
Submitted:
30 June 2023
Posted:
06 July 2023
You are already at the latest version
Abstract
In Brazil, sucrose-rich broths (cane juice and/or molasses) are used to produce billions of liters of both fuel ethanol and cachaça per year using selected Saccharomyces cerevisiae industrial strains. Aiming to improve this disaccharide fermentation, we have first determined the genetic characteristics, by array comparative genomic hybridization, of a group of 8 fuel ethanol and 5 cachaça industrial yeasts that tend to dominate the fermentors during the production season. The widespread presence of SUC genes encoding invertase at multiple telomeres has been shown to be a common feature of both baker’s and distillers’ yeast strains and is postulated to be an adaptation to sucrose-rich broths. Our results show that only two strains (one fuel ethanol and one cachaça yeast) have amplification of the SUC genes, with high invertase activity. The other industrial yeast strains had a single SUC locus (SUC2) in their genome, with different patterns of invertase activity. These results indicate that invertase activity probably does not limit sucrose fermentation during fuel ethanol and cachaça production by these industrial strains. Using this knowledge, we changed the mode of sucrose metabolism of an industrial strain by avoiding extracellular invertase activity, overexpressing the intracellular invertase, and increasing its transport through the AGT1 permease. This approach allowed higher ethanol production from sucrose by the modified industrial yeast when compared to its parental strain.

Keywords:
bioethanol
; cachaça
; sucrose
; invertase
; thiamin
; Saccharomyces cerevisiae
1. Introduction
Sugarcane is a predominant crop in tropical countries and is used as raw material for several fermentation processes employing Saccharomyces cerevisiae yeasts. Bioethanol, mainly used as an automotive fuel, is by far the most common renewable fuel, and it is thus poised to contribute greatly toward goals of energy independence and environmental sustainability [1]. Fuel ethanol production from sugarcane represents the major large-scale technology capable of producing bioethanol efficiently and economically. Indeed, Brazil is one of the most competitive producers of bioethanol in the world, having the lowest cost of production worldwide and accounting for approximately ~30% of the global production, with almost 27 billion liters produced in 2022 from sugarcane [2,3,4]. Sugarcane juice and molasses can also be used for the production of baker’s yeast and several distilled alcoholic beverages. For example, in Brazil, the sugarcane juice is used to produce cachaça, a distilled spirit that nowadays is the third most popular distilled beverage in the world, with an annual production reaching almost 2 billion liters, performed by industrial and thousands of artisanal producers [5].
The Brazilian industrial process of bioethanol production is characterized by fermentations in very large tanks (0.5 to 3 million liters) using very high yeast cell densities (10-15% w/v) in a fed-batch mode (the so-called Melle–Boinot process) to ferment sugarcane juice and/or diluted molasses that contain up to 150-200 g/L of total sugar [6,7]. The yeast cells are collected by centrifugation after each fermentation cycle of 8-10 h, treated with diluted sulphuric acid for 1-2 h, and more than 90% of the yeasts are reused from one cycle to the next. This ensures the high cell density that contributes to very short fermentation times and thus high productivity of the process. Cachaça production also involves the fermentation of sugarcane juice containing 140-160 g/L of total sugar with a starter culture from a previous fermentation that normally represents 20% of the total volume of the fermentation vat [8]. Both bioethanol and cachaça production are non-sterile processes, and thus subject not only to contamination by microorganisms, but also to a continuous succession of yeast strains [9,10,11,12], some of which can dominate the fermenters and contribute to efficient and stable fermentations [12,13,14].
The disaccharide sucrose (α-d-glucopyranosyl-(1→2)-β-d-fructofuranoside) is by far one of the cheapest sugars in the industrial utilization of the yeast S. cerevisiae [15]. It is well known that yeast cells harbor an extracellular invertase (β-d-fructosidase) that hydrolyzes sucrose into glucose and fructose that are transported by facilitated diffusion into the cells by hexose transporters and metabolized through glycolysis. Invertase is encoded by one or several telomeric SUC genes (SUC1 to SUC5 and SUC7 to SUC10), SUC2 being the most common locus found in the sub-telomeric region of chromosome IX of almost all S. cerevisiae strains, including other closely related yeast species, and thus it is considered the ancestral locus of this gene family [16,17]. Since the amplification of telomeric SUC genes is only observed in industrial (e.g., bakers’, brewers’ and distillers’ yeasts) strains, it was suggested that this trait was under selection during the process of domestication of these yeasts [18,19]. These SUC genes generate two different mRNAs: a larger transcript encoding an invertase with a signal sequence required for its secretion from the cell, and a shorter transcript lacking this signal sequence and thus coding for an intracellular form of the enzyme [20]. While the former mRNA is repressed by high concentrations of sucrose or its hydrolysis products (glucose and fructose), the intracellular invertase is expressed constitutively. Finally, efficient extracellular invertase expression requires low levels of glucose or fructose in the medium [21]. The transcriptional activator of this gene is still unknown despite significant improvements in our knowledge regarding the molecular mechanisms involved in the repression of SUC expression [22].
Although high invertase activities would be expected to ensure efficient fermentation of sucrose, several reports have shown poor (or even negative) correlations between the levels of this enzyme at the yeast surface and the fermentative performance of the cells [23,24]. Indeed, it has been shown [25,26] that the vast majority (>90%) of the glucose and fructose molecules resulting from sucrose hydrolysis diffuse away into the medium before they can be imported into the yeast cell, which may allow the growth of other contaminant microorganisms, including those lacking invertase or who are unable to ferment this disaccharide. However, it is important to note that sucrose is both a β-fructoside and a α-glucoside, and thus it can also be metabolized through the active transporters and enzymes responsible for maltose (and other α-glucosides) fermentation in yeasts [27,28,29,30,31]. This alternative pathway is interesting from a biotechnological point of view due to the energy required for the H+-sucrose symport activity, which consumes 1 ATP molecule per sucrose transported. This results in a lower biomass that can be compensated by enhanced consumption of the sugar, leading to higher ethanol yields when compared with strains hydrolyzing sucrose outside the cells [32,33,34].
In the present report we analyzed the phenotypic and genetic characteristics of a group of 8 fuel ethanol and 5 cachaça industrial yeasts to develop a strategy to genetically modify an industrial fuel ethanol strain for improved ethanol yield from sucrose. Our array comparative genomic hybridization (array-CGH) results show that only two strains (one fuel ethanol and one cachaça yeast) have amplification of the SUC genes, reflected by a high invertase activity. The other industrial yeast strains had a single SUC locus (SUC2) in their genome, although with different patterns of invertase activity. Based on these results we chose one strain and developed an integrative genomic modification approach to avoid extracellular invertase activity, overexpress the intracellular form of invertase, and improve the activity of the AGT1 permease responsible for high-affinity H+-sucrose symport activity [27,29]. The modified industrial yeast strain consumed sucrose directly, without releasing glucose or fructose into the medium, and produced more ethanol when compared to the parental unmodified yeast strain.
2. Materials and Methods
2.1. Strains, Media, and Growth Conditions
The S. cerevisiae strains analyzed in this study are listed in Table 1. Yeasts were grown on rich YP medium containing 20 g/L peptone and 10 g/L yeast extract supplemented with 20 g/L glucose, sucrose, or maltotriose. When required, 20 g/L agar, 0,5 g/L zeocin (Invivogen, San Diego, CA, USA), 3 mg/L Antimycin A, or 200 mg/L geneticin (G-418) sulfate (both from Invitrogen, Thermo Fisher Scientific Inc., Sinapse Biotecnologia, São Paulo, SP, Brazil) were added to the medium. The pH of the medium was adjusted to pH 5.0 with HCl, except when using zeocin where the pH was adjusted to pH 8.0 with NaOH. Cells were grown aerobically at 28°C with shaking (160 rpm) in cotton-plugged Erlenmeyer flasks filled to 20% of the volume with medium, and cellular growth was followed by turbidity measurements at 570 nm (OD570nm).
2.2. Array-CGH Protocol
The array comparative genome hybridization analysis of the industrial yeast strains was performed as described previously [36,39]. We used microarrays onto which had been spotted PCR products corresponding to full-length ORFs from the laboratory S288C strain [40], and thus the reference DNA used in all hybridizations was isolated from this yeast. Genomic DNA was isolated with YeaStar columns (Zymo Research, Irvine, CA, USA), cut with HaeIII (New England Biolabs, Ipswich, MA, USA), and 1 mg of this DNA was labeled with fluorescently tagged nucleotides (Perkin-Elmer, Waltham, MA, USA), usually Cy3-dUTP for the reference strain and Cy5-dUTP for the industrial strains, using the BioPrime random-prime labeling system (Invitrogen). After labeling, the reactions were heat-inactivated, the experimental and reference DNAs were mixed, purified away from unincorporated label using Zymo Clean&Concentrate columns (Zymo Research), and then hybridized to the microarrays at 65°C as described [36,39]. Arrays were scanned with an Axon 4000A scanner and the data were extracted using GenePix (Molecular Devices Corp., Union City, CA, USA) software. The array-CGH data were treated and analyzed as described previously [36,39]. Supplementary Table S1 lists all genetic elements present in the microarrays and their significant (or not) red/green (R/G) ratios, obtained with the CGH-Miner program [41].
2.3. PFGE, Chromosome Blotting and Hybridization
Yeast chromosomes were prepared from 1 ml of yeast cells pre-grown in YP-20 g/L glucose medium and collected at the stationary phase of growth. Cells were treated with Zymolyase 20T and proteinase K in low-melting-point agarose blocks, transferred to a 1% agarose gel, and pulsed-field gel electrophoresis (PFGE) was performed at 8°C using a Gene Navigator system (Amersham Pharmacia Biotech do Brasil Ltda., São Paulo, SP, Brazil) as previously described [29,35,36,39]. Following electrophoresis, the gel was stained with ethidium bromide and photographed (Gel Doc™ XR, BioRad Laboratories, Hercules, CA, USA). The chromosomes separated by PFGE were transferred to a nylon membrane (Hybond-N+, GE Healthcare, Barueri, SP, Brazil) by capillary blotting. Labeling of DNA probes (see below), pre-hybridization, hybridization, stringency washes, and chemiluminescent signal generation and detection were performed with an AlkPhos kit (GE Healthcare) as recommended by the manufacturer. After hybridization, an autoradiography film (Hiperfilm™ ECL, Kodak, GE Healthcare) was exposed to the membrane for 1 to 2 h before it was developed. Images were obtained with Image Lab Software (Gel Doc™ XR) and annotated with Microsoft PowerPoint. Probes were generated by PCR using DNA from strain S288C as template with primers (Table 2) SUC100-F and SUC1320-R for the SUC2 gene.
2.4. Determination of Invertase Activity
Yeast cells were incubated for 12 h in rich YP medium containing 20 g/L sucrose, 20 g/L ethanol plus 30 g/L glycerol, or this last medium supplemented with 1 g/L glucose, and centrifuged (3,000 g, 2 min). The extracellular (periplasmic) invertase activity was determined with whole cells pre-incubated with 50 mM NaF to block glycolysis as described [42], using 100 mM of sucrose. The total invertase activity (extracellular plus intracellular) was determined with permeabilized yeast cells [43] and also 100 mM of sucrose as substrate. The intracellular invertase activity was calculated by subtracting the periplasmic invertase from the total invertase activity. The invertase activity was expressed as nmol of glucose produced (mg dry cell weight-DCW)-1 min-1.
2.5. Determination of the Activity of the AGT1 Permease
The activity of the AGT1 permease was determined with a colorimetric assay using p-nitrophenyl-α-D-glucopyranoside (pNPαG) as substrate [44]. The pNPαG transport activity was expressed as nmol of p-nitrophenol produced (mg DCW)-1 min-1.

Table 2.
Plasmids and primers used in this study.
| Relevant Features or Sequence (5’ → 3’) | |
|---|---|
| Plasmids: | |
| pGRSd-AGT1 | ori ampr CEN6 URA3 PGPD-AGT1-TPGK [45] |
| pUG66 | ori ampr LoxP-BleR-LoxP [46] |
| Primers: 1 | |
| SUC100-F | GCGATAGACCTTTGGTCCAC |
| SUC1320-R | GGACCGTGGTAACTCTAAGG |
| V-SUC2F | GAAATTATCCGGGGGCGAAG |
| V2-SUC2F | GAGTTGTTGTCCTAGCGTAG |
| V2-SUC2R | TCCATTTCCCTCACTACTTC |
| V3-SUC2F | GCATCCACACGTCACAATCT |
| SUC100-R | GTGGACCAAAGGTCTATCGC |
| ssSUC2-F1 | ATGCTTTTGCAAGCTTTCCTTTTCCTTTTGGCTGGTTTTGCAGCCAAAATATCTGCATCAGCCAGCTGAAGCTTCGTACGC |
| 551SUC2-R1 | ATTCGTATTGGTAGCCTAAGAAACCTTCATTGGCAAATGCAGATTCTAGCTTCCAGGACTGCATAGGCCACTAGTGGATC |
| GPD-AGT1F | GCCATAGATTCTACTCGGTCTATCTATCATGTAACACTCCGTTGATGCGTACTAGAGAGTTTATCATTATCAATAC |
| AGT1-389R | GAAAAACTGGCAGGGCATAC |
| V-GPDF | CAACCATCAGTTCATAGGTC |
| AGT1-I505-R | ACGGGCCAGCACTATAGTCTTAGTTCTC |
| V-AGT1F1 | GAATTTTCGGTTGGTG |
2.6. Molecular Biology Techniques
Standard methods for bacterial transformation, DNA manipulation, and analysis were employed [47]. To overexpress the intracellular form of invertase in S. cerevisiae, we had previously transformed a laboratory strain with the kanMX-PADH1 module from plasmid pFA6a-kanMX6-PADH1 [46] that introduced the strong constitutive ADH1 promoter in front of the second codon encoding methionine of the SUC2 gene, after 60 bp of the first start codon, removing the 20 N-terminal amino acids of the resulting protein [32]. The DNA of strain BSY21-34B3 (Table 1) overexpressing the intracellular form of invertase was used to amplify a DNA fragment containing long regions of homology (635 bp and 459 bp, respectively) to the upstream and downstream region of the kanMX6-PADH1::iSUC2 module using primers V2-SUC2F and V2-SUC2R. After transforming [48] cells of the industrial strain CAT-1 with this module, the cells were plated on YP-20 g/L glucose medium containing G-418 and incubated at 28°C. The G-418-resistant isolates were tested for proper genomic integration of the kanMX6-PADH1::iSUC2 cassette at the SUC2 locus by diagnostic colony PCR using primers V3-SUC2F and SUC100-R (Table 2), that amplified a 3,274 bp fragment from the kanMX6-PADH1::iSUC2 locus, and a 774 bp fragment from a normal SUC2 gene (since CAT-1 is diploid, both fragments were obtained). Thus, we next deleted part of the normal copy of the remaining SUC2 gene using primers ssSUC2-F1 and 551SUC2-R1, which allowed the amplification of the BleR gene from plasmid pUG66 (Table 2) having 60 bp of homology to the beginning of the SUC2 gene (absent in the kanMX6-PADH1::iSUC2 locus) and to the position 551 in the coding region of the SUC2 gene, thus truncating almost 1/3 of the normal SUC2 open-reading frame. The resulting PCR product was used to transform competent yeast cells, and zeocin–resistant isolates were tested for proper genomic integration of the BleR gene at the suc2Δ locus by analytical colony PCR using primers V-SUC2F and SUC100R (Table 2) that amplified a ~2,700 bp fragment from the kanMX6-PADH1::iSUC2 locus, but failed to amplify a 550 bp that will be obtained if the strain had a normal SUC2 gene (e.g., strain CAT-1).
Finally, to overexpress the high-affinity sucrose-H+ symporter encoded by the AGT1 gene [27,28,29] we used primers GPD-AGT1F and AGT1-389R (Table 2) to amplify from plasmid pGRSd-AGT1 (Table 2 [45]) the PGPD (PTDH3) promoter region controlling the AGT1 gene present in the plasmid. The linear DNA fragment contained an upstream sequence with 55 bp of homology to the upstream region of the AGT1 gene present in chromosome VII (~279 bp from the start of the gene), the PTDH3 promoter, and 409 bp of the beginning of the AGT1 ORF (and thus acting as a long region of homology for integration). The isolation of transformants was done using YP-20 g/L maltotriose plates containing 3 mg/L Antimycin A as selecting media, since in the presence of Antimycin A strains that do not have a functional AGT1 permease would not grow (e.g. strain CAT-1), since they grow aerobically on this carbon source [39]. The correct integration of the PTDH3 promoter upstream of the AGT1 gene was verified in the selected strains using primers V-GPDF and AGT1-I505R (Table 2) that allowed amplification of a 1,944 bp fragment from strain GMY08 (but not from strain CAT-1), confirming the correct modification of the promoter region of the AGT1 gene. Since the strains are diploids, we used two other primers (V-AGT1F1 and AGT1-I505R, Table 2) to check if the two copies of the AGT1 were modified. The region with homology to primer V-AGT1F1 is removed with the PTDH3-AGT1 module, but since strain GMY08 still had a 1,583 bp fragment amplified with these two primers, it indicated that only one copy of the AGT1 gene was modified (as expected, strain CAT-1 had also this fragment amplified when analyzed by colony PCR).
2.7. Sucrose Batch Fermentations
Yeast strains were grown in rich YP medium containing 20 g/L sucrose, collected at the end of the log phase (<1 g DCW/L), centrifuged (3,000 g, 5 min), washed twice with ice-cold distilled water, and resuspended in ice-cold distilled water in order to obtain a suspension with 20 g DCW/L. A volume of this suspension was mixed with the same volume of 2x YP medium (40 g/L peptone and 20 g/L yeast extract) containing 400 g/L sucrose in order to simulate industrial conditions: high cell densities (10 g DCW/L) and high sucrose concentrations (~200 g/L). The fermentations were carried out in cotton plugged Erlenmeyer flasks filled to 1/5 of the volume, with shaking (160 rpm) and at 28°C. One mL culture samples were harvested regularly, the OD570nm determined, centrifuged (5,000 g, 2 min), and their supernatants stored at -20°C for the determination of sugars and ethanol. The chemical and biochemical methods used to quantify the concentrations of sucrose, glucose, fructose, and ethanol have been described in detail before [49]. For the comparison between strains CAT-1 and GMY08, the concentrations of sucrose, glucose, fructose, ethanol, and glycerol were determined by high-performance liquid chromatography (HPLC) equipped with a refractive index detector (RI-2031 Plus; JASCO, Tokyo, Japan) using an HyperREZ XP Organic Acid Column (Thermo Scientific, Waltham, MA, USA). The HPLC apparatus was operated at 20°C using 5 mM H2SO4 as the mobile phase at a flow rate of 0.2 ml/min.
3. Results
3.1. Microarray Karyotyping of Industrial Yeast Strains Used in Sugarcane-Based Fermentation Processes
We performed aCGH analysis of thirteen industrial yeast strains that are used in sugarcane-based fermentation process in Brazil (Table 1): 8 industrial fuel ethanol strains − six from the State of São Paulo (strains BG-1, CAT-1, PE-2, SA-1, and VR-1, previously analyzed by Stambuk and co-workers [36]) plus strain BAT, and two yeast strains isolated from the State of Paraíba (strains UFPE-135 and UFPE-179) − and five cachaça yeast strains isolated from the State of Minas Gerais (strains UFMG-829, UFMG-905, UFMG-1007, UFMG-2097, and UFMG-2439). Although strain BAT showed some undesired industrial characteristics, such as excessive foam production, flocculation, and premature sedimentation [35], it is a yeast strain that dominates the industrial process. Strain UFMG-905, besides being used as a starter in cachaça production [13,37], is also a yeast strain with interesting probiotic characteristics [50,51].
Visualization of the data with the Java TreeView program [52] revealed that all yeast strains analyzed showed hybridization patterns with signals across most of the chromosomes corresponding to R/G ratios near a value of 1.0, indicating that the genomes of these industrial diploid yeast strains lacked chromosomal aneuploidies (data not shown). Figure 1A shows the array-CGH data for a selection of genes involved in sugar fermentation, including the SUC2 gene encoding invertase and 20 genes involved in sugar transport (HXT1 to HXT17, plus GAL2 and the transceptors SNF3 and RGT2). In addition, Figure 1B shows the data for genes involved in thiamine and pyridoxine biosynthesis (SNO/SNZ genes), which were previously shown to be amplified in the 5 fuel ethanol industrial strains from São Paulo [36], as well as genes involved in the transport of thiamine and its precursors (THI7, NRT1 and THI72 genes).
From the data shown in Figure 1A it is evident that, with the exception of the fuel ethanol strain UFPE-135 and the cachaça strain UFMG-829 that showed amplification of the SUC2 gene, all the other strains seem to have the usual copy number of this gene in their genome. Almost all strains also have regular copy numbers of the major hexose transporters HXT1 to HXT7 [53,54,55]. On the other hand, a lower copy number was detected for the telomeric HXT genes HXT8, HXT9, HXT11, HXT12, and HXT14 to HXT17 for almost all of the industrial strains analyzed. Note that HXT10 is not telomeric, while HXT13 is located at one of the telomeres of chromosome V. The genes HXT13, HXT15, HXT16 and HXT17 lost the status of hexose transporters since they were reported as polyol (xylitol, mannitol and sorbitol) transporters [56]. Although all strains seem to have a lower copy number of the galactose-inducible GAL2 permease, growth on this carbon source by many of the strains was not impaired by the presence of the mitochondrial inhibitor antimycin A (data not shown), and thus the strains still have a functional GAL2 transporter in their genomes [57]. Many of the telomeric HXT transporters absent in the genome of the industrial strains are probably the consequence of their replacement by the amplified SNO/SNZ genes (also telomeric) involved in pyridoxine and thiamin biosynthesis (Figure 1B) [36,58]. In strain S288C, these genes are located in the left telomere of chromosome VI (SNO3/SNZ3) and chromosome XIV (SNO2/SNZ2). The industrial sugarcane strains, which lack the genes in chromosome VI, have these genes spread in the telomeres of chromosomes IV, VII, IX, X, XI and/or XIV [36,59]. The amplified SNO/SNZ genes present in chromosome IV are replacing the telomeric region containing the HXT15 gene. In the case of chromosome X, there is the particular situation in some industrial strains in which both telomeres might contain the SNO/SNZ genes, in the right telomere (replacing the region containing the HXT16 gene) and also in the left telomere (replacing the region containing the HXT8 and/or HXT9 genes) [59,60]. Figure 1 also shows that, in some industrial strains (e.g., the cachaça yeasts), the SNO/SNZ genes (particularly the SNZ2/SNZ3 genes) were not amplified as much as in the case of the fuel-ethanol strains. Interestingly, these strains do show amplification of transporters involved in the uptake of thiamin (THI7 gene) and its precursors 2-methyl-4-amino-5-hydroxymethylpyrimidine (derived from vitamin B6) and/or 4-methyl-5-β-hydroxyethylthiazole, the NRT1 (also known as THI71) and THI72 genes [61,62]. These results highlight the importance that thiamin (and also pyridoxine) has for yeast strains used in sugarcane-based fermentation processes, as well as other industrial fermentations that normally use high sugar concentrations [35,63].
3.2. Invertase Activity of Industrial Yeast Strains Used in Sugarcane-Based Fermentation Processes
The aCGH analysis of the thirteen industrial yeast strains indicated that only two strains (strain UFPE-135 and UFMG-829) showed amplification of the SUC2 gene encoding invertase. Figure 2 shows a chromosomal blot of the SUC loci found in the laboratory S288C strain (SUC2), as well as in some of the fuel-ethanol yeast strains, including strain UFPE-135 that has a higher copy-number of SUC genes in its genome. It is evident that the majority of sugarcane yeast strains harbors only the SUC2 gene in their genomes (strains CAT-1, PE-2, BG-1, and SA-1), while strain UFPE-135 has, besides this gene, also the SUC1 gene located in chromosome VII. The identity of the VII chromosome copy was confirmed by hybridization with a probe based on the AGT1 gene (data not shown). Other strains (e.g., UFMG-1007, UFPE-179, and VR-1) also presented only the SUC2 loci in their genomes (see Supplementary Figure S1). When the invertase activity was determined in these yeast strains, many of them (the fuel ethanol strains CAT-1, PE-2, VR-1, and UFPE-179, and the cachaça yeast strain UFMG-1007) had a similar profile to the laboratory strain S288C: higher activity on sucrose compared to the derepressed conditions (growth with ethanol/glycerol), and even higher activity after growth on the derepressed conditions supplemented with 0,1% glucose, one of the best conditions for induction of invertase expression [64]. Some strains showed a slightly different pattern, with a high invertase activity after derepression conditions (strains BG-1 and SA-1), and as expected, the strains that had amplification of the SUC genes (strains UFMG-829 and UFPE-135) were the ones with the highest invertase activity under all conditions tested (Figure 3).
3.3. Sucrose Batch Fermentation by the Industrial Yeast Strains Used in Sugarcane-Based Fermentation Processes
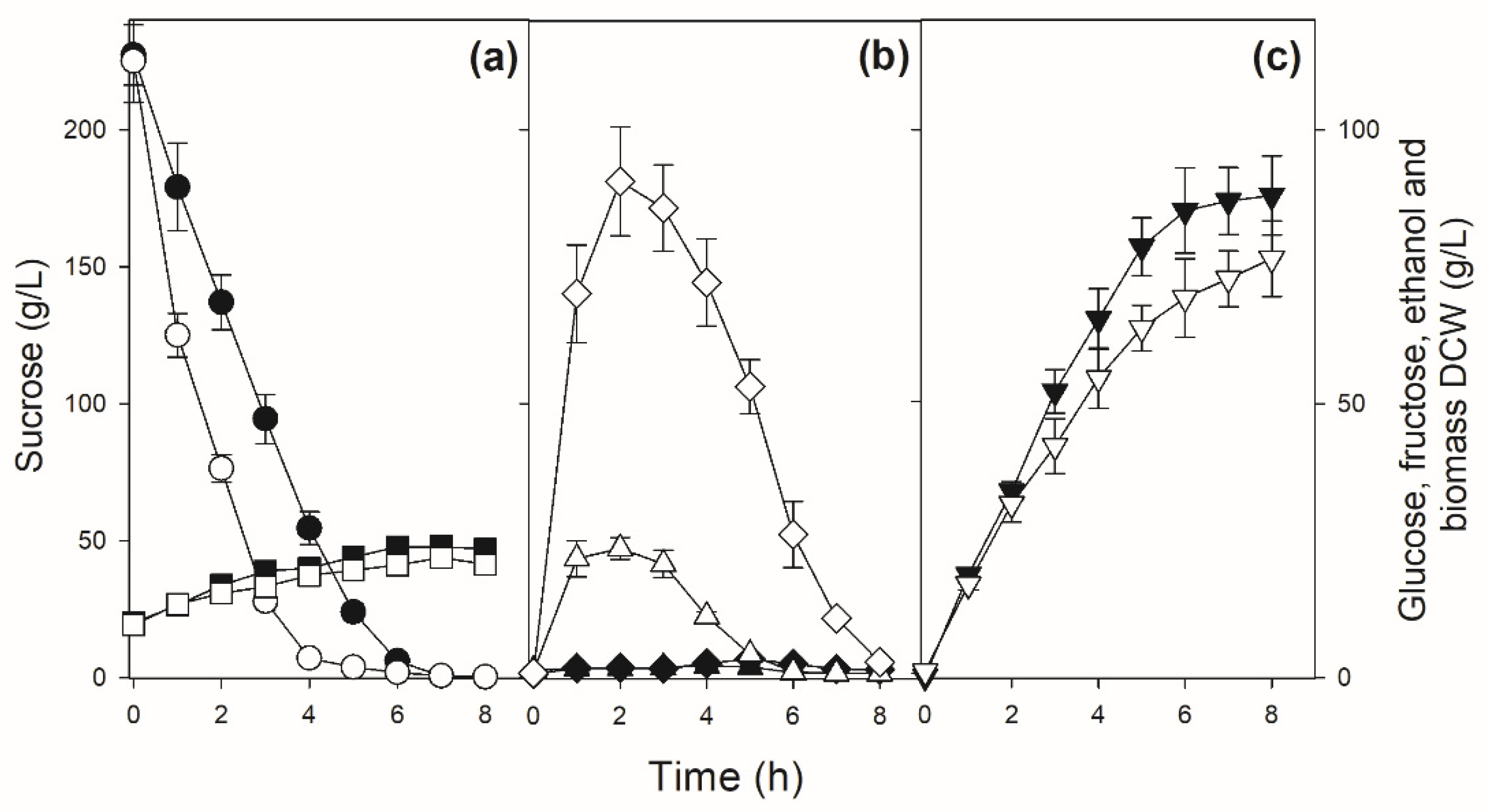
We performed sucrose batch fermentations trying to simulate industrial conditions: initial high (>200 g/L) sucrose concentrations to be fermented by high (10 g/L DCW) cellular concentrations. Figure 4 shows the results for some strains (PE-2, BG-1, UFPE-179 and UFMG-1007) that have a typical pattern of sucrose consumption: sucrose is hydrolyzed rapidly and totally consumed in ~6 h (strains PE-2, UFPE-179 and UFMG-1007), releasing significant amounts of glucose and fructose into the medium that will be consumed by ~8 h of fermentation to produce 68-78 g/L ethanol. Note that due to higher invertase activity, strain BG-1 hydrolyzes almost all sucrose in ~4 h, releasing higher amounts of glucose and especially fructose into the medium. In general, fructose accumulates to higher concentrations and takes longer to be consumed by the yeast cells, when compared to the glucose released into the medium (Figure 4). In the case of the yeast strains UFMG-829 and UFPE-135 with amplification of the SUC genes (Figure 1), with high invertase activity (Figure 3), the hydrolysis of sucrose is so fast that during the initial mixing and sampling of the fermentation broth (which included a 2 min centrifugation) a significant release of glucose, fructose and ethanol occurs, and sucrose is totally hydrolyzed in less than 4 h (Figure 5), producing 74-77 g/L of ethanol.
Figure 4.
Sucrose batch fermentation by selected sugarcane industrial yeast strains. Panel (a) shows the concentrations of sucrose (circles) and biomass (squares), panel (b) shows the concentrations of glucose (triangles) and fructose (diamonds), while panel (c) shows the concentrations of ethanol (inverted triangles) during the fermentation by strain PE-2 (white symbols), UFPE-179 (light gray symbols), BG-1 (gray symbols) and UFMG-1007 (black symbols).
Figure 4.
Sucrose batch fermentation by selected sugarcane industrial yeast strains. Panel (a) shows the concentrations of sucrose (circles) and biomass (squares), panel (b) shows the concentrations of glucose (triangles) and fructose (diamonds), while panel (c) shows the concentrations of ethanol (inverted triangles) during the fermentation by strain PE-2 (white symbols), UFPE-179 (light gray symbols), BG-1 (gray symbols) and UFMG-1007 (black symbols).
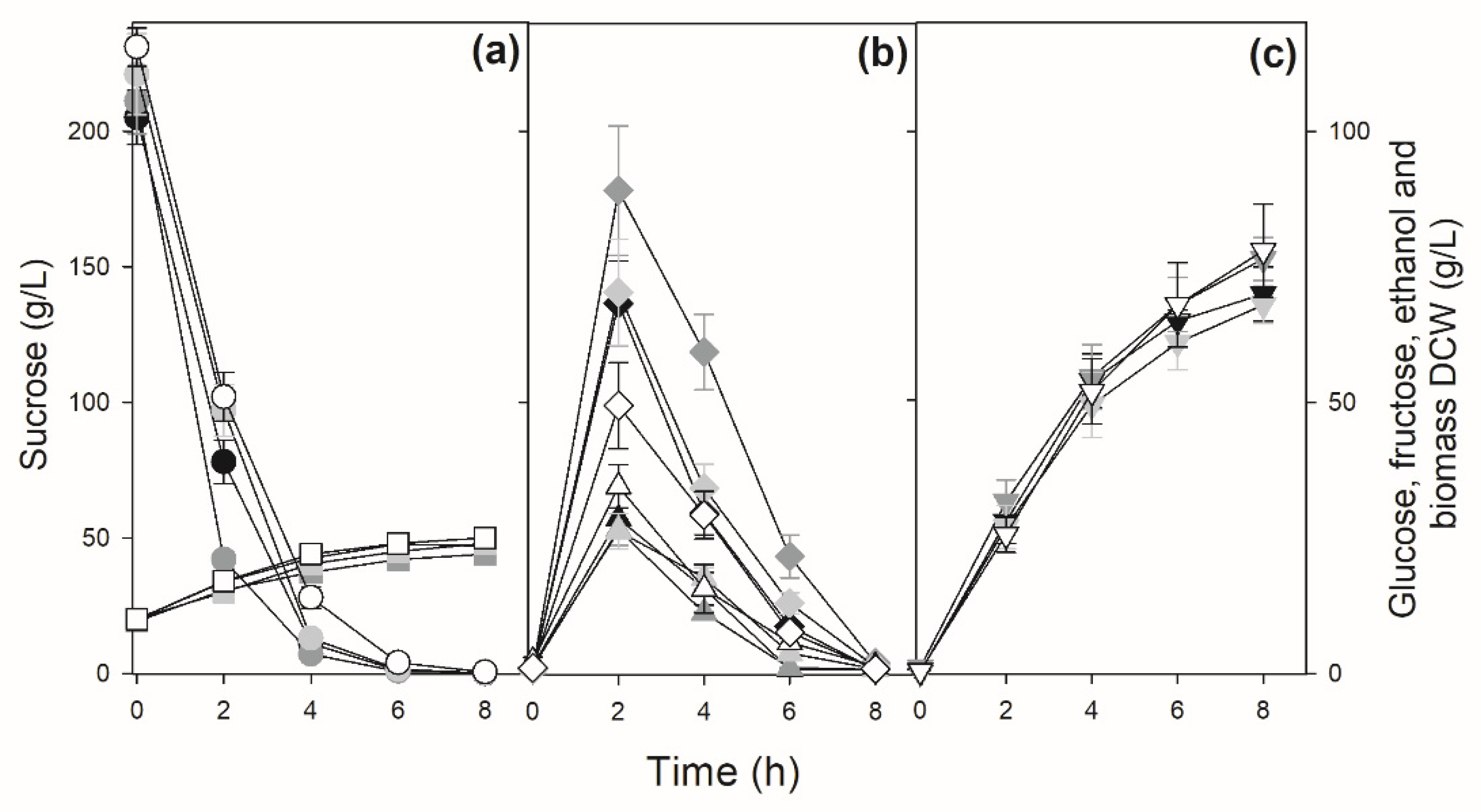
Figure 5.
Sucrose batch fermentation by sugarcane industrial yeast strains with amplification of the SUC genes. Panel (a) shows the concentrations of sucrose (circles) and biomass (squares), panel (b) shows the concentrations of glucose (triangles) and fructose (diamonds), while panel (c) shows the concentrations of ethanol (inverted triangles) during the fermentation by strain UFMG-829 (white symbols), and UFPE-135 (black symbols).
Figure 5.
Sucrose batch fermentation by sugarcane industrial yeast strains with amplification of the SUC genes. Panel (a) shows the concentrations of sucrose (circles) and biomass (squares), panel (b) shows the concentrations of glucose (triangles) and fructose (diamonds), while panel (c) shows the concentrations of ethanol (inverted triangles) during the fermentation by strain UFMG-829 (white symbols), and UFPE-135 (black symbols).
3.4. Modifying the Mode of Sucrose Fermentation by an Industrial Fuel-Ethanol Yeast Strain
The results shown in the previous sections indicate that all the industrial strains analyzed contain common genomic features, many of them probably contributing to their dominance in the sugarcane-based industrial processes. Regarding sucrose fermentation, the majority of strains have a single SUC locus (SUC2 gene) in their genome, which is an important characteristic if we wish to modify the way that the yeast cells will ferment sucrose: instead of its extracellular hydrolysis, the disaccharide will be actively transported into the cell and hydrolyzed intracellularly [32]. To achieve these goals, initially it was necessary to modify the genes encoding invertase; given that the industrial strains are diploid, that requires modifying 2 copies, even in those strains having a single SUC2 locus. Thus, we decided to perform the genetic engineering strategies with strain CAT-1, which has previously been very well characterized in many aspects, including its genome sequence [64]. First, one copy of the SUC2 gene had its promoter region modified in order to overexpress the intracellular form of invertase (kanMX6-PADH1::iSUC2), placing the constitutive PADH1 promoter controlling the expression of an invertase lacking the 20 first amino acids, the signal peptide that drives the protein to the endoplasmic reticulum and Golgi for its secretion [20]. The other copy of the SUC2 gene was partially deleted (suc2Δ::BleR), thus avoiding the expression of the extracellular form of invertase. Table 3 shows that strain CAT-1 had an extracellular invertase activity ~4 times higher than the intracellular invertase activity. On the other hand, the opposite trend was observed for the modified strain GMY08 with a large increase in intracellular invertase activity and a significant decrease in the extracellular invertase activity. Note that the residual “extracellular invertase” activity of strain GMY08 is probably the consequence of sucrose entrance into the cells, and its intracellular hydrolysis, releasing glucose molecules that cannot be metabolized by the yeast cells due to blockage of glycolysis by sodium fluoride [42].
It was evident from our previous work engineering on the mode of sucrose fermentation in laboratory yeast strains that sucrose transport by the AGT1 permease was a limiting factor [32]. Only after a laboratory evolution approach in anaerobic sucrose-limited chemostats it was possible to isolate an evolved strain with higher transport activity, due to a duplication of the AGT1 gene, that showed the expected improved sucrose fermentation performance [32]. Strain CAT-1 has the AGT1 gene in chromosome VII [65], but this strain does not ferment efficiently maltotriose (another substrate of the AGT1 permease) [39] due to the presence of a divergent and non-functional promoter region, also found in other industrial yeast strains unable to ferment maltotriose [66]. Instead, strain CAT-1 uses maltotriose only after an extensive lag phase, and it metabolizes this sugar aerobically due to its extracellular hydrolysis mediated by the isomaltase encoded by the IMA5 gene [39,67]. Indeed, as can be seen in Table 3, strain CAT-1 had no pNPαG transport activity, a measurement of this permease´s activity, indicating that it was also necessary to overexpress the AGT1 gene. For this purpose, the promoter region of the AGT1 gene present in CAT-1 was replaced by the constitutive PTDH3 promoter using the same strategy of Vidgren and co-workers [68], and the transformants were selected in maltotriose plates containing Antimycin A. Although only one copy of the AGT1 gene had its promoter modified in strain GMY08 (PTDH3::AGT1), it clearly had an increased activity of this transporter (Table 3).
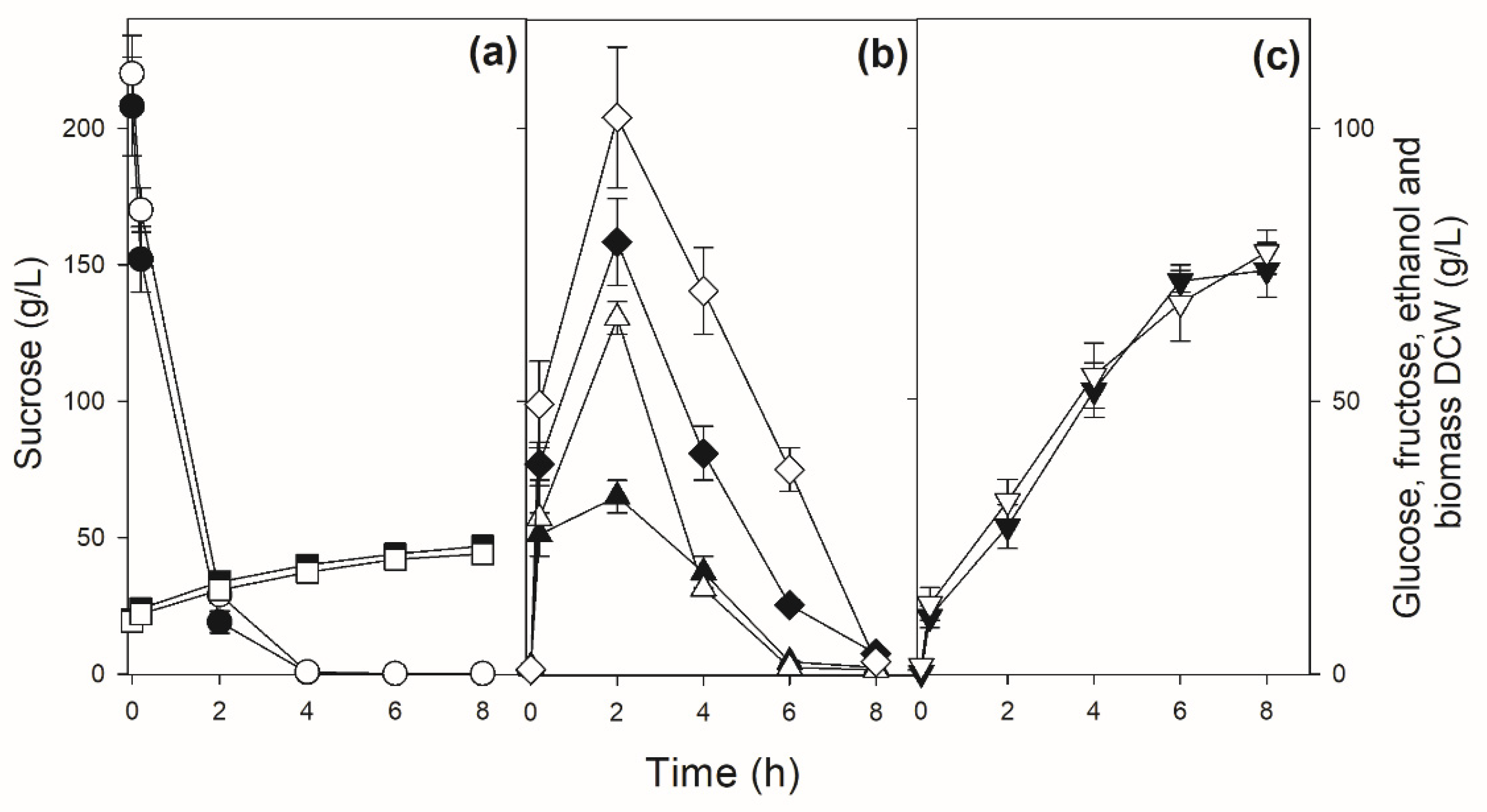
The physiological performance of the GMY08 strain was evaluated in sucrose batch fermentation and compared to the parental strain (Figure 6). Strain CAT-1 showed a fermentation performance similar to other industrial sugarcane strains (see Figure 4), hydrolyzing sucrose in up to 5-6 h and releasing huge amounts of hexoses that took up to 8 h to be completely consumed. The modified strain GMY08 showed a slower sucrose consumption for 6-7 h, without releasing hexoses in the medium (Figure 6). Because of the energy demand of the active sucrose-H+ symport [32], it produced approximately ~11% more ethanol than the parental CAT-1 strain. In addition, the amount of glycerol produced by strain GMY08 during sucrose batch fermentation was half of that produced by the parental strain CAT-1 (Table 3).
Figure 5.
Sucrose batch fermentation by strains CAT-1 and GMY08. Panel (a) shows the concentrations of sucrose (circles) and biomass (squares), panel (b) shows the concentrations of glucose (triangles) and fructose (diamonds), while panel (c) shows the concentrations of ethanol (inverted triangles) during the fermentation by strain CAT-1 (white symbols) or GMY08 (black symbols).
Figure 5.
Sucrose batch fermentation by strains CAT-1 and GMY08. Panel (a) shows the concentrations of sucrose (circles) and biomass (squares), panel (b) shows the concentrations of glucose (triangles) and fructose (diamonds), while panel (c) shows the concentrations of ethanol (inverted triangles) during the fermentation by strain CAT-1 (white symbols) or GMY08 (black symbols).
4. Discussion
Analysis of genomic variation among a large number of S. cerevisiae strains has revealed not only significant genetic variability but has also shown that, in general, these strains cluster according to their technological application rather than geographical origin [69,70,71], indicating that wild and domesticated populations have evolved different life strategies for adaptation to generally different environments. Although there are different driving forces that shape the genome structure and gene content of domesticated and wild yeasts, variations in gene copy number have a greater phenotypic effect than do single nucleotide polymorphisms or other genomic changes [71,72]. In the case of the Brazilian cachaça yeast strains, their genomic signatures indicate that they derive from wine yeasts that have undergone an additional round of domestication, or “secondary domestication” [73], while the industrial fuel ethanol yeast strains are also closely related to cachaça strains forming a single monophyletic clade proximal to wine strains [59,71,74]. Our array-CGH results indeed confirm the close proximity of the cachaça and fuel ethanol strains, sharing, for example, common loss of some telomeric HXT genes, while genes involved in thiamine (B1) and pyridoxine (B6) biosynthesis and uptake from the media are amplified in these sugarcane fermenting yeasts.
The aCGH analysis of the thirteen industrial yeast strains indicated that only two strains (one cachaça and one fuel ethanol) showed amplification of the SUC2 gene encoding invertase. This characteristic was very promising for modifying of the mode of sucrose fermentation by an industrial strain aiming its direct transport into the cell and its intracellular hydrolysis. If the industrial strain has several SUC loci, several genes encoding invertase needed to be modified, but with strain CAT-1 only three genomic engineering strategies were performed: overexpression of the intracellular invertase (kanMX6-PADH1::iSUC2), inactivation of the other SUC2 gene to prevent the expression of the extracellular invertase (suc2Δ::BleR), and overexpression of the sucrose-H+ symporter encoded by the AGT1 permease (PTDH3::AGT1). It is important to note that overexpression of the AGT1 permease is certainly a required modification in the fuel ethanol yeasts, since this gene is normally regulated by maltose or maltotriose through the MAL regulatory system [75], and not by the presence of sucrose. Only if the strain is MAL constitutive, as some laboratory strains used previously (strains CEN.PK2-1C or 1403-7A, see [29,32]), will allow the expression of the AGT1 permease during growth on sucrose. When strain CAT-1 was modified with these three genomic engineering approaches, the resulting strain GMY08 fermented sucrose directly through its active transport and intracellular hydrolysis. Besides the expected higher ethanol production as a consequence of the energetics of sucrose transport [32], strain GMY08 did not released in the medium any glucose or fructose, sugars that can favor microbial contamination even by microorganisms that would not normally use sucrose as a carbon source [25,26,76]. Another consequence of the hydrolysis of high sucrose concentrations by invertase is a sudden osmotic shock promoted by the high glucose plus fructose concentration in the medium. Yeast cells respond to the osmotic stress increasing the production of glycerol from the glycolytic intermediate dihydroxyacetone phosphate, which will lead to a decrease in the ethanol yield [77,78,79,80]. Indeed, strain GMY08 produced half the amount of the glycerol produced by strain CAT-1 during batch sucrose fermentation. While the engineered industrial strain developed in this study showed good fermentation performance on sucrose when compared to the wild-type strain, it would be important to evaluate its fermentation performance under industrial process conditions, including its possible effect on the microbial dynamics on the fermentation tanks.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Table S1: CGH-Miner data of 13 yeast strains showing red to green ratios of genetic elements; and Figure S1: Detection of SUC genes in additional industrial sugarcane yeasts.
Author Contributions
Conceptualization, G.M. and B.U.S.; investigation, methodology and formal analysis, G.M., V.R.G., M.G.D., E.H.D., S.L.A.J. and A.B.; resources, B.D., G.S. and C.A.R.; writing—review and editing, B.D., G.S., C.A.R. and B.U.S.; project administration and funding acquisition, B.U.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by grants and fellowships from the Brazilian agencies Coordination for the Improvement of Higher Education Personnel (CAPES), National Council for Scientific and Technological Development (CNPq, process 507123/2004-2, 552877/2007-7, 551392/2010-0, and 308389/2019-0), Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG, process CBB-APQ-03071-17), Financier of Studies and Projects (FINEP, process 01.07.0761.00/1372-07), and by Grupo Cerradinho Açúcar Álcool e Energia S/A (São Paulo, Brazil). This work is part of the project “INCT Yeasts: Biodiversity, preservation and biotechnological innovation”, supported by fellowships from CNPq (process 406564/2022-1). This work was also supported in part by a NSF ADVANCE grant DBI-0340856 (B.D.) and NIH R01 HG003328 (G.S.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable. All data is contained within the article and supplementary material.
Acknowledgments
Donation of the HPLC system by the Japanese International Cooperation Agency (JICA, Japan) is acknowledged. We also thank M. Longtine (University of North Carolina, USA) and J. François (Centre de Bioingénierie Gilbert Durand, France) for providing plasmids.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Goldemberg J. Ethanol for a sustainable energy future. Science 2007, 315, 808-810. [CrossRef]
- CONAB – Companhia Nacional de Abastecimento - Acompanhamento da safra brasileira de cana-de-açúcar - Safra 2022/23. Available online: https://www.conab.gov.br/info-agro/safras/cana/boletim-da-safra-de-cana-de-acucar (accessed on 02 March 2023).
- Jaiswal, D.; de Souza, A.P.; Larsen, S.; LeBauer, D.S.; Miguez, F.E.; Sparovek, G.; Bollero, G.; Buckeridge, M.S.; Long, S.P. Brazilian sugarcane ethanol as an expandable green alternative to crude oil use. Nat. Clim. Change 2017, 7, 788-792. [CrossRef]
- Jacobus, A.P.; Gross, J.; Evans, J.H.; Ceccato-Antonini, S.R.; Gombert, A.K. Saccharomyces cerevisiae strains used industrially for bioethanol production. Essays Biochem. 2021, 65, 147-161. [CrossRef]
- do Nascimento e Silva, J.H.; Verruma-Bernardi, M.R.; de Oliveira, A.L Cachaça production in Brazil and its main contaminant (ethyl carbamate). Sci. Agric. 2018, 77, e20180135. [CrossRef]
- Amorim, H.V.; Lopes, M.L.; Oliveira, J.V.C.; Buckeridge, M.S.; Goldman, G.H. Scientific challenges of bioethanol production in Brazil. Appl. Microbiol. Biotechnol. 2011, 91, 1267-1275. [CrossRef]
- Lopes, M.L.; Paulillo, S.C.; Godoy, A.; Cherubin, R.A., Lorenzi, M.S., Giometti, F.H., Bernardino, C.D., Amorim Neto, H.B., Amorim, H.V. Ethanol production in Brazil: a bridge between science and industry. Braz. J. Microbiol. 2016, 47(Suppl 1), 64-76. [CrossRef]
- Badotti, F.; Gomes, F.C.O.; Rosa, C.A. Brazilian cachaça: fermentation and production. In: Handbook of Plant-Based Fermented Food and Beverage Technology, Second ed.; Hui, Y.H.; Evranuz, E.Ö. Eds; CRC Press, Florida, USA; 2012; pp 639–648.
- Guerra, J.B.; Araújo, R.A.; Pataro, C.; Franco, G.R.; Moreira, E.S.; Mendonça-Hagler, L.C.; Rosa, C.A. Genetic diversity of Saccharomyces cerevisiae strains during the 24 h fermentative cycle for the production of the artisanal Brazilian cachaça. Lett. Appl. Microbiol. 2001, 33, 106-111. [CrossRef]
- da Silva-Filho, E.; Santos, S.K.; Resende, A.M.; Morais, J.O.; de Morais-Jr, M.A.; Simões, D.A. Yeast population dynamics of industrial fuel-ethanol fermentation process assessed by PCR-fingerprinting. Antonie van Leeuwenhoeck 2005, 88, 13-23. [CrossRef]
- Badotti, F.; Rosa, C.A.; Stambuk, B.U. Biochemical and molecular characterization of Saccharomyces cerevisiae strains isolated from artisan-type fermentations of sugarcane and molasses in the production of sugarcane brandy in Florianópolis-SC, Brazil. Braz. J. Food Technol. 2010, 13, 205-213. [CrossRef]
- Basso, L.C.; de Amorim, H.V.; de Oliveira, A.J.; Lopes, M.L. Yeast selection for fuel ethanol production in Brazil. FEMS Yeast Res. 2008, 8, 1155-1163. [CrossRef]
- Gomes, F.C.; Silva, C.L.; Marini, M.M.; Oliveira, E.S.; Rosa, C.A. Use of selected indigenous Saccharomyces cerevisiae strains for the production of the traditional cachaça in Brazil. J. Appl. Microbiol. 2007, 103, 2438-2447. [CrossRef]
- Campos, C.R.; Silva, C.F.; Dias, D.R.; Basso, L.C.; Amorim, H.V.; Schwan, R.F. Features of Saccharomyces cerevisiae as a culture starter for the production of the distilled sugar cane beverage, cachaça in Brazil. J. Appl. Microbiol. 2010, 108, 1871-1879. [CrossRef]
- Marques, W.L.; Raghavendran, V.; Stambuk, B.U.; Gombert, A.K. Sucrose and Saccharomyces cerevisiae: A relationship most sweet. FEMS Yeast Res. 2016, 16, fov107. [CrossRef]
- Korshunova, I.V.; Naumova, E.S.; Naumov, G.I. Comparative molecular genetic analysis of β-fructosidases of yeasts Saccharomyces. Mol. Biol. 2005, 39, 413-419. [CrossRef]
- Naumova, E.S.; Sadykova, A.Z.; Martynenko, N.N.; Naumov, G.I. Molecular polymorphism of β-fructosidase SUC genes in the Saccharomyces yeasts. Mol. Biol. 2014, 48, 573–582. [CrossRef]
- Codón, A.C.; Benítez, T.; Korhola, M. Chromosomal polymorphism and adaptation to specific industrial environments of Saccharomyces strains. Appl. Microbiol. Biotechnol. 1998, 49, 154-163. [CrossRef]
- Naumova, E.S.; Sadykova, A.Z.; Martynenko, N.N.; Naumov, G.I. Molecular genetic characteristics of Saccharomyces cerevisiae distillers’ yeasts. Microbiology 2013, 82, 175–185. [CrossRef]
- Carlson, M.; Botstein, D. Two differentially regulated mRNAs with different 5′ ends encode secreted with intracellular forms of yeast invertase. Cell 1982, 28, 145-154. [CrossRef]
- Herwing, C.; Doerries, C.; Marison, I.; von Stockar, U. Quantitative analysis of the regulation scheme of invertase expression in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2001, 76, 247-258. [CrossRef]
- Belinchón, M.M.; Gancedo J.M. Different signaling pathways mediate glucose induction of SUC2, HXT1 and pyruvate decarboxylase in yeast. FEMS Yeast Res. 2007, 7, 40-47. [CrossRef]
- Takeshige, K.; Ouchi, K. Effects of yeast invertase on ethanol production in molasses. J. Ferment. Bioeng. 1995, 79, 513-515. [CrossRef]
- Myers, D.K.; Lawlor, D.T.M.; Attfield, P.V. Influence of invertase activity and glycerol synthesis and retention on fermentation of media with a high sugar concentration by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1997, 63, 145-150. [CrossRef]
- Greig, D.; Travisano, M. The Prisoner’s Dilemma and polymorphism in yeast SUC genes. Proc. Biol. Sci. 2004, 271(Suppl 3), S25-6. [CrossRef]
- Gore, J.; Youk, H.; Oudenaarden, A. Snowdrift game dynamics and facultative cheating in yeast. Nature 2009, 459, 253-256. [CrossRef]
- Stambuk, B.U.; Batista, A.S.; de Araujo, P.S. Kinetics of active sucrose transport in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2000, 89, 212-214. [CrossRef]
- Batista, A.S.; Miletti, L.C.; Stambuk, B.U. Sucrose fermentation by Saccharomyces cerevisiae lacking hexose transport. J. Mol. Microbiol. Biotechnol. 2004, 8, 26-33. [CrossRef]
- Badotti, F.; Dário, M.G.; Alves-Jr, S.L.; Cordioli, M.L.; Miletti, L.C.; de Araujo, P.S.; Stambuk, B.U. Switching the mode of sucrose utilization by Saccharomyces cerevisiae. Microb. Cell. Fact. 2008, 7, 4. [CrossRef]
- Brown, C.A.; Murray, A.W.; Verstrepen, K.J. Rapid expansion and functional divergence of subtelomeric gene families in yeasts. Curr. Biol. 2010, 20, 895-903. [CrossRef]
- Marques, W.L.; Mans, R.; Marella, E.R.; Cordeiro, R.L.; van den Broek, M.; Daran, J.G.; Pronk, J.T.; Gombert, A.K.; van Maris, A.J. Elimination of sucrose transport and hydrolysis in Saccharomyces cerevisiae: a platform strain for engineering sucrose metabolism. FEMS Yeast Res. 2017, 17, fox006. [CrossRef]
- Basso, T.O.; de Kok, S.; Dario, M.; do Espirito-Santo, J.C.; Müller, G.; Schlölg, P.S.; Silva, C.P.; Tonso, A.; Daran, J.M.; Gombert, A.K.; van Maris, A.J.; Pronk, J.T.; Stambuk, B.U. Engineering topology and kinetics of sucrose metabolism in Saccharomyces cerevisiae for improved ethanol yield. Metab. Eng. 2011, 13, 694-703. [CrossRef]
- Gombert, A.K.; van Maris, A.J. Improving conversion yield of fermentable sugars into fuel ethanol in 1st generation yeast-based production processes. Curr. Opin. Biotechnol. 2015, 33, 81-86. [CrossRef]
- van Aalst, A.C.A.; de Valk, S.C.; van Gulik, W.M.; Jansen, M.L.A.; Pronk, J.T.; Mans, R. Pathway engineering strategies for improved product yield in yeast-based industrial ethanol production. Synth. Syst. Biotechnol. 2022, 7, 554-566. [CrossRef]
- de Figueiredo, C.M.; Hock, D.H.; Trichez, D.; Magalhães, M.d.L.B.; Lopes, M.L.; de Amorim, H.V.; Stambuk, B.U. High foam phenotypic diversity and variability in flocculant gene observed for various yeast cell surfaces present as industrial contaminants. Fermentation 2021, 7, 127. [CrossRef]
- Stambuk, B.U.; Dunn, B.; Alves-Jr, S.L.; Duval, E.H.; Sherlock, G. Industrial fuel ethanol yeasts contain adaptive copy number changes in genes involved in vitamin B1 and B6 biosynthesis. Genome Res. 2009, 19, 2271-2278. [CrossRef]
- Oliveira, E.S.; Rosa, C.A.; Morgano, M.A.; Fermentation characteristics as criteria for selection of cachaça yeast. World J. Microbiol. Biotechnol, 2004, 20, 19–24. [CrossRef]
- Marini, M.M.; Gomes, F.C.O.; Silva, C.L.C.; Cadete, R.M.; Badotti, F.; Oliveira, E.S.; Cardoso, C.R.; Rosa, C.A. The use of selected starter Saccharomyces cerevisiae strains to produce traditional and industrial cachaça: a comparative study. World J. Microbiol. Biotechnol. 2009, 25, 235–242. [CrossRef]
- Duval, E.H.; Alves-Jr, S.L.; Dunn, B.; Sherlock, G.; Stambuk, B.U. Microarray karyotyping of maltose-fermenting Saccharomyces yeasts with differing maltotriose utilization profiles reveals copy number variation in genes involved in maltose and maltotriose utilization. J. Appl. Microbiol. 2010, 109, 248-259. [CrossRef]
- DeRisi, J.L.; Iyer, V.R.; Brown, P.O. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science 1997, 278, 680-686. [CrossRef]
- Wang, P.; Kim, Y.; Pollack, J.; Narasimhan, B.; Tibshirani, R. A method for calling gains and losses in array CGH data. Biostatistics 2005, 6, 45–58. [CrossRef]
- Silveira, M.C.; Carvajal, E.; Bom, E.P. Assay for in vivo yeast invertase activity using NaF. Anal. Biochem. 1996, 238, 26-28. [CrossRef]
- Stambuk, B.U. A simple experiment illustrating metabolic regulation: induction versus repression of yeast α-glucosidase, Biochem. Educ. 1999, 27, 177-180. [CrossRef]
- Hollatz, C.; Stambuk, B.U. Colorimetric determination of active α-glucoside transport in Saccharomyces cerevisiae. J. Microbiol. Methods. 2001, 46, 253-259. [CrossRef]
- Jules, M.; Guillou, V.; François, J.; Parrou, J.L. Two distinct pathways for trehalose assimilation in the yeast Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2004, 70, 2771-2778. [CrossRef]
- Petracek, M.E.; Longtine, M.S. PCR-based engineering of yeast genome. Methods Enzymol. 2002, 350, 445-469. [CrossRef]
- Ausubel, F.M.; Brent, R.; Kingston, R.E.; Moore, D.D.; Seidman, J.G.; Smith, J.A.; Struhl, K. Short Protocols in Molecular Biology, 3rd ed.; John Wiley & Sons: New York, USA, 1995.
- Gietz, D.; St Jean, A.; Woods, R.A.; Schiestl, R.H. Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Res. 1992, 20, 1425. [CrossRef]
- Badotti, F.; Batista, A.S.; Stambuk, B.U. Sucrose active transport and fermentation by Saccharomyces cerevisiae. Braz. Arch. Biol. Techn. 2006, 49(Supl 1), 115–123.
- Bastos, R.W.; Pedroso, S.H.; Vieira, A.T.; Moreira, L.M.; França, C.S.; Cartelle, C.T.; Arantes, R.M.; Generoso, S.V.; Cardoso, V.N.; Neves, M.J.; Nicoli, J.R.; Martins, F.S. Saccharomyces cerevisiae UFMG A-905 treatment reduces intestinal damage in a murine model of irinotecan-induced mucositis. Benef. Microbes. 2016, 7, 549-557. [CrossRef]
- Miranda, V.C.; Santos, S.S.; Assis, H.C.; Faria, A.M.C.; Quintanilha, M.F.; Morão, R.P.; Nicoli, J.R.; Cara, D.C.; Martins, F.S. Effect of Saccharomyces cerevisiae UFMG A-905 in a murine model of food allergy. Benef. Microbes. 2020, 11, 255-268. [CrossRef]
- Saldanha, A.J. Java Treeview - extensible visualization of microarray data. Bioinformatics 2004, 20, 3246-3248. [CrossRef]
- Reifenberger, E.; Freidel, K.; Ciriacy, M. Identification of novel HXT genes in Saccharomyces cerevisiae reveals the impact of individual hexose transporters on glycolytic flux. Mol. Microbiol. 1995, 16, 157-167. [CrossRef]
- Diderich, J.A.; Schepper, M.; van Hoek, P.; Luttik, M.A.; van Dijken, J.P.; Pronk, J.T.; Klaassen, P.; Boelens, H.F.; de Mattos, M.J.; van Dam, K.; Kruckeberg, A.L. Glucose uptake kinetics and transcription of HXT genes in chemostat cultures of Saccharomyces cerevisiae. J. Biol. Chem. 1999, 274, 15350-15359. [CrossRef]
- Ozcan, S.; Johnston, M. Function and regulation of yeast hexose transporters. Microbiol. Mol. Biol. Rev. 1999, 63, 554-569. [CrossRef]
- Jordan, P.; Choe, J.Y.; Boles, E.; Oreb, M. Hxt13, Hxt15, Hxt16 and Hxt17 from Saccharomyces cerevisiae represent a novel type of polyol transporters. Sci. Rep. 2016, 6, 23502. [CrossRef]
- Donnini, C.; Lodi, T.; Ferrero, I.; Algeri, A.; Puglisi, P.P. Allelism of IMP1 and GAL2 genes of Saccharomyces cerevisiae. J. Bacteriol. 1992, 174, 3411-3415. [CrossRef]
- Paxhia, M.D.; Downs, D.M. SNZ3 encodes a PLP synthase involved in thiamine synthesis in Saccharomyces cerevisiae. G3: Gen. Genom. Genet. 2019, 9, 335-344. [CrossRef]
- Jacobus, A.P.; Stephens, T.G.; Youssef, P.; González-Pech, R.; Ciccotosto-Camp, M.M.; Dougan, K.E.; Chen, Y.; Basso, L.C.; Frazzon, J.; Chan, C.X.; Gross, J. Comparative genomics supports that Brazilian bioethanol Saccharomyces cerevisiae comprise a unified group of domesticated strains related to cachaça spirit yeasts. Front. Microbiol. 2021, 12, 644089. [CrossRef]
- McIlwain, S.J.; Peris, D.; Sardi, M.; Moskvin, O.V.; Zhan, F.; Myers, K.S.; Riley, N.M.; Buzzell, A.; Parreiras, L.S.; Ong, I.M.; Landick, R.; Coon, J.J.; Gasch, A.P.; Sato, T.K.; Hittinger, C.T. Genome sequence and analysis of a stress-tolerant, wild-derived strain of Saccharomyces cerevisiae used in biofuels research. G3: Gen. Genom. Genet. 2016, 6, 1757-1766. [CrossRef]
- Nosaka, K. Recent progress in understanding thiamin biosynthesis and its genetic regulation in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2006, 72, 30-40. [CrossRef]
- Mojzita, D.; Hohmann, S. Pdc2 coordinates expression of the THI regulon in the yeast Saccharomyces cerevisiae. Mol. Genet. Genomics. 2006, 276, 147-161. [CrossRef]
- Labuschagne, P.; Divol, B. Thiamine: a key nutrient for yeasts during wine alcoholic fermentation. Appl. Microbiol. Biotechnol. 2021, 105, 953-973. [CrossRef]
- Ozcan, S.; Vallier, L.G.; Flick, J.S.; Carlson, M.; Johnston, M. Expression of the SUC2 gene of Saccharomyces cerevisiae is induced by low levels of glucose. Yeast 1997, 13, 127-137. [CrossRef]
- Babrzadeh, F.; Jalili, R.; Wang, C.; Shokralla, S.; Pierce, S.; Robinson-Mosher, A.; Nyren, P.; Shafer, R.W.; Basso, L.C.; de Amorim, H.V.; de Oliveira, A.J.; Davis, R.W.; Ronaghi, M.; Gharizadeh, B.; Stambuk, B.U. Whole-genome sequencing of the efficient industrial fuel-ethanol fermentative Saccharomyces cerevisiae strain CAT-1. Mol. Genet. Genomics 2012, 287, 485-494. [CrossRef]
- Vidgren, V.; Kankainen, M.; Londesborough. J.; Ruohonen, L. Identification of regulatory elements in the AGT1 promoter of ale and lager strains of brewer’s yeast. Yeast 2011, 28, 579-594. [CrossRef]
- Alves-Jr, S.L.; Thevelein, J.M.; Stambuk, B.U. Extracellular maltotriose hydrolysis by Saccharomyces cerevisiae cells lacking the AGT1 permease. Lett. Appl. Microbiol. 2018, 67, 377-383. [CrossRef]
- Vidgren, V.; Huuskonen, A.; Virtanen, H.; Ruohonen, L.; Londesborough, J. Improved fermentation performance of a lager yeast after repair of its AGT1 maltose and maltotriose transporter genes. Appl. Environ. Microbiol. 2009, 75, 2333-2345. [CrossRef]
- Legras, J.L.; Merdinoglu, D.; Cornuet, J.M.; Karst, F. Bread, beer and wine: Saccharomyces cerevisiae diversity reflects human history. Mol. Ecol. 2007, 16, 2091–2102. [CrossRef]
- Liti, G.; Carter, D.M.; Moses, A.M.; Warringer, J.; Parts, L.; James, S.A.; Davey, R.P.; Roberts, I.N.; Burt, A.; Koufopanou, V.; Tsai, I.J.; Bergman, C.M.; Bensasson, D.; O’Kelly, M.J.; van Oudenaarden, A.; Barton, D.B.; Bailes, E.; Nguyen, A.N.; Jones, M.; Quail, M.A.; Goodhead, I.; Sims, S.; Smith, F.; Blomberg, A.; Durbin, R.; Louis, E.J. Population genomics of domestic and wild yeasts. Nature 2009, 458, 337–341. [CrossRef]
- Peter, J.; de Chiara, M.; Friedrich, A.; Yue, J.X.; Pflieger, D.; Bergström, A.; Sigwalt, A.; Barre, B.; Freel, K.; Llored, A.; Cruaud, C.; Labadie, K.; Aury, J.M.; Istace, B.; Lebrigand, K.; Barbry, P.; Engelen, S.; Lemainque, A.; Wincker, P.; Liti, G.; Schacherer, J. Genome evolution across 1,011 Saccharomyces cerevisiae isolates. Nature 2018, 556, 339-344. [CrossRef]
- Liu, Z.L.; Huang, X. Copy number variants impact phenotype-genotype relationships for adaptation of industrial yeast Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2022, 106, 6611-6623. [CrossRef]
- Barbosa, R.; Pontes, A.; Santos, R.O.; Montandon, G.G.; de Ponzzes-Gomes, C.M.; Morais, P.B.; Gonçalves, P.; Rosa, C.A.; Sampaio, J.P. Multiple rounds of artificial selection promote microbe secondary domestication-The case of cachaça yeasts. Genome Biol. Evol. 2018, 10, 1939-1955. [CrossRef]
- Rego-Costa, A.; Huang, I.T.; Desai, M.M.; Gombert, A.K. Yeast population dynamics in Brazilian bioethanol production. G3: Gen. Genom. Genet. 2023, 13, kad104. [CrossRef]
- Alves-Jr, S.L.; Herberts, R.A.; Hollatz, C.; Miletti, L.C.; Stambuk, B.U. Maltose and maltotriose active transport and fermentation by Saccharomyces cerevisiae. J. Am. Soc. Brew. Chem. 2007, 65, 99-104. [CrossRef]
- Pereira, L.F.; Lucatti, E.; Basso, L.C.; de Morais-Jr, M.A. The fermentation of sugarcane molasses by Dekkera bruxellensis and the mobilization of reserve carbohydrates. Antonie Van Leeuwenhoek 2014, 105, 481-489. [CrossRef]
- Erasmus, D.J.; van der Merwe, G.K.; van Vuuren, H.J. Genome-wide expression analyses: Metabolic adaptation of Saccharomyces cerevisiae to high sugar stress. FEMS Yeast Res. 2003, 3, 375-399. [CrossRef]
- Parmar, J.H.; Bhartiya, S.; Venkatesh, K.V. Characterization of the adaptive response and growth upon hyperosmotic shock in Saccharomyces cerevisiae. Mol. Biosyst. 2011, 7, 1138-1148. [CrossRef]
- Petelenz-Kurdziel, E.; Kuehn, C.; Nordlander, B.; Klein, D.; Hong, K.K.; Jacobson, T.; Dahl, P.; Schaber, J.; Nielsen, J.; Hohmann, S.; Klipp, E. Quantitative analysis of glycerol accumulation, glycolysis and growth under hyper osmotic stress. PLoS Comput. Biol. 2013, 9, e1003084. [CrossRef]
- Blomberg, A. Yeast osmoregulation - glycerol still in pole position. FEMS Yeast Res. 2022, 22, foac035. [CrossRef]
Figure 1.
Copy number differences and similarities among industrial sugarcane yeast strains for genes involved in the metabolism and transport of sugar, and of vitamins B1 and B6. (A) array-CGH data showing the copy number of genes involved in sugar fermentation among the different strains. (B) array-CGH data showing the copy number of genes involved in thiamine (vitamin B1) and pyridoxine (vitamin B6) biosynthesis and transport among the different strains. A scale of relative gene copy number is shown at the bottom.
Figure 1.
Copy number differences and similarities among industrial sugarcane yeast strains for genes involved in the metabolism and transport of sugar, and of vitamins B1 and B6. (A) array-CGH data showing the copy number of genes involved in sugar fermentation among the different strains. (B) array-CGH data showing the copy number of genes involved in thiamine (vitamin B1) and pyridoxine (vitamin B6) biosynthesis and transport among the different strains. A scale of relative gene copy number is shown at the bottom.
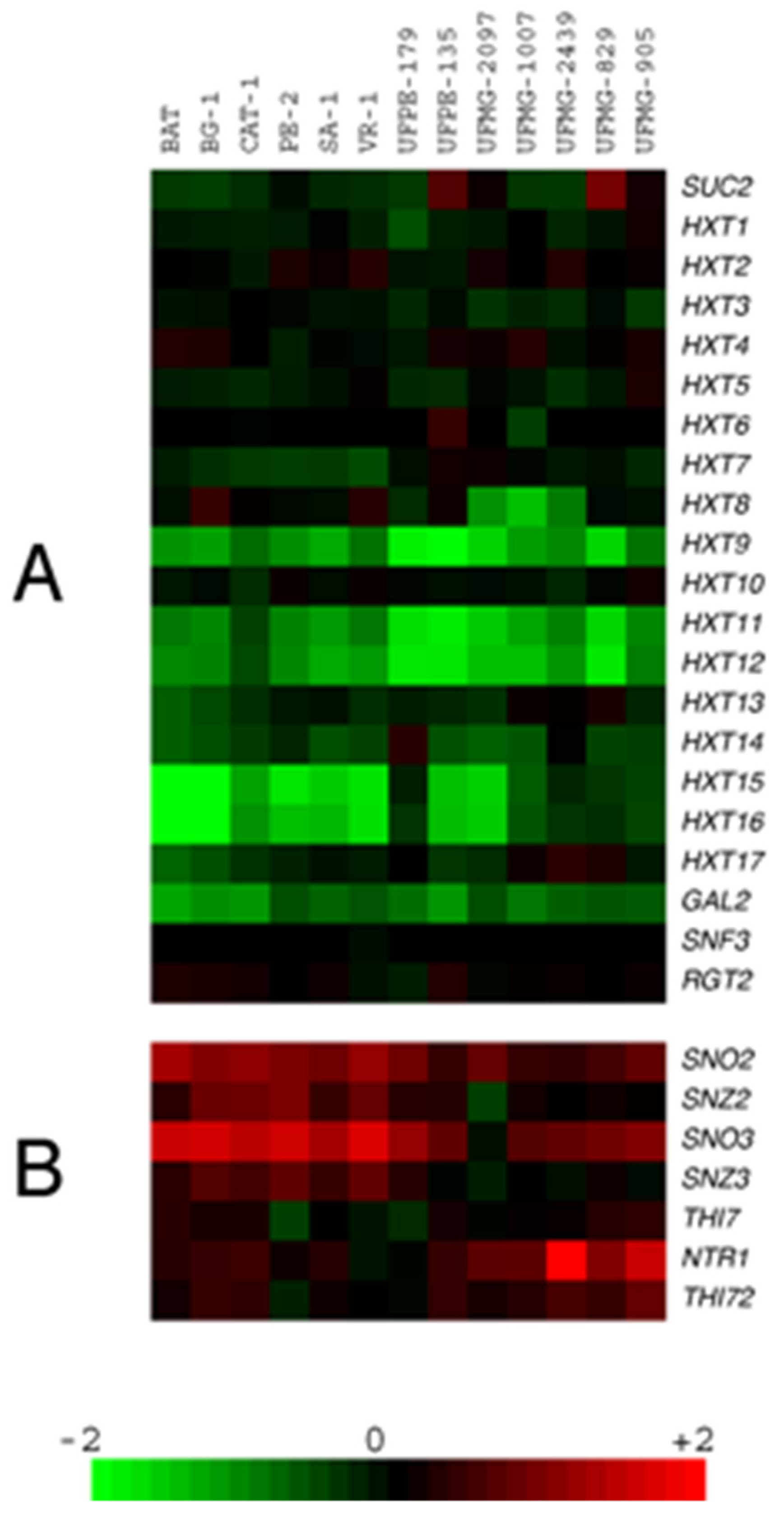
Figure 2.
Detection of SUC genes in the industrial sugarcane yeasts. (a) PFGE separation of yeast chromosomes (ethidium-bromide stained). (b) Southern blot of gel shown in panel (a), hybridized with a probe for SUC2 to detect which chromosomes carry SUC genes. (Lane 1) Reference laboratory strain S288C, which contains SUC2 on chromosome IX (indicated to the left of panel a); (lanes 2–6) strains CAT-1, PE-2, BG-1, SA-1, and UFPE-136, respectively. This last strain has both the SUC2 gene in chromosome IX and the SUC1 gene in chromosome VII.
Figure 2.
Detection of SUC genes in the industrial sugarcane yeasts. (a) PFGE separation of yeast chromosomes (ethidium-bromide stained). (b) Southern blot of gel shown in panel (a), hybridized with a probe for SUC2 to detect which chromosomes carry SUC genes. (Lane 1) Reference laboratory strain S288C, which contains SUC2 on chromosome IX (indicated to the left of panel a); (lanes 2–6) strains CAT-1, PE-2, BG-1, SA-1, and UFPE-136, respectively. This last strain has both the SUC2 gene in chromosome IX and the SUC1 gene in chromosome VII.
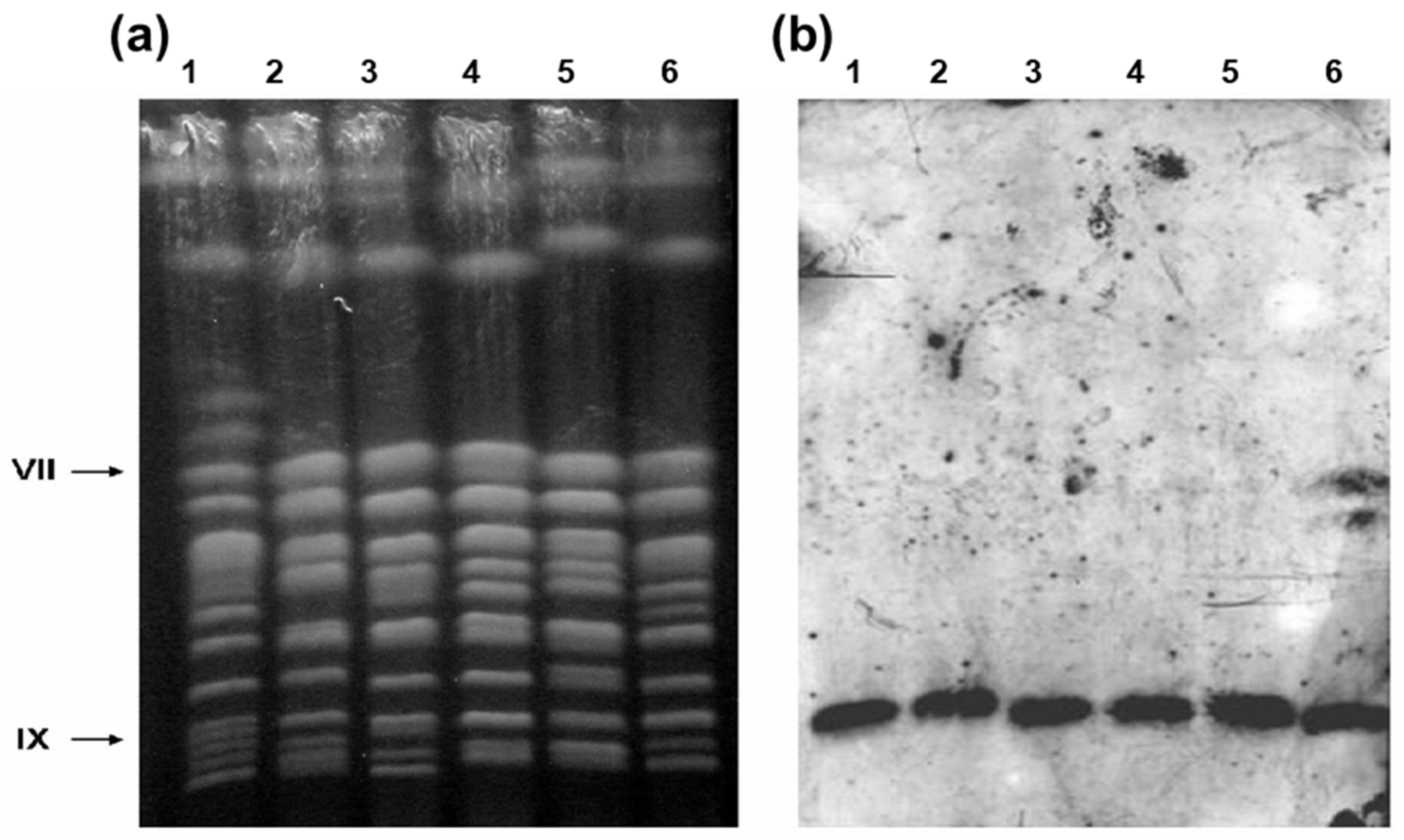
Figure 3.
Invertase activity of the sugarcane industrial yeast strains. The extracellular invertase activity was determined after the growth of the cells in rich medium containing 20 g/L sucrose, 20 g/L ethanol plus 30 g/L glycerol, or this last medium supplemented with 1 g/L glucose.
Figure 3.
Invertase activity of the sugarcane industrial yeast strains. The extracellular invertase activity was determined after the growth of the cells in rich medium containing 20 g/L sucrose, 20 g/L ethanol plus 30 g/L glycerol, or this last medium supplemented with 1 g/L glucose.
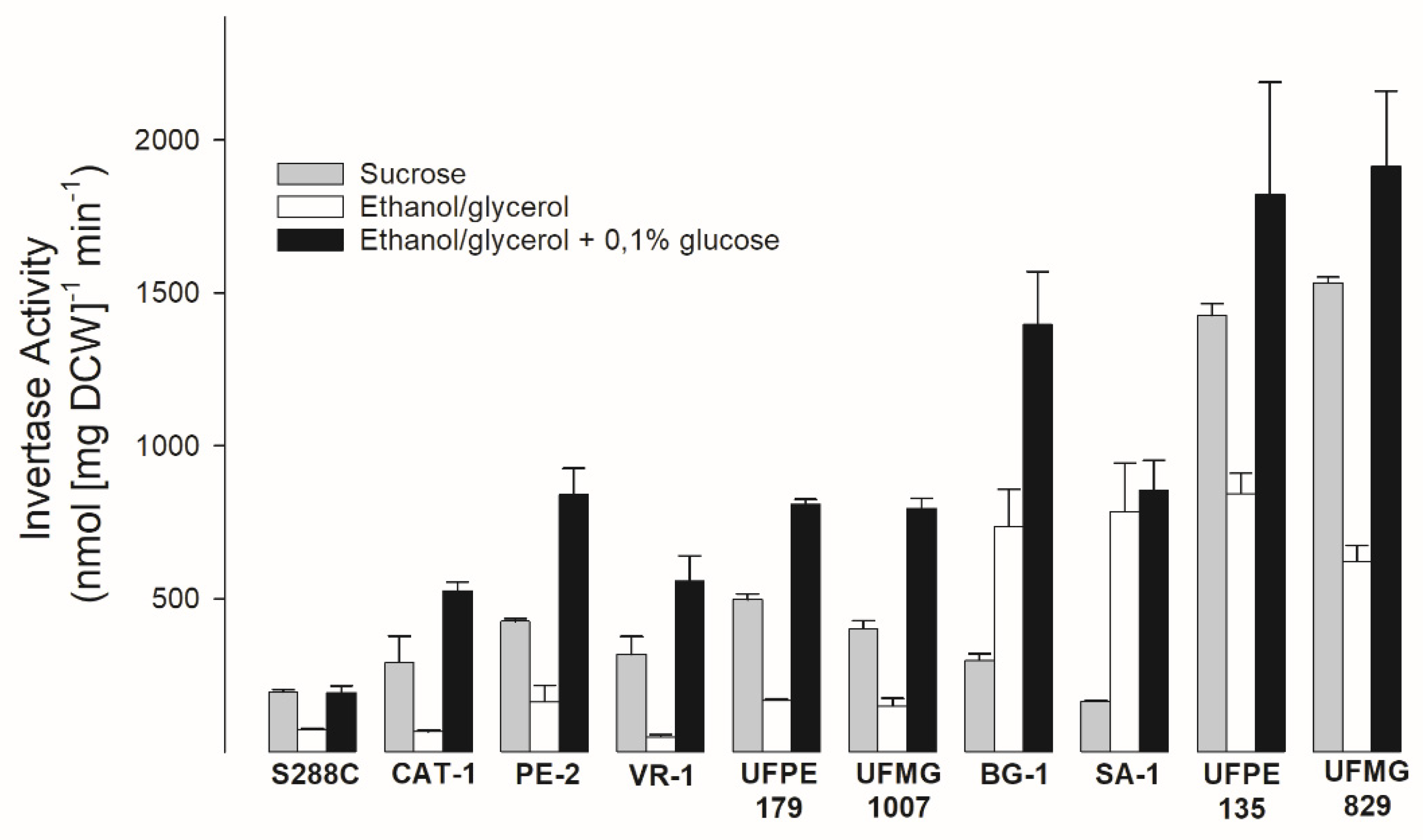
Table 1.
Yeast strains used in this study.
| Yeast strains | Relevant features | Source |
|---|---|---|
| BAT | Industrial fuel ethanol strain isolated in 2011 from Usina Batatais, São Paulo, Brazil. | [35] |
| BG-1 | Industrial fuel ethanol strain isolated in 1989/1990 from Usina Barra Grande, Sao Paulo, Brazil. | [36] |
| CAT-1 | Industrial fuel ethanol strain isolated in 1998/1999 from Usina VO Catanduva, São Paulo, Brazil. | [12,36] |
| PE-2 | Industrial fuel ethanol strain isolated in 1993/1994 from Usina da Pedra, Sao Paulo, Brazil. | [12,36] |
| SA-1 | Industrial fuel ethanol strain isolated in 1989/1990 from Usina Santa Adelia, Sao Paulo, Brazil. | [36] |
| S288C | MATα mal gal2 mel flo1 flo8-1 hap1 SUC2 | [35] |
| BSY21-34B3 | MATa ura3-52 trp-289 kanMX-PADH1::iSUC2 | [32] |
| UFMG-829 | Cachaça strain isolated in 1996 from a distillery in Porto Firme, Minas Gerais, Brazil. | [13] |
| UFMG-905 | Cachaça strain isolated in 1996 from a distillery in Nova União, Minas Gerais, Brazil. | [13,37] |
| UFMG-1007 | Cachaça strain isolated in 1996 from a distillery in Salinas, Minas Gerais, Brazil. | [13] |
| UFMG-2097 | Cachaça strain isolated in 1999 from a distillery in Salinas, Minas Gerais, Brazil. | [38] |
| UFMG-2439 | Cachaça strain isolated in 1999 from a distillery in Salinas, Minas Gerais, Brazil. | [38] |
| UFPE-135 | Industrial fuel ethanol strain isolated in 1998/1999 from Japungu distillery, Paraíba, Brazil | [10] |
| UFPE-179 | Industrial fuel ethanol strain isolated in 1998/1999 from Miriri distillery, Paraíba, Brazil | [10] |
| VR-1 | Industrial fuel ethanol strain isolated in 1993/1994 from Usina Vale do Rosario, Sao Paulo, Brazil | [12,36] |
| GMY08 | Isogenic to CAT-1, but kanMX-PADH1::iSUC2 / suc2Δ::BleR PTDH3::AGT1 / AGT1 | This work |
Table 3.
Invertase activity, activity of the AGT1 permease, and glycerol produced by the indicated yeast strains.
Table 3.
Invertase activity, activity of the AGT1 permease, and glycerol produced by the indicated yeast strains.
| Strain | Activity1 (nmol of product [mg DCW]-1 min-1) | Glycerol produced at the end of fermentation2 (g/L) | ||
|---|---|---|---|---|
| Extracellular invertase | Intracellular invertase | pNPαG transport | ||
| CAT-1 | 295 ± 70 | 74 ± 6 | 0.1 ± 0.1 | 10.5 ± 1.2 |
| GMY08 | 56 ± 2 | 2,223 ± 45 | 4.5 ± 0.3 | 5.4 ± 0.7 |
1 Determined with cells grown with 20 g/L sucrose. 2 Fermentation of >200 g/L sucrose.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



