux. Atherosclerosis 2009, 207, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Regnström, J.; Nilsson, J.; Tornvall, P.; Landou, C.; Hamsten, A. Susceptibility to Low-Density Lipoprotein Oxidation and Coronary Atherosclerosis in Man. Lancet 1992, 339, 1183–1186. [Google Scholar] [CrossRef] [PubMed]
- Heinecke, J.W. Oxidants and Antioxidants in the Pathogenesis of Atherosclerosis: Implications for the Oxidized Low Density Lipoprotein Hypothesis. Atherosclerosis 1998, 141, 1–15. [Google Scholar] [CrossRef]
- Li, J.; Zhong, Z.; Yuan, J.; Chen, X.; Huang, Z.; Wu, Z. Resveratrol Improves Endothelial Dysfunction and Attenuates Atherogenesis in Apolipoprotein E-Deficient Mice. J. Nutr. Biochem. 2019, 67, 63–71. [Google Scholar] [CrossRef]
- Yang, H.; Yang, T.; Heng, C.; Zhou, Y.; Jiang, Z.; Qian, X.; Du, L.; Mao, S.; Yin, X.; Lu, Q. Quercetin Improves Nonalcoholic Fatty Liver by Ameliorating Inflammation, Oxidative Stress, and Lipid Metabolism in Db/Db Mice. Phytother. Res. 2019, 33, 3140–3152. [Google Scholar] [CrossRef]
- Hosseini, A.; Razavi, B.M.; Banach, M.; Hosseinzadeh, H. Quercetin and Metabolic Syndrome: A Review. Phytother. Res. 2021, 35, 5352–5364. [Google Scholar] [CrossRef]
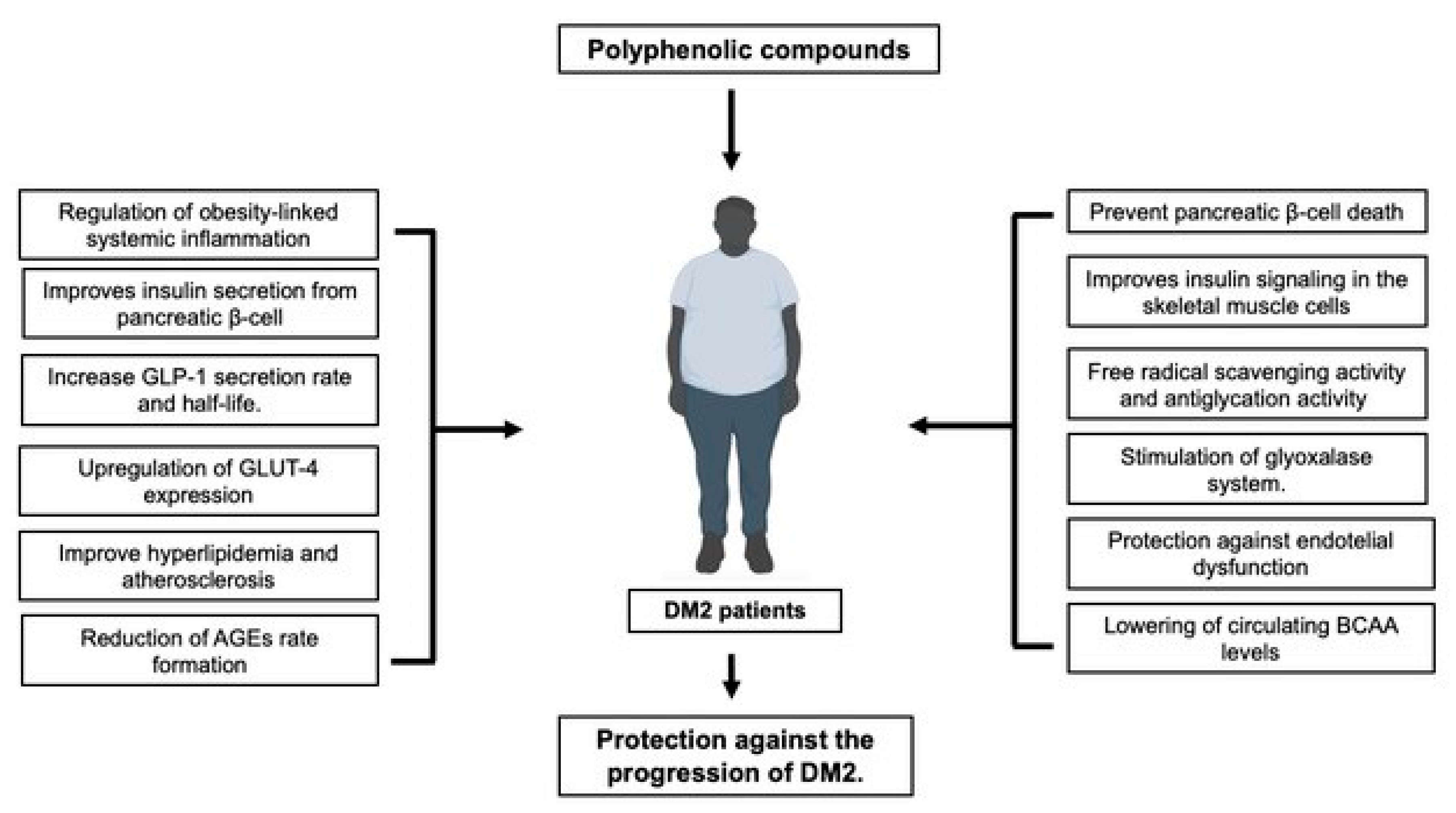
Figure 1.
Polyphenols have multifaceted actions to support their use in the management of DM2, due to their effects on pancreatic islet β-cell functioning and promotion of insulin production and signaling, protection against micro-and microvascular complications, protection against the progression of T2DM-associated obesity, management of dyslipidemia and gut microbiome dysbiosis.
Figure 1.
Polyphenols have multifaceted actions to support their use in the management of DM2, due to their effects on pancreatic islet β-cell functioning and promotion of insulin production and signaling, protection against micro-and microvascular complications, protection against the progression of T2DM-associated obesity, management of dyslipidemia and gut microbiome dysbiosis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



