
Submitted:
03 July 2023
Posted:
04 July 2023
You are already at the latest version
Abstract
Changes in food quality can dramatically impair zooplankton fitness, especially in eutrophic water bodies where cyanobacteria are usually predominant. Cyanobacteria are considered a food with low nutritional value, besides some species can produce bioactive secondary metabolites reported as toxic to zooplankton. Considering that cyanobacteria can limit survival, growth and reproduction of zooplankton, we hypothesized that the adverse effects of the dietary exposure of Daphnia species to saxitoxin-producing cyanobacteria depend on the relative abundance of nutitious food source in the diet. Life table experiments were conducted with different diets: 1) without nutritional restriction, where neonates were fed with diets at a constant green algae (Monoraphidium capricornutum) biomass as a nutritious food source, and increasing cyanobacterial concentration (STX-producing Raphidiopsis raciborskii), and 2) with diets consisting of different proportions of M. capricornutum (nutritious) and R. raciborskii (poor food) at a total biomass 1.0 mg C L 1. In general, the presence of high proportions of cyanobacteria promoted a decrease in Daphnia somatic growth, reproduction and intrinsic rate of population increase (r) in both diets with more pronounced effects in the nutritionally restricted diet (90% R. raciborskii). Two-way ANOVA revealed significant effects of species/clone and treatments in both assays, with significant interaction between those factors only in the second assay. Regarding the grazing assay, only D. laevis was negatively affected by increased cyanobacterial proportions in the diet. In the life table assay with constant edible food, a reduction in the reproduction parameters and the intrinsic rate of population increase (r) of all species was observed, In conclusion, we found adverse effects of the toxic cyanobacterial strain R. raciborskii on Daphnia fitness, regardless of the constant amount of nutritious food available, proving the toxic effect of R. raciborskii and the nutritional quality of the food has a greater influence on the fitness of these animals.
Keywords:
zooplankton
; cyanotoxins
; nutritional constraint
; cyanobacteria
1. Introduction
Factors such as intensified industrialization, urban growth, and agricultural activities lead to increased inputs of nitrogen and phosphate nutrients in aquatic ecosystems, causing eutrophication (Chorus & Welker, 2021). This process has favored the occurrence of cyanobacterial harmful algal blooms (CyanoHABs) which have been boosted by climate changes (Huisman et al., 2018). Bloom-forming cyanobacteria can cause negative impacts in aquatic ecosystems by cyanotoxins production, oxygen depletion, shading of benthic habitats and trophic decoupling (Havens & Hudnel, 2008; Pearl & Huisman, 2008; Moustaka-Gouni & Sommer, 2020).
Cyanobacteria are photosynthetic prokaryotes that usually compose marine and freshwater phytoplankton, and contribute to the primary production in aquatic food webs as a significant food source for zooplankton consumers, especially in eutrophic water bodies (Lürling & Verschoor, 2003; Nogueira et al., 2004; Ger et al., 2016). However, dietary exposure to cyanobacteria can affect zooplankton due to low nutritional quality, such as the lack of polyunsaturated fatty acids (PUFAs) and sterols (Martin-Creuzeburg et al., 2008); difficulted digestibility (DeMott & Van Donk, 2013) or by the ability of some cyanobacterial genera on producing bioactive secondary metabolites (cyanotoxins) which can affect zooplankton motility, feeding behavior, survival and life cycle (Ferrão-Filho & Kozlowsky-Suzuki, 2011; Vilar, Ferrão-Filho & Azevedo, 2022).
Among cyanobacteria, Raphidiopsis raciborskii (formerly Cylindrospermopsis raciborskii) (Aguilera et al., 2018) stands out as a N-fixing and bloom-forming species which has been recently reported for higher latitudes as an invasive aquatic microbe. The ability of R. raciborskii on spreading over several regions is attributed to its high phenotypic plasticity regarding tolerance to light, salinity, ability to storage phosphorus, toxin production, and resistance to herbivory (Antunes et al. 2015; Burford et al., 2016; Vilar et al., 2021). Toxin-producing strains of this species can produce cytotoxic (cylindrospermopsins) (Hawkins et al., 1997; Wood & Stirling, 2003; Lei et al., 2014) and neurotoxic (saxitoxins, goniautoxins, and C toxins, hereafter STXs) alkaloids; being the latter toxins produced up to date only by south American strains (Lagos et al.,1999; Molica et al., 2005; Pomati et al., 2003; Ferrão-Filho et al., 2007; Piccini et al., 2011). R. raciborskii blooms are increasingly frequent in tropical lakes and reservoirs which makes this species a representative component of phytoplankton communities in these water bodies (Soares et al., 2013; Rzymski et al., 2014; Yang et al., 2017; Pagni et al., 2020).
Overall, the establishment of cyanoHABs usually represents a change from diverse and edible phytoplankton to a less diverse and inedible one (large colonies and filaments, and production of harmful metabolites) (Hansson et al., 2007; Moustaka-Gouni & Sommer, 2020). In turn, it can affect zooplankton composition, which shifts from a community dominated by large generalist filter feeders such as cladocerans (e.g., Daphnia), toward small-bodied selective organisms such as copepods, rotifers, and planktonic protozoans (Przytulska et al., 2015; Moustaka-Gouni & Sommer, 2020). However, although there is a consensus that cyanobacteria play a role in the trophic decoupling of freshwater food webs (Heathcote et al., 2016), mainly due to defensive chemical traits against phytoplankton consumers (Rangel et al., 2020; Vilar et al., 2021), little is known whether the increased cyanobacterial biomass affects the zooplankton due to a relatively low abundance of nutritious algae or if cyanobacterial toxicity overrides nutritional constraint regardless of high quality food availability at a non-limiting biomass concentration. Therefore, we evaluated whether the negative impacts of dietary exposure of Daphnia to toxic cyanobacteria are mostly ruled out by nutritional constraints due to nutritious algae availability. Therefore, we tested the hypothesis that the toxicity of STXs-producing R. raciborskii impairs Daphnia feeding behavior and fitness regardless of the high availability of nutritious algae.
2. Methods
2.1. Phytoplankton and zooplankton culture conditions
The saxitoxin-producing strain Raphidiopsis raciborskii CYLCAM-01 (Mesquita et al., 2019) was kindly provided by Dr. Marcelo M. Marinho (Laboratory of Ecology and Physiology of Phytoplankton – Rio de Janeiro State University). The cyanobacterium was isolated from the Camorim reservoir; a eutrophic water body located at the Pedra Branca State Park (PEPB) – Rio de Janeiro, Brazil. The strain was cultivated in a WC medium (Guillard, 1975) with initial pH adjusted to 8.0 and maintained at a temperature of 23±1 °C, the light intensity of 50 μmol m-2s-1 and a 12:12 h light/dark photoperiod.
The tested zooplankton consisted of neotropical Daphnia species isolated from different water bodies: D. laevis (Lagoa do Ibirité-Minas Gerais), D. laevis (Lagoa Jacaré-Minas Gerais) and D. gessneri (Reservatório do Apertado-Bahia). Animals were kept in 1000 mL beakers filled with artificial RT medium (Tollrian, 1993) enriched with 30% filtered (filter 1-2 µm pore size, Sartorius) natural water from decantation tanks of the CEDAE treatment plant (State Company for Water and Sewage, Camorim Reservoir) and supplied with commercial 0.05% humic acid extract (~1.125 mg C L-1, Microbe Lift® Amazon Black & Soft Water Conditioner, USA) at an initial pH 7.6. The animals were incubated at 23.5 ± 1 °C under low light intensity (20 μmol m-2s-1) and a 12:12h light/dark photoperiod. To feed the animals, the green microalgae Monoraphidium capricornutum and Ankistrodesmus stiptatus were cultivated in oligo medium (Afnor, 1980) with initial pH adjusted to 7.0. For cultures maintenance, the animals are usually fed at each two days with a total carbon biomass of 0.4 mg L-1 of both algal species (1:1).
2.2. Phytoplankton biomass estimative and saxitoxin analysis
Cyanobacterial filaments and microalgal cells were counted in a Fuchs-Rosenthal chamber under an optical microscope to determine the cell/filament density (unit mL-1). Subsequently, the population density was converted to biovolume (mm3 L-1) from the average cell/filament volume (µm3) (Hillebrand et al., 2002; Sun & Liu, 2003) obtained by cells and filaments dimensions (length and width) measurements under an optical microscope with the aid of CellB software (Soft Imaging System GmbH, Münster, Germany). Finally, the carbon biomass (mgC L-1) was estimated from cell volume measurements as described by Rocha & Duncan (1985).
STX was analyzed by ELISA (Enzyme-Linked Immunoabsorbent Assay) using a Beacon saxitoxin kit (Beacon Analytical Systems, Inc.) according to the manufacturer’s recommendation.
2.3. Life table experiments
Life-table experiments were run to evaluate whether the amount of saxitoxin-producing cyanobacteria in the diet could affect zooplankton fitness, biomass acquisition, and survivorship, despite the availability of nutritious green microalgae (hereafter nutritious food) as follows:
2.4. Dietary exposure to STX-producing cyanobacteria under constant nutritious food
In the first experiment, neonates (<24h) of the species D. laevis (RD clone), D. laevis (IB clone) e D. gessneri were exposed to increased concentrations of the toxic R. raciborskii CYLCAM-01 (0, 0.5, 1.0 and 1.5 mg C L-1) and a constant biomass concentration of nutritious food (0.4 mg C L-1 of M. capricornutum) during 15 days. The experiment was run with one individual per test tube (n=15) filled with 30 mL of the phytoplankton cell suspensions at the different biomass concentrations (experimental diets) and incubated over 15 days. Furthermore, we assumed to keep a constant nutritious food concentration in an attempt to separate the effects of nutritional inadequacy and toxicity (Jónasdóttir et al. 1998).
2.5. Dietary exposure to STX-producing cyanobacteria under variable nutritious food
In the second experiment, neonates (<24h) from each Daphnia species/clone were exposed to increased cyanobacterial proportions (25-90%) relative to nutritious food (75-10%). For the nutritious food (green algae), we kept a higher proportion of A. stiptatus (90%) relative to M. capricornutum since this microalgae seems to be more suitable for zooplankton nutrition regarding its lipid composition (as well as phosphate content) which consists mainly of unsaturated fatty acids (C18 chains such as alpha-linolenic acid, oleic acid, and linoleic acid) (Miranda et al., 2016). Thus, the Daphnia dietary exposure to toxic R. raciborskii CYLCAM-01 was set into three experimental diets: i) 25% CYLCAM-01 + 75% nutritious food; ii) 50% CYLCAM-01 + 50% nutritious food; and iii) 90% CYLCAM-01 + 10% nutritious food. The control group consisted of 100% nutritious food. All proportions were relative to a total carbon biomass concentration of 1.0 mg C L-1. The experiment was run in test tubes (n=15) filled with 30 mL of phytoplankton cell suspensions and incubated over 15 days. For all experimental diets, the medium was daily renewed over the experiment duration.
For the life table parameters, neonates individually exposed to the different diets were assessed from newborn to maturity during 15 days or up to at least the third brood, to assess the (i) age at first reproduction (primiparous) and the number of neonates produced per female in each brood. The survival (lx) and mean fecundity (mx) were used to estimate the mean intrinsic rate of population increase (r) by bootstrap analysis (Software Rm 2.0, Taberner et al., 1993) using the following equation:
where lx is age-specific survival, mx is age-specific fecundity and x is the age in days.
At the beginning and the end of the experiments, the organisms of each treatment were measured for the body length from the top of the head to the base of the caudal spine under an Olympus magnifying glass (SZ61) with 20× magnification to estimate the somatic growth rate by the following formula:
where Li is the initial length, Lf is the final length and t is the incubation time in days.
2.6. Grazing assays
Grazing assays was performed to evaluate whether an increased proportion of saxitoxin-producing cyanobacteria in the diet, at a constant non-limiting biomass concentration, affect or not zooplankton feeding behavior.
Adult individuals (~6 days) of all Daphnia clones were dietary exposed to phytoplankton cell suspensions at increased proportions of R. raciborskii CYLCAM-01 (25, 50, and 90%) relative to the nutritious food (=green microalgae M. capricornutum), at a total carbon biomass concentration of 1.0 mg C L-1. The animals were starved (~12h) before the beginning of the experiment and then conditioned in 10 mL test tubes (n=4; 2 individuals/test tube) filled with the different experimental diets. A cellular suspension composed of 100% nutritious food was assumed as the negative control for cyanobacterial impact by dietary exposure. Alongside, all experimental diets were also established without animals as the negative control for cell loss by Daphnia grazing.
The Daphnia clones were incubated in the dark for 3 hours and subsequently, the clearance rate (CR, in mL ind-1 h-1) was estimated from chlorophyll fluorescence (ChlF0) emitted from phytoplankton cells in the experimental diets and detected by a PHYTO-PAM fluorimeter (Walz, Germany) (Lürling & Verschoor, 2003). The clearance rate was estimated using the following equation:
with Chlacontrol and Chlatreatment as the difference in the algal concentration (as chlorophyll-a specific for Cyanobacteria and green algae; µg L-1) at the initial and final time in the control (without animals) and treatments (with animals), respectively. Δt is the incubation time (h), V is the volume of culture medium (mL) and N is the number of animals.
The calculation of clearance rate was based on the fluorescence signals given: green (for green algae), blue (for cyanobacteria) and total chlorophyll (for the sum of both signals).
2.7. Data analysis
CL50 (Lethal Concentration to 50% of individuals) and CE50 (Effective Concentration where 50% of the population exhibits a response) were estimated in the online platform MOSAIC (Charles et al., 2018).
Population parameters (reproduction parameters and intrinsic rate of population increase (r)), as well as feeding behavior were examined for normality and variance homoscedasticity. Once the parametric premises were assumed, the data were compared using one-way ANOVA, and differences between treatments were analyzed by Tukey HSD test (p<0.05) using GraphPad Prism 5.0 software. To verify differences regarding the population parameters between Daphnia species/clones and treatments we used the two-way ANOVA, with species/clone and treatment as categorical factors. The intrinsic rate of population increase (r) was analyzed by Student's t-test (p<0.05), using Primer 1.0 program.
3. Results
3.1. Dietary exposure to STX-producing cyanobacteria under constant nutritious food
In this assay, the animals were exposed to a cell-bound STX concentration ranging from 1.5 to 4.5 ng L-1 as shown in Table 1. Only D. gessneri showed reduced survival by the increasing concentrations of cyanobacteria, with a reduction in the number of individuals throughout the experiment. The estimates of LC50 and CE50 are shown in Table 2. No effects were observed on the survival of D. laevis clones. There was a significant reduction in the somatic growth rate of all species tested (Figure 1).
The two-way ANOVA revealed a significant effect of species/clone and treatment in all population parameters, with no interaction between those factors (Table 3).
Despite of the constant amount of nutritious food, the presence of STX-producing cyanobacteria in the diet promoted adverse effects on Daphnia populational parameters such as age at first reproduction, fecundity, total offspring and intrinsic population growth rate (r) of all species (Figure 2). In the parameter age of first reproduction, a significant delay in reproduction (ANOVA, Tukey test, p= 0.009) was observed only at the highest concentration of cyanobacteria (Figure 2A). In the average fecundity, only D. laevis (RD) and D. gessneri suffered a significant negative effect with increasing concentrations of cyanobacteria in the diet (Figure 2B). The total number of neonates produced by D. laevis (RD) and D. gessneri were reduced significantly at all concentrations, with a greater impact on the D. laevis (RD) clone. Although the clone D. laevis (IB) was not significantly affected in the parameters of mean fecundity and total neonates produced in any of the treatments (Fig. 2B and 2C), there was a reduction in the intrinsic rate of population increase (r) of this clone as well as in the other clones (Figure 2D). Again, the clone D. laevis (RD) was most affected, with reductions of r-value in the order of 21 to 33%.
3.2. Dietary exposure to STX-producing cyanobacteria under variable nutritional food
The analysis detected STX concentrations ranging from 5.0 to 15.2 ng L-1 (Table 1). As in the previous life table assay with fixed nutritious food, D. laevis clones showed no reduction in survival. On the other hand, D. gessneri experienced a reduction in survival over the 15 days of exposure, with a concentration-dependent decline in the number of individuals and mortality of all individuals in the treatment 90% R. raciborskii from day 11 and the treatment 50% on day 14. The LC50 and EC50 values can be seen in Table 4.
For D. gessneri, a dietary exposure to 25% of cyanobacteria caused a 50% reduction in survivorship, while about 56% cyanobacteria in the diet caused a 50% reduction in the production of offspring (Table 4). For both D. laevis clones, this effect on reproduction was caused at a higher proportion of cyanobacteria in the diet (81 and 89%, respectively; Table 4).
A significant reduction in somatic growth rate was observed in the treatment with 90% cyanobacteria in the diet (0.9 mg C L-1) for D. laevis (RD) and 50% and 90% (0.5 and 0.9 mg C L-1) for D. laevis (IB) (p>0.0001) (Figure 3). It was not possible to calculate the growth rate for D. gessneri once all organisms died in the treatments with cyanobacteria prior to end of the life table experiment.
Overall, the two-way ANOVA revealed a significant effect of species/clone and treatment only for fecundity and also a significant interaction between those factors (Table 5). The other parameters showed significant effect only for treatment, besides a interaction between those factors (Table 5).
Population parameters as well as somatic growth rate were negatively affected only at 90% R. raciborskii for all clones, with D. gessneri presenting a greater impact in the total offspring production and in the intrinsic rate of population increase (r-values) (Figure 4). Total offspring likewise was significantly reduced only at the 90% treatment for D. laevis (RD; p<0.001) and D. laevis (IB; p=0.006), and for D. gessneri p<0.001 there was a more drastic reduction in total offspring, being significant also in 25% and 50% R. raciborskii treatments (Fig. 4C). The intrinsic rate of population increase (r) was indeed negative for D. gessneri on the 90% cyanobacterial diet (p<0.05), due to the reduced number of neonates produced per female (only one female reproduced in this treatment) (Figure 4D).
3.3. Cyanobacterial effects on feeding behavior
The animals showed a significant reduction in the total clearance rate (CR) in the 90% cyanobacterial diet only for D. laevis (IB clone) (Fig. 5A). However, when green algae (M. capricornutum) clearance rate alone was analyzed separately, a significant reduction occurred with negative clearance rate for both D. laevis clones in the 90% treatment (Fig. 5B). D. gessneri had a totally different behavior, with an increase in clearance rate of M. capricornutum with increasing concentrations of cyanobacteria (Fig. 5B). When only the cyanobacterial signal was analyzed, with the exception of the D. laevis (RD) clone, clearance rates were in general lower than the green algae. However, significant reductions were observed only in 50% for D. laevis (IB) and in 90% cyanobacterial diet for D. gessneri (Figure 5C).
No cyanobacterial signal was detected in the 25% treatment for D. laevis (RD) and D. gessneri so it was not possible to quantify the cyanobacterial clearance rate at this concentration for those species.
4. Discussion
The results corroborated our main hypothesis that toxicity of STXs-producing R. raciborskii impairs Daphnia feeding behavior and fitness regardless of the high availability of nutritious algae. This effect was, however, species/clone dependent. In both life table assays, there was a reduction in the somatic growth and population increase rates of all species exposed to cyanobacterial concentrations relative to the control. Probably, the energetic cost invested in survival may have decreased the fitness of these animals, causing reduced fecundity and growth. The quality of food is determined by a variety of substances in natural environments. Cyanobacteria are generally considered a poor food due to lack of specific micronutrients, such as polyunsaturated fatty acids and sterols (Martin-Creuzburg et al., 2008; Von Elert et al., 2003). If diets are deficient in essential substances necessary for growth, such as sterols and fatty acids, growth rates may be affected (Sarrazin & Sperfeld, 2022). However, species/clones may respond differently to the variable diets, according to their sensitivity and nutritional requirements (Fileto et al., 2007; Ferrão-Filho et al., 2019).
The two experimental designs had some important differences that should be pointed out. In the first one we had the purpose to exclude que nutritional effect by providing a non-limiting amount of nutritious food (green algae) at a constant ratio while varying the concentration of cyanobacteria (0 to 1.5 mg C L-1), thus the total food concentration varied (from 0.4 to 1.9 mg C L-1) and so the proportion of nutritious food in the diet varied (100 to 21%). The second one was performed with variable proportions between cyanobacteria and green algae so the total food concentration remained constant at 1.0 mg C L-1. Therefore, the equivalent concentration of cyanobacteria in both assays do not represent the same proportion of the total food in each assay. For example, at 1.0 mg C L-1 of cyanobacteria in assay 1 there was ~30% of nutritious food, while at 0.9 mg C L-1 in assay 2 there was only 10% of nutritious food (see Table 1). Thus, at equivalent cyanobacterial concentrations, nutritional restriction was much higher in assay 2.
The negative effects on reproductive parameters and somatic growth of D. laevis (RD) were more pronounced in the experiment without nutritional restriction. Therefore, only the low nutritional value of cyanobacteria does not explain the reduction in fecundity and somatic growth, since these organisms received a sufficient amount of food with high nutritional quality, probably evidencing a toxic effect due to the presence of STX. Also, the fact that at equivalent concentrations of cyanobacteria there was a higher concentration of nutritious food in assay 1 than in assay 2 corroborates this hypothesis. In a previous study, a D. laevis clone isolated from Lagoa da Pampulha-MG showed reduced fecundity when chronically exposed to CYRF-1 strain (STXs producer) and NPCS-1 strain (non-STXs producer), both strains of the species R. raciborskii. However, the effects of the CYRF-1 strain were more pronounced than those of the NPCS-1 strain, showing that besides toxicity, nutritional constraints must play a role on the reduced fecundity (Restani & Fonseca, 2014).
On the other hand, in the life table assay with variable nutritional food availability, D. laevis clones did not show much difference in reproductive parameters, both being equally affected only in high share of cyanobacteria (90%). At lower share of R. raciborskii (25 – 50%) D. laevis seems to cope with the presence of this poor food resource, showing good growth rates. This species can coexist for long periods with cyanobacterial blooms, but its total lipid reserves decrease (Pinto-Coelho et al., 2003), which could explain the reduced fitness of these animals when they were exposed to treatments with higher proportions of cyanobacteria. In studies with cylindrospermopsin-producing (R. raciborskii and Aphanizomenon ovalisporum) and non-producing (R. raciborskii) strains, effects on survival and somatic growth of D. magna, as well as damage to the digestive epithelium were also observed (Nogueira et al., 2006).
Among the species, D. gessneri showed the most contrasting responses between the two life table assays performed. While in the assay without nutritional restriction D. gessneri was the most tolerant among the species tested, in the trial with the nutritional restriction it suffered the most deleterious effects on reproduction, having extremely low fecundity, total offspring production and negative rate of population increase in the highest proportion (90%) of cyanobacteria in the diet.
Regarding survival, in both life table assays, the results did not show resistance of D. gessneri as in previous studies where this species showed high survival. Using the R. raciborskii T3 strain (STXs producer), authors found that it did not interfere with the survival or reproduction of D. gessneri, while the cladocerans D. pulex and M. micrura, had reductions in survivorship and population increase rate (Costa et al., 2013). Therefore, it is possible that D. gessneri is more tolerant to toxins and sensitive to the lack of nutritious food.
Using mixtures of cyanobacteria and green algae (Scenedesmus obliquus), effects of the CYRF-1 strain on D. magna, such as reductions in survival, body growth, population growth rate, and clearance rate, have been reported (Soares et al., 2009). As for D. laevis in our experiment, a proportion of 1:1 (50:50%) between green algae and cyanobacteria was enough to reverse the effects of CYRF strain on D. magna. In an assay with D. pulicaria being exposed to treatments containing S. obliquus or filaments of a non-toxic strain of R. raciborskii, the presence of a non-toxic strain affected the reproductive success of Daphnia, not only through reduced fecundity but also through an increase in the number of aborted eggs (Bednarska & Slusarczyk, 2013). This study, as well as that of Restani and Fonseca (2014) with the non-toxic strain NPCS-1, reinforce the fact that the observed effects on reproduction are not explained only by the toxins and nutritional issues are involved.
In a study with another R. raciborskii strain (CYLCAM-2) isolated from the same reservoir and with the same experimental design of our study, but with more limited food condition (total food = 200 g C L-1), the four cladocerans tested showed decreased fitness even in the lower proportion of cyanobacteria (25%), with differences in sensitivity between species/clones (Ferrão-Filho et al., 2019). Apparently, D. laevis (IB clone) was one of the most resistant species being more affected only at the highest proportion of cyanobacteria in the diet (100%). Thus, this clone seems to be more affected by nutritional deficiency than toxicity. In the present study, D. gessneri showed to be very sensitive to lack of nutritious food, having its fitness severely depressed in the highest proportion of cyanobacteria (90% of the diet).
The grazing assay showed decreased total clearance rate only for D. laevis (IB) clone when exposed to a diet with cyanobacteria. When the clearance rate of green algae was analyzed, there was a significant reduction for both D. laevis clones. Probably, those clones reduced its clearance rate due to the presence of toxins, poor nutritional quality, and/or morphology of R. raciborskii. In a previous study, D. magna, even presenting feeding inhibition, continued with a positive population increase rate (r), indicating that these organisms can thrive in eutrophic environments dominated by cyanobacteria (Soares et al., 2009). Similarly, it is possible that despite the reduction in clearance rate D. laevis maintained its fitness, since in the life table assay with variable food availability both clones were less affected in reproductive parameters. Curiously, the clearance rate of cyanobacteria for the clone RD was higher in the 90% treatment, showing that this clone is capable of ingesting high amounts of cyanobacterial filaments. This may have compensated the decreased ingestion of green algae.
It has been suggested that filaments (with an average length of 426.7 ± 208.4 µm in this study) could impair the ingestion of food by Daphnia (Wilson et al., 2006; Panosso & Lürling, 2010; Bednarska et al., 2014). Although the D. laevis clones are medium-sized (1.8 mm), ingestion of large filaments has been observed in other studies in cladocerans of similar size. D. pulex (0.6 mm) when exposed to STXs-producing strains, was significantly more affected in its clearance rate by ingesting long filaments of MVCC19 (863.5 ± 483.3 µm) and CYRF-01 (338.7 ± 190.3 µm) than the copepod Notodiaptomus iheringi (1.1 mm) (Fabre et al., 2017). Copepods seem to cope better with the presence of filamentous cyanobacteria (Ger et al., 2011). They are able to cut long filaments (Bouvy et al., 2000) and can avoid toxic strains (Panosso et al., 2003). Cladocerans, however, are generalists filter-feeders and filamentous cyanobacteria can clog their filtering apparatus (DeMott et al., 2001). Nevertheless, D. laevis (RD) seems to ingest the filaments of R. raciborskii, becoming more exposed to the toxins of the strain,. This is the reason it may have been more affected in the life table experiment with no food restriction. In addition, toxin production of R. raciborskii can also reduce the fitness of copepods, especially when exposed to nutritionally restricted diets, indicating that toxins interfere more with herbivory than filament size (Rangel et al., 2016; 2020).
In the grazing assay, D. gessneri showed a tendency to increase the total clearance rate in the treatments with cyanobacteria, which may be attributed to an increase in the ingestion of green algae. By the other hand, this species showed a reduced clearance rate of cyanobacteria in the 50% treatment, but a comparable rate to green algae in the 90% treatment, which may explain its reduced fitness, as it may be able to ingest the poor food in an environment with low nutritious food availability. DeMott (1999) has shown that Daphnia spp. exposed to toxic Microcystis exhibited inhibition of feeding after 1 h of exposure to toxic food, but had substantially recovered after 24 h in the same mixtures (“hunger effect”). Thus, it is possible that D. gessneri elevated its feeding rate in 90% cyanobacterial diet to decrease the risk of starvation, as the availability of nutritious food was too low.
Some studies have already demonstrated selectivity in cladocerans. Although in general they are considered non-selective filter-feeders, experiments with D. ambigua revealed that they can avoid toxic strains of M. aeruginosa (Tillmanns et al., 2011). Using video recording, D. pulicaria was able to discriminate between toxic Microcystis strains and the green algae Scenedesmus, slowing its mandible movement in the presence of this cyanobacterium (Ghadouani et al., 2004). Higher clearance rates of the green algae S. capricornutum were observed in treatments with a mixture of Microcystis aeruginosa (MIRF-1) and R. raciborskii (CYRF-1) than in all other treatments for D. laevis and D. similis, suggesting no feeding inhibition of these cladocerans and perhaps selective clearance (Ferrão-Filho et al., 2017). Therefore, the possibility that D. gessneri has the ability to discriminate toxic cyanobacteria at the same time that it increases it is clearance rate of green algae cannot be discarded.
Differences between Daphnia clones may be dependent on the genotype and life history of these organisms (Tillmanns et al., 2008; Bednarska et al., 2014). The D. gessneri clone was isolated from the Apertado Reservoir, located in Mucugê (BA), where there are no reported occurrences of cyanobacterial blooms. The D. laevis clone (IB) was isolated from Ibirité Lagoon (MG), where phytoplankton is dominated by Microcystis spp. (Garcia et al., 2009). The D. laevis clone (RD) was isolated from Lake Jacaré (Rio Doce State Park, MG) where R. raciborskii has already been reported to occur (Miranda et al., 2013). Cyanobacteria dominating environments for long periods can act as a natural selection force on Daphnia populations, with genotypes with higher growth and reproduction rates being favored (Mello & Maia-Barbosa, 2015). Thus, differences in sensitivity among Daphnia clones to cyanobacteria may be a selective factor, determining the composition of zooplankton, favoring the competitive ability of some tolerant species over other sensitive species (Ferrão-Filho et al., 2007).
In conclusion, we found adverse effects of toxic R. raciborskii CYLCAM-1 on Daphnia fitness, even when exposed to a diet without food restriction. Particularly, in the life table assay with constant nutritious food, a reduction in the reproduction parameters for all species was observed, evidencing that the toxic effect of CYLCAM-1 overrides the nutritional effect. In the assay with variable nutritious food, D. gessneri presented as the most sensitive species in the diet with the highest proportion of cyanobacteria, which suggests a strongest nutritional effect in that clone. Daphnia clones that showed a high clearance rate (i.e. continued feeding even in diets with high cyanobacterial ratios) appeared to be less tolerant to exposure to CYLCAM-01. While clones with low clearance rate that reduced their clearance rate in the presence of cyanobacteria, were more tolerant and seem to avoid ingesting filaments, being able to coexist with R. raciborskii in conditions where dominance of this cyanobacteria occurs.
In a scenario with climate change and increased dominance of cyanobacteria, the zooplankton community may suffer alterations of its composition due to different levels of sensitivity among organisms. In a natural environment, reduced predation of zooplankton on phytoplankton may favor the dominance of toxic cyanobacteria, causing the extinction of sensitive species and reduction in the diversity of a natural ecosystem. Studies investigating behavioral resistance (such as rejecting filaments) or physiological tolerance to STX-producing strains are needed for more elucidation on the feeding selectivity and ecophysiology of different Daphnia clones.
Author Contributions
Conceptualization, Aloysio Ferrão-Filho; Funding acquisition, Sandra Azevedo; Investigation, Gabriele dos Reis; Methodology, Mauro Vilar; Supervision, Aloysio Ferrão-Filho; Writing – original draft, Gabriele dos Reis; Writing – review & editing, Mauro Vilar, Sandra Azevedo and Aloysio Ferrão-Filho.
Funding
CNPq and CAPES.
Acknowledgments
We thank CAPES for the first author fellowship and CNPq for financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- AFNOR - Association Française of Normalization (1980). Essais des eaux. Determination of inhibition of Scenedesmus subspicatus par une substance. Norme Experimentale T90-304.
- Aguilera, A.; Gómez, E.B.; Kaštovský, J.; Echenique, R.O.; Salerno, G.L. The polyphasic analysis of two native Raphidiopsis isolates supports the unification of the genera Raphidiopsis and Cylindrospermopsis (Nostocales, Cyanobacteria). Phycologia 2018, 57, 130–146. [Google Scholar] [CrossRef]
- Antunes, J.T.; Leã£O, P.N.; Vasconcelos, V.M. Cylindrospermopsis raciborskii: review of the distribution, phylogeography, and ecophysiology of a global invasive species. Front. Microbiol. 2015, 6, 473. [Google Scholar] [CrossRef] [PubMed]
- Bednarska, A.; Pietrzak, B.; Pijanowska, J. Effect of poor manageability and low nutritional value of cyanobacteria on Daphnia magna life history performance. J Plankton Res 2014, 36, 838–847. [Google Scholar] [CrossRef]
- Bednarska, A.; Slusarczyk, M. Effect of non-toxic, filamentous cyanobacteria on egg abortion in Daphnia under various thermal conditions. Hydrobiologia 2012, 715, 151–157. [Google Scholar] [CrossRef]
- Bonilla, S.; Aubriot, L.; Soares, M.C.S.; et al. What drives the distribution of the bloom-forming cyanobacteria Planktothrix agardhii and Cylindrospermopsis raciborskii? FEMS Microbiol Ecol 2012, 79, 594–607. [Google Scholar] [CrossRef]
- Bouvy, M.; Falcão, D.; Marinho, M.; Pagano, M.; Moura, A. Occurrence of Cylindrospermopsis (Cyanobacteria) in 39 Brazilian tropical reservoirs during the 1998 drought. Aquat. Microb. Ecol. 2000, 23, 13–27. [Google Scholar] [CrossRef]
- Bouvy, M.; Molica, R.; Oliveira, S.; Marinho, M.; Becker, B. Dynamics of a toxic cyanobacterial bloom (Cylindrospermopsis raciborskii) in a shallow reservoir in the semi-arid region of Northeast Brazil. Aquat. Microb. Ecol. 1999, 20, 285–297. [Google Scholar] [CrossRef]
- Bunioto, T.C.; Arcifa, M.S. Effects of food limitation and temperature on cladocerans from a tropical Brazilian lake. Aquat Ecol 2007, 41, 569–578. [Google Scholar] [CrossRef]
- Burford, M.A.; Beardall, J.; Willis, A.; Orr, P.T.; Magalhaes, V.F.; Rangel, L.M.; Azevedo, S.M.F.O.E.; Neilan, B.A. Understanding the winning strategies used by the bloom-forming cyanobacterium Cylindrospermopsis raciborskii. Harmful Algae 2016, 54, 44–53. [Google Scholar] [CrossRef]
- Costa, S.M.; Ferrão-Filho, A.S.; Azevedo, S.M.F.O. Effects of saxitoxin- and non-saxitoxin-producing strains of the cyanobacterium Cylindrospermopsis raciborskii on the fitness of temperate and tropical cladocerans. Harmful Algae 2013, 28, 55–63. [Google Scholar] [CrossRef]
- Charles S, Veber P, Delignette-Muller ML MOSAIC: a web-interface for statistical analyses in ecotoxicology. Env. Sci Pollut Res. 2018, 25, 11295. [CrossRef] [PubMed]
- Chorus I, Bartram J. Toxic cyanobacteria in water: a guide to their public health consequences, monitoring and management, 3rd ed.; WHO - World Health Organization: Geneva, 1999. [Google Scholar]
- DeMott, W.R. Foraging strategies and growth inhibition in five daphnids feeding on mixtures of a toxic cyanobacterium and a green alga. Freshwat Biol 1999, 42, 263–274. [Google Scholar] [CrossRef]
- DeMott, W.R.; Van Donk, E. Strong interactions between stoichiometric constraints and algal defenses: evidence from population dynamics of Daphnia and algae in phosphorus-limited microcosms. Oecologia 2013, 171, 175–186. [Google Scholar] [CrossRef] [PubMed]
- DeMott, W.R.; Gulati, R.D.; Van Donk, E. Daphnia food limitation in three hypereutrophic Dutch lakes: Evidence for exclusion of large-bodied species by interfering filaments of cyanobacteria. Limnol Ocean. 2001, 46, 2054–2060. [Google Scholar] [CrossRef]
- Fabre, A.; Lacerot, G.; de Paiva, R.R.; Soares, M.C.S.; Magalhães, V.F.; Bonilla, S. South American PSP toxin-producing Cylindrospermopsis raciborskii (Cyanobacteria) decreases clearance rates of cladocerans more than copepods. Hydrobiologia 2017, 785, 61. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.S. Bioacumulação de cianotoxinas e seus efeitos em organismos aquáticos. Oecol. Bras. 2009, 13, 272–312. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.S.; Abreu, S.S.D.; Oliveira, T.; Magalhães, V.F.; Pflugmacher, S.; Silva, E.M. Single and combined effects of microcystin and saxitoxin producing cyanobacteria on the fitness and antioxidant defenses of cladocerans. Environ. Toxicol. Chem. 2017, 9999, 1–9. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.S.; Costa, S.M.; Ribeiro, M.G.L.; Azevedo, S.M.F.O. Effects of a saxitoxin-producer strain of Cylindrospermopsis raciborskii (cyanobacteria) on the swimming movements of cladocerans. Environ. Toxicol. 2008, 23, 161–168. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.S.; Cunha, R.; Magalhães, V.F.; Soares, M.C.S.; Baptista, D.F. Evaluation of sub-lethal toxicity of Cyanobacteria on the swimming activity of aquatic organisms by image analysis. J. Braz. Soc. Ecotoxicol. 2007, 2, 93–99. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.S.; Dias, T.M.; Pereira, U.J.; dos Santos, J.A.A.; Kozlowsky-Suzuki, B. Nutritional and toxicity constraints of phytoplankton from a Brazilian reservoir to the fitness of cladoceran species. Environ. Sci. Pollut. Res. 2019, 26, 12881–12893. [Google Scholar] [CrossRef]
- Ferrão-Filho AS, Koslowsky-Suzuki B Cyanotoxins: Bioaccumulation and Effects on Aquatic Animals. Mar. Drugs 2011, 9, 2729–2772. [CrossRef]
- Ferrão-Filho, Soares MC, Magalhães VF, Azevedo SM. A rapid bioassay for detecting saxitoxins using a Daphnia acute toxicity test. Environ. Pollut. 2010, 158, 2084–2093. [Google Scholar] [CrossRef] [PubMed]
- Ferrão-Filho AS, Soares MCS, Lima RS, Magalhães VF Effects of Cylindrospermopsis raciborskii (Cyanobacteria) on the swimming behavior of Daphnia (Cladocera). Envirn Toxicol Chem 2014, 31, 223–229.
- Garcia FC, Barbosa FAR, Braz S, Petrucio MM, Faria B Water quality of an urban reservoir subjected to periodic applications of copper sulphate: the case of Ibirité reservoir, Southeast Brazil. Acta Limnol Bras 2009, 21, 235–243.
- Ghadouani, A.; Pinel-Alloul, B.; Plath, K. Effects of Microcystis aeruginosa and purified microcystin-LR on the feeding behavior of Daphnia pulicaria. Limnol Ocean. 2004, 49, 666–679. [Google Scholar] [CrossRef]
- Ger, K.A.; Urrutia-Cordero, P.; Frost, P.C.; Hansson, L.-A.; Sarnelle, O.; Wilson, A.E.; Lürling, M. The interaction between cyanobacteria and zooplankton in a more eutrophic world. Harmful Algae 2016, 54, 128–144. [Google Scholar] [CrossRef]
- Ger, K.A.; Panosso, R.; Lürling, M. Consequences of acclimation to Microcystis on the selective feeding behavior of the calanoid copepod Eudiaptomus gracilis. Limnol. Oceanogr. 2011, 56, 2103–2114. [Google Scholar] [CrossRef]
- Gliwicz, Z.M. Food thresholds and body size in cladocerans. Nature 1990, 343, 638–640. [Google Scholar] [CrossRef]
- Guillard, R.R. Cultures of phytoplankton for feeding of marine invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum: New York, 1975; pp. 29–60. [Google Scholar]
- Hansson, L.; Gustafsson, S.; Rengefors, K.; Bomark, L. Cyanobacterial chemical warfare affects zooplankton community composition. Freshw. Biol. 2007, 52, 1290–1301. [Google Scholar] [CrossRef]
- Hardy, E. R.; Duncan, A. Food concentration and temperature effects on life-cycle characteristics of tropical Cladocera Daphnia gessneri Herbst, Diaphanosoma sarsi Richard, and Moina reticulata Daday. I. Development time. Acta Amaz. 1994, 24, 119–134. [Google Scholar] [CrossRef]
- Hart, R.C.; Bychek, E.A. Body syze in freshwater planktonic crustaceans: an overview of extrinsic determinants and modifying influences of bioctic interactions. Hydrobiologia 2011, 668, 61–108. [Google Scholar] [CrossRef]
- Havens, K.E.; Hudnell, H.K. Cyanobacteria blooms: effects on aquatic ecosystems, Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Springer: New York, 2008; pp. 733–747. [Google Scholar]
- Hawkins, P.R.; Chandrasena, N.R.; Jones, G.J.; Humpage, A.R.; Falconer, I.R. Isolation and toxicity of Cylindrospermopsis raciborskii from an ornamental lake. Toxicon 1997, 35, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Heathcote, A.J.; Filstrup, C.T.; Kendall, D.; Downing, J.A. Biomass pyramids in lake plankton: influence of Cyanobacteria size and abundance. Inland Waters 2016, 6. [Google Scholar] [CrossRef]
- Hillebrand, H.; Dürselen, C.D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 2002, 35, 403–424. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Huszar, V.L.M.; Silva, L.H.S.; Marinho, M.M.; Domingos, P.; Sant’Anna, C.L. Cyanoprokaryote assemblages in eight productive tropical Brazilian waters. Hydrobiologia 2000, 424, 67–77. [Google Scholar] [CrossRef]
- Jónasdóttir, S.H.; Kiørboe, T.; Tang, K.W.; St John, M.; Visser, A.W.; Saiz, E.; Dam, H.G. Role of diatoms in copepod production: good, harmlessor toxic? Mar Ecol Prog Ser 1998, 172, 305–308. [Google Scholar] [CrossRef]
- Lagos, N.; Onodera, H.; Zagatto, P.A.; Andrinolo, D.; Azevedo, S.M.F.O.; Oshima, Y. The first evidence of paralytic shelfish toxins in the freshwater cyanobacterium Cylindrospermopsis raciborskii, isolated from Brazil. Toxicon 1999, 37, 1359–1373. [Google Scholar] [CrossRef]
- Lei, L.; Peng, L.; Huang, X.; Han, B.P. Occurrence and dominance of Cylindrospermopsis raciborskii and dissolved cylindrospermopsin in urban reservoirs used for drinking water supply. South China, Environ. Monit. 2014.
- Leonard, J.A.; Pearl, H.W. Zooplâncton community structure, micro-zooplâncton grazing impact, and seston energy content in the St. Johns river system, Florida as influenced by the toxic cyanobacterium Cylindrospermopsis raciborskii. Hydrobiologia 2005, 537, 89–97. [Google Scholar] [CrossRef]
- Lürling, M.; Verschoor, A.M. FO-spectra of chlorophyll fluorescence for the determination of zooplankton grazing. Hydrobiologia 2003, 491, 145–157. [Google Scholar] [CrossRef]
- Pagni, R.L.; de Falco, P.B.; dos Santos, A.C.A. Autecology of Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju. Acta Limnol. Bras. 2020, 32. [Google Scholar] [CrossRef]
- Pearl, H.; Huisman, J. Blooms Like It Hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Martin-Creuzburg, D.; von Elert, E.; Hoffmann, K.H. Nutritional constraints at the cyanobacteria-Daphnia magna interface: The role of sterols. Limnol. Oceanogr. 2008, 53, 456–468. [Google Scholar] [CrossRef]
- Mello, N.A.S.T.; Maia-Barbosa, P.M. Cyanobacteria bloom: selective filter for zooplankton? Braz J Biol 2015, 75, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, M.; Lürling, M.; Dorr, F.; Pinto, E.; Marinho, M. Combined Effect of Light and Temperature on the Production of Saxitoxins in Cylindrospermopsis raciborskii Strains. Toxins 2019, 11, 38. [Google Scholar] [CrossRef]
- Miranda, F.S.; Pinto-Coelho, R.M.; Gonzaga, A.V. Redução da riqueza de organismos do zooplâncton (com ênfase em Copepoda e Cladocera) nas lagoas do médio Rio Doce/MG. Rev. Bras. De Zoociências 2013, 15, 69–90. [Google Scholar]
- Miranda, C.T.; de Lima, D.V.N.; Atella, G.C.; de Aguiar, P.F.; Azevedo, S.M.F.O. Optimization of Nitrogen, Phosphorus and Salt for Lipid Accumulation of Microalgae: Towards the Viability of Microalgae Biodiesel. Nat. Sci. 2016, 08, 557–573. [Google Scholar] [CrossRef]
- Molica, R.J.R.; Oliveira, E.J.A.; Carvalho, P.V.V.C.; Costa, A.P.N.S.F.; Cunha, M.C.C.; Melo, G.L. Ocurrence of saxitoxins and anatoxin-a(s)-like anticholinesterase in Brazilian drinking water supply. Harmful Algae 2005, 4, 743–753. [Google Scholar] [CrossRef]
- Moustaka-Gouni, M.; Sommer, U. Effects of Harmful Blooms of Large-Sized and Colonial Cyanobacteria on Aquatic Food Webs. Water 2020, 12, 1587. [Google Scholar] [CrossRef]
- Nogueira, I.C.; Lobo-Da-Cunha, A.; Vasconcelos, V.M. Effects of Cylindrospermopsis raciborskii and Aphanizomenon ovalisporum (cyanobacteria) ingestion on Daphnia magna midgut and associated diverticula epithelium. Aquat. Toxicol. 2006, 80, 194–203. [Google Scholar] [CrossRef]
- Nogueira, I.C.G.; Saker, M.L.; Pflugmacher, S.; Wiegand, C.; Vasconcelos, V.M. Toxicity of the cyanobacteriumCylindrospermopsis raciborskii toDaphnia magna. Environ. Toxicol. 2004, 19, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Panosso, R.; Carlsson, P.; Kozlowsky-Suzuki, B.; Azevedo, S.M.F.O.; Granéli, E. Effect of grazing by a neotropical copepod, Notodiaptomus, on a natural cyanobacterial assemblage and on toxic and non-toxic cyanobacterial strains. J. Plankton Res. 2003, 25, 1169–1175. [Google Scholar] [CrossRef]
- Piccini, C.; Aubriot, L.; Fabre, A.; Amaral, V.; González-Piana, M.; Giani, A.; Figueredo, C.C.; Vidal, L.; Kruk, C.; Bonilla, S. Genetic and eco-physiological differences of South American Cylindrospermopsis raciborskii isolates support the hypothesis of multiple ecotypes. Harmful Algae 2011, 10, 644–653. [Google Scholar] [CrossRef]
- Pinto-Coelho, R.M.; Bezerra-Neto, J.F.; Giane, A.; Macedo, C.F.; Figueiredo, C.C.; Carvalho, E.A. The Collapse of a Daphnia laevis (Birge, 1878) population in Pampulha Resevoir, Brasil. Acta Limnol. Bras. 2003, 3, 53–70. [Google Scholar]
- Pomati, F.; Neilan, B.A.; Suzuki, T.; Manarolla, G.; Rossett, C. Enhancement of intracellular saxitoxin accumulation by lidocaine hydrocloride in the cyanobacterium Cylindrospermopsis raciborskii T3 (Nostocales). J. Phycol. 2003, 39, 535–542. [Google Scholar] [CrossRef]
- Rangel, L.M.; Ger, K.A.; Silva, L.H.S.; Soares, M.C.S.; Faassen, E.J.; Lürling, M. Toxicity Overrides Morphology on Cylindrospermopsis raciborskii Grazing Resistance to the Calanoid Copepod Eudiaptomus gracilis. Microb. Ecol. 2016, 71, 835–844. [Google Scholar] [CrossRef]
- Rangel, L.M.; Silva, L.H.S.; Faassen, E.J.; Lürling, M.; Ger, K.A. Copepod Prey Selection and Grazing Efficiency Mediated by Chemical and Morphological Defensive Traits of Cyanobacteria. Toxins 2020, 12, 465. [Google Scholar] [CrossRef]
- Restani, G.C.; Fonseca, A.L. Effects of Cylindrospermopsis raciborskii strains (Woloszynska, 1912) Senayya & Subba Raju on the mobility of Daphnia laevis (Cladocera, Daphniidae). Braz. J. Biol. 2014, 74, 23–31. [Google Scholar]
- Rzymski, P.; Poniedziałek, B. In search of environmental role of cylindrospermopsin: a review on global distribution and ecology of its producers. Water Res. 2014, 66, 320–327. [Google Scholar] [CrossRef]
- Rocha, O.; Duncan, A. The relationship between cell carbono and cell volume in freshwater algal species used in zooplanktonic studies. J Plankton Res. 1985, 7, 279–294. [Google Scholar] [CrossRef]
- Sarnelle, O.; Gustafsson, S.; Hansson, L.A. Effects of cyanobacteria on fitness components of the herbivore Daphnia. J. Plankton Res. 2010, 32, 471–477. [Google Scholar] [CrossRef]
- Sikora, A.; Dawidowicz, P. Breakage of cyanobacterial filaments by small- and large-sized Daphnia: are there any temperature dependent differences? Hydrobiologia 2017, 798, 119–126. [Google Scholar] [CrossRef]
- Soares, M.C.S.; Lürling, M.; Panosso, R.; Huszar, V. Effects of the cyanobacterium Cylindrospermopsis raciborskii on feeding and life history characteristics of the grazer Daphnia magna. Ecotoxicol. Environ. 2009, 72, 1183–1189. [Google Scholar] [CrossRef]
- Soares, M.C.S.; Huszar, V.; Miranda, M.N.; Melo, M.M.; Roland, F.; Lürling, M. Cyanobacterial dominance in Brazil: distribution and environmental preferences. Hydrobiologia 2013, 717, 1–12. [Google Scholar] [CrossRef]
- Sun, J.; Liu, D. Geometric models for calculating cell biovolume and surface area for phytoplankton. J. Plankton Res. 2003, 25, 1331–1346. [Google Scholar] [CrossRef]
- Taberner, A.; Castañera, P.; Silvertre, E.; Dopazo, J. Estimation of the intrinsic rate of natural increase and its error by both algebraic and resampling Approaches. Comput. Appl. Biosci. 1993, 9, 535–540. [Google Scholar] [CrossRef]
- Tillmanns, A.R.; Burton, S.K.; Pick, F.R. Daphnia Pre-Exposed to Toxic Microcystis Exhibit Feeding Selectivity. Hydrobiology 2011, 96, 20–28. [Google Scholar] [CrossRef]
- Tillmanns, A.R.; Wilson, A.E.; Pick, F.R.; Sarnelle, O. Meta-analysis of cyanobacterial effects on zooplankton population growth rate: species-specific responses. Hydrobiology 2008, 171/4, 285–285. [Google Scholar] [CrossRef]
- Urabe, J.; Shimizu, Y.; Yamaguchi, T. Understanding the stoichiometric limitation of herbivore growth: the importance of feeding and assimilation flexibilities. Ecol. Lett. 2018, 21, 197–206. [Google Scholar] [CrossRef]
- Vilar, MCP. ; Rodrigues, TFCP.; Silva, LO; Pacheco, ABF; Ferrão-Filho, AS; Azevedo, SMFO. Ecophysiological Aspects and sxt Genes Expression Underlying Induced Chemical Defense in STX-Producing Raphidiopsis raciborskii (Cyanobacteria) against the Zooplankter Daphnia gessneri. Toxins 2021, 13, 406. [Google Scholar]
- Vilar, M.C.P.; Ferrão-Filho, A.d.S.; Azevedo, S.M. Single and mixed diets of the toxic Cyanobacteria Microcystis aeruginosa and Raphidiopsis raciborskii differently affect Daphnia feeding behavior. Food Webs 2022, 32. [Google Scholar] [CrossRef]
- Wilson AE, Sarnelle O, Tillmanns AR Effects of cyanobacterial toxicity and morphology on the population growth of freshwater zooplankton: Meta-analyses of laboratory experiments. Limnol. Oceanogr. 2006, 51, 1915–1924. [CrossRef]
- Wood, S.A.; Stirling, D.J. First identification of the cylindrospermopsin-producing cyanobacterium Cylindrospermopsis raciborskii in New Zealand. New Zeal. J. Mar. Fresh. Res. 2003, 37, 821–828. [Google Scholar] [CrossRef]
- Yang, J.R.; Hong, L.V.; Isabwe, A.; Liu, L.; Yu, X.; Chen, H.; Yang, J. Disturbance-induced phytoplankton regime shifts and recovery of cyanobacteria dominance in two subtropical reservoirs. Water Res. 2017, 120, 52–63. [Google Scholar] [CrossRef]
Figure 1.
Somatic growth rate of D. laevis (IB), D. laevis (RD) and D. gessneri in assay 1, exposed to cyanobacteria and without nutritional restriction (constant concentration of green algae at 0.4 mg C L-1). Control refers to a concentration of only green algae (0.4 mg C L-1). Different letters indicate significant differences (ANOVA, Tukey test p<0.05).
Figure 1.
Somatic growth rate of D. laevis (IB), D. laevis (RD) and D. gessneri in assay 1, exposed to cyanobacteria and without nutritional restriction (constant concentration of green algae at 0.4 mg C L-1). Control refers to a concentration of only green algae (0.4 mg C L-1). Different letters indicate significant differences (ANOVA, Tukey test p<0.05).
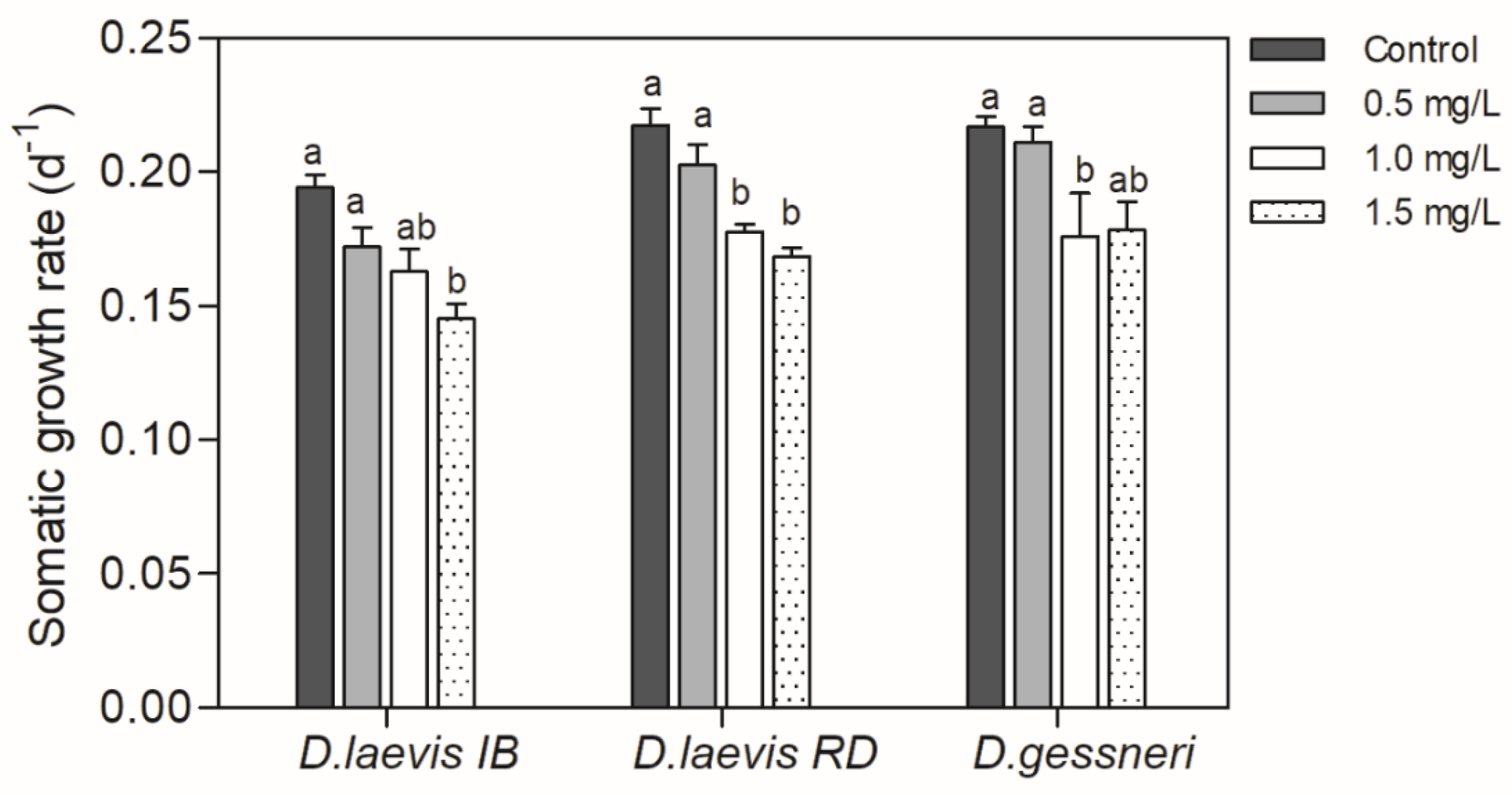
Figure 2.
Population parameters of D. laevis (RD), D. laevis (IB) and D. gessneri in the assay 1, without nutritional restriction. A) Age of first reproduction; B) Fecundity; C) Total offspring; D) Intrinsic rate of population increase (r). Treatments have a constant concentration of green algae (0.4 mg C L-1) and a variable concentration of cyanobacteria. Control refers to a concentration of only green algae (0.4 mg C L-1). Different letters indicate significant differences (ANOVA, Tukey's test, p<0.05).
Figure 2.
Population parameters of D. laevis (RD), D. laevis (IB) and D. gessneri in the assay 1, without nutritional restriction. A) Age of first reproduction; B) Fecundity; C) Total offspring; D) Intrinsic rate of population increase (r). Treatments have a constant concentration of green algae (0.4 mg C L-1) and a variable concentration of cyanobacteria. Control refers to a concentration of only green algae (0.4 mg C L-1). Different letters indicate significant differences (ANOVA, Tukey's test, p<0.05).
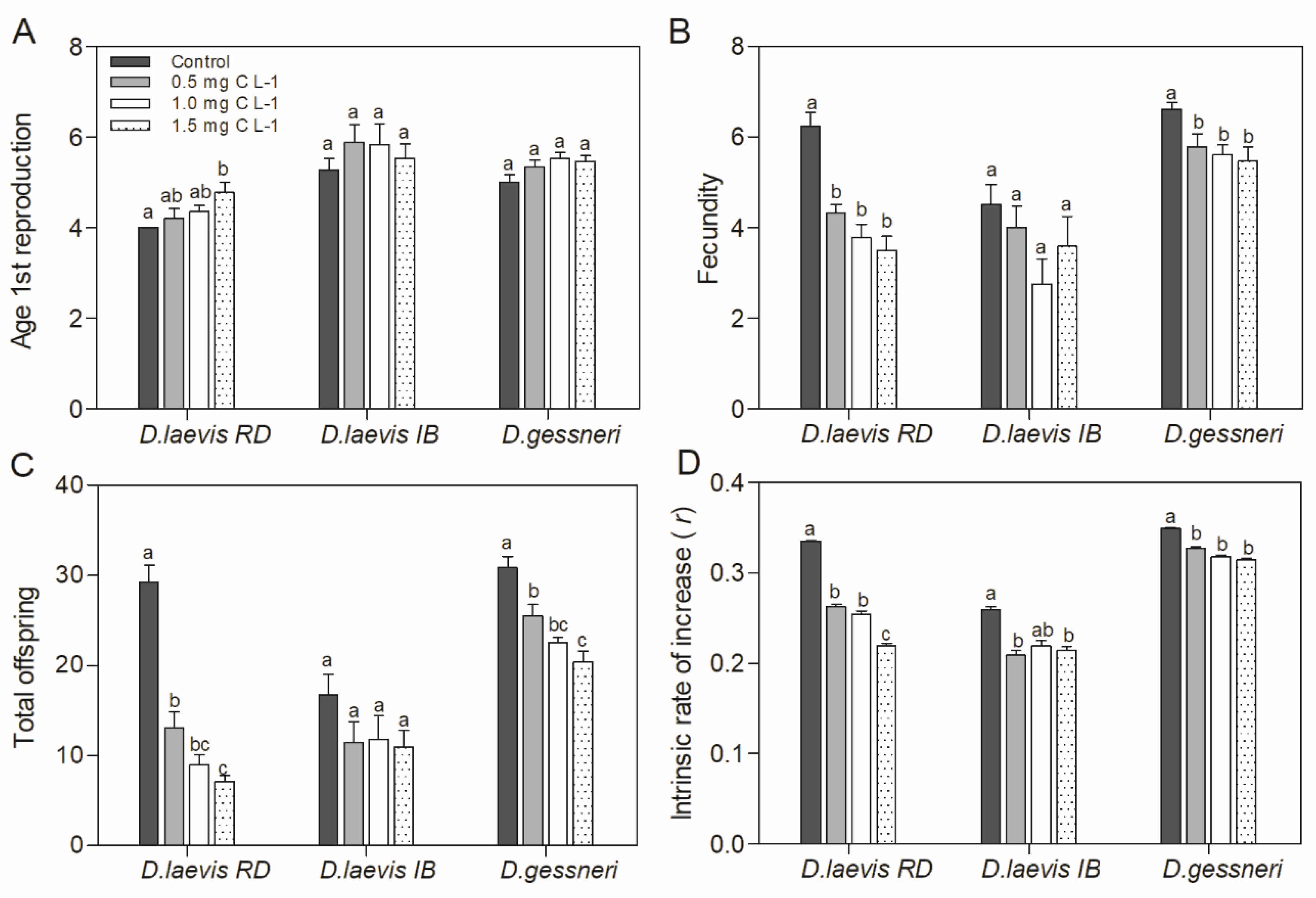
Figure 3.
Somatic growth rate of D. laevis (RD) and D. laevis (IB) in assay 2, exposed to treatments with different proportions of green algae (100-10%) and cyanobacteria (0-90%) at a total concentration of 1.0 mg C L-1. Control refers to 100% green algae. Different letters indicate significant differences (ANOVA, Tukey test p<0.05).
Figure 3.
Somatic growth rate of D. laevis (RD) and D. laevis (IB) in assay 2, exposed to treatments with different proportions of green algae (100-10%) and cyanobacteria (0-90%) at a total concentration of 1.0 mg C L-1. Control refers to 100% green algae. Different letters indicate significant differences (ANOVA, Tukey test p<0.05).
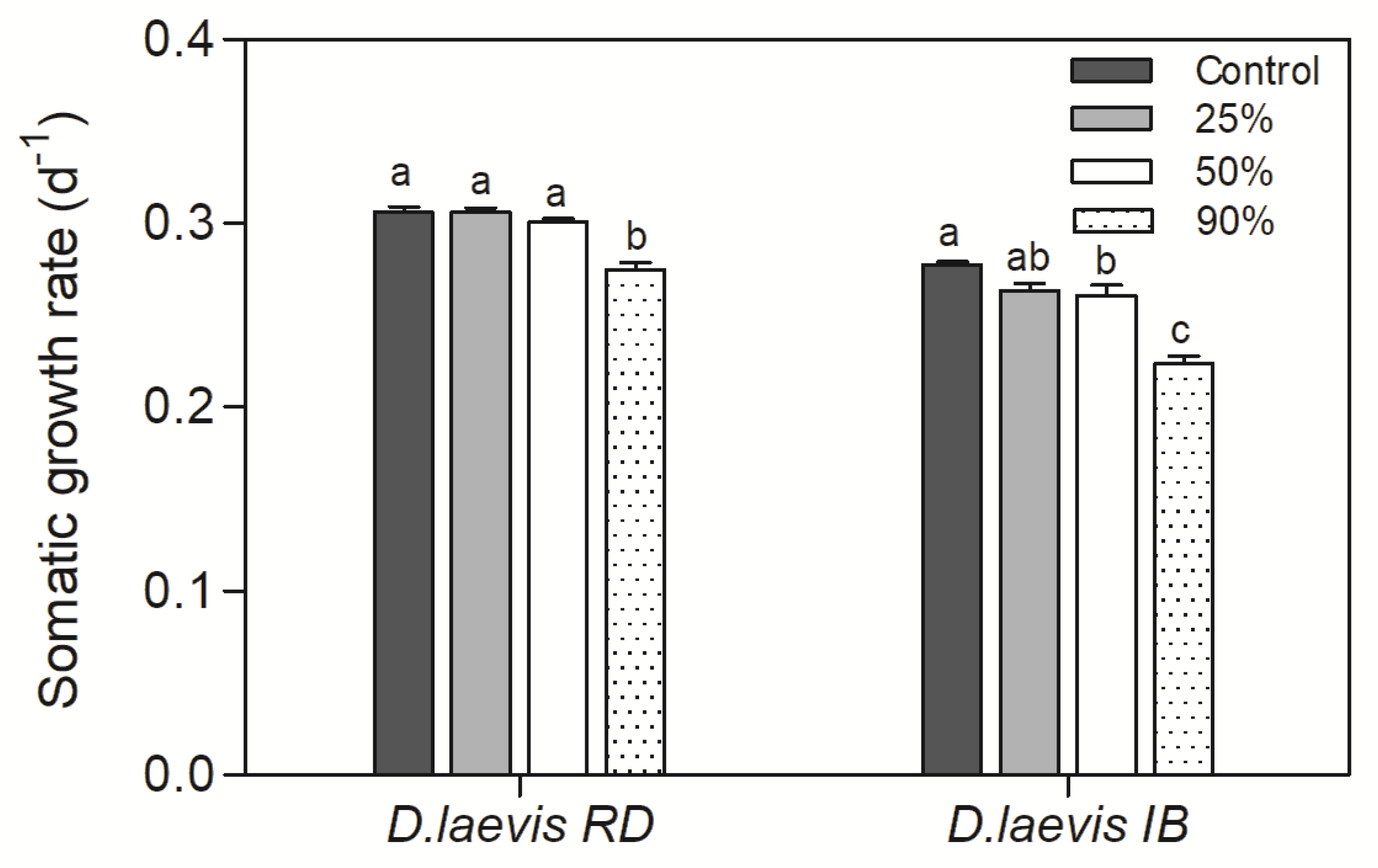
Figure 4.
Population parameters of D. laevis (RD), D. laevis (IB) and D. gessneri in the assay 2, with different proportions of green algae (100-10%) and cyanobacteria (0-90%) at a total concentration of 1.0 mg C L-1. Control refers to 100% green algae. A) Age of first reproduction; B) Fecundity; C) Total offspring; D) Intrinsic rate of population increase (r). Different letters indicate significant differences (ANOVA, Tukey's test, p<0.05).
Figure 4.
Population parameters of D. laevis (RD), D. laevis (IB) and D. gessneri in the assay 2, with different proportions of green algae (100-10%) and cyanobacteria (0-90%) at a total concentration of 1.0 mg C L-1. Control refers to 100% green algae. A) Age of first reproduction; B) Fecundity; C) Total offspring; D) Intrinsic rate of population increase (r). Different letters indicate significant differences (ANOVA, Tukey's test, p<0.05).
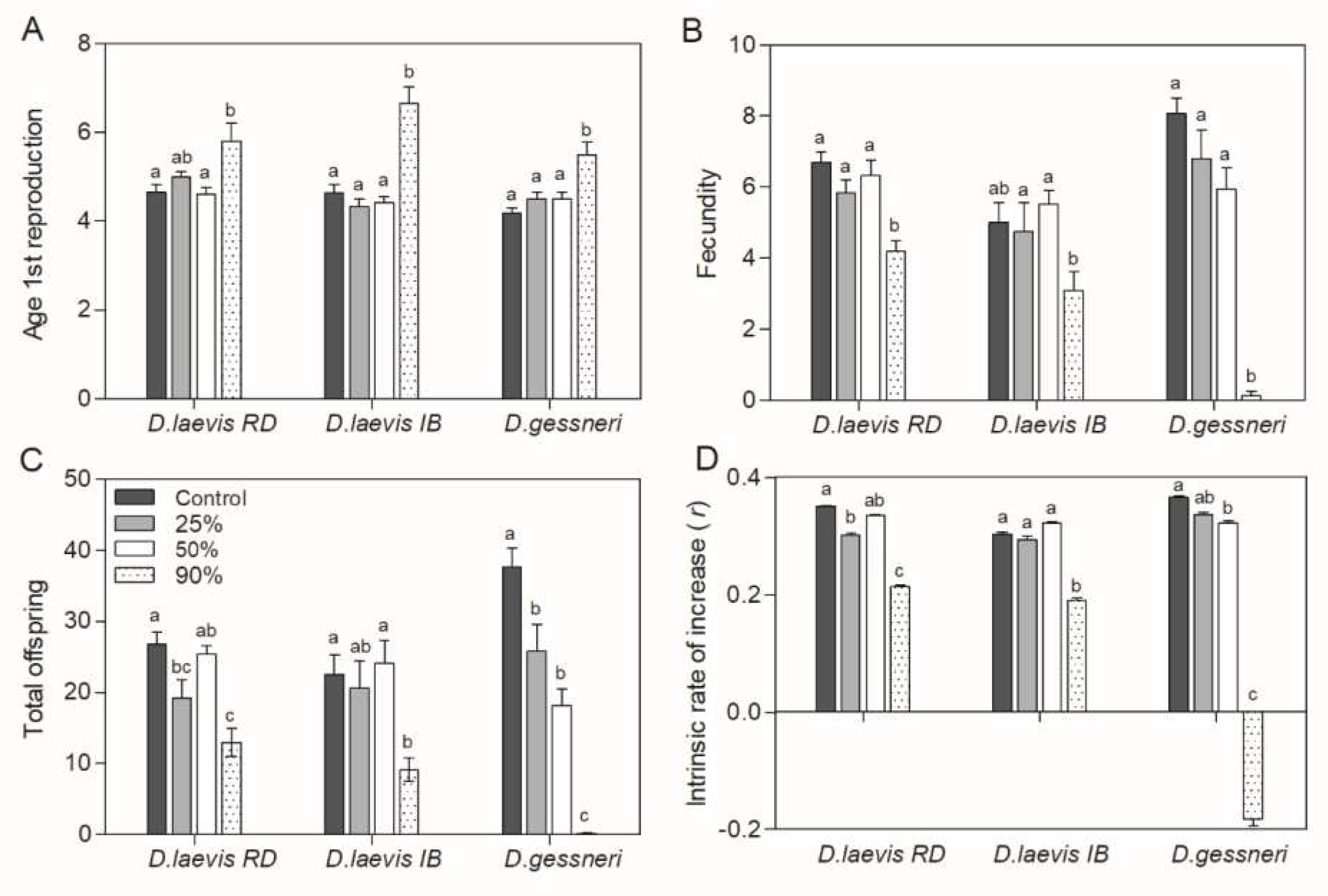
Figure 5.
A. Total clearance rate; B. Clearance rate of M. capricornutum and C. Clearance rate of R. raciborskii of D. laevis (RD), D. laevis (IB) and D. gessneri exposed to treatments with varying proportions of green algae (100-10%) and cyanobacteria (0-90%) at a total concentration of 1.0 mg C L-1. Different letters indicate significant differences (ANOVA, Tukey test p<0.05).
Figure 5.
A. Total clearance rate; B. Clearance rate of M. capricornutum and C. Clearance rate of R. raciborskii of D. laevis (RD), D. laevis (IB) and D. gessneri exposed to treatments with varying proportions of green algae (100-10%) and cyanobacteria (0-90%) at a total concentration of 1.0 mg C L-1. Different letters indicate significant differences (ANOVA, Tukey test p<0.05).
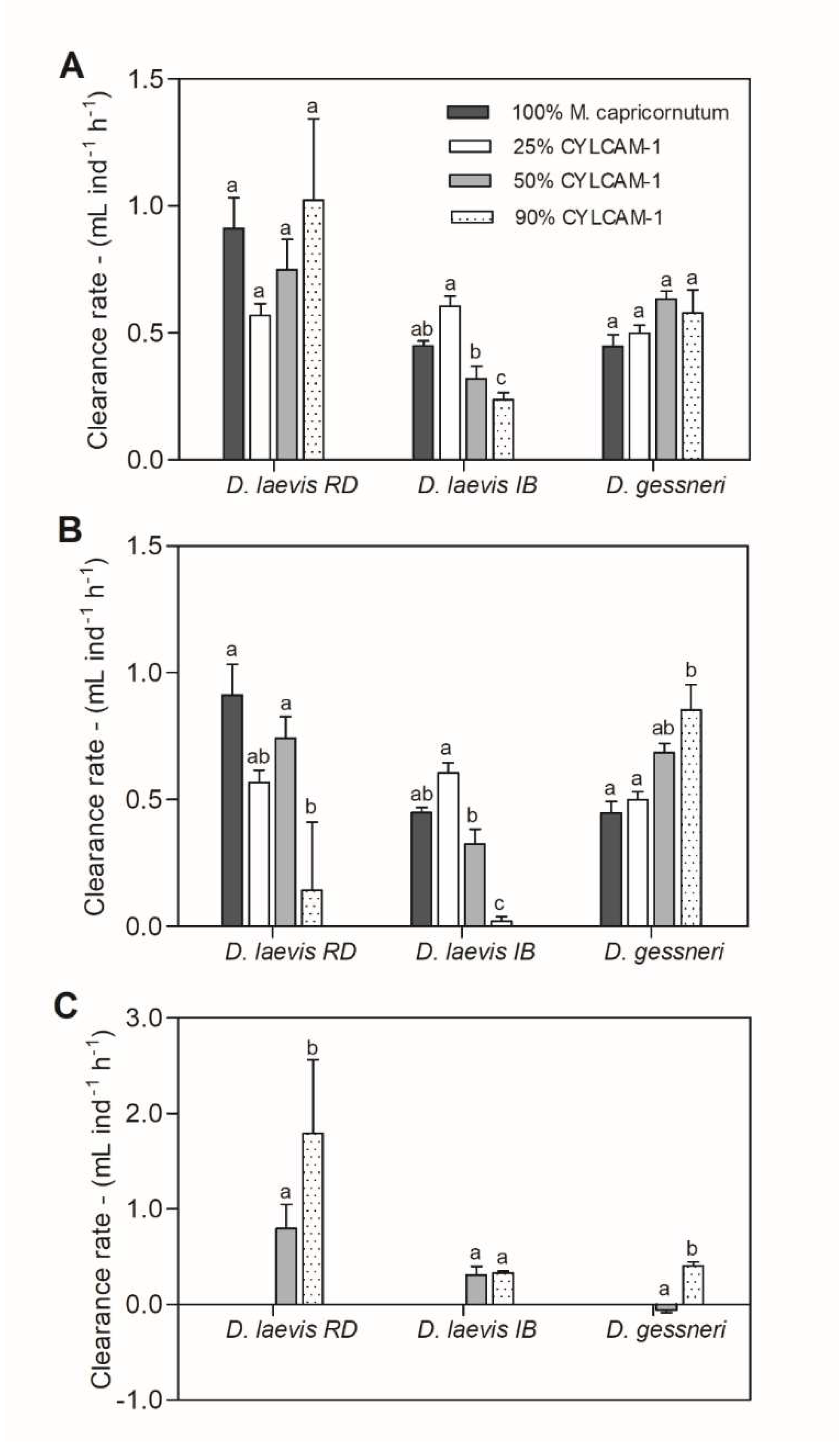

Table 1.
Biomass (as mg C L-1 and %) and STX concentrations in the life table assays without nutritional restriction and with variable proportions of cyanobacteria and nutritious food, and in the grazing assay with clones of D. laevis and D. gessneri.
Table 1.
Biomass (as mg C L-1 and %) and STX concentrations in the life table assays without nutritional restriction and with variable proportions of cyanobacteria and nutritious food, and in the grazing assay with clones of D. laevis and D. gessneri.
| Assay | Date | Cyanobacteria concentration (mg C L-1) | Green algae concentration (mg C L-1) | % nutritious food | STX (ng/L) |
|---|---|---|---|---|---|
| 1. Life table (Constant nutritious food) | 16/05/2017 | 0.00 | 0.40 | 100.0 | --- |
| 0.50 | 0.40 | 44.4 | 1,5 | ||
| 1.00 | 0.40 | 28.6 | 3,0 | ||
| 1.50 | 0.40 | 21.1 | 4,5 | ||
| 2. Life table (Variable nutritious food) | 12/03/2018 | 0.00 | 1.00 | 100.0 | --- |
| 0.25 | 0.75 | 75.0 | 5,0 | ||
| 0.50 | 0.50 | 50.0 | 9,9 | ||
| 0.90 | 0.10 | 10.0 | 15,2 | ||
| 3. Grazing (Variable nutritious food) | 16/02/2018 | 0.00 | 1.00 | 100.0 | --- |
| 0.25 | 0.75 | 75.0 | 7,2 | ||
| 0.50 | 0.50 | 50.0 | 14,4 | ||
| 0.90 | 0.10 | 10.0 | 25,9 | ||
| 4. Grazing (Variable nutritious food) | 03/05/2018 | 0.00 | 1.00 | 100.0 | --- |
| 0.25 | 0.75 | 75.0 | 3,3 | ||
| 0.50 | 0.50 | 50.0 | 6,6 | ||
| 0.90 | 0.10 | 10.0 | 11,9 |
Table 2.
LC50 and EC50 values and 95% confidence intervals of D. laevis (IB), D. laevis (RD) and D. gessneri in the life table assay 1, with constant nutritious food. The symbols (-) indicate that it was not possible to calculate the estimates for the species.
Table 2.
LC50 and EC50 values and 95% confidence intervals of D. laevis (IB), D. laevis (RD) and D. gessneri in the life table assay 1, with constant nutritious food. The symbols (-) indicate that it was not possible to calculate the estimates for the species.
| Species | LC50 (mg C L-1) | EC50 (mg C L-1) |
|---|---|---|
| D. laevis (Ibirité) | - | 1,24 (1,02 - 1,52) |
| D. laevis (Rio Doce) | - | 0,98 (0,82 - 1,21) |
| D. gessneri | 1,33 (0,75 - 1,92) | 1,68 (1,47 - 1,92) |
Table 3.
Results of two-way ANOVA for age at first reproduction, fecundity and total offspring in the assay 1, with constant nutritious food.
Table 3.
Results of two-way ANOVA for age at first reproduction, fecundity and total offspring in the assay 1, with constant nutritious food.
| Factor | df | F | P |
|---|---|---|---|
| Age at first reproduction | |||
| Species | 6 | 35,42 | < 0,0001 |
| Treatment | 3 | 3,32 | 0,02 |
| Species x treatment | 6 | 0,74 | 0,60 |
| Fecundity | |||
| Species | 2 | 37,04 | < 0,001 |
| Treatment | 3 | 15,30 | < 0,001 |
| Species x treatment | 6 | 1,87 | 0,09 |
| Total offspring | |||
| Species | 2 | 67,35 | < 0,001 |
| Treatment | 3 | 15,30 | < 0,001 |
| Species x treatment | 6 | 1,87 | 0,09 |
Table 4.
Estimates of the LC50 and EC50 of D. laevis (IB), D. laevis (RD) and D. gessneri in the life table assay 2, with variable proportions of cyanobacteria and nutritious food. The symbols (-) indicate that it was not possible to calculate the estimates for the species.
Table 4.
Estimates of the LC50 and EC50 of D. laevis (IB), D. laevis (RD) and D. gessneri in the life table assay 2, with variable proportions of cyanobacteria and nutritious food. The symbols (-) indicate that it was not possible to calculate the estimates for the species.
| Species | LC50 (mg C L-1) | EC50 (mg C L-1) |
|---|---|---|
| D. laevis (Ibirité) | - | 0,81 (0,23 - 0,95) |
| D. laevis (Rio Doce) | - | 0,89 (0,76 - 0,97) |
| D. gessneri | 0,25 (0,17 - 0,34) | 0,56 (0,53 - 0,61) |
Table 5.
Results of two-way ANOVA for age at first reproduction, fecundity and total offspring in assay 2, with variable proportions of cyanobacteria and nutritious food.
Table 5.
Results of two-way ANOVA for age at first reproduction, fecundity and total offspring in assay 2, with variable proportions of cyanobacteria and nutritious food.
| Factor | df | F | P |
|---|---|---|---|
| Age at first reproduction | |||
| Species | 2 | 2,24 | 0,11 |
| Treatment | 3 | 20,45 | < 0,001 |
| Species x treatment | 6 | 2,19 | 0,04 |
| Fecundity | |||
| Species | 2 | 5,87 | 0,003 |
| Treatment | 3 | 38,33 | < 0,001 |
| Species x treatment | 6 | 7,54 | < 0,001 |
| Total offspring | |||
| Species | 2 | 0,68 | 0,50 |
| Treatment | 3 | 36,97 | < 0,001 |
| Species x treatment | 6 | 6,05 | < 0,001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



