
Submitted:
04 July 2023
Posted:
05 July 2023
You are already at the latest version
Abstract
Heart is composed of muscle cells called cardiomyocytes, including a specialized population, named pacemaker cells, that form the Cardiac Conduction System (CCS), responsible for generating the action potential dictating heart contractions. Failure of the CCS system leads to cardiac arrhythmias requiring complicated therapies and often surgical implantation of electrical pacemakers. However, recent research focusses on development of novel therapies using biological pacemakers aiming to substitute electrical devices. While most signalling pathways and transcription factors involved in the development of the pacemaker cells are known, the upstream regulatory networks need to be predicted through computer-based databases, mathematical modelling as well as functional testing of the regulatory elements in vivo, indicating the need for further research. Here we summarise the current knowledge about the vertebrate myocardial CCS system and development of the pacemaker cells and emphasise areas of future research to clarify the regulation of muscle pacemaker cells and ease development of biological therapies.
Keywords:
Cardiac conduction system
; pacemaker
; gene regulatory network
1. Contracting Cardiac Muscle
The heart is a muscular organ that pumps to circulate the blood throughout the body with the ability to initiate and coordinate electrical impulses to begin contractions of the atria and ventricles. The production of electrical impulses by the heart is regulated by the Cardiac Conduction System (CCS), specifically the Sinoatrial Node (SAN) (Mohan et al., 2017; van Weerd and Christoffels, 2016). These contractions are accomplished by the well-timed coordination of the fast and slow components of the CCS, which are of myogenic origin (Cheng et al., 1999; Meilhac et al., 2004). In higher vertebrates, the CCS is classified into the SAN or the primary pacemaker site, atrioventricular node (AVN), the secondary pacemaker site, and the “wiring” of the ventricles, the His-Purkinje fibres (Davies, 1942; Mohan et al., 2017). Electrical impulses are first generated in the SAN, located between the superior vena cava and the right atrium, and are rapidly propagated through the cardiomyocytes (CMs) of the atrium, resulting in an atrial contraction (Kennedy et al., 2016). After the initiation of an atrial contraction, the electrical impulses reach the slow conducting tissues of the secondary pacemaker site, the AVN, located within the atrioventricular septum, causing a delay before leading on to the His-Purkinje fibres, resulting in a ventricular contraction (Moorman et al., 1998; van Weerd and Christoffels, 2016) (Figure 1).
Within a human embryo, the first signs of heart muscle contraction typically begin around embryonic day 22 during the third week of gestation, when the first heart field develops into the heart tube (Tan and Lewandowski, 2020). Comparatively, contraction of the heart muscle in mice begins earlier around embryonic days E8 to E9 (Andres-Delgado and Mercader, 2016; Lindsey et al., 2014). However, in zebrafish, with a different anatomical structure of the heart muscle compared to human, mouse or chicken, contraction begins at 22 hours post fertilisation with contraction beginning in chicken at around HH10 to HH11 (Martinsen, 2005; Wittig and Munsterberg, 2020; Xia et al., 2020). Once the heart tube fully develops, the rate of contraction of the heart muscle increases with the heart forming the ability to pump blood efficiently throughout the body (Kennedy et al., 2016; Moorman et al., 1998; van Weerd and Christoffels, 2016).
Studies from mammalian, avian and fish model systems have shown that each CCS component consists of a specialized group of CMs with distinctive morphological and electrophysiological properties and transcriptional profiles (Anderson et al., 2009). The dysfunction of any of CCS components can lead to cardiac arrhythmias like Brugada syndrome, long QT syndrome and sudden cardiac death (Barbuti et al., 2007; Cohle et al., 2002). Some of these arrhythmias complicate therapy for congenital cardiac conditions. As a result, comprehension of the CCS gene regulatory work is essential for the creation of therapeutics for cardiac arrhythmias.
Treatment of various types of arrhythmias need the installation of electrical pacemakers. Current and older generations of electrical pacemakers have been known to have a variety of limitations to their design and function including issues with battery life, generator failure and risk of infection with materials used (Alasti et al., 2018; Cingolani et al., 2017). The limitations and complications with electrical pacemakers led to biological pacemakers being researched and developed to be used in alternative (Cingolani et al., 2017). Although heart development has been studied for more than a century, little is known about the development of its pacemaker cells mainly due to the absence of a complete understanding of the genetic mechanism regulating its development. The development of biological pacemakers needs the translation of already known knowledge of the signaling pathways, transcription factors, and their gene regulatory networks to increase the reprogramming efforts.
2. Key Transcription factors involved in Cardiac Conduction System development
Cardiac Conduction System development and homeostasis is reliant on transcriptional and regulatory networks that are embryonic-stage-dependent, dose-dependent, and tissue-dependent (Li et al., 2022; Liang et al., 2020; Schram et al., 2002). A cascade of transcription factors including SHOX2, BMP4, SCN5A, TBX3, TBX5, NKX2-5, ID2 and more has been established as instrumental to this spatial divergence in myocytes development, as will be evidenced in the sub-sections below.
2.1. Short stature homeobox 2 (SHOX2) and bone morphogenic protein 4 (BMP4)
SHOX2 and BMP4 are important for SAN formation (Liang et al., 2015; van Weerd and Christoffels, 2016); van Weerd and Christoffels, 2016). Dysfunction of the SAN can lead to a variety of cardiac arrhythmias including bradycardic arrhythmias (Hu et al., 2018). SHOX2 transcription factor is essential for SAN formation and differentiation with SHOX2 being widely expressed throughout the body including the heart muscle (Hu et al., 2018). BMPs are a group of signalling molecules that belongs to the Transforming Growth Factor β superfamily of proteins that have a key role in pacemaker development. The key role of BMP4 in embryonic heart development is the promotion of fibroblast reprogramming to cardiomyocytes with pacemaker activity (Efe et al., 2011). BMP4 is also a target of SHOX2 with the expression of both SHOX2 and BMP4 overlapping (Puskaric et al., 2010). One of the main functions of BMP4 is its role in the differentiation of the cardiac pacemaker cells (Wu et al., 2019). A series of epistatic genetic experiments in Xenopus have elucidated that SHOX2 interacts directly with BMP4 promoter and the expression patterns of BMP4 and SHOX2 overlap in the SAN during embryonic development (Puskaric et al., 2010).
2.2. T-box transcription factor 5 (TBX5), NK2 Homeobox 5 (NDX2-5) and Inhibitor of DNA binding 2 (ID2)
TBX5, NKX2-5 and ID2 are essential for atrioventricular bundle and bundle branch development (Moskowitz et al., 2007; van Weerd and Christoffels, 2016). TBX5 has a variety of roles throughout the body but specifically has main function within cardiac development (Moskowitz et al., 2007). Mutations of the TBX5 transcription factor is known to lead to cardiac defects at the cardiac septa and cardiac conduction system. Within early embryonic cardiac development, TBX5 works as a transcriptional activator for genes working in the maturation of cardiomyocytes (Moskowitz et al., 2007). Later in cardiac development, TBX5 role changes and focusses on the structure of the cardiac conduction system as well as the maintenance of cardiomyocyte maturation (Moskowitz et al., 2007). NKX2-5 is a cardiac homeobox transcription factor that is expressed throughout the cardiac system (Nakashima et al., 2014). Mutations within NKX2-5 lead to cardiac defects and atrioventricular conduction abnormalities. Throughout cardiac development, NKX2-5 has been found to have roles in the regulation of the working and conducting myocytes within the atria whilst working alongside the Notch signalling pathway (Nakashima et al., 2014). ID2 is a cardiac transcription factor with expression being detected around the neural crest, inflow and outflow tract as well as within the neurons around the aorta and pulmonary artery (Fraidenraich et al., 2004; Hu et al., 2019). Later in development ID2 is expressed in the atrioventricular bundle at E12.5 and within the bundle branches around E16.5 (Hu et al., 2019; Moskowitz et al., 2007).
2.3. T-Box Transcription Factor 3 (TBX3)
TBX3 is expressed in the cardiac conduction system of the heart muscle and is essential for repressing atrial differentiation (Wiese et al., 2008). Various studies have identified noncoding variants near expression of TBX3 that is linked to PR interval and QRS duration, suggesting that changes to TBX3 expression has the ability to affect the function of the atrioventricular conduction system (Eif et al., 2018; Harst et al., 2016; Mohan et al., 2017; Pfeufer et al., 2010; Setten et al., 2018; Sotoodehnia et al., 2010; Verweij et al., 2014).
2.4. T-box transcription factor 18 (TBX18)
TBX18 has key roles in heart muscle development, particularly in the formation and structure of the SAN (Greulich et al., 2011; Wu et al., 2019). Furthermore, expression of TBX18 is essential for early SAN specification and specifically produces pacemaker activity within early development of embryonic heart muscle formation (Cho, 2015; McNally and svensson, 2009; Wiese et al., 2008). Additionally, one study successfully converted ventricular myocytes to induced pacemaker cells within postnatal rats via TBX18 expression potentially providing a novel method for biological pacemakers (Kapoor et al., 2013).
2.5. ISLET-1 (ISL1)
ISL1 is a transcription factor with roles in multiple organs during embryonic development with roles within cardiac development specifically as a marker for second heart field progenitors (Ren et al., 2021). ISL1 expression is detected as early as E7 in mouse heart development however, expression of ISL1 changes throughout development with expression being seen in the SAN from postnatal to adulthood (Zhou et al., 2019).
2.6. GATA4
GATA4 is a regulator of cardiomyocyte proliferation, and differentiation with expression being high until birth but remains detectable in all cardiomyocytes (Whitcomb et al., 2020). Mutations with GATA4 results in cardia bifida which signifies the function of GATA4 in early heart formation (Molkentin et al., 1997; Whitcomb et al., 2020). GATA6 is a regulator of SAN development with mutations with GATA6 leading to dysfunction with SAN patterning and size leading to arrhythmias (Gharibeh et al., 2021).
2.7. HAND1
HAND1 plays an essential role in the specification and differentiation of embryonic structures including the cardiac muscle of the heart (Zheng et al., 2021). HAND1 is an essential regulator for cardiac precursor cell fate decision and morphogenesis regulated by the signalling pathway, BMP (Firulli et al., 2020). Mutations within HAND1 have been shown to be associated with cogenital heart disease (Firulli et al., 2014; Vincentz et al., 2017). Further research found that BMP can activate BMP signalling-HAND1 regulation within heart muscle development (Zheng et al., 2021).
2.8. IRX3
IRX3 is essential for the regulation of rapid electrical propagation in the ventricular conduction system by transcription of Cx40 and Cx43 (Kim et al., 2016). Development of the ventricular conduction system is regulated by the activation of transcription factors including NKX2-5, TBX3, TBX5, and ID2 (Bakker et al., 2008; Briggs et al., 2008; Hoogaars et al., 2004; Jay et al., 2004; Kim et al., 2016; Moskowitz et al., 2007; Moskowitz et al., 2004). Loss of the transcription factors leads to many cardiac defects specifically the loss of NKX2-5 and TBX5 leading to the increased chance of developing arrhythmias (Kim et al., 2016). IRX5 is expressed in a gradient in the ventricular myocardium with the epicardium having the lowest expression and the endocardium having the highest expression (Munshi, 2012). IRX5 mutations leads to increased chance of developing arrhythmias due to having abnormal repolarisation within the ventricular conduction system in the absence of a homeostatic Kv4.2 gradient (Costantini et al., 2005; Munshi, 2012).
3. Key Signalling pathways involved in CCS development
The conserved signalling pathways that have been found to be crucial for CCS specialization are Notch, BMP, Wnt, and NKX2-5 (Hoogaars et al., 2004; Liang et al., 2015; Luxan et al., 2016; Wang et al., 2020).
3.1. Notch signalling
Notch signalling pathway is a conserved pathway that has critical roles in the regulation of cell fate specification and their differentiation, and tissue patterning (Luxan et al., 2016; Nakano et al., 2018; Rentschler et al., 2012). Similarly, a knock-out study revealed Notch1 to be lethal around E9.5 to E11.5 as it controls the development of the sinus venous valve and the SAN by coordinating myocardial Wnt and NRG1 signalling functions (Luxan et al., 2016; Wang et al., 2020).
3.2. BMP signalling pathway
The BMP (bone morphogenetic protein) signalling pathway is involved in the differentiation of the sinoatrial node (SAN) and the atrioventricular node (AVN) and regulates cardiac progenitor development (Wang et al., 2010). Regulation of the BMP signalling pathway is coordinated by the SMAD proteins with the BMPs being associated with the TGFβ superfamily (Callis et al., 2005). The TGFβ pathway is involved in processes throughout the heart muscle as well as in the formation and patterning of the cardiac conduction system (Hoogaars et al., 2004; Yousefi et al., 2020). Various studies have shown that TGFβ signalling is essential for heart muscle development specifically TGFβ1, 2, and 3 being expressed in specific regions and stages of the cardiac conduction system development of the heart muscle (Ramos-Mondragon et al., 2008).
3.3. Wnt signalling
The Wnt signalling pathway regulates the proliferation and differentiation of cardiac progenitor cells during cardiac development and the formation of the conduction system. Recently, Liang and colleagues have shown that canonical Wnt signalling promotes pacemaker cell specification of cardiac mesodermal cells derived from mouse and human embryonic stem cells (Liang et al., 2020). They have shown that one of the key canonical Wnt/β-catenin ligand, Wnt3a, enhances the expression of a chamber of cardiomyocyte genes NKX2-5. This rises the number of pacemaker-like myocytes while reducing cardiac troponin T-positive pan-cardiac differentiation (Liang et al., 2020).
The signalling pathways involved with the development of the cardiac conduction system of the heart muscle interact with each other and with various other factors to regulate the development and function of the CCS. Reprogramming efforts in human induced pluripotent stem cells have shown the impact of other signalling pathways like FGF and retinoic acid which reprogram the cardiac mesoderm to generate SAN-like cells (Liu et al., 2020b). Transcriptome analysis of mouse and human sinoatrial node cells and sinoatrial ring (SAR) in zebrafish reveals a conserved genetic program (Burkhard and Bakkers, 2018; Efe et al., 2011; Liang et al., 2020; Minhas et al., 2021; Puskaric et al., 2010; van Eif et al., 2019) (Liu et al., 2020a).
4. The genetic network of CCS development
A unique gene expression mechanism enables cardiac pacemaker cells in the SAN to fire autonomously and initiate the heartbeat. The CCS is evolutionarily conserved in the building plan of the heart, and this indicates that the cellular and molecular mechanisms that drive the formation of pacemaker tissues are almost similar among vertebrates. Studies have shown that mammalian pacemaker CMs exhibit typical pacemaker action potentials and express molecular markers such as Isl1, Shox2 and Hcn4 (Blaschke et al., 2007; Minhas et al., 2021; Tessadori et al., 2012; Vedantham et al., 2015). These mammalian genes are conserved in zebrafish and other teleost species. Knocking down of these genes in zebrafish leads to bradycardia, a phenotype indicating defects in cardiac pacemaker activity, further supporting their conserved roles for regulating pacemaker development in zebrafish (Blaschke et al., 2007; de Pater et al., 2009; Tessadori et al., 2012). The transcription factors (TFs) and the signaling pathways that control CCS specification constitute a gene regulatory network, and at the core of these networks are the cis-acting regulatory regions that are bound by TFs. Recent studies in mice have reported an Isl1 specific enhancer which has not been identified in zebrafish (Galang et al., 2020). However, there is limited understanding of the underlying gene regulatory network of these critical cells that are responsible for the electrical conduction of the heart.
For better understanding of such GRNs, the identification of cis-regulatory modules involved in the development of the cardiac conduction system will pave the way to elucidate molecular mechanisms underlying their regulation of expression. Tissue-specific gene expression obliges long-range regulatory regions, such as enhancers, which dictate the precisely spatial-temporal and dosage-dependent expression of their target genes (Minhas et al., 2019). The availability of publicly available genomics data, well-established protocols for chromosomal conformation capture followed by next-generation sequencing in isolated hearts (3C, 4C-Seq, 5C, Hi-Seq), or derived methods like FAIRE-Seq and ATAC-Seq, single-cell sequencing, multiple genome-wide Chip-seq datasets and evolutionary conservation studies across various vertebrate model species (for example mice, chicken or zebrafish) can identify multiple regulatory landscapes which act as cardiac conduction-specific enhancers (Moskowitz et al., 2007). These identified enhancers can then be functionally tested in vivo, and mutants generated using CRISPR/Cas9 (Figure 2). Mutant generation by genome-editing techniques like CRISPR/Cas9 can be used to: (1) study the role of the identified enhancers in development of CCS, and (2) gene expression patterns of the regulated genes can provide insights into diseases like cardiac arrythmias.
Thus, the GRN for key TFs involved in CCS like NKX2-5, TBX3, TBX5, ISL1, GATA4, GATA6, HAND1, SHOX2, IRX3 and IRX5 needs to be investigated systematically to build the CCS-specific gene regulatory network.
5. Conclusion and future directions
Building a CCS-Specific GRNs needs computational predictions through publicly accessible databases, mathematically modelling, followed by functional testing of the regulatory elements in vivo. This will be further validated by knock-out studies. This needs a lot of seriously multi-disciplinary collaborative work (including work from system biologists, developmental biologists, and molecular biologists, and computational experts) and funding to work in parallel to work and highlight and predict the important genes and their putative regulatory regions to test them functionally.
References
- Alasti, M. , Machado, C., Rangasamy, K., Bittinger, L., Healy, S., Kotschet, E., Adam, D., Alison, J., 2018. Pacemaker-mediated arrhythmias. J Arrhythm 34, 485-492.
- Anderson, R.H.; Yanni, J.; Boyett, M.R.; Chandler, N.J.; Dobrzynski, H. The anatomy of the cardiac conduction system. Clin. Anat. 2009, 22, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Delgado, L.; Mercader, N. Interplay between cardiac function and heart development. Biochim. et Biophys. Acta (BBA) - Mol. Cell Res. 2016, 1863, 1707–1716. [Google Scholar] [CrossRef] [PubMed]
- Bakker, M. , Boukens, B., Mommersteeg, M., Brons, J., Wakker, V., Moorman, A., Christoffels, V.M., 2008. Transcription Factor TBX3 Is Required for the Specification of the Atrioventricular Conduction System. Circulation Research 102, 1340-1349.
- Barbuti, A.; Baruscotti, M.; Difrancesco, D. The Pacemaker Current: From Basics to the Clinics. J. Cardiovasc. Electrophysiol. 2007, 18, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Blaschke, R.J.; Hahurij, N.D.; Kuijper, S.; Just, S.; Wisse, L.J.; Deissler, K.; Maxelon, T.; Anastassiadis, K.; Spitzer, J.; Hardt, S.E.; et al. Targeted Mutation Reveals Essential Functions of the Homeodomain Transcription Factor Shox2 in Sinoatrial and Pacemaking Development. Circulation 2007, 115, 1830–1838. [Google Scholar] [CrossRef]
- Briggs, L.E.; Takeda, M.; Cuadra, A.E.; Wakimoto, H.; Marks, M.H.; Walker, A.J.; Seki, T.; Oh, S.P.; Lu, J.T.; Sumners, C.; et al. Perinatal Loss of Nkx2-5 Results in Rapid Conduction and Contraction Defects. Circ. Res. 2008, 103, 580–590. [Google Scholar] [CrossRef]
- Burkhard, S.B. , Bakkers, J., 2018. Spatially resolved RNA-sequencing of the embryonic heart identifies a role for Wnt/beta-catenin signaling in autonomic control of heart rate. Elife 7.
- Callis, T.E. , Cao, D., Wang, D.-Z. Bone Morphogenetic Protein Signaling Modulates Myocardin Transactivation of Cardiac Genes. Circ. Res. 2005, 97, 992–1000. [Google Scholar] [CrossRef]
- Cheng, G.; Litchenberg, W.H.; Cole, G.J.; Mikawa, T.; Thompson, R.P.; Gourdie, R.G. Development of the cardiac conduction system involves recruitment within a multipotent cardiomyogenic lineage. Development 1999, 126, 5041–5049. [Google Scholar] [CrossRef]
- Cho, H.C. Pacing the Heart with Genes: Recent Progress in Biological Pacing. Curr. Cardiol. Rep. 2015, 17, 65–65. [Google Scholar] [CrossRef]
- Cingolani, E.; Goldhaber, J.I.; Marbán, E. Next-generation pacemakers: from small devices to biological pacemakers. Nat. Rev. Cardiol. 2017, 15, 139–150. [Google Scholar] [CrossRef]
- Cohle, S.D.; Suarez-Mier, M.P.; Aguilera, B. Sudden Death Resulting from Lesions of the Cardiac Conduction System. Am. J. Forensic Med. Pathol. 2002, 23, 83–89. [Google Scholar] [CrossRef]
- Costantini, D.L.; Arruda, E.P.; Agarwal, P.; Kim, K.-H.; Zhu, Y.; Zhu, W.; Lebel, M.; Cheng, C.W.; Park, C.Y.; Pierce, S.A.; et al. The Homeodomain Transcription Factor Irx5 Establishes the Mouse Cardiac Ventricular Repolarization Gradient. Cell 2005, 123, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Davies, F. THE CONDUCTING SYSTEM OF THE VERTEBRATE HEART. Hear. 1942, 4, 66–76. [Google Scholar] [CrossRef] [PubMed]
- de Pater, E.; Clijsters, L.; Marques, S.R.; Lin, Y.-F.; Garavito-Aguilar, Z.V.; Yelon, D.; Bakkers, J. Distinct phases of cardiomyocyte differentiation regulate growth of the zebrafish heart. Development 2009, 136, 1633–1641. [Google Scholar] [CrossRef]
- Efe, J.A.; Hilcove, S.; Kim, J.; Zhou, H.; Ouyang, K.; Wang, G.; Chen, J.; Ding, S. Conversion of mouse fibroblasts into cardiomyocytes using a direct reprogramming strategy. Nature 2011, 13, 215–222. [Google Scholar] [CrossRef] [PubMed]
- van Eif, V.W.W.; Devalla, H.D.; Boink, G.J.J.; Christoffels, V.M. Transcriptional regulation of the cardiac conduction system. Nat. Rev. Cardiol. 2018, 15, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Firulli, B.A.; Fuchs, R.K.; Vincentz, J.W.; Clouthier, D.E.; Firulli, A.B. Hand1 phosphoregulation within the distal arch neural crest is essential for craniofacial morphogenesis. Development 2014, 141, 3050–3061. [Google Scholar] [CrossRef]
- Firulli, B.A.; George, R.M.; Harkin, J.; Toolan, K.P.; Gao, H.; Liu, Y.; Zhang, W.; Field, L.J.; Liu, Y.; Shou, W.; et al. HAND1 loss-of-function within the embryonic myocardium reveals survivable congenital cardiac defects and adult heart failure. Cardiovasc. Res. 2019, 116, 605–618. [Google Scholar] [CrossRef]
- Fraidenraich, D.; Stillwell, E.; Romero, E.; Wilkes, D.; Manova, K.; Basson, C.T.; Benezra, R.; Lisci, M.; Barton, P.R.; Lezmy, J.; et al. Rescue of Cardiac Defects in Id Knockout Embryos by Injection of Embryonic Stem Cells. Science 2004, 306, 247–252. [Google Scholar] [CrossRef]
- Galang, G.; Mandla, R.; Ruan, H.; Jung, C.; Sinha, T.; Stone, N.R.; Wu, R.S.; Mannion, B.J.; Allu, P.K.; Chang, K.; et al. ATAC-Seq Reveals an Isl1 Enhancer That Regulates Sinoatrial Node Development and Function. Circ. Res. 2020, 127, 1502–1518. [Google Scholar] [CrossRef]
- Gharibeh, L.; Yamak, A.; Whitcomb, J.; Lu, A.; Joyal, M.; Komati, H.; Liang, W.; Fiset, C.; Nemer, M. GATA6 is a regulator of sinus node development and heart rhythm. Proc. Natl. Acad. Sci. 2020, 118. [Google Scholar] [CrossRef]
- Greulich, F.; Rudat, C.; Kispert, A. Mechanisms of T-box gene function in the developing heart. Cardiovasc. Res. 2011, 91, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Harst, P.v.d. , Setten, J.v., Verweij, N., Vogler, G., Franke, L., Maurano, M.T., Wang, X., Leach, I.M., Eijgelsheim, M., Sotoodehnia, N., et al. 2016. 52 Genetic Loci Influencing Myocardial Mass. Journal of the American College of Cardiology 68, 1435-1448.
- Hoogaars, W.M.H. , Tessari, A., Moorman, A.F.M., Boer, P.A.J.d., Hagoort, J., Soufan, A.T., Campione, M., Christoffels, V.M., 2004. The transcriptional repressor Tbx3 delineates the developing central conduction system of the heart. Cardiovascular Research 62, 489-499.
- Hu, W.; Xin, Y.; Zhao, Y.; Hu, J. Shox2: The Role in Differentiation and Development of Cardiac Conduction System. Tohoku J. Exp. Med. 2018, 244, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Xin, Y.; Hu, J.; Sun, Y.; Zhao, Y. Inhibitor of DNA binding in heart development and cardiovascular diseases. Cell Commun. Signal. 2019, 17, 51. [Google Scholar] [CrossRef] [PubMed]
- Jay, P.Y. , Harris, B.S., Maguire, C.T., Buerger, A., Wakimoto, H., Tanaka, M., Kupershmidt, S., Roden, D.M., Schultheiss, T.M., O’Brien, T.X., Gourdie, R.G., Berul, C.I., Izumo, S., 2004. Nkx2-5 mutation causes anatomic hypoplasia of the cardiac conduction system. The Journal of Clinical Investigation 113, 1130-1137.
- Kapoor, N.; Liang, W.; Marbán, E.; Cho, H.C. Direct conversion of quiescent cardiomyocytes to pacemaker cells by expression of Tbx18. Nat. Biotechnol. 2012, 31, 54–62. [Google Scholar] [CrossRef]
- Kennedy, A. , Finlay, D.D., Guldenring, D., Bond, R., Moran, K., McLaughlin, J., 2016. The Cardiac Conduction System: Generation and Conduction of the Cardiac Impulse. Critical Care Nursing Clinics of North America 28, 269-279.
- Kim, K.-H.; Rosen, A.; Hussein, S.M.I.; Puviindran, V.; Korogyi, A.S.; Chiarello, C.; Nagy, A.; Hui, C.-C.; Backx, P.H. Irx3 is required for postnatal maturation of the mouse ventricular conduction system. Sci. Rep. 2016, 6, srep19197–19197. [Google Scholar] [CrossRef]
- Li, K.Y.C.; Cook, A.C.; Lovering, R.C. GOing Forward With the Cardiac Conduction System Using Gene Ontology. Front. Genet. 2022, 13, 802393. [Google Scholar] [CrossRef]
- Liang, W.; Han, P.; Kim, E.H.; Mak, J.; Zhang, R.; Torrente, A.G.; Goldhaber, J.I.; Marbán, E.; Cho, H.C. Canonical Wnt signaling promotes pacemaker cell specification of cardiac mesodermal cells derived from mouse and human embryonic stem cells. STEM CELLS 2019, 38, 352–368. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, Q.; Cattaneo, P.; Zhuang, S.; Gong, X.; Spann, N.J.; Jiang, C.; Cao, X.; Zhao, X.; Zhang, X.; et al. Transcription factor ISL1 is essential for pacemaker development and function. J. Clin. Investig. 2015, 125, 3256–3268. [Google Scholar] [CrossRef]
- Lindsey, S.E.; Butcher, J.T.; Yalcin, H.C. Mechanical regulation of cardiac development. Front. Physiol. 2014, 5, 318. [Google Scholar] [CrossRef]
- Liu, F.; Fang, Y.; Hou, X.; Yan, Y.; Xiao, H.; Zuo, D.; Wen, J.; Wang, L.; Zhou, Z.; Dang, X.; et al. Enrichment differentiation of human induced pluripotent stem cells into sinoatrial node-like cells by combined modulation of BMP, FGF, and RA signaling pathways. Stem Cell Res. Ther. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Liu, F.; Fang, Y.; Hou, X.; Yan, Y.; Xiao, H.; Zuo, D.; Wen, J.; Wang, L.; Zhou, Z.; Dang, X.; et al. Enrichment differentiation of human induced pluripotent stem cells into sinoatrial node-like cells by combined modulation of BMP, FGF, and RA signaling pathways. Stem Cell Res. Ther. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Luxan, G. , D’Amato, G., Pompa, J.L.d.l., Nakanishi, T., Markwald, R.R., Baldwin, H.S., Keller, B.B., Srivastava, D., Yamagishi, H., 2016. Intercellular Signaling in Cardiac Development and Disease: The NOTCH pathway, Etiology and Morphogenesis of Congenital Heart Disease: From Gene Function and Cellular Interaction to Morphology, 1 ed. Springer, Tokyo.
- Martinsen, B.J. Reference guide to the stages of chick heart embryology. Dev. Dyn. 2005, 233, 1217–1237. [Google Scholar] [CrossRef] [PubMed]
- McNally, E.M.; Svensson, E.C. Setting the pace: Tbx3 and Tbx18 in cardiac conduction system development. Circulation Research 2009, 104, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Meilhac, S.M.; Esner, M.; Kelly, R.G.; Nicolas, J.-F.; Buckingham, M.E. The Clonal Origin of Myocardial Cells in Different Regions of the Embryonic Mouse Heart. Dev. Cell 2004, 6, 685–698. [Google Scholar] [CrossRef]
- Minhas, R.; Loeffler-Wirth, H.; Siddiqui, Y.H.; Obrębski, T.; Vashisht, S.; Abu Nahia, K.; Paterek, A.; Brzozowska, A.; Bugajski, L.; Piwocka, K.; et al. Transcriptome profile of the sinoatrial ring reveals conserved and novel genetic programs of the zebrafish pacemaker. BMC Genom. 2021, 22, 1–15. [Google Scholar] [CrossRef]
- Minhas, R.; Paterek, A.; Łapiński, M.; Bazała, M.; Korzh, V.; Winata, C.L. A novel conserved enhancer at zebrafish zic3 and zic6 loci drives neural expression. Dev. Dyn. 2019, 248, 837–849. [Google Scholar] [CrossRef]
- Mohan, R. , Boukens, B.J., Christoffels, V.M., 2017. Lineages of the Cardiac Conduction System. Journal of Cardiovascular Development and Disease 4, 5.
- Molkentin, J.D.; Lin, Q.; Duncan, S.A.; Olson, E.N. Requirement of the transcription factor GATA4 for heart tube formation and ventral morphogenesis. Genes Dev. 1997, 11, 1061–1072. [Google Scholar] [CrossRef] [PubMed]
- Moorman, A.F.; de Jong, F.; Denyn, M.M.; Lamers, W.H. Development of the Cardiac Conduction System. Circ. Res. 1998, 82, 629–644. [Google Scholar] [CrossRef]
- Moskowitz, I.P.; Kim, J.B.; Moore, M.L.; Wolf, C.M.; Peterson, M.A.; Shendure, J.; Nobrega, M.A.; Yokota, Y.; Berul, C.; Izumo, S.; et al. A Molecular Pathway Including Id2, Tbx5, and Nkx2-5 Required for Cardiac Conduction System Development. Cell 2007, 129, 1365–1376. [Google Scholar] [CrossRef]
- Moskowitz, I.P.G.; Pizard, A.; Patel, V.V.; Bruneau, B.G.; Kim, J.B.; Kupershmidt, S.; Roden, D.; Berul, C.I.; Seidman, C.E.; Seidman, J.G. The T-Box transcription factor Tbx5 is required for the patterning and maturation of the murine cardiac conduction system. Development 2004, 131, 4107–4116. [Google Scholar] [CrossRef]
- Munshi, N.V. Gene Regulatory Networks in Cardiac Conduction System Development. Circ. Res. 2012, 110, 1525–1537. [Google Scholar] [CrossRef] [PubMed]
- Nakano, A.; Nakashima, Y.; Yanez, D.A.; Touma, M.; Nakano, H.; Jarodzewicz, A.; Jordan, M.C.; Pellegrini, M.; Roos, K.P. Abstract 13: Nkx2-5-notch Signaling Axis Regulates The Proliferation Of The Atrial Myocytes And Conduction System. Circ. Res. 2014, 115. [Google Scholar] [CrossRef]
- Pfeufer, A.; van Noord, C.; Marciante, K.D.; Arking, D.E.; Larson, M.G.; Smith, A.V.; Tarasov, K.V.; Müller, M.; Sotoodehnia, N.; Sinner, M.F.; et al. Genome-wide association study of PR interval. Nat. Genet. 2010, 42, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Puskaric, S.; Schmitteckert, S.; Mori, A.D.; Glaser, A.; Schneider, K.U.; Bruneau, B.G.; Blaschke, R.J.; Steinbeisser, H.; Rappold, G. Shox2 mediates Tbx5 activity by regulating Bmp4 in the pacemaker region of the developing heart. Hum. Mol. Genet. 2010, 19, 4625–4633. [Google Scholar] [CrossRef]
- Ramos-Mondragon, R. , Galindo, C.A., Avila, G., 2008. Role of TGF-β on cardiac structural and electrical remodeling. Vascular Health and Risk Management 4, 1289-1300.
- Ren, J.; Miao, D.; Li, Y.; Gao, R. Spotlight on Isl1: A Key Player in Cardiovascular Development and Diseases. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef]
- Rentschler, S.; Yen, A.H.; Lu, J.; Petrenko, N.B.; Lu, M.M.; Manderfield, L.J.; Patel, V.V.; Fishman, G.I.; Epstein, J.A. Myocardial Notch Signaling Reprograms Cardiomyocytes to a Conduction-Like Phenotype. Circulation 2012, 126, 1058–1066. [Google Scholar] [CrossRef]
- Schram, G. , Pourrier, M., Melnyk, P., Nattel, S., 2002. Differential distribution of cardiac ion channel expression as a basis for regional specialization in electrical function. Circ Res 90, 939-950.
- van Setten, J.; Brody, J.A.; Jamshidi, Y.; Swenson, B.R.; Butler, A.M.; Campbell, H.; Del Greco, F.M.; Evans, D.S.; Gibson, Q.; Gudbjartsson, D.F.; et al. PR interval genome-wide association meta-analysis identifies 50 loci associated with atrial and atrioventricular electrical activity. Nat. Commun. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Sotoodehnia, N.; Isaacs, A.; Bakker, P.I.W.d.; Dörr, M.; Newton-Cheh, C.; Nolte, I.M.; van der Harst, P.; Müller, M.; Eijgelsheim, M.; Alonso, A.; et al. Common variants in 22 loci are associated with QRS duration and cardiac ventricular conduction. Nat. Genet. 2010, 42, 1068–1076. [Google Scholar] [CrossRef]
- Tessadori, F.; van Weerd, J.H.; Burkhard, S.B.; Verkerk, A.O.; de Pater, E.; Boukens, B.J.; Vink, A.; Christoffels, V.M.; Bakkers, J. Identification and Functional Characterization of Cardiac Pacemaker Cells in Zebrafish. PLoS ONE 2012, 7, e47644. [Google Scholar] [CrossRef]
- van Eif, V.W.W.; Stefanovic, S.; van Duijvenboden, K.; Bakker, M.; Wakker, V.; Vries, C.d.G.-D.; Zaffran, S.; Verkerk, A.O.; Boukens, B.J.; Christoffels, V.M. Transcriptome analysis of mouse and human sinoatrial node cells reveals a conserved genetic program. Development 2019, 146. [Google Scholar] [CrossRef]
- van Weerd, J.H. , Christoffels, V.M., 2016. The formation and function of the cardiac conduction system. Development 143, 197-210.
- Vedantham, V. , Galang, G., Evangelista, M., Deo, R.C., Srivastava, D. RNA Sequencing of Mouse Sinoatrial Node Reveals an Upstream Regulatory Role for Islet-1 in Cardiac Pacemaker Cells. Circ. Res. 2015, 116, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Verweij, N. , Mateo Leach, I., van den Boogaard, M., van Veldhuisen, D.J., Christoffels, V.M., LifeLines Cohort, S., Hillege, H.L., van Gilst, W.H., Barnett, P., de Boer, R.A., van der Harst, P., 2014. Genetic determinants of P wave duration and PR segment. Circ Cardiovasc Genet 7, 475-481.
- Vincentz, J.W.; Toolan, K.P.; Zhang, W.; Firulli, A.B. Hand factor ablation causes defective left ventricular chamber development and compromised adult cardiac function. PLOS Genet. 2017, 13, e1006922. [Google Scholar] [CrossRef]
- Wang, J.; Greene, S.B.; Bonilla-Claudio, M.; Tao, Y.; Zhang, J.; Bai, Y.; Huang, Z.; Black, B.L.; Wang, F.; Martin, J.F. Bmp Signaling Regulates Myocardial Differentiation from Cardiac Progenitors Through a MicroRNA-Mediated Mechanism. Dev. Cell 2010, 19, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, P.; Jiang, L.; Wu, B.; Zhou, B. Control of sinus venous valve and sinoatrial node development by endocardial NOTCH1. Cardiovasc. Res. 2019, 116, 1473–1486. [Google Scholar] [CrossRef]
- Whitcomb, J.; Gharibeh, L.; Nemer, M. From embryogenesis to adulthood: Critical role for GATA factors in heart development and function. IUBMB Life 2019, 72, 53–67. [Google Scholar] [CrossRef]
- Wiese, C.; Grieskamp, T.; Airik, R.; Mommersteeg, M.T.M.; Gardiwal, A.; Vries, C.D.G.-D.; Schuster-Gossler, K.; Moorman, A.F.M.; Kispert, A.; Christoffels, V.M. Formation of the Sinus Node Head and Differentiation of Sinus Node Myocardium Are Independently Regulated by Tbx18 and Tbx3. Circ. Res. 2009, 104, 388–397. [Google Scholar] [CrossRef]
- Wittig, J.G.; Münsterberg, A. The Chicken as a Model Organism to Study Heart Development. Cold Spring Harb. Perspect. Biol. 2019, 12, a037218. [Google Scholar] [CrossRef]
- Wu, L.; Du, J.; Jing, X.; Yan, Y.; Deng, S.; Hao, Z.; She, Q. Bone morphogenetic protein 4 promotes the differentiation of Tbx18-positive epicardial progenitor cells to pacemaker-like cells. Exp. Ther. Med. 2019, 17, 2648–2656. [Google Scholar] [CrossRef]
- Xia, J.; Meng, Z.; Ruan, H.; Yin, W.; Xu, Y.; Zhang, T. Heart Development and Regeneration in Non-mammalian Model Organisms. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, F.; Shabaninejad, Z.; Vakili, S.; Derakhshan, M.; Movahedpour, A.; Dabiri, H.; Ghasemi, Y.; Mahjoubin-Tehran, M.; Nikoozadeh, A.; Savardashtaki, A.; et al. TGF-β and WNT signaling pathways in cardiac fibrosis: non-coding RNAs come into focus. Cell Commun. Signal. 2020, 18, 1–16. [Google Scholar] [CrossRef]
- Zheng, M.; Erhardt, S.; Ai, D.; Wang, J. Bmp Signaling Regulates Hand1 in a Dose-Dependent Manner during Heart Development. Int. J. Mol. Sci. 2021, 22, 9835. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yang, H.; Shi, J.; Zhang, M.; Yang, S.; Wang, N.; Sun, R.; Wang, Z.; Fei, J. Fate Tracing of Isl1+Cells in Adult Mouse Hearts under Physiological and Exercise Conditions. Int. J. Sports Med. 2019, 40, 921–930. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Cardiac contraction and histology of cardiac muscle. (A) Schematic representation of the components of the cardiac conduction system (CCS) in a human heart. In the inset picture, a cross-section of the human heart muscle is shown, with binucleated cardiomyocyte. The various components of the CCS (in green) are labelled: Sinoatrial node (SAN) located at the junction of the superior caval vein and right atrium, generates the impulse that then travels to the atrioventricular node (AVN). Propagation occurs through the left and right bundle branches of His-Purkinje leading to ventricular contraction. (B) An electrocardiogram representing the recording of the electrical activity of the heart. The upper chambers of the heart (atria) begin to beat when the first wave of the ECG, labelled P, appears. The lower chambers of the heart (ventricles) are represented by the QRS complex as electrical current flow. The electrical current spreads back over the ventricles in the opposite direction during the recovery phase, which is represented by the T wave.
Figure 1.
Cardiac contraction and histology of cardiac muscle. (A) Schematic representation of the components of the cardiac conduction system (CCS) in a human heart. In the inset picture, a cross-section of the human heart muscle is shown, with binucleated cardiomyocyte. The various components of the CCS (in green) are labelled: Sinoatrial node (SAN) located at the junction of the superior caval vein and right atrium, generates the impulse that then travels to the atrioventricular node (AVN). Propagation occurs through the left and right bundle branches of His-Purkinje leading to ventricular contraction. (B) An electrocardiogram representing the recording of the electrical activity of the heart. The upper chambers of the heart (atria) begin to beat when the first wave of the ECG, labelled P, appears. The lower chambers of the heart (ventricles) are represented by the QRS complex as electrical current flow. The electrical current spreads back over the ventricles in the opposite direction during the recovery phase, which is represented by the T wave.
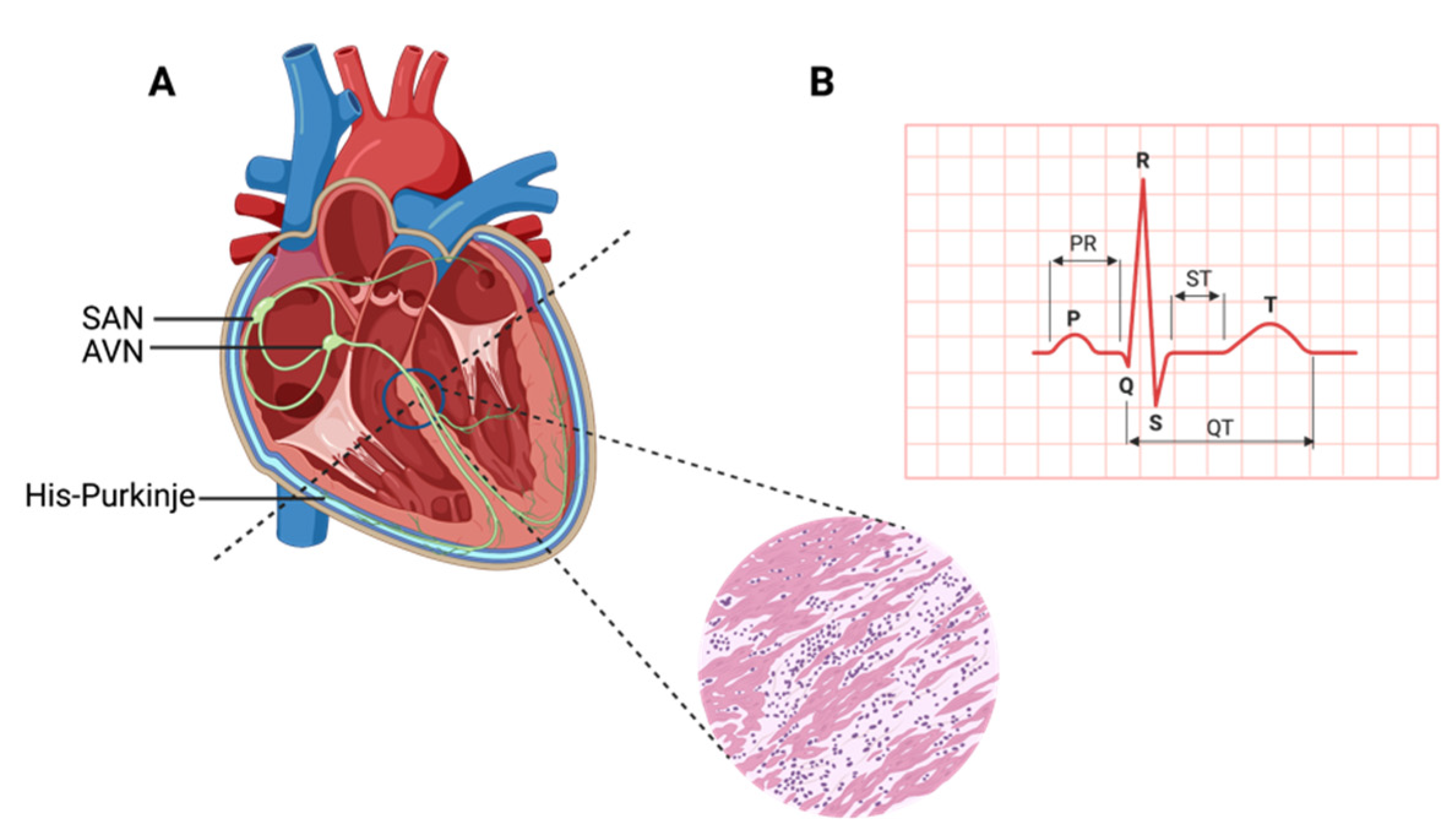
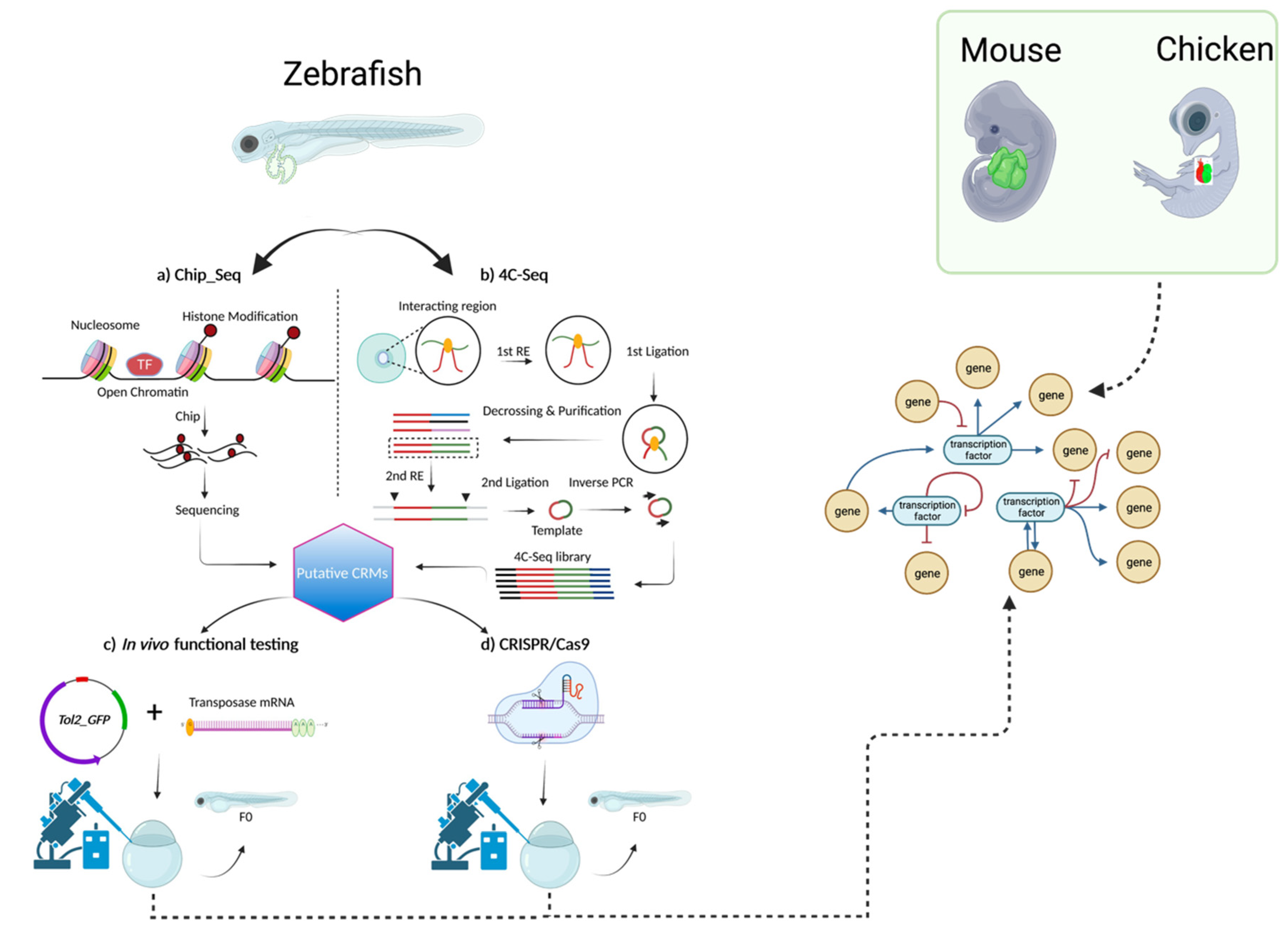
Figure 2.
Proposed methodology to further investigate CCS-specific enhancers leading to building an informative GRN. Schematic of methods that can be used to identify cis-regulatory modules (CRMs) using isolated hearts of various key developmental models (zebrafish, mice or chicken). For each model, in this case zebrafish is shown, various chromatin capture methods like ChIP-Seq or 4C-Seq can be employed to get a list of putative CRMs. These CRMs can be functionally tested and investigated further using genome-editing techniques like CRISPR/Cas9. A functionally validated GRN can then be generated by intersecting data obtained from several developmental models.
Figure 2.
Proposed methodology to further investigate CCS-specific enhancers leading to building an informative GRN. Schematic of methods that can be used to identify cis-regulatory modules (CRMs) using isolated hearts of various key developmental models (zebrafish, mice or chicken). For each model, in this case zebrafish is shown, various chromatin capture methods like ChIP-Seq or 4C-Seq can be employed to get a list of putative CRMs. These CRMs can be functionally tested and investigated further using genome-editing techniques like CRISPR/Cas9. A functionally validated GRN can then be generated by intersecting data obtained from several developmental models.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



