
Submitted:
30 June 2023
Posted:
04 July 2023
You are already at the latest version
Abstract
The accommodative response of the human eye is predominantly driven by foveal vision but reacts also to off-foveal stimuli. Here, we report on monocular accommodation measurements using parafoveal and perifoveal annular stimuli centered around the fovea and extending up to 8 degree radial eccentricity for young emmetropic and myopic subjects. The stimuli were presented through a sequence of random defocus step changes induced by a pupil-conjugated tunable lens. A Hartmann-Shack wavefront sensor with an infrared beacon was used to measure real-time changes of ocular aberrations up to and including the 4th radial order across a 3 mm pupil at 20 Hz. Our findings show a significant reduction in accommodative response with increased radial eccentricity.
Keywords:
Accommodation
; Sign of Defocus
; Aberrations
; Wavefront sensor
; Tunable lens
; Parafovea
; Perifovea
; Emmetropization
; Myopia
1. Introduction
Accommodation is the change in optical power of the crystalline lens of the human eye to compensate for defocus induced by different viewing distances. The amplitude of accommodation reduces with age and becomes almost zero at around 52 years of age [1], although some studies have found that this reduction can be extended up to 60 to 70 years [1,2,3]. The accommodative response also depends on stimulus properties, such as spatial frequency content [4] and the location in the visual field [5]. In this study, we investigate the ability of the human eye when accommodating to annular stimuli presented in the parafoveal and perifoveal visual field.
Campbell concluded that foveal cones are responsible for accommodation. i.e., only when the luminance equals the cone threshold of visibility will the accommodative reflex be triggered [6]. In the case of parafoveal vision, Fincham reported that observers were unable to accommodate to a white spot of light when its width exceeded 10 arcmin and the monocular viewing angle was more than 10 arcmin away from the spot [7]. Yet, Semmlow and Tinor measured parafoveal accommodative vergence response while viewing a 6 arcmin dark target at eccentricities up to 6° and found an approximately linear drop-off in response with eccentricity [8]. Similarly, Gu and Legge measured parafoveal accommodation by varying the size of a dark disc on a bright background and instructing observers to view the contour of the circle as sharp as possible at eccentricities of 1°, 7°, 15° and 30° [9]. They concluded that peripheral vision evokes accommodative responses albeit with reduced magnitude. The same conclusion was reached by Hartwig, Charman and Radhakrishnan when measuring the accommodative response to peripheral stimuli in myopes and emmetropes [10]. When both foveal and parafoveal accommodative cues are present, a compromise in response has been reported [11,12].
The question of parafoveal accommodation is not only of fundamental interest but also directly relevant for subjects affected by central vision loss who rely partially or entirely on parafoveal vision and vision aids [13,14]. Furthermore, peripheral optics are believed to play a role in emmetropization. The image on the peripheral retina depends on the optical errors, the state of accommodation, and environmental factors since the surroundings are not dioptrically uniform [15,16]. One such environmental factor is an increase in indoor activities, which has been found to be a major reason for myopia prevalence in the young population [17]. Sprague et al. [16] estimated that an average 2.1 refocusing/s throughout the day is required by the eye for daily tasks with a threshold of 0.4 diopters. This is close to 121,000 refocusing tasks per day. Thus, understanding the sensitivity to defocus at different eccentricities, and the corresponding effect on accommodation in various settings, may provide vital insight into mechanisms that stimulate eye growth and myopia [18].
We recently introduced an automated method to study foveal-driven accommodation using a current-driven tunable lens (TL) in a conjugated pupil plane to introduce a random sequence of defocus step changes within the accommodative range of each observer [19]. Randomness is used to avoid subjects from learning the required response that can otherwise mask the desired signal [20]. We expand on our earlier study and report here on measurements of the accommodative response across the macula from the fovea and parafovea to the perifovea for young observers to green-colored annular stimuli with visual angle radii from 1° to 8°. Other studies have used other smaller targets to examine the peripheral retina [9,12]. They have the advantage of including higher spatial frequencies but are localized to just a small area of the retina at any time. In turn, the use of annular targets benefits from including all equidistant points from the fovea simultaneously. See the end of the document for further details on references.
2. Materials and Methods
Figure 1 shows a schematic of the monocular vision system used to measure parafoveal accommodation. The TL (OptotuneTM EL-16-40-TC-VIS-5D-C) was used to generate a sequence of negative defocus steps within the comfortable accommodative range of each subject. This range is determined manually before measurements by asking each subject to look at the target while adjusting the power of the TL via software. The TL has a nominal response time of 5 ms and a settling time of 25 ms. The sequence of defocus steps was a random pattern that was chosen to avoid memorization of the expected response. The same sequence was used for each subject scaled to his/her accommodative range. The TL provided step defocus changes every 10 s while the subject accommodated to an annular target. The procedure was repeated for each subject with nine different radii of the annular target, viz., 1°, 2°, 3°, 4°, 5°, 6°, 7°, 7.5°, and 8°. No measurements were performed beyond 8° retinal eccentricity for any subjects.
2.1. Participants
Six participants with ages in the range of 23 to 34 years old took part in this study: 1 emmetrope (≤ −0.5 D), 2 mild myopes (≤ −3 D), and 3 moderate-to-high myopes (≤ −6 D). Subjects were provided with an information leaflet and signed an informed consent form prior to measurements. Myopic subjects wore their spectacles during measurements. Table 1 shows the age, refractive error (right eye) and experimental-determined accommodative range for each subject. The refractive error was determined with an EyeNetraTM autorefractor having a nominal error of 0.35 diopters.
2.2. Procedures
Subjects viewed with their right eye a green annular target on a dark background on a computer monitor while their left eye was covered with a dark patch. The annular target, with a line width of 1.2 mm, was chosen as it simultaneously covers all points at a fixed radial distance from the fovea. For annular targets with 1°, 2°, 3°, 4°, 5°, 6°, 7°, 7.5° and 8° retinal eccentricity, the corresponding retinal illuminance was 1.51, 1.80, 1.98, 2.11, 2.21, 2.29, 2.35, 2.38 and 2.41 log Td. The monitor was placed at a viewing distance of 1-meter from the conjugated pupil plane corresponding to an initial +1 diopter baseline of accommodation. The Zernike defocus coefficient C20 determined with the wavefront sensor was converted from micrometers (µm) to diopters (D) as follows:
where, d is the pupil size in diameter. The factor of 16 is due to the scaling of the wavefront at the pupil onto the Hartmann-Shack wavefront sensor (HS-WFS). A near-IR laser source of 850 nm wavelength (Edmund OpticsTM) was focused onto the retina and served as probe to sense real-time ocular aberrations at 20 Hz including up to the 4th radial order of the Zernike aberrations with a CMOS-based HS-WFS (ThorlabsTM WFS20-5C). The system was computer controlled via a LabviewTM interface. A 3 mm iris was placed in a conjugated pupil plane to limit the effective pupil diameter to keep the measurement situation identical in all cases.
A small pupil limits the impact of higher-order aberrations and increases the depth of focus resembling vision outdoors. In our earlier study, we found no significant difference in accommodative amplitude between pupil sizes of 2.5, 3.5 and 4.5 mm and therefore negligible impact of higher-order aberrations [19]. Subjects were not dilated, but the room was dark to ensure a sufficiently large natural pupil. As only changes in accommodative state were studied, a possible myopia shift caused by the dark environment, i.e., night myopia [21] was of no concern as it would provide a constant offset. A bite bar was used to limit unwanted head motion during measurements. For each of the 9 annular stimuli the predetermined random sequence of defocus steps was induced by the TL and the subjects were instructed to look straight onto the wavefront sensing beam (red dot) while keeping the circular accommodation target in focus as well as they could. To exclude disturbance by the IR beacon, separate measurements were performed without the green annular targets where the subjects were instructed to look at the red laser dot, and it was verified that the initially measured defocus of the eye did not change when the TL cycled through the defocus sequence. Thus, all subjects were able to suppress the IR beacon and concentrate on the green annular targets while using the red dot as a fixation point.
A sequence of random defocus steps was generated, and data acquisition was in each case limited to 110 s to limit fatigue. Every 5 to 10 min. subjects would rest to relax their vision and longer breaks were given if they found it difficult to accommodate. To increase comfort, data collection took place across 2 to 3 days for each subject and measurements were repeated between 2 - 3 times at certain circumstances, i.e., if the HS-WFS signal was degraded by unwanted eye motion (due to subject’s head position movement) or tiredness of subjects. In all the presented data, blinks were removed numerically using MatlabTM processing without otherwise affecting the temporal response (shown in Figure S6) and defocus was determined as specified in Equation (1).
3. Experimental Results
The response to the random step sequence for subjects #1 and #2 (emmetrope and mild myope) is shown in Figure 2. For both observers, defocus changes are followed closely both during accommodation and relaxation at 1° radial eccentricity. From 2° to 6° eccentricity, accommodation and relaxation are in the expected direction but with a reduced response with increasing eccentricity which agrees with earlier studies [10,12,22,23]. At and beyond 7° eccentricity the accommodative response becomes negligible. The other mild myopic subject #3, showed the same tendency (see Figure S1 in the supplementary document). An extra step (7.5°) between 7° and 8° radial eccentricity was taken to monitor the response closely at the point where accommodation is absent.
Figure 3 shows the mean accommodative response determined for the interval 11 - 19 s (induced defocus ranged from −1.16 D to −2.10 D depending on the individual) as a function of stimuli eccentricity for all six subjects. The accommodative response decreases with eccentricity although some variations are noticeable, e.g., for subjects #2 and #3, at 3° eccentricity defocus went up compared to 2° and in case of myopic subject #6, more variations were noticed at 4°, 6° and 7.5°, which can be due to overshooting or lead of accommodation. The accommodative range and initial position differ between subjects as expected based on Table 1.
To estimate the difference between accommodative response and induced defocus further analysis was performed. Table 2 shows these results at time interval 0 - 110 s and radial eccentricities in the range of 1° - 8° together with the average for all subjects. A low value means that the response followed the induced defocus well. Except for subject #3, all subjects showed a gradual decrease in accommodative response with eccentricity. Subject #3 is the oldest of the subjects and therefore the range and drop-off with eccentricity is smaller. On average, the accommodative response flattens out for radial eccentricities from 7° and beyond.
To determine whether other Zernike terms were affected, selected coefficients are shown in Figure S2 for subject #1. As expected with a small 3 mm pupil, the defocus term dominates the response although careful comparison does reveal that astigmatism Z3 changed with the same sign as Z4 whereas astigmatism Z5 and spherical aberration Z12 changed with the opposite sign. This tendency was also noticed with foveal accommodation for pupil sizes up to 4.5 mm [19]. In some cases, coma Z7 and Z8 (for simplicity not shown) changed too but only slightly (less than 0.2%) with the same sign as Z4. In all cases, defocus alone accounts for more than 99% of the total response with the small pupil.
Figure 4 shows the accommodative response for myopic subjects #4 and #6. As emmetropes and mild myopes, myopic subjects were also able to track changes of the TL in the correct direction at up to 6° perifoveal vision albeit with reduced response. At 7° and beyond no response was observed (same for myopic subject #5; shown in Figure S3). The refraction by the spectacles impacts on the field of view. However, for most subjects this is a modest effect. With a vertex distance of 14 mm and for the most myopia subject (#6) wearing −6 D correction equals an 8% reduction in field size (or 7.3° for the 8° target). If fully corrected with contact lenses, this will not be perceived as a reduction by the subject. All subject groups showed a similar accommodative response with smaller amplitude at increasing eccentricity and reduced range with higher age. Some accommodative overshooting (lead of accommodation) was noticed, although this occurred mostly for the untrained subjects and may thus be a result of uncertainty and increased errors.
All subjects showed a similar tendency in case of reaction and response time determined following the same procedure as in Ref. [19]. Figure 5 shows the temporal response of accommodation and relaxation for subject #1 at 1° and 5° eccentricity. The reaction time (i.e., latency or delay in the onset of accommodation) differs little between (a) lower and (b) higher eccentricities while the response time (i.e., time required to complete accommodation before stabilizing at a new accommodative level) almost doubles at 5° eccentricity when compared to 1° as shown in Figure 5 (and with a reduced accommodation amplitude). For foveal stimuli the reaction time was in the range of 300 – 700 ms and the response time 200 – 800 ms where far-to-near accommodation response was faster than near-to-far relaxation [19,24,25]. For higher eccentricities (5° and beyond) the reaction time increased slightly (400 – 800 ms) though some variations may happen, e.g., relaxation reaction time is approximately 320 ms in Figure 5(b), but the response time increased substantially (900 ms – 2 s) for most of the cases. The response varied with the magnitude of the defocus step, but the oscillatory behavior was in good correspondence with earlier findings [26].
Figure 6 shows examples of (a) reaction and (b) response times for subject #1 for 1° to 6° retinal eccentricity arranged from lower to higher dioptric stimulus value. Figure 6(a) shows an increase in reaction time at 6° retinal eccentricity but there is also some variation between reaction times at other eccentricities and for different defocus stimuli. Figure 6(b) shows that the response time was almost 70% higher for 5° and 6° retinal eccentricity when compared to lower eccentricities. The longer times relate to the more challenging accommodation at increased eccentricity. Yet, as for the foveal study [19], there is no clear relation between reaction and response times for different defocus stimuli. Similar observations were made for the other subjects studied (for simplicity not shown).
Histograms for (a) reaction and (b) response times for all 6 subjects are shown in Figure 7 in intervals of 100 ms up to 2.0 s. As can be seen, the tendency of the highest numbers of reaction and response times are similar and mostly in the range of 200 – 900 ms for 1° to 5° retinal eccentricity. Fatigue as well as untrained subjects would typically have increased reaction and response times.
Figure 8 shows the mean (a) reaction and (b) response times with standard deviation for 10 step changes (measurement time 110 s) for all 6 subjects from 1° to 6° retinal eccentricity (7°, 7.5° and 8° were excluded as there was no noticeable accommodation). Both mean reaction and response times were higher for myopic subject #6 who was an untrained subject while for rest of the subjects both times were mostly under 1000 ms. In Figure 8(a) the error is the largest at 6° for subject #1, at 5° and 6° for subject #4 and #5; and at 4°, 5° and 6° for subject #6. Similarly, Figure 8(b) shows largest errors at 6°, at 5° and at 4°, 5° and 6° for subjects #1, #2 and #6, respectively. Thus, increased stimulus eccentricity increases the accommodative uncertainty and required time.
4. Discussion
For all six subjects, a reduction in parafoveal accommodation response at large eccentricity was noted, which agrees with earlier studies [8]. At 7° and beyond there was no longer a clear correspondence between defocus stimuli and the ocular response. This is smaller than in some studies [9] plausibly due to the reduction of accommodative cues using a TL. All subjects showed similar peripheral accommodative tendencies, and therefore no more subjects were recruited in this study. A related study [12] found that at 7° retinal eccentricity (14° diameter) an accommodative response was present when subjects looked onto a black-and-white textured target made with a circle containing a Maltese cross target at the center while experiencing sinusoidal induced defocus (±1.5 D) by a TL at frequencies 0.1, 0.2, 0.5 and 1 Hz. The presence of the central Maltese cross target can possibly be the reason why an accommodative response was still present. This differs from the methodology employed in the present study where concentric annular targets were displayed for increasing retinal eccentricity. Finally, the TL induced random defocus instead of a well-known periodic defocus change, thereby reducing possible accommodative cues. Small fluctuations were noticed throughout accommodation, and it is plausible that these may provide a cue to the accommodative response [27]. Blur adaptation can improve visual acuity by approximately 0.2 logMAR within approximately an hour [28,29] and a related acuity reduction occurs after removing a prescribed correction [30]. This adaptation may partially be of optical origin as photoreceptors dynamically adapt to the direction of incident light [22,23,31]. A change in monocular vergence due to accommodation could result in reduced image contrast due to leakage from the cone outer segments [11,19,32]. This would be most apparent for foveal vision due to the high density of cones whereas in the periphery the reduced cone density would make such an optical mechanism less sensitive to defocus. The cone photoreceptor density decreases with eccentricity to approximately 7000 cones/mm2 in the parafovea [33]. For a low-density hexagonal packing this translates to a cone-to-cone spacing of 12.8 µm [34]. Using simple trigonometry this suggests that the retina would be insensitive to defocus below 0.4 diopters with a 3 mm pupil. In turn, a foveal density of 160,000 cones/mm2 [33] and a hexagonal packing corresponds to a cone-to-cone spacing of 2.7 µm and a sensitivity to just 0.08 diopters of defocus. The latter is similar in magnitude to the fluctuations of accommodation. Thus, it is possible that the reduced accommodative response (see Figure 3) with increasing eccentricity may be linked to cone photoreceptor density, leakage of light from outer segments [35] and reduced sensibility to defocus in the periphery. When it comes to contrast of the stimuli, previous studies found that it does not have any influence in the accommodative response while reading on an electronic micro-display [36] or for near vision [37], and it has no significant impact on the accommodative lag [38]. Yet, other studies claimed that pupil size and higher-order aberrations may have some influence in the accommodation stimulus-response [39] and illumination intensity will impact on the amplitude of accommodation [40].
Another key parameter is the retinal ganglion cell density that also drops off with increasing eccentricity [41]. The parafoveal results are similar between emmetropic and myopic subjects. Thus, refractive error may matter less [42] than monocular vergence that possibly limits outer-segment leakage of light [32,35].
5. Conclusions
In this study we reported on an automated monocular system to measure accommodation to annular stimuli projected on the parafovea and the perifovea in response to random step defocus changes induced by a current-driven tunable lens. The accommodative response time increased gradually with increasing eccentricity while the amplitude decreased gradually and became negligible at 7° radial distance and beyond. For a 3 mm limited eye pupil, defocus is the dominating Zernike coefficient with no significant contribution from higher-order aberrations. Our experimental results clearly show that the amplitude of accommodation reduces with increased eccentricity, which can possibly be linked to the reduction in cone density and thus to a contrast reduction caused by leakage of light from the outer segments [35]. Moreover, no evidence of a different accommodative response was found between subjects of different refractive errors in this study which agrees well with previous findings [10,43]. We are conducting further research to understand the mechanisms that trigger the correct direction of accommodation and thus the detection of the sign of defocus in the young adult eye both in foveal and parafoveal vision, as this ultimately may relate to myopia and the emmetropization process [44,45].
Author Contributions
Conceptualization, B.V., N. S. and L.L.; methodology, B.V. and N.S.; software, N.S. and B.V.; validation, B.V., N.S., L.L., P.P. and D.R.; formal analysis, N.S., B.V.; investigation, N.S. and B.V.; resources, B.V., N.S.; data curation, N.S. and B.V.; writing—original draft preparation, N.S.; writing—review and editing, N.S., B.V., L.L., P.P. and D.R; visualization, B.V. and N.S.; supervision, B.V. and L.L.; project administration, B.V. and N.S.; funding acquisition, B.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by European Union’s H2020 ITN network “MyFUN” under Marie Skłodowska-Curie grant agreement no. 675137.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of UNIVERSITY COLLEGE DUBLIN Human Research Ethics Committee – Sciences (protocol code LS-16-75-Vohnsen and date of approval December 12th 2016).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study. Written informed consent has been obtained from the patient(s) to publish this paper.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors are grateful to all subjects who kindly participated in this study and to Peter Unsbo for discussions on study protocol.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Hamasaki, D.; Ong, J.; Marg, E. The amplitude of accommodation in presbyopia. Am. J. Optom. Arch. Am. Acad. Optom. 1956, 33, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Mordi, J.A.; Ciuffreda, K.J. Static aspects of accommodation: age and presbyopia. Vis. Res. 1998, 38, 1643–1653. [Google Scholar] [CrossRef] [PubMed]
- Jackson, E. Amplitude of accommodation at different periods of life. Cal. State. J. Med. 1907, 5, 163–166. [Google Scholar]
- Taylor, J.; Charman, W.N.; O'Donnell, C.; Radhakrishnan, H. Effect of target spatial frequency on accommodative response in myopes and emmetropes. J. Vision 2009, 9, 1–14. [Google Scholar] [CrossRef]
- Liu, T.; Sreenivasan, V.; Thibos, L.N. Uniformity of accommodation across the visual field. J. Vision 2016, 16, 1–16. [Google Scholar] [CrossRef]
- Campbell, F.W. The minimum quantity of light required to elicit the accommodation reflex in man. J. Physiol. (London) 1954, 123, 357–366. [Google Scholar] [CrossRef]
- Fincham, E.F. The accommodation reflex and its stimulus. Br. J. Ophthalmol. 1951, 35, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Semmlow, J.L.; Tinor, T. Accommodative convergence response to off-foveal retinal images. J. Opt. Soc. Am. 1978, 68, 1497–1501. [Google Scholar] [CrossRef]
- Gu, Y.; Legge, G.E. Accommodation to stimuli in peripheral vision. J. Opt. Soc. Am. A 1987, 4, 1681–1687. [Google Scholar] [CrossRef]
- Hartwig, A.; Charman, W.N.; Radhakrishnan, H. Accommodative response to peripheral stimuli in myopes and emmetropes. Ophthalmic Physiol. Opt. 2011, 31, 91–99. [Google Scholar] [CrossRef]
- Hennessy, R.T.; Leibowitz, H.W. The effect of a peripheral stimulus on accommodation. Percept. Psychophys. 1971, 10, 129–132. [Google Scholar] [CrossRef]
- Labhishetty, V.; Cholewiak, S.A.; Banks, M.S. Contributions of foveal and non-foveal retina to the human eye's focusing response. J. Vision 2019, 19, 1–15. [Google Scholar] [CrossRef]
- Timberlake, G.T.; Mainster, M.A.; Peli, E.; Augliere, R.A.; Essock, E.A. , Arend, L.E. Reading with a macular scotoma. I. Retinal location of scotoma and fixation area. Invest. Ophthal. Vis. Sci. 1986, 27, 1137–1147. [Google Scholar] [PubMed]
- Chung, S.T.L. Enhancing visual performance for people with central vision loss. Optom. Vis. Sci. 2010, 87, 276–284. [Google Scholar] [CrossRef]
- García, M.G.; Ohlendorf, A.; Schaeffel, F.; Wahl, S. Dioptric defocus maps across the visual field for different indoor environments. Biomed Opt Express. 2017, 9, 347–359. [Google Scholar]
- Sprague, W.W.; Cooper, E.A.; Reissier, S.; Yellapragada, B.; Banks, M.S. The natural statistics of blur. J. Vision 2016, 16, 1–27. [Google Scholar]
- Öner, V.; Bulut, A.; Oruç, Y.; Özgür, G. Influence of indoor and outdoor activities on progression of myopia during puberty. Int. Ophthalmol. 2016, 36, 121–125. [Google Scholar] [CrossRef]
- Wallman, J.; Winawer, J. Homeostasis of eye growth and the question of myopia. Neuron. 2004, 43, 447–468. [Google Scholar] [CrossRef]
- Sharmin, N.; Vohnsen, B. Monocular accommodation response to random defocus changes induced by a tuneable lens. Vis. Res. 2019, 165, 45–53. [Google Scholar] [CrossRef]
- Cornsweet, T.N.; Crane, H.D. Training the visual accommodation system. Vis. Res. 1973, 13, 713–715. [Google Scholar] [CrossRef]
- Lopez-Gil, N.; Peixoto-de-Matos, S.C.; Thibos, L.N.; González-Méijome, J.M. Shedding light on night myopia. J. Vision 2012, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Eckmiller, M.S. Defective cone photoreceptor cytoskeleton, alignment, feedback, and energetics can lead to energy depletion in macular degeneration. Prog. Ret. and Eye Res. 2004, 23, 495–522. [Google Scholar] [CrossRef]
- Vohnsen, B. Directional sensitivity of the retina: A layered scattering model of outer-segment photoreceptor pigments. Biomed. Opt. Express 2014, 5, 1569–1587. [Google Scholar] [CrossRef] [PubMed]
- Charman, W.N. The eye in focus: accommodation and presbyopia. Clin. Exp. Optom. 2008, 91, 207–225. [Google Scholar] [CrossRef] [PubMed]
- Tucker, J.; Charman, W.N. Reaction and response times for accommodation. Am. J. Optom. Physiol. Opt. 1979, 56, 490–503. [Google Scholar] [CrossRef]
- Campbell, F.W.; Westheimer, G.; Robson, J.G. Significance of fluctuations of accommodation. J. Opt. Soc. Am. 1958, 48, 669. [Google Scholar] [CrossRef]
- Charman, W.N.; Heron, G. Microfluctuations in accommodation: an update on their characteristics and possible role. Ophthalmic. Physiol. Opt. 2015, 35, 476–499. [Google Scholar] [CrossRef]
- Williams, M.M.; Tresilian, J.R.; Strang, N.C.; Kochhar, P.; Wann, J.P. Improving vision: neural compensation for optical defocus. Proc. Biol. Sci. 1998, 265, 71–77. [Google Scholar] [CrossRef]
- Cufflin, M.P.; Mallen, E.A.H. Blur adaptation: clinical and refractive considerations. Clin. Exp. Optometry 2020, 103, 104–111. [Google Scholar] [CrossRef]
- Pesudovs, K.; Brennan, N. Decreased uncorrected vision after a period of distance fixation with spectacle wear. Optom. Vis. Sci. 1993, 70, 528–531. [Google Scholar] [CrossRef]
- Carmichael Martins, A.; Vohnsen, B. Directional light-capture efficiency of the foveal and parafoveal photoreceptors at different luminance levels: an experimental and analytical study. Biomed. Opt. Express 2019, 10, 3760–3772. [Google Scholar] [CrossRef] [PubMed]
- Vohnsen, B.; Carmichael, A.; Sharmin, N.; Qaysi, S.; Valente, D. Volumetric integration model of the Stiles-Crawford effect of the first kind and its experimental verification. J. Vision 2017, 17, 1–11. [Google Scholar] [CrossRef]
- Curcio, C.A.; Sloan, K.R.; Kalina, R.E.; Hendrickson, A.E. Human photoreceptor topography. J. Comp. Neurol. 1990, 292, 497–523. [Google Scholar] [CrossRef] [PubMed]
- Vohnsen, B.; Iglesias, I.; Artal, P. Directional light scanning laser ophthalmoscope. J. Opt. Soc. Am. A 2005, 22, 2606–2612. [Google Scholar] [CrossRef] [PubMed]
- Vohnsen, B. Geometrical scaling of the developing eye and photoreceptors and a possible relation to emmetropization and myopia. Vis. Res. 2019, 189, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Bernal-Molina, P.; Esteve-Taboada, J.J.; Ferrer-Blasco, T.; Montés-Micó, R. Influence of contrast polarity on the accommodative response. J. Optom. 2019, 12, 38–43. [Google Scholar] [CrossRef]
- Ward, P.A. The effect of stimulus contrast on the accommodation response. Ophthal. Physiol. Opt. 1987, 7, 9–15. [Google Scholar] [CrossRef]
- Bakaraju, R.C.; Yeotikar, N.S.; Rao, V.S. Accommodative lag versus different stimuli. J. Mod. Opt. 2007, 54, 1299–1305. [Google Scholar] [CrossRef]
- Buehren, T.; Collins, M.J. Accommodation stimulus-response function and retinal image quality. Vis. Res. 2006, 46, 46,1633–1645. [Google Scholar] [CrossRef]
- Kim, D.Y.; Kim, S.Y.; Hyun, H.G.; Moon, B.Y. Effects of variation of illumination on visual function factors. J. Korean Ophthalmic. Opt. Soc. 2015, 20, 195–200. [Google Scholar] [CrossRef]
- Watson, A.B. A formula for human retinal ganglion cell receptive field density as a function of visual field location. J. Vision 2014, 14, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lundström, L.; Mira-Agudelo, A.; Artal, P. Peripheral optical errors and their change with accommodation differ between emmetropic and myopic eyes. J. Vision 2009, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Abbott, M.L.; Schmid, K.L.; Strang, N.C. Differences in the accommodation stimulus response curves of adult myopes and emmetropes. Ophthalmol. Physiol. Opt. 1998, 18, 13–22. [Google Scholar] [CrossRef]
- Swiatczak, B.; Schaeffel, F. Myopia: why the retina stops inhibiting eye growth. Sci. Rep. 2022, 12, 21704. [Google Scholar] [CrossRef]
- Rozema, J.; Dankert, S.; Iribarren, R. Emmetropization and non-myopic eye growth. Surv. Ophthalmol. 2023, 68, 759–783. [Google Scholar] [CrossRef]
Figure 1.
Schematic of the monocular vision system used to measure foveal, parafoveal and perifoveal accommodation responses to a sequence of random defocus steps. Three 4-f telescopes were used to place the iris, HS-WFS and tunable lens in conjugated pupil planes. Beam splitter 1 is a hot mirror whereas beam splitter 2 is a 50/50 coated plate. Only a single green ring in the annular accommodation target was shown at any given time.
Figure 1.
Schematic of the monocular vision system used to measure foveal, parafoveal and perifoveal accommodation responses to a sequence of random defocus steps. Three 4-f telescopes were used to place the iris, HS-WFS and tunable lens in conjugated pupil planes. Beam splitter 1 is a hot mirror whereas beam splitter 2 is a 50/50 coated plate. Only a single green ring in the annular accommodation target was shown at any given time.
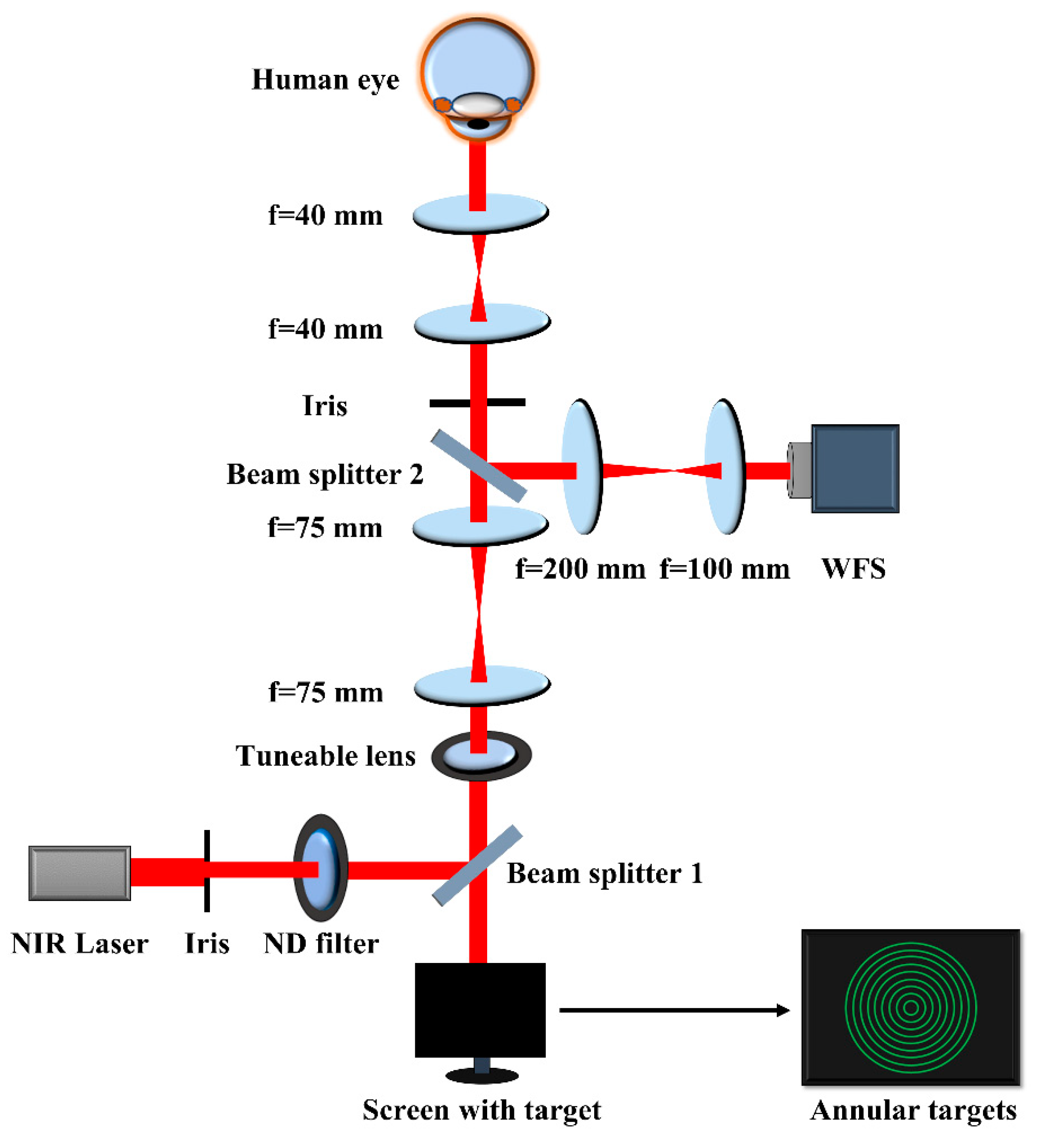
Figure 2.
Induced defocus sequence by the TL (black line) and accommodation response (red line) as a function of time for subjects (a) #1 and (b) #2 with increasing radial eccentricity from 1° to 8°. The induced defocus is shown with opposite sign to ease comparison.
Figure 2.
Induced defocus sequence by the TL (black line) and accommodation response (red line) as a function of time for subjects (a) #1 and (b) #2 with increasing radial eccentricity from 1° to 8°. The induced defocus is shown with opposite sign to ease comparison.
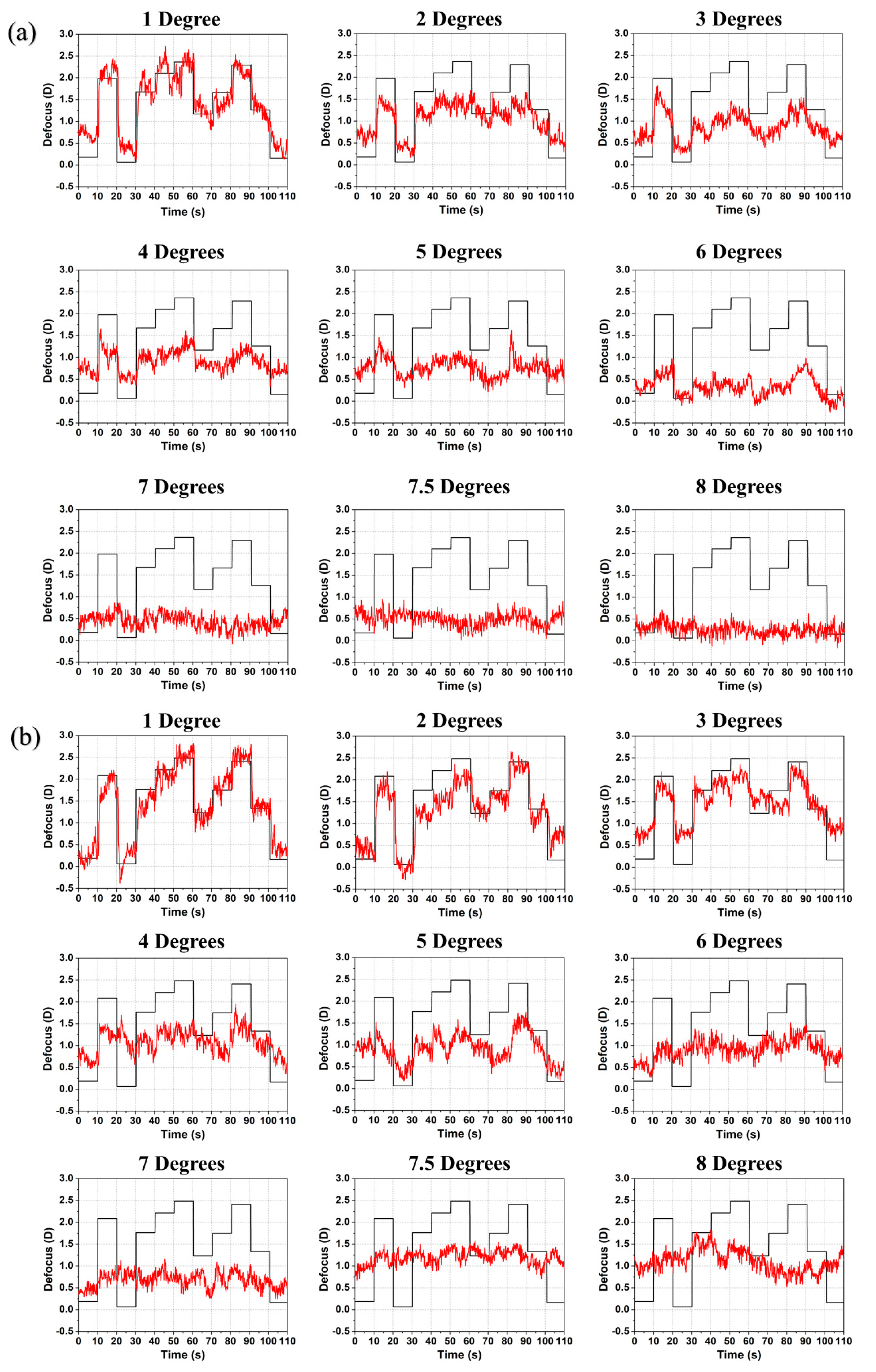
Figure 3.
Mean accommodative response as a function of eccentricity for all subjects at time interval 11-19 s where the induced defocus by the TL was (#1) −1.98, (#2) −2.08, (#3) −1.16, (#4) −1.93, (#5) −2.10, and (#6) −1.87 diopters.
Figure 3.
Mean accommodative response as a function of eccentricity for all subjects at time interval 11-19 s where the induced defocus by the TL was (#1) −1.98, (#2) −2.08, (#3) −1.16, (#4) −1.93, (#5) −2.10, and (#6) −1.87 diopters.
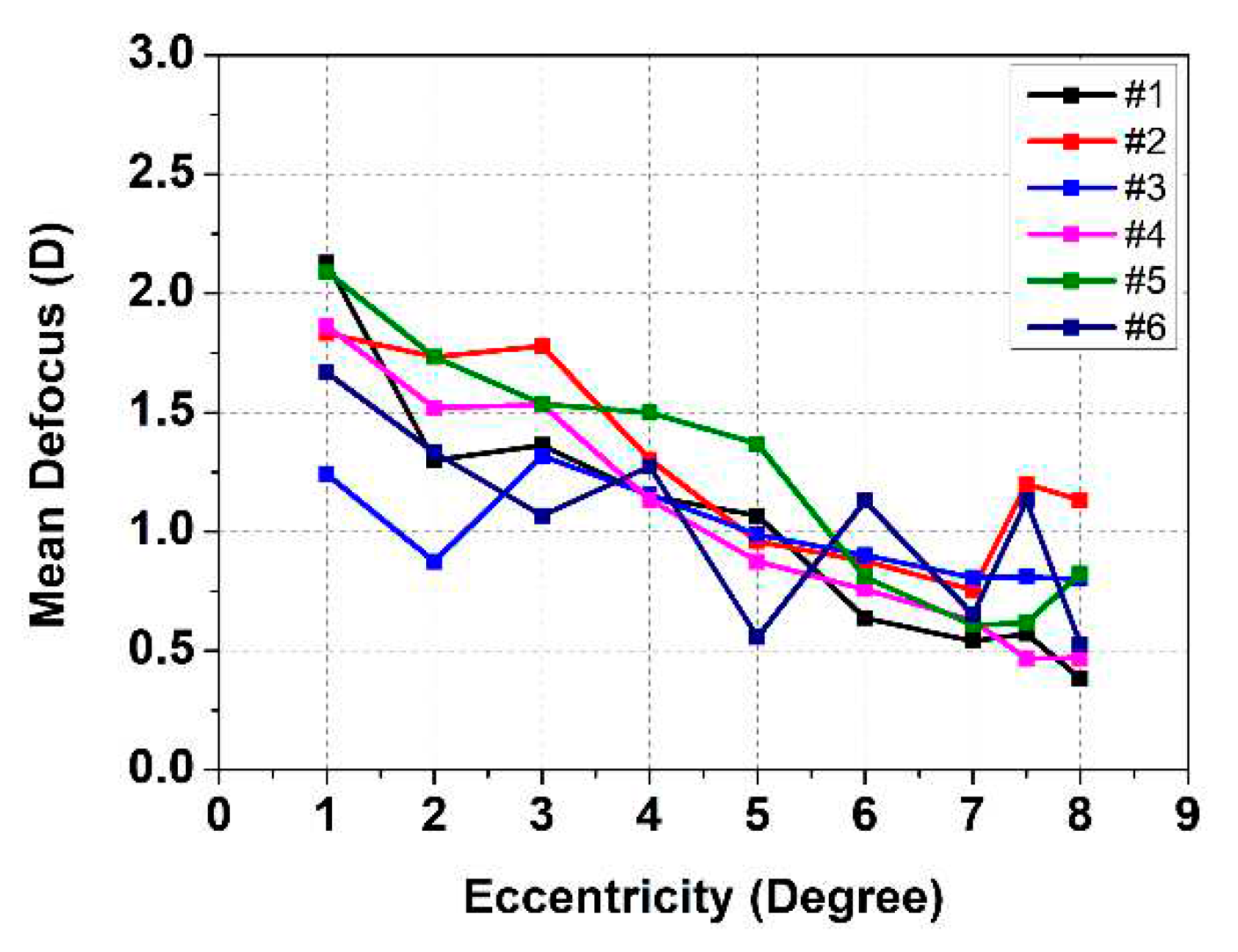
Figure 4.
Induced defocus sequence by the TL (black line) and accommodation response (red line) as a function of time for subjects (a) #4 and (b) #6 with increasing radial eccentricity from 1° to 8°. The induced defocus is shown with opposite sign to ease comparison.
Figure 4.
Induced defocus sequence by the TL (black line) and accommodation response (red line) as a function of time for subjects (a) #4 and (b) #6 with increasing radial eccentricity from 1° to 8°. The induced defocus is shown with opposite sign to ease comparison.
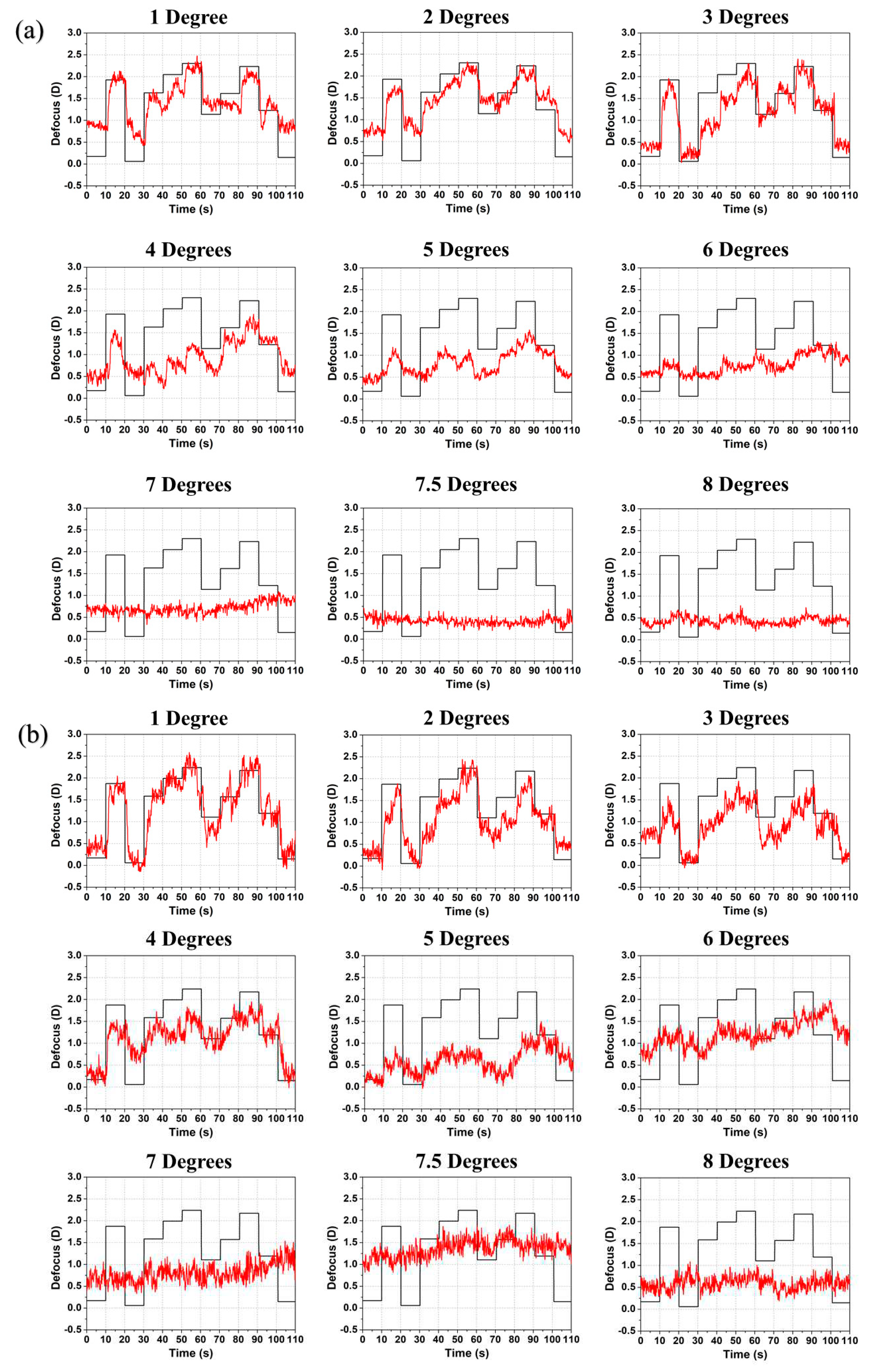
Figure 5.
Examples of temporal dynamics with two stimulus radii (a) 1° and (b) 5°: defocus (TL: black line) and accommodation (eye: red line) following an accommodative step change (left) and relaxation (right) for subject #1. Determined reaction time (brown rectangle) and response time (green rectangle) are indicated.
Figure 5.
Examples of temporal dynamics with two stimulus radii (a) 1° and (b) 5°: defocus (TL: black line) and accommodation (eye: red line) following an accommodative step change (left) and relaxation (right) for subject #1. Determined reaction time (brown rectangle) and response time (green rectangle) are indicated.
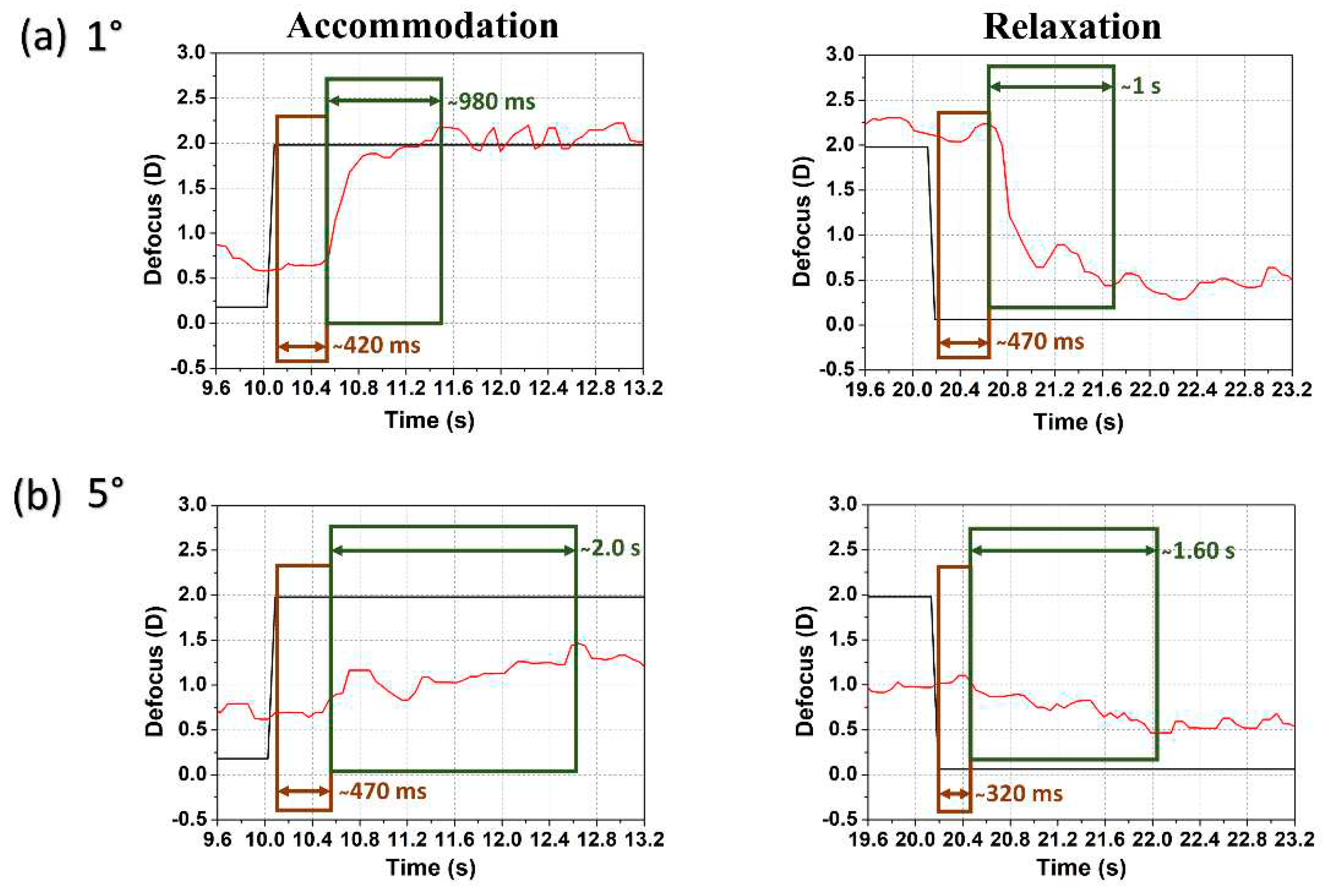
Figure 6.
Parafoveal and perifoveal (a) reaction and (b) response times for different defocus step changes in the TL (arranged from lower to higher steps) for subject #1.
Figure 6.
Parafoveal and perifoveal (a) reaction and (b) response times for different defocus step changes in the TL (arranged from lower to higher steps) for subject #1.
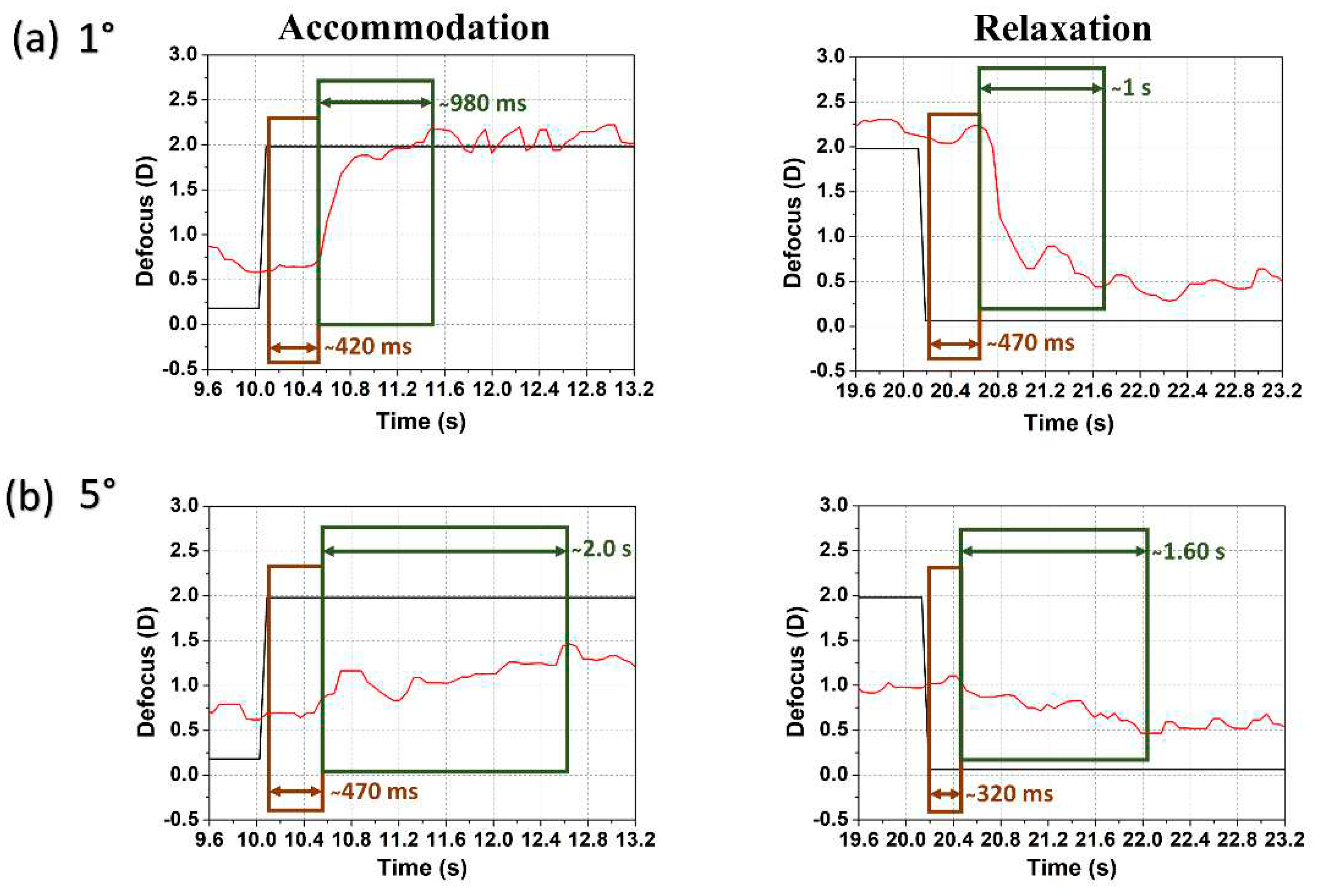
Figure 7.
Total number of cases for (a) reaction and (b) response times for all 6 subjects from 1° to 6° retinal eccentricity.
Figure 7.
Total number of cases for (a) reaction and (b) response times for all 6 subjects from 1° to 6° retinal eccentricity.
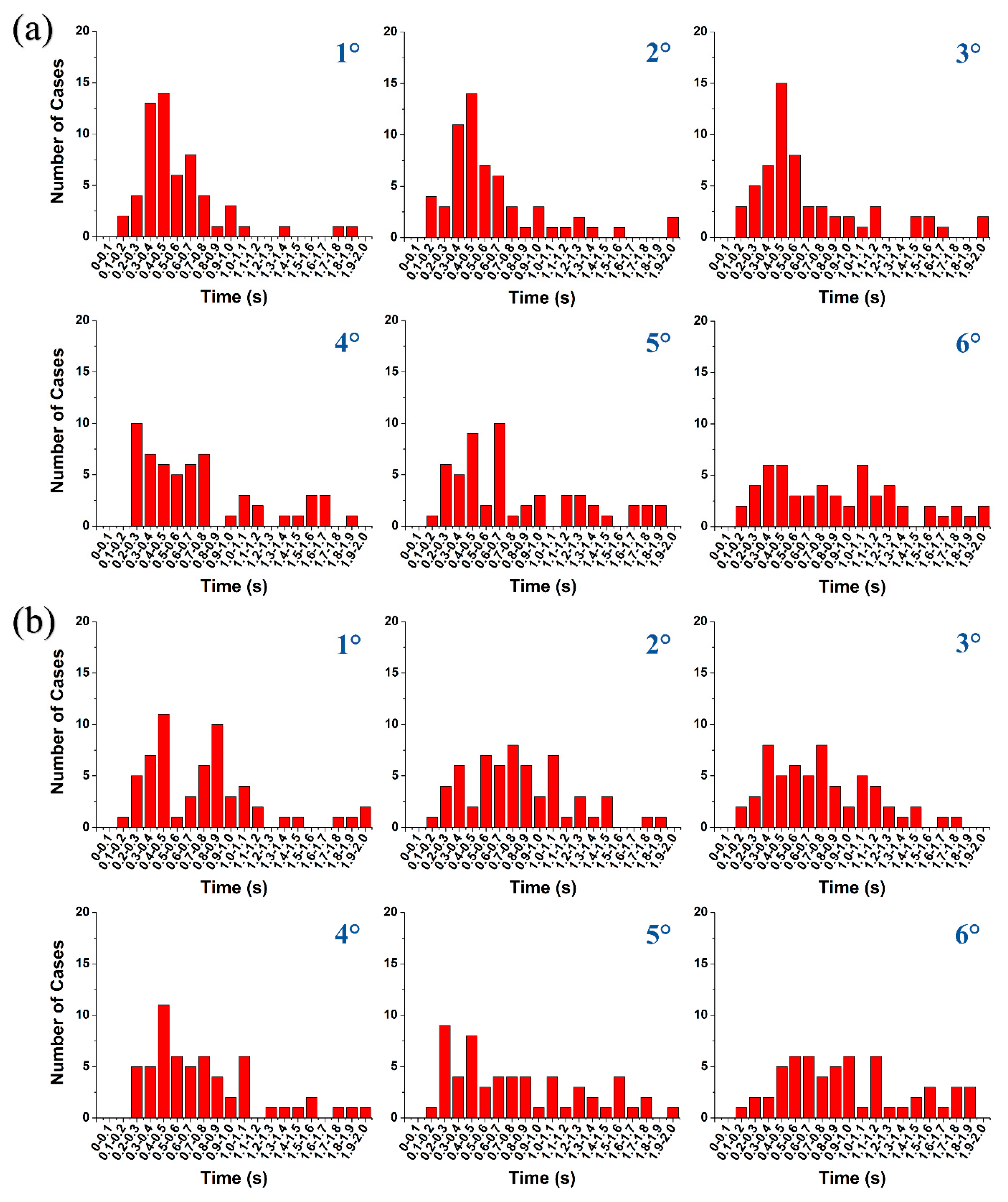
Figure 8.
Mean (a) reaction and (b) response times with standard deviation (blue lines) for all 6 subjects from 1° to 6° retinal eccentricity.
Figure 8.
Mean (a) reaction and (b) response times with standard deviation (blue lines) for all 6 subjects from 1° to 6° retinal eccentricity.
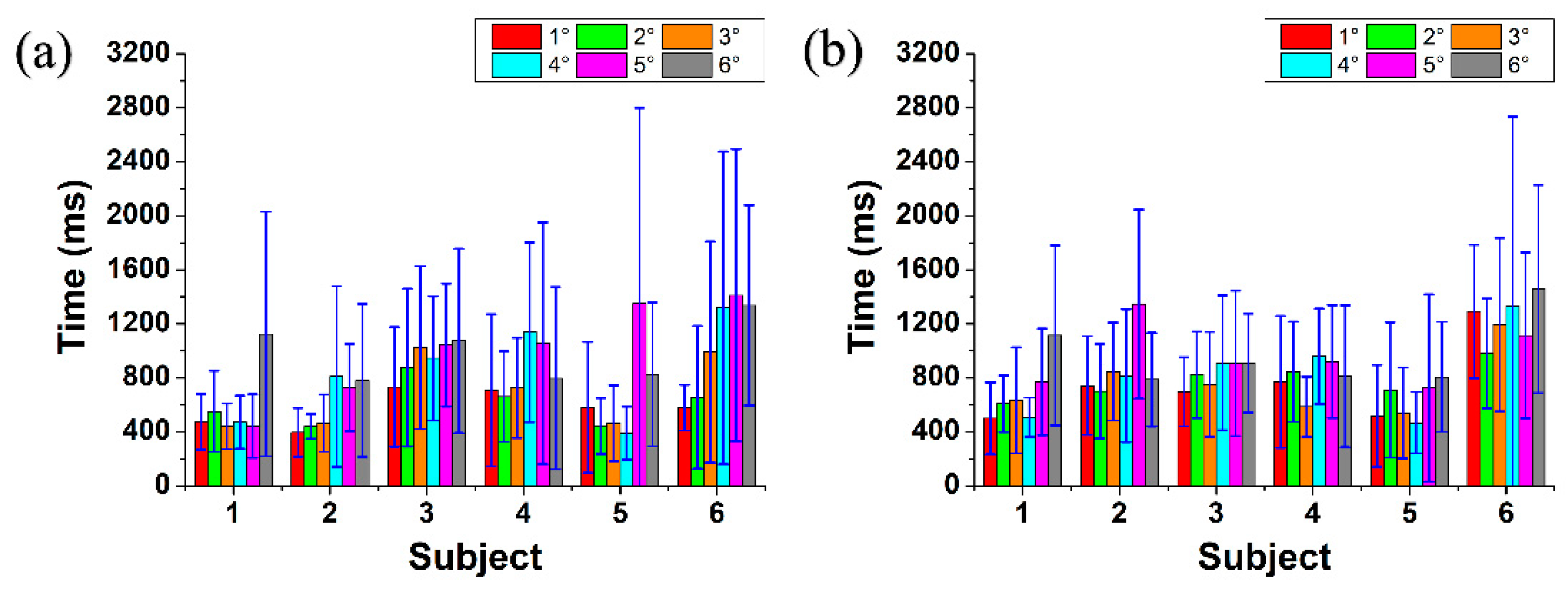

Table 1.
Refractive error in diopters (D) of the subject’s right eye. The comfortable accommodative (Ac) range was determined manually with the TL by each subject. Subject #1 is emmetrope, subjects #2 and #3 are mild myopes and subjects #4 - #6 are myopes who all wore their spectacles during measurements. The visual targets were displayed on a screen at 1-meter distance from the conjugated pupil plane (1 diopter initial bias).
Table 1.
Refractive error in diopters (D) of the subject’s right eye. The comfortable accommodative (Ac) range was determined manually with the TL by each subject. Subject #1 is emmetrope, subjects #2 and #3 are mild myopes and subjects #4 - #6 are myopes who all wore their spectacles during measurements. The visual targets were displayed on a screen at 1-meter distance from the conjugated pupil plane (1 diopter initial bias).
| Subject | Age (Years) | Sphere (D) | Cylinder (D) | Ac. Range (D) |
|---|---|---|---|---|
| #1 | 29 | −0.50 | 0.00 | 3.40 |
| #2 | 28 | −0.75 | 0.00 | 3.53 |
| #3 | 34 | −0.75 | 0.00 | 2.42 |
| #4 | 24 | −4.75 | 0.00 | 3.35 |
| #5 | 23 | −5.75 | 0.75 (Axis 30°) | 3.55 |
| #6 | 25 | −6.00 | −2.25 (Axis 15°) | 3.28 |
Table 2.
The average root-mean-square (RMS) deviation between induced defocus and accommodative response for all subjects for stimuli at eccentricities from 1° to 8°. The time interval was 0 to 110 s.
Table 2.
The average root-mean-square (RMS) deviation between induced defocus and accommodative response for all subjects for stimuli at eccentricities from 1° to 8°. The time interval was 0 to 110 s.
| Subject | 1° | 2° | 3° | 4° | 5° | 6° | 7° | 7.5° | 8° |
|---|---|---|---|---|---|---|---|---|---|
| #1 | 0.388 | 0.628 | 0.802 | 0.805 | 0.956 | 1.259 | 1.247 | 1.222 | 0.925 |
| #2 | 0.378 | 0.482 | 0.504 | 0.816 | 0.883 | 0.936 | 1.116 | 0.855 | 0.935 |
| #3 | 0.415 | 0.435 | 0.676 | 0.444 | 0.418 | 0.425 | 0.497 | 0.469 | 0.499 |
| #4 | 0.510 | 0.476 | 0.481 | 0.797 | 0.845 | 0.967 | 1.039 | 1.224 | 1.199 |
| #5 | 0.476 | 0.499 | 0.719 | 0.671 | 0.842 | 1.112 | 1.294 | 1.258 | 1.185 |
| #6 | 0.401 | 0.544 | 0.634 | 0.564 | 0.997 | 0.760 | 0.954 | 0.752 | 1.059 |
| Average | 0.428 | 0.510 | 0.636 | 0.683 | 0.823 | 0.910 | 1.024 | 0.963 | 0.967 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



