
Submitted:
03 July 2023
Posted:
04 July 2023
You are already at the latest version
Abstract
Drought and salinity are among the most important abiotic stress factors that limit the productivity in economically important plants. Although genes from wild relatives or unadapted germplasm have often been used for stress tolerance breeding, only few studies have examined wild relatives to understand the genetic and molecular basis of stress tolerance. In this study, we monitored the changes in gene expression profiles of leaf and root tissues of two wild barleys (Hordeum spontaneum L.) accessions from Israel after treatment with salt (NaCl) and drought (PEG) stress.
We identified a total of 641 differentially expressed genes across 24 conditions (2 accessions, 2 stress conditions, 2 tissues and 3 time points). Our gene expression analyses revealed large numbers of differentially expressed genes in different accessions by different stress treatments. In addition, the number of genes altered in the leaves of stress-exposed plants of both genotypes was higher than those altered in the roots. Interestingly, there was a relatively little overlap between the leaf and the root stress-responsive gene expression patterns, suggesting that different stress-associated processes might be operating in these tissues during stress adaptation.
Overall, our results revealed a number of candidate genes and plant processes associated with stress tolerance in wild barley. Wild barley would be a useful source of new genetic variation for drought and salt stress tolerance. Our results could provide new insights into the mechanisms of drought and salt stress tolerance in wild barley and should be useful for genetic improvement of salt-drought tolerance in cultivated barley.
Keywords:
wild barley
; (Hordeum spontaneum L.)
; drought and salinity
; transcriptome analysis
1. Introduction
Abiotic stress factors such as drought and salinity are global issues that adversely affect yield potential of many crop plants throughout the world [1,2]. Therefore, improving abiotic stress tolerance is an important aim of many breeding programs. Importantly, these two seemingly different stress factors can have many commonalities as far as the physiological effects of stress on essential cellular processes are concerned. For instance, both water and salt stress negatively influence the ability of plants to take up water and can therefore induce similar or overlapping physiological effects [3].
Causing the big losses in crop productivity [4], negative impacts of the drought stress are emerged during the various growing phases of plants [5]. Humidity loss ratio from the leaves as a result of transpiration tends to be higher than the water absorbance ratio by the roots [6], so that decrease in the water potential of the soil followed by the plant is coinsided leading to the turgor pressure drop in severe conditions [7]. In such conditions, abscisic acid (ABA) which is synthesized in roots and translocated to the leaves via transpiration, causes the stomata closure resulting in the decrease in CO2 concentration, Rubisco activity and photosynthesis rate [8]. In plants, physiologic, morphologic, and biochemical responces in various levels are emerged across the drought stress [9]. By slowing biosynthetic reactions, need to ATP is decreased, excess electron in electron transport system of mitochondria and chloroplasts is encountered, and synthesis of compounds like superoxide, hydrogen peroxide (H2O2), reactive oxygen species (ROS), superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), and ascorbate peroxidase (APX) is expected [8].
To resist destructive effects of ROS and unfavorable environmental conditions, plants possesse enzymatic/enzymatic non-antioxidant defense system and variousely are adapted based on genus and genotype, water loss rate, growth phase, organ and cell type [10,11]. Furthermore, stressed plants accumulate sugar, amino acid (prolin), polyamine, glysine betaine, and inorganic ion (K+) to adjust osmotic potential [12]. In plants exposed to these stresses, essential cellular constituents such as proteins/enzymes and nucleic acids can be inactivated through the accumulation of reactive oxygen caused by cellular water loss [13].
Soil salinity has been accepted to be an important environmental stress and continuously changing global climatic factors tend to intensify its effects. Salinity stress commonly is referred to excess sodium ion accumulation in the soil [14], and this situation results in the osmotic stress and ion toxisity [15]. In relation to the plant response to salt stress, hyper-osmotic stress, ion toxicity caused by cellular ion homeostasis imbalance, nutrition imbalance, and oxidative damages of excess ROS rates might be emerged [16,17]. Barley as the forth main crop over the world after wheat, rice, and maize is produced for animal fedding and human nutrition [18]. Relative to other major cereal crops such as wheat (Triticum aestivum L.), barley (Hordeum vulgare L.) is a moderately drought- and salt-tolerant species cultivated as a food and feed crop in many arid and semi-arid regions of the world [19,20].
In different growth phases of barley, spike and grain formation and anthesis are among the most sensitive yield-related features negatively affected by drought stress [21,22] so that the remarkable yield loss of about % 49-87 has been emphasized. In spite of beeing the most tolerant cereal crop against the salinity, retarded growth and yield loss under the sever salinity conditions is observed in barley [23]. In this crop, salinity tolerance varies depending on the growth stages and genotypes. For instance, seed germination and seedling stages are among the most sensitive ones in salinity conditions [24]. Nevertheless, salinity tolerance is very complicated process. This involves osmo-protectant biosynthesis, photosynthesis regulation, hormone signals, adjustment of ion homeostasis, and antioxidant metabolism by which, water uptake and various regulatory interaction pathways like osmotic tolerance, take place. All these pathways percept salt stress complicatedly and remain activated during the salinity signals, along with the various stress related regulatory gene expression families [15,17].
Several physiological [25,26], cytogenetic [27], and genetic studies have been performed in recent years to dissect the molecular and physiological basis of salt and drought tolerance in barley [20,28,29,30,31].
Wild relatives of crop plants often adapted to harsher environments than their cultivated counterparts can contain many genes associated with stress tolerance [32,33]. Wild barley survives severe environmental conditions for a long time developing suitable adaptation system. Due to the natural tolerance of this crop against drought and salinty stresses and fungal diseases [34,35]. and on account of whole identification of its genome sequence [36], the crop has been accepted to be the model plant in stress biology studies. On the other hand, due to the fast disappearence of genetic variation, modern barley cultivars tend to be more sensitive to biotic and abiotic stresses [37]. On account of the rich genetic variation of the wild barley [37,38], it is considered the primary gene basin in modern breeding attempts [39]. Furthermore, wild and cultured barley genotypes have been accepted to possess various drought stress tolerance abilities [40,41], so, identifying the drought tolerance related genes/features in germplasm of the tolerant wild barley, is of great importance in improving tolerant cultivars [38,42]. For example, Hordeum spontaneum C. Koch, the wild progenitor of cultivated barley found in a wide-range of habitats in the Middle East is thought to contain many genes that can be exploited for genetic improvement of stress tolerance [37]. Profiling of plant gene expression under different environmental conditions enables the identification of candidate stress tolerance genes [43]. In recent years, both cultivated and wild barley genotypes have been subjected to stress-asssociated transcriptome analyses. In earlier studies, Walia et al. [44] performed an expression analysis of barley under salinity stress and identified large numbers of differentially-expressed genes. Talame et al. [29] found that the genes upregulated in cultivated barley under salt and drought stress encode proteins with roles in defence and stress acclimation pathways. Several recent studies have also identified differentially expressed genes in wild barley accessions [45,46,47]. For instance, Bedada et al. [46] reported various genes differentially expressed between a salt-tolerant and a salt-sensitive H. spontaneum accession. Similarly, Bahieldin et al. [47] identified diffentially expressed genes in the leaves of H. spontaneum exposed to salt stress while Wu et al. [45] compared gene expression profiles of H. vulgare and H. spontaneum. More recently, Liang et al. [33] examined dehydration-responsive gene expression patterns in the Tibetan hulless barley (Hordeum vulgare var. nudum), a barley genotype adapted to stressfull environmental conditions found in the Qinghai-Tibetan plateau. However, to the best of our knowledge, salt- and drought stress responsive gene expression profiles have not been comparatively analysed in different tissues and genotypes of H. spontaneum.
In the present study, we have studied the expression patterns of genes responding to dehydration and salinity in the leaf and root tissues of two wild barley accessions using the Affymetrix Barley 1 GeneChip containing 22.400 unigenes. Our results from these analyses identified a number of candidate genes and physiological processes potentially associated with stress tolerance. Surprisingly, the wild barley accessions studied here showed largely independent responses to stress treatments. We found very little overlap, if any, between the differentially expressed genes identified in the roots and the leaves of the same genotypes by different stress treatments. Future functional analyses of candidate genes identified in this study will improve our overall understanding of the molecular basis of stress tolerance in barley.
2. Materials and methods
2.1. Plant material and Growth
Two wild barley (Hordeum spontaneum L.) accessions (20-40 and 23-70) obtained from the Genebank of Haifa University located at the Institute of Evolution in Israel, were used for leaf water potential and gene expression analyses. Seeds were surface-sterilized with 5% sodium hypochlorite for 20 min, washed with distilled water and pre-germinated in petri dishes at room temperature in the dark. Wild barley seedlings at a similar germination stage were then grown hydroponically in pots containing 0.2 L of modified 1/10 Hoagland’s solution (K2SO4, KH2PO4, MgSO4.7H2O, Ca(NO3)2.4H2O and KCl) and micronutrients (H3BO3, MnSO4, CuSO4.5H2O, NH4Mo, ZnSO4.7H2O) with a final concentration of the following ions, 2mM Ca, 10-6 M Mn, 4mM NO3, 2.10-7 M Cu, 1mM Mg, 10-8 M NH4, 2mM K, 10-6 M Zn, 0.2 mM P, 10-6M B and 10-4 M Fe) in a growth chamber under 16 h daylight at 28 °C and an 8 h dark period at 23 °C for 15 days.
2.2. Stress Applications
Stress treatments were applied to 15-day-old seedlings. Salt stress (NaCl) was applied gradually as described by Walia et al. [20] in barley. Initially, 25mM of salt was applied to seedlings for a day and the salt concentration used was gradually elevated daily first to 50mM, then to 75 mM and finally to 100mM during the next three days (Figure 1). CaCl2 was also added together with NaCl to maintain a Na+/Ca2+ concentration ratio of 10:1 on a molar basis to prevent a salt-induced Ca2+ deficiency [48].
PEG 6000 (polyethylene glycol-6000; MW:5000-7000 g/mol, Bio-word) was added (100mM PEG, 20%) to the nutrient solution until the osmolality of the solution, as measured using a vapor pressure osmometer, became identical to that of the 100mM salt solutions to match stem water potentials of plants exposed to either treatment. In gene expression analyses, plants were subjected to stress treatments as 0h (T0: control for control + salt+ PEG), 3h (T3), and 27h (T27) at concentrations of 100 mM salt or 100 mM PEG. Both stress control plants (without any treatment) were irrigated with Hoagland's solution at the same time and at the same rate every day. After 24 h salt or PEG exposure, root, leaf and controls samples (0h, 3h and 27h) for gene expression studies (Figure 1), immediately frozen in liquid nitrogen and stored at −80°C until further use.
2.3. Leaf Water Potential Measurements
Leaf water potentials (MPas) were measured on fully expanded leaves using a pressure chamber according to the manufacturer’s instructions at 0 (T0), 3(T3) and 27(T27) h following salt and PEG treatments using a Model 600, PMS Instrument Company (Albany, USA). All measurements were replicated 5 times. The MPa values were compared to the control values using the t-test at the p <0.05 significance level.
2.4. RNA Extractions and Barley Genechip Array Hybridizations
This study included two treatments (PEG and salt); two wild barley genotypes (23-70 and 20-40); two tissues (leaf and root); three time-points (0h (T0), 3h (T3), and 27h (T27)) and three biological replicates per treatment. In total, 108 Affymetrix Barley1 Gene Chips, (a total of 18 chips with 3 biological replicates of both root and leaf tissue (control chips: for both stresses (PEG and salt)) were used.
Total RNA was isolated from salt- or PEG-treated and control plants using a Qiagen RNeasy Plant Mini Kit (Qiagen, Valencia, USA) according to the manufacturer’s instructions. RNA integrity was confirmed by electrophoresis on 1% agarose gels containing formaldehyde and RNA quality was confirmed with an Agilent 2100 Bioanalyzer (Santa Clara, CA) using RNA LabChip® assays according to the manufacturer’s instructions.
Sample labellings and hybridizations to Affymetrix Gene Chips were conducted according to the manufacturer’s instructions. In brief, cDNA was synthesized from 3 μg of total RNA, followed by second-strand DNA synthesis using a 3′one-cycle cDNA Synthesis kit (Affymetrix, Santa Clara, CA). Biotin labeled cRNA was synthesized using MEGAscript® IVT labeling kit (Affymetrix), fragmented by Mg2+ hydrolysis and hybridized for 16 h at 45°C. Arrays were then washed using a GeneChip® Fluidics Station 450 and scanned using a GeneChip® 3000 Scanner. Spiking controls were added to total RNA before cDNA synthesis. Additional controls were also added to the resulting aRNA (Poly-A RNA) before hybridizations.
2.5. Microarray Data Processing and Analyzes
The expression data were first subjected to a series of rigorous quality control steps to ensure data reproducibility and overall quality, following general Affymetrix guidelines (Affymetrix, Santa Clara, CA). All computations associated to the microarray data were performed using the R computational and statistical language (http://cran.r-project.org), Version 2.15.0. Raw intensity values were processed first by RMA (Robust Multi-array Average) [49]. Upon application of pre-processing and normalization, all 108 arrays exhibited consistent expression distributions. To ensure strict reproducibility standards, all sets of triplicated expression measures were examined for the remaining probe sets. Any set of triplicates in which one of the measures exhibited a standard deviation of more than 1.14 (the maximum possible standard deviation for three measures is 1.1547), and a coefficient of variation in the upper 10 percentiles of all triplicate measures was scrutinized. If one single measure was near 1.1547, this indicated that the remaining two measures were very similar, and that the third triplicate was as its maximum outlying capacity, and thus this one triplicate was removed. This procedure left two replicates within the set of which the mean was used for subsequent analyses. Only 3.5 % of all measurements were excluded for this rule. Additionally, any remaining triplicates that exhibited a coefficient of variation of greater than 0.6 were removed. This included only 544 triplicates (0.2 % of all triplicated measurements) and these thresholds were used to identify gross outlying individual measurements within a triplicate set [50]. The Affymetrix control probe sets and the 4.371 (19 %) probe sets with three or less Present calls (as computed by the Affymetrix MAS 5 presence/absence detection algorithm) were excluded from further analysis.
Relative expression values of normalized and processed data of these 18.427 probe sets were then computed as log-transformed ratios (base 2) of each experimental treatment vs. the treatment’s control. A simple two-way fixed effect ANOVA (Analysis of Variance) was performed to examine probe sets with significant cultivar and developmental stage effects. ANOVA was performed separately for leaf under PEG treatment; leaf under salt treatment; root under PEG treatment; root under salt treatment. The false discovery rate (FDR) [51] multiple testing correction method was applied to p-values of the interaction effect F-statistics. The 1.368 probe sets with and adjusted interaction p-value of p < 0.001 were identified for further study. Further selection yielded a subset of 641 probe sets that had a differential expression of more than 2-fold between any two successive developmental states. This list is included as Table S1. These probe sets were clustered via simple agglomerative hierarchical clustering, using the average agglomerative method, and a function of the Pearson correlation coefficient as distance metric.
Gene ontology (GO) enrichment analyses for both stresses (PEG and salt) were performed using http://geneontology.org/ web server to assign probes to the domains, cellular component, molecular function and biological process. Our ontology analysis was performed considering fact that a probe could be assigned to more than one knowledge domain in the ontology [52,53].
For pathway analysis, protein ids corresponding to the gene ids in the chip from the annotation file (in all plant species) are obtained from the NCBI (National Center for Biotechnology Information) website. The protein list and corresponding (available) gene sequences were downloaded from the server as fasta files. Pathway analysis was performed in the KEGG (Kyoto Encyclopedia of Genes and Genomes) database for both stresses (PEG and salt) with this protein list and also selected different plant species (there was no barley). As it is known, KEGG is a database used to understand the functions of biological systems against stress.
2.7. Validation of Microarray Results Using qRT-PCR
Total RNA was extracted, and its integrity verified as described above. cDNA was synthesized using a Transcriptor first strand cDNA synthesis kit (Roche), according to the manufacturer’s instructions with 2 μg RNA reverse-transcribed. Primers for genes assayed by quantitative real-time reverse transcription (qRT-PCR) were selected using Primer3 software [54]. qRT-PCR reactions were prepared using a SYBR Green I Master (Roche) and performed using the LightCycler®480 Real Time PCR system (Roche). Gene expression was determined for triplicate technical replicates by using serial six dilutions (i.e. 1/10 to 1/100000) of control cDNA standard curves per gene. A Tubulin gene (Hordeum vulgare, alpha tubulin (tubA; α-tubulin); Gen bank accession number: U40042.1) was used for normalization (as a housekeeping gene) of qRT-PCR data. The specificity of qRT-PCR amplifications were checked by a melting curve analysis. All qRT-PCR reactions were repeated three times using biological replicates at each time point. Probe set IDs, primer sequences of related genes and fold induction information are given in Table S2. The relative expression levels (Ct: cycle threshold values) were calculated by using the REST 2009 online software according to the 2-ΔΔCT (the delta-delta-Ct or ddCt) algorithm [55].
3. Results
3.1. Optimizations of Stress Applications
In this study, we comparatively examined physiological and molecular responses of the two wild barley accessions (20-40 and 23-70) originated from a desert and xeric region of Israel, respectively). These two accessions are known to have good adaptation to extreme environmental conditions like high temperature, and salt and drought stress [56]. Of the two wild barley accessions, 20-40 was collected from SedeBoqer, the Northern Negev desert of Israel, which receives only 91 mm of average annual rainfall. Similarly, 23-70 was collected from Wadi Quilt located in the Judean desert region of Israel with very low annual precipitations [57,58,59].
For stress application, we mainly followed the procedure developed originally by Walia et al. [20] for barley. Briefly, salt or PEG were applied to 15-day-old seedlings, a plant development stage known to be highly susceptible to stress with serious consequences on yield [60]. Secondly, as shown in Figure 1, the intensity of salt and PEG stress were increased gradually so that the physiological effects of these stress treatments could be separated from those their “shock” causing effects [29]. As described in Materials and Methods, relatively lower concentrations of salt or PEG (25 mM salt and approximately 25 mM PEG) applied initially were gradually raised to 50 mM salt and 50 mM PEG in the second day and to 75 mM salt and 75 mM PEG on the third day of stress treatments. For leaf water potential measurements and gene expression analyses, plants grown under 100 mM salt or 100 mM PEG concentrations were used for RNA isolations at 0h, 3h and 27h of stress treatment. As described previously, the stress applied at this stage does not cause any visible symptom development (e.g. senescence, wilting or chlorosis) [60].
3.2. Leaf Water Potentials of Wild Barley Accessions Under Salt or PEG Stress
It is widely known that the stress caused by salt and PEG reduces water potential of plant tissues [61] and stress-tolerant genotypes show relatively smaller reductions in their leaf water potentials under stress than those of non-tolerant genotypes within the same species [28]. To determine whether the stress treatments worked as expected and whether the wild barley accessions used in this study differentially respond to salt and drought, we first measured the leaf water potential of the wild barley genotypes after salt or PEG treatment. In these experiments, the leaf water potential of 23-70 declined from -0.425 to -0.675 at 3 h and to -0.879 MPa at 27 h of salt stress. In contrast, the leaf water potential of 20-40 did not show any significant decrease in response to salt stress at either time-points examined (Figure 2A). However, leaf water potentials of both genotypes showed similar reductions at 3h and 27h after the PEG treatment and controls showed MPa values between -0.2 and -0.3. As shown in Figure 2B, the leaf water potential of 20-40 decreased from -0.410 to -0.846 at 3h, and to -0.91 at 27h while that of 23-70 decreased from -0.417 to -0.782 at 3h, and to -0.9 at 27h. Together, these experiments showed that wild barley accessions used in this study were responding to salt and PEG treatments. However, it is likely that the genotypes differ in their response to salt treatment with 20-40 being less sensitive to these treatments than 23-70.
3.3. Expression Profiling of Wild Barley Accessions Under Salt and PEG Stress
To determine the effect of stress on gene expression of the wild barley genotypes, we analyzed gene expression profiles following salt- or PEG-stress using the Affymetrix 22K Barley 1 chip. We have identified 641 transcripts with significantly different expression patterns across three time-points (0h, 3h and 27h) for at least one stress (PEG or salt)/tissue (root or leaf) combination in one genotype than the other in the ANOVA analysis. The annotations of these 641 genes were conducted according to "Barley1 Annotations, CSV format, Release 36 (1.8 MB, 4/13/16)" (http://www.affymetrix.com/Auth/analysis/ downloads/na36/ivt/Barley1.na36.annot.csv.zip) (Table S3). A clustering algorithm using the correlation across all 24 conditions was applied to these 641 genes to identify groups of genes with similar expression patterns. This analysis identified 15 major clusters (Figure S1). The genes with distinct expression patterns were mostly found in Cluster 3, Cluster 5 and Cluster 9. In fact, Cluster 9 and Cluster 1 include nearly half (50.3%) of the 641 differentially expressed genes (Figure S2).
Differentially expressed genes found in Cluster 9 were enriched for the following metabolic processes (18.46%), organic substance metabolic process (16.92%), localization (12,31%), cellular protein metabolic process (10.77%), cellular biosynthetic process (9,23%), gene expression (6,15%), oxidation-reduction process (6.15%), organic acid (4.62%), signal transduction (4,62%), cellular process (4.62%), cellular response to stimulus (3.08%), cellular component organization or biogenesis (3.08%) (Figure 3).
Interestingly, expression profiles trajectories observed for Cluster 1 (61 genes) (Figure S2B) were somewhat similar to Cluster 9 (263 genes) (Figure S2A) with a steady increase at transcript levels for the 3th hour salt and PEG treatment in 23-70. Together, these two inversely correlated clusters represent about 50.5% of all salt and drought responsive genes.
3.4. Validation of Microarray Results Using qRT-PCR
To validate the microarray data, expression profiles of selected genes up- or down-regulated in response to drought and salinity were analyzed by qRT-PCR. The qRT-PCR data obtained from these experiments were consistent with the expression ratios obtained from the Barley GeneChip® microarray for these selected genes (Table S2).
3.5. Biological Processes Impacted by Salinity and Drought Stress in Wild Barley
Among 641 differentially expressed genes identified in this study had identifiable homologies to entries in Barley1 Annotations, CSV format, Release 36. Of these 156 and 141 genes could be assigned a function based either on molecular function- (156 genes) or Biological process- (141 genes) based GO annotations as shown in Figures S3 and S4.
In Biological Process analyzes (Figure S3) it was determined that common transcripts whose expression changes in both salt and drought stress are associated with catabolic processes (15%), oxidation-reduction (12%) and organic substance metabolic process (9%) at high rates. Transcripts associated with oxidoreductase activity (13%) and nucleotide binding (13%) were significantly detected in Molecular Function analyzes (Figure S4).
We identified genes that were regulated by both stresses. These included genes encoding a 60 kDa jasmonate-induced protein (Contig3501_at), a cold-regulated protein (Contig4281_s_at), a thionin precursor (Contig1568_x_at), an abscisic acid- and stress-induced protein (Contig8961_at), a dehydrin 9 (Contig1718_s_at), a safener-induced In2.1-like protein (Contig6546_at), an oxalate oxidase (Contig3017_at), a physical impedance induced protein (Contig3783_at), a hypersensitive-induced reaction protein 3 (Contig3783_s_at), a cold acclimation protein WCOR413 (Contig3112_at), a peroxidase (rbah13p07_s_at), a putative I-box binding factor (Contig15729_at), a MAP kinase kinase (Contig15489_at), a putative heat shock protein (Contig17190_at), a MYB-related transcription factor (Contig3783_at), a putative receptor protein kinase ZMPK1 precursor (Contig18873_at), a TNP2-like protein (EBro03_SQ007_C24_at), a defensin (Contig3216_at), and a putative glutathione synthetase (Contig21604_at) (Table 1 and Table 2, Table S3).
The genes whose expression decreased under drought and salt stress encode a peroxidase (Contig2112_at, rbah13p07_s_at), a putative I-box binding factor (Contig15729_at), a germin-like protein, a C2-HC type zinc finger protein (Contig5588_at) and a hexose transporter-like protein (Contig9662_at). The genes whose expressions increased under both drought and salt stress encode a jasmonate-induced protein (Contig3501_at), a canalicular multispecific organic anion transporter 2-like protein (Contig6152_at), an aldehyde dehydrogenase family protein (Contig4991_s_at), and a P-glycoprotein, putative (Contig17453_at) (Table 1, Table 2, Table S3).
We performed KEGG pathway analysis of differentially expressed genes for both PEG and salt stress which were divided into different enriched categories, significantly including ribosome, plant hormone signal transduction, protein processing in endoplasmic reticulum, amino sugar and nucleotide sugar metabolism, oxidative phosphorylation, circadian rhythm–plant, plant-pathogen interaction and glycolysis/gluconeogenesis (Figure 4).
3.6. Comparison of Transcriptomes Between Genotypes, Tissues and Stress Treatments
We have compared the transcriptomes of two barley accessions as well as those of different tissues and treatments to identify differentially expressed genes (DEG) that were common to both genotypes. As shown in Figure 5, in response to salt treatment, we identified 195 DEG in the leaf tissue of 23-70 and 20-40 across all time points tested. Similarly, in response to PEG, 162 genes were differentially expressed in the leaves of these two accessions (Figure 5C). The numbers of differentially expressed genes in the roots of the accessions were much higher than those in the leaves. For example, 778 and 307 genes were differentially expressed between 23-70 and 20-40 roots across all three time points in response to salt or PEG treatments (Figure 5A,B) whereas the number of differentially expressed genes was only 195 and 162 for leaves.
Interestingly, largely different genes were differentially expressed between different tissues by the same treatment. For instance, only 5 and 6 differentially expressed genes were shared between PEG- and salt-stressed leaves and roots of 23-70 and 20-40, respectively (Figure 5C,D). Remarkably, there was very little overlap between salt and PEG treatments as far as differentially expressed genes in different tissues and accessions are concerned. For instance, only 8 differentially expressed genes in response to both salt and PEG treatments were shared in the leaves of the accessions across all time points. Similarly, out of 778 differentially expressed genes between 23-70 and 20-40 roots, 731 genes were differentially expressed only between the salt-stressed roots of 23-70 and 20-40. There were only 41 genes differentially expressed in the roots of the accessions in response to both salt and PEG treatment (Figure 5D).
4. Discussion
It is well-established that wild relatives of crop plants can harbor genes essential in biotic and abiotic stress tolerance [62]. Previous studies have investigated the effects of dehydration shock, drought, salinity and osmotic stress on the transcriptome of wild and cultivated barley genotypes mostly using single genotypes [28,29,30,31,63,64]. However, to the best of our knowledge, there has not been any report on the responses of different wild barley genotypes and different tissues to abiotic stress conditions.
To understand how wild barley genotypes, tolerate adverse environmental conditions, in this study, expression profiles of two wild barley accessions from Israel were studied under salt and drought stress. To obtain a comprehensive picture (especially common genes of drought and salinity stress), our gene expression analyses were conducted on different tissues (roots and leaves) and at different time-points after stress application in two separate wild barley accessions. In contrast, other studies on H. spontaneum have examined either a single genotype or a single tissue (i.e. mostly leaves) [28,31,63]. In this study, the comparison of differentially expressed genes between different tissues, treatments and accessions across different time points revealed interesting insights. Firstly, there was a very little overlap between the differentially expressed genes of each accessions by salt and PEG stress in both leaf and root tissue. This was unexpected, given that both salt and PEG stress are known to cause similar physiological effects [65]. However, it is likely that, in contrast to cultivated barley, wild barley accessions found even in close geographical locations can harbor significant levels of genetic diversity [66]. Supporting this hypothesis, Wang et al. [31] identified different sets up drought-associated genes and traits associated with drought tolerance from wild barley populations from Israel.
Secondly, larger number of genes were differentially expressed in the roots than in the leaves in response to stress treatments. Although the reasons for this are not known, one possibility would be that, in contrast to leaves, roots are directly affected by the stress treatment. In addition, under our experimental conditions, very little, if any overlap, was found between the gene expression profiles of root and leaves tissues. Finally, differentially expressed genes in each accession under stress conditions were largely different. Although both 20-40 and 23-70 were originated from dry environments and thus expected to show good adaptation to stress conditions, based on our leaf water potential measurements, it appears that 23-70 is more sensitive to salt than 20-40, suggesting that natural variation for salt stress might exist between the accessions despite their overall ability to adapt to adverse environmental conditions. This is consistent with our gene expression analyses, which showed large numbers of genes differentially expressed between these genotypes (Fig.7, Table S1).
4.1. Leaf Water Potentials of Wild Barley Accessions Under Salt or PEG Stress
In some studies, it is stated that the osmotic potential of the plant decreases at a high rate during the decrease of leaf water potential [67] while it is still unclear whether this is due to the high concentration of organic solutes or the change in the adaptation of the cellular environment through the plant development. On the other hand, osmotic adjustment under drought stress is thought to be a drought tolerance mechanism that causes a decrease in osmotic potential through the solute accumulation [68]. Studies [22,25,69] have proven that barley cultivars with high drought adaptation show smaller water potential reduction (-MPa). According to this, in drought stress treatments (7, 14, 21 and 28 days), drought tolerant “Rum” barley cultivar had lower water potential reduction (-1.8 MPa) than “Athroh” barley cultivar (-2.2 MPa) [22]. Similarly, under no watering conditions (25 days), water potential reduction was found to be lower in drought tolerant “Martin” (-1.5 MPa) than the sensitive barley cultivar “Roho” (-2.5 MPa) [69]. In current study, the range of -0.4 to -0.9 MPA values determined depending on the time points in both wild barley genotypes under drought stress is similar to the mentioned studies. However, significant reductions in water potential (WP) and osmotic potential were observed in Tibetan wild barley genotypes (4% soil moisture content, 30 days) in both drought-tolerant and drought-sensitive genotypes that these parameters did not make a clear distinction between genotypes in terms of drought tolerance feature [37].
In terms of salt stress, as it has been found in 23-70 wild barley accession, as water potential reduction is continued, the salt concentration increases in plant cells [70,71]. Since reduction in MPa values induces salt accumulation in the cell, higher decline is occurred in MPa values and hence, it is thought that cultivars salt sensitivity might be increased. On the other hand, in our study, in the short-term measurements were made 3 hours after NaCl treatment (75 mM) there was no significant changes in MPA values of 2 wild barley accessions (T1: wild barley line and 20-45: salt-sensitive line), similar to the 20-40 accession and the measurement results were reported to be similar [72]. In our study, 23-70 accession showed similar MPA values in both PEG and salt stress treatments between the 3h and 27h time points. Similarly, Ueada et al. [28]. showed that similar rates were observed in leaf water potential values under PEG (20% w/v) and salt stress (200 mM NaCl) during the first 10 hours of treatments and this confirms that barley plants are exposed to the same degree of water stress under both PEG and salt stress during the first 10 hours.
4.2. Expression Profiling of Wild Barley Accessions Under Salt and PEG Stress
The proteins associated with abiotic stress tolerance can be classified in three major groups [73]. The first group consists of heat shock proteins (HSPs), LEA proteins, osmoprotectans and antioxidant enzymes that protect plant cells against damaging effects [74]. The second group includes proteins involved in signaling and transcriptional control such as mitogen-activated protein kinases (MAPKs), Calcium-dependent protein kinases (CDPKs) [75] and SOS kinase [76], phospholipases [77] and transcription factors [78]. The third group includes the proteins involved in water an ion uptake and transport [79]. Most genes identified in our microarray fall into these general categories. In the following sections, various examples of these differentially expressed genes were briefly discussed.
Jasmonic acid (JA) is an important plant hormone associated with abiotic stress tolerance [80]. We identified differentially expressed genes involved in JA responses after salt treatment such as the genes encoding a jasmonate-induced protein and a thionin. The gene encoding a jasmonate-induced protein, 60kDa (JIPS60) (Contig3501_at) was induced in the leaves (3h, 27h) of both accessions. In barley, the JIPS60 gene plays a role as cellular messenger for translation in stress, and in recent studies, it has been stated that JIP60 may be a candidate Quantitative trait locus (QTL) for biotic and abiotic stress tolerance [81]. This gene was up-regulated in the roots of 23-70, proving that up-regulated JIP60 during salt stress inhibits plant growth and protects against stress [82,83].
Another gene encoding a thionin precursor (Contig1568_x_at) was up-regulated in the leaves (3h,27h) 23-70 by both stresses. Thionins are defined as components of plant immunity against environmental stress factors [84]. Our results are consistent with those by Gao et al. [85] who reported that the JA pathway was activated in the leaves of a salt tolerant barley genotype.
Abscisic acid (ABA) is another plant hormone associated with stress tolerance in plants. ABA levels increase under salt and drought stress in plants [86]. Our analysis identified a gene encoding an abscisic acid- and stress-induced protein (Contig8961_at) in the leaves of 20-40 by both salt (3h) and PEG (3h,27h). Interestingly, this gene was downregulated in 20-40 and 23-70 roots (3h) by salt. No alterations at the transcript levels of this gene in the leaves of 23-70 could be observed. However, this gene was down-regulated by both salt (3h) and PEG (3h, 27h) in the roots of 23-70. Abscisic acid responsive element-binding factor (ABF3) gene encodes the transcription factor associated with the expressions of ABA responsive genes and controls expression of stress-responsive genes [87]. Overexpression of ABF3 increases tolerance to drought stress [88,89]. It is hypothesized that, the drought tolerance provided by the overexpression of ABF3 is associated with the decrease in transpiration, the decrease in ROS accumulation and also with the increase in the amount of chlorophyll [89]. Similarly, in this study, abscisic acid responsive element binding factor (ABF3) (Contig15982_at) was induced in 20-40 leaves treated with PEG (3h). Also, several ABA signaling related genes associated with drought response were also identified from both cultivated [85] and other wild barley populations from Israel [31].
Dehydrins (DHNs) constitute a distinct biochemical group of Late Embryogenesis Abundant (LEA) proteins, which is known as group 2 LEA (or LEA II) proteins [90]. These proteins are known to accumulate during late embryogenesis or can be induced in vegetative tissues by drought, salinity, cold, and heat [91]. In studies, it has been reported that while dehydrin 9 gene is generally affected by drought stress, it has no expression changes in salt stress [92]. Interestingly, in our study, a gene encoding a dehydrin 9 protein (Contig1718_s_at) was induced in the leaves (3h,27h) and the roots (3h,27h) of 20-40 by both stresses. In a study conducted on two wild barley genotypes, it was reported that the gene expression of some dehydrin transcripts (Dehydrin 1,3,5,6,9) including the dehydrin 9 gene, could reveal quantitative differences according to differential drought tolerance [93]. On the other hand, it is also stated that the expression of especially Dehydrin 3 and Dehydrin 9 genes is positively correlated with osmotic regulation, chlorophyll a, chlorophyll b contents and plant biomass [94]. Similarly, in our study, this gene was induced only in the roots (27h) of 23-70 and also in the leaves (3h,27h) and the roots (3h,27h) of 20-40 treated with PEG.
ROS (reactive oxygen species) accumulate in plants as a result of stress exposure leading to cellular damage, if not removed timely [95]. Some peroxidases are involved in removing ROS while others can use ROS as substrate [95]. Peroxidase tends to activate related signal pathways in plant resistance, against biotic and abiotic stresses. Recent studies show that the interactions between these peroxidases with various antioxidant properties play a critical role in regulating stress responses [96]. Our analysis has identified several peroxidase encoding genes differentially expressed in either the roots or the leaves of the two wild barley accessions. For instance, a peroxidase (rbah13p07_s_at) encoding gene was down-regulated by salt treatment in the leaves of both (23-40:3h, 20-40:3h,27h) accessions. In addition, this gene was down-regulated by PEG in the leaves (3h,27h) of both accessions. Similarly, a gene encoding a peroxidase 2 precursor (Contig2112_at) was down-regulated in the leaves (3h) and roots (3h) of 23-40 and in the roots (3h) of 20-40 by PEG. Based on the drought stress researches, existence of peroxidase gene among the significant down regulated genes in leaf tissue of drought tolerant barley genotype (Otis) has been identified [97]. İn our work, down regulation of peroxidase genes in leaf tissues of both 20-40 and 23-70, under PEG treatment has shown parallel results with this research. According to many researches on salt stress [98,99] enzymatic and non enzymatic activity and expression of antioxidants has been shown to increase in salt stress resistant plants after rising the ROS following the salt treatment. Similarly, in a study [15] in Tunisian barley accessions (Boulifa: salt-tolerant and Testour: salt-sensitive) treated with 200mM salt stress (2, 8, and 24 h), the expression of several peroxidase genes increased in a genotype specific manner in leaf tissues.
In another study [100], it was reported that these protective enzyme activities produced against ROS are more in leaves than in roots, as water transpiration can be reduced by stomata adjusting in leaves against drought, while roots are more exposed to water stress than leaves. In our study, similar to the Ge et al. [100], it was revealed that a decrease in gene expression of protective enzymes could be observed as a result of disruptions in cell membranes. In this study, another gene encoding a putative peroxidase (Contig1865_at) was up-regulated in 23-70 leaves (3h) only by PEG, suggesting that similarly, several ROS-related genes associated with drought response were identified from other wild barley populations from Israel [31] and Egypt [63].
Germins and germin-like proteins (GLPs)/oxalate oxidase-like protein constitute a ubiquitous family of plant proteins that are involved in many developmental and abiotic stress-related processes [101] For instance, an oxalate oxidase (OxO) encoding gene (Contig3017_at) was upregulated in the roots (3h,27h) of both accessions by both stresses. In studies, it has been reported that abiotic stresses such as salt and heat can increase the expression and activation of oxalate oxidase in barley root tissue [102,103]. On the other hand, the knowledge that H2O2 production via oxalate oxidase is necessary for drought stress tolerance has also been reported [104]. Similarly, in other studies carried out on salt stress [105] and drought stress [106] in barley, it was also determined that oxalate oxidase genes are highly expressed in root tissue, and these results are similar to those found in root tissue.
Glutathione (GSH) acts as an anti-oxidant by quenching reactive oxygen species and is involved in the ascorbate–glutathione cycle that eliminates damaging peroxides. Glutathione synthetase is an enzyme involved in glutathione biosynthesis [107,108]. A gene encoding a putative glutathione synthetase (Contig21604_at) was down-regulated by both salt (23-70:3h, 20-40:3h,27h) and PEG (3h,27h) in both accessions while in 23-70 this gene was up-regulated by PEG (3h,27h). Glutathione transferases (GSTs) can also be induced by biotic or abiotic stresses [109] and plays a key role in the antioxidant enzyme system [110]. Our analysis identified several glutathione transferase encoding genes up-regulated in wild barley during stress. For instance, Glutathione-S-transferase 2 (Contig21640_at and Contig5838_at), Glutathione S-transferase GST 37 (Contig15282_at) were up-regulated in the roots (3h) of 23-70 by both stresses while in 20-40 roots (3h), Glutathione-S-transferase 2 (Contig21640_at) and Glutathione S-transferase GST 37 (Contig15282_at) were up-regulated by salt and PEG, respectively. Some studies have reported that, under ABA osmotic stress, glutathione S-transferase AtGSTU17 knockout mutants of Arabidopsis, show drought and salt stress tolerance [111]. In recent studies, it was determined that overexpression of the GST gene in Arabidopsis [112] and maize [110] significantly reduced drought tolerance, while overexpression of the 3 orthologous populus GST genes increased salt and drought tolerance in Arabidopsis [113]. From this point of view, in our study, the up regulation profile of the GST gene determined in root tissue did not show a clear distinction between 23-70 and 20-40, in terms of drought and salt stress tolerance.
Protein kinases are involved in protein phosphorylation events associated with signal transduction during stress adaptation [114]. Protein kinases can alter the amino acids activities by catalyzing the γ-phosphate transfer from adenosine triphosphate (ATP) to a specific amino acid. These findings suggest that protein kinases are involved in many aspects of cellular regulation and metabolism [115]. A gene encoding a putative leucine rich repeat containing protein kinase (Contig24926_at) was up-regulated in the leaves (27h) of 20-40 by salt.In the study on drought-tolerant (Otis) and drought-sensitive (Baroni) genotypes of barley, similar to the results of our study, it was emphasized that under drought stress, protein kinases (serine/threonine protein kinase, leucine-rich receptor-like protein kinase family protein isoform) are common reduced genes in the leaf tissue of both genotypes and also it was reported that 6 different protein kinase genes were significantly down-regulated in drought-sensitive ʺBaronesseʺ [97]. Similarly, a gene encoding a serine/threonine kinase-like protein (Contig 25448_at) was down regulated in the leaves (3h,27h) of 23-70 by PEG. In the same study [97], it was described that many protein kinases were down-regulated in Baronesse compared to Otis, and protein kinase genes may be one of the candidate factors related to drought tolerance. From this point of view, it can be considered that the drought tolerance of 20-40 is higher than that of 23-70. Dehydration and high salinity were reported to induce the expression of SPK3 (serine/threonine protein kinase 3) [116]. Our analysis also identified a gene encoding a putative protein kinase SPK-3 (HR01N22u_s_at) up-regulated in the leaves (3h,27h) of 20-40 by PEG. In addition, a gene encoding a serine/threonine kinase-like protein (Contig 25448_at) was down-regulated in the leaves (3h,27h) of 23-70 by PEG. In another study [117], conducted to detect important genes for drought tolerance in barley through transcriptomic data mining, it was emphasized that the serine/threonine protein kinase/threonine-specific protein kinase gene is among the top candidate genes.
In recent studies in plants, it has been reported that MAPK (Mitogen-activated protein kinase) cascades are involved in the regulation of many biological processes such as cell division, plant growth, and hormonal response, as well as in the response to stress factors such as drought, salt, heat and pathogen infection [118]. We identified a gene encoding a MAP kinase (Contig15489_at) down-regulated by both stresses in the leaves (3h) of 23-70 and in the roots (3h) of 20-40. Dudziak et al. [119] reported that some MAPK genes were significantly expressed in short-term (3h and 6h) under drought stress. The MAPK gene expression in both stresses in our study supports these studies. Similarly, several genes encoding protein-phosphorylation-related proteins that may be associated with drought tolerance were also identified from other wild barley populations from Israel [31].
Transcription factors (TFs) that control gene expression under stress conditions play an important role during stress adaptation. For instance, the bZIP TFs regulate several biochemical processes that protect plants under drought and high salinity [120]. Basic leucine zippers (bZIPs) have been shown to confer stress resistance via triggering expression of stress-responsive genes, especially by binding to ABA-responsive elements in their promoters [120,121]. Abscisic Acid Responsive Elements-Binding Factor (ABF3) is a bZIP protein that regulates key aspects of plant seed development and abscisic acid signaling (Kang et al. 2002). In many studies, it has been reported that the ABF3 gene is induced by ABA application and osmotic stress, especially in vegetative organs [122,123], and the ABF3 gene activates the relevant stress responsive genes by binding to the ABA-responsive element. ABF3 overexpression in alfafa [89] and rice [89] increased drought tolerance. Parallel to these studies, our analyses identified an ABF3-like gene (Contig15982_at) up-regulated in 20-40 leaves (3h) by PEG. As another transcription factor, MYB transcription factors are implicated in stress and developmental responses [125]. Overexpression of OsMYB3R-2 increased tolerance to multiple abiotic stresses in Arabidopsis [126], while heterologous expression of TaMYB3R1 increased drought and salt tolerance in transgenic Arabidopsis plants [127]. In recent years, it has been reported that ectopic expression of ZmMYB3R in Arabidopsis can lead to higher ABA content, increasing stomatal closure to minimize water loss, thereby increasing stress tolerance [125]. A gene encoding a MYB-related transcription factor (Contig3875_s_at) was down-regulated by both stresses (3h) only in 20-40.
DnaJ gene expression has been shown as a potential index of drought tolerance [128]. DnaJ proteins, also known as heat shock protein 40 (Hsp40), are important partners of Hsp70 proteins [128]. DnaJ (Hsp40), a heat shock protein, is a molecular chaperone responsive to various environmental stresses [129]. DnaK (Hsp70) proteins are also induced by changing various environmental stresses and believed to function in the protection and recovery of cells from the ill effects of stress [130]. A gene encoding a DnaK-type molecular chaperone HSP70 (L32165_s_at) was up-regulated in the roots (3h,27h) of 23-70 by both stresses. Similarly, Isayenkov et al. [131]. found that some DnaJ genes were up-regulated in response to both salt and PEG stress in barley (Hordeum marinum ssp. marinum) roots. Another gene encoding a DnaJ protein putative (Contig11487_at) was down-regulated both in the leaves (3h) and the roots (3h) of 23-70 by both stresses while a gene encoding another Chaperone protein DnaJ 3 (Contig632_s_at) was down-regulated only in the leaves (3h) of 23-70.
Expression of cold-regulated (COR) genes in plants play a critical role in cold stress tolerance and adaptation to cold stress. However, these COR genes (COR78/RD29A, COR47, COR15a) in Arabidopsis plant are also induced by dehydration (drought, salt stress) or ABA [132]. In a study on tomato leaves, it was reported that the COR gene (LeCOR413PM2) increased at different levels in response to salt, drought and cold stress [133]. In the same study, it was reported that overexpression of the COR gene not only reduces cell membrane damage in response to stress, but also preserves ROS accumulation, high activity of antioxidant enzymes and content of osmotic regulators [133]. Similarly, the expression of a gene encoding a cold-regulated protein (COR) was reported to increase in response to drought stress [134]. Parallel to these studies, we identified a gene encoding a cold-regulated protein (Contig4281_s_at) up-regulated in the leaves of 23-70 by both stresses and in the roots by PEG only. This gene was also up-regulated in the leaves of 20-40 by PEG.
5. Conclusion
İn this study, we comparatively analyzed the stress-associated transcriptomes of the two wild barley accessions under salinity and drought stress imposed gradually to mimic the effect of natural stresses. A general data summary in our study revealed that most of the genes expressed at the 3h time point, regardless of stress type, continued to be expressed at the 27h time point of treatment. In 27h time point of PEG application, especially Dehydrin 9, Cold-regulated protein, Putative heat shock protein and Germin-like protein genes were determined to be expressed only in root tissue while, SERK2 protein and Safener-induced In2.1-like protein genes were expressed only in leaf tissue at this time point. In salt stress, especially Safener-induced in 2.1-like protein, Glutathione-S-transferase 2, Cold acclimation protein WCOR413, Signal Recognition Particle 54 KD Protein3 (SRP54), Abscisic acid- and stress-induced protein and TNP2-like protein genes were expressed only in the root tissue at the 3h time point. These different gene expression profiles determined for leaf and root tissue, similar to other studies [135], prove that there are differences in the regulatory mechanisms involved in these tissues.
Another important finding is that some genes are co-expressed with both PEG and salt stress, regardless of tissue. Accordingly, it was determined that especially Thionine precursor, 60 kDa jasmonate-induced protein, Putative glutathione synthetase, Abscisic acid- and stress-induced protein, Cold-regulated protein, Abscisic acid responsive elements-binding factor (ABF3) genes were up-regulated in both PEG and salt stress while, Peroxidase MAP kinase, Abscisic acid- and stress-induced protein and MYB-related transcription factor genes were observed to be down-regulated. It is thought that these detected genes will help to identify common genes involved in drought and salt stress and obtaining the higher productivity in crop plants through genetic engineering.
In conclusion, we identified several novel salt and drought stress-related genes which have not been previously associated with these stresses, in addition, some important common genes (drought and salt) have been identified. Future functional analyses of these genes should reveal new insights into the molecular mechanisms underpinning stress tolerance in plants.
Supplementary Materials
Table S1: 641 probe sets that showed differential expression of 2-fold or more by salt and drought stress in two different time-points, Table S2: Validation of gene expression (fold-induction or repression relative to corresponding controls) obtained from Affymetrix array experiments by qRT-PCR for selected genes, Table S3: Probe set IDs, gene titles, gene symbols, target description, gene Ontology Biological Process, Gene Ontology (Cellular Component, Molecular Function) of 641 probe set that show differential expression in at least one comparison, Figure. S1: Assignment of 15 top-level clusters (marked by red lines) based on the expression profiles of 641 salt- and drought-responsive genes in 20-40 and 23-70 wild barley accessions (A). A heatmap of 15 clusters according to the 641 salt and drought responsive genes in 20-40 and 23-70 wild barley genotypes (X axis genotype-tissue-treatment-time) (B), Figure S2: Expression patterns of Cluster 9 (A) and Cluster 1 (B) which include 50.3% of all differentially expressed genes (641 genes) when all 24 conditions were considered, Figure S3: Gene ontology (GO) annotations (Biological Process) of salt and drought responsive probe sets on the microarray that had identifiable homologies to Affy annotation file, Figure S4: GO annotations (Molecular Function) of salt and drought responsive probe sets on the microarray that had identifiable homologies to Affy annotation file.
Author Contributions
Methodology, M.B., A.A.U., C.Y.Ö., F.Y.B. and U.K.; Investigation, M.B., A.A.U., C.Y.Ö., F.Y.B. and U.K.; Writing–original draft, A.A.U., K.K., C.Y.Ö., F.Y.B. and K.S.; Writing – review & editing, K.K., J.C.C. and A.E.; Software, U.K. and K.S.; Visualization, K.K., K.S., Validation, K.S.; Conceptualization, J.C.C and A.E.; Formal analysis, K.S.; Funding acquisition, A.E; Supervision, A.E.
Funding
This research was supported by the DPT (Ministry of Development of Turkey) project (Grant no: 2001 K 120240).
Data Availability Statement
All data are included in the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tattersall, E.A.; Grimplet, J.; DeLuc, L.; Wheatley, M.D.; Vincent, D.; Osborne, C.; Ergül, A.; Lomen, E.; Blank, R.R.; Schlauch, K.A.; et al. Transcript abundance profiles reveal larger and more complex responses of grapevine to chilling compared to osmotic and salinity stress. Funct. Integr. Genom. 2007, 7, 317–333. [Google Scholar] [CrossRef]
- Boscaiu, M.; Fita, A. Physiological and molecular characterization of crop resistance to abiotic stresses. Agronomy 2020, 10, 1308. [Google Scholar] [CrossRef]
- Munns, R. Plant Adaptations to Salt and Water Stress: Differences and Commonalities, Adv. Bot. Res. 2011, 57, 2–32. [Google Scholar]
- Shehzad, M.A.; Nawaz, F.; Ahmad, F.; Ahmad, N.; Masood, S. Protective effect of potassium and chitosan supply on growth, physiological processes and antioxidative machinery in sunflower (Helianthus annuus L.) under drought stress. Ecotoxicol Environ. Saf. 2020, 187, 109841. [Google Scholar] [CrossRef]
- Iwuala, E.; Odjegba, V.; Sharma, V.; Alam, A. Drought stress modulates expression of aquaporin gene and photosynthetic efficiency in Pennisetum glaucum (L.) R. Br. genotypes. Curr. Plant Biol 2019, 21, 100131. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought stress in plants: causes, consequences, and tolerance. In Drought Stress Tolerance in Plants; Hossain, M., Wani, S., Bhattacharjee, S., Burritt, D., Tran, LS., Eds.; Springer: Cham, 2016; Vol 1, pp. 1–16. [Google Scholar]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Kulkarni, K.P.; Imran, Q.M.; Mun, B.G.; Yun, B.W. Nitric oxide regulates plant responses to drought, salinity, and heavy metal stress. Environ. Exp. Bot. 2019, 161, 120–133. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, H.; Chen, T.; Pen, J.; Yu, S.; Zhao, X. Morphological and physiological responses of cotton (Gossypium hirsutum L.) plants to salinity. PLoS One 2014, 9, e112807. [Google Scholar] [CrossRef]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic stress in crop species: improving tolerance by applying plant metabolites. Plants 2021, 10, 186. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, J.G. Osmotic adjustment and plant adaptation to environmental changes related to drought and salinity. Environ. Rev. 2010, 18, 309–319. [Google Scholar] [CrossRef]
- Bray, E.A. Plant Responses to Water Deficit. Trends Plant Sci. 1997, 2, 48–54. [Google Scholar] [CrossRef]
- Hernández, J.A. Salinity tolerance in plants: Trends and perspectives. Int. J. Mol Sci. 2019, 20, 2408. [Google Scholar] [CrossRef]
- Nefissi Ouertani, R.; Arasappan, D.; Ruhlman, T.A.; Ben Chikha, M.; Abid, G.; Mejri, S.; Ghorbel, A.; Jansen, R.K. Effects of salt stress on transcriptional and physiological responses in barley leaves with contrasting salt tolerance. Int. J. Mol. Sci. 2022, 23, 5006. [Google Scholar] [CrossRef]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Naghdyzadegan Jahromi, M.; Razzaghi, F.; Zand-Parsa, S. Strategies to increase barley production and water use efficiency by combining deficit irrigation and nitrogen fertilizer. Irrig. Sci. 2022, 41, 261–275. [Google Scholar] [CrossRef]
- Al-Karaki, G.N. Germination, sodium, and potassium concentrations of barley seeds as influenced by salinity. J. Plant Nutr. 2001, 24, 511–522. [Google Scholar] [CrossRef]
- Walia, H.; Wilson, C.; Wahid, A.; Condamine, P.; Cui, X.; Close, T.J. Expression analysis of barley (Hordeum vulgare L.) during salinity stress. Funct. Integr. Genom. 2006, 6, 143–156. [Google Scholar] [CrossRef]
- Samarah, N.H. Effects of drought stress on growth and yield of barley. Agron. Sustain Dev. 2005, 25, 145–149. [Google Scholar] [CrossRef]
- Samarah, N.H.; Alqudah, A.M.; Amayreh, J.A.; McAndrews, G. M. The effect of late-terminal drought stress on yield components of four barley cultivars. J. Agron. Crop Sci. 2009, 195, 427–441. [Google Scholar] [CrossRef]
- Jamshidi, A.; Javanmard, H.R. Evaluation of barley (Hordeum vulgare L.) genotypes for salinity tolerance under field conditions using the stress indices. Ain Shams Eng. 2018, J9, 2093–2099. [Google Scholar] [CrossRef]
- Hamzeh-Kahnoji, Z.; Ebrahimi, A.; Sharifi-Sirchi, G.R.; Majidi-Hervan, E. Monitoring of morphological, biochemical and molecular responses of four contrasting barley genotypes under salinity stress. J. Saudi Soc. Agric. Sci. 2022, 21, 187–196. [Google Scholar] [CrossRef]
- Cramer, G.R.; Epstein, E.; Lauchli, A. Effects of sodium, potassium and calcium on salt-stressed barley. Physiol. Plant 1990, 80, 83–88. [Google Scholar]
- Munns, R.; Rawson, H.M. Effect of salinity on salt accumulation and reproductive development in the apical meristem of wheat and barley. Aust. J. Plant Physiol. 1999, 26, 459–464. [Google Scholar] [CrossRef]
- Forster, B.P.; Russell, J.R.; Ellis, R.P.; Handley, L.L.; Robinson, D.; Hackett, C.A.; Nevo, E.; Waugh, R.; Gordon, D.C.; Keıth, R.; et al. Locating genotypes and genes for abiotic stress tolerance in barley: a strategy using maps, markers and the wild species. New Phytol. 1997, 137, 141–147. [Google Scholar] [CrossRef]
- Ueda, A.; Kathiresan, A.; Inada, M.; Narita, Y.; Nakamura, T.; Shi, W.; Takabe, T.; Bennett, C. Osmotic stress in barley regulates expression of a different set of genes than salt stress does. J. Exp. Bot. 2004, 55, 2213–2218. [Google Scholar] [CrossRef] [PubMed]
- Talame, V.; Ozturk, N.Z.; Bohnert, H.J.; Tuberosa, R. Barley transcript profiles under dehydration shock and drought stress treatments: a comparative analysis. J. Exp. Bot. 2007, 58, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Zhou, L.; Zhang, Z.; Xian, W.; Han, B. Effect of water stress on physiological and growth charaters of Prunella vulgaris at the vegetative stage. China. J. Chin. Mater. Med. 2009, 34, 1761–1764. [Google Scholar]
- Wang, X.; Chen, Z.H.; Yang, C.; Zhang, X.; Jin, G.; Chen, G.; Wang, Y.; Holford, P.; Nevo, E.; Zhang, G. Genomic adaptation to drought in wild barley is driven by edaphic natural selection at the Tabigha Evolution Slope. PNAS 2018, 115, 5223–5228. [Google Scholar] [CrossRef]
- Nevo, E. Origin, evolution, population genetics and resources for breeding of wild barley, Hordeum spontaneum, in the Fertile Crescent. In: Shewry P (ed) Barley: genetics, biochemistry, molecular biology and biotechnology. CAB International, Wallingford, UK, 1992, pp. 19–43.
- Liang, J.; Chen, X.; Deng, G.; Pan, Z.; Zhang, H.; Li, Q.; Yang, K.; Long, H.; Yu, M. Dehydration induced transcriptomic responses in two Tibetan hulless barley (Hordeum vulgare var. nudum) accessions distinguished by drought tolerance. BMC Genom. 2017, 11, 775. [Google Scholar] [CrossRef] [PubMed]
- Gürel, F.; Öztürk, N.Z.; Yörük, E.; Uçarlı, C.; Poyraz, N. Comparison of expression patterns of selected drought-responsive genes in barley (Hordeum vulgare L.) under shock-dehydration and slow drought treatments. Plant Growth Regul. 2016a, 80, 183–193. [Google Scholar] [CrossRef]
- Gürel, F.; Öztürk, N.Z; Uçarlı, C.; Rosellini, D. Barley genes as tools to confer abiotic stress tolerance in crops. Front. Plant Sci. 2016b, 7, 1137. [Google Scholar] [CrossRef]
- Daszkowska-Golec, A.; Collin, A.; Sitko, K.; Janiak, A.; Kalaji, H.M.; Szarejko, I. Genetic and physiological dissection of photosynthesis in barley exposed to drought stress. Int. J. Mol. Sci. 2019, 20, 6341. [Google Scholar] [CrossRef]
- Zhang, M.; Jin, Z.Q.; Zhao, J.; Zhang, G.; Wu, F. Physiological and biochemical responses to drought stress in cultivated and Tibetan wild barley. Plant Growth Regul. 2015, 75, 567–574. [Google Scholar] [CrossRef]
- Ellis, R.P.; Forster, B.P.; Robinson, D.; Handley, L.L.; Gordon, D.C.; Russell, J.R.; Powell, W. Wild barley: a source of genes for crop improvement in the 21st century? J. Exp. Bot. 2000, 51, 9–17. [Google Scholar] [CrossRef]
- Zhao, J.; Sun, H.; Dai, H.; Zhang, G.; Wu, F. Difference in response to drought stress among Tibet wild barley genotypes. Euphytica 2010, 172, 395–403. [Google Scholar] [CrossRef]
- Barati, M.; Majidi, M.M.; Mirlohi, A. Response of cultivated and wild barley germplasm to drought stress at different developmental stages. Crop. Sci. 2015, 55, 2668–2681. [Google Scholar] [CrossRef]
- Barati, M.; Majidi, M.M.; Mostafavi, F.; Mirlohi, A.; Safari, M.; Karami, Z. Evaluation of wild barley species as possible sources of drought tolerance for arid environments. Plant Genet. Resour. 2017, 16, 209–217. [Google Scholar] [CrossRef]
- Cai, K.; Chen, X.; Han, Z.; Wu, X.; Zhang, S.; Li, Q.; Nazir, M.M.; Zhang, G.; Zeng, F. Screening of worldwide barley collection for drought tolerance: The assessment of various physiological measures as the selection criteria. Front. Plant Sci. 2020, 11, 1159. [Google Scholar] [CrossRef] [PubMed]
- Rensink, W.A.; Buell, C.R. Microarray expression profiling resources for plant genomics. Trends Plant Sci. 2005, 10, 603–609. [Google Scholar] [CrossRef]
- Walia, H.; Wilson, C.; Timothy, J. Comparative Transcriptional Profiling of Barley Cultivar Maythorpe and Its Derived Mutant Golden Promise under Salinity Stress. In Plant & Animal Genome XIII International Conference, San Diego, CA. 2005, 234.
- Wu, D.; Cai, S.; Chen, M.; Ye, L.; Chen, Z.; Zhang, H.; Dai, F.; Wu, F.; Zhang, G. Tissue metabolic responses to salt stress in wild and cultivated barley. PLoS one 2013, 8, e55431. [Google Scholar] [CrossRef]
- Bedada, G.; Westerbergh, A.; Müller, T.; Galkin, E.; Bdolach, E.; Moshelion, M.; Fridman, E.; Schmid, K.J. Transcriptome sequencing of two wild barley (Hordeum spontaneum L.) ecotypes differentially adapted to drought stress reveals ecotype-specific transcripts. BMC Genom. 2014a, 15, 1–20. [Google Scholar] [CrossRef]
- Bahieldin, A.; Atef, A.; Sabir, J.S.; Gadalla, N.O.; Edris, S.; Alzohairy, A.M.; Radhwan, N.A.; Baeshen, M.N.; Ramadan, A.M.; Eissa, H.F.; et al. RNA-Seq analysis of the wild barley (H. spontaneum) leaf transcriptome under salt stress. CR. Biol. 2015, 338, 285–97. [Google Scholar] [CrossRef]
- Cramer, G.R.; Läuchli, A.; Epstein, E. Effects of NaCl and CaCl2 on ion activities in complex nutrient solutions and root growth of cotton. Plant Physiol. 1986, 81, 792–797. [Google Scholar] [CrossRef]
- Irizarry, R.A.; Hobbs, B.; Beazer-barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization and summaries of high density oligonucleotide array probe level data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Aw, T.; Schlauch, K.; Keeling, C.I.; Young, S.; Bearfield, J.C.; Blomquist, G.J.; Tittiger, C. Functional genomics of mountain pine beetle (Dendroctonus ponderosae) midguts and fat bodies. BMC Genom. 2010, 11, 1–12. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. Roy. Statist. Soc. Ser. 1995, B57, 289–300. [Google Scholar] [CrossRef]
- Gene Ontology Consortium. Gene ontology consortium: going forward. Nucleic Acids Res. 2015, 43, D1049–D1056. [Google Scholar] [CrossRef]
- Gene Ontology Consortium. Gene Ontology annotations and resources. Nucleic Acids Res. 2012, 41, D530–D535. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. In Bioinformatics methods and protocols, Humana Press, Totowa, N.J. 200, pp. 365–386.
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lavie, B.; Stow, V.; Krugman, T.; Beiles, A.; Nevo, E. Fitness in wild barley from two opposing slopes of a Mediterranean microsite at Mount Carmel, Israel. Barley Genetics Newslett. 1993, 23, 12–14. [Google Scholar]
- Nevo, E.; Beiles, A.; Gutterman, Y.; Storch, N.; Kaplan, D. Genetic resources of wild cereals in Israel and vicinity. I. Phenotypic variation within and between populations of wild wheat, Triticum dicoccoides. Euphytica 1984, 33, 717–735. [Google Scholar] [CrossRef]
- Chen, G.; Krugman, T.; Fahima, T.; Zhang, F.; Korol, A.B.; Nevo, E. Differential patterns of germination and desiccation tolerance of mesic and xeric wild barley (Hordeum spontaneum L.) in Israel. J. Arid Environ. 2004, 56, 95–105. [Google Scholar] [CrossRef]
- Lupu, A.; Nevo, E.; Zamorzaeva, I.; Korol, A. Ecological–genetic feedback in DNA repair in wild barley, Hordeum spontaneum. Genetica 2006, 127, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Walia, H.; Wilson, C.; Condamine, P.; Liu, X. , Ismail, A. M.; Close, T.J. Large-scale expression profiling and physiological characterization of jasmonic acid-mediated adaptation of barley to salinity stress. Plant Cell Environ. 2007, 30, 410–421. [Google Scholar] [CrossRef]
- Cramer, G.R.; Ergül, A.; Grimplet, J.; Tillett, R.L.; Tattersall, E.A.R.; Bohlman, M.C.; Vincent, D.; Sonderegger, J.; Evans, C.; Osborne, C.; et al. Water and salinity stress in grapevines: early and late changes in transcript and metabolite profiles. Funct. Integr. Genom. 2007, 7, 111–134. [Google Scholar] [CrossRef]
- Boscai, M.; Donat, P.M.; Llınares, J.; Vicent, P.; Vicente Meana, Ó. Stress-tolerant Wild Plants: a Source of Knowledge and Biotechnological Tools for the Genetic Improvement of Stress Tolerance in Crop Plants. Not. Bot. Hortı. Agrobo. 2012, 40, 323–327. [Google Scholar] [CrossRef]
- Ashoub, A.; Müller, N.; Jiménez-Gómez, J.M.; Brüggemann, W. Prominent alterations of wild barley leaf transcriptome in response to individual and combined drought acclimation and heat shock conditions. Physiol. Plant 2018, 163, 18–29. [Google Scholar]
- Zhang, M.; Fu, M.M.; Qiu, C.W.; Cao, F.; Chen, Z.H.; Zhang, G.; Wu, F. Response of tibetan wild barley genotypes to drought stress and identification of quantitative trait loci by genome-wide association analysis. Int. J. Mol. Sci. 2019, 20, 791. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Bedada, G.; Westerbergh, A.; Nevo, E.; Korol, A.; Schmid, K.J. DNA sequence variation of wild barley Hordeum spontaneum (L.) across environmental gradients in Israel. Heredity 2014b, 112, 646–655. [Google Scholar] [CrossRef]
- Turner, N.C. Turgor maintenance by osmotic adjustment: 40 years of progress. J. Exp. Bot. 2018, 69, 3223–3233. [Google Scholar] [CrossRef] [PubMed]
- Nio, S.A.; Ludong, D.P.M.; Wade, L.J.J.A. Comparison of leaf osmotic adjustment expression in wheat (Triticum aestivum L.) under water deficit between the whole plant and tissue levels. Agric. Nat. Resour. 2018, 52, 33–38. [Google Scholar] [CrossRef]
- Naceur, B.; Paul, R.; Ben Salah, H. Screening of six barley varieties for drought resistance by using leaf water potential, membrane stability, and phosphatasic activity parameters. Prospettive e proposte mediterranee-Rivista di Economia. Agricoltura e Ambiente 1997, 4, 51–54. [Google Scholar]
- González, A.; Martı́n, I.; Ayerbe, L. Barley yield in water-stress conditions.: The influence of precocity, osmotic adjustment and stomatal conductance. Field Crops Res. 1999, 62, 23–34. [Google Scholar] [CrossRef]
- Nayyar, H.; Walia, D.P. Genotypic variation in wheat in response to water stress and abscisic acid-induced accumulation of osmolytes in developing grains. J. Agron. Crop Sci. 2004, 190, 39–45. [Google Scholar] [CrossRef]
- Vysotskaya, L.; Hedley, P.E.; Sharipova, G.; Veselov, D.; Kudoyarova, G.; Morris, J.; Jones, H.G. Effect of salinity on water relations of wild barley plants differing in salt tolerance. AoB Plants 2010, 2010, plq006. [Google Scholar] [CrossRef]
- Ciarmiello, L.F.; Woodrow, P.; Fuggi, A.; Pontecorvo, G.; Carillo, P. Plant Genes for Abiotic Stress, Abiotic Stress in Plants - Mechanisms and Adaptations. In Arun Shanker, B. Venkateswarlu (eds) 2011, 978–953.
- Wang, W.X.; Vinocur, B.; Shoseyov, O.; Altman, A. Biotechnology of plant osmotic stress toelrance and physiological and moleucalr considerations. Acta Hort. 2001, 560, 285–292. [Google Scholar] [CrossRef]
- Ludwig, A.A.; Romeis, T.; Jones, J.D. CDPK-mediated signalling pathways: specificity and cross-talk. J. Exp. Bot. 2004, 55, 181–188. [Google Scholar] [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Frank, W.; Munnik, T.; Kerkmann, K.; Salamini, F.; Bartels, D. Water deficit triggers phospholipase D activity in the resurrection plant Craterostigma plantagineum. Plant Cell 2000, 12, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular responses to dehydration and low temperature: differences and cross-talk between two stress signaling pathways. Curr. Opin. Plant Biol. 2000, 3, 217–223. [Google Scholar] [CrossRef]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Kazan, K. Diverse roles of jasmonates and ethylene in abiotic stress tolerance. Trends Plant Sci. 2015, 20, 219–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Stevanato, P.; Lv, C.; Li, R.; Geng, G. Comparative physiological and proteomic analysis of two sugar beet genotypes with contrasting salt tolerance. J. Agric. Food Chem. 2019, 67, 6056–6073. [Google Scholar] [CrossRef]
- Görschen, E.; Dunaeva, M.; Hause, B.; Reeh, I.; Wasternack, C.; Parthier, B. Expression of the ribosome-inactivating protein JIP60 from barley in transgenic tobacco leads to an abnormal phenotype and alterations on the level of translation. Planta 1995, 202, 470–478. [Google Scholar]
- Szypulska, E.; Jankowski, K.; Weidner, S. ABA pretreatment can limit salinity-induced proteome changes in growing barley sprouts. Acta Physiol. Plant. 2017, 39, 1–18. [Google Scholar] [CrossRef]
- Barashkova, A.S.; Sadykova, V.S.; Salo, V.A.; Zavriev, S. K.; Rogozhin, E.A. Nigellothionins from black cumin (Nigella sativa l.) seeds demonstrate strong antifungal and cytotoxic activity. Antibiotics 2021, 10, 166. [Google Scholar] [CrossRef]
- Gao, R.; Duan, K.; Guo, G.; Zhizhao, D.; Zhiwei, C.; Liang, L.; Ting, H.; Ruiju, L.; Jianhua, H. Comparative Transcriptional Profiling of Two Contrasting Barley Genotypes under Salinity Stress during the Seedling Stage. Int. J. Genomics 2013, 139, 822–835. [Google Scholar] [CrossRef]
- Lim, J.; Lim, C.W.; Lee, S.C. Core Components of Abscisic Acid Signaling and Their Post-Translational Modification. Front. Plant Sci. 2022, 13, 895698. [Google Scholar] [CrossRef] [PubMed]
- Collin, A.; Daszkowska-Golec, A.; Szarejko, I. Updates on the role of abscisic acid insensitive 5 (abi5) and abscisic acid-responsive element binding factors (ABFs) in ABA signaling in different developmental stages in plants. Cells 2021, 10, 1996. [Google Scholar] [CrossRef]
- Vanjildorj, E.; Bae, T.W.; Riu, K.Z.; Kim, S.Y.; Lee, H.Y. Overexpression of Arabidopsis ABF3 gene enhances tolerance to drought and cold in transgenic lettuce (Lactuca sativa). PCTOC 2005, 83, 41–50. [Google Scholar] [CrossRef]
- Wang, Z.; Su, G.; Li, M.; Ke, Q.; Soo Young, K.; Hongbing, L.; Jin, H.; Bingcheng, X.; Xi-Ping, D.; Sang-Soo, K. Overexpressing Arabidopsis ABF3 increases tolerance to multiple abiotic stresses and reduces leaf size in alfalfa. Plant Physiol. Biochem. 2016, 109, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Ingram, J.; Bartels, D. The molecular basis of dehydration tolerance in plants. Annu. Rev. Plant Biol. 1996, 47, 377–403. [Google Scholar] [CrossRef]
- Hanin, M.; Brini, F.; Ebel, C.; Toda, Y.; Takeda, S.; Masmoudi, K. Plant dehydrins and stress tolerance: versatile proteins for complex mechanisms. Plant Signal Behav. 2011, 6, 1503–1509. [Google Scholar] [CrossRef]
- Binott, J.J.; Owuoche, J.O.; Bartels, D. Physiological and molecular characterization of Kenyan barley (Hordeum vulgare L.) seedlings for salinity and drought tolerance. Euphytica 2017, 213, 1–23. [Google Scholar] [CrossRef]
- Suprunova, T.; Krugman, T.; Fahima, T.; Chen, G.; Shams, I.; Korol, A.; Nevo, E. Differential expression of dehydrin genes in wild barley, Hordeum spontaneum, associated with resistance to water deficit. Plant Cell Environ. 2004, 27, 1297–1308. [Google Scholar] [CrossRef]
- Kosová, K.; Vítámvás, P.; Prášil, I.T. Wheat and barley dehydrins under cold, drought, and salinity–what can LEA-II proteins tell us about plant stress response? Front. Plant Sci. 2014, 5, 343. [Google Scholar] [CrossRef]
- Sewelam, N.; Kazan, K.; Schenk, P.M. Global Plant Stress Signaling: Reactive Oxygen Species at the Cross-Road. Front. Plant Sci. 2016, 23, 187. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Kim, H.S.; Park, S.C.; Ji, C.Y.; Yang, J.W.; Lee, H.U.; Kwak, S.S. Downregulation of swpa4 peroxidase expression in transgenic sweet potato plants decreases abiotic stress tolerance and reduces stress-related peroxidase expression. Plant Biotechnol. Rep. 2021, 15, 69–76. [Google Scholar] [CrossRef]
- Harb, A.; Simpson, C.; Guo, W.; Govindan, G.; Kakani, V.G.; Sunkar, R. The effect of drought on transcriptome and hormonal profiles in barley genotypes with contrasting drought tolerance. Front. Plant Sci. 2020, 11, 618491. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Ge, T.; Suı, F-G. ; Baı, L. et al. Effects of Water Stress on the Protective Enzyme Activities and Lipid Peroxidation in Roots and Leaves of Summer Maize. Agric. Sci. China 2006, 5, 291–298. [Google Scholar] [CrossRef]
- Anum, J.; O’Shea, C.; Zeeshan Hyder, M.; Farrukh, S.; Skriver, K.; Malik, S.I.; Yasmin, T. Germin like protein genes exhibit modular expression during salt and drought stress in elite rice cultivars. Mol. Biol. Rep. 2022, 49, 293–302. [Google Scholar] [CrossRef]
- Hurkman, W.; Tanaka, C.K. Efect of salt stress on germin gene expression in barley roots. Plant Physiol. 1996, 110, 971–7. [Google Scholar] [CrossRef]
- Valentovičová, K.; Halušková, Ľ.; Huttová, J.; Mistrík, I.; Tamás, L. Efect of heavy metals and temperature on the oxalate oxidase activity and lignifcation of metaxylem vessels in barley roots. Environ. Exp. Bot. 2009, 66, 457–62. [Google Scholar] [CrossRef]
- Voothuluru, P.; Sharp, R.E. Apoplastic hydrogen peroxide in the growth zone of the maize primary root under water stress. I. Increased levels are specific to the apical region of growth maintenance. J. Exp. Bot. 2013, 64, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Ueda, A.; Shi, W.; Nakamura, T.; Takabe, T. Analysis of salt-inducible genes in barley roots by differential display. J. Plant Res. 2002, 115, 0119–0130. [Google Scholar] [CrossRef] [PubMed]
- Chmielewska, K.; Rodziewicz, P.; Swarcewicz, B.; et al. Analysis of drought-induced proteomic and metabolomic changes in barley (Hordeum vulgare L.) leaves and roots unravels some aspects of biochemical mechanisms involved in drought tolerance. Front. Plant Sci. 2016, 7, 1108. [Google Scholar] [CrossRef]
- Anders, J.V.; Davis, G.D. Abiotic stress alters transcript profiles and activity of glutathione S-transferase, glutathione peroxidase, and glutathione reductase in Euphorbia esula. Physiol. Plant 2004, 120, 421–433. [Google Scholar] [CrossRef]
- Rezaei, M.K.; Shobbar, Z.S.; Shahbazi, M.; Abedini, R.; Zare, S. Glutathione S-transferase (GST) family in barley: Identification of members, enzyme activity, and gene expression pattern. J. Plant Physiol. 2013, 170, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; Lapthorn, A.; Edwards, R. Plant glutathione transferases. Genome Biol. 2002, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, Y.; Duan, R.; Fan, J.; Jiao, P.; Sun, H.; Guan, S.; Liu, S. Overexpression of Maize Glutathione S-Transferase ZmGST26 Decreases Drought Resistance of Arabidopsis. Agronomy 2022, 12, 2948. [Google Scholar] [CrossRef]
- Chen, J.H.; Jiang, H.W.; Hsieh, E.J.; Chen, H. Y.; Chien, C.T.; Hsieh, H.L.; Lin, T.P. Drought and salt stress tolerance of an Arabidopsis glutathione S-transferase U17 knockout mutant are attributed to the combined effect of glutathione and abscisic acid. Plant Physiol. 2012, 158, 340–351. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, N.; Liu, Z.; Liu, S.; Liu, C.; Lin, J.; Yang, H.; Li, S.; Yukawa, Y. The AtGSTU7 gene influences glutathione-dependent seed germination under ABA and osmotic stress in Arabidopsis. Biochem. Biophys. Res. Commun. 2020, 528, 538–544. [Google Scholar] [CrossRef]
- Yang, Q.; Liu, Y.J.; Zeng, Q.Y. Overexpression of three orthologous glutathione S-transferases from Populus increased salt and drought resistance in Arabidopsis. Biochem. Syst. Ecol. 2019, 83, 57–61. [Google Scholar] [CrossRef]
- Bartels, D.; Sunkar, R. Drought and Salt Tolerance in Plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Chen, X.; Ding, Y.; Yang, Y.; Song, C.; Wang, B.; Yang, S.; Guo, Y.; Gong, Z. Protein kinases in plant responses to drought, salt, and cold stress. J. Integr. Plant Biol. 2021, 63, 53–78. [Google Scholar] [CrossRef]
- Yoon, H.W.; Kim, M.C.; Shin, P.G.; et al. Differential expression of two functional serine/threonine protein kinases from soybean that have an unusual acidic domain at the carboxy terminus. Mol. Gen. Genet. 1997, 255, 359–371. [Google Scholar] [CrossRef]
- Zinati, Z.; Sazegari, S. Identification of important genes involved in priming induced drought tolerance in barley through transcriptomic data mining. Crop Pasture Sci. 2022, 73, 1011–1025. [Google Scholar] [CrossRef]
- Cui, L.; Yang, G.; Yan, J.; Pan, Y.; Nie, X. Genome-wide identification, expression profiles and regulatory network of MAPK cascade gene family in barley. BMC Genom. 2019, 20, 750. [Google Scholar] [CrossRef]
- Dudziak, K.; Zapalska, M.; Börner, A.; Szczerba, H.; Kowalczyk, K.; Nowak, M. Analysis of wheat gene expression related to the oxidative stress response and signal transduction under short-term osmotic stress. Sci. Rep. 2019, 9, 1–14. [Google Scholar]
- Ying, S.; Zhang, D.F.; Fu, J.; Shi, Y.S.; Song, Y.C.; Wang, T.Y.; Li, Y. Cloning and characterization of a maize bZIP transcription factor, ZmbZIP72, confers drought and salt tolerance in transgenic Arabidopsis. Planta 2012, 235, 253–266. [Google Scholar] [CrossRef]
- Liao, Y.; Zou, H.F.; Wei, W.; Hao, Y.J.; Tian, A.G.; Huang, J.; Liu, Y.F.; Zhang, J.S.; Chen, S.Y. Soybean GmbZIP44, GmbZIP62 and GmbZIP78 genes function as negative regulator of ABA signaling and confer salt and freezing tolerance in transgenic Arabidopsis. Planta 2008, 228, 225–240. [Google Scholar] [CrossRef]
- Xu, Y.; Zhao, X.; Aiwaili, P.; Mu, X.; Zhao, M.; Zhao, J.; Cheng, L.; Ma, C.; Gao, J.; Hong, B. A zinc finger protein BBX19 interacts with ABF3 to affect drought tolerance negatively in chrysanthemum. Plant J. 2020, 103, 1783–1795. [Google Scholar] [CrossRef]
- Zhou, M.; Cheng, H.; Chiang, V.L.; Li, W.; Yang, C.; Wang, C. PtrbZIP3 transcription factor regulates drought tolerance of Populus trichocarpa. Environ. Exp. Bot. 2023, 208, 105231. [Google Scholar] [CrossRef]
- Oh, S.J.; Song, S.I.; Kim, Y.S.; et al. Arabidopsis CBF3/DREB1A and ABF3 in transgenic rice increased tolerance to abiotic stress without stunting growth. Plant Physiol. 2005, 138, 341–351. [Google Scholar] [CrossRef]
- Wu, J.; Jiang, Y.; Liang, Y.; Chen, L.; Chen, W.; Cheng, B. Expression of the maize MYB transcription factor ZmMYB3R enhances drought and salt stress tolerance in transgenic plants. Plant Physiol. Biochem. 2019, 137, 179–188. [Google Scholar] [CrossRef]
- Ma, Q.; Dai, X.; Xu, Y.; et al. Enhanced tolerance to chilling stress in OsMYB3R-2 transgenic rice is mediated by alteration in cell cycle and ectopic expression of stress genes. Plant Physiol. 2009, 150, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Tian, S.; Dong, H.; Guo, C. Pleiotropic effects of TaMYB3R1 on plant development and response to osmotic stress in transgenic Arabidopsis. Gene 2015, 558, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Shan, Z.; Zhou, S.; Yang, Q.; Gai, J.; Li, Y. GmDNAJC7 from Soybean Is Involved in Plant Tolerance to Alkaline-Salt, Salt, and Drought Stresses. Agronomy 2022, 12, 1419. [Google Scholar] [CrossRef]
- Zhichang, Z.; Wanrong, Z.; Jinping, Y.; Jianjun, Z.; Xufeng, L.Z.L.; Yang, Y. Over-expression of Arabidopsis DnaJ (Hsp40) contributes to NaCl-stress tolerance. Afr. J. Biotechnol. 2010, 9, 972–978. [Google Scholar]
- Sugino, M.; Hibino, T.; Tanaka, Y.; Nii, N.; Takabe, T. Overexpression of DnaK from a halotolerant cyanobacterium Aphanothece halophytica acquires resistance to salt stress in transgenic tobacco plants. Plant Sci. 1999, 146, 81–88. [Google Scholar] [CrossRef]
- Isayenkov, S.; Hilo, A.; Rizzo, P.; Tandron Moya, Y.A.; Rolletschek, H.; Borisjuk, L.; Radchuk, V. Adaptation strategies of halophytic barley Hordeum marinum ssp. marinum to high salinity and osmotic stress. Int. J. Mol. Sci. 2020, 21, 9019. [Google Scholar] [CrossRef]
- Huang, G.T.; Ma, S.L.; Bai, L.P.; Zhang, L.; Ma, H.; Jia, P.; Liu, J.; Zhong, M.; Guo, Z.F. Signal transduction during cold, salt, and drought stresses in plants. Mol. Biol. Rep. 2012, 39, 969–987. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, X.; Zhang, Z.; Wang, A.; Zhu, J. Cold-regulated gene LeCOR413PM2 confers cold stress tolerance in tomato plants. Gene 2021, 764, 145097. [Google Scholar] [CrossRef]
- Lee, K.W.; Choi, G.J.; Kim, K.Y.; Ji, H.C.; Zaman, R.; Lee, S. H. Identification of drought induced differentially expressed genes in barley leaves using the annealing control-primer-based Gene Fishing technique. Aust. J. Crop Sci. 2011, 5, 1364–1369. [Google Scholar]
- Das, R.R.; Pradhan, S.; Parida, A. De-novo transcriptome analysis unveils differentially expressed genes regulating drought and salt stress response in Panicum sumatrense. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
Figure 1.
Experimental setup of stress treatments in wild barley. Salinity and PEG stress were imposed on 15-day-old plants in a stepwise manner (marked with an asterisk). Black dots on the graph represent the time points of stress treatments. Time-points where tissue samples were harvested from control and stressed plants are indicated with asterisks. Initially, 25 mM salt was applied to the seedlings for a day and salt concentrations were gradually elevated daily first to 50 mM, then to 75 mM and finally to 100 mM during the next three days.
Figure 1.
Experimental setup of stress treatments in wild barley. Salinity and PEG stress were imposed on 15-day-old plants in a stepwise manner (marked with an asterisk). Black dots on the graph represent the time points of stress treatments. Time-points where tissue samples were harvested from control and stressed plants are indicated with asterisks. Initially, 25 mM salt was applied to the seedlings for a day and salt concentrations were gradually elevated daily first to 50 mM, then to 75 mM and finally to 100 mM during the next three days.
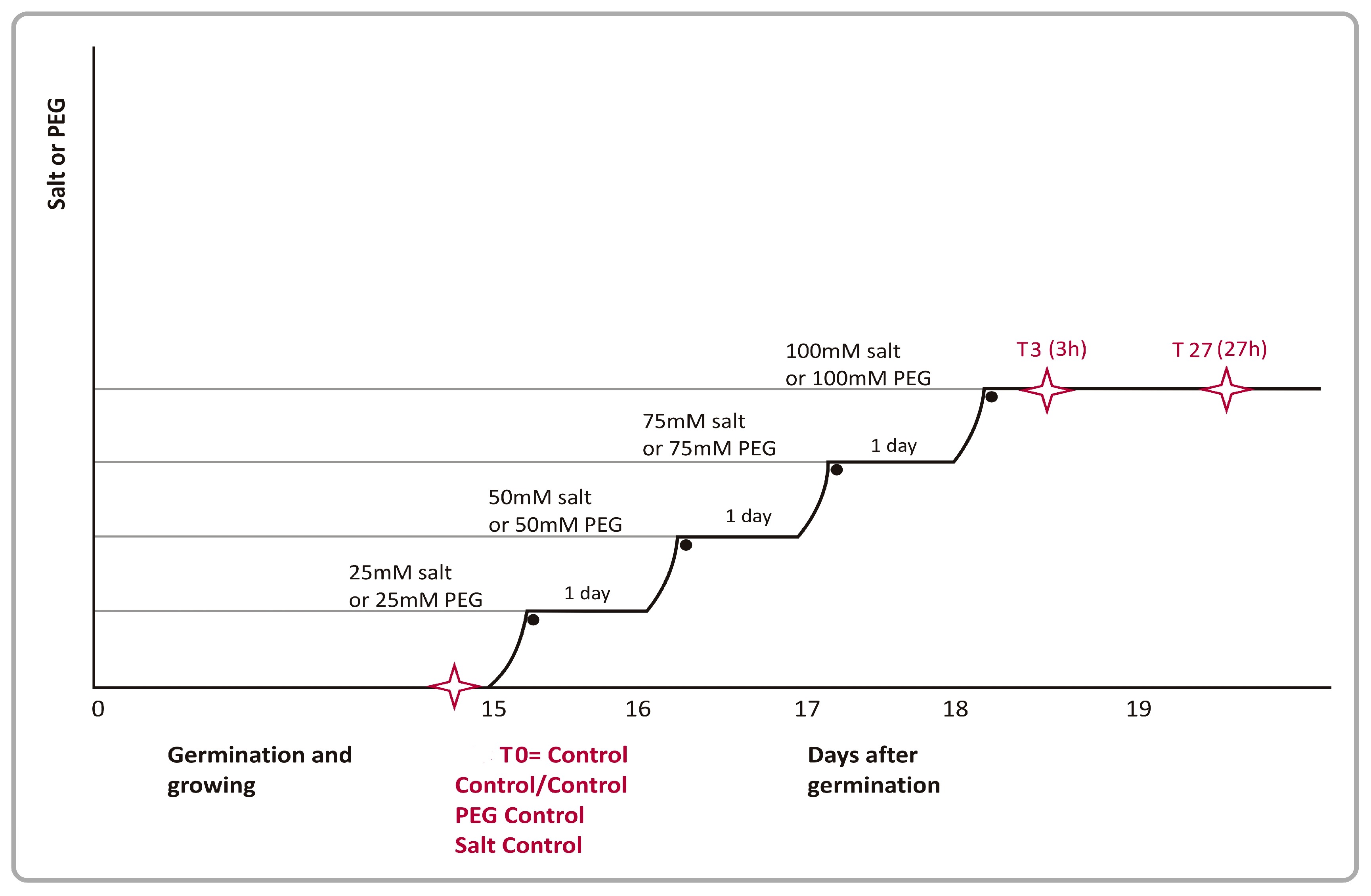
Figure 2.
Leaf water potentials of 20-40 and 23-70 wild barley genotypes under PEG (A) and salt (B) stress. Results are presented as mean +/-SD of five individual measurements.
Figure 2.
Leaf water potentials of 20-40 and 23-70 wild barley genotypes under PEG (A) and salt (B) stress. Results are presented as mean +/-SD of five individual measurements.
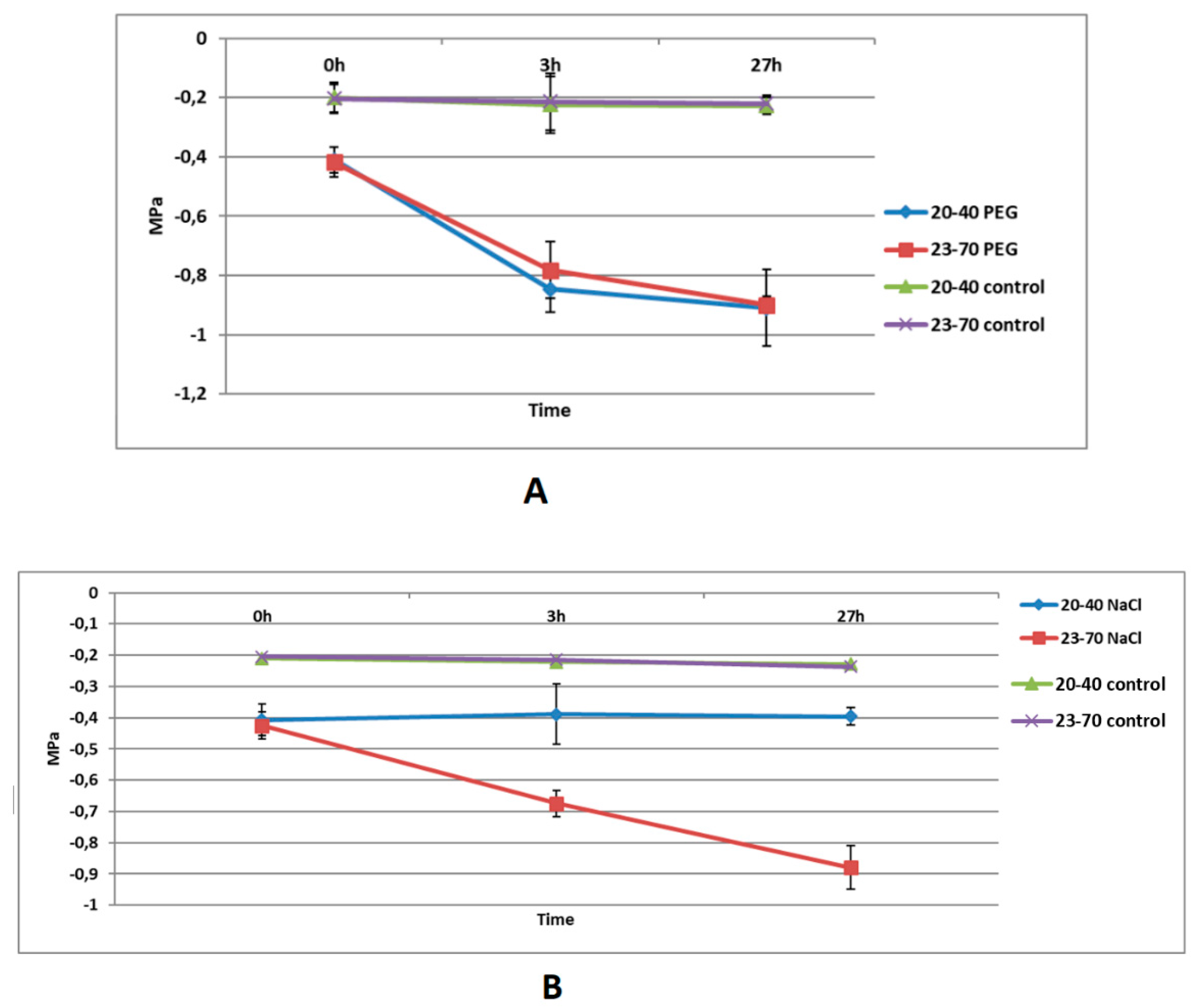
Figure 3.
Gene ontology (GO) annotations of Cluster 9 (Biological Process) of salt and drought responsive probe sets on the microarray that had identifiable homologies to Affy annotation file.
Figure 3.
Gene ontology (GO) annotations of Cluster 9 (Biological Process) of salt and drought responsive probe sets on the microarray that had identifiable homologies to Affy annotation file.
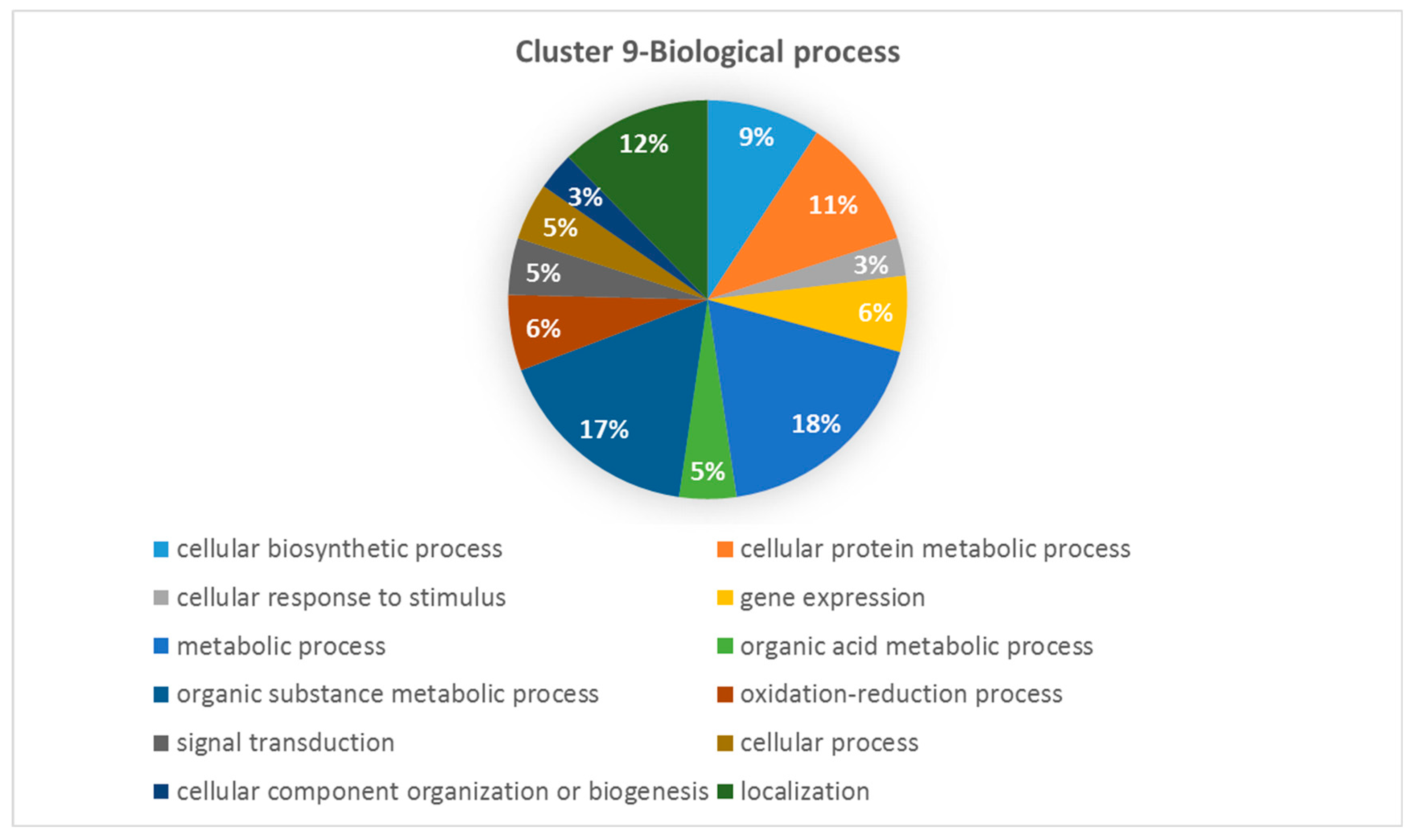
Figure 4.
The most significantly enriched KEGG pathways for PEG and salt stress genes.

Figure 5.
Venn diagrams (A, B, C, D) of differentially expressed genes across two different wild barley accessions (20-40 and 23-70), two tissues (leaf and root), two treatments (salt and PEG) and three time points (0, 3 and 27h).
Figure 5.
Venn diagrams (A, B, C, D) of differentially expressed genes across two different wild barley accessions (20-40 and 23-70), two tissues (leaf and root), two treatments (salt and PEG) and three time points (0, 3 and 27h).
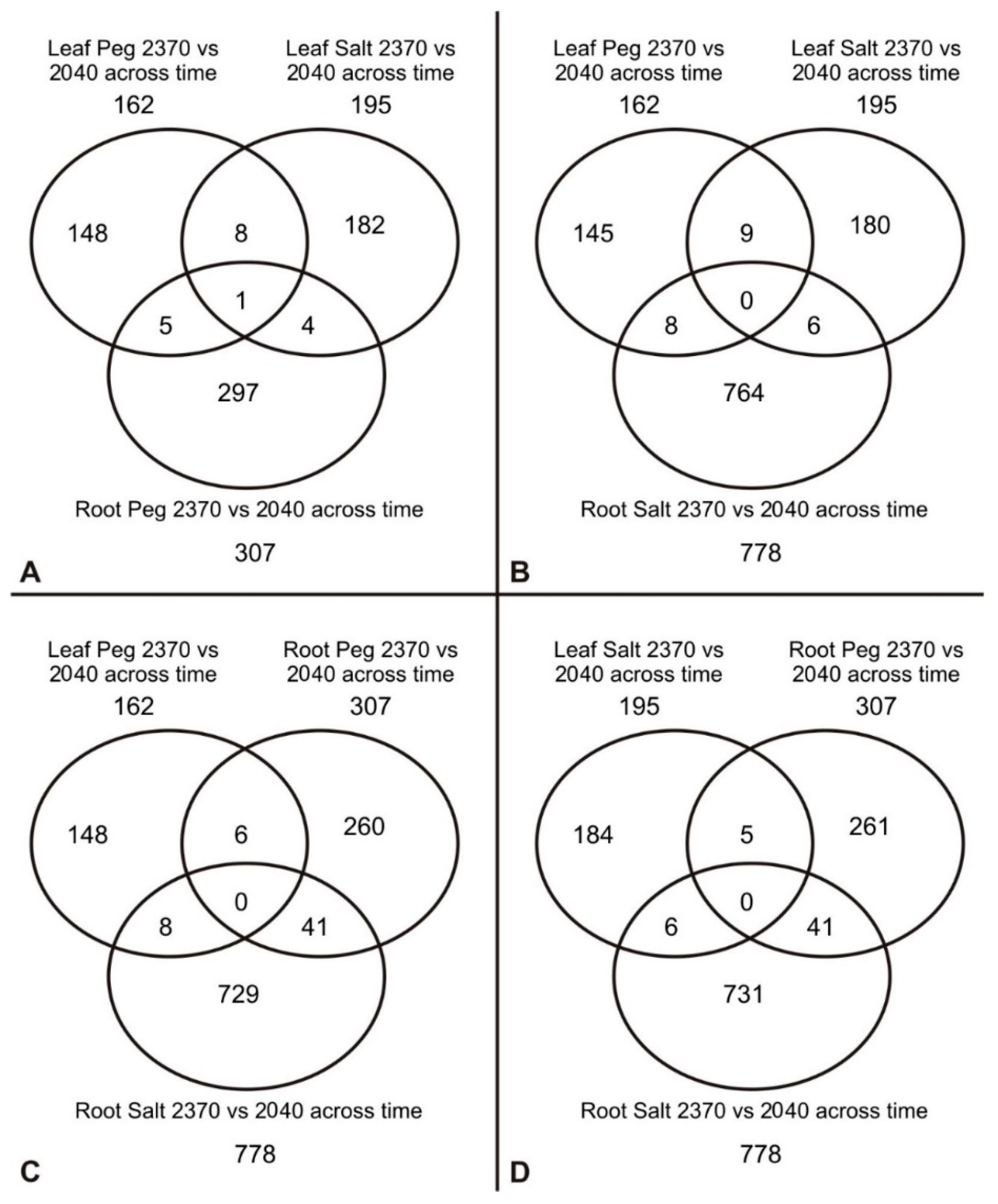

Table 1.
Examples of genes that were differentially expressed (up-regulated or down-regulated) in wild-barley in response to PEG treatment (*:co-expressed gene with salt stress).
Table 1.
Examples of genes that were differentially expressed (up-regulated or down-regulated) in wild-barley in response to PEG treatment (*:co-expressed gene with salt stress).
| Up-regulated genes | ||||||
|---|---|---|---|---|---|---|
| Probe Set ID | Accession No | E-valuec | Annotation | Genotype | Tissue | Time Point |
| Contig1568_x_at* | S22515 | 2,00E-65 | Thionin precursor | 23-70 | Leaf | 3h-27h |
| Contig3501_at* | Q00531 | e-108 | 60 kDa jasmonate-induced protein | 23-70 | Leaf | 3h |
| 20-40 | Leaf | 27h | ||||
| HV06O23u_at | NP_567835.1 | mRNA cleavage factor subunit - like protein | 23-70 | Leaf | 3h-27h | |
| 23-70 | Root | 3h-27h | ||||
| 20-40 | Leaf | 3h-27h | ||||
| 20-40 | Root | 3h-27h | ||||
| Contig13483_at | NP_188806.1 | 5,00E-70 | Integral membrane protein, putative | 23-70 | Leaf | 3h-27h |
| Contig21604_at* | CAC83006.1 | 1,00E-42 | Putative glutathione synthetase | 23-70 | Leaf | 3h-27h |
| Contig1865_at | AAM08517.1 | 3,00E-90 | Putative peroxidase | 23-70 | Leaf | 3h |
| Contig1718_s_at* | AAD02260.1 | 3,00E-30 | Dehydrin 9 | 23-70 | Root | 27h |
| 20-40 | Leaf | 3h-27h | ||||
| 20-40 | Root | 3h-27h | ||||
| Contig8961_at* | T02663 | 9,00E-17 | Abscisic acid- and stress-induced protein | 20-40 | Leaf | 3h-27h |
| Contig9774_s_at | Unknown | 20-40 | Leaf | 3h-27h | ||
| Contig4281_s_at* | CAC12881.1 | 2,00E-77 | Cold-regulated protein | 23-70 | Leaf | 3h |
| 23-70 | Root | 27h | ||||
| 20-40 | Leaf | 3h-27h | ||||
| Contig3112_at* | T06810 | 2,00E-87 | Cold acclimation protein WCOR413 | 20-40 | Leaf | 3h-27h |
| 20-40 | Root | 3h | ||||
| Contig15982_at | NP_567949.1 | 1,00E-31 | Abscisic acid responsive elements-binding factor(ABF3) | 20-40 | Leaf | 3h |
| HR01N22u_s_at | BAB62578.1 | 1,00E-15 | Putative protein kinase SPK-3 | 20-40 | Leaf | 3h-27h |
| Contig9239_s_at | Unknown | 23-70 | Leaf | 27h | ||
| 20-40 | Leaf | 27h | ||||
| Contig6546_at* | T06480 | e-101 | Safener-induced In2.1-like protein | 23-70 | Root | 3h |
| Contig3017_at* | CAA74595.1 | e-126 | Oxalate oxidase | 23-70 | Root | 3h-27 |
| 20-40 | Root | 27h | ||||
| Contig4213_at | CAB85629.1 | 1,00E-50 | Putative ripening-related protein | 23-70 | Root | 3h-27 |
| Contig5838_at | AAG40562.1 | 4,00E-88 | Glutathione-S-transferase 2 | 23-70 | Root | 3h |
| Contig17190_at* | AAL83988.1 | 2,00E-36 | Putative heat shock protein | 23-70 | Root | 27h |
| Contig3783_s_at* | AAC31615.1 | 2,00E-17 | Hypersensitive-induced reaction protein 3 | 20-40 | Root | 3h-27 |
| Contig15282_at | AAG34845.1 | 6,00E-41 | Glutathione S-transferase GST 37 | 23-70 | Root | 3h |
| 20-40 | Root | 3h | ||||
| Down-regulated genes | ||||||
| rbah13p07_s_at* | AAM76682.1 | 2,00E-24 | Peroxidase | 23-70 | Leaf | 3h-27h |
| 23-70 | Root | 3h | ||||
| 20-40 | Leaf | 3h-27h | ||||
| 20-40 | Root | 3h | ||||
| Contig25448_at | BAC20673.1 | 3,00E+38 | Serine/threonine kinase-like protein | 23-70 | Leaf | 3h-27h |
| Contig15729_at* | BAB63804.1 | 7,00E-30 | Putative I-box binding factor | 23-70 | Leaf | 3h |
| 23-70 | Root | 3h-27h | ||||
| 20-40 | Root | 3h | ||||
| Contig15489_at* | AAK73104.1 | 3,00E-33 | MAP kinase kinase | 23-70 | Leaf | 3h |
| 20-40 | Root | 3h | ||||
| Contig2112_at | S61406 | e-103 | Peroxidase 2 precursor | 23-70 | Leaf | 3h |
| 23-70 | Root | 3h | ||||
| 20-40 | Root | 3h | ||||
| Contig17190_at* | AAL83988.1 | 2,00E-36 | Putative heat shock protein | 23-70 | Leaf | 3h |
| Contig3783_at* | AAC31615.1 | 2,00E-17 | Physical impedance induced protein | 20-40 | Leaf | 3h-27h |
| Contig13932_at | AAL34131.1 | 6,00E-27 | Unknown | 23-70 | Leaf | 3h-27h |
| 20-40 | Leaf | 3h-27h | ||||
| Contig3636_at | CAC37639.1 | 8,00E-60 | SERK2 protein | 20-40 | Leaf | 27h |
| Contig6546_at* | T064480 | e-101 | Safener-induced In2.1-like protein | 20-40 | Leaf | 27h |
| Contig3875_s_at* | NP_566088.2 | 1,00E-23 | MYB-related transcription factor | 20-40 | Leaf | 3h |
| Contig21604_at* | CAC83006.1 | 1,00E-42 | Putative glutathione synthetase | 23-70 | Root | 3h-27h |
| 20-40 | Root | 3h-27h | ||||
| Contig3216_at | BAC10287.1 | 1,00E-20 | Defensin | 23-70 | Root | 3h-27h |
| Contig18873_at | P17801 | 4,00E-53 | Putative receptor protein kinase ZMPK1 precursor | 23-70 | Root | 3h-27h |
| 20-40 | Root | 3h | ||||
| EBro03_SQ007_C24_at | AAL73531.1 | 1,00E-18 | TNP2-like protein | 23-70 | Root | 3h |
| Contig8961_at* | T02663 | 9,00E-17 | Abscisic acid- and stress-induced protein | 23-70 | Root | 3h-27h |
| Contig10179_s_at | Unknown | 20-40 | Root | 3h | ||
| Contig25015_at* | NP_680449.1 | 2,00E-08 | Unknown | 23-70 | Root | 3h |
| 20-40 | Root | 3h | ||||
| HC109A09_T3_at | NP_079900.1 | 6,00E-12 | Unknown | 23-70 | Root | 3h-27 |
| 20-40 | Root | 3h-27 | ||||
| Contig3156_s_at | T05956 | 7,00E-36 | Germin-like protein | 23-70 | Leaf | 27h |
| 23-70 | Root | 3h | ||||
| 20-40 | Root | 27h |
Table 2.
Examples of genes that were differentially expressed (up-regulated or down-regulated) in wild-barley in response to salt. (*:co-expressed gene with PEG stress).
Table 2.
Examples of genes that were differentially expressed (up-regulated or down-regulated) in wild-barley in response to salt. (*:co-expressed gene with PEG stress).
| Up-regulated genes | ||||||||
|---|---|---|---|---|---|---|---|---|
| Probe Set ID | Accession No | E-valuec | Annotation | Genotype | Tissue | Time Point | ||
| Contig3501_at* | Q00531 | e-108 | 60 kDa jasmonate-induced protein | 23-70 | Leaf | 3h | ||
| 23-70 | Root | 3h | ||||||
| 20-40 | Leaf | 3h-27h | ||||||
| Contig6004_x_at | NP_564147.1 | 2,00E-10 | Unknown | 23-70 | Leaf | 3h-27h | ||
| Contig4281_s_at* | CAC12881.1 | 2,00E-77 | Cold-regulated protein | 23-70 | Leaf | 27h | ||
| Contig10808_at | BAB02515.1 | 8,00E-45 | Transporter-like protein | 23-70 | Leaf | 27h | ||
| 23-70 | Root | 3h-27h | ||||||
| 20-40 | Root | 3h-27h | ||||||
| Contig1568_x_at* | S22515 | 2,00E-65 | Thionin precursor | 23-70 | Leaf | 3h-27h | ||
| Contig3967_at | AAK62365.1 | 5,00E-23 | Unknown | 23-70 | Root | 27h | ||
| 20-40 | Leaf | 3h | ||||||
| Contig8961_at* | T02663 | 9,00E-17 | Abscisic acid- and stress-induced protein | 20-40 | Leaf | 3h | ||
| Contig1718_s_at* | AAD02260.1 | 3,00E-30 | Dehydrin 9 | 20-40 | Leaf | 3h | ||
| 20-40 | Root | 3h-27h | ||||||
| Contig24926_at | BAC10698.1 | 4,00E-13 | Putative leucine rich repeat containing protein kinase | 20-40 | Leaf | 27h | ||
| Contig6546_at* | T06480 | e-101 | Safener-induced In2.1-like protein | 23-70 | Root | 3h | ||
| 20-40 | Root | 3h | ||||||
| Contig21640_at | AAG40562.1 | 1,00E-61 | Glutathione-S-transferase 2 | 23-70 | Root | 3h | ||
| 20-40 | Root | 3h | ||||||
| Contig3017_at* | CAA74595.1 | e-126 | Oxalate oxidase | 23-70 | Root | 3h-27 | ||
| 20-40 | Root | 3h-27h | ||||||
| HW06F04u_s_at | Unknown | 23-70 | Root | 3h | ||||
| Contig3783_at* | AAC31615.1 | 2,00E-17 | Physical impedance induced protein | 20-40 | Root | 3h-27h | ||
| Contig3783_s_at* | AAN17464.1 | e-126 | Hypersensitive-induced reaction protein 3 | 23-70 | Root | 3h-27h | ||
| 20-40 | Root | 3h-27h | ||||||
| Contig3112_at* | T06810 | 2,00E-87 | Cold acclimation protein WCOR413 | 20-40 | Root | 3h | ||
| Down-regulated genes | ||||||||
| Contig ID | Accession No | E-valuec | Annotation | Genotype | Tissue | Time Point | ||
| rbah13p07_s_at* | AAM76682.1 | 2,00E-24 | Peroxidase | 23-70 | Leaf | 3h | ||
| 20-40 | Leaf | 3h-27h | ||||||
| Contig15729_at* | BAB63804.1 | 7,00E-30 | Putative I-box binding factor | 23-70 | Leaf | 3h | ||
| 23-70 | Root | 3h-27h | ||||||
| 20-40 | Root | 3h | ||||||
| Contig6953_at | Unknown | 23-70 | Leaf | 3h-27h | ||||
| 20-40 | Leaf | 27h | ||||||
| Contig15489_at* | AAK73104.1 | 3,00E-33 | MAP kinase kinase | 23-70 | Leaf | 3h | ||
| 20-40 | Root | 3h-27h | ||||||
| Contig11487_at | AAM63624.1 | 2,00E-33 | DnaJ protein, putative | 23-70 | Leaf | 3h | ||
| 23-70 | Root | 3h | ||||||
| Contig17190_at* | AAL83988.1 | 2,00E-36 | Putative heat shock protein | 23-70 | Leaf | 3h | ||
| 23-70 | Root | 3h | ||||||
| Contig3875_s_at* | NP_566088.2 | 1,00E-23 | MYB-related transcription factor | 20-40 | Leaf | 3h | ||
| Contig3783_at* | AAC31615.1 | 2,00E-17 | Physical impedance induced protein | 20-40 | Leaf | 3h-27h | ||
| Contig18873_at* | P17801 | 4,00E-53 | Putative receptor protein kinase ZMPK1 precursor | 23-70 | Root | 3h-27h | ||
| Contig9042_s_at | Unknown | 23-70 | Root | 3h | ||||
| 20-40 | Root | 3h | ||||||
| HV05A09u_s_at | P49970 | 9,00E-23 | Signal Recognition Particle 54 KD Protein3 (SRP54) | 23-70 | Root | 3h | ||
| Contig8961_at* | T02663 | 9,00E-17 | Abscisic acid- and stress-induced protein | 23-70 | Root | 3h | ||
| 20-40 | Root | 3h | ||||||
| EBro03_SQ007_C24_at | AAL73531.1 | 1,00E-18 | TNP2-like protein | 23-70 | Root | 3h | ||
| Contig3216_at | BAC10287.1 | 1,00E-20 | Defensin | 23-70 | Root | 3h-27h | ||
| Contig25015_at* | NP_680449.1 | 2,00E-08 | Unknown | 23-70 | Root | 3h-27h | ||
| 20-40 | Root | 3h-27h | ||||||
| Contig21604_at* | CAC83006.1 | 1,00E-42 | Putative glutathione synthetase | 23-70 | Root | 3h | ||
| 20-40 | Root | 3h-27h | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



