
Submitted:
04 July 2023
Posted:
10 July 2023
You are already at the latest version
Abstract
The aquaculture sector has become increasingly reliant on soybean meal (SBM) and soy-derived proteins, because of their large abundance, low price and favorable amino acid profile. However, for Atlantic salmon, a major aquaculture species, the inclusion of soybean meal, and soy protein concentrate (SPC) in certain combinations has adverse impacts on its gut health and welfare. This study evaluated two enhanced SBM targeting improved gut health for inclusion in freshwater phase salmon diets, enzyme pre-treatment (ETS) and addition of fructose oligosaccharide (USP), respectively in comparison to untreated soybean meal (US) and fish meal (FM). The impact on growth performance, gut microbiome, and behaviors relevant to welfare was investigated. Both enhancements of SBM were found to support comparable growth performance to FM. Both targeted treatments altered the gut microbiome community. Fish fed SBM showed a tendency for more reactive behavior compared to those fed the FM-based control. All fish had a similarly low response to elicited stress, although ETS fed fish responded more actively than those fed the US diet. SBM fed fish also had lower repeatability of behavior, which may have implications for welfare. Both enhancements provide a promising basis to optimize the application of this widely used protein source.
Keywords:
Soybean
; coping style
; welfare
; prebiotic
; non-starch polysaccharides
1. Introduction
In 21st century, Atlantic salmon (Salmo salar) has become one of the most intensively farmed finfish in the world and the industry continues to expand as consumer demand and global population grow [1]. Unlike many intensively farmed fin-fish, Atlantic salmon is a carnivore and has a high dietary protein demand [2]. Sourcing protein for inclusion in salmon aquafeeds has become a major challenge and bottleneck for the aquaculture sector [3]. Historically fish meal dominated the aquafeed market, but increasing and unsustainable pressure on wild-capture fisheries and rising prices have resulted in the commercial uptake of soy protein sources [3,4]. Soybean meal (SBM) gained initial popularity because it is highly available, cheap and has a favourable amino acid profile for farmed fish [5]. However, for Atlantic salmon, SBM can have adverse consequences to gut health, and triggers soybean meal induced enteritis (SBMIE) attributed to the presence of saponins, non-starch polysaccharides (NSP) and other anti-nutritional factors (ANFs) [6,7,8,9,10]. These ANFs can adversely alter the distal gut morphology of salmon [11,12], reduce nutrient absorption and efficient digestion [13], adversely alter the gut microbiome community [14] and consequently compromises the immune resilience and welfare of fish [15,16]. To address this problem, the salmon industry now primarily uses soy protein concentrate (SPC) an alcohol extracted soy product that reduces the inflammatory effect of SBM but has still been observed to have negative impact on gut health and gut microbiome of Atlantic salmon [17,18,19]. Applications of such treatments increase the cost of the aquafeed sector, thus there is a need to ensure tangible benefits for salmon production and welfare [20].
The gut microbiome of fish has been linked to key traditional measures of aquaculture productivity, including, nutrient digestibility and availability, growth performance [21], the metabolism, immune development, and disease resistance [22,23,24], the metabolism [25,26,27]. The composition of the gut microbiome is in turn influenced by the feed, host´s environment [28], and genetics [29,30]. In juvenile salmonids, the gut microbiome is particularly malleable to alterations in dietary protein sources [31,32] and this can influence the establishment and development of the gut microbiome into the adult developmental stage [33]. When salmonids are fed diets containing SBM or SPC, the composition of the gut microbiome is distinct from fish fed marine diets. There is a greater presence of bacteria associated with inflammation, and an imbalance in the community which has been linked to poor intestinal health [17,18]. Increasingly, aquaculture research is highlighting the potential of dietary treatments that can have functional benefits to the host by altering the gut microbiome. Such treatments include the application of probiotics, prebiotics, synbiotics, fermentation and enzyme-treatment of ingredients [34,35,36,37,38]. Prebiotics are non-digestible fibers that are not directly used by the host but act as a nutritional source to support the proliferation of desirable gut microbiota [39], they are widely used in human foods, terrestrial farming and have high acceptance for application to aquaculture diets [40]. In aquaculture, fructose oligosaccharides (FOS), and mannan oligosaccharides (MOS) have been used in feeds to provide gut microbiome-enhancing substrates and even growth performance [41,42]. Enzyme treatment has also become a popular processing method to improve the value of proteins for animal and aquaculture feeds [35]. Enzyme treatment of plant proteins such as SBM has been used to breakdown long-chain carbohydrates such as NSPs to improve their nutritional value for dietary inclusion [37,43]. This has a potential added benefit, that the breakdown product of long-chain carbohydrates includes shorter chain oligosaccharides, which in fish diets are not utilised by the host but may have a prebiotic value to the gut microbiome of the host [44].
There is a growing understanding of the role of gut microbiome not just in traditional measures of productivity for aquaculture but also in the observation of the bi-directional link between the gut microbiome and the central nervous system of animals including fish, which has been dubbed, the gut-brain-axis [45,46]. In fish there is evidence that the gut microbiome influences swimming behaviour [47], feeding behaviour [21] and social behaviour [48]. Behaviour is an early indicator of changes in environment [49]. Thus, monitoring behaviour is a valuable tool for managing welfare [50], a growing concern for the aquaculture sector [51,52].
In fish, five behavioural types relevant for welfare in aquaculture have been identified along five major axes: boldness-shyness, exploration-avoidance, sociability, activity, and aggression [53,54]. When such behavioural differences are consistent over time and context, they can be referred to as personality [55,56]. Boldness is a measure of predisposition to take risks [57] and individuals can be categorised on a continuum from bold to shy. Exploration is a measure of predisposition to engage with a novel environment or object [49] and individuals can be categorised on a continuum from active explorers to cautious explorers [58]. Coping style, closely related to personality, refers to a coherent set of behavioural and physiological stress responses, which is consistent over time and which is characteristic to a certain group of individuals [55]. This approach allows to classify the individuals of a given population into reactive and proactive categories. Proactive fish are bold individuals taking more risks and exploring their environment faster (less cautiously) when exposed to novelty [59,60] and they are more aggressive, dominant [60,61], and display low variability and flexibility in their behavioural responses with environmental changes [62]. By contrast, reactive fish are shyer individuals tending to be risk-averse and are generally neophobic, show higher behavioural flexibility and are more responsive to their environment [63], and are more sociable than proactive individuals [64]. If there is a correlation between different behaviour traits at the individual level, this is known as the behavioural syndrome which can be associated with welfare and freedom to express natural behaviour [51,54,65]. Behavioural traits can be altered by dietary ingredients making them important indicators to monitor [15,47,66]. In juvenile Senegalese sole (Solea senegalensis), replacing fish oil with vegetable oils resulted in significantly more reactive individuals [67]. It will be important to understand how dietary treatments affect the coping style and stress response of juvenile Atlantic salmon since it could have impacts on the productivity, fitness and welfare of farmed fish [53].
The objective of this study was to optimise the application of SBM in farmed Atlantic salmon diets and to elucidate the impact of novel enhancements of SBM (enzyme pre-treatment and addition of prebiotic respectively) on growth performance, gut microbiome, and behaviour. This study hypothesises that these novel enhancements will benefit salmonid aquaculture by supporting good growth performance, increasing the presence of desirable bacteria in the gastro-intestinal tract and improve fish welfare compared with unenhanced SBM. This study addresses important knowledge gaps for both the aquaculture industry and the field of applied microbiology in a commercially relevant model organism.
2. Materials and Methods
2.1. Experimental animals and study design
Atlantic salmon (Salmo salar) hatched by Stofnfiskur Ltd. (Vogar, Iceland) were incubated at 5.5°C and eyed eggs were transferred to Laxar ehf. (Kópavogur, Iceland). Fish were raised to first feeding using standard commercial techniques and start-feed diet BioMar Inicio-plus (United Kingdom) of 0.5mm pellet size and 12°C water temperature. Fry were transferred to Verid aquaculture station of Hólar University (Saudárkrókur, Iceland) where they were acclimated for 6 weeks prior to the start of the experiment. All fish within the experiment were individually weighed and measured under anaesthetic (2-phenyoxyethanol of 300ppm) following a 24-hour fasting period. Fish were split into 12 identical 20L-White cylindrical PVC tanks, in triplicate for each feed treatment. Each tank contained 40 fish with initial weight of 2.2±0.4g. Fish were maintained at 11±1.9°C under continuous light of 250±50 lux and 90±10% oxygen saturation. Fish were fed with experimental feed treatments for 70 days. The experiment was performed following Icelandic guidelines and within the permits and licenses of Verid aquaculture station.
2.2. Experimental feeds and feeding
A fish meal-based control (FM) diet, an untreated soybean meal (US) diet, an enzyme treated soybean meal (ETS), and an untreated soybean meal with added fructose oligosaccharide (USP) were used. The chemical composition of protein sources used in this study is presented in Table 1. For each of the soy diets, US, ETS and USP were included at 25%, replacing fish meal, the diet formulation and nutritional composition of each diet are presented in Table 2. The enzyme treated soybean for the ETS diet was produced by treatment with a 50:50 blend of Hostazyme X (main activity: Endo – 1,4 – β-xylanase, side enzyme activity: End – 1,4 -β-glucanase (cellulase), endo – 1,3(4) – β-glucanase, α-amylase, protease) and Hostazyme C (main activity; End-1,4 – β-glucanase, side enzyme activity: Endo – 1,4 – β-xylanase, endo – 1,3(4) – β-glucanase, α-amylase). The enzyme preparation was blended in 50°C water. The solution was added to 2L of water and 1kg of soybean meal and mixed for 15 minutes. The mix was then incubated for 3 hours at 50°C and manually mixed every 30 minutes. Following incubation, excess water was removed through pressing. The material was then further dried for 80°C. The diets were produced by cold pelletisation at Matís ohf. (Iceland). All dry ingredients were milled to homogenise particle size (IPHARMACHINE, Germany). Dry ingredients were homogenised in a standard food mixer (KitchenAid, USA) and the mix was milled again to improve the homogeneity of the feed. The dry mix was returned to the food mixer and fish oil was added and 200ml of water to produce the optimal consistency for processing in a pasta machine set to 0.5mm strings (ADE, Germany). Strings were dried in a commercial food dryer (Kreuzmayr, Austria) to <10% moisture content. During the 70-day feeding trial, fish were fed continuously by electric belt-feed with identical feed volumes at 15% excess fed based on the feed requirements for this developmental stage.
2.3. Growth performance
The fish were not fed for 24hours prior to measuring. After 70-days of feeding, all individual fish from each replicate tank within each treatment were anesthetised (2-phenoxyethanol 300ppm) and their wet weight (g) and total length (cm) measured. The specific growth rate (SGR) (%) over the study period was calculated: SGR = ((Ln(Final Weight)-Ln(Initial Weight)) x 100)/t, where t is the number of days of the experimental period. Mortality was monitored daily during the trial period.
2.4. Gut sampling
Following the final measurements of growth performance, all fish were left for one week to recover where they were maintained on their respective experimental diets. Fish were then fasted for 12-hours. Three fish per tank n = 9 per treatment were randomly selected for gut microbiome analysis. Fish were humanely euthanised with a lethal dose of anaesthetic (2-phenoxyethanol 600ppm). The outside of the fish was washed in 90% ethanol followed by sterile distilled water. The gastro-intestinal (GI) tract and contents (from the start of the mid-gut, just below the pyloric caeca to the end of the distal gut) was sampled under sterile conditions and directly frozen at -80°C.
2.5. DNA extraction, PCR amplification and sequencing
GI tract samples were individually manually homogenised in a sterile petri dish with a sterile scalpel to physically break up the material. Sample material was transferred to a sterile 2ml Eppendorf tube with 300µl of sterile 1mm diameter silica beads (BioSpec Products, United States) and 800µl of CD1 solution from the QIAamp PowerFecal Pro DNA kit (QIAGEN, Germany). Samples were vortexed for 5 seconds and shaken at maximum speed (30Hz) in a laboratory mixer mill (Retsch MM400) for 1 minute. The supernatant (~800µl) was transferred to the PowerBead Pro tube from the QIAGEN QIAamp PowerFecal Pro DNA kit. The protocol for this DNA extraction kit was then followed and final DNA was eluted with 80µl of C6 solution. A DNA negative (no material added) was also run to ensure no contamination occurred during DNA extraction. DNA concentration was measured for each sample with 2µl with Invitrogen Qubit dsDNA BR Assay kit (Invitrogen, Carlsbad, CA, USA). DNA was diluted to 4ng µl-1 in 50µl aliquots. Samples were subjected to PCR of the V3-V4 regions of the 16S rRNA gene with a universal bacterial primer pair S-D-Bact-0341-b-S-17 (5′−CCTACGGGNGGCWGCAG-3′)/S-D-Bact-0785-a-A-21(5′-GACT- ACHVGGGTATCTAATCC-3′) (Klindworth et al., 2013). PCR mater mix included diluted DNA, nuclease-free water, Q5 High Fidelity DNA polymerase (New England Biolabs, Ipswich, USA), Q5 GC Enhancer, 0.5µM of each primer containing Illumina overhand adapters, and 1 x Q5 Reaction buffer, 200µM dNTPs (New England Biolabs, Ipswich, USA). Both positive and negative samples were also run in the PCR to monitor for absence of contamination and successful amplification. The thermocycling protocol had an initial denaturation step (90°C for 30s), followed by 35 cycles of: denaturation (90°C for 10s), annealing (52°C for 30s), and extension step (72°C for 30s), with a final extension (72°C for 2 min). Libraries were multiplexed with Nextera XT v2 barcodes (Illumina, USA), normalised using Sequel-Prep normalisation places (ThermoFisher Scientific, USA) and sequenced on MiSeq desktop sequencer (Illumina, USA) using v3 chemistry and 2 x 300 cycles.
2.6. Behaviour
Following gut microbiome sampling, eight (8) fish from the remaining tank population were randomly selected to be uniquely tagged with Visible Implant Elastomer (VIE) colour tags (Leblanc and Noakes, 2012) under anaesthetic (2-phenoxyethanol 300ppm). Each fish was injected parallel to the dorsal fin, below the surface skin layer with two 1cm strips of colour with unique combinations so each of the 8 selected fish per tank (n=24 per treatment) could be identified and followed through all behavioural observations. All remaining untagged fish per tank were retained in tank to maintain a consistent stocking density but were not included in behavioural observations. Each tagged fish was observed in two different behaviour contexts (a swimming test and an open field test with shelter) and each of these tests was performed two times. There was always a one-week recovery period between each behavioural observation. Behaviour observations were performed in a dedicated room to minimise external disturbance. Fish were individually tested in all behavioural observations. Behaviour was recorded using a monochrome camera (Basler Ace acA1920-150um, Germany) with a frame rate of 30Hz and resolution of 1280 x 1024 pixels placed 112cm (swimming test arena) and 110cm (open-field test) above the water level of the respective behavioural arena. Video recordings were analysed with EthoVision XT software (Noldus, The Netherlands), which was used to track fish in each arena. The VIE tags were recorded for each fish after each behaviour observation so their individual data could be tracked across the different time and contexts of the behavioural observations.
2.6.1. Swimming test for assessing response to stress and exploration.
A swimming test was used to collect behaviour observation data for both stress response and exploration-avoidance axis. The apparatus for assessing these traits consisted of 4 circular arenas (diameter = 25cm, water depth 7.8cm (4L), height= 15cm). The arena was illuminated from below to provide uniform light intensity (260 lux) (Figure 1). Tagged fish from each tank were collected from the trial tank and transferred in identical white transfer buckets with closed tops and then placed into individual arenas in random order. All four arenas were filmed simultaneously and continuously for 20 minutes. For the purposes of this analysis, this was separated into four virtual periods. The 0-5 minutes was the acclimation period (AC), 5-10 minutes was the normal swimming period (NS). At the 10-minute mark a stress was elicited (bottom lighting was switched off for 3 seconds and then switched back on again), minutes 10-15 was thus the post light stress period (PLS), and 15-20 minutes was the recovery period (RC). Behaviour was not recorded while the light was off.
Variables collected through the swimming tests were, mean distance from centre-point (cm): the distance of the centre-point of the fish body from arena centre (DisCent). Total distance moved (cm): the distance travelled by a fish measured from the centre-point of body between consecutive X-Y coordinates (TotDis). Mean velocity (BL s-1): the distance moved by the centre-point of body per unit time between consecutive X-Y coordinates normalised to body lengths per second (Vel). Absolute angular velocity (° s-1): expressed in degrees per second was calculated by EthoVision XT software. Vangn = RTAn/tn – (tn-1), where RTAn represents the relative turn angle for sample n, and tn-(tn-1) is the time difference between the current and previous sample (AngVel).). Mobility state, the cumulative duration for which the fish body is changing, highly mobile (s) when cumulative duration is 60% (HiMob). Moderate mobile (s) when cumulative duration was between 20-60% (MedMob). Immobile (s) when cumulative duration was below 20% (Immob).
2.6.2. Open-field test for assessing boldness
An open-field test (OFT) with a shelter was used to assess boldness [68,69,70]. The apparatus for assessing this trait consisted of 4 rectangular arenas (40 x 30 x 25 cm), water depth 6cm (7L) with an identical shelter placed in the bottom-right of each arena (14 x 6.5 x 6.5 cm). The arenas were illuminated from below to provide uniform light intensity (260 lux) (Figure 2A). Tagged fish from each tank were collected from the trial tank and transferred in identical white transfer buckets with closed tops and then placed in random order into individual arena shelters through a top compartment (4cm diameter) which was then closed, and the main shelter door (a sliding opaque trapdoor) was kept shut, closing off access to the arena. All four arenas were filmed simultaneously and continuously for 25 minutes. The first 5 minutes the fish remained shut inside the shelter acting as an acclimation period, after which the door to each arena shelter was lifted simultaneously and filming continued for a further 20 minutes uninterrupted. Each individual arena was virtually divided into four zones using the EthoVision XT software. These zones were called Shelter, Entry, Border and Centre (Figure 2B). The centre zone is considered high-risk and staying close to the border edges of a space is considered more cautious and an indicator of a shyer individual [57,71]. Variables used to characterise OFT behaviour for the trait of boldness were as follows: Latency (s) to emerge-time taken to exit the shelter (Lat). Time spent in each zone (s) (Shelter, Entry, Border, Centre) respectively (Shelt, Ent, Bord, Cent). Mean distance from shelter (cm) (DisShelt). Number of returns to shelter (Ret). Cumulative duration spent highly mobile (s) (HiMob). Distance moved: the distance travelled by a fish measured from the centre-point of body between consecutive X-Y coordinates. Mean velocity: the distance moved by the centre-point of body per unit time between consecutive X-Y coordinates normalised to body lengths per second. Absolute angular velocity: expressed in degrees per second.
2.7. Statistical methods
Statistical analyses were performed in Rstudio version 3.6.1 (2019-07-05). Tests for growth performance, gut microbiome and behavioural characteristics were two-tailed with a significance level set to α = 0.05. We fitted multivariate linear mixed models to estimate the repeatabilities and the correlations between the personality estimates using Bayesian statistical modelling.
2.7.1. Growth performance
SGR (%) was used to assess growth performance across the different dietary treatments in this study. A Linear Mixed Model (LMM) with the package lme4 [72] was selected defining feed treatment (FM, US, ETS, USP) as a fixed factor and Tank as a random nested factor of feed treatment. Since the random nested factor of tank did not cause significant variation in SGR (%), a simplified statistical test was adopted. A Linear Model (LM) with the package nmle [73] was used, where feed treatment was a fixed factor and the model residuals had a normal distribution. A Tukey post-hoc test was applied to assess pairwise differences between feed treatments.
2.7.2. Gut microbiome
To assess the gut microbiome of fish fed different dietary treatments in this study, demultiplexed FASTQ files from Illumina were processed to produce amplicon sequence variants (ASVs) with DADA2 package version 1.16.0 [74]. The filterAndTrim variables used were as follows: truncLen=c(280,250), trimLeft=21, maxN=0, maxEE=c(2,2), truncQ=2. Taxonomy was assigned to ASVs from version 138 of the SILVA database and the function assignTaxonomy [75]. The R packages, phyloseq [76], microbiome [77] and vegan [78] were used to analyse the microbiome community and ggplot2 was sued to visualise key data [79]. The average read number output from the DADA2 pipeline was 20643±12702, and two samples were removed as they did not contain any reads after processing. Five PCR negative samples were also sequenced to control for any contamination that may have occurred during sample amplification. These controls were used to remove suspected contamination from samples using the decontam package with prevalence method and threshold of 0.5 [80]. Read depth was normalised across all samples using the function rarefy_even_depth. Raw 16S rRNA gene amplicon reads can be found in the Sequence Read Archive following submission.
Alpha and beta diversity indices were used to quantitatively analyse the gut microbiome. Alpha diversity measures selected were, observed richness of AVSs, Shannon diversity, Chao1 diversity and Pielou´s evenness. A GLMM was used to assess if there was a significant difference in the alpha diversity measures between the different feed treatments in this study. In this model, dietary treatment was a fixed factor and tank was a random nested factor of feed treatment (which was tested by a Likelihood Ratio Test [81]). A Tukey test was applied for post-hoc testing. Beta diversity was assessed by transforming microbiome community data using a Bray-Curtis dissimilarity matrix. An Analysis of Similarity (ANOSIM) was performed to determine if there was any significant difference between and within the gut microbiome community of fish fed the different diets. Relative abundance of taxa at the phylum level and genera level as a proportion was calculated. Taxa present in the community at <1% relative abundance were grouped into a category labelled “Other”.
2.7.3. Behavioural characteristics
To assess stress response, all swimming test variables were analysed using Linear mixed effect models (LMM). For each analysis, the explanatory variables included in the full model were, feed treatment (FM, US, ETS, USP), period (AC, NS, PLS, RC), and TL, and interactions between feed treatment and period were considered. Random effects considered in the model were trial replicate number (1, 2), and Tank (1,2,3) nested in feed treatment, and individual ID.
To assess the behaviour trait of exploration, TotDis recorded for AC and NS only, was used as a proxy [82]. Both traits i.e., boldness and exploration were then compared between feed treatments in two respective LMM models where boldness score or TotDis were the response variables. The full models were reduced by backward selection based on the Akaike Information Criterion (AIC) [83]. Diagnostics based on residuals of the model were performed to assess the adequacy of the reduced model and compliance with the underlying assumptions. Dependent variables were transformed whenever necessary to ensure that the residuals followed the assumed error distribution. Finally, the effects of the independent variables were estimated from the reduced models and their significance was tested by likelihood ratio tests (LRT) between models respecting marginality of the effects that are supposed to follow a χ2 distribution under the null hypothesis (type II tests; [81]). This analysis was followed by a post-hoc multiple comparison test [84] to assess pairwise differences between models.
2.7.4. Repeatability of behaviour traits and correlations
A Bayesian multivariate linear model using Stan [85] was run using the ‘brms’ package [86]. We used one model for each feed treatment (four models in total). Each model simultaneously regressed each dependent variable (i.e., boldness score and TotDis) against a set of fixed factors (Tank, Trial, SGR and TL). The random-effects structure included individual fish identity (ID) as a grouping variable, allowing us to calculate the repeatability of Boldness and TotDis as the ratio of the among individual variance and the sum of the among individual and residual-level variances (i.e., personality) [87]. Fish that did not exit the shelter for at least one of two the trial were removed from the analysis. Moreover, the model estimated covariances between Boldness and TotDis at both the ID and residual levels. The among individual covariance quantified the degree to which Boldness and TotDis was correlated among individuals across multiple trials (i.e., behavioural syndrome). The model was run for 64000 iterations (32000 for warmup and 32000 for sampling), four chains, adapt delta was set to 0.9, the max tree depth to 40, and All other parameters were set to their defaults. Convergence was assessed using the standard diagnostics provided by Stan, including the potential scale reduction factor (̂R), effective sample size, and visual inspection of trace plots and histograms for each model parameter. Unless otherwise noted, we used posterior modes for point estimates and higher posterior density with 95 % coverage for uncertainty intervals (UI95 %), respectively calculated using the map_estimate and hdi functions from the “coda” package [88].
3. Results
3.1. Growth performance
There was a significant difference in SGR % between dietary treatments in this study (Figure 3). Fish fed FM had significantly higher SGR% than those fed US. Fish fed the enhanced soy diets, ETS and USP had slightly higher SGR% than those fed the unenhanced, US diets, but no significant differences were found between fish fed the FM and US diets.
3.2. Gut microbiome
There was a significant difference in the gut microbiome community alpha diversity measures for Shannon diversity (Figure 4B) and for Pielou evenness (Figure 4D) but not for the observed richness of ASVs (Figure 4A) or Chao1 diversity (Figure 4C) between the dietary treatments. For both Shannon diversity and Pielou´s evenness, the highest community diversity and evenness values were observed for the FM diet, which was significantly higher than the values for all other treatments, followed by the unenhanced US diet which had comparable Shannon diversity and evenness to the enhanced USP fed fish. Fish fed the enhanced ETS diet had significantly lower gut microbiome community Shannon diversity and evenness than fish fed any other diet.
The gut microbiome communities were significantly different between dietary treatments. There was a much greater similarity between individual fish from the same treatment than between individuals from different treatments (ANOSIM P = 0.001, R=0.84). The NMDS plot (Figure 5) shows distinct clustering by feed treatment.
At the phylum level, for all dietary treatments, the phyla with the largest proportion presence were Firmicutes (FM=0.81±0.1, US=0.93±0.03, ETS=0.99±0.01, USP=0.95±0.03), followed by lower proportions of Actinobacteria (FM=0.1±0.06, US=0.05±0.03, ETS=0.01±0.01, USP=0.03±0.01) and Proteobacteria (FM=0.09±0.12, US=0.01±0.01, ETS=0.01±0.01, USP=0.02±0.03) which were at similar levels.
At the taxonomic level of genus, there were nine genera with a relative abundance >1% of the community (Figure 6). All other genera were present at very low relative abundance. The genera composition between the FM dietary treatment and the three dietary treatments containing SBM showed distinct difference. The genera, Anaerosalibacter, Clostridium_sensu_stricto_18, Clostridium_sensu_stricto_7, Hathewaya and Peptosteptococcus all had a greater relative abundance in FM fed fish gut microbiome than US, ETS or USP fed fish gut microbiomes. In all these genera, however, there was a trend that relative abundance was higher in US and USP than the proportion observed in ETS fish gut microbiome. The LABs, Leuconostoc and Weissella had the lowest relative abundance in FM fish gut microbiomes, slightly higher in the ETS fish gut microbiomes and highest in the US and USP fish gut microbiomes. For the LAB Enterococcus there was a different trend, the relative abundance was very high in the ETS fish gut microbiome, low but present in the FM and USP fish gut microbiome and absent in the US fish gut microbiome. Staphylococcus was observed in similar relative abundance for FM, US and USP fish gut microbiomes but much lower in ETS fish gut microbiome.
3.3. Behavioural characteristics
3.3.1. Swimming test
There was no significant difference in any of the swimming variables between any of the dietary treatments. There was a significant difference between periods for all variables (Table S1: Swimming Activity) but this did not appear to be driven by the elicited light stress but instead by time spent in the arena. The interaction between feed treatment and period was significant for AngVel (° s-1) for USP feed during PLS period (χ2=17.53, df=9, p=0.03), Vel and it was significant for US feed during RC period (χ2=17.54, df=9, p=0.041). There was a significant effect of swimming test replicate (1,2) for TotDis (cm) and DisCent (cm). For all variables there was a significant effect of TL (cm), for the variables DisCent (cm), TotDis (cm), and Vel (BL s-1) i.e., there was an inverse relationship with TL (cm) (larger fish had lower values for these variables). For AngVel there was a positive relationship with TL (cm) (larger fish had higher values for these variables).
3.3.2. Exploration trait mean value
TotDis (proxy for exploration) did not significantly vary between feed treatments (χ2 = 0.46, df = 3, p = 0.93, Figure 7) but there was a highly significant effect of TL (χ2 = 71.4, df = 1, p <0.0001) i.e., the higher TL, the lower TotDis.
3.3.3. Boldness trait mean value
PC1 explained 48% of variation in the data. For the PC1 , a high value indicates a greater total distance travelled (cm) (loading = 0.43), higher swimming velocity (BL s-1) (loading = 0.41), greater number of returns to the shelter (loading = 0.3), greater time spent in the entry zone (s) (loading = 0.12), greater time spent in the border zone (s) (loading = 0.41), greater time in the center zone (s) (loading = 0.21), and higher mobility (s) (loading = 0.31). A low value indicates, greater time spent in the shelter (s) (loading = -0.35), Latency to exit shelter (s) (loading = -0.25) and greater absolute angular velocity (° s-1) (loading = -0.21). PC1 therefore presents a gradient from shyer (low values) to bolder (high values) and was used hereafter as proxy for boldness score. There was no significant difference in the boldness score between feed treatments, although there was a trend visible showing FM fed fish to be bolder than fish fed any of the soy diets, and the US fed fish to be shyer than any other fish (Figure 8). Total length (cm) was significant (P=<0.001, df=1, χ²=31.92, S.E =0.11, Estimate= -0.6).The random factor test repeat number was not significant, but the random factors individual ID and tank were significant.
3.3.4. Repeatability and correlation of boldness and exploration traits
Among individual variances of both boldness and exploration traits were unambiguously different from zero in all diet treatments but US and USP for boldness (Table 3) , indicating individual repeatability in these behaviours. Highest repeatability for boldness trait was reported in the FM diet, and in USP for exploration. The large confidence interval in the US for exploration shows a high degree of uncertainty of the repeatability estimate (Table 3). The repeatability of these traits was still high in the ETS diet but much lower for boldness trait and with a large confidence interval (Table 3). The covariance of boldness and exploration for each diet treatment but the ETS was close to zero with UI95% strongly overlapping zero (Figure 9). In the ETS treatment neither ID nor residuals overlapped with 0 showing a positive correlation between boldness and exploration (Figure 9).
4. Discussion
The objective of the present multi-disciplinary study was to optimise the application of soybean meal (SBM) in formulated aquafeeds for salmonids. This study reports the impact of two novel enhancements of SBM, an enzyme pre-treated SBM (ETS), and a SBM with addition of the prebiotic (fructose oligosaccharides, FOS) (USP) compared with an untreated SBM (US) and a fish meal control (FM) on the growth performance, gut microbiome, and individual behaviour traits relevant to the culture of juvenile Atlantic salmon (Salmo salar).
The growth performance of Atlantic salmon in this study across all treatments was within a normal range for the freshwater stage at the experiment water temperature [89,90]. The findings were consistent with the existing literature, showing that FM diets support higher growth rates than untreated SBM for Atlantic salmon, both at inclusion levels of 20% SBM [12], and at higher levels of 31% SBM [91]. However, salmon parr fed diets containing 16.7% SMB [92] or 40% SBM [93] performed comparably to those fed a FM diets. In seawater phase, inclusion levels of 20% SBM and higher show reduced growth rate in Atlantic salmon compared to those fed FM diets [11], although inconsistencies in the literature exist [94]. The two novel enhancements of SBM applied in this study supported comparable growth performance to FM, suggesting both enzyme pre-treatment and addition of FOS have potential benefits for improving SBM application for growth performance of salmon during the freshwater stage. Similar improvements have also been seen with SBM treated to reduce the content of oligosaccharides and ANFs compared with an untreated SBM at a 40% crude protein supply [95]. In Japanese seabass (Lateolabrax japonicus) NSP enzymes applied to feed also had beneficial effects on growth performance [96] and in the white-spotted snapper (Lutjanus stellatus) NSP targeted enzyme treatment of Gracilaria lemaneiformis growth performance was also enhanced [97]. On the contrary, [98] found no growth performance benefits of NSP-targeted enzyme treatment of SBM for Atlantic salmon. Furthermore, phytase pre-treatment on SPC replacing 60% of FM in SW Atlantic salmon did not improve growth performance [98]. The addition of FOS to diets improved growth performance of Rainbow trout (Oncorhynchus mykiss) [99], however, when FOS was added to FM diets for Atlantic salmon, no effect on growth performance was observed [42].
The differences in the gut microbiome community assemblage between fish fed different diets have been observed in many studies with salmonids, between FM and SBM [30,32] and between different treatments of conventional protein sources [18]. Other studies have also found FM based diets to support greater diversity and community evenness in fish gut microbiome compared with other protein sources [100,101]. The findings of this study that USP fed fish had comparable diversity to US, and that ETS fed fish had lower diversity than all other treatments, were similar to what was found in white sea bream (Diplodus sargus) fed FOS [102]. However, research has shown increased gut microbiome diversity with the additions of prebiotics for juvenile hybrid Tilapia (Oreochromis niloticus♀ × Oreochromis aureus ♂) [103]. The lack of difference in the present study may be driven by the need for longer term continuous administration to detect positive gut health impacts of feed additives in fish [104]. It has been noted that diets with SBM which contains natural oligosaccharide sources may mask any beneficial effects of additional prebiotics [102,105]. The impact of low values for diversity and evenness found for ETS fed fish will be important to elucidate, since high gut microbiome community evenness and diversity values have been associated with improved productivity in aquaculture [106]. In the present study, it seems that this lower diversity and evenness is driven by a dominance of the lactic acid bacteria (LAB) Enterococcus, a genus that has been associated with growth and immune benefits to fish [107]. Because the presence of this genus has not excluded other taxa establishing in the gut, reduction in evenness and diversity may not adversely affect fish gut health. Although community dominance at such an early development stage should be monitored to ensure that an undesirable dysbiosis does not establish itself [21]. This study suggests that the application of FOS has quite different impacts than the enzyme-treatment at the genus taxonomic level, both may be having a prebiotic effect, but stimulate the growth of different communities. The increased presence of the LABs in SBM diets compared to FM is consistent with the existing literature for salmonids [30], as is the addition of prebiotic ingredients increasing levels of LAB in the common carp (Cyprinus carpio) [108]. LABs have been associated with improved digestive function, improved gut health, and disease resistance [38,109,110]. Since the levels are lower for these two taxa in the ETS fed fish, it suggests that the enzyme treatment may have broken down part of the SBM favoured by these two taxa, a phenomenon that warrants further investigation. The greater presence of Clostridium_senu_stricto_7, Clostridium_senu_stricto_18 and Peptostreptococcus in FM fed fish have been found in faster growing individual Rainbow trout [111], which is consistent with the growth performance findings for this trial. These bacteria have been linked to fermentation of different amino acids [112], suggesting their growth may be facilitated by amino acids present in FM that are not present or less present in SBM. There is a prevalence of Hathewaya and Anaerosalibacter in FM fed fish compared with SBM fed fish in the study, these taxa have been found in Chinook salmon (Oncorhynchus tshawytscha) gut microbiomes associated with presence in feed [113] suggesting an association with FM. There is a need for future research to address the functional role of LAB for Atlantic salmon and to establish taxa associated with growth performance and gut health benefits to improve the application of feed additives and dietary enhancements.
This study is the first to investigate behavioural traits relevant to welfare related to diets containing different protein sources, and enhancements of dietary protein for Atlantic salmon. The light stress did not appear to produce a response in the fish tested, suggesting the need for a different stress source, as the fish may have been habituated to light stress as they were kept under continuous light [114]. It is also possible that broodstock selection and process of domestication of Atlantic salmon genetic lines could already have selected for more proactive styles [62,115] which may also explain why there was low responsiveness to the elicited stress. While the scores of the traits of exploration and boldness behaviour traits did not differ significantly between the diets in this study, there was an interesting trend that FM fed fish displayed a more proactive coping strategy i.e., higher boldness and exploration values than fish fed any of the SBM diets. For ETS fed fish, there was indication of more proactive-type behaviour compared to US fed fish. This is consistent with results from juvenile Rainbow trout, where fish fed a plant-based diet, exhibit an increase in apathetic behaviour and an increased stress response (both traits of reactive coping styles [53] compared to a marine diet [15]. This goes along with other results in juvenile Senegalese sole (Solea senegalensis) individuals fed diets enriched with fish oil from Cod liver, compared with vegetable oils from Linseed, Soybean and Olive, that were more proactive [116].
For boldness, the repeatability was highest in FM fed fish showing the existence of personality trait which could indicate that they may be more suited with the aquaculture environment. However, regarding the exploration, high repeatability was shown in all treatments, which could indicate lesser impact of the diet treatment on this trait i.e., highly canalised trait [117]. Existing studies have indicated that more proactive individuals do show a higher degree of repeatability for behaviour traits which matches well with the findings of the present study. Interestingly, there was a positive link between growth and exploration in the FM treatment only which is in accordance with previous studies showing higher fitness in proactive fish e.g., higher reproductive success in gilthead seabream (Sparus aurata) [67], higher growth rates for salmon [118], faster feeding recovery after stress [119] and lesser sensitivity to environmental stress [120] for Rainbow trout, all of which may make them better suited to the intensive farming environment of salmonid aquaculture [115] as these individuals may be more productive and also experience better welfare. Repeatability of both traits was also high in the ETS which could indicate a benefit of the enzyme treatment whereas the US (and the USP to a lesser extent) showed no evidence for any pronounced personality trait. This could indicate an impact of these diets on welfare. In summary, high level of repeatability of both traits may show consistent freedom to express natural behaviour which may indicate a good welfare condition [65,115].
In the present study a clear behavioural syndrome was detected only for ETS but surprisingly not for FM which may indicate the fish fed ETS experiencing different conditions or pressures than the fish fed other diets [58]. Domestication of salmon may also reduce the presence of behavioural syndromes. In a study of urban song sparrows no correlation between traits of boldness and aggression could be found, whereas there was a correlation, and thus behaviour syndrome in their wild counterparts [121].This also may suggest that the behaviour of US fed fish in this study was more impacted by the culture conditions and have adjusted by becoming more behaviourally plastic or showing adapted behaviours to cope [58]. There is a need for further research attention on the impact of dietary proteins and treatments on behaviour and welfare for farmed Atlantic salmon to support the optimisation of existing feed materials and the industry uptake of alternative proteins. This study also highlights the need for greater understanding on how domestication may alter salmon behaviour and therefore welfare in intensive farms which will help inform selection programs to optimise coping styles in farmed fish [122,123].
Although, this study did not specifically address the link between gut-microbiome and behaviour at the individual level, our results suggest potential impacts of diets on the gut microbiome that in turn influence behaviour. Indeed, fish fed SBM diets displayed an altered gut-microbiome, lower boldness repeatability and tended to have lower mean boldness values. Studies have shown that manipulating fish microbiome can modulate the behaviour via the gut-brain axis [47] and a recent study on farmed Arctic charr (Salvelinus alpinus) [124] showed evidence of bacterial strains influencing the response to stress and growth where fish fed with E. thailandicus 04-394 and L.brevis ISCAR-07433 displayed an increase in motility and slower growth which can be interpreted as lower stress coping ability. Using personality has been suggested to be a valuable tool to reduce chronic stress in captive fish where bolder individuals are usually more resistant to chronic stress [125]. Therefore, the gut alteration by SBM diets could directly impact boldness in Atlantic salmon, which in turn could affect reaction to stress and ultimately the growth. In this study, FM fed fish were the treatment group that grew the fastest, had a distinctly different gut microbiome community visible at the taxonomic level and tended toward more proactive behaviour and potentially better welfare compared to any of the SBM treatment. Conversely, the US fish differed the most from the FM fish in all aspects of this multi-discipline approach, showing poorer growth, a lower diversity and evenness of the gut microbiome community and behaviours that indicated differing impact compared with any other diet. While enhancements of SBM ameliorated some of these traits, there were still difference with FM fish, highlighting the importance of such multi-disciplinary studies to elucidate the impact both to traditional productivity measures for aquaculture but also to measures that explore indirect impacts of nutrition on immune development and welfare. Future studies should aim to establish functional links between these different fields to optimise Atlantic salmon aquaculture. This will be especially important as new feed enhancements and alternative proteins enter the market to ensure they are safe and effective for the aquaculture sector and to support optimal broodstock selection programs.
5. Conclusion
The results of this multi-disciplinary study suggest that the SBM enhancements tested provide growth performance benefits compared to unenhanced SBM, alter the gut microbiome community assemblage compared to both unenhanced SBM and FM and in the case of enzyme pre-treatment, support a strong dominance of the LAB Enterococcus. This study demonstrates for the first time the impact of SBM on juvenile Atlantic salmon behaviour traits, showing there is a trend that a plant-based diet may increase reactive coping styles, which may have adverse consequences for welfare of fish in intensive farm systems. The two enhancements of SBM offer some improvements on unenhanced SBM which provide a promising basis for continued investigation to optimise the application of this widely used protein source. Future studies should attempt to elucidate the functional link between nutritional composition of protein sources, composition of the gut microbiome and their influence on behaviour traits relevant to the aquaculture sector.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Table S1: Swimming Activity.
Author Contributions
“Funding Acquisition, Methodology and Conceptualization, A.L, D.B, W.K, and M.Ø.; Project administration A.L.; investigation and formal analysis, A.L, D.B, C.S, S.K, S.S, B.B and G.L.; writing—original draft preparation, A.L, D.B, M.Ø, and C.S.; writing—review and editing, A.L, D.B, MØ; W.P, R.B, and G.L.; supervision, D.B, M.Ø, W.P and R.B.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by AVS – Research Fund, grant number 62497 for the project “Enhanced soy protein for salmonid diets”.
Institutional Review Board Statement
“The study was conducted in accordance with the regulations of the Icelandic Food and Veterinary Authority and the Icelandic Environment Agency and by the Hólar University ethical committee represented by David Benhaïm.
Data Availability Statement
Gut microbiome data is available as a BioProject on the Sequence Read Archive.
Acknowledgments
The authors of this study would like to acknowledge the valuable advice and support of the Verid, Iceland Aquaculture husbandry team and Dr. Camille Leblanc for value support during the project development. The authors would also like to acknowledge the support and facilities of Matís microbiology and nutritional laboratory teams. The authors would further like to thank the NMBU Biovit departments for valuable feedback on the development of this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1. FAO The State of World Fisheries and Aquaculture. Sustain. action. Rome. 2020. [CrossRef]
- National Research Council (NRC) Nutrient Requirements of Fish. National Academy Press, Washington DC. 2011.
- Naylor, R.L.; Hardy, R.W.; Bureau, D.P.; Chiu, A.; Elliott, M.; Farrell, A.P.; Forster, I.; Gatlin, D.M.; Goldburg, R.J.; Hua, K.; et al. Feeding Aquaculture in an Era of Finite Resources. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 15103–15110. [Google Scholar] [CrossRef]
- Tacon, A.G.J.; Metian, M. Feed Matters: Satisfying the Feed Demand of Aquaculture. Rev. Fish. Sci. Aquac. 2015, 23, 1–10. [Google Scholar] [CrossRef]
- Gatlin, D.M.; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, T.G.; Hardy, R.W.; Herman, E.; Hu, G.; Krogdahl, Å.; Nelson, R.; et al. Expanding the Utilization of Sustainable Plant Products in Aquafeeds: A Review. Aquac. Res. 2007, 38, 551–579. [Google Scholar] [CrossRef]
- Ao, Z.; Choct, M. Oligosaccharides Affect Performance and Gut Development of Broiler Chickens. Asian-Australasian J. Anim. Sci. 2013, 26, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Baeverfjord, G.; Krogdahl, Å. Development and Regression of Soybean Meal Induced Enteritis in Atlantic Salmon, Salmo salar L., Distal Intestine: A Comparison with the Intestines of Fasted Fish. J. Fish Dis. 1996, 19, 375–387. [Google Scholar] [CrossRef]
- Hedrera, M.I.; Galdames, J.A.; Jimenez-Reyes, M.F.; Reyes, A.E.; Avendaño-Herrera, R.; Romero, J.; Feijóo, C.G. Soybean Meal Induces Intestinal Inflammation in Zebrafish Larvae. PLoS One 2013, 8, 1–10. [Google Scholar] [CrossRef]
- Hu, H.; Kortner, T.M.; Gajardo, K.; Chikwati, E.; Tinsley, J.; Krogdahl, A. Intestinal Fluid Permeability in Atlantic Salmon (Salmo salar L.) Is Affected by Dietary Protein Source. PLoS One 2016, 11, 1–18. [Google Scholar] [CrossRef]
- Knudsen, D.; Jutfelt, F.; Sundh, H.; Sundell, K.; Koppe, W.; Frøkiær, H. Dietary Soya Saponins Increase Gut Permeability and Play a Key Role in the Onset of Soyabean-Induced Enteritis in Atlantic Salmon (Salmo salar L.). Br. J. Nutr. 2008, 100, 120–129. [Google Scholar] [CrossRef]
- Krogdahl, Å.; Bakke-McKellep, A.M.; Baeverfjord, G. Effects of Graded Levels of Standard Soybean Meal on Intestinal Structure, Mucosal Enzyme Activities, and Pancreatic Response in Atlantic Salmon (Salmo salar L.). Aquac. Nutr. 2003, 9, 361–371. [Google Scholar] [CrossRef]
- Romarheim, O.H.; Øverland, M.; Mydland, L.T.; Skrede, A.; Landsverk, T. Bacteria Grown on Natural Gas Prevent Soybean Meal-Induced Enteritis in Atlantic Salmon1-3. J. Nutr. 2011, 141, 124–130. [Google Scholar] [CrossRef]
- Booman, M.; Forster, I.; Vederas, J.C.; Groman, D.B.; Jones, S.R.M. Soybean Meal-Induced Enteritis in Atlantic Salmon (Salmo salar) and Chinook Salmon (Oncorhynchus tshawytscha) but Not in Pink Salmon (O. gorbuscha). Aquaculture 2018, 483, 238–243. [Google Scholar] [CrossRef]
- Kononova, S. V.; Zinchenko, D. V.; Muranova, T.A.; Belova, N.A.; Miroshnikov, A.I. Intestinal Microbiota of Salmonids and Its Changes upon Introduction of Soy Proteins to Fish Feed. Aquac. Int. 2019, 27, 475–496. [Google Scholar] [CrossRef]
- Sadoul, B.; Foucard, A.; Valotaire, C.; Labbé, L.; Goardon, L.; Lecalvez, J.M.; Médale, F.; Quillet, E.; Dupont-Nivet, M.; Geurden, I.; et al. Adaptive Capacities from Survival to Stress Responses of Two Isogenic Lines of Rainbow Trout Fed a Plant-Based Diet. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Talwar, C.; Nagar, S.; Lal, R.; Negi, R.K. Fish Gut Microbiome: Current Approaches and Future Perspectives. Indian J. Microbiol. 2018, 58, 397–414. [Google Scholar] [CrossRef]
- Desai, A.R.; Links, M.G.; Collins, S.A.; Mansfield, G.S.; Drew, M.D.; Van Kessel, A.G.; Hill, J.E. Effects of Plant-Based Diets on the Distal Gut Microbiome of Rainbow Trout (Oncorhynchus mykiss). Aquaculture 2012, 350–353, 134–142. [Google Scholar] [CrossRef]
- Green, T.J.; Smullen, R.; Barnes, A.C. Dietary Soybean Protein Concentrate-Induced Intestinal Disorder in Marine Farmed Atlantic Salmon, Salmo salar Is Associated with Alterations in Gut Microbiota. Vet. Microbiol. 2013, 166, 286–292. [Google Scholar] [CrossRef]
- Metochis, C.P.; Spanos, I.; Auchinachie, N.; Crampton, V.O.; Bell, J.G.; Adams, A.; Thompson, K.D. The Effects of Increasing Dietary Levels of Soy Protein Concentrate (SPC) on the Immune Responses and Disease Resistance (Furunculosis) of Vaccinated and Non-Vaccinated Atlantic Salmon (Salmo salar L.) Parr. Fish Shellfish Immunol. 2016, 59, 83–94. [Google Scholar] [CrossRef]
- Denstadli, V.; Hillestad, M.; Verlhac, V.; Klausen, M.; Øverland, M. Enzyme Pretreatment of Fibrous Ingredients for Carnivorous Fish: Effects on Nutrient Utilisation and Technical Feed Quality in Rainbow Trout (Oncurhynchus mykiss). Aquaculture 2011, 319, 391–397. [Google Scholar] [CrossRef]
- Perry, W.B.; Lindsay, E.; Payne, C.J.; Brodie, C.; Kazlauskaite, R. The Role of the Gut Microbiome in Sustainable Teleost Aquaculture. Proc. R. Soc. B Biol. Sci. 2020, 287. [Google Scholar] [CrossRef]
- Gajardo, K.; Rodiles, A.; Kortner, T.M.; Krogdahl, Å.; Bakke, A.M.; Merrifield, D.L.; Sørum, H. A High-Resolution Map of the Gut Microbiota in Atlantic Salmon (Salmo salar): A Basis for Comparative Gut Microbial Research. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- López Nadal, A.; Ikeda-Ohtsubo, W.; Sipkema, D.; Peggs, D.; McGurk, C.; Forlenza, M.; Wiegertjes, G.F.; Brugman, S. Feed, Microbiota, and Gut Immunity: Using the Zebrafish Model to Understand Fish Health. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Yukgehnaish, K.; Kumar, P.; Sivachandran, P.; Marimuthu, K.; Arshad, A.; Paray, B.A.; Arockiaraj, J. Gut Microbiota Metagenomics in Aquaculture: Factors Influencing Gut Microbiome and Its Physiological Role in Fish. Rev. Aquac. 2020, 12, 1903–1927. [Google Scholar] [CrossRef]
- Dvergedal, H.; Sandve, S.R.; Angell, I.L.; Klemetsdal, G.; Rudi, K. Association of Gut Microbiota with Metabolism in Juvenile Atlantic Salmon. Microbiome 2020, 8, 1–8. [Google Scholar] [CrossRef]
- Ghanbari, M.; Kneifel, W.; Domig, K.J. A New View of the Fish Gut Microbiome: Advances from next-Generation Sequencing. Aquaculture 2015, 448, 464–475. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost Microbiomes: The State of the Art in Their Characterization, Manipulation and Importance in Aquaculture and Fisheries. Front. Microbiol. 2014, 5, 1–1. [Google Scholar] [CrossRef]
- Dehler, C.E.; Secombes, C.J.; Martin, S.A.M. Environmental and Physiological Factors Shape the Gut Microbiota of Atlantic Salmon Parr (Salmo salar L.). Aquaculture 2017, 467, 149–157. [Google Scholar] [CrossRef]
- Smith, C.C.R.; Snowberg, L.K.; Gregory Caporaso, J.; Knight, R.; Bolnick, D.I. Dietary Input of Microbes and Host Genetic Variation Shape Among-Population Differences in Stickleback Gut Microbiota. ISME J. 2015, 9, 2515–2526. [Google Scholar] [CrossRef]
- Gajardo, K.; Jaramillo-Torres, A.; Kortner, T.M.; Merrifield, D.L.; Tinsley, J.; Bakke, A.M.; Krogdahl, Å. Alternative Protein Sources in the Diet Modulate Microbiota and Functionality in the Distal Intestine of Atlantic Salmon (Salmo salar). Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef]
- Michl, S.C.; Beyer, M.; Ratten, J.M.; Hasler, M.; LaRoche, J.; Schulz, C. A Diet-Change Modulates the Previously Established Bacterial Gut Community in Juvenile Brown Trout (Salmo trutta). Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Michl, S.C.; Ratten, J.M.; Beyer, M.; Hasler, M.; La Roche, J.; Schulz, C. The Malleable Gut Microbiome of Juvenile Rainbow Trout (Oncorhynchus mykiss): Dietdependent Shifts of Bacterial Community Structures. PLoS One 2017, 12, 1–21. [Google Scholar] [CrossRef]
- Dehler, C.E.; Secombes, C.J.; Martin, S.A.M. Seawater Transfer Alters the Intestinal Microbiota Profiles of Atlantic Salmon (Salmo salar L.). Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Drew, M.D.; Borgeson, T.L.; Thiessen, D.L. A Review of Processing of Feed Ingredients to Enhance Diet Digestibility in Finfish. Anim. Feed Sci. Technol. 2007, 138, 118–136. [Google Scholar] [CrossRef]
- Kirk, O.; Borchert, T.V.; Fuglsang, C.C. Industrial Enzyme Applications. Curr. Opin. Biotechnol. 2002, 13, 345–351. [Google Scholar] [CrossRef]
- Kiron, V. Fish Immune System and Its Nutritional Modulation for Preventive Health Care. Anim. Feed Sci. Technol. 2012, 173, 111–133. [Google Scholar] [CrossRef]
- Li, C.; Zhang, B.; Liu, C.; Zhou, H.; Wang, X.; Mai, K.; He, G. Effects of Dietary Raw or Enterococcus faecium Fermented Soybean Meal on Growth, Antioxidant Status, Intestinal Microbiota, Morphology, and Inflammatory Responses in Turbot (Scophthalmus maximus L.). Fish Shellfish Immunol. 2020, 100, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E.; Van Doan, H.; Lee, S.H.; Soltani, M.; Hoseinifar, S.H.; Harikrishnan, R.; Song, S.K. Probiotics, Lactic Acid Bacteria and Bacilli: Interesting Supplementation for Aquaculture. J. Appl. Microbiol. 2020, 129, 116–136. [Google Scholar] [CrossRef]
- Ganguly, S.; Dora, K.C.; Sarkar, S.; Chowdhury, S. Supplementation of Prebiotics in Fish Feed: A Review. Rev. Fish Biol. Fish. 2013, 23, 195–199. [Google Scholar] [CrossRef]
- Ringø, E.; Olsen, R.E.; Gifstad, T.; Dalmo, R.A.; Amlund, H.; Hemre, G.I.; Bakke, A.M. Prebiotics in Aquaculture: A Review. Aquac. Nutr. 2010, 16, 117–136. [Google Scholar] [CrossRef]
- Akrami, R.; Iri, Y.; Khoshbavar Rostami, H.; Razeghi Mansour, M. Effect of Dietary Supplementation of Fructooligosaccharide (FOS) on Growth Performance, Survival, Lactobacillus Bacterial Population and Hemato-Immunological Parameters of Stellate Sturgeon (Acipenser stellatus) Juvenile. Fish Shellfish Immunol. 2013, 35, 1235–1239. [Google Scholar] [CrossRef]
- Grisdale-Helland, B.; Helland, S.J.; Gatlin, D.M. The Effects of Dietary Supplementation with Mannanoligosaccharide, Fructooligosaccharide or Galactooligosaccharide on the Growth and Feed Utilization of Atlantic Salmon (Salmo salar). Aquaculture 2008, 283, 163–167. [Google Scholar] [CrossRef]
- Jacobsen, H.J.; Kousoulaki, K.; Sandberg, A.S.; Carlsson, N.G.; Ahlstrøm, Ø.; Oterhals, Å. Enzyme Pre-Treatment of Soybean Meal: Effects on Non-Starch Carbohydrates, Protein, Phytic Acid, and Saponin Biotransformation and Digestibility in Mink (Neovison vison). Anim. Feed Sci. Technol. 2018, 236, 1–13. [Google Scholar] [CrossRef]
- Bedford, M.R. Future Prospects for Non-Starch Polysaccharide Degrading Enzymes Development in Monogastric Nutrition; Gonzáles-Ortiz, G., Bedford, M.R., Bach Knudsen, K.E., Courtin, C.M., Classen, H.L., Eds.; 2019; ISBN 9789086868933.
- Foster, J.A.; Rinaman, L.; Cryan, J.F. Stress & the Gut-Brain Axis: Regulation by the Microbiome. Neurobiol. Stress 2017, 7, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.R.; Mayer, E.A. Gut-Brain Axis and Behavior. Nestle Nutr Inst Work. Ser. 2017, 88, 45–53. [Google Scholar] [CrossRef]
- Borrelli, L.; Aceto, S.; Agnisola, C.; De Paolo, S.; Dipineto, L.; Stilling, R.M.; Dinan, T.G.; Cryan, J.F.; Menna, L.F.; Fioretti, A. Probiotic Modulation of the Microbiota-Gut-Brain Axis and Behaviour in Zebrafish. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.C.; Cable, J.; Lima-Maximino, M.G.; Maximino, C.; Xavier, R. Using Fish Models to Investigate the Links between Microbiome and Social Behaviour: The next Step for Translational Microbiome Research? Fish Fish. 2019, 20, 640–652. [Google Scholar] [CrossRef]
- Martins, C.I.M.; Galhardo, L.; Noble, C.; Damsgård, B.; Spedicato, M.T.; Zupa, W.; Beauchaud, M.; Kulczykowska, E.; Massabuau, J.C.; Carter, T.; et al. Behavioural Indicators of Welfare in Farmed Fish. Fish Physiol. Biochem. 2012, 38, 17–41. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, L.U. Fish Behaviour and Welfare. Appl. Anim. Behav. Sci. 2007, 3, 173–175. [Google Scholar] [CrossRef]
- Huntingford, F.A.; Adams, C.; Braithwaite, V.A.; Kadri, S.; Pottinger, T.G.; Sandøe, P.; Turnbull, J.F. Current Issues in Fish Welfare. J. Fish Biol. 2006, 68, 332–372. [Google Scholar] [CrossRef]
- Solgaard, H.S.; Yang, Y. Consumers’ Perception of Farmed Fish and Willingness to Pay for Fish Welfare. Br. Food J. 2011, 113, 997–1010. [Google Scholar] [CrossRef]
- Castanheira, M.F.; Conceição, L.E.C.; Millot, S.; Rey, S.; Bégout, M.L.; Damsgård, B.; Kristiansen, T.; Höglund, E.; Øverli, Ø.; Martins, C.I.M. Coping Styles in Farmed Fish: Consequences for Aquaculture. Rev. Aquac. 2017, 9, 23–41. [Google Scholar] [CrossRef]
- Conrad, J.L.; Weinersmith, K.L.; Brodin, T.; Saltz, J.B.; Sih, A. Behavioural Syndromes in Fishes: A Review with Implications for Ecology and Fisheries Management. J. Fish Biol. 2011, 78, 395–435. [Google Scholar] [CrossRef] [PubMed]
- Koolhaas, J.M.; Korte, S.M.; De Boer, S.F.; Van Der Vegt, B.J.; Van Reenen, C.G.; Hopster, H.; De Jong, I.C.; Ruis, M.A.W.; Blokhuis, H.J. Coping Styles in Animals: Current Status in Behavior and Stress- Physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Réale, D.; Reader, S.M.; Sol, D.; McDougall, P.T.; Dingemanse, N.J. Integrating Animal Temperament within Ecology and Evolution. Biol. Rev. Camb. Philos. Soc. 2007, 82, 291–318. [Google Scholar] [CrossRef] [PubMed]
- Benhaïm, D.; Leblanc, C.A.L.; Horri, K.; Mannion, K.; Galloway, M.; Leeper, A.; Knobloch, S.; Sigurgeirsson, Ó.; Thorarensen, H. The Effect of Triploidy on the Performance, Gut Microbiome and Behaviour of Juvenile Atlantic Salmon (Salmo salar) Raised at Low Temperature. Appl. Anim. Behav. Sci. 2020, 229, 105031. [Google Scholar] [CrossRef]
- Sih, A.; Bell, A.; Johnson, J.C. Behavioral Syndromes: An Ecological and Evolutionary Overview. Trends Ecol. Evol. 2004, 19, 372–378. [Google Scholar] [CrossRef]
- MacKenzie, S.; Ribas, L.; Pilarczyk, M.; Capdevila, D.M.; Kadri, S.; Huntingford, F.A. Screening for Coping Style Increases the Power of Gene Expression Studies. PLoS One 2009, 4, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Øverli, Ø.; Korzan, W.J.; Larson, E.T.; Winberg, S.; Lepage, O.; Pottinger, T.G.; Renner, K.J.; Summers, C.H. Behavioral and Neuroendocrine Correlates of Displaced Aggression in Trout. Horm. Behav. 2004, 45, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.F.; Herrera, M.; Costas, B.; Conceição, L.E.C.; Martins, C.I.M. Can We Predict Personality in Fish? Searching for Consistency over Time and across Contexts. PLoS One 2013, 8, 1–9. [Google Scholar] [CrossRef]
- Ruiz-Gomez, M. de L.; Kittilsen, S.; Höglund, E.; Huntingford, F.A.; Sørensen, C.; Pottinger, T.G.; Bakken, M.; Winberg, S.; Korzan, W.J.; Øverli, Ø. Behavioral Plasticity in Rainbow Trout (Oncorhynchus mykiss) with Divergent Coping Styles: When Doves Become Hawks. Horm. Behav. 2008, 54, 534–538. [Google Scholar] [CrossRef]
- Sneddon, L.U. The Bold and the Shy: Individual Differences in Rainbow Trout. J. Fish Biol. 2003, 62, 971–975. [Google Scholar] [CrossRef]
- Ward, A.J.W.; Thomas, P.; Hart, P.J.B.; Krause, J. Correlates of Boldness in Three-Spined Sticklebacks (Gasterosteus aculeatus). Behav. Ecol. Sociobiol. 2004, 55, 561–568. [Google Scholar] [CrossRef]
- Webster, A.J.F. Farm Animal Welfare: The Five Freedoms and the Free Market. Vet. J. 2001, 161, 229–237. [Google Scholar] [CrossRef]
- Ishizaki, Y.; Masuda, R.; Uematsu, K.; Shimizu, K.; Arimoto, M.; Takeuchi, T. The Effect of Dietary Docosahexaenoic Acid on Schooling Behaviour and Brain Development in Larval Yellowtail. J. Fish Biol. 2001, 58, 1691–1703. [Google Scholar] [CrossRef]
- Ibarra-Zatarain, Z.; Parati, K.; Cenadelli, S.; Duncan, N. Reproductive Success of a Marine Teleost Was Correlated with Proactive and Reactive Stress-Coping Styles. J. Fish Biol. 2019, 94, 402–413. [Google Scholar] [CrossRef]
- Masayuki Yoshida, M.N. and K.U.G. Comparison of Behavioral Responses to a Novel Environment between Three Teleosts, Bluegill. Fish. Sci. 2005, 71, 314–319. [Google Scholar] [CrossRef]
- Budaev, S. V.; Zworykin, D.D.; Mochek, A.D. Individual Differences in Parental Care and Behaviour Profile in the Convict Cichlid: A Correlation Study. Anim. Behav. 1999, 58, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Toms, C.N.; Echevarria, D.J.; Jouandot, D.J. A Methodological Review of Personality-Related Studies in Fish: Focus on the Shy-Bold Axis of Behavior. Int. J. Comp. Psychol. 2010, 23, 0–25. [Google Scholar] [CrossRef]
- Dahlbom, S.J.; Lagman, D.; Lundstedt-Enkel, K.; Sundström, L.F.; Winberg, S. Boldness Predicts Social Status in Zebrafish (Danio rerio). PLoS One 2011, 6, 2–8. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. ; R Core Team Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-137. Http://CRAN.R-Project.Org/Package=nlme. 2018.
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Lahti, L.; Shetty, S. Microbiome R Package. Bioconductor 2017. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package. 2022.
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag New York. ISBN 978-3-319-24277-4. https://ggplot2.tidyverse.org 2016.
- Davis, N.M.; Proctor, Di.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple Statistical Identification and Removal of Contaminant Sequences in Marker-Gene and Metagenomics Data. Microbiome 2018, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Second Edi.; SAGE Publications. 2011. [Google Scholar]
- Beukeboom, R.; Morel, A.; Phillips, J.S.; Ólafsdóttir, G.Á.; Benhaïm, D. Activity vs Exploration: Locomotion in a Known and Unknown Environment Differs in Atlantic Cod Juveniles (Gadus morhua). Behav. Processes 2022, 202, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Zero-Truncated and Zero-Inflated Models for Count Data BT - Mixed Effects Models and Extensions in Ecology with R; 2009; ISBN 978-0-387-87458-6.
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biometrical J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Carpenter, B.; Gelman, A.; Hoffman, M.D.; Lee, D.; Goodrich, B.; Betancourt, M.; Brubaker, M.A.; Guo, J.; Li, P.; Riddell, A. Stan: A Probabilistic Programming Language. J. Stat. Softw. 2017, 76. [Google Scholar] [CrossRef] [PubMed]
- Bürkner, P.C. Brms: An R Package for Bayesian Multilevel Models Using Stan. J. Stat. Softw. 2017, 80. [Google Scholar] [CrossRef]
- Johnson, W.D.; Koch, G.G. Intraclass Correlation Coefficient. Lovric, M. Int. Encycl. Stat. Sci. Springer, Berlin, Heidelberg, 2011. [CrossRef]
- Plummer, M.; Best, N.; Cowles, K.; Vines, K. {CODA}: Convergence Diagnosis and Output Analysis for {MCMC}. R News 2006, 6, 7–11. [Google Scholar]
- Nathanailides, C.; Lopez-Albors, O.; Stickland, N.C. Influence of Prehatch Temperature on the Development of Muscle Cellularity in Posthatch Atlantic Salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 1995, 52, 675–680. [Google Scholar] [CrossRef]
- Sørensen, M.; Gong, Y.; Bjarnason, F.; Vasanth, G.K.; Dahle, D.; Huntley, M.; Kiron, V. Nannochloropsis Oceania-Derived Defatted Meal as an Alternative to Fishmeal in Atlantic Salmon Feeds. PLoS One 2017, 12, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Storebakken, T.; Kvien, I.S.; Shearer, K.D.; Grisdale-Helland, B.; Helland, S.J.; Berge, G.M. The Apparent Digestibility of Diets Containing Fish Meal, Soybean Meal or Bacterial Meal Fed to Atlantic Salmon (Salmo salar): Evaluation of Different Faecal Collection Methods. Aquaculture 1998, 169, 195–210. [Google Scholar] [CrossRef]
- Sahlmann, C.; Gu, J.; Kortner, T.M.; Lein, I.; Krogdahl, Å.; Bakke, A.M. Ontogeny of the Digestive System of Atlantic Salmon (Salmo salar L.) and Effects of Soybean Meal from Start-Feeding. PLoS One 2015, 10, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Øvrum Hansen, J.; Hofossæter, M.; Sahlmann, C.; Ånestad, R.; Reveco-Urzua, F.E.; Press, C.M.L.; Mydland, L.T.; Øverland, M. Effect of Candida utilis on Growth and Intestinal Health of Atlantic Salmon (Salmo salar) Parr. Aquaculture 2019, 511, 734239. [Google Scholar] [CrossRef]
- Olli, J.J.; Krogdahl; Våbenø, A. Dehulled Solvent-extracted Soybean Meal as a Protein Source in Diets for Atlantic Salmon, Salmo salar L. Aquac. Res. 1995, 26, 167–174. [Google Scholar] [CrossRef]
- Refstie, S.; Storebakken, T.; Roem, A. Feed Consumption and Conversion in Atlantic Salmon (Salmo salar) Fed Diets with Fish Meal, Extracted Soybean Meal or Soybean Meal with Reduced Content Of. Aquaculture 1998, 301–312. [Google Scholar] [CrossRef]
- Ai, Q.; Mai, K.; Zhang, W.; Xu, W.; Tan, B.; Zhang, C.; Li, H. Effects of Exogenous Enzymes (Phytase, Non-Starch Polysaccharide Enzyme) in Diets on Growth, Feed Utilization, Nitrogen and Phosphorus Excretion of Japanese Seabass, Lateolabrax japonicus. Comp. Biochem. Physiol. - A Mol. Integr. Physiol. 2007, 147, 502–508. [Google Scholar] [CrossRef]
- Zhu, D.; Wen, X.; Li, S.; Xuan, X.; Li, Y. Effects of Exogenous Non-Starch Polysaccharide-Degrading Enzymes in Diets Containing Gracilaria lemaneiformis on White-Spotted Snapper Lutjanus stellatus Akazaki. Aquac. Int. 2016, 24, 491–502. [Google Scholar] [CrossRef]
- Denstadli, V.; Storebakken, T.; Svihus, B.; Skrede, A. A Comparison of Online Phytase Pre-Treatment of Vegetable Feed Ingredients and Phytase Coating in Diets for Atlantic Salmon (Salmo salar L.) Reared in Cold Water. Aquaculture 2007, 269, 414–426. [Google Scholar] [CrossRef]
- Ortiz, L.T.; Rebolé, A.; Velasco, S.; Rodríguez, M.L.; Treviño, J.; Tejedor, J.L.; Alzueta, C. Effects of Inulin and Fructooligosaccharides on Growth Performance, Body Chemical Composition and Intestinal Microbiota of Farmed Rainbow Trout (Oncorhynchus mykiss). Aquac. Nutr. 2013, 19, 475–482. [Google Scholar] [CrossRef]
- Bruce, T.J.; Neiger, R.D.; Brown, M.L. Gut Histology, Immunology and the Intestinal Microbiota of Rainbow Trout, Oncorhynchus mykiss (Walbaum), Fed Process Variants of Soybean Meal. Aquac. Res. 2018, 49, 492–504. [Google Scholar] [CrossRef]
- Reveco, F.E.; Øverland, M.; Romarheim, O.H.; Mydland, L.T. Intestinal Bacterial Community Structure Differs between Healthy and Inflamed Intestines in Atlantic Salmon (Salmo salar L.). Aquaculture 2014, 420–421, 262–269. [Google Scholar] [CrossRef]
- Guerreiro, I.; Serra, C.R.; Pousão-Ferreira, P.; Oliva-Teles, A.; Enes, P. Prebiotics Effect on Growth Performance, Hepatic Intermediary Metabolism, Gut Microbiota and Digestive Enzymes of White Sea Bream (Diplodus sargus). Aquac. Nutr. 2018, 24, 153–163. [Google Scholar] [CrossRef]
- Liu, W.; Wang, W.; Ran, C.; He, S.; Yang, Y.; Zhou, Z. Effects of Dietary ScFOS and Lactobacilli on Survival, Growth, and Disease Resistance of Hybrid Tilapia. Aquaculture 2017, 470, 50–55. [Google Scholar] [CrossRef]
- Burgos, F.A.; Ray, C.L.; Arias, C.R. Bacterial Diversity and Community Structure of the Intestinal Microbiome of Channel Catfish (Ictalurus punctatus) during Ontogenesis. Syst. Appl. Microbiol. 2018, 41, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Roberfroid, M.B. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Infante-Villamil, S.; Huerlimann, R.; Jerry, D.R. Microbiome Diversity and Dysbiosis in Aquaculture. Rev. Aquac. 2021, 13, 1077–1096. [Google Scholar] [CrossRef]
- Alshammari, E.; Patel, M.; Sachidanandan, M.; Kumar, P.; Adnan, M. Potential Evaluation and Health Fostering Intrinsic Traits of Novel Probiotic Strain Enterococcus durans F3 Isolated from the Gut of Fresh Water Fish Catla catla. Food Sci. Anim. Resour. 2019, 39, 844–861. [Google Scholar] [CrossRef]
- Momeni-Moghaddam, P.; Keyvanshokooh, S.; Ziaei-Nejad, S.; Parviz Salati, A.; Pasha-Zanoosi, H. Effects of Mannan Oligosaccharide Supplementation on Growth, Some Immune Responses and Gut Lactic Acid Bacteria of Common Carp (Cyprinus carpio) Fingerlings. Vet. Res. forum an Int. Q. J. 2015, 6, 239–244. [Google Scholar]
- De Vuyst, L.; Leroy, F. Bacteriocins from Lactic Acid Bacteria: Production, Purification, and Food Applications. J. Mol. Microbiol. Biotechnol. 2007, 13, 194–199. [Google Scholar] [CrossRef]
- Ringø, E.; Hoseinifar, S.H.; Ghosh, K.; Doan, H. Van; Beck, B.R.; Song, S.K. Lactic Acid Bacteria in Finfish-An Update. Front. Microbiol. 2018, 9, 1–37. [Google Scholar] [CrossRef]
- Chapagain, P.; Arivett, B.; Cleveland, B.M.; Walker, D.M.; Salem, M. Analysis of the Fecal Microbiota of Fast-and Slow-Growing Rainbow Trout (Oncorhynchus mykiss). BMC Genomics 2019, 20, 1–11. [Google Scholar] [CrossRef]
- Neis, E.P.J.G.; Dejong, C.H.C.; Rensen, S.S. The Role of Microbial Amino Acid Metabolism in Host Metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef]
- Steiner, K.; Heasman, K.; Laroche, O.; Pochon, X.; Preece, M.; Bowman, J.P.; Walker, S.P.; Symonds, J.E. The Microbiome of Chinook Salmon (Oncorhynchus tshawytscha) in a Recirculation Aquaculture System. Aquaculture 2021, 534, 736227. [Google Scholar] [CrossRef]
- Madaro, A.; Fernö, A.; Kristiansen, T.S.; Olsen, R.E.; Gorissen, M.; Flik, G.; Nilsson, J. Effect of Predictability on the Stress Response to Chasing in Atlantic Salmon (Salmo salar L.) Parr. Physiol. Behav. 2016, 153, 1–6. [Google Scholar] [CrossRef]
- Huntingford, F.; Adams, C. Behavioural Syndromes in Farmed Fish: Implications for Production and Welfare. Behaviour 2005, 142, 1207–1221. [Google Scholar] [CrossRef]
- Ibarra-Zatarain, Z.; Morais, S.; Bonacic, K.; Campoverde, C.; Duncan, N. Dietary Fatty Acid Composition Significantly Influenced the Proactive-Reactive Behaviour of Senegalese Sole (Solea senegalensis) Post-Larvae. Appl. Anim. Behav. Sci. 2015, 171, 233–240. [Google Scholar] [CrossRef]
- Bell, A.M.; Hankison, S.J.; Laskowski, K.L. The Repeatability of Behaviour: A Meta-Analysis Alison. Anim. Behav. 2009, 77, 771–783. [Google Scholar] [CrossRef]
- Damsgard, B.; Evensen, T.H.; Øverli, Ø.; Gorissen, M.; Ebbesson, L.O.E.; Rey, S.; Höglund, E. Proactive Avoidance Behaviour and Pace-of-Life Syndrome in Atlantic Salmon. R. Soc. Open Sci. 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Øverli, Ø.; Sørensen, C.; Pulman, K.G.T.; Pottinger, T.G.; Korzan, W.; Summers, C.H.; Nilsson, G.E. Evolutionary Background for Stress-Coping Styles: Relationships between Physiological, Behavioral, and Cognitive Traits in Non-Mammalian Vertebrates. Neurosci. Biobehav. Rev. 2007, 31, 396–412. [Google Scholar] [CrossRef]
- Höglund, E.; Gjøen, H.M.; Pottinger, T.G. ; Øverli Parental Stress-Coping Styles Affect the Behaviour of Rainbow Trout Oncorhynchus mykiss at Early Developmental Stages. J. Fish Biol. 2008, 73, 1764–1769. [Google Scholar] [CrossRef]
- Evans, J.; Boudreau, K.; Hyman, J. Behavioural Syndromes in Urban and Rural Populations of Song Sparrows. Ethology 2010, 116, 588–595. [Google Scholar] [CrossRef]
- Baker, M.R.; Goodman, A.C.; Santo, J.B.; Wong, R.Y. Repeatability and Reliability of Exploratory Behavior in Proactive and Reactive Zebrafish, Danio rerio. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Horri, K.; Allal, F.; Vergnet, A.; Benhaim, D.; Vandeputte, M.; Chatain, B.; Bégout, M.L. Heritability of Boldness and Hypoxia Avoidance in European Seabass, Dicentrarchus labrax. PLoS One 2016, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Knobloch, S.; Skírnisdóttir, S.; Dubois, M.; Kolypczuk, L.; Leroi, F.; Leeper, A.; Passerini, D.; Marteinsson, V. Impact of Putative Probiotics on Growth, Behavior, and the Gut Microbiome of Farmed Arctic Char (Salvelinus alpinus). Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef]
- Prentice, P.M.; Houslay, T.M.; Wilson, A.J. Exploiting Animal Personality to Reduce Chronic Stress in Captive Fish Populations. Front. Vet. Sci. 2022, 9. [Google Scholar] [CrossRef]
Figure 1.
Swimming test arenas used for observation of behavioural traits of exploration and stress response.
Figure 1.
Swimming test arenas used for observation of behavioural traits of exploration and stress response.

Figure 2.
(A). Open Field test arenas used for observation of the behavioural trait of boldness. (B). An arena with four virtual zones outlined in the EthoVision XT software, Shelter, Entry, Border, and Centre.
Figure 2.
(A). Open Field test arenas used for observation of the behavioural trait of boldness. (B). An arena with four virtual zones outlined in the EthoVision XT software, Shelter, Entry, Border, and Centre.

Figure 3.
Average Individual Specific Growth Rate (%) for each dietary treatment. Different letters above the bars indicate significant differences (Tukey post-hoc test, p<0.05). FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
Figure 3.
Average Individual Specific Growth Rate (%) for each dietary treatment. Different letters above the bars indicate significant differences (Tukey post-hoc test, p<0.05). FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
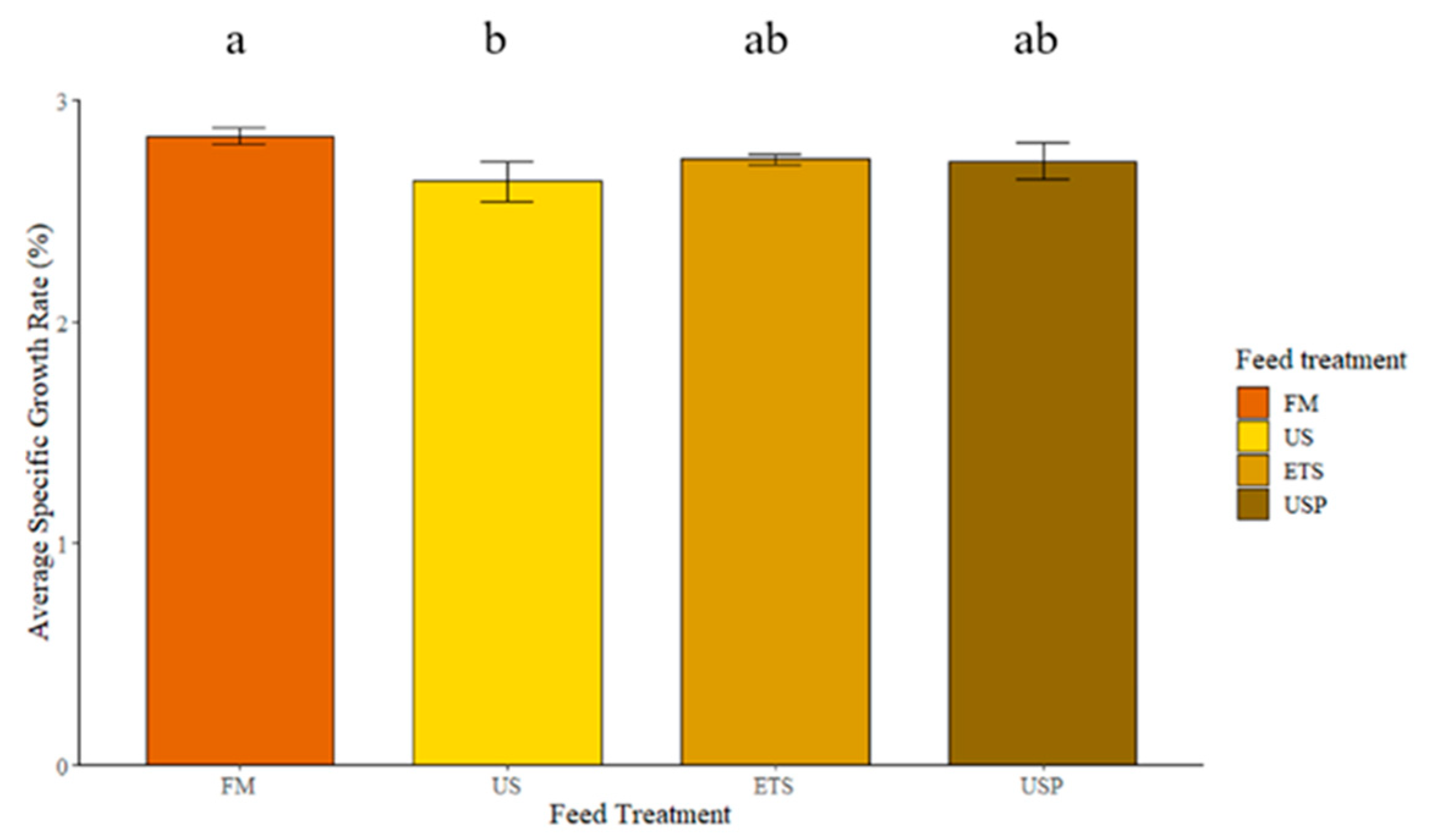
Figure 4.
Alpha diversity measures of fish fed different dietary treatments: (A) observed richness of ASVs, (B) Shannon diversity, (C) Chao1 diversity, (D) Pielou evenness. Different letters above the bars indicate significant differences (Tukey post-hoc test, p<0.05). FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
Figure 4.
Alpha diversity measures of fish fed different dietary treatments: (A) observed richness of ASVs, (B) Shannon diversity, (C) Chao1 diversity, (D) Pielou evenness. Different letters above the bars indicate significant differences (Tukey post-hoc test, p<0.05). FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
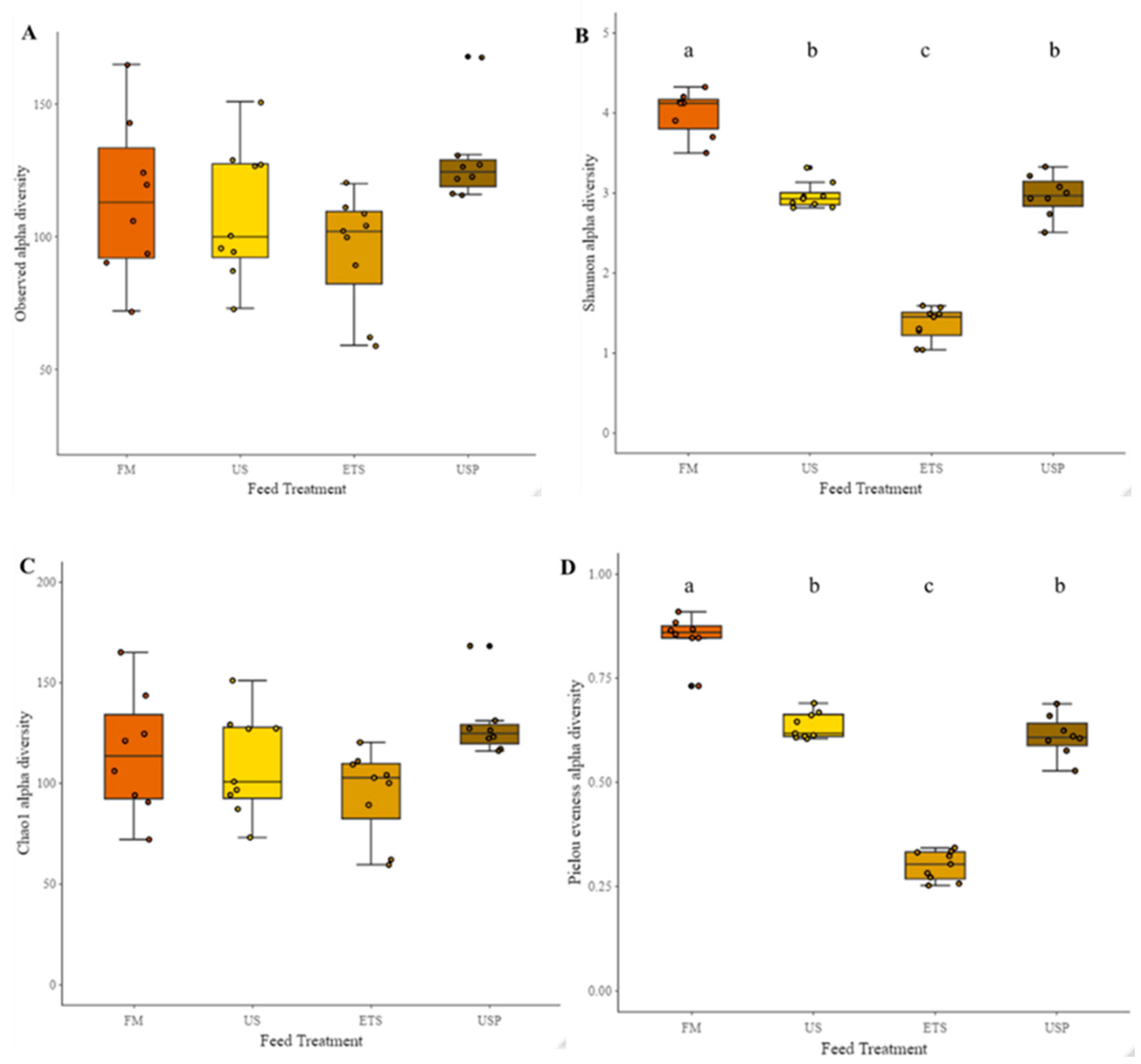
Figure 5.
NMDS plot of individual fish gut microbiome composition for each feed treatment. FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
Figure 5.
NMDS plot of individual fish gut microbiome composition for each feed treatment. FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
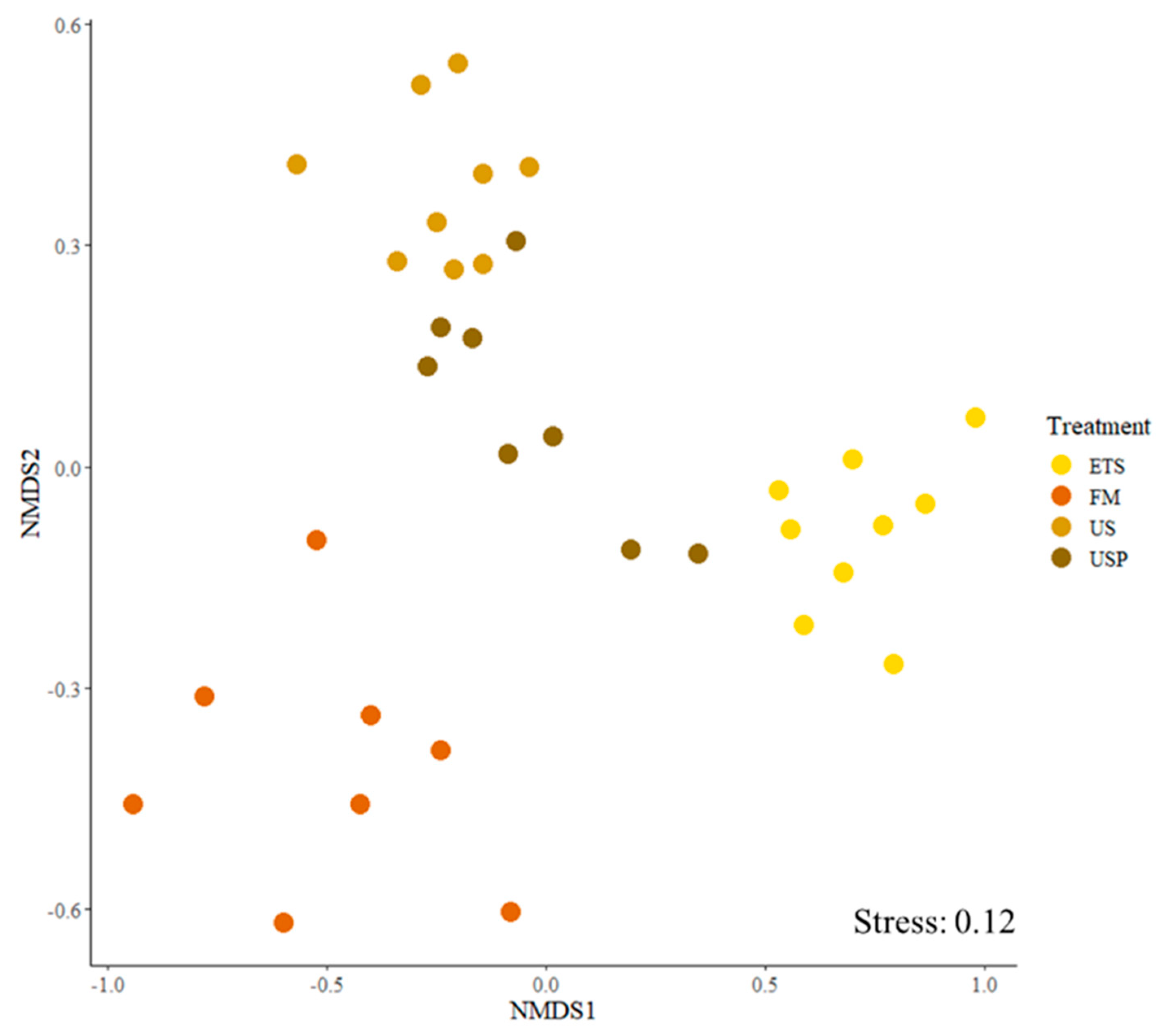
Figure 6.
Relative abundance of genera of taxa present in gut microbiome community of fish fed each dietary treatment. FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
Figure 6.
Relative abundance of genera of taxa present in gut microbiome community of fish fed each dietary treatment. FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
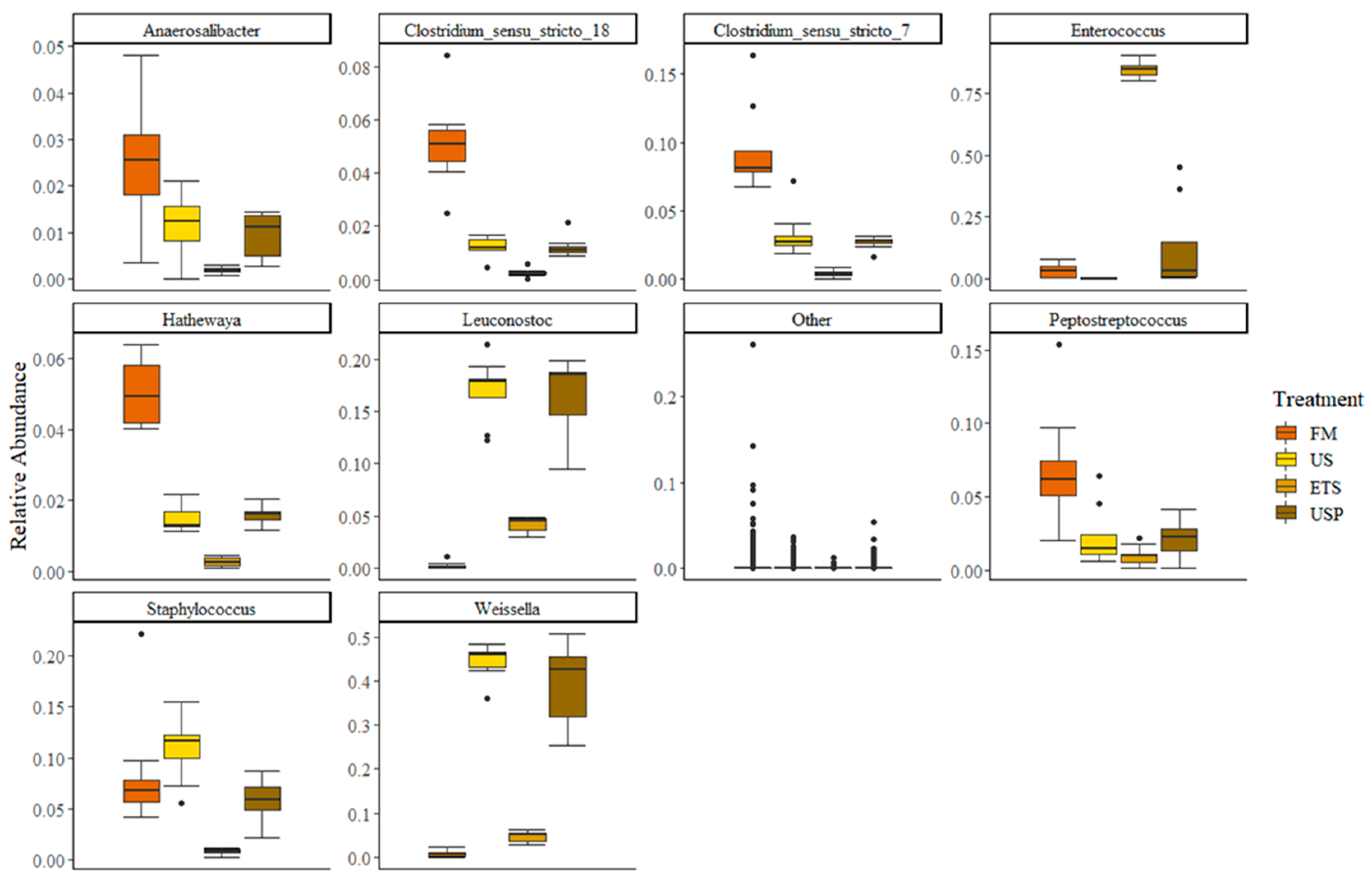
Figure 7.
Distance travelled (i.e., proxy for exploration) of fish fed different dietary treatments. FM is fish-meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
Figure 7.
Distance travelled (i.e., proxy for exploration) of fish fed different dietary treatments. FM is fish-meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
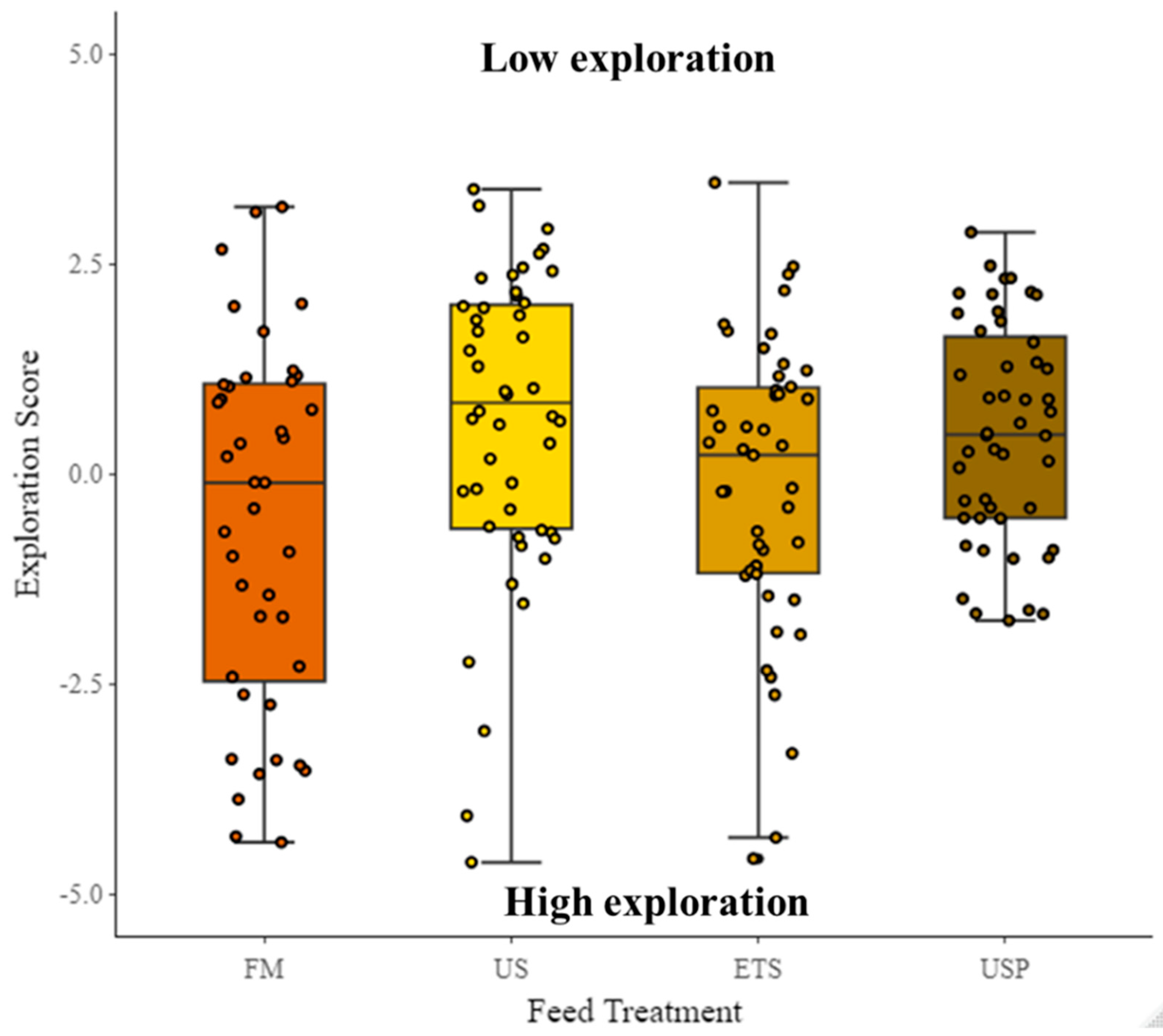
Figure 8.
Boldness score of fish fed different feed treatments where lower values indicate shyer individuals and high values indicate bolder individuals. FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
Figure 8.
Boldness score of fish fed different feed treatments where lower values indicate shyer individuals and high values indicate bolder individuals. FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
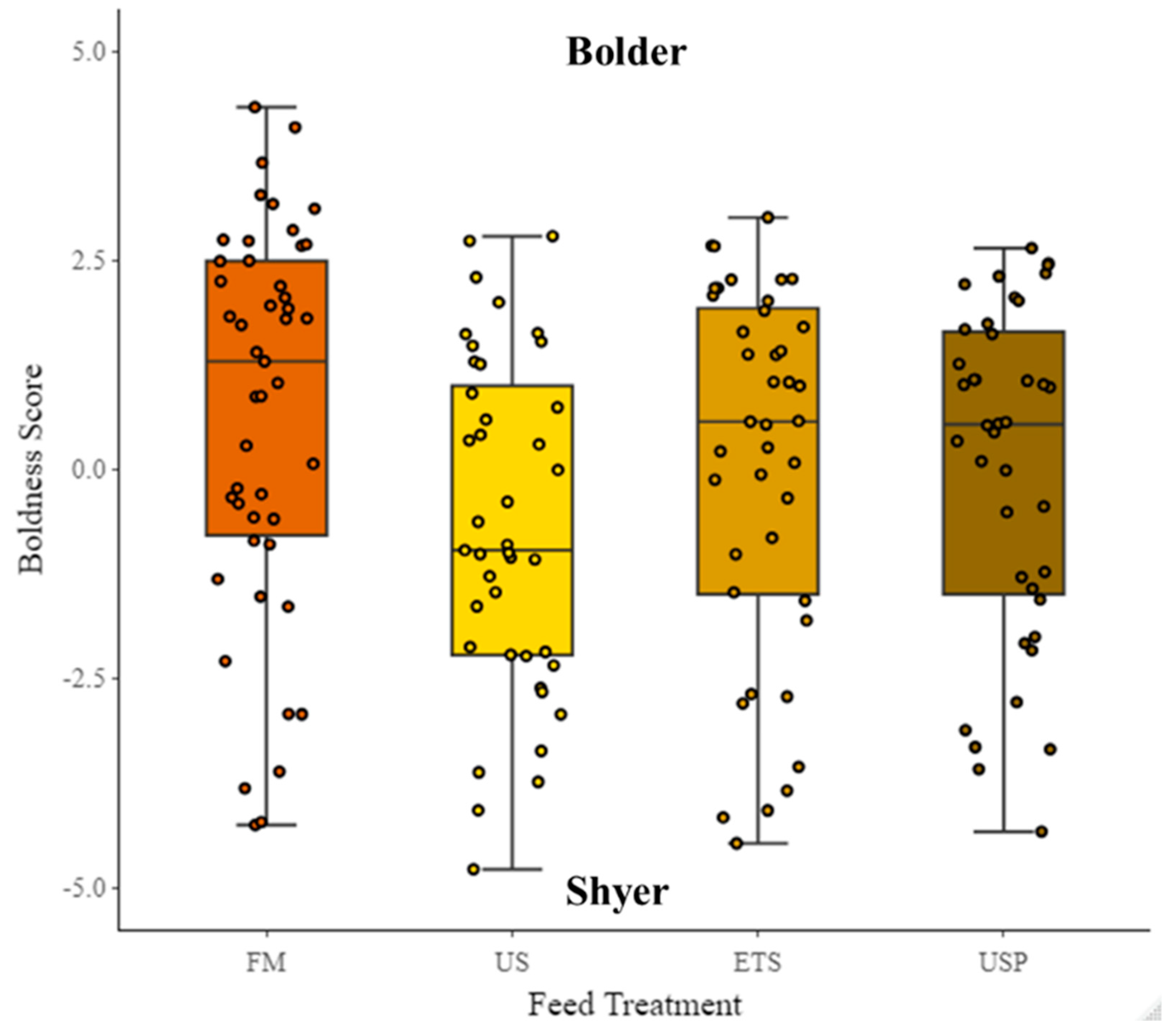
Figure 9.
Left side: Posterior modes and their corresponding uncertainty intervals for the variances and pairwise covariance of boldness and exploration for each diet treatment, on both the ID level and residual level. The black dashed line indicates zero. Right side: Posterior modes and their corresponding 95 % uncertainty intervals for the fixed effects SGR, TL, Tank and Trial.TL, SGR and Trial. Dashed line indicates zero. Because the non-categorial fixed effects were z-scored, the coefficient values are presented in units of standard deviation. FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
Figure 9.
Left side: Posterior modes and their corresponding uncertainty intervals for the variances and pairwise covariance of boldness and exploration for each diet treatment, on both the ID level and residual level. The black dashed line indicates zero. Right side: Posterior modes and their corresponding 95 % uncertainty intervals for the fixed effects SGR, TL, Tank and Trial.TL, SGR and Trial. Dashed line indicates zero. Because the non-categorial fixed effects were z-scored, the coefficient values are presented in units of standard deviation. FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide.
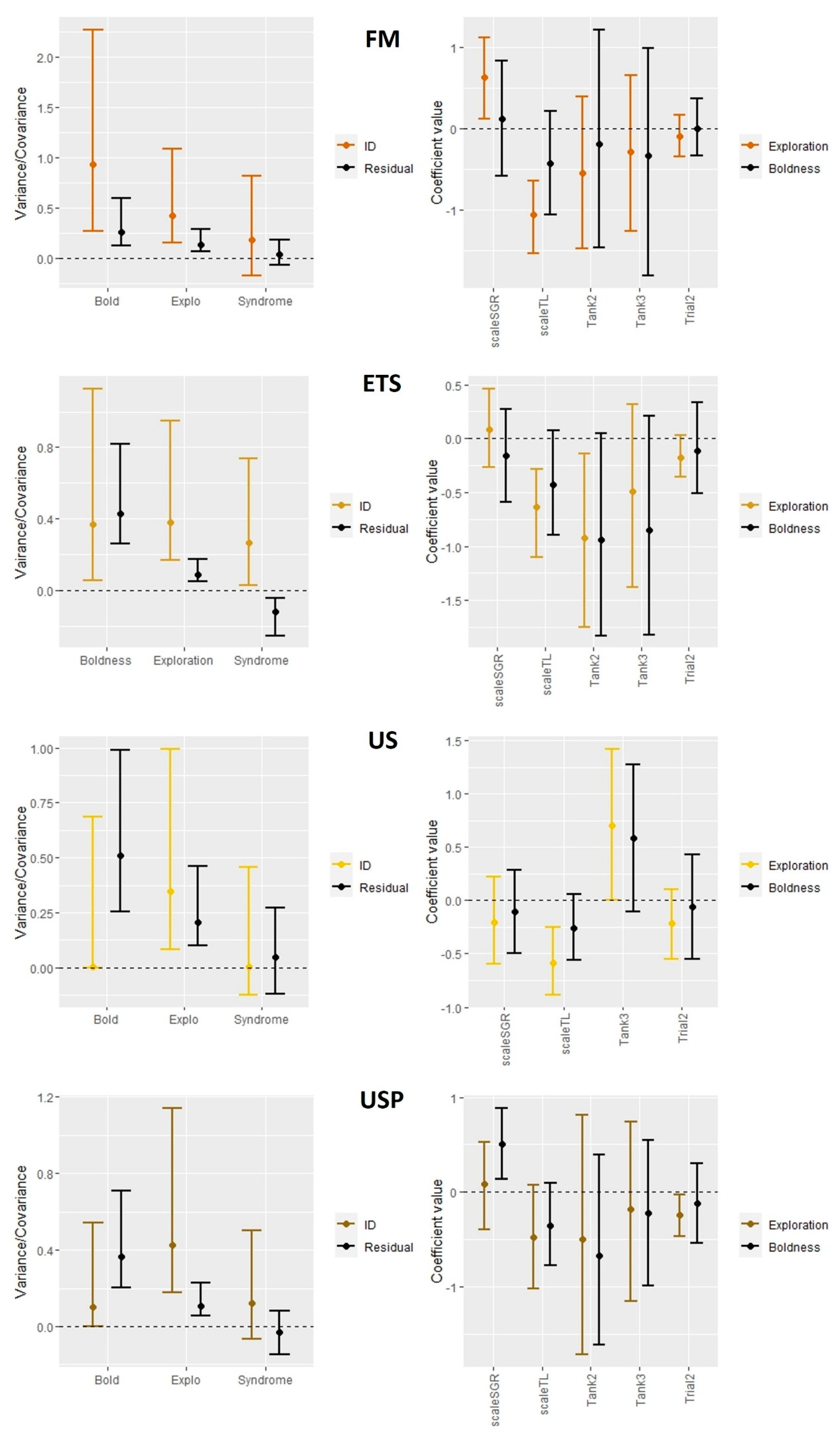

Table 1.
Chemical composition of the protein sources used in feed treatments for this study.
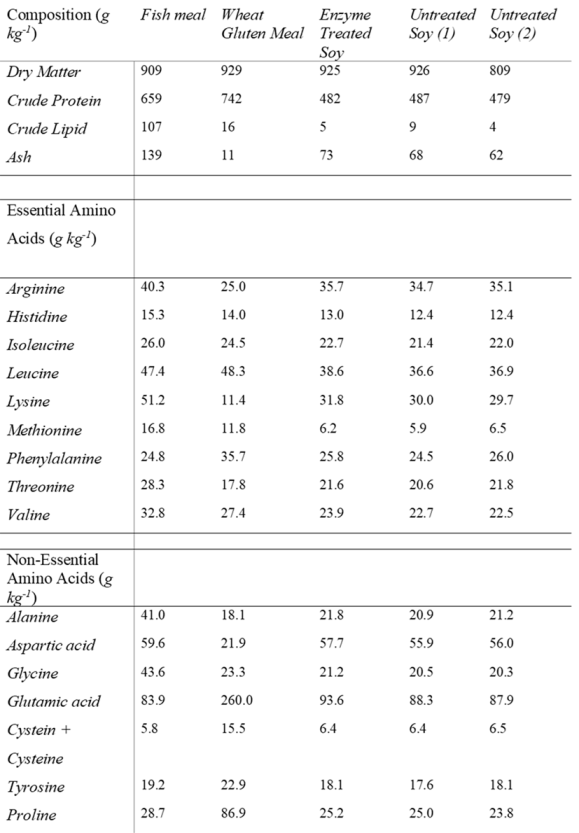
Table 2.
Feed formulation and chemical composition for feed treatments for this study.
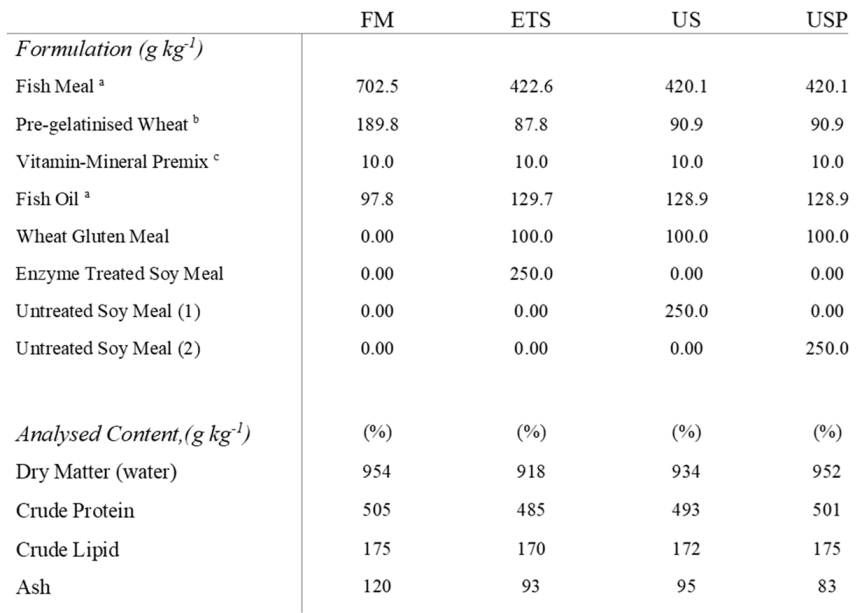
a Laxá hf, Iceland. b Emmelev A/S, Denmark. c Laxa salmon premix 2006, Trouw Nutrition, Holland.
Table 3.
Repeatability estimates (R) in the four dietary treatments. FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide. Boldness scores and exploration were obtained from two consecutive open field tests repeated 7 days apart. CI: confidence interval.
Table 3.
Repeatability estimates (R) in the four dietary treatments. FM is fish meal-based control diet, US is an untreated soybean meal diet, ETS is an enzyme treated soybean meal, and USP is an untreated soybean meal with fructose oligosaccharide. Boldness scores and exploration were obtained from two consecutive open field tests repeated 7 days apart. CI: confidence interval.
| Diet treatment | R | CI |
|---|---|---|
| FM | ||
| Boldness | 0.84 | [0.49-0.94] |
| Exploration | 0.82 | [0.52-0.94] |
| ETS | ||
| Boldness | 0.49 | [0.16-0.77] |
| Exploration | 0.84 | [0.64-0.94] |
| US | ||
| Boldness | 0.00 | [0.00-0.62] |
| Exploration | 0.72 | [0.30-0.89] |
| USP | ||
| Boldness | 0.30 | [0.00-0.61] |
| Exploration | 0.85 | [0.60-0.94] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



