Submitted:
03 July 2023
Posted:
04 July 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Balanced fear supports human rational decision making and useful behavioral responses. In contrast, overwhelming, persistent, and unbalanced fear can paralyze the individual and result in heightened anxiety, lack of cognitive flexibility, fear-based public compliance and serious mental health issues. Psychobiotics research has established that a healthy microbiome is required for balanced fear and mental health protection via control of fear extinction. The recent Covid-19 pandemic featured daily, persistent, fear-of-a single contagion conditioning on a global scale paired with various behavioral mandates (e.g., lockdowns of the healthy, masks, isolation from environmental microbes and each other) that degraded the human microbiome and isolated us from each other and useful environmental microbes. It also ignored the historic role of secondary bacterial pathobionts, in pandemic deaths. This narrative review examines how institutional promotion of fear-of-a single contagion, lack of balanced risk communication, and appalling disregard of our fundamental nature (as majority-microbial human superorganisms) resulted in problems rather than solutions. This review concludes that government-public health-media promotion of pervasive fear and microbiome-degrading behaviors: 1) increased public compliance, 2) reduced cognitive flexibility, and 3) increased risk of mental health conditions. It further argues that microbiome-first public health should be embraced to ensure that human freedom, rather than paralyzing fear, dominates our future.
Keywords:
microbiome
; human superorganism
; fear conditioning
; fear-of-contagion
; fear extinction
; Covid-19
; pandemic
; risk communication
; mental health challenges
; self-empowerment
1. Introduction
To date, the twenty-first century could be labeled as the age of the microbes. This is not because of the recent SARS-CoV-2 (Covid-19) pandemic but rather because of recognition of the keystone role for microbiomes in human, animal, plant, complex ecosystems, and even planetary health. It is now clear that the fundamental biology of the human superorganism (holobiont) could be termed as ”mainly microbial” based on gene and cell numbers. Additionally, it is clear that looking across species and within the individual organism, microbiome-immunological compatibility is essential for both body integrity/health and even maintenance of the species [1].
During at least the last 60 years, pharma-driven medicine and much of public health have delivered to those they serve ever-increasing, co-morbid chronic diseases with polypharmacy as we age [2,3]. As a result, the lifespan has been increasingly filled with disease beginning earlier and earlier in life (e.g., childhood asthma and obesity). The healthspan (those decades of life spent largely free of disease) is all too brief if not rare among the population. Virtually all chronic diseases (previously termed non-communicable diseases) require: 1) misregulated, ongoing inflammation and 2) a dysfunctional microimmunosome [4]. At the heart of these diseases is a dysbiotic microbiome [5]. Disease treatments in the absence of rebalancing the microbiome are usually incomplete.
Beyond the role of the human microbiome in protecting against or promoting chronic diseases, it is the front-line defense against pathobionts. Given the human microbiome locations at portals of pathogen entry, the status of the microbiome can determine the effectiveness of colonization resistance and whether pathogens gain a foothold and an opportunity to produce disease [6]. Therefore, the microbiome should be our first priority in human healthcare rather than an afterthought, or worse excluded from public health initiatives altogether.
The recent Covid-19 pandemic demonstrated the massive gap between microbiome science and application to human public health. Rather than utilizing the human microbiome to aid in the pandemic (support for the immune system), the public was pushed into a deeper fear of microbes than already existed. This deep fear supported public compliance with a series of mandates [7]. In hindsight, many of the mandates were questionable at best and had as an outcome, degradation of the human microbiome [8,9,10,11].
This narrative review examines select gut microbiota-brain-neurological processes that are regulated by the microbiome and are important for human resiliency as well as neuropsychological and behavioral health. Dysbiosis of the human microbiome is known to increase the risk of myriad neurodevelopmental, neurodegenerative and psychiatric diseases, disorders, and conditions [12,13]. Examples of these include Parkinson’s Disease [14], Alzheimer’s Disease [15], autism [16,17], schizophrenia [18], major depressive disorder [19], and attention deficit/hyperactivity disorder [20]. Given the significance of gut microbiota-brain interconnections, this review also considers the role of flawed/incomplete public risk communication during the recent pandemic.
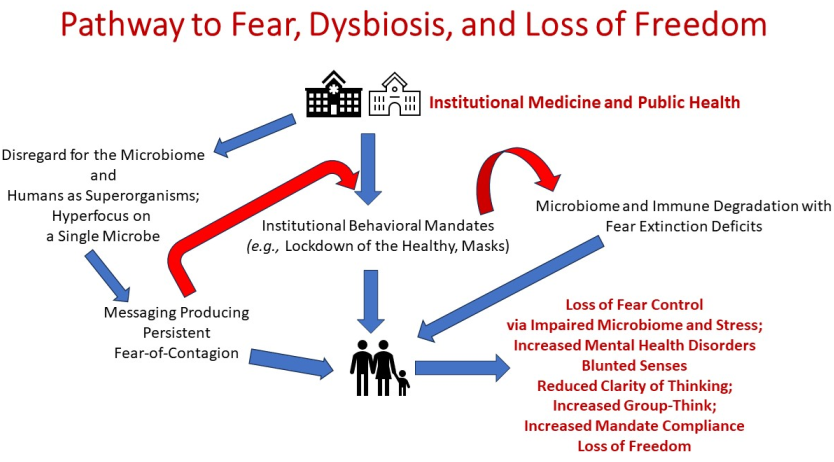
The premise is that daily, obsessive focus on a single viral pathobiont (the Covid-19 virus) to the exclusion of bacterial risk factors, the human microbiome, and whole human health led entire populations on a path to eroded human freedoms and a degraded human microbiome. There are six factors in the freedom vs. fear tug-of war that are discussed in sequence this paper. These are: 1) why misguided, misdirected, and/or inadequate public risk-benefit communication is central to fear-driven domination over human freedom, 2) the significance of a healthy gut microbiome for neurochemical/brain interconnectivity, balance, and mental health, 3) the Covid-19 pandemic as an example of fear-of-microbes conditioning, anxiety provoking and microbiome damaging mandates, and impaired mental health status, 4) the role of a healthy microbiome to extinguish fear, promote flexible-thinking, promote optimism via neurochemical balance, and facilitate brain and other physiological systems development, 5) the capacity of the microbiome to affect human senses, and 6) the importance of the microbiome to support societal and environmental interactions vs. isolation.
Finally, the paper describes why priority inclusion of the microbiome in the Covid-19 pandemic medical and published health initiatives could have resulted in a more free-thinking, cognitively flexible, and connected society overall in contrast with a fear conditioned, sensory isolated, and compliant populace. Healthcare for the public should never again exclude the microbiome.
2. Clear Risk-Benefit Communication with the Public for Self-Empowerment and Fear Reduction
One of the hallmarks of information-based decision-making (personal, family, community or even global) is use of precise risk-benefit analysis and communication. It is so important that most people do not make any major decisions without performing some level of risk-benefit comparisons even if they may not be aware they are doing that.
In recent publications the present authors and collaborators detailed why accurate, evidence-based, risk-benefit analysis and public communication involving our microbes is essential. It is critical for health-promoting actions, individual self-empowerment, and benefits for the whole human, the human superorganism. When risk communication is misdirected, inaccurate, or inadvertently or intentionally incomplete (e.g., missing the microbiome, additional pathobionts, risk of chronic diseases, the likely success of public health initiatives), damage rather than support of human health is likely. Defective risk communication (one that keeps critical details from the public) can even result in mass death rather than promotion of health as was seen with the example of Lysenkoism [21]. Such risk communication also encourages group-think rather than self-empowering, individual decision making.
In 2010 we published a book titled “Strategies for Protecting Your Child’s Immune System [22]. As the book was developing it became clear that we needed to educate and communicate fundamental information on safety/toxicity (Chapter 1) followed not only by an introduction to the scientific concept of risk (Chapter 2) but also, by an entire Chapter (3) titled Risk Exercises. This showed readers that the public actually operated based on benefit-risk decision making without even being conscious of it. But also, that risk communication information was vital in making key health decisions. In fact, Chapter 2 was titled “What’s the Risk,” because we encouraged parents and parents-to-be to pose the ‘What’s the Risk’ question to healthcare providers at every possible opportunity. If the risk of a prescription drug, medical procedure, or even a lifestyle change is completely unknown, then an informed decision cannot be made. The data are incomplete and the patient healthcare consumer lacks critical information on whether to accept a given medical treatment. Ironically, risk-based decision making is a fundamental consideration in financial planning. But this is not always given the same priority when it comes to health clients and decisions to be made.
The human microbiome is a major factor in risk of both infectious and chronic diseases [5,23,24,25]. Given the significance of the microbiome’s role in disease prevention and host resistance including against the Covid-19 virus [11,26,27], pandemic risk communication was a stunning failure in its myopic approach and exclusion of the microbiome. Broader risk communication concerning: 1) the use of beneficial microbes as a front-line microbial and immunological defense against life threatening infections including the Covid-19 virus, and 2) the use of microbiota to dampen excessive inflammation would have enabled the public not only to distinguish among good vs. bad microbes, but would also have facilitated pandemic health strategies beyond just experimental drugs. Instead, the hyperfocus on risk from a single viral pathogen left a locked down populace with an us vs. them (microbes) mentality and exacerbated a “fear of contagion” psychology [28,29,30].
2.1. Microbiota and Risk of Viral and/or Bacterial Pathogenesis
One of the seeming risk communication failures during the pandemic involved the importance of protecting against Covid-19 related complications and/or death. A significant beneficial factor is effective colonization resistance within the microimmunosome. This involves 1): viral loading issues as well as immune status in reducing the primary viral infection and carriage of potentially dangerous pathobionts in the nasal or even gut microbiome that can promote secondary bacterial infections or even specific chronic diseases. During the Covid-19 pandemic people were generally not made aware that co-secondary bacterial infections are a significant cause of complications and death during respiratory virus infection. This was despite the fact that researchers advocated for microbiota-based risk reduction to protect against severe Covid-19 disease [31].
Early during the Covid-19 pandemic, Di Stadio et al. [32] argued that the nasal microbiome was important for immunomodulatory protection against the Covid-19 infection. Smith et al. [33] and Jochems et al. [34] reported that carriage of specific nasal bacteria was important in the host response and resistance/vulnerability to Covid-19 infection. Specific nasal bacteria were directly connected with specific cytokine production and, thereby, affected whether airway damaging mucosal inflammation occurred during the response to Covid-19. Additionally, Jochems et al. [34] considered the role of specific bacteria in promoting or protecting against systemic inflammation. In fact, Coleman et al. [16] including a present author reviewed the importance of nasal carriage of Staphylococcus aureus in infants and the capacity to promote not only respiratory bacterial infection but also to directly instigate childhood asthma in this journal [16]. S. aureus is among the potentially life-threatening pathobionts involved with several body locations with pathogenesis potential easily affected by the body’s changes (e.g., viral infections, skin disruption, microbiome degradation) [35]. It needed to be included in risk-benefit considerations and communications.
The link between nasal microbiota and risk of respiratory virus infections is not limited to Covid-19. In the case of viral loading, Schippa and colleagues [36] found evidence that perturbed nasal microbiota profiles were linked with the course of Infant Respiratory Syncytial Virus (RSV) infection and bronchiolitis. The authors posited a colonization resistance relationship where a resilient basal microbiota may reduce viral load and severity of the respiratory disease.
2.2. History of Secondary Bacterial Pneumonia Deaths in Human Pandemics and Animal Coronavirus Infections
There is a second problem with the idea that Covid-19 virus was the sole contagion in the recent pandemic pathway to death. History tells us that many respiratory virus pandemics had secondary bacterial and/or fungal infections as a significant, if not leading cause of death. For example, in 2008 Morens, Taubenberger, and Fauci [37] published a modern analysis of a sample from the 1918-1919 influenza epidemic (also known as the “Spanish Flu”). Morris et al. [38] reviewed influenza pandemics through the ages and concluded that specific bacterial pathobionts such as Streptococcus pneumoniae, Haemophilus influenzae, and Staphylococcus aureus were responsible secondary bacterial infections resulting in death. The step-wise mechanisms through which primary viral infections result in bacterial pneumonia has been reviewed [39]. These researchers concluded that a majority of deaths were likely caused by secondary bacterial pneumonia. They further warned that preparedness for future viral pandemics must go beyond viral vaccine and anti-viral medications to focus on bacterial agents and by prevention of fatal bacterial pneumonia. Stunningly, the historic 2008 warning from researchers [37] about threat of death from secondary bacterial infections was largely forgotten during the Covid-19 pandemic where public health officials restricted focus to the single viral pathogen.
Would respiratory coronavirus infections be expected to result in significant bacterial pneumonia? Absolutely. Among animal respiratory coronaviruses the spike protein from bovine had among the highest percent identity comparisons with the SARS-CoV-2 spike protein exceeding that of even the bat coronavirus spike protein [40]. Bovine coronavirus has been shown to enhance bacterial adherence [41]. Calves are particularly susceptible to a respiratory disease complex where Bovine coronavirus is usually a mild symptom-producing primary infection that predisposes for more serious secondary bacterial infections from among several species (e.g., Pasteurella multocida, Mannheimia haemolytica, Histophilus somni, and Mycoplasma bovis). It is the respiratory bacterial infections that are more likely to be life threatening [42].
Among other animal respiratory coronaviruses, Infectious Bronchitis Virus (IBV) (a respiratory gamma coronavirus) in poultry has been suggested as a useful model for SARS-CoV-2 (Covid-19 virus) dissemination and infection [43]. An examination of decades of IBV research in poultry reveals that IBV-promoted secondary bacterial infection and resulting bacterial pneumonia is the leading cause of deaths rather than the primary respiratory coronavirus infection [44,45,46]. This example parallels what is seen with Influenza pandemics in human and bovine coronavirus infection in calves. These examples show us that tunnel-vision focus on a single respiratory viral pathogen is often not sufficient to encompass the risk of death from pathobiont-induced respiratory failure. With SARS-CoV-2, that is an equally important lesson.
2.3. The Covid-19 Example with Bacterial and/or Fungal Infections Causing Death
Considering that it is: 1) more than century since an influenza pandemic produced high levels of death by bacterial pneumonia, and 2) we know that prominent animal respiratory coronaviruses often produce death via bacterial pneumonia, it is not surprising that bacterial and/or fungal infections increased Covid-19 case severity and often resulted in death. [47]. Using the K18-hACE2 mouse model of COVID-19, Smith et al. [33] performed a study examining the time-sensitive immunological mechanisms through which the SARS-CoV-2 virus increases the susceptibility and pathogenicity of bacterial co-infection. Immunologically, Peng et al. 2023 [48] found that coronavirus impairs the host’s ability to clear bacterial pathogens by interfering with lysosomal function. Taken together these studies suggest that the public should have been educated and better protected against not just SARS-CoV-2 but also against the bacterial pathogen complex that results in a higher rate of death.
Cohen et al. [49] found that there was a higher rate of bacterial co-infection among moderate to severe Covid-19 hospital admitted patients than milder cases. The most common bacteria were Haemophilus influenzae (36%), Staphylococcus aureus (23%), Streptococcus pneumoniae (10%) and Enterobacter cloacae (10%) [49]. In a New York City hospital study, those hospitalized with Covid-19 had a 1.5 fold higher prevalence of positive bacterial culture tests vs. non-Covid-19 patients. Furthermore, certain hospital procedures for Covid -19 patients (e.g., intubation, ventilators) increased the frequency of bacterial pneumonia cases. Among the bacteria detected were methicillin-resistant Staphylococcus aureus, Klebsiella pneumoniae, and antibiotic-resistant organisms [50]. De Bruyn et al. [51] also reported that a majority of Covid-19 patients had secondary bacterial infections during their hospital ICU stay. In a systematic review of ventilator associated bacterial pneumonia among Covid-19 patients, Velásquez-Garcia et al. [52] found that gram-negative bacteria were the most frequent causes of pneumonia with Staphylococcus aureus being the most common organism isolated among those patients.
For a risk-benefit communication viewpoint, the general public was not made aware that a significant percentage of death associated with Covid-19 was not from the virus but rather from bacterial or fungal infections. Nor is it clear that the general public was educated about the potential role of ICU Covid-19 protocols in death by bacterial pneumonia. Given that secondary bacterial pneumonia was not a surprising development in this pandemic, the lack of risk communication beyond the single instigating virus (Covid-19) would have to be considered as a significant public health failure.
2.4. Adverse Risk Communication Regarding of Mask Mandates
Mask mandates were pervasive and compliance expected during the Covid-19 pandemic. But clear risk-benefit public communication that included likely adverse effects of prolonged wearing of facial masks was generally absent. Colonization resistance afforded by useful nose microbiota is important as a front-line defense against Covid-19 [32] and also potentially dangerous against bacterial infections [53]. Among the risks of loss of colonization resistance is the enhanced trafficking of the opportunistic pathogen Staphylococcus aureus [54]. One demonstrated adverse risk of prolonged face mask wearing was the concentration of bacteria on the face side of the face masks. This included potentially pathogenic bacteria including S. aureus that were found to be concentrated [55]. This is actually an expected outcome if face masks were filtering bacteria, which is one of their designed functions. In additional to concern over intra-individual load of potential bacterial pathogens, a recent study found that a significant risk exists for cross-contamination with S. aureus with mask disposal, particularly in the hospital setting [56].
Adverse risk from the mask mandates extended beyond loss of colonization resistance against bacterial infections, increased pathobiont loading and increased risks of pathobiont transmission (used masks as a biohazard). Pervasive wearing of face masks has been shown to impair both speech recognition and language development in young children [57].
2.5. Risk of Microbiome and Immune (Microimmunosome) Degradation
The microimmunosome (the systems biology combination of the microbiome and immune system) is a controlling factor in our health and well-being [4,58]. An obvious omission in risk communication during the Covid-19 pandemic was public assistance to maintain and optimize the microimmunsome to facilitate natural immunity, enhance colonization resistance and support natural health against Covid-19 and myriad other pathobionts. Finlay et al. [10] discuss the fact that implementing far more stringent hygiene practices come at a microbial cost to the human superorganism. In fact, an examination of pandemic mandates showed that many public health directives promoted microimmunsome degradation vs. optimized host defense function [10]. As a medical and public approach this was stunning considering two major realities: 1) Covid-19 was not the only significant human pathogen on the planet and 2) there would be human life after the pandemic. Effective public health strategies and risk communication should recognize the need for natural human health and resiliency beyond today’s apparent “emergency.”
The risk of microimmunsome degradation is one concern of many when the microbiome becomes compromised. An equally important concern is the function of the microbiome in fear extinction. If that role of the microbiome is compromised, then the population experiencing daily almost constant fear-of-contagion messaging may be less able to escape the fear mode. Finlay et al. [10] discuss the importance of a balanced approach to microbes where the public recognizes that we need beneficial microbes for our better health. The question is: Did the pandemic public health messaging fail to account for humans as holobionts and effectively throw the baby out with the bath water?
3. The Covid-19 Pandemic, Fear-of-Contagion and Compliance
Human fear conditioning and its relationship to human behavior and mental health has received extensive examination [59]. The recent Covid-19 pandemic provided a prime example of how human fear conditioning can be applied on a global scale. Mertens et al. [60] studied what they termed “Pandemic Panic” comparing populations in North America vs. Europe over the course of the Covid-19 pandemic. In this comparison the researchers found that predictive factors influenced the fear of Covid-19 and included region, anxious personality traits, gender, and media use. The prevalence of fear was higher in North America than Europe across the time frame examined.
Hauck et al. [61] discuss how the recent Covid-19 pandemic presented significant global populations with a persistent fear of illness. The authors discuss how control measures (e.g., mandates) presented other adverse health challenges that included both physical and mental health challenges. These authors list: being in quarantine, being overwhelmed or bored, feeling helpless, losing money, and the perception of inadequate information as stressors that added to the fear of illness (e.g., contagion). These researchers examined a healthy university population and found that those students with high Covid-pandemic anxiety had problems in distinguishing between fear and safety cues. This is consistent with the findings of Cannito et al. [62] who found that people who believed in severe consequences of the Covid-19 contagion were more likely to have hyper health anxiety toward the virus along with obsessive attention devoted to the contagion.
Among the factors predicting persistent anxiety and depression during the pandemic was insomnia [63]. The effect of a sleep disorder on pandemic anxiety and depression was significantly greater than that of other known risk factors (age, sex, and previous psychiatric diagnoses). This is important when the role of a healthy microbiome is considered. In an early study of chronic disease comorbidities, Dietert et al. [64] showed that pro-inflammatory driven sleep disorders and depression were at the epicenter of the most common comorbidities of otherwise diverse chronic diseases. Importantly, multi-systems biology dysfunction such as sleep disorders and disrupted circadian clocks have been shown to be affected by the status of the microbiome and are difficult to correct in the absence of microbiome rebalancing [reviewed in 58]. Sleep and circadian cycle regulation is another pathway through which human microbiome status affected the potential greater mental health burden arising from the “Pandemic Panic.”
Table 1 provides research examples of government, public health and media promotion of fear-of-contagion during the Covid-19 pandemic. It also illustrates the most vulnerable populations and the range of adverse mental health outcomes linked with the fear promotion [60,61,62,63,64,65,66,67,68,69,70,71,72].
With fear conditioning and institutional messaging to enhance fear of contagion being critical factors in ensuring global population compliance, it has become clear why maintaining a healthy human microbiome was absent from major public health recommendations during the Covid-19 pandemic. As will be discussed in the next section, a focus on the microbiome and particularly the gut microbiome would have been likely to dampen Covid-19-related fear and anxiety and would have potentially eroded mandate population compliance.
4. Regulation of Fear Extinction by the Microbiome and Public Health Implications
Both fear acquisition or conditioning and fear extinction are learned behaviors. In the lab standard Pavlovian conditioning is used where the subject animal is presented with a trigger (i.e., odor, tone) called the conditioned stimulus (CS) and then subjected to an adverse stimulus (i.e., foot shock) called the unconditioned stimulus (US). Once the animal has learned the fear response (i.e., freezing upon CS), the fear memory and reaction have been consolidated. The animal will now produce that behavior when the CS is presented [73,74]. The entire response has been acquired by associating the CS with a US.
The fear extinction process is something of the reverse of this in which the CS is repeatedly presented to the animal, but the US is absent. Over time the animal learns that it is safe and no longer produces the fear response [73,74]. The animal models have proven to be generalizable to human fear conditioning as well.
While a plethora of brain and neurological sites appear to be involved with these two processes, the core hub appears to be made up of the hippocampus, basolateral amygdala (BLA) and ventromedial prefrontal cortex (vmPFC or vPFC) [75]. The hippocampus plays a vital role in memory, learning, behavior, and spatial navigation [76,77,78]. It is central to the system of consolidating and encoding memories [79] and also converts short-term memories into long-term memories [80]. The BLA is part of the amygdala involved with input and the processing of emotion [81]. On the other hand, the vmPFC seems to act largely as a relay station. Gage and Baars [82] termed it the “integrative hub for emotional, sensory, social memory and self-related information processing” [82]. Because the vmPFC is highly interconnected, it binds large-scale networks together that promote emotional processing and social cognition [82]. These three brain areas have been identified by a number of researchers as being core to fear conditioning and extinction. Evidence suggests that gut microbiota are important in regulating these brain regions [83,84].
Linkages have been made between the neurological systems involved, dopamine and the gut microbiome, which all play into these learned behaviors. Salinas-Hernández and Duvarci [74] stress that fear extinction appears to represent new learning rather than forgetting or erasure of the original fear memory, and that it may be promoted by the reward learning system []. In their model, midbrain dopamine neurons encode reward prediction error (RPE) signals to drive reward learning [74]. Not getting a foot shock is the RPE and that triggers the reward center in the brain of which dopamine is a crucial component. In fact, dopamine increases in the mPFC, which is crucial for consolidating fear extinction memories. The dopamine levels remain high during and after extinction learning [74]. Together, these brain regions and their complex interactions both enable conditioned fear and then help create fear extinction memories to bring the individual back to homeostasis.
Even more central to the entire fear cascade process and most likely preceding fear acquisition is the status of the gut microbiome. Preclinical research showed that changes in gut microbiota composition are able to affect fear recall/fear extinction and risk of PTSD–relevant phenotypes [85]. Gut microbiota alterations/degradation can induce dopaminergic dysfunctions that help to determine not only the risk of developing PTSD but also its severity [85]. One of the microbial metabolites affecting the dopaminergic system is the neurotoxic metabolite p-cresol produced by the Ruminococcaceae and Lachnospiraceae families of bacteria [85,86,87]. Hamamah et al. [88] provided a comprehensive review of the distinct roles of numerous gut microbiome genera (e.g., Prevotella, Bacteroides, Lactobacillus, Bifidobacterium, Clostridium, Enterococcus, and Ruminococcus) in regulating dopaminergic signaling in the brain and beyond.
A hallmark of the human microbiome is its capacity to regulate not only most physiological systems [89,90,91,92] but also complex inter-system global functions [93]. Among the systems under microbiome regulation is the brain/neurological system [94]. In humans and many other mammals, the gut microbiome is a powerful regulator of neurodevelopment, brain chemistry, and responses to stimuli [95]. This can affect not only brain function but also human behavior [96]. While microbiome status can drive fear, anxiety, stress, depression, and mental health status at virtually any time of life, early life represents a critical window for programming of later life responses [97]. The importance of the gut-brain axis generally and between the gut microbiome and brain status specifically is so important that an entire field of study (known as psychobiotics) has emerged to guide improved human mental health via the gut microbiome [98,99,100]. In fact, Zielińska et al. [101] argued that psychobiotics and the manipulation of microbiota should have been used during the Covid-19 pandemic to protect our mental health.
Among the pivotal roles of the gut microbiome is the regulation of fear via control of fear extinction. While microbiome dysbiosis can result in many different comorbid chronic diseases [91], among the most life changing are those conditions driven by unrelenting fear and anxiety [102]. This is one of the reasons why gut microbiome dysbiosis is such a significant health threat.
Fear extinction is a learned experience that results in the production of new memories and the effective suppression of adverse memories. But it appears to be far from a simple inhibition process. As previously mentioned, dopamine appears to be involved in major aspects of fear extinction [74]. Fear extinction is critically important to good mental health. As discussed by Beckers et al. [59] and Sangha et al. [103], safety can be learned through fear extinction. But Beckers et al. [59] describe the reality that a characteristic of anxiety disorders is that unrealistic fear persists even in the face of obvious safety. Deficits in fear extinction are a risk factor for anxiety disorders since safety is not learned.
In a recent review study of 137 volunteers, Wen et al. [104] performed mapping of fear extinction among widely diverse brain regions using regional connectivity measures. They found increased connectivity among 133 out of 432 regions that were associated with fear extinction learning. These reported findings are in contrast with earlier studies focusing more on the brain areas of the BLA, hippocampus and vPFC that comprise the central hub of fear conditioning and extinction. Wen et al. 2021 [104] stressed that fear extinction is an inherently dynamic and complex process that begins at the core and reaches out across a vast area of the brain.
Table 2 provides examples of human and animal studies establishing the effects of microbiota on the regulation of fear extinction [84,105,106,107,108,109,110,111,112,113,114,115].
Maintenance of a healthy microbiome throughout the pandemic would have benefited colonization resistance against pathobionts. Table 2 illustrates that pandemic focus on the microbiome is likely to have optimized fear extinction and/or reduced challenges with anxiety or depression. This would have been a highly useful outcome both for the period of the Covid-19 pandemic and also the aftermath. However, a lingering question is whether optimized fear extinction across large populations would have reduced: 1) the impact of fear-of contagion messaging and/or 2) the levels of compliance with pandemic mandates? What is clear is that erosion of the microbiome before or during the pandemic would increase the risk of fear-related mental health challenges. Examples of fear-extinction deficit-connected mental health challenges are discussed in the following section.
5. Defective Fear Extinction and Mental Health Implications
Deficits in fear extinction and imbalanced fear have been identified as the cornerstone for a variety of psychiatric conditions. Importantly, Lebois et al. [116] reviewed the linkage of impaired fear extinction to psychiatric conditions, neurochemical and anatomical mediators involved in fear extinction as well as some augmentation strategies. Cryan and colleagues reviewed: 1) the connections between the microbiome, anxiety and depression, and microbiota-based therapeutic opportunities [117] as well as 2) how our microbiota help to mold our social brain [118].
Several factors can be involved in fear extinction deficits and elevated risk of mental health conditions. But most of these are also affected by microbiome status. Stress can be an important factor in fear extinction deficits contributing to elevated risk of mental health problems [119]. Sleep can support fear extinction while sleep disorders promote deficits [120]. As previously discussed, problems with dopamine regulation is also a factor in effective fear extinction learning [74]. Levels of sex hormone such as estrogen are important in fear extinction and gut microbiome status and are thought to affect both hormone and neurochemical (e.g., dopamine) regulation, in turn, affecting fear extinction [111,121].
6. Lockdown of the Healthy: Sensory Dulling, Microbiota, and Mental Health
During the Covid-19 pandemic a “lockdown of the healthy” was mandated in many counties that included school closures and limited social interactions. In particular, children and adolescents were adversely affected in terms of both mental and physical health [128,129,130]. Lockdown stress appeared to have the greatest adverse effects in single parent and/or single child households [131]. However, the elderly were also adversely affected by the lockdowns. A study in Italy found that the elderly experienced adverse changes in sleep quality and patterns [132].
In a recent article [133], we considered the internet of microbes and how mandated lockdown/isolation of the healthy during the Covid-19 pandemic and separation from the internet of microbes (e.g., each other, microbe-rich soil, and farm animal environments) could contribute to microbiome degradation. Such negative effects on the microbiome have been suggested [10] and researchers have even suggested that anti-anxiety, depression therapy should have been initiated early in the pandemic to minimize fear and stress based adverse outcomes [134].
Here we consider the potential effect of a degraded microbiome on the human senses. Several studies indicate that impaired senses are linked with anxiety and/or depression [135,136,137]. For this reason, sensory decline is an additional health risk for fear/stress-related anxiety disorders. This section illustrates that a “lockdown of the healthy” carries its own adverse health risk, which is likely overlooked when/if the microbiome is ignored.
The five basic human senses (taste, smell, touch, hearing, and sight) are vital components of our perception of the world around us. They provide us with information that is the cornerstone of human consciousness [138]. Among the regulatory capabilities of the human microbiome is the capacity to influence the senses. This is likely to be an interconnection that is less well known among the general public. One of the authors discusses microbiome control of taste and smell perception within a prior article [139]. Here, we update the literature on taste and smell and extend the discussion to include vision, hearing and skin sensitivity (touch). It is useful to note that microbiome regulation of senses seems to go beyond the basic five. Proprioception, or the knowledge of the body in space/location, is an awareness that can be affected by microbiota. For example, magnetogenic bacteria contain a specialized organelle that can detect and orient to magnetic fields. This helps the bacteria navigate magnetic fields and may play a role in magnetic field migration in higher vertebrates. For example, when migrating Eurasian reed warblers are treated with antibiotics, they lose their directional awareness [140]. This supports the hypothesis that a symbiotic relationship for magnetic field awareness exists. Interestingly, the human gut microbiome contains magnetotactic bacteria as well [141]. Hence, we may need to look beyond the five basic senses to know the full impact of the human microbiome on our human conscious and awareness.
7. Conclusions
As superorganisms (holobionts), humans have an overwhelming number of microbial genes that dwarf, in comparison, the number of chromosomal genes. These microbial genes are no less significant in impact than chromosomal genes. This biological reality alone is sufficient to expect that any major medical or public health initiative would be directed at the whole human and not at some misrepresentation of humans. But in the case of the most recent pandemic (Covid-19), government-supported medical and public health initiatives went a step further than to simply ignore human reality. As is evident in this narrative review, the protocols and daily public health messages flooding media in the name of the pandemic reflected two assumptions: 1) we exist as a microbe-free version of humans, 2) we needed to fight the one and only pathobiont that exists, a respiratory coronavirus, SARS-CoV-2, Covid-19. Risk communication for the public was virtually devoid of information on beneficial microbes and the human microbiome. Instead, the pandemic protocols and messaging were aimed at near total public compliance, and this was engendered through persistent fear-of contagion conditioning in what has been termed the “Pandemic Panic.” Such persistent fear conditioning if not controlled can result in metal health disorders.
Fortunately, we possess an inherent microbiome-driven brake on the fear-conditioning-mental health disorder life course: microbiota-regulated fear extinction learning. As discussed in this paper, fear extinction is thought to involve massive brain-neural connections that, through a largely parallel learning process requiring our microbiome, protects us against conditions such as anxiety disorders, depression, and PTSD. A significant problem is that many of the public health mandates directly degraded the human microbiome. We are not the first microbiome researchers to point this out [10]. As this review illustrates, the mandates degraded not only the human microbiome, but also our personal freedoms. Normal human clarity of thinking was replaced by a group-think pandemic fog. In particular, the “lockdown of the healthy” blocked us from microbiome-replenishing reservoirs of microbiota and negatively impacted host resistance via the microimmunosome. And it is now clear that the mandates also dulled human senses. The microbiome plays a significant role in the regulation of these senses. Degraded senses are also co-morbid for chronic diseases and can promote mental health conditions. Hence, microbiome degradation through public health mandates facilitated what is now an epidemic of post-pandemic mental health challenges with children among the most vulnerable populations.
Through the application of microbiology, we must insist that beneficial microbes are at the forefront of our healthful solutions.
Author Contributions
For this narrative review, R.R.D. led the drafting of each section, content pertaining to risk-benefit analysis and communication, the microbiome, and various pandemics. R.R.D. also drafted all tables and figures. J.M.D. oversaw content involving human behavior, psychiatric, and mental health conditions. J.M.D. also led editing and organization of the paper.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare they have no conflict of interest.
References
- Bell, K.; Bordenstein, S.R. , 2022. A margulian view of symbiosis and speciation: the Nasonia wasp system. Symbiosis, 2022, 87, 3–10. [Google Scholar] [CrossRef]
- Menditto, E.; Gimeno Miguel, A.; Moreno Juste, A.; Poblador Plou, B.; Aza Pascual-Salcedo, M.; Orlando, V.; González Rubio, F.; Prados Torres, A. Patterns of multimorbidity and polypharmacy in young and adult population: Systematic associations among chronic diseases and drugs using factor analysis. PLoS One 2019, 14, e0210701. [Google Scholar] [CrossRef] [PubMed]
- Almodóvar, A.S.; Nahata, M.C. Associations between chronic disease, polypharmacy, and medication-related problems among medicare beneficiaries. J. Manag. Care Special. Phar. 2019 25, 573–577. [CrossRef]
- Dietert, R.R. The microbiome-immune-host defense barrier complex (microimmunosome) and developmental programming of noncommunicable diseases. Reprod. Toxicol. 2017, 68, 49–58. [Google Scholar] [CrossRef]
- Vijay, A.; Valdes, A.M. Role of the gut microbiome in chronic diseases: a narrative review. Eur. J. Clin. Nutr. 2022, 76, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Flores, G.; Pickard, J.M.; Núñez, G. Microbiota-mediated colonization resistance: mechanisms and regulation. Nat. Rev. Microbiol. 2023, 21, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Harper, C.A. , Satchell, L.P., Fido, D. et al. Functional Fear Predicts Public Health Compliance in the COVID-19 Pandemic. Int. J. Ment. Health Addiction 2021, 19, 1875–1888. [Google Scholar] [CrossRef] [PubMed]
- Domingues, C.P.; Rebelo, J.S.; Dionisio, F.; Botelho, A.; Nogueira, T. 2020. The social distancing imposed to contain COVID-19 can affect our microbiome: a double-edged sword in human health. Msphere 2020, 5, e00716–20. [Google Scholar] [CrossRef] [PubMed]
- Ejtahed, HS.; Hasani-Ranjbar, S.; Siadat, S.D.; Larijani, B. The most important challenges ahead of microbiome pattern in the post era of the COVID-19 pandemic. J. Diabetes Metab. Disord. 2020, 19, 2031–2033. [Google Scholar] [CrossRef]
- Finlay, B.B.; Amato, K.R.; Azad, M.; Blaser, M.J.; Bosch, T.C.; Chu, H.; Dominguez-Bello, M.G.; Ehrlich, S.D.; Elinav, E.; Geva-Zatorsky, N.; Gros, P. The hygiene hypothesis, the COVID pandemic, and consequences for the human microbiome. Proc. Natl. Acad. Sci. U S A. 2021, 118, e2010217118. [Google Scholar] [CrossRef]
- Bacorn, M.; Romero-Soto, H.N.; Levy, S.; Chen, Q.; Hourigan, S.K. The Gut Microbiome of Children during the COVID-19 Pandemic. Microorganisms 2022, 10, 2460. [Google Scholar] [CrossRef]
- Dash, S.; Syed, Y.A.; Khan, M.R. Understanding the Role of the Gut Microbiome in Brain Development and Its Association With Neurodevelopmental Psychiatric Disorders. Front. Cell. Dev. Biol. 2022, 10, 880544. [Google Scholar] [CrossRef] [PubMed]
- k Ghezzi, L.; Cantoni., C.; Rotondo, E.; Galimberti, D. The Gut Microbiome-Brain Crosstalk in Neurodegenerative Diseases. Biomedicines 2022, 10, 1486. [Google Scholar] [CrossRef] [PubMed]
- Hashish, S.; Salama, M. The Role of an Altered Gut Microbiome in Parkinson’s Disease: A Narrative Review. Appl. Microbiol. 2023, 3, 429–447. [Google Scholar] [CrossRef]
- Chen, C.; Liao, J.; Xia, Y.; Liu, X.; Jones, R.; Haran, J.; McCormick, B.; Sampson, T.R.; Alam, A.; Ye, K. Gut microbiota regulate Alzheimer’s disease pathologies and cognitive disorders via PUFA-associated neuroinflammation. Gut 2022, 71, 2233–2252. [Google Scholar] [CrossRef]
- Coleman, M.E.; Dietert, R.R.; North, D.W.; Stephenson, M.M. Enhancing Human Superorganism Ecosystem Resilience by Holistically ‘Managing Our Microbes’. Appl. Microbiol. 2021, 1, 471–497. [Google Scholar] [CrossRef]
- Nirmalkar, K.; Qureshi, F.; Kang, D.W.; Hahn, J.; Adams, J.B.; Krajmalnik-Brown, R. Shotgun Metagenomics Study Suggests Alteration in Sulfur Metabolism and Oxidative Stress in Children with Autism and Improvement after Microbiota Transfer Therapy. Int. J. Mol. Sci. 2022, 23, 13481. [Google Scholar] [CrossRef]
- Thirion, F.; Speyer, H.; Hansen, T.H.; Nielsen, T.; Fan, Y.; Le Chatelier, E.; Fromentin, S.; Berland, M.; Plaza Oñate, F.; Pons, N.; et al. Alteration of Gut Microbiome in Patients With Schizophrenia Indicates Links Between Bacterial Tyrosine Biosynthesis and Cognitive Dysfunction. Biol. Psychiatry Glob. Open Sci. 2022, 3, 283–291. [Google Scholar] [CrossRef]
- Ortega, M.A.; Alvarez-Mon, M.A.; García-Montero, C.; Fraile-Martinez, O.; Guijarro, L.G.; Lahera, G.; Monserrat, J.; Valls, P.; Mora, F.; Rodríguez-Jiménez, R.; et al. Gut Microbiota Metabolites in Major Depressive Disorder-Deep Insights into Their Pathophysiological Role and Potential Translational Applications. Metabolites 2022, 12, 50. [Google Scholar] [CrossRef]
- Gkougka, D.; Mitropoulos, K.; Tzanakaki, G.; Panagouli, E.; Psaltopoulou, T.; Thomaidis, L.; Tsolia, M.; Sergentanis, T.N.; Tsitsika, A. Gut microbiome and attention deficit/hyperactivity disorder: a systematic review. Pediatr. Res. 2022, 92, 1507–1519. [Google Scholar] [CrossRef]
- Dietert, R.R. 2021. The microbiological basis of human superorganism freedom. Am. J. Biomed. Sci. Res. 2021 13, 653-662. Available online: https://biomedgrid.com/fulltext/volume13/the-microbiological-basis-of-human-superorganism-freedom.001933.php.
- Dietert, R.R.; Dietert, J.M. Strategies For Protecting Your Child's Immune System: Tools for Parents and Parents-To-Be. 1st ed.; World Scientific Publishing. Singapore, Singapore, 2010; pp. 1–26. Available online: https://www.worldscientific.com/worldscibooks/10.1142/7444#t=aboutBook.
- Dietert, R.R.; Dietert, J.M. The Completed Self: An Immunological View of the Human-Microbiome Superorganism and Risk of Chronic Diseases. Entropy 2012, 14, 2036–2065. [Google Scholar] [CrossRef]
- Dietert, R.R. A Focus on Microbiome Completeness and Optimized Colonization Resistance in Neonatology. NeoReviews 2018, 19, e78–e88. [Google Scholar] [CrossRef]
- Dietert, R.R. Microbiome First Medicine in Health and Safety. Biomedicines 2021, 9, 1099. [Google Scholar] [CrossRef] [PubMed]
- Rossini, V.; Tolosa-Enguis, V.; Frances-Cuesta, C. Sanz, Y. Gut microbiome and anti-viral immunity in COVID-19. Crit. Rev. Food Sci. Nutr. 2022, 16, 1–16. [Google Scholar] [CrossRef]
- Merra, G.; Capacci, A.; Cenname, G.; Esposito, E.; Dri, M.; Di Renzo, L.; Marchetti. M. The "Microbiome": A Protagonist in COVID-19 Era. Microorganisms 2022, 10, 296. [Google Scholar] [CrossRef] [PubMed]
- Mercader Rubio, I.; Sánchez-López, P.; Ángel, N.G.; Ruiz, N.F.O. Psychological Consequences of Fear of COVID-19: Symptom Analysis of Triggered Anxiety and Depression Disorders in Adolescents and Young Adults. Int. J. Environ. Res. Public Health 2022, 19, 14171. [Google Scholar] [CrossRef] [PubMed]
- Rania, N.; Coppola, I. The fear of contagion and the attitude toward the restrictive measures imposed to face COVID-19 in Italy: The psychological consequences caused by the pandemic one year after it began. Front. Psychol. 2022, 13, 118. [Google Scholar] [CrossRef] [PubMed]
- Baldi, E.; Savastano, S. 2021. Fear of contagion: one of the most devious enemies to fight during the COVID-19 pandemic. Disaster Med. Public Health Prep. 2021, 15, e8–e9. [Google Scholar] [CrossRef] [PubMed]
- Ngo, V.L.; Gewirtz, A.T. Microbiota as a potentially-modifiable factor influencing COVID-19. Curr. Opin. Virol. 2021, 49, 21–26. [Google Scholar] [CrossRef]
- Di Stadio, A.; Costantini, C.; Renga, G.; Pariano, M.; Ricci, G.; Romani, L. The microbiota/host immune system interaction in the nose to protect from COVID-19. Life 2020, 10, 345. [Google Scholar] [CrossRef]
- Smith, A.P.; Williams, E.P.; Plunkett, T.R.; Selvaraj, M.; Lane, L.C.; Zalduondo, L.; Xue, Y.; Vogel, P.; Channappanavar, R.; Jonsson, C.B.; et al. Time-Dependent Increase in Susceptibility and Severity of Secondary Bacterial Infections During SARS-CoV-2. Front. Immunol. 2022, 13, 894534. [Google Scholar] [CrossRef] [PubMed]
- Jochems, S.P.; Ferreira, D.M.; Smits, H.H. Microbiota and compartment matter in the COVID-19 response. Nat. Immunol. 2021, 22, 1350–1352. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.; Weinberger, D.M.; Satzke, C. 2023. Thematic issue on bacterial–viral co-infections. FEMS Microbes 2023, 4, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Schippa, S.; Frassanito, A.; Marazzato, M.; Nenna, R.; Petrarca, L.; Neroni, B.; Bonfiglio, G.; Guerrieri, F.; Frasca, F.; Oliveto, G.; et al. Nasal microbiota in RSV bronchiolitis. Microorganisms 2020, 8, 731. [Google Scholar] [CrossRef] [PubMed]
- Morens, D.M.; Taubenberger, J.K.; Fauci, A.S. Predominant role of bacterial pneumonia as a cause of death in pandemic influenza: implications for pandemic influenza preparedness. J. Infect. Dis. 2008, 198, 962–970. [Google Scholar] [CrossRef]
- Morris, D.E.; Cleary, D.W.; Clarke, S.C. 2017. Secondary bacterial infections associated with influenza pandemics. Front. Microbiol. 2017, 8, 1041. [Google Scholar] [CrossRef]
- Prasso, J.E.; Deng, J.C. Postviral complications: bacterial pneumonia. Clin. Chest Med. 2017, 38, 127–138. [Google Scholar] [CrossRef]
- Tilocca, B.; Soggiu, A.; Musella, V.; Britti, D.; Sanguinetti, M.; Urbani, A.; Roncada, P. Molecular basis of COVID-19 relationships in different species: a one health perspective. Microbes Infect. 22, 218–220. [CrossRef]
- Fahkrajang, W.; Sudaryatma, P.E.; Mekata, H.; Hamabe, S.; Saito. A.; Okabayashi, T. Bovine respiratory coronavirus enhances bacterial adherence by upregulating expression of cellular receptors on bovine respiratory epithelial cells. Vet. Microbiol. 2021, 255, 109017. [Google Scholar] [CrossRef]
- Frucchi, A.P.S.; Dall Agnol, A.M.; Bronkhorst, D.E.; Beuttemmuller, E.A.; Alfieri, A.A.; Alfieri, A.F. Bovine coronavirus co-infection and molecular characterization in dairy calves with or without clinical respiratory disease. Front. Vet. Sci. 2022, 9, 895492. [Google Scholar] [CrossRef]
- Nefedova, E.; Koptev, V.; Bobikova, A.S.; Cherepushkina, V.; Mironova, T.; Afonyushkin, V.; Shkil, N.; Donchenko, N.; Kozlova, Y.; Sigareva, N.; Davidova, N. The infectious bronchitis coronavirus pneumonia model presenting a novel insight for the SARS-CoV-2 dissemination route. Vet. Sci. 2021, 8, 239. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.H.; Cook, J.K.; Parsell, Z.E. The experimental infection of chickens with mixtures of infectious bronchitis virus and Escherichia coli. J. Gen. Virol. 1985, 66, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Cook, J.K.; Frazier, J.A.; Narita, M. Escherichia coli multiplication and lesions in the respiratory tract of chickens inoculated with infectious bronchitis virus and/or E. coli. Avian Dis. 1992, 881–890. [Google Scholar] [CrossRef]
- Hoerr, F.J. The pathology of infectious bronchitis. Avian Dis. 2021, 65, 600–611. [Google Scholar] [CrossRef] [PubMed]
- Dudoignon, E.; Caméléna, F.; Deniau, B.; Habay, A.; Coutrot, M.; Ressaire, Q.; Plaud, B.; Berçot, B.; Dépret, F. Bacterial pneumonia in COVID-19 critically ill patients: a case series. Clin. Infect. Dis. 2021, 72, 905–906. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Dela Cruz, C.S.; Sharma, L. Coronaviruses, Lysosomes, and Secondary Bacterial Infections: Coronaviruses Outsmart the Host. DNA Cell Biol. 2023, 42, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Cohen, R.; Finn, T.; Babushkin, F.; Geller, K.; Alexander, H.; Shapiro, M.; Uda, M.; Mostrchy, A.R.; Amash, R.; Shimoni, Z.; Paikin, S. High rate of bacterial respiratory tract co-infections upon admission amongst moderate to severe COVID-19 patients. Infect. Dis. 2022, 54, 134–144. [Google Scholar] [CrossRef]
- Weidmann, M.D.; Berry, G.J.; Zucker, J.E.; Huang, S.; Sobieszczyk, M.E.; Green, D.A. Bacterial Pneumonia and Respiratory Culture Utilization among Hospitalized Patients with and without COVID-19 in a New York City Hospital. J. Clin. Microbiol. 2022, 60, e00174–22. [Google Scholar] [CrossRef]
- De Bruyn, A.; Verellen, S.; Bruckers, L.; Geebelen, L.; Callebaut, I.; De Pauw, I.; Stessel, B.; and Dubois, J. Secondary infection in COVID-19 critically ill patients: a retrospective single-center evaluation. BMC Infect. Dis. 2022, 22, 207. [Google Scholar] [CrossRef]
- Velásquez-Garcia, L.; Mejia-Sanjuanelo, A.; Viasus, D.; Carratalà, J. Causative Agents of Ventilator-Associated Pneumonia and Resistance to Antibiotics in COVID-19 Patients: A Systematic Review. Biomedicines 2022, 10, 1226. [Google Scholar] [CrossRef]
- Ortega-Peña, S.; Rodríguez-Martínez, S.; Cancino-Diaz, M.E.; Cancino-Diaz, J.C. Staphylococcus epidermidis Controls Opportunistic Pathogens in the Nose, Could It Help to Regulate SARS-CoV-2 (COVID-19) Infection? Life 2022, 12, 341. [Google Scholar] [CrossRef] [PubMed]
- Raineri, E.J.; Altulea, D.; van Dijl, J.M. Staphylococcal trafficking and infection—from ‘nose to gut’and back. FEMS Microbiol. Rev. 2022, 46, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Park, A.M.; Khadka, S.; Sato, F.; Omura, S.; Fujita, M.; Hashiwaki, K.; Tsunoda, I. Bacterial and fungal isolation from face masks under the COVID-19 pandemic. Sci. Rep. 2022, 12, 11361. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, P., Masiuk, H., Kulig, P., Skoryk, A., Wcisłek, A., Jursa-Kulesza, J., Sarna, A., Sławiński, M., Kotowski, M., Tejchman, K. and Kotfis, K. Medical Face Masks Do Not Affect Acid–Base Balance Yet Might Facilitate the Transmission of Staphylococcus aureus in Hospital Settings during the COVID-19 Pandemic. Int. J. Environ. Res. Public Health 2023, 20, 2474. [CrossRef]
- Kwon, M.; Yang, W. Effects of face masks and acoustical environments on speech recognition by preschool children in an auralised classroom. Appl. Acoust. 2023, 202, 109149. [Google Scholar] [CrossRef]
- Dietert, R.R.; Dietert, J.M. Using microbiome-based approaches to deprogram chronic disorders and extend the healthspan following adverse childhood experiences. Microorganisms 2022, 10, 229. [Google Scholar] [CrossRef]
- Beckers, T.; Hermans, D.; Lange, I.; Luyten, L.; Scheveneels, S.; Vervliet, B. Understanding clinical fear and anxiety through the lens of human fear conditioning. Nat. Rev. Psychol. 2023, 2, 233–245. [Google Scholar] [CrossRef]
- Mertens, G.; Lodder, P.; Smeets, T.; Duijndam, S. Pandemic panic? Results of a 14-month longitudinal study on fear of COVID-19. J. Affect. Disord. 2023, 322, 15–23. [Google Scholar] [CrossRef]
- Hauck, A.; Michael, T.; Ferreira de Sá, D.S. Fear learning and generalization during pandemic fear: How COVID-19-related anxiety affects classical fear conditioning with traumatic film clips. J. Psychiatr. Res. 2022, 155, 90–99. [Google Scholar] [CrossRef]
- Cannito, L.; Di Crosta, A.; Palumbo, R.; Ceccato, I.; Anzani, S.; La Malva, P.; Palumbo, R.; Di Domenico, A. Health anxiety and attentional bias toward virus-related stimuli during the COVID-19 pandemic. Sci. Rep. 2020, 10, 16476. [Google Scholar] [CrossRef]
- Meaklim, H.; Saunders, W.J.; Byrne, M.L.; Junge, M.F.; Varma, P.; Finck, W.A.; Jackson, M.L. Insomnia is a key risk factor for persistent anxiety and depressive symptoms: A 12-month longitudinal cohort study during the COVID-19 pandemic. J. Affect. Disord. 2023, 322, 52–62. [Google Scholar] [CrossRef]
- Dietert, R.R.; DeWitt, J.C.; Germolec, D.R.; Zelikoff., J.T. Breaking patterns of environmentally influenced disease for health risk reduction: immune perspectives. Environ. Health Perspect. 2010, 118, 1091–1099. [Google Scholar] [CrossRef]
- Lerouge, R., Lema, M.D. and Arnaboldi, M., 2023. The role played by government communication on the level of public fear in social media: An investigation into the Covid-19 crisis in Italy. Government Information Quarterly 2023, p.101798. [CrossRef]
- Feng, P.; Chen, Z.; Becker, B.; Liu, X.; Zhou, F.; He, Q.; Qiu, J.; Lei, X.; Chen, H.; Feng, T. Predisposing Variations in Fear-Related Brain Networks Prospectively Predict Fearful Feelings during the 2019 Coronavirus (COVID-19) Pandemic. Cereb. Cortex. 2022. 32. 540-553. [CrossRef]
- Józefacka, N.M., Karpiński, E.A., Superson, B., Kołek, M.F., Skrzypczak, A.R. and Kania, G., 2023. Potential Factors Conditioning the Compliance to Mandatory Face Covering in the Public Space Due to SARS-CoV-2 Pandemic. Int. J. Environ. Res. Public Health 2023, 20, 726. [CrossRef]
- Altintas, E., El Haj, M., Boudoukha, A.H., Olivier, C., Lizio, A., Luyat, M. and Gallouj, K., 2022. Emotional exhaustion and fear of COVID-19 in geriatric facilities during the COVID-19 pandemic. Int. J. Geriatr. Psychiatry 2022, 37. [CrossRef]
- Baldi, E.; Savastano, S. Fear of Contagion: One of the Most Devious Enemies to Fight During the COVID-19 Pandemic. Disaster Med. Public Health. Prep. 2021, 15, e8–e9. [Google Scholar] [CrossRef]
- Muñoz-Vela, F.J.; Rodríguez-Díaz, L.; Gómez-Salgado, J.; Fernández-Carrasco, F.J.; Allande-Cussó, R.; Vázquez-Lara, J.M.; Fagundo-Rivera, J. Fear and Anxiety in Pregnant Women During the COVID-19 Pandemic: A Systematic Review. Int. J. Public Health 2023, 68, 1605587. [Google Scholar] [CrossRef]
- Albikawi, Z.F. Fear Related to COVID-19, Mental Health Issues, and Predictors of Insomnia among Female Nursing College Students during the Pandemic. Healthcare 2023, 11, 174. [Google Scholar] [CrossRef] [PubMed]
- Kerker, B.D.; Willheim, E.; Weis, J.R. The COVID-19 pandemic: implications for maternal mental health and early childhood development. Am. J. Health Promot. 2023, 37, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Shechner, T.; Hong, M.; Britton, J.C.; Pine, D.S.; Fox, N.A. Fear conditioning and extinction across development: evidence from human studies and animal models. Biological Psychol. 2014, 100, 1–12. [Google Scholar] [CrossRef]
- Salinas-Hernández, X.I.; and Duvarci, S. 2021. Dopamine in fear extinction. Front. Synaptic Neurosci. 2021, 13, 635879. [Google Scholar] [CrossRef] [PubMed]
- Furini, C.; Myskiw, J.; Izquierdo, I. The learning of fear extinction. Neurosci. Biobehav. Rev. 47, 670–683. [CrossRef] [PubMed]
- Leblanc, H.; Ramirez, S. Linking Social Cognition to Learning and Memory. J. Neurosci. 2020, 40, 8782–8798. [Google Scholar] [CrossRef]
- Donato, F.; Alberini, C.M.; Amso, D.; Dragoim, G.; Dranovskym, A.; Newcombe, N.S. The Ontogeny of Hippocampus-Dependent Memories. J. Neurosci. 2021, 41, 920–926. [Google Scholar] [CrossRef]
- Sosa, M.; Giocomo, L.M. Navigating for reward. Nat. Rev. Neurosci. 2021, 22, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Takehara-Nishiuchi, K. Neurobiology of systems memory consolidation. Eur. J. Neurosci. 2021, 54, 6850–6863. [Google Scholar] [CrossRef]
- Banquet, J.P.; Gaussier, P.; Cuperlier, N.; Hok, V.; Save, E.; Poucet, B.; Quoy, M.; Wiener, S.I. Time as the fourth dimension in the hippocampus. Prog. Neurobiol. 2021, 199, 101920. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, J.Z. From structure to behavior in basolateral amygdala-hippocampus circuits. Front. Neural Circuits 2017, 11, 86. [Google Scholar] [CrossRef]
- Gage, N.M.; Baars, B.J. 2018. Humans Are Social Beings. Fundamentals of Cognitive Neuroscience: A Beginners Guide, 2nd ed.; Academic Press: Cambridge, Massachusetts, USA, 2018; pp. 321–356. [Google Scholar]
- Tang, W.; Meng, Z.; Li, N.; Liu, Y.; Li, L.; Chen, D.; Yang, Y. 2021. Roles of gut microbiota in the regulation of hippocampal plasticity, inflammation, and hippocampus-dependent behaviors. Front. Cell. Infect. Microbiol. 2021, 10, 611014. [Google Scholar] [CrossRef]
- Sharvin, B.L.; Aburto, M.R.; Cryan, J.F. Decoding the neurocircuitry of gut feelings: Region-specific microbiome-mediated brain alterations. Neurobiol. Dis. 2023, 179, 106033. [Google Scholar] [CrossRef]
- Laudani, S.; Torrisi, S.A.; Alboni, S.; Bastiaanssen, T.F.; Benatti, C.; Rivi, V.; Moloney, R.D.; Fuochi, V.; Furneri, P.M.; Drago, F.; Salomone, S. Gut microbiota alterations promote traumatic stress susceptibility associated with p-cresol-induced dopaminergic dysfunctions. Brain Behav. Immun. 2023, 107, 385–396. [Google Scholar] [CrossRef]
- Swer, N.M.; Venkidesh, B.; Murali, T.S.; et al. Gut microbiota-derived metabolites and their importance in neurological disorders. Mol. Biol. Rep. 2023, 50, 1663–1675. [Google Scholar] [CrossRef]
- Gacias, M.; Gaspari, S.; Santos, P.M.G.; Tamburini, S.; Andrade, M.; Zhang, F.; Shen, N.; Tolstikov, V.; Kiebish, M.A.; Dupree, J.L.; and Zachariou, V. 2016. Microbiota-driven transcriptional changes in prefrontal cortex override genetic differences in social behavior. Elife 2016, 5, e13442. [Google Scholar] [CrossRef]
- Hamamah, S.; Aghazarian, A.; Nazaryan, A.; Hajnal, A.; Covasa, M. Role of microbiota-gut-brain axis in regulating dopaminergic signaling. Biomedicines 2022, 10, 436. [Google Scholar] [CrossRef]
- Cryan, J.F.; O'Riordan, K.J.; Cowan, C.S.; Sandhu, K.V.; Bastiaanssen, T.F.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The microbiota-gut-brain axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; and Cong, Y. 2021. Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseases. Cell. Mol. Immunol. 2021, 18, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in health and diseases. Signal Transduct. Target Ther. 2022, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.L.; Garcia-Reyero, N.; Martyniuk, C.J.; Tubbs, C.W.; and Bisesi Jr, J.H. Regulation of endocrine systems by the microbiome: perspectives from comparative animal models. Gen. Comp. Endocrinol. 2020, 2020 292, 113437. [Google Scholar] [CrossRef]
- Cheng, W.Y.; Ho, Y.S.; and Chang, R.C.C. Linking circadian rhythms to microbiome-gut-brain axis in aging-associated neurodegenerative diseases. Ageing Res. Rev. 2022, 78, 101620. [Google Scholar] [CrossRef]
- Sorboni, S.G.; Moghaddam, H.S.; Jafarzadeh-Esfehani, R.; Soleimanpour, S. A comprehensive review on the role of the gut microbiome in human neurological disorders. Clin. Microbiol. Rev. 2022, 35, e00338–20. [Google Scholar] [CrossRef]
- Miri, S.; Yeo, J.; Abubaker, S.; Hammami, R. Neuromicrobiology, an emerging neurometabolic facet of the gut microbiome? Front. Microbiol. 2023, 14, 1098412. [Google Scholar] [CrossRef]
- Fox, M.; Lee, S.M.; Wiley, K.S.; Lagishetty, V.; Sandman, C.A.; Jacobs, J.P.; Glynn, L.M. Development of the infant gut microbiome predicts temperament across the first year of life. Dev. Psychopathol. 2022, 34, 1914–1925. [Google Scholar] [CrossRef]
- Lynch, C.M.; Cowan, C.S.; Bastiaanssen, T.F.; Moloney, G.M.; Theune, N.; van de Wouw, M.; Zanuy, E.F.; Ventura-Silva, A.P.; Codagnone, M.G.; Villalobos-Manríquez, F.; et al. Critical windows of early-life microbiota disruption on behaviour, neuroimmune function, and neurodevelopment. Bran, Behav. Immun. 2023, 108, 309–327. [Google Scholar] [CrossRef]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W. Psychobiotics and the manipulation of bacteria–gut–brain signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef]
- Deepika, A.K. Shukla; Kumari, A.; and Kumar, A. Gut brain regulation using psychobiotics for improved neuropsychological illness. Developmental Psychobiology, 2023, 65, e22404. [Google Scholar] [CrossRef] [PubMed]
- Berding, K.; Bastiaanssen, T.F.; Moloney, G.M.; Boscaini, S.; Strain, C.R.; Anesi, A.; Long-Smith, C.; Mattivi, F.; Stanton, C.; Clarke, G.; Dinan, T.G. Feed your microbes to deal with stress: A psychobiotic diet impacts microbial stability and perceived stress in a healthy adult population. Mol. Psychiatry 2023, 28, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, D.; Karbowiak, M.; and Brzezicka, A. The Role of Psychobiotics to Ensure Mental Health during the COVID-19 Pandemic—A Current State of Knowledge. International Journal of Environmental Research and Public Health, 19(17), p.11022. Int. J. Environ. Res. Public Health 2022, 19, 11022. [Google Scholar] [CrossRef] [PubMed]
- Dursun, P.; Alyagut, P.; Yılmaz, I. Meaning in life, psychological hardiness and death anxiety: individuals with or without generalized anxiety disorder (GAD). Curr. Psychol. 2022, 41, 3299–3317. [Google Scholar] [CrossRef] [PubMed]
- Sangha, S.; Diehl, M.M.; Bergstrom, H.C.; Drew, M.R. 2020. Know safety, no fear. Neurosci. Biobehav. Rev. 2020, 108, 218–230. [Google Scholar] [CrossRef]
- Wen, Z.; Chen, Z.S.; Milad, M.R. Fear extinction learning modulates large-scale brain connectivity. Neuroimage 2021, 238, 118261. [Google Scholar] [CrossRef]
- Crumeyrolle-Arias, M.; Jaglin, M.; Bruneau, A.; Vancassel, S.; Cardona, A.; Daugé, V.; Naudon, L.; Rabot, S. Absence of the gut microbiota enhances anxiety-like behavior and neuroendocrine response to acute stress in rats. Psychoneuroendocrinology 2014, 42, 207–217. [Google Scholar] [CrossRef]
- Lach, G.; Fülling, C.; Bastiaanssen, T.F.; Fouhy, F.; Donovan, A.N.O.; Ventura-Silva, A.P.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Enduring neurobehavioral effects induced by microbiota depletion during the adolescent period. Transl. Psychiatry 2020, 10, 382. [Google Scholar] [CrossRef]
- Cui, X.; Zhou, S.; Xia, G.; Chen, J.; Jiang, L.; Huang, J.; and Tong, J. A multispecies probiotic accelerates fear extinction and inhibits relapse in mice: role of microglia. Neuropharmacology 2021, 193, 108613. [Google Scholar] [CrossRef]
- Chu, C.; Murdock, M.H.; Jing, D.; Won, T.H.; Chung, H.; Kressel, A.M.; Tsaava, T.; Addorisio, M.E.; Putzel, G.G.; Zhou, L.; Bessman, N.J. The microbiota regulate neuronal function and fear extinction learning. Nature 2019, 574, 543–548. [Google Scholar] [CrossRef]
- Cowan, C.S.; Stylianakis, A.A.; Richardson, R. Early-life stress, microbiota, and brain development: probiotics reverse the effects of maternal separation on neural circuits underpinning fear expression and extinction in infant rats. Dev. Cogn. Neurosci. 2019, 37, 100627. [Google Scholar] [CrossRef]
- Fox, J.H.; Hassell Jr, J.E.; Siebler, P.H.; Arnold, M.R.; Lamb, A.K.; Smith, D.G.; Day, H.E.; Smith, T.M.; Simmerman, E.M.; Outzen, A.A.; et al. Preimmunization with a heat-killed preparation of Mycobacterium vaccae enhances fear extinction in the fear-potentiated startle paradigm. Brain Behave. Immun. 2017, 66, 70–84. [Google Scholar] [CrossRef]
- Maeng, L.Y.; and Beumer, A. Never fear, the gut bacteria are here: Estrogen and gut microbiome-brain axis interactions in fear extinction. Int. J. Psychophysiol. 2023, 189, 66–75. [Google Scholar] [CrossRef]
- Chbeir, S.; Carrión, V. 2023. Resilience by design: How nature, nurture, environment, and microbiome mitigate stress and allostatic load. World J. Psychiatry 2023, 13, 144. [Google Scholar] [CrossRef]
- Tyagi, P.; Tasleem, M.; Prakash, S.; Chouhan, G. Intermingling of gut microbiota with brain: Exploring the role of probiotics in battle against depressive disorders. Food Research International 2020, 137, 109489. [Google Scholar] [CrossRef]
- Zhu, F.; Tu, H.; Chen, T. The Microbiota–Gut–Brain Axis in Depression: The Potential Pathophysiological Mechanisms and Microbiota Combined Antidepression Effect. Nutrients 2022, 14, 2081. [Google Scholar] [CrossRef]
- Kumar, A.; Pramanik, J.; Goyal, N.; Chauhan, D.; Sivamaruthi, B.S.; Prajapati, B.G.; Chaiyasut, C. 2023. Gut Microbiota in Anxiety and Depression: Unveiling the Relationships and Management Options. Pharmaceuticals 2023, 16, 565. [Google Scholar] [CrossRef]
- Lebois, L.A.; Seligowski, A.V.; Wolff, J.D.; Hill, S.B.; Ressler, K.J. Augmentation of extinction and inhibitory learning in anxiety and trauma-related disorders. Annu. Rev. Clin. Psychol. 2019, 15, 257–284. [Google Scholar] [CrossRef]
- Berding, K.; and Cryan, J.F. Microbiota-targeted interventions for mental health. Curr. Opin. Psychiatry 2022, 35, 3. [Google Scholar] [CrossRef]
- Sherwin, E., Bordenstein, S.R., Quinn, J.L., Dinan, T.G. and Cryan, J.F., 2019. Microbiota and the social brain. Science 2019, 366, eaar2016. [CrossRef]
- Maren, S.; Holmes, A. Stress and Fear Extinction. Neuropsychopharmacol. 2016, 41, 58–79. [Google Scholar] [CrossRef]
- Pace-Schott, E.F.; Seo, J.; Bottary, R. The influence of sleep on fear extinction in trauma-related disorders. Neurobiol. Stress 2023, 22, 100500. [Google Scholar] [CrossRef]
- Wilson, D.R.; Binford, L.; Hickson, S. The Gut Microbiome and Mental Health. J. Holist Nurs. 2023, Apr 20:8980101231170487. [CrossRef]
- Cooper, S.E.; Dunsmoor, J.E. 2021. Fear conditioning and extinction in obsessive-compulsive disorder: a systematic review. Neurosci. Biobehav. Rev. 2021, 129, 75–94. [Google Scholar] [CrossRef]
- Zuj, D.V.; Palmer, M.A.; Lommen, M.J.; Felmingham, K.L. The centrality of fear extinction in linking risk factors to PTSD: A narrative review. Neurosci. Biobehav. Rev. 2016, 69, 15–35. [Google Scholar] [CrossRef]
- Cohn, M.D.; van Lith, K.; Kindt, M.; Pape, L.E.; Doreleijers, T.A.; van den Brink, W.; Veltman, D.J.; Popma, A. Fear extinction, persistent disruptive behavior and psychopathic traits: fMRI in late adolescence. Soc. Cogn. Affect. Neurosci. 2016, 11, 1027–1035. [Google Scholar] [CrossRef]
- Muench, C.l.; Charlet, K.; Balderston, N.L.; Grillon, C.; Heilig, M.; Cortes, C.R.; Momenan, R.; Lohoff, F.W. Fear conditioning and extinction in alcohol dependence: evidence for abnormal amygdala reactivity. Addict. Biol. 2021, 26, e12835. [Google Scholar] [CrossRef]
- Spencer, A.E.; Marin, M.F.; Milad, M.R.; Spencer, T.J.; Bogucki, O.E.; Pope, A.L.; Plasencia, N.; Hughes, B.; Pace-Schott, E.F.; Fitzgerald, M.; et al. Abnormal fear circuitry in attention deficit hyperactivity disorder: a controlled magnetic resonance imaging study. Psychiatry Research: Neuroimaging 2017, 262, 55–62. [Google Scholar] [CrossRef]
- Craske, M.G.; Sandman, C.F.; Stein, M.B. 2022. How can neurobiology of fear extinction inform treatment? Neurosci. Biobehav. Rev. 2022, 143, 104923. [Google Scholar] [CrossRef]
- Luijten, M.A.; van Muilekom, M.M.; Teela, L.; Polderman, T.J.; Terwee, C.B.; Zijlmans, J.; Klaufus, L.; Popma, A.; Oostrom, K.J.; van Oers, H.A.; Haverman, L. The impact of lockdown during the COVID-19 pandemic on mental and social health of children and adolescents. Qual. Life Res. 30, 2795–2804. [CrossRef]
- Panchal, U.; Salazar de Pablo, G.; Franco, M.; Moreno, C.; Parellada, M.; Arango, C.; Fusar-Poli, P. The impact of COVID-19 lockdown on child and adolescent mental health: systematic review. Eur. Child Adolesc. Psychiatry 2021, 1–27. [Google Scholar] [CrossRef]
- Cellini, N.; Di Giorgio, E.; Mioni, G.; Di Riso, D. Sleep and psychological difficulties in Italian school-age children during COVID-19 lockdown. J. Pediatr. Psychol. 2021, 46, 153–167. [Google Scholar] [CrossRef]
- Christner, N.; Essler, S.; Hazzam, A.; Paulus, M. Children’s psychological well-being and problem behavior during the COVID-19 pandemic: An online study during the lockdown period in Germany. PloS One 2021, 16, e0253473. [Google Scholar] [CrossRef]
- Amicucci, G.; Salfi, F.; D’Atri, A.; Viselli, L.; Ferrara, M. The differential impact of COVID-19 lockdown on sleep quality, insomnia, depression, stress, and anxiety among late adolescents and elderly in Italy. Brain Sci. 2021, 11, 1336. [Google Scholar] [CrossRef]
- Dietert, R.R.; Dietert, J.M. Using the Internet of Microbes to Survive the Assault on the Human Microbiome. Am. J. Biomed. Biol. Res. 2023, 19, 71–76. [Google Scholar] [CrossRef]
- Ghannoum, M.A.; Ford, M.; Bonomo, R.A.; Gamal, A.; McCormick, T.S. A microbiome-driven approach to combating depression during the COVID-19 pandemic. Front. Nutr. 2021, 8, 576. [Google Scholar] [CrossRef]
- Athanassi, A.; Dorado Doncel, R.; Bath, K.G.; Mandairon, N. Relationship between depression and olfactory sensory function: a review. Chem. Senses 2021, 46, 1–12. [Google Scholar] [CrossRef]
- Lawrence, B.J.; Jayakody, D.M.; Bennett, R.J.; Eikelboom, R.H.; Gasson, N.; Friedland, P.L. Hearing loss and depression in older adults: a systematic review and meta-analysis. Gerontologist 2020, 60, e137–e154. [Google Scholar] [CrossRef]
- Hur, K.; Choi, J.S.; Zheng, M.; Shen, J.; Wrobel, B. Association of alterations in smell and taste with depression in older adults. Laryngoscope Investing. Otolaryngol. 2018, 3, 94–99. [Google Scholar] [CrossRef]
- Vithoulkas, G.; Muresanu, D.F. Conscience and consciousness: a definition. J. Med. Life 2014, 7, 104–108. [Google Scholar]
- Dietert, R.R. Microbiome First Approaches to Rescue Public Health and Reduce Human Suffering. Biomedicines 2021, 9, 1581. [Google Scholar] [CrossRef]
- Werber, Y.; Natan, E.; Lavner, Y.; and Vortman, Y. Antibiotics affect migratory restlessness orientation. J. Ethol. 2022, 40, 175–180. [Google Scholar] [CrossRef]
- Simon, R.A.; Ranasinghe, P.D.; Barazanji, N.; Jungeström, M.B.; Xu, J.; Bednarska, O.; Serrander, L.; Engström, M.; Bazylinski, D.A.; Keita, Å.V.; Walter, S. Magnetotactic bacteria from the human gut microbiome associated with orientation and navigation regions of the brain. J. of Oceanol. Limnol. 2021, 39, 2044–2052. [Google Scholar] [CrossRef]
- Schamarek, I.; Anders, L.; Chakaroun, R.M.; Kovacs, P.; Rohde-Zimmermann, K. The role of the oral microbiome in obesity and metabolic disease: potential systemic implications and effects on taste perception. Nutr. J. 2023, 22, 1–13. [Google Scholar] [CrossRef]
- Leung, R.; and Covasa, M. Do gut microbes taste? Nutrients 2021, 13, 2581. [Google Scholar] [CrossRef]
- Doyle, M.E.; Premathilake, H.U.; Yao, Q.; Mazucanti, C.H.; and Egan, J.M. 2023. Physiology of the tongue with emphasis on taste transduction. Physiol. Rev. 2023, 103, 1193–1246. [Google Scholar] [CrossRef]
- Thangaleela, S.; Sivamaruthi, B.S.; Kesika, P.; Bharathi, M.; Chaiyasut, C. Nasal Microbiota, Olfactory Health, Neurological Disorders and Aging—A Review. Microorganisms 2022, 10, 1405. [Google Scholar] [CrossRef]
- López-Dávalos, P.C.; Requena, T.; Pozo-Bayón, M.Á.; and Muñoz-González, C. Decreased retronasal olfaction and taste perception in obesity are related to saliva biochemical and microbiota composition. Food Research International 2023, 167, 112660. [Google Scholar] [CrossRef]
- Kociszewska, D.; Chan, J.; Thorne, P.R.; and Vlajkovic, S.M. The link between gut dysbiosis caused by a high-fat diet and hearing loss. Int. J. of Mol. Sci. 2021, 22, 13177. [Google Scholar] [CrossRef]
- Kociszewska, D.; and Vlajkovic, S. 2022. Age-related hearing loss: the link between inflammaging, immunosenescence, and gut dysbiosis. International Journal of Molecular Sciences 2022, 23, 7348. [Google Scholar] [CrossRef]
- Lee, J.S.; Lee, S.M.; Son, H.S.; Yoon, Y.N.; Shin, J.E.; Sul, W.J.; Yu, D.; Choe, Y.B.; Lee, Y.W. Analysis of the Microbiome of the Ear Canal in Normal Individuals and Patients with Chronic Otitis Externa. Ann. Dermatol. 2022, 34, 461–471. [Google Scholar] [CrossRef]
- Jörissen, J.; van den Broek, M.F.; De Boeck, I.; Van Beeck, W.; Wittouck, S.; Boudewyns, A.; Van de Heyning, P.; Topsakal, V.; Van Rompaey, V.; Wouters, I.; and Van Heirstraeten, L. Case-Control Microbiome Study of Chronic Otitis Media with Effusion in Children Points at Streptococcus salivarius as a Pathobiont-Inhibiting Species. Msystems 2021, 6, e00056–21. [Google Scholar] [CrossRef]
- Lin, B.; Wang, Y.; Zhang, P.; Yuan, Y.; Zhang, Y.; Chen, G. Gut microbiota regulates neuropathic pain: potential mechanisms and therapeutic strategy. J. Headache Pain 2020, 21, 1–16. [Google Scholar] [CrossRef]
- Keum, H.L.; Kim, H.; Kim, H.J.; Park, T.; Kim, S.; An, S.; Sul, W.J. Structures of the skin microbiome and mycobiome depending on skin sensitivity. Microorganisms 2020, 8, 1032. [Google Scholar] [CrossRef]
- Skowron, K.; Bauza-Kaszewska, J.; Kraszewska, Z.; Wiktorczyk-Kapischke, N.; Grudlewska-Buda, K.; Kwiecińska-Piróg, J.; Wałecka-Zacharska, E.; Radtke, L.; Gospodarek-Komkowska, E. Human skin microbiome: Impact of intrinsic and extrinsic factors on skin microbiota. Microorganisms 2021, 9, 543. [Google Scholar] [CrossRef]
- Olunoiki, E.; Rehner, J.; Bischoff, M.; Koshel, E.; Vogt, T.; Reichrath, J.; Becker, S.L. Characteristics of the Skin Microbiome in Selected Dermatological Conditions: A Narrative Review. Life 2022, 12, 1420. [Google Scholar] [CrossRef]
- Zysset-Burri, D.C.; Morandi, S.; Herzog, E.L.; Berger, L.E.; Zinkernagel, M.S. The role of the gut microbiome in eye diseases. Progr. Retin. Eye Res. 2023, 92, 101117. [Google Scholar] [CrossRef]
- Peter, V.G.; Morandi, S.C.; Herzog, E.L.; Zinkernagel, M.S.; Zysset-Burri, D.C. Investigating the Ocular Surface Microbiome: What Can It Tell Us? Clin. Ophthalmol. 2023, 17, 259–271. [Google Scholar] [CrossRef]
- Aragona, P.; Baudouin, C.; Del Castillo, J.M.B.; Messmer, E.; Barabino, S.; Merayo-Lloves, J.; Brignole-Baudouin, F.; Inferrera, L.; Rolando, M.; Mencucci, R.; Rescigno, M. The ocular microbiome and microbiota and their effects on ocular surface pathophysiology and disorders. Surv. Ophthalmol. 2021, 66, 907–925. [Google Scholar] [CrossRef]
- Borroni, D.; Paytuví-Gallart, A.; Sanseverino, W.; Gómez-Huertas, C.; Bonci, P.; Romano, V.; Giannaccare, G.; Rechichi, M.; Meduri, A.; Oliverio, G.W.; et al. Exploring the healthy eye microbiota niche in a multicenter study. Int. J. Mol. Sci. 2022, 23, 10229. [Google Scholar] [CrossRef]
- An, Q.; Zou, H. Ocular surface microbiota dysbiosis contributes to the high prevalence of dry eye disease in diabetic patients. Crit. Rev. Microbiol. 2022, 1–10. [Google Scholar] [CrossRef]
- Deng, Y.; Ge, X.; Li, Y.; Zou, B.; Wen, X.; Chen, W.; Lu, L.; Zhang, M.; Zhang, X.; Li, C.; Zhao, C. Identification of an intraocular microbiota. Cell Discov. 2021, 7, 13. [Google Scholar] [CrossRef]

Table 1.
Examples of Institutionally-Promoted Covid-19 Fear-of-Contagion, Vulnerable Populations, and Adverse Outcomes.
Table 1.
Examples of Institutionally-Promoted Covid-19 Fear-of-Contagion, Vulnerable Populations, and Adverse Outcomes.
| Group Studied | Findings | Reference(s) |
|---|---|---|
| Italian government and media |
Messaging emphasizing life-changing scenarios produced the greatest fear among the public. The researchers emphasized that government must balance the level of fear with proper causation or risk public psychological harm. | [65] |
|
Study of international population with the majority being European young adults. |
Found that those who consulted regular media, professional websites, and social media for additional pandemic information experienced the greatest fear. |
[60] |
| Study of Chinese population using pre-pandemic MRIs compared with later pandemic fear responses. |
Study revealed severe impact resulting from fear of contagion during the pandemic. The pre-pandemic neural connectome pattern could predict those who would experience the most pandemic fear. | [66] |
| An online study with majority female participants. |
Researchers observed a significant moderate positive correlation between anxiety around Covid-19 and adherence to the mandates. | [67] |
| Study of young adult university students in Germany | Researchers found that those students with high COVID-Anxiety exhibited poorer discrimination performance between fear and safety cues. |
[61] |
| International study with 2,069 majority female participants |
Pre-existing or new-onset insomnia elevated the risk of affective disorder outcomes (anxiety and/or depression symptoms) during the pandemic. |
[63] |
| Study included 132 Italian individuals with the majority female. |
Researchers found that higher health anxiety in general predicted attentional bias toward a fear of Covid-19 contagion. | [62] |
| Study of 118 healthcare workers in French geriatric facilities | Researchers found a significant increase in emotional exhaustion among the workers and this was related to both increased demands at work and increased fear of contagion. | [68] |
| Review concerning Milan Italy region fear of hospital contagion and reduction in available emergency services for cardiac events |
Discussion of the high rate of acute coronary syndrome observed out of the hospital with low hospital emergency service use | [69] |
| Review of research studies on fear and anxiety among pregnant women during the Covid-19 pandemic |
The researchers reported a high prevalence of fear and anxiety with significant impact on mental health. Intolerance of uncertainty was identified as one of the risk factors. | [70] |
| A study of Covid-19 fear and mental health issues among 154 female nursing students in Saudi Arabia |
Fear of Covid-19 contagion, depression, anxiety and insomnia were highly prevalent among the students with fear of contagion the most prevalent (79.3%). | [71] |
| Review article focused on highly vulnerable populations for pandemic fear-based mental health challenges | Women and young children were identified as highly vulnerable with mandates such as social isolation and school closures exacerbating fear-based anxiety and depression. | [72] |
Table 2.
Examples of Studies and Reviews on Microbiota Regulation of Fear Extinction.
| Study | Significant Findings | References(s) |
|---|---|---|
| Early study on germ-free and specific pathogen free rats | Absence of a microbiome produced HPA axis disruption, brain function changes and significantly heightened anxiety behavior and lack of cognitive flexibility. | [105] |
| Research on mice concerning the neurobehavioral effects of transitory microbiome depletion in different age groups. |
Adolescent mice were more sensitive to transitory microbiome depletion than adults. Short-term depletion produced long-lasting shifts in fear-based learning, heightened anxiety-like behaviors and changes in amygdala gene expression in adolescents. | [106] |
| Study examined the effects of multi-species probiotic (Bifidobacterium longum Lactobacillus acidophilus and Enterococcus faecalis) administration on recovery of fear memory after fear conditioning |
Probiotic administration in mice modulated fear conditioning-induced microbial dysbiosis, promoted long-term fear extinction, alleviated hippocampal synapse loss induced by fear conditioning, and limited microglia activation |
[107] |
| Study in mice examined the role of microbiota in fear extinction |
Researchers found that antibiotic treatment of adult mice resulting in microbiome dysbiosis produced impaired fear extinction. They also found that extinction learning and learning-related plasticity require microbiota-derived signals. Microbial deficits in early life produced deficits in fear extinction learning in adulthood. | [108] |
| Study of maternal separation stress in rats and protective effects of probiotics |
Researchers found that maternally-separated male rat pups experienced inappropriate, accelerated development of fear circuitry/behavior and that probiotics protected against unbalanced fear. | [109] |
| Study of the effects of the environmental/soil bacterium, Mycobacterium vaccae, on fear extinction in adult rats |
Researchers found that injection with M. vaccae preparations produced long-lasting enhancement of the rate of within-session fear extinction. | [110] |
| Review of the relationship between gut microbiota, fear extinction, and mental illness with an emphasis on sex-based differences in microbiota. |
Researchers stressed that female stress-related, mental illness is significantly more prevalent than that in men. They emphasized the importance of sex hormones in the gut microbiota-brain regulation, gut microbiota differences based on sex and the need for female focused studies. | [111] |
| Review of gut microbiota functionality of brain regions | Effects of antibiotics and probiotics on fear extinction is included in this recent review | [84] |
| Review concerning how nature, nurture, and microbiota mitigate stress. |
This review includes information on microbiota and fear extinction within the broader subject | [112] |
| Review of probiotics as anti-anxiety and anti-depression psychobiotics |
This review emphasizes specific beneficial microbial metabolism | [113] |
| Review of the microbiota-gut-brain axis in depression | This review emphasizes pathophysiological mechanisms and processes through which microbiota have an anti-depressant function. | [114] |
| Comprehensive review of gut microbiota in anxiety and depression |
This review considers a variety of strategies for gut rebiosis as an anti-anxiety, anti-depression tool. | [115] |
Table 3.
Examples of Fear Learning-Fear Extinction Problems and Mental Health.
| Title 1 | Title 2 | Title 3 |
|---|---|---|
| Obsessive-Compulsive Disorder | Systematic review revealing importance of defective fear extinction | [122] |
| Posttraumatic Stress Disorder | Review of fear extinction as a predictor of PTSD | [123] |
| Disruptive Behavior Disorder | Fear extinction deficits reported to be involved in late adolescent endotype. | [124] |
| Alcohol Dependency | Fear conditioning and extinction with a focus on the role of amygdalain FCE-involved addiction | [125] |
| Attention Deficit Hyperactivity Disorder | Abnormal circuits for fear extinction detected in adults with ADHD | [126] |
| Anxiety Disorders | Focus on neurobiology of fear extinction for treatment of anxiety disorders | [127] |
Table 4.
Role of the Microbiome in Human Senses1.
| Sense | Findings | Reference(s) |
|---|---|---|
| Taste | Review including the role of the microbiome in regulation of taste | [139] |
| Taste | Recent review of the role of oral microbiota in taste perception | [142] |
| Taste | A review detailing specific bacteria and their metabolism as it influences taste perception. The review also covered gut microbiome dysbiosis and linked pathologies. | [143] |
| Taste | Review of the tongue including the role of the tongue microbiome in taste perception. | [144] |
| Smell | Review including the role of the microbiome in regulation of smell | [139] |
| Smell | Recent review describing the pivotal role of nasal microbiota in olfactory development, function, and dysfunctions. | [145] |
| Taste and Smell | Human study reported that obese population displayed reductions in odor and taste preferences some of which were related to oral microbiota difference. | [146] |
| Hearing |
Reviews provide evidence supporting a link between gut microbiome dysbiosis, inner ear inflammation, and sensorineural hearing loss. | [147,148] |
| Hearing | Study of the ear canal microbiota from healthy individuals vs. those with chronic otitis externa | [149] |
| Hearing | Study of 70 otitis media effusion children and two control groups revealed Streptococcus salivarius as a commensal with effective colonization resistance capacity against several key pathobionts. | [150] |
| Neuropathic Pain (including |
Review of microbiome regulation of neuropathic pain | [151] |
| Somatosensory function) | ||
| Skin Sensitivity Syndrome |
Study examining the skin microbiome/mycobiome of 23 patients with sensitivity syndrome compared against control groups. | [152] |
| Skin (sensitivity) | Review covering most aspects of the skin microbiome including both intrinsic and extrinsic factors. | [153] |
| Skin (sensitivity) | Review of skin microbiome differences among healthy vs. diseased/sensitive skin | [154] |
| Sight | Review of dysbiotic gut microbiome-driven eye diseases (age-related macular degeneration, retinal artery occlusion, central serous chorioretinopathy and uveitis) via microbial metabolites and the immune system |
[155] |
| Sight | Introduction to the recently discovered ocular surface microbiome. | [156] |
| Sight | Review of the role of the ocular microbiome in eye disease | [157] |
| Sight | Study of the characterization of healthy eye microbiomes | [158] |
| Sight | Study describes the role of specific ocular surface microbiota in contributing to dry eye disease in diabetic patients. | [159] |
| Sight | Review of intraocular microbiota | [160] |
1 Touch is one of the least directly studied senses as per regulation by microbiota. The references listed in this table on neuropathic pain and skin sensitivity intersect with the issue of somatosensory function.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



