
Submitted:
05 July 2023
Posted:
06 July 2023
You are already at the latest version
Abstract
Synaptic zinc ions (Zn2+) play an important role in the development of vascular dementia (VD) and Parkinson’s disease (PD). In this article, based on our study and many others, we review the mo-lecular pathways by which Zn2+ causes neurotoxicity. Zn2+ influences calcium homeostasis, energy production pathway, production of reactive oxygen species, endoplasmic reticulum stress pathway, and activated protein kinase/c-Jun amino terminal kinase (SAPK/JNK) pathway and consequently exerts neurotoxicity. Furthermore, we searched various crops for substances that protect neurones from neurotoxicity caused by Zn2+ and clarified that carnosine (β-alanylhistidine) may be a therapeutic drug for VD and PD. Here, we also review the molecular mechanisms underlying the role of carnosine as an endogenous protector and its protective effect against Zn2+-induced cyto-toxicity and discuss prospects for future neurodegenerative diseases therapeutic applications of this dipeptide.
Keywords:
apoptosis
; carnosine
; endoplasmic reticulum stress
; Parkinson’s disease
; synapse
; vascular dementia
; zinc
1. Introduction
Zinc (Zn) plays an important role in a variety of physiological functions, including cell mitosis, immune system, and protein and nucleic acid synthesis, and acts as a cofac-tor for over 300 enzymes or metalloproteins, contributing to normal brain function [1]. Nevertheless, Zn is widely known to play an important role in the development of post-ischaemic neurodegeneration and vascular dementia (VD) [2]. Recently, it has been reported that Zn is involved in the mechanism of Parkinson’s disease (PD) pathogenesis [3].
Senile dementia is characterised by severe memory loss and inability to form new memories in the elderly, and its prevalence increases with age. A fact sheet published by the World Health Organization in 2023 states that there are 55 million people with dementia worldwide, an increase of nearly 10 million each year [4]. Dementia is a serious social problem in rapidly aging societies. Senile dementia is classified into Alzheimer’s disease (AD), VD, and dementia with Lewy bodies (DLB). VD is considered the second most common type of dementia, accounting for approximately 20%-40% of senile dementias [5]. Both AD and DLB are characterised by abnormally accumulated protein deposits (β-amyloid protein (AβP) in AD and α-synuclein in DLB), whereas VD is a degenerative cerebrovascular disease with a series of strokes or ischaemia [6,7,8]. PD is a progressive neurodegenerative disease that presents with motor deficits, such as resting tremors, muscle rigidity, akinesia, and impaired postural reflexes and develops in people over the age of 60 years at a rate of approximately 1 in 100 [9,10]. Although its pathogenesis is unclear, it is caused by the loss of dopamine neurones in the substantia nigra and promotion of inflammatory responses by microglia at the lesion site [11,12].
We revealed the influence of energy production pathways, disruption of calcium (Ca) homeostasis, and endoplasmic reticulum (ER) stress pathways on the molecular mechanisms of Zn2+-induced neurotoxicity [13]. Based on our recent findings regarding the involvement of the stress-activated protein kinase/c-Jun amino terminal kinase (SAPK/JNK) pathway in the generation of reactive oxygen species (ROS) [14], we hypothesised that these molecular pathways are related to Zn2+-induced neurotoxicity.
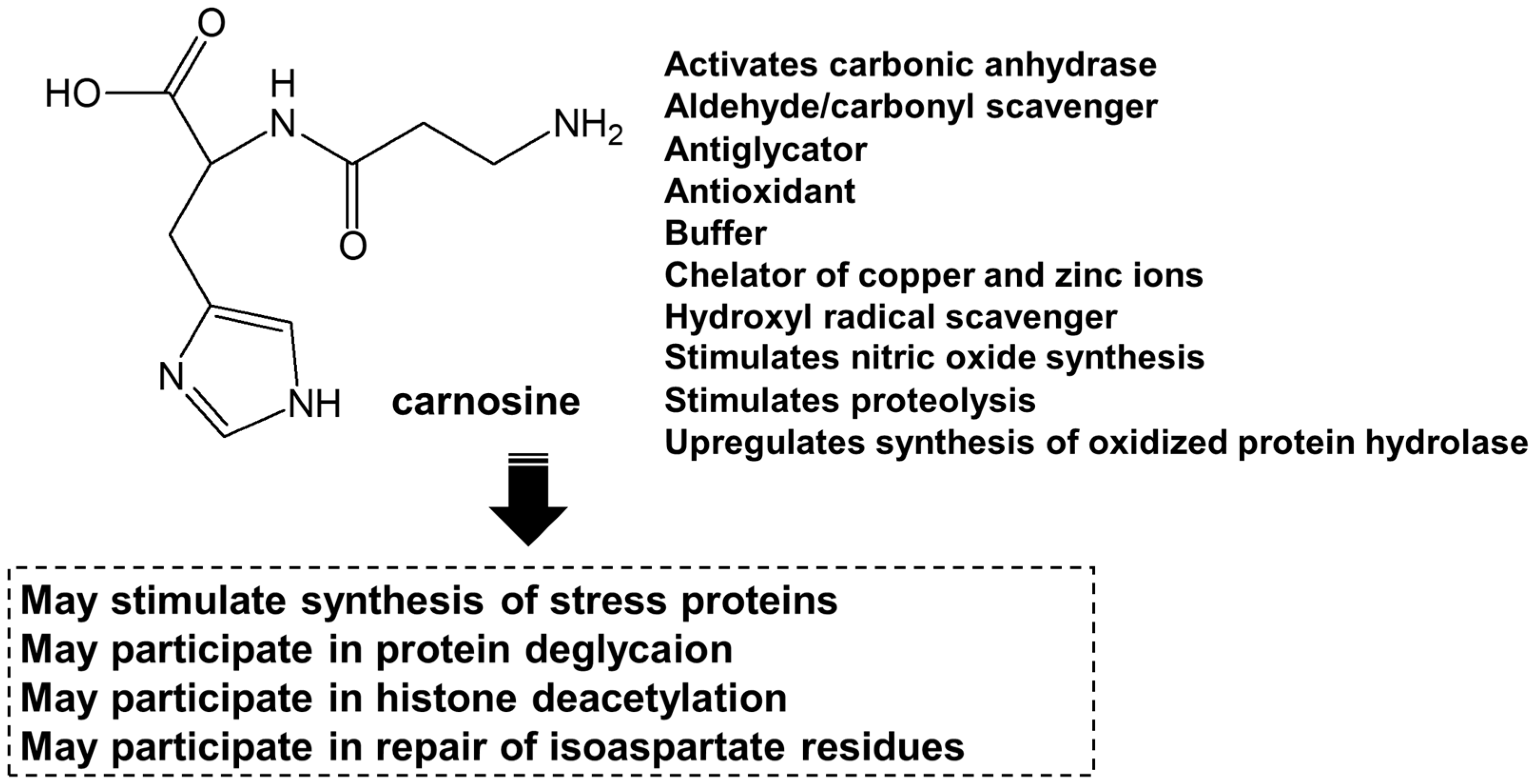
Substances that reduce Zn2+-induced neurotoxicity are potential agents for the treatment or prevention of VD [15]. During the search for a protective substance against Zn2+-induced neurotoxicity, we found that carnosine (β-alanylhistidine) exhibited a marked inhibitory effect on Zn2+-induced neurotoxicity and proposed that it is a candidate drug for the treatment of VD [16]. Carnosine is an endogenous dipeptide with various useful properties such as antioxidant, anti-glycation and anti-cross-linking (Figure 1) [17,18]. Carnosine accumulates abundantly in the skeletal muscle and olfactory bulb. As the olfactory bulb is the gateway for external information and substances, it likely protects neurones from external toxins and acts as an endogenous protector against damage and aging. Carnosine levels change during development and decrease with age [19]. This may be associated with an increased risk of neuropathy with aging. Carnosine supplementation in older adults may reduce this risk. In this article, based on our own and other previous studies, we focus on the molecular mechanisms of Zn2+-induced neurotoxicity and the properties of carnosine against this disease and discuss potential agents for VD.
Figure 1.
Structures of carnosine and its related compounds, and roles of carnosine. The various functions of carnosine in mammals.
Figure 1.
Structures of carnosine and its related compounds, and roles of carnosine. The various functions of carnosine in mammals.
2. Zn neurotoxicity
2.1. Usefulness of GT1-7 cells in the study of Zn2+-induced neurotoxicity
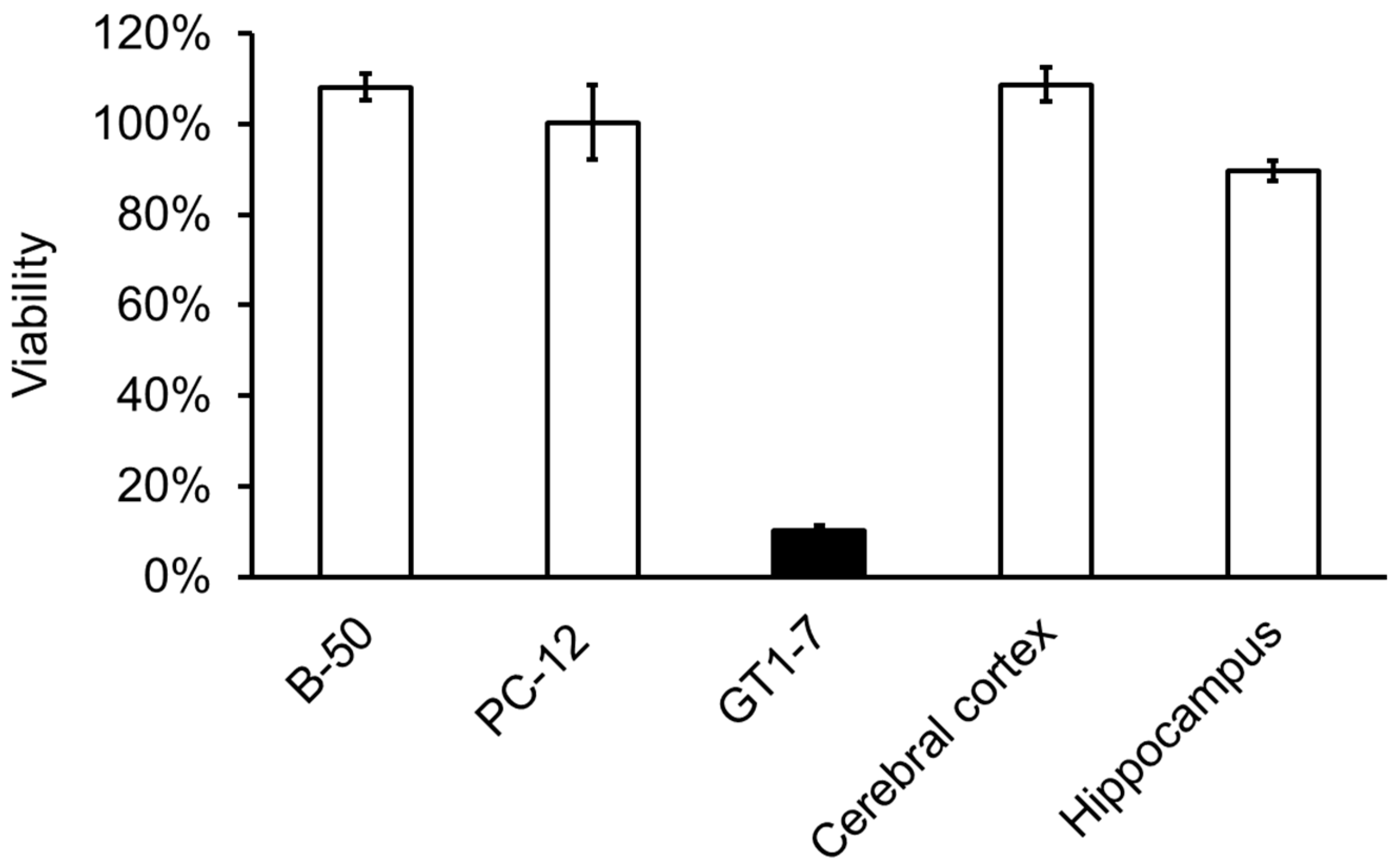
It has been recognised that abnormalities in Zn homeostasis (e. g., excess or deficiency) are involved in neurological diseases such as VD [2]. Understanding the molecular mechanism of neuronal cell death induced by Zn2+, which accounts for a large amount of Zn in the brain, is important for elucidating the pathogenesis of VD and developing therapeutic agents. However, it is difficult to examine Zn2+-induced neuronal cell death using primary cultured neurones of the cerebral cortex and hippocampus, which are used to study cell death in the brain, or PC-12 cells, a pheochromocytoma cell line. Zn2+ is released along with glutamate upon glutamatergic neuronal excitation [20]. As glutamate also causes neurotoxicity, it is difficult to distinguish between the effects of Zn2+ and glutamate in cells with glutamate receptors. We have shown that Zn2+ causes cell death in immortalised hypothalamic neurone GT1-7 cells [21,22]. In these studies, GT1-7 cells were more vulnerable to Zn2+ cytotoxicity than other neuronal cells, including primary cultures of rat cortical and hippocampal neurones, PC-12 cells, and B-50 cells. (Figure 2). GT1-7 cells possess neuronal characteristics such as neurite extension, secretion of gonadotropin-releasing hormone (GnRH), and expression of neurone-specific proteins and receptors, including microtubule-associated protein 2, tau protein, neurofilament, synaptophysin, GABAA receptors, dopamine receptors, and L-type Ca2+ channels [23]. In contrast, GT1-7 cells lack or possess low levels of ionotropic glutamate receptors and exhibit little cytotoxicity to glutamate [24]. These properties suggest that the GT1-7 cell line is an excellent model for investigating Zn2+-induced neurotoxicity.
Figure 2.
Apoptotic death of GT1-7 cells after exposure to Zn2+. Viability of various neuronal cells after exposure to Zn2+. Cultured neuronal cells (GT1-7, PC-12, and B-50 cells [a neuroblastoma cell line], primary cultured neurones of the rat cerebral cortex, and primary cultured neurones of the rat hippocampus) were administered 50 μM of ZnCl2. After 24 h, cell viability was analysed by the WST-1 method.
Figure 2.
Apoptotic death of GT1-7 cells after exposure to Zn2+. Viability of various neuronal cells after exposure to Zn2+. Cultured neuronal cells (GT1-7, PC-12, and B-50 cells [a neuroblastoma cell line], primary cultured neurones of the rat cerebral cortex, and primary cultured neurones of the rat hippocampus) were administered 50 μM of ZnCl2. After 24 h, cell viability was analysed by the WST-1 method.
2.2. Molecular mechanism of Zn2+-induced GT1-7 cytotoxicity
Zn2+-treated GT1-7 cells are positive for transferase-mediated biotinylated UTP nick-end labelling, indicating that Zn2+ induces apoptosis in GT1-7 cells [21,22]. Studies using DNA microarray analysis and real-time PCR (RT-PCR) have revealed that the administration of Zn2+ to GT1-7 cells induces the expression of various genes, including in metal-related genes (Zn transporter 1 [ZnT-1], metallothionein [MT]1), and MT2), ER stress-related genes, and Ca2+ signalling transmission-related genes [25]. The administration of Zn2+ to GT1-7 cells also increased intracellular Ca2+ levels ([Ca2+]i). Apoptosis is inhibited by pyruvate, citrate, and Ca2+ channel antagonists (nifedipine, conotoxin, and Al3+) [21,22,26]. Using a high-resolution multisite video imaging system with Fura-2 as a fluorescent probe for cytosolic Ca2+, we observed the changes in [Ca2+]i after exposure to Zn2+ [22]. This revealed that pretreatment with Al3+, which acts as various kinds of Ca2+ channel blocker [27], suppressed the elevation of the [Ca2+]i levels in Zn2+-treated GT1-7 cells. Al has difficulty entering cells without a membrane-permeable chelator, and Al3+ does not affect the viability of GT1-7 cells under these experimental conditions [28]. Zn2+-induced GT1-7 cell death may be attenuated by Al3+, which suppress the elevation of the [Ca2+]i level. Therefore, Ca2+ homeostasis is likely to be involved in Zn2+ neurotoxicity.
We previously showed that the energy substrates pyruvate and citrate salts inhibit Cu2+- and Zn2+-induced GT1-7 cell death [29]. The coexistence of pyruvate and citrate did not affect the [Ca2+]i, intracellular Zn2+ levels ([Zn2+]i), or MT mRNA levels. Therefore, it is unlikely that pyruvate and citrate attenuated Cu/Zn-induced neurotoxicity by chelating Cu2+ and Zn2+. Shelline et al. reported that Zn exposure decreased the nicotinamide adenine dinucleotide (NAD+) and ATP levels in cultured cortical neurones, and treatment with pyruvate restored the NAD+ levels [30,31]. Pyruvate administration also attenuated post-ischaemic neuronal cell death in vivo [32]. Imaging studies using Zn2+-sensitive fluorescent dyes and mitochondrial markers have revealed that Zn2+ is localised within mitochondria [33]. Zn2+ has been reported to inhibit various mitochondrial enzymes and intracellular trafficking of mitochondria. Taken together, these results suggest that energy deficiency and inhibition of mitochondrial glycolysis are involved in Zn2+ neurotoxicity [28].
DNA microarray analysis revealed that Zn2+ markedly increased the expression of ER stress-related genes, including CCAAT enhancer-binding protein homologous protein (CHOP) and growth arrest and DNA damage-induced gene 34 (GADD34) [34,35]. The ER acts as an intracellular Ca2+ reservoir and is involved in the regulation of [Ca2+]i. Therefore, an increase in the [Ca2+]i is associated with a decrease in the Ca2+ level in the ER. A decreased Ca2+ level in the ER is thought to cause ER stress because it leads to the decreased function of molecular chaperones and enzymes that bind Ca2+ [36]. The increase in the [Ca2+]i in GT1-7 cells induced by Zn2+ administration appears to be closely related to the upregulation of these ER stress-related factors. ER stress is associated with the accumulation of unfolded and misfolded proteins and is involved in various neurological diseases such as cerebral ischaemia, AD, and prion disease (PD) [37]. Three signalling proteins (ER stress sensors) —inositol-requiring enzyme 1α (IRE1α), protein kinase R-like ER kinase (PERK), and activating transcription factor (ATF) 6— are activated by sensing ER stress [38]. IRE1α, PERK, and ATF6 activate diverse signalling pathways. Phosphorylation of the α-subunit of eukaryotic translation initiation factor 2α regulates ATF4 translation via PERK. ATF4 is a transcription factor that promotes CHOP and GADD34. In addition to these factors, whose expression was confirmed to be induced by Zn2+ administration in DNA microarray studies, we examined the expression of other ER stress-related genes such as immunoglobulin binding protein (Bip), ER degradation-enhancing α-mannosidase-like protein (EDEM), spliced X-box binding protein-1 (sXBP1), glucose-regulated protein 94 (GRP94), and protein disulfide isomerase (PDI) by Zn2+ using RT-PCR. RT-PCR confirmed that Zn2+ administration to GT1-7 cells induced the expression of activity-regulated cytoskeleton (Arc), CHOP, GADD34, and ATF4. The induction of the expression of metal-related genes ZnT-1, MT1, and MT2 was also confirmed, whereas other ER stress-related genes, including Bip, EDEM, sXBP1, GRP94, and PDI, showed no significant changes. Furthermore, dantrolene, an inhibitor of ER stress, attenuated Zn2+-induced GT1-7 cytotoxicity [25]. These results strongly suggested that PERK-related pathways are involved in Zn2+-induced ER stress.
In addition to Zn, trace elements such as iron (Fe), copper (Cu), and manganese (Mn) are distributed at different levels in various parts of the brain and maintain the normal structure and function [39,40]. Among these metals, we showed that the presence of Cu2+ markedly exacerbated Zn2+-induced GT1-7 cytotoxicity [25]. Cu2+ alone did not affect the gene expression levels of Arc, CHOP, and GADD34 but significantly enhanced the induction of these factors by Zn2+. Furthermore, western blotting showed that the co-administration of Zn2+ and Cu2+ significantly increased the amount of CHOP protein. CHOP is involved in the initiation of the apoptotic cascade [41] and the activation of GADD34, which reportedly increases after traumatic brain injury [42]. We also found that the antioxidant thioredoxin-conjugated human serum albumin (HSA-Trx) attenuated Cu2+- and Zn2+-induced neuronal cell death [43]. Zn exists only as Zn2+, whereas Cu is a redox-active metal that exists as oxidised Cu2+ and reduced Cu+. Cu2+ administration induces ROS generation in GT1-7 cells, whereas Zn2+ alone does not induce ROS generation or affect Cu2+-induced ROS generation [14]. It is widely known that oxidative stress is involved in various neurodegenerative diseases, and ROS that cause oxidative stress adversely affects many signalling pathways, such as SAPK/JNK-related and ER-related pathways [44,45,46,47]. The co-administration of Cu2+ and Zn2+ to GT1-7 cells activated SAPK/JNK, phospho-c-Jun, and phospho-ATF2 downstream of the SAPK/JNK pathway. Furthermore, SP600125, an inhibitor of the SAPK/JNK signalling pathway, significantly suppresses Cu2+- and Zn2+-induced SAPK/JNK signalling pathway activation and neuronal cell death [14]. In addition, suppression of Cu2+ and Zn2+ cytotoxicity by HSA-Trx inhibits SAPK/JNK signalling pathway activation and ROS production [43]. Furthermore, the endogenous selenium (Se)-containing amino acid selenomethionine (Se-Met) induces glutathione peroxidase, blocks ROS production, significantly inhibited CHOP induction, and inhibites Cu2+- and Zn2+-mediated cytotoxicity [48]. Cu2+ triggers ROS production, which may enhance Zn2+ cytotoxicity by inducing the SAPK pathway and ER stress. These findings indicated that Zn2+ may be significantly involved in the ER stress pathway. Figure 3 shows our hypotheses regarding Zn2+-induced neuronal death (and the protective effect of carnosine, which will be discussed later).
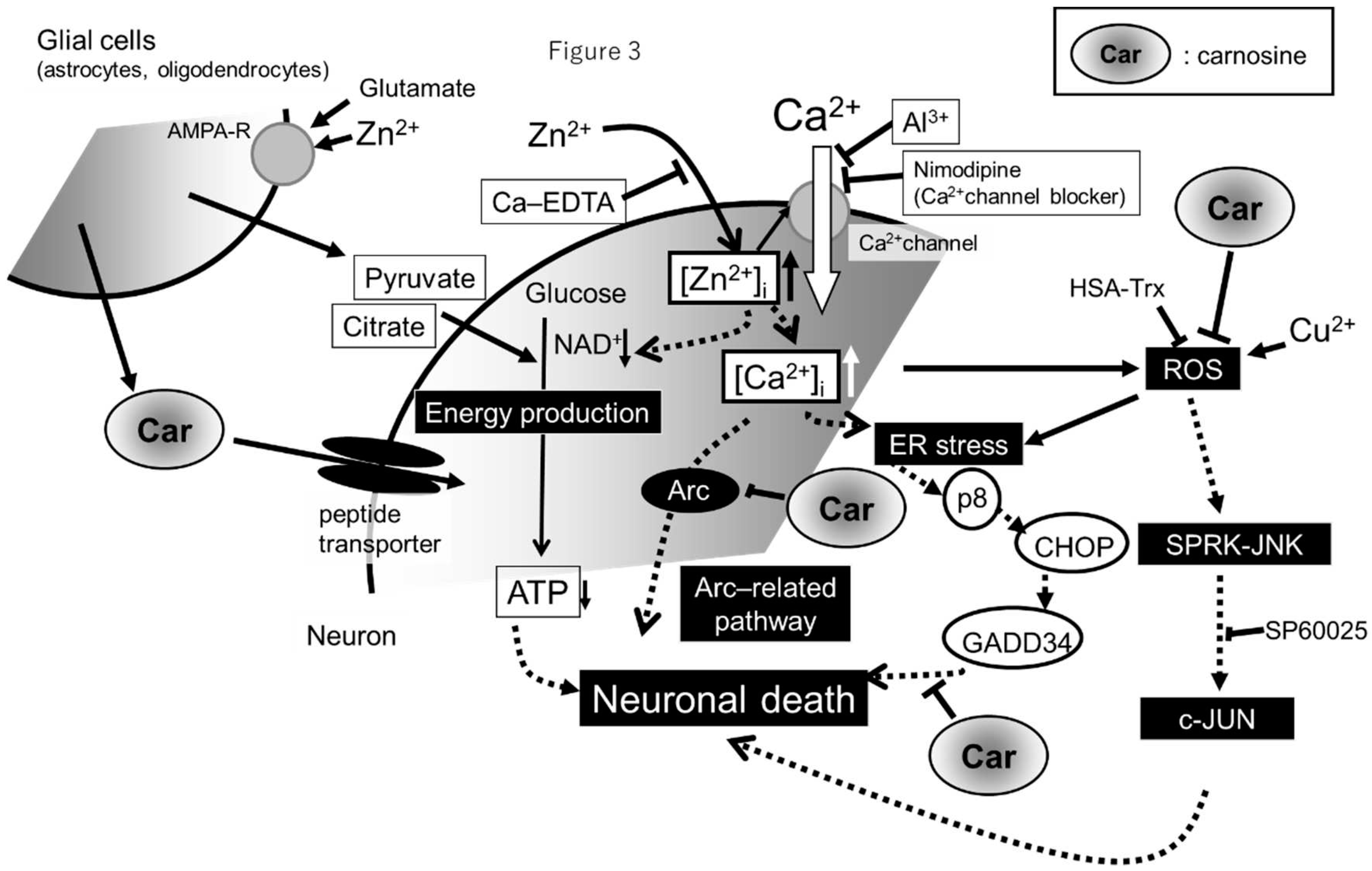
Figure 3.
Hypothetical illustration of the molecular mechanism underlying the protective effect of carnosine in preventing zinc-induced neuronal death. Zn2+ is stored in presynaptic vesicles and is secreted into the synaptic cleft during ischaemia. Excess secreted Zn2+ translocates into the cell and can cause disruption of Ca2+ homeostasis, mitochondrial energy failure, endoplasmic reticulum (ER), and oxidative stress and consequently apoptotic neuronal death. Co-exposure to Zn2+ and Cu2+, which are stored in presynaptic vesicles and secreted during ischaemia, potentiates these effects. These pathways are inhibited by Zn2+ chelators (Ca-EDTA), Ca2+ channel blockers (Al3+ and nimodipine), energy substrates (pyruvate and citrate), SAPK/JNK signalling pathway inhibitor (SP600125), and antioxidants (HSA-Trx). Carnosine inhibits the ER stress-related, Arc-related apoptotic, and ROS pathways. Carnosine is synthesised in glial cells, secreted in response to glutamate and Zn2+ stimulation, and protects neurones from Zn2+ neurotoxicity.
Figure 3.
Hypothetical illustration of the molecular mechanism underlying the protective effect of carnosine in preventing zinc-induced neuronal death. Zn2+ is stored in presynaptic vesicles and is secreted into the synaptic cleft during ischaemia. Excess secreted Zn2+ translocates into the cell and can cause disruption of Ca2+ homeostasis, mitochondrial energy failure, endoplasmic reticulum (ER), and oxidative stress and consequently apoptotic neuronal death. Co-exposure to Zn2+ and Cu2+, which are stored in presynaptic vesicles and secreted during ischaemia, potentiates these effects. These pathways are inhibited by Zn2+ chelators (Ca-EDTA), Ca2+ channel blockers (Al3+ and nimodipine), energy substrates (pyruvate and citrate), SAPK/JNK signalling pathway inhibitor (SP600125), and antioxidants (HSA-Trx). Carnosine inhibits the ER stress-related, Arc-related apoptotic, and ROS pathways. Carnosine is synthesised in glial cells, secreted in response to glutamate and Zn2+ stimulation, and protects neurones from Zn2+ neurotoxicity.
2.3. neurodegenerative diseases and Zn
Based on the aforementioned results, the hypotheses regarding Zn2+-related neurotoxicity are as follows (Figure 3). Normally, Zn2+ and Cu2+ are released into the synaptic cleft upon neuronal excitation and regulate signal transduction [20]. Secreted Zn2+ and Cu2+ undergo rapid reuptake into presynaptic neurones via the Zn transporter or CTR1, thereby maintaining the level of these ions in the synaptic cleft. However, under conditions such as transient global cerebral ischaemia, prolonged neuronal excitation occurs in major parts of the brain, and Zn2+ and Cu2+ are released from synaptic vesicles into the synaptic cleft and translocate to the same neurones in large amounts. Increased [Zn2+]i inhibit of mitochondrial energy production mechanisms and increases the [Ca2+]i. Impaired cellular protein folding due to energy depletion causes the accumulation of defective proteins in the ER. An increase in the [Ca2+]i induces ROS generation. This potentiates ER stress and/or the SAPK/JNK pathways leading to apoptotic neuronal cell death.
There is increasing evidence that Zn2+-mediated postischaemic neuronal cell death is involved in neurodegeneration after stroke or ischaemia [39,49,50,51]. VD is a disease associated with such neurodegeneration [8,52]. After transient global ischaemia, blockage of blood flow and concomitant oxygen and glucose deprivation induce abnormal neuronal excitation in most parts of the brain, followed by an excessive release of glutamate into synaptic clefts. A continuous influx of large amounts of Ca2+ is triggered, causing delayed cell death of vulnerable neurones in the hippocampus or cerebral cortex, leading to the development of infarcts and, ultimately cognitive impairment and VD. Epidemiological studies have reported that approximately 30% of patients with stroke develop symptoms of dementia 3 years later [53]. Under ischaemic conditions, significant amounts of Zn2+ (approximately 300 μM) have been reported to be released into the synaptic cleft along with glutamate after membrane depolarisation [54]. Furthermore, there is a report showing that Zn accumulates in apoptotic neurones in the hippocampus after ischaemia [55]. The administration of calcium ethylenediaminetetraacetate (Ca-EDTA), a membrane-impermeable Zn2+ chelator, protected hippocampal neurones and reduced infarct volume after transient global ischaemia in experimental animals [56]. Kitamura et al. revealed an increase in the extracellular Zn2+ levels in rats with transient middle cerebral artery occlusion using microdialysis [57]. Additionally, Zn2+ contributes to increased blood-brain barrier (BBB) permeability following ischaemia [58].
Zn2+-mediated neuronal cell death may be involved in the pathogenesis of PD [3]. Dopaminergic neuronal shedding and microglial activation, which are implicated in the pathogenesis of PD, have been observed in animal models of PD established using 6-hydroxydopamine (6-OHDA) or 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine [59]. ROS derived from 6-OHDA, which are taken up through dopamine transporters, intraneuronal 6-OHDA autoxidation, extracellular 6-OHDA autoxidation, and microglial activation are the molecular mechanisms responsible for 6-OHDA-induced dopaminergic degeneration [60]. 6-OHDA is readily oxidised and generates several reactive and cytotoxic products, such as quinones, H2O2, 5,6-dihydroxyindole, superoxide anion radicals, hydroxyl radicals, and singlet oxygen [61]. ROS derived from paraquat, an herbicide that is also taken up by dopamine transporters, lead to glutamate exocytosis via transient receptor potential melastatin 2 cation channel activation in the substantia nigra, followed by nigral dopaminergic degeneration via intracellular Zn2+ dysregulation [62,63]. From the intracellular ROS derived from 6-OHDA and PQ, H2O2 readily passes through cell membranes via aquaporin channels [64]. H2O2 elevation in the extracellular compartment excites glutamatergic neurone terminals [65], and Zn2+ is released along with glutamate. Additionally, we showed that treatment with 6-OHDA induces integrated stress-related genes, such as CHOP, GADD34, and ATF4 in GT1-7 cells [66]. Increase of Zn2+ release by 6-OHDA may be related to induction of these factors.
Zn2+ entry and an increased [Zn2+]i in other words ‘Zn translocation’ are key events in Zn2+-induced neurotoxicity. There are three major pathways for Zn2+ entry: voltage-gated Ca2+ channels, N-methyl-D-aspartic acid (NMDA)-type glutamate receptors, and AMPA/kainate-type glutamate receptors (A/K-R) [28]. Under normal conditions, most hippocampal neurones express AMPA receptors containing GluR2 subunits that are poorly permeable to Ca2+ and Zn2+. However, after ischaemia, there is a sharp decrease in GluR2 expression, and neurones express Ca2+-permeable AMPA receptors (Ca-A/KR). As the permeability of Zn2+ and Ca2+ through the Ca-A/KR channels is greater than that through the NMDA receptor channels, the increased expression of Ca-A/KR channels enhances Ca2+ and Zn2+ toxicity. Zn2+ is also thought to be involved in the transcriptional regulation of Ca-A/KR channels, as Ca-EDTA attenuates ischaemia-induced downregulation of the GluR2 gene [56]. These Zn2+-mediated neuronal cell death events can be explained by the Zn2+-mediated neurotoxicity hypothesis. Considering the involvement of Zn2+ in transient global ischaemia, substances that prevent Zn2+-induced neuronal cell death may be potential candidates for the prevention or treatment of postischaemic neurodegeneration and, ultimately, the treatment of VD and PD. We used extracts of various agricultural products, such as vegetables, fruits and fish, and found that extracts of the Japanese eel (Anguilla japonica), mango fruit (Mangifera indica L.), and round herring (Etrumeus teres) protected GT1-7 cells from Zn2+-induced neurotoxicity. The active fractions were separated from these extracts using high-performance liquid chromatography (HPLC) and the structures of the components were analysed by LC mass spectrometry. The active compounds included carnosine, citric acid, and histidine [16,34,67].
3. Carnosine can be a therapeutic agent for cerebrovascular dementia
3.1. Carnosine
Carnosine is a natural dipeptide composed of β-alanine and L-histidine (His). Carnosine and its analogues (anserine [1-methylcarnosine] and homocarnosine) are present in most vertebrate tissues, including those of birds, fish, and mammals including humans [17,18]. In particular, carnosine is present in high levels in animals and fish that exercise frequently, such as horses, chickens, bonitos, and whales. In humans, the carnosine levels have been reported to be higher in males, decrease with age, and be diet dependent, with a vegetarian diet reducing carnosine levels in the skeletal muscle [68,69]. Similar to creatine and ATP, this dipeptide is also found in some muscle net at 50-200 mM [70,71]. In animals, factors such as trauma, shock, starvation, and injection adversely affect the carnosine levels in muscle tissue. Infection and trauma may be associated with dysregulation of cellular Ca and myocardial depression. Carnosine may also play a role in the regulation of cardiac cell contractility and [Ca2+]i [72].
Carnosine is abundant in animals and fish that exercise frequently, such as horses, chickens, bonito, and whales. For example, intramuscular carnosine concentrations are 6–10 times higher in horses than in humans [73]. We analysed the carnosine content in thoroughbred horse muscle and found that carnosine content was associated with the muscle fibre type [74]. Among the five equine muscle tissues (radius flexor, gill triceps brachii, masseter, gluteus medius, and sternocleidomastoid), the gluteus medius had the highest carnosine concentration. The glutaeus medius is enriched with type IIa (fast-twitch oxidative glycolytic muscle fibres) and IIx (fast-twitch glycolytic muscle fibre) fibres [75]. As these muscle fibres are primarily used during high-intensity exercise, carnosine may play an important role in high-intensity exercise. Due to the alkaline nature of carnosine (pKa = 7.01), it is thought to play a significant role in intracellular buffering [76]. This buffering action is thought to play an important role in maintaining pH balance against the production of lactic acid, which causes muscle contraction fatigue due to acidosis during high-intensity anaerobic exercise and a decrease in intracellular pH. Therefore, the muscle carnosine concentration may be positively related to exercise performance [73,77]. Highly trained athletes have higher carnosine levels compared to untrained individuals [78]. Furthermore, dietary supplementation with carnosine or β-alanine increases the concentration of muscle carnosine and delays fatigue during high-intensity exercise [78].
In addition, carnosine possesses various functions, such as anti-oxidation, anti-glycation, anti-cross-linking, and metal chelation, which mediate its beneficial effects in vivo [18]. Carnosine scavenges both reactive oxygen- and nitrogen-containing unpaired electrons and inhibits lipid oxidation through a combination of free radical scavenging and metal chelation. It also inhibits the Maillard reaction, which produces many end-products, especially advanced glycation end-products, that contribute to the development of various senile diseases, such as AD, vascular sclerosis, atherosclerosis, and osteoarthritis. In addition, carnosine exhibits anti-crosslinking properties that inhibit protein oligomerisation. N-acetylcarnosine is used as a treatment for cataracts because carnosine inhibits α-crystal fibrosis of the lens [79]. Polaprezinc, a Zn-carnosine complex, is effective in repairing gastrointestinal ulcers and other lesions [80]. Polaprezinc is also used in Zn supplementation therapy and shows protective effects against cadmium-induced lung injury [81].
3.2. Carnosine in the brain
Carnosine and homocarnosine have been detected in the mammalian brain, but anserine has not yet been detected [82]. β-alanine is readily transported throughout the brain by Na+-dependent-amino acid transport system(s) and acts as a neuromodulator/neurotransmitter or might, in theory, form carnosine [83]. Additionally, the carnosine transporter (peptide transporter 2) is expressed in some rat neuronal cells [84,85]. This suggests the putative ability of carnosine to cross the BBB. In the brain, carnosine is present in the olfactory bulb [86] and has been reported to be secreted from oligodendrocytes upon stimulation with glutamate [87]. Boldyrev et al. reported that carnosine is mainly present in the neurones or glial cells of the olfactory bulb, with levels in the olfactory bulb exceeding 1,000 μmol/kg [18]. We developed a quantitative analysis method for carnosine and its analogues using an HPLC system equipped with a carbon column (Hypercarb™) [88] and investigated the distribution of these compounds in the rat brain [89]. The rat brain contains significant amounts of carnosine in the olfactory bulb but less carnosine in the cerebral cortex and cerebellum, and anserine was not detected in any region tested. It has also been revealed that the carnosine levels in the olfactory bulb increase from the foetal stage to maturity. In contrast, the homocarnosine levels show no change with postnatal age. These results are similar to those of the previous studies described earlier. Biffo et al. showed that carnosine is located in olfactory receptor neurones, specifically in the perinuclear membrane and neurites, including the axons and synaptic terminals of the olfactory bulb [90]. Carnosine is rapidly synthesised and transported to the olfactory bulb via axonal transport [91]. In primary olfactory neurones, carnosine synthase activity is decreased by denervation and is restored by regeneration [92,93]. Carnosine in the olfactory bulb may be localised mainly in the sensory neurones [19].
Because carnosine forms complexes with Ca2+, Cu2+, and Zn2+ [94,95], it plays an important role in regulating Zn2+ homeostasis at synapses in neural tissues, especially in the carnosine- and Zn-rich olfactory lobes [18]. Disease-associated proteins (e.g., AβP, prion protein and α-synuclein) are thought to be central to the pathogenesis of various neurodegenerative diseases known as ‘structural diseases’ including AD, DLB, and PD. Carnosine interferes with cross-linking and subsequent conformational changes through its anti-cross-linking action [40]. Corona et al. reported that the administration of carnosine inhibited AβP deposition and improved the learning ability of AD model mice [96]. Carnosine prevents oxidative stress and inflammation induced by AβP [97]. We previously reported that carnosine alleviates neuronal cell death by changing the conformation of the prion protein fragment peptide (PrP106-126) [98]. In addition, carnosine has been reported to reduce Mn-induced neurotoxicity [99]. Based on these beneficial properties, carnosine is thought to act as a ‘gatekeeper’ or a ‘neuroprotectant’ in the brain [100].
3.3. Carnosine suppresses Zn-induced neuronal death
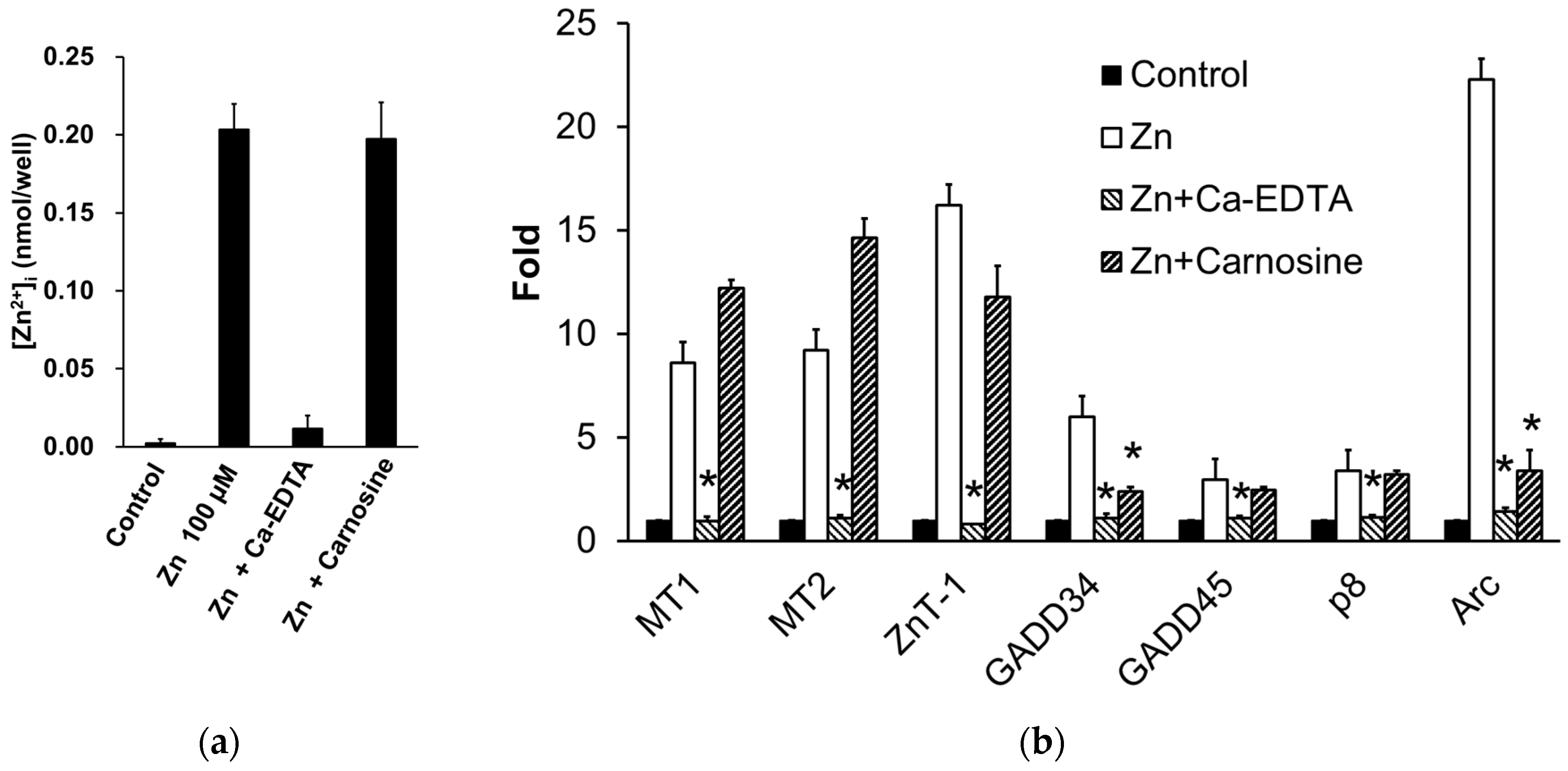
We found that carnosine has a protective effect on neurones from Zn2+-induced neurotoxicity (Figure 4) and are investigating the mechanism. Our previous study showed that carnosine did not affect [Zn2+]i or the expression of metal-related genes, such as ZnT-1 (Figure 5a,b) [35]. Although carnosine can chelate Zn2+, it does not inhibit Zn2+ translocation by binding to extracellular Zn2+. In contrast, we found that carnosine inhibited the Zn2+-induced expression of ER stress-related genes, such as GADD34 and CHOP, and the Ca2+-related gene Arc (activity-related cytoskeletal protein) (Figure 5b) [35]. ER stress has been implicated in the development of various neurodegenerative diseases such as AD, PD, and ischaemia-induced neurodegeneration [34,101]. GADD34, a gene encoding a sensor protein for ER stress, is induced by DNA damage and is thought to be involved in DNA repair and tumorigenesis [102]. Arc resides in the dendrites and encodes a protein that plays an important role in synaptic plasticity and memory consolidation. Arc expression is induced by increased neuronal activity in response to learning and brain-derived neurotrophic factor [103]. Carnosine attenuates neurodegeneration induced by ER stressors such as thapsigargin and tunicamycin [35]. Thus, carnosine may protect neurones from Zn2+, not by inhibiting Zn2+ translocation, but by affecting ER stress and Arc-related pathways (Figure 3). Studies in experimental animals have suggested that carnosine protects against ischaemia-induced neurodegeneration in vivo [104,105,106,107]. Carnosine reduced 6-OHDA-induced neuronal cell death and inflammatory responses in GT1-7 cells. Specifically, carnosine markedly inhibited 6-OHDA-induced upregulation of the stress-related genes, such as Chop, GADD34, and Atf4. Furthermore, it suppresses 6-OHDA-induced activation of the SAPK/JNK signalling pathway by inhibiting ROS production [66]. Therefore, it may effectively prevent the onset and/or exacerbation of PD.
Figure 4.
Protective activity against Zn2+-induced neurotoxicity in GT1-7 cells. GT1-7 cells are treated with ZnCl2 (30 μM) with or without various carnosine levels. After 24 h, the viability of GT1-7 cells is measured using the WST-1 assay. The data are presented as means ± S.E.M., n = 6. * p < 0.01, ** p < 0.005.
Figure 4.
Protective activity against Zn2+-induced neurotoxicity in GT1-7 cells. GT1-7 cells are treated with ZnCl2 (30 μM) with or without various carnosine levels. After 24 h, the viability of GT1-7 cells is measured using the WST-1 assay. The data are presented as means ± S.E.M., n = 6. * p < 0.01, ** p < 0.005.
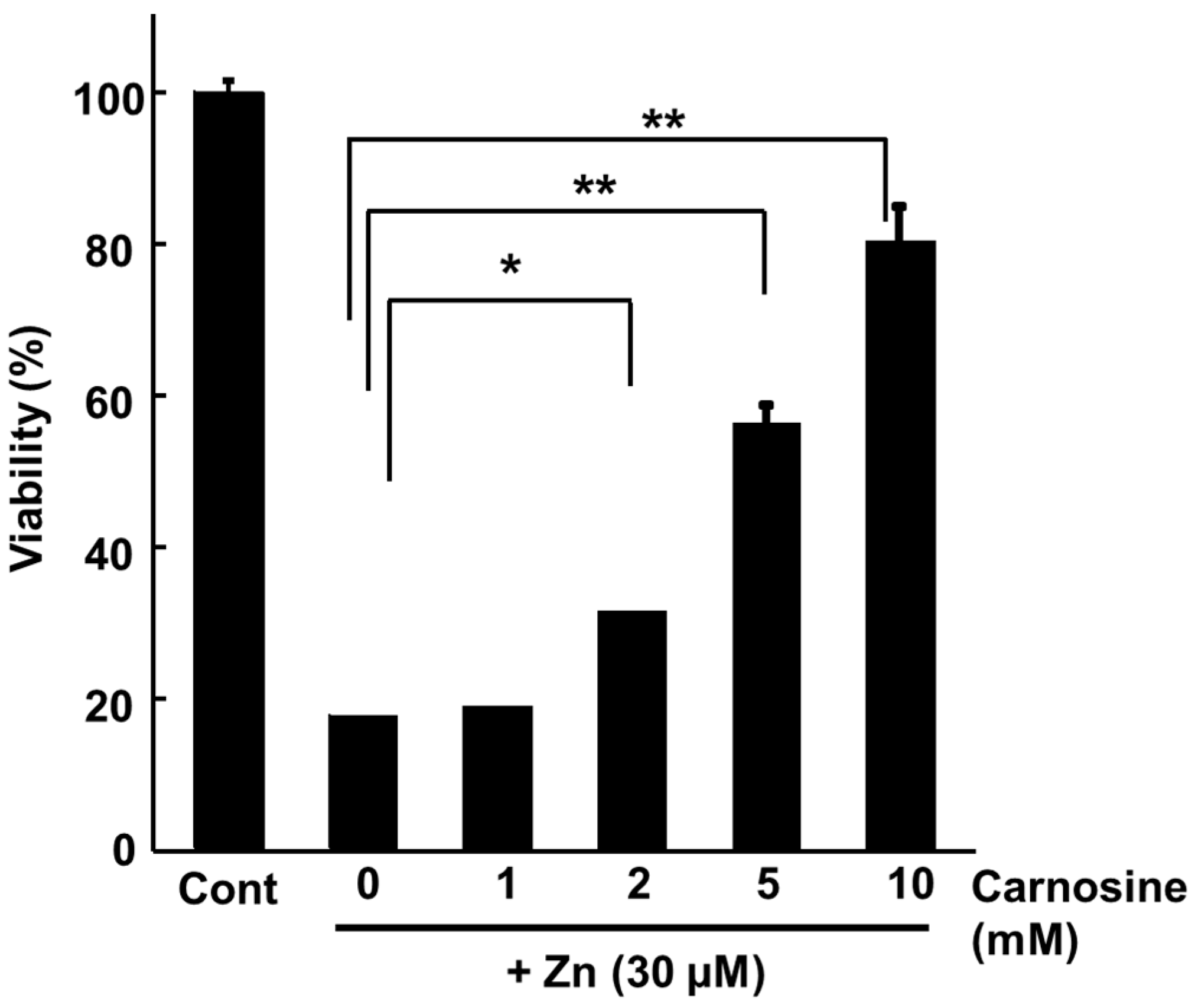
Figure 5.
Effects of carnosine on the expression of Zn-induced factors. (a) Effects of carnosine and anserine on Zn influx into GT1-7 cells. GT1-7 cells are treated with 100 μM of ZnCl2 for 30 min in the presence or absence of Ca-EDTA (0.2 mM) or carnosine (2.0 mM), and [Zn2+]i was measured using a Metallo Assay Zinc LS kit (Metallogenics, Chiba, Japan) according to the manufacturer’s instructions. Data are presented as means ± S.E.M., n = 3. (b) Effects of carnosine on Zn-induced gene expression. GT1-7 cells are treated with 50 μM ZnCl2 for 6 h in the presence or absence of Ca-EDTA (0.5 mM) or carnosine (5.0 mM). The expression of metal-, ER stress-, and Arc-related genes is analysed by RT-PCR, and the gene expression levels are normalised to β-actin. Data are presented as the mean ± S.E.M., n = 3. ** p < 0.01 versus the Zn group.
Figure 5.
Effects of carnosine on the expression of Zn-induced factors. (a) Effects of carnosine and anserine on Zn influx into GT1-7 cells. GT1-7 cells are treated with 100 μM of ZnCl2 for 30 min in the presence or absence of Ca-EDTA (0.2 mM) or carnosine (2.0 mM), and [Zn2+]i was measured using a Metallo Assay Zinc LS kit (Metallogenics, Chiba, Japan) according to the manufacturer’s instructions. Data are presented as means ± S.E.M., n = 3. (b) Effects of carnosine on Zn-induced gene expression. GT1-7 cells are treated with 50 μM ZnCl2 for 6 h in the presence or absence of Ca-EDTA (0.5 mM) or carnosine (5.0 mM). The expression of metal-, ER stress-, and Arc-related genes is analysed by RT-PCR, and the gene expression levels are normalised to β-actin. Data are presented as the mean ± S.E.M., n = 3. ** p < 0.01 versus the Zn group.
3.4. Potential uses of carnosine and its derivative as supplements
Orally administered carnosine is widely believed to be rapidly degraded to β-alanine and histidine by circulating carnosinases (CN1). However, carnosine or β-alanine supplementation has been reported to increase the carnosine levels in the brain [108,109]. Carnosine supplementation (40 mg/day) has been shown to be effective in treating patients with severe depressive disorder [110]. In addition, carnosine/anserine supplementation (750 mg of anserine and 250 mg of carnosine per day) has been reported to improve episodic memory [111] and mild cognitive impairment [112] in older adults. Therefore, dietary carnosine or related amino acids may be synthesised into carnosine in the brain and taken up into cells by oligopeptide transporters such as PEPT2, PHT1, and PHT2 [113]. A recent epidemiological study reported an inverse correlation between the serum β-alanine levels and aetiology of dementia [114]. Considering these factors and the fact that the carnosine levels in the body decrease with age [115], carnosine replacement therapy may be beneficial for VD, AD, and other neurological disorders. Finding derivatives or analogues of carnosine that are resistant to degradation by CN1 is important for exploiting its full potential. Pharmacological variants of carnosine, such as carnosinol, a reduced carnosine derivative that is resistant to CN1, have been developed and show promise for use in the treatment of metabolic diseases, such as obesity and diabetes [116]. Additionally, balenine, found in marine mammals and reptiles, is a more stable natural analogue than carnosine in vivo and has potential uses as a dietary or ergogenic supplement [117].
4. Conclusions
Our hypothesis regarding the molecular pathways involved in Zn2+-induced neurotoxicity may aid in the development of preventive and therapeutic agents for VD and PD. Based on the activity of carnosine, we published two patents for carnosine and its related compounds (D-histidine) as drugs or supplements for the prevention and/or treatment of VD [118,119]. Carnosine has many beneficial properties such as water solubility, heat inactivation, and non-toxicity, making it an excellent neuroprotective drug or supplement that benefits human health. Further studies are required to elucidate the molecular mechanisms by which carnosine prevents neurotoxicity.
Author Contributions
Conceptualization, D.M., M. K. and K.M.; Resources, D.M., M. K., K.M and K.Y.; Draft preparation, review and editing, D.M., M.K., T.O. and K.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partially supported by a Grant-in Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology of Japan. (JSPS Kakennhi grant numbers. JP 26460177 and JP 17H03197).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data supporting the findings of this study are available from the corresponding author, Dai Mizuno, upon reasonable request.
Acknowledgments
We would like to thank Editage (www.editage.com) for the English language editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hambidge, M. Human zinc deficiency. J. Nutr. 2000, 130, 1344S–1349S. [Google Scholar] [CrossRef]
- Weiss, J.H., Sensi, S.L., Koh, J.Y. Zn(2+): a novel ionic mediator of neural injury in brain disease. Trends Pharmacol. Sci. 2000, 21, 395–401. [CrossRef]
- Nishio, R., Morioka, H., Takeuchi, A., Saeki, N., Furuhata, R., Katahira, M., Chinenn, T., Tamura, H., Tamano, H., Takeda, A. Intracellular hydrogen peroxide produced by 6-hydroxydopamine is a trigger for nigral dopaminergic degeneration of rats via rapid influx of extracellular Zn2. Neurotoxicology 2022, 89, 1–8.
- World Health Organization. Dementia. Fact sheets of WHO. Available online: https://www.who.int/news-room/fact-sheets/detail/dementia (accessed on 1 June 2023).
- Cao, Q., Tan, C.-C., Xu, W., Hu, H., Cao, X.-P., Dong, Q., Tan, L., Yu, J.-T. The prevalence of dementia: a systematic review and meta-analysis. J. Alzheimer’s Dis. 2020, 73, 1157–1166. [CrossRef] [PubMed]
- Selkoe, D.J. The molecular pathology of Alzheimer’s disease. Neuron 1991, 6, 487–498. [Google Scholar] [CrossRef]
- Brás, I.C., Dominguez-Meijide, A., Gerhardt, E., Koss, D., Lázaro, D.F., Santos, P.I., Vasili, E., Xylaki, M., Outeiro, T.F. Synucleinopathies: where we are and where we need to go. J. Neurochem. 2020, 153, 433–454.
- Iadecola, C. The pathobiology of vascular dementia. Neuron 2013, 80, 844–866. [Google Scholar] [CrossRef]
- Pagano, G., Ferrara, N., Brooks, DJ., Pavese, N. Age at onset and Parkinson disease phenotype. Neurology 2016, 86, 1400–1407. [CrossRef] [PubMed]
- Kalia, LV., Lang, AE. Parkinson’s disease. Lancet 2015, 386, 896–912. [CrossRef]
- Erekat, NS. Apoptosis and its Role in Parkinson’s Disease. In Stoker, TB., Greenland, JC., Eds.; Parkinson’s Disease: Pathogenesis and Clinical Aspects. Codon Publications: Brisbane, Australia. 2018; pp. 65–82. [Google Scholar]
- Macchi, B., Di Paola, R., Marino-Merlo, F., Felice, MR., Cuzzocrea, S., Mastino, A. Inflammatory and cell death pathways in brain and peripheral blood in Parkinson’s disease. CNS Neurol. Disord. Drug Targets. 2015, 14, 313–324. [CrossRef]
- Kawahara, M., Tanaka, K.I., Kato-Negishi, M. Zinc, carnosine, and neurodegenerative diseases. Nutrients 2018, 10, 147. [CrossRef] [PubMed]
- Tanaka, K.I., Shimoda, M., Kasai, M., Ikeda, M., Ishima, Y., Kawahara, M. Involvement of SAPK/JNK signaling pathway in copper enhanced zinc-induced neuronal cell death. Toxicol. Sci. 2019, 169, 293–302. [CrossRef]
- Kawahara, M., Konoha, K., Nagata, T., Sadakane, Y. Protective substances against zinc-induced neuronal death after ischemia: carnosine as a target for drug of vascular type of dementia. Recent Pat. CNS Drug Discov. 2007, 2, 145–149.
- Sadakane, Y., Konoha, K., Nagata, T., Kawahara, M. Protective activity of the extracts from Japanese eel (Anguilla japonica) against zinc-induced neuronal cell death: carnosine and an unknown substance. Trace Nutrient Res. 2007, 24, 98–105.
- Hipkiss, A.R. On the relationship between energy metabolism, proteostasis, aging and Parkinson’s disease: possible causative role of methylglyoxal and alleviative potential of carnosine. Aging Dis. 2017, 8, 334–345. [Google Scholar] [CrossRef]
- Boldyrev, A.A., Aldini, G., Derave, W. Physiology and pathophysiology of carnosine. Physiol Rev. 2013, 93, 1803–1845. [CrossRef] [PubMed]
- De Marchis, S., Modena, C., Peretto, P., Migheli, A., Margolis, F.L., Fasolo, A. Carnosine-related dipeptides in neurons and glia. Biochemistry (Mosc) 2000, 65, 824–833.
- Frederickson, C.J., Suh, S.W, Silva, D., Frederickson, C.J., Thompson, R.B. Importance of zinc in the central nervous system: the zinc-containing neuron. J. Nutr. 2000, 130, 1471S–1483S. [CrossRef]
- Kawahara, M., Kato-Negishi, M., Kuroda, Y. Pyruvate blocks zinc-induced neurotoxicity in immortalized hypothalamic neurons. Cell Mol. Neurobiol., 2002, 22, 87–93. [CrossRef]
- Koyama, H., Konoha, K., Sadakane, Y., Ohkawara, S., Kawahara, M. Zinc neurotoxicity and the pathogenesis of vascular-type dementia: Involvement of calcium dyshomeostasis and carnosine. J. Clin. Toxicol., 2012, S3, 002.
- Mellon, P.L., Windle J.J., Goldsmith P.C., Padula C.A., Roberts J.L., Weiner R.I. Immortalization of hypothalamic GnRH neurons by genetically targeted tumorigenesis. Neuron 1990, 5, 1–10. [CrossRef]
- Mahesh, V.B., Zamorano, P., De Sevilla, L., Lewis, D., Brann, D.W. Characterization of ionotropic glutamate receptors in rat hypothalamus, pituitary and immortalized gonadotropin-releasing hormone (GnRH) neurons (GT1-7 cells). Neuroendocrinology 1999, 69, 397–407. [CrossRef]
- Tanaka, K., Kawahara, M. Copper enhances zinc-induced neurotoxicity and the endoplasmic reticulum stress response in a neuronal model of vascular dementia. Front Neurosci. 2017, 11, 58.
- Konoha, K., Sadakane, Y., Kawahara, M. Effects of gadolinium and other metal on the neurotoxicity of immortalized hypothalamic neurons induced by zinc. Biomed. Res. Trace Elem. 2004, 15, 275–277.
- Büsselberg, D., Platt, B., Haas, H.L., Carpenter, D.O. Voltage gated calcium channel currents of rat dorsal root ganglion (DRG) cells are blocked by Al3+. Brain Res. 1993, 622, 163–168. [CrossRef] [PubMed]
- Kawahara, M., Kato-Negishi, M., Hosoda, R., Imamura, L., Tsuda, M., Kuroda, Y. Brain-derived neurotrophic factor protects cultured rat hippocampal neurons from aluminum maltolate neurotoxicity. J. Inorg. Biochem. 2003, 97, 124–131. [CrossRef] [PubMed]
- Tanaka, K.-I., Shimoda, M., Kawahara, M. Pyruvic acid prevents Cu2+/Zn2+-induced neurotoxicity by suppressing mitochondrial injury. Biochem. Biophys. Res. Commun. 2018, 495, 1335–1341. [CrossRef]
- Cai, A.L., Zipfel, G.J., Sheline, C.T. Zinc neurotoxicity is dependent on intracellular NAD levels and the sirtuin pathway. Eur. J. Neurosci. 2006, 24, 2169–2176. [CrossRef]
- Lee, J.Y., Kim, Y.H., Koh, J.Y. Protection by pyruvate against transient forebrain ischemia in rats. J. Neurosci. 2001, 21, RC171. [CrossRef]
- Sheline, C.T., Behrens, M.M., Choi, D.W. Zinc-induced cortical neuronal death: Contribution of energy failure attributable to loss of NAD(+) and inhibition of glycolysis. J. Neurosci. 2000, 20, 3139–3154. [CrossRef]
- Sensi, S.L., Ton-That, D., Sullivan, P.G., Jonas, E.A., Gee, K.R., Kaczmarek, L.K., Weiss, J.H. Modulation of mitochondrial function by endogenous Zn2+ pools. Proc. Natl. Acad. Sci. USA 2003, 100, 6157–6162.
- Kawahara, M., Sadakane, Y., Koyama, H., Konoha, K., Ohkawara, S. D-histidine and L-histidine attenuate zinc-induced neuronal death in GT1-7 cells. Metallomics 2013, 5, 453–460. [CrossRef] [PubMed]
- Mizuno, D., Konoha-Mizuno, D., Mori, M., Sadakane, Y., Koyama, H., Ohkawara, S., Kawahara, M. Protective activity of carnosine and anserine against zinc-induced neurotoxicity: a possible treatment for vascular dementia. Metallomics 2015, 7, 1233–1239. [CrossRef]
- Higo, T., Kozo, H., Hisatsune, C., Nukina, N., Hashikawa, T., Hattori, M., Nakamura, T., Mikoshiba, K. Mechanism of ER stress-induced brain damage by IP(3) receptor. Neuron 2010, 68, 865–878. [CrossRef]
- Kaneko, M., Imaizumi, K., Saito, A., Kanemoto, S., Asada, R., Matsuhisa, K., Ohtake, Y. ER stress and disease: toward prevention and treatment. Biol. Pharm. Bull. 2017, 40, 1337–1343.
- Sano, R., Reed, J.C. ER stress-induced cell death mechanisms. Biochim. Biophys. Acta 2013, 1833, 3460–3470. [CrossRef]
- Becker J.S., Matusch A, Palm C, Salber D, Morton K.A., Becker J.S. Bioimaging of metals in brain tissue by laser ablation inductively coupled plasma mass spectrometry (LA-ICP-MS) and metallomics. Metallomics 2010, 2, 104–111. [CrossRef]
- Kawahara M., Kato-Negishi M, Tanaka K. Cross talk between neurometals and amyloidogenic proteins at the synapse and the pathogenesis of neurodegenerative diseases. Metallomics 2017, 9, 619–633. [CrossRef] [PubMed]
- Yang, Y., Liu, L., Naik, I., Braunstein, Z., Zhong, J., Ren, B. Transcription factor C/EBP homologous protein in health and diseases. Front Immunol. 2017, 8, 1612. [CrossRef]
- Paschen, W., Hayashi, T., Saito, A., Chan, P.H. GADD34 protein levels increase after transient ischemia in the cortex but not in the CA1 subfield: implications for post-ischemic recovery of protein synthesis in ischemia-resistant cells. J. Neurochem. 2004, 90, 694–701. [CrossRef]
- Tanaka, K.I., Shimoda, M., Chuang, V.T.G., Nishida, K., Kawahara, M., Ishida, T., Otagiri, M., Maruyama, T., Ishima, Y. Thioredoxin-albumin fusion protein prevents copper enhanced zinc-induced neurotoxicity via its antioxidative activity. Int. J. Pharm. 2018, 535, 140–147. [CrossRef] [PubMed]
- Dent, P., Yacoub, A., Contessa, J., Caron, R., Amorino, G., Valerie, K., Hagan, M.P., Grant, S., Schmidt-Ullrich, R. Stress and radiation-induced activation of multiple intracellular signaling pathways. Radiat. Res. 2003, 159, 283–300. [CrossRef]
- Dandekar, A., Mendez, R., Zhang, K. Cross talk between ER stress, oxidative stress, and inflammation in health and disease. Methods Mol. Biol. 2015, 1292, 205–214.
- Papaconstantinou, J. The role of signaling pathways of inflammation and oxidative stress in development of senescence and aging phenotypes in cardiovascular disease. Cells 2019, 8, 1383. [CrossRef]
- Konno, T., Melo, E.P., Chambers, J.E., Avezov, E. Intracellular sources of ROS/H2O2 in health and neurodegeneration: spotlight on endoplasmic reticulum. Cells 2021, 10, 233. [CrossRef] [PubMed]
- Nakano, Y., Shimoda, M., Okudomi, S., Kawaraya, S., Kawahara, M., Tanaka, K.-I. Seleno-L-methionine suppresses copper-enhanced zinc-induced neuronal cell death via induction of glutathione peroxidase. Metallomics 2020, 12, 1693–1701. [CrossRef]
- Jiang, H., Song, N., Jiao, Q., Shi, L., Du, X. Iron pathophysiology in parkinson diseases. Adv. Exp. Med. Biol. 2019, 1173, 45–66.
- Kawahara, M., Mizuno, D., Koyama, H., Konoha, K., Ohkawara, S., Sadakane, Y. Disruption of zinc homeostasis and the pathogenesis of senile dementia. Metallomics 2014, 6, 209–219. [CrossRef]
- Stork, C.J., Li, Y.V. Elevated cytoplasmic free zinc and increased reactive oxygen species generation in the context of brain injury. Acta Neurochir. Suppl. 2016, 121, 347–353.
- Kalaria, R.N. The pathology and pathophysiology of vascular dementia. Neuropharmacology 2018, 134, 226–239. [Google Scholar] [CrossRef]
- Pendlebury, S.T., Rothwell, P.M. Prevalence, incidence, and factors associated with pre-stroke and post-stroke dementia: a systematic review and meta-analysis. Lancet Neurol. 2009, 8, 1006–1018. [CrossRef] [PubMed]
- Frederickson, C.J., Klitenick, M.A., Manton, W.I., Kirkpatrick, J.B. Cytoarchitectonic distribution of zinc in the hippocampus of man and the rat. Brain Res. 1983, 27, 335–339.
- Koh, J.Y., Suh, S.W., Gwag, B.J., He, Y.Y., Hsu, C.Y., Choi, D.W. The role of zinc in selective neuronal death after transient global cerebral ischemia. Science 1996, 272, 1013–1016. [CrossRef] [PubMed]
- Calderone, A., Jover, T., Mashiko, T., Noh, K.M., Tanaka, H., Bennett, M.V., Zukin, R.S. Late calcium EDTA rescues hippocampal CA1 neurons from global ischemia-induced death. J. Neurosci. 2004, 24, 9903–9913. [CrossRef] [PubMed]
- Kitamura, Y., Iida, Y., Abe, J., Mifune, M., Kasuya, F., Ohta, M., Igarashi, K., Saito, Y., Saji, H. Release of vesicular Zn2+ in a rat transient middle cerebral artery occlusion model. Brain Res. Bull. 2006, 69, 622–625. [CrossRef]
- Qi, Z., Shi, W., Zhao, Y., Ji, X., Liu, K.J. Zinc accumulation in mitochondria promotes ischemia-induced BBB disruption through Drp1-dependent mitochondria fission. Toxicol. Appl. Pharmacol. 2019, 377, 114601. [CrossRef]
- Konnova, EA., Swanberg M. Animal Models of Parkinson’s Disease. In Stoker, TB., Greenland, JC., Eds.; Parkinson’s Disease: Pathogenesis and Clinical Aspects. Codon Publications: Brisbane, Australia. 2018; pp. 83–106.
- Rodriguez-Pallares, J., Parga, J.A., Joglar, B., Guerra, M.J., Labandeira-Garcia J.L. The mitochondrial ATP-sensitive potassium channel blocker 5-hydroxydecanoate inhibits toxicity of 6-hydroxydopamine on dopaminergic neurons. Neurotox. Res. 2009, 15, 82–95. [CrossRef]
- Jonsson, G. Chemical neurotoxins as denervation tools in neurobiology. Annu. Rev. Neurosci. 1980, 3, 169–187. [Google Scholar] [CrossRef]
- Tamano, H., Morioka, H., Nishio, R., Takeuchi, A., Takeda. A. Blockade of rapid influx of extracellular Zn2+ into nigral dopaminergic neurons overcomes Paraquat-induced Parkinson’s disease in rats. Mol. Neurobiol. 2019, 56, 4539–4548. [CrossRef]
- Tamano, H., Nishio, R., H. Morioka, Furuhata, R., Komata, Y., Takeda. A. Paraquat as an environmental risk factor in Parkinson’s disease accelerates age-related degeneration via rapid influx of extracellular Zn2+ into nigral dopaminergic neurons. Mol. Neurobiol. 2019, 56, 7789–7799. [CrossRef]
- Fisher, A.B. Redox signaling across cell membranes. Antioxid. Redox Signal. 2009, 11, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Hara, Y., Wakamori, M., Ishii, M., Maeno, E., Nishida, M., Yoshida, T., Yamada, H., Shimizu, S., Mori, E., Kudoh, J., Shimizu, N., Kurose, H., Okada, Y., Imoto, K., Mori. Y. LTRPC2 Ca2+-permeable channel activated by changes in redox status confers susceptibility to cell death. Mol. Cell 2002, 9, 163–173.
- Kubota, M., Kobayashi, N., Sugizaki, T., Shimoda, M., Kawahara M., Tanaka K. Carnosine suppresses neuronal cell death and inflammation induced by 6-hydroxydopamine in an in vitro model of Parkinson’s disease. PLoS One 2020, 15, e0240448.
- Sadakane, Y., Konoha, K., Kawahara, M. Protective activity of mango (Mangifera indica L.) fruit against a zinc-induced neuronal cell death is independent of its antioxidant activity. Trace Nutr. Res. 2005, 22, 73–79.
- Everaert, I., Mooyaart, A., Baguet, A., Zutinic, A., Baelde, H., Achten, E., Taes, Y., De Heer, E., Derav, W. Vegetarianism, female gender and increasing age, but not CNDP1 genotype, are associated with reduced muscle carnosine levels in humans. Amino Acids. 2011, 40, 1221–1229. [CrossRef]
- Derave, W., Everaert, I., Beeckman, S., Baguet, A. Muscle carnosine metabolism and beta-alanine supplementation in relation to exercise and training. Sports. Med. 2010, 40, 247–263. [CrossRef]
- Bakardjiev, A., Bauer, K. Biosynthesis, release, and uptake of carnosine in primary cultures. Biochemistry (Mosc) 2000, 65, 779–782.
- Harris, R.C., Wise, J.A., Price, K.A., Kim, H.J., Kim, C.K., Sale, C. Determinants of muscle carnosine content. Amino Acids 2012, 43, 5–12.
- Gariballa, A.E., Sinclair, A.J. Carnosine: physiological properties and therapeutic potential. Age Ageing 2000, 29, 207–210. [CrossRef]
- A Boldyrev, A., Aldini, G., Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [CrossRef]
- Mori, M., Mizuno, D., Konoha-Mizuno, K., Sadakane, Y., Kawahara M. Carnosine concentration in the muscle of thoroughbred horses and its implications in exercise performance. Trace Nutrients Res. 2015, 32, 49–53.
- Kawai, M., Minami, Y., Sayama, Y., Kuwano, A., Hiraga, A., Miyata H. Muscle Fiber Population and Biochemical Properties of Whole Body Muscles in Thoroughbred Horses. Anat. Rec. (Hoboken) 2009, 292, 1663–1669. [CrossRef] [PubMed]
- Abe, H. Role of histidine-related compounds as intracellular proton buffering constituents in vertebrate muscle. Biochemistry (Mosc) 2000, 65, 757–765. [Google Scholar] [PubMed]
- Sale, C., G Artioli, G., Gualano, B., Saunders., M Hobson, R., C Harris, R.Carnosine: from exercise performance to health. Amino Acids 2013, 44, 1477–1491.
- Quesnele, JJ., Laframboise, MA., Wong, JJ., Kim, P., Wells, GD., The effects of beta-alanine supplementation on performance: a systematic review of the literature. Int. J. Sport. Nutr. Exerc. Metab. 2014, 24, 14–27.
- Dubois, V.D., Bastawrous, A. N-acetylcarnosine (NAC) drops for age-related cataract. Cochrane Database Syst. Rev. 2017, 2, CD009493.
- Matsukura, T., Tanaka, H. Applicability of zinc complex of L-carnosine for medical use. Biochemistry (Mosc) 2000, 65, 817–823.
- Kimura, K., Nakano, Y., Sugizaki, T., Shimoda, M., Kobayashi, N., Kawahara, M., Tanaka, K.I. Protective effect of polaprezinc on cadmium-induced injury of lung epithelium. Metallomics 2019, 11, 1310–1320.
- Baslow, M.H., Suckow, R.F., Berg, M.J., Marks, N., Saito, M., Bhakoo, K.K. Differential expression of carnosine, homocarnosine and N-acetyl-L-histidine hydrolytic activities in cultured rat macroglial cells. J. Mol. Neurosci. 2001, 17, 351–359. [CrossRef]
- Tiedje, KE., Stevens, K., Barnes, S., Weaver, DF. β-Alanine as a small molecule neurotransmitter. Neurochem. Int. 2010, 57, 177–188. [CrossRef]
- Wang, H., Fei, YJ., Ganapathy, V., Leibach FH. Electrophysiological characteristics of the proton-coupled peptide transporter PEPT2 cloned from rat brain. Am. J. Physiol. 1998, 275, C967–C975. [CrossRef] [PubMed]
- Lopachev, AV., Abaimov, DA., Filimonov, IS., Kulichenkova, KN., Fedorova, TN. An assessment of the transport mechanism and intraneuronal stability of L-carnosine. Amino Acids 2022, 54, 1115–1122. [CrossRef]
- Bonfanti, L., Peretto, P., De Marchis, S., Fasolo, A. Carnosine-related dipeptides in the mammalian brain. Prog. Neurobiol. 1999, 59, 333–353. [CrossRef] [PubMed]
- Bakardjiev, A. Carnosine and beta-alanine release is stimulated by glutamatergic receptors in cultured rat oligodendrocytes. Glia 1998, 24, 346–351. [Google Scholar] [CrossRef]
- Mori, M., Mizuno, D., Konoha-Mizuno, K., Sadakane, Y., Kawahara, M. Quantitative analysis of carnosine and anserine in foods by performing high performance liquid chromatography. Biomed. Res. Trace Elem. 2015, 26, 147–152.
- Kawahara, M., Sadakane Y., Mizuno K., Kato-Negishi M., Tanaka K. Carnosine as a possible drug for zinc-induced neurotoxicity and vascular dementia. Int. J. Mol. Sci. 2020, 21, 2570. [CrossRef]
- Biffo, S., Grillo, M., Margolis, F.L. Cellular localization of carnosine-like and anserine-like immunoreactivities in rodent and avian central nervous system. Neuroscience 1990, 35, 637–651. [CrossRef]
- Margolis, F.L., Grillo, M. Axoplasmic transport of carnosine (β-alanyl-L-histidine) in the mouse olfactory pathway. Neurochem. Res. 1977, 2, 507–519. [CrossRef]
- Harding, J., Graziadei, P.P., Monti Graziadei, G.A., Margolis, F.L. Denervation in the primary olfactory pathway of mice. IV. Biochemical and morphological evidence for neuronal replacement following nerve section. Brain Res. 1977, 132, 11–28. [CrossRef]
- Harding, J., Margolis, F.L. Denervation in the primary olfactory pathway of mice. III. Effect on enzymes of carnosine metabolism. Brain Res. 1976, 110, 351–360. [CrossRef]
- Baran, E.J. Metal complexes of carnosine. Biochemistry (Mosc) 2000, 65, 789–797. [Google Scholar] [PubMed]
- Trombley, P.Q., Horning, M.S., Blakemore, L.J. Interactions between carnosine and zinc and copper: implications for neuromodulation and neuroprotection. Biochemistry (Mosc) 2000, 65, 807–816.
- Corona, C., Frazzini, V., Silvestri, E., Lattanzio, R., La Sorda, R., Piantelli, M., Canzoniero, L.M., Ciavardelli, D., Rizzarelli, E., Sensi, S.L. Effects of dietary supplementation of carnosine on mitochondrial dysfunction, amyloid pathology, and cognitive deficits in 3xTg-AD mice. PLoS ONE 2011, 6, e17971.
- Caruso, G., Fresta, C.G., Musso, N., Giambirtone, M., Grasso, M., Spampinato, S.F., Merlo, S., Drago, F., Lazzarino, G., Sortinom, M.A., et al. Carnosine prevents Aβ-induced oxidative stress and inflammation in microglial cells: a key role of TGF-β. Cells 2019, 8, 64. [CrossRef] [PubMed]
- Kawahara, M., Koyama, H., Nagata, T., Sadakane, Y. Zinc, copper, and carnosine attenuate neurotoxicity of prion fragment PrP106-126. Metallomics 2011, 3, 726–734. [CrossRef]
- Ommati, M.M., Heidari, R., Ghanbarinejad, V., Aminian, A., Abdoli, N., Niknahad, H. The neuroprotective properties of carnosine in a mouse model of manganism is mediated via mitochondria regulating and antioxidative mechanisms. Nutr. Neurosci. 2020, 23, 731–743. [CrossRef]
- Berezhnoy, D.S., Stvolinsky, S.L., Lopachev, A.V., Devyatov, A.A., Lopacheva, O.M., Kulikova, O.I., Abaimov, D.A., Fedorova, T.N. Carnosine as an effective neuroprotector in brain pathology and potential neuromodulator in normal conditions. Amino Acids 2019, 51, 139–150. [CrossRef]
- Ferreiro, E., Baldeiras, I., Ferreira, I.L., Costa, R.O., Rego, A.C., Pereira C.F., Oliveira C.R. Mitochondrial- and endoplasmic reticulum-associated oxidative stress in Alzheimer’s disease: from pathogenesis to biomarkers. Int. J. Cell. Biol. 2012, 2012, 735206.
- Moskalev, A.A., Smit-McBride, Z., Shaposhnikov, M.V., Plyusnina, E.N., Zhavoronkov, A., Budovsky, A., Tacutu, R., E Fraifeld, V. Gadd45 proteins: relevance to aging, longevity and age-related pathologies. Ageing Res. Rev. 2012, 11, 51–66.
- Korb, E., Finkbeiner S. Arc in synaptic plasticity: from gene to behavior. Trends Neurosci. 2011, 34, 591–598. [CrossRef]
- Zhang, X., Song, L., Cheng, X., Yang, Y., Luan, B., Jia, L., Xu, F., Zhang, Z. Carnosine pretreatment protects against hypoxia-ischemia brain damage in the neonatal rat model. Eur. J. Pharmacol. 2011, 667, 202–207. [CrossRef] [PubMed]
- Park, H.S., Han, K.H., Shin, J.A., Park, J.H., Song, K.Y., Kim, D.H. The neuroprotective effects of carnosine in early stage of focal ischemia rodent model. J. Korean Neurosurg. Soc. 2014, 55, 125–130. [CrossRef] [PubMed]
- Noguchi, K., Ali, T.F.S., Miyoshi, J., Orito, K., Negoto, T., Biswas, T., Taira, N., Koga, R., Okamoto, Y., Fujita, M., et al. Neuroprotective effects of a novel carnosine-hydrazide derivative on hippocampal CA1 damage after transient cerebral ischemia. Eur. J. Med. Chem. 2019, 163, 207–214. [CrossRef] [PubMed]
- Davis, C.K., Laud, P.J., Bahor, Z., Rajanikant, G.K., Majid, A. Systematic review and stratified meta-analysis of the efficacy of carnosine in animal models of ischemic stroke. J. Cereb. Blood Flow Metab. 2016, 36, 1686–1694. [CrossRef]
- Tomonaga, S., Hayakawa, T., Yamane, H., Maemura, H., Sato, M., Takahata, Y., Morimatsu, F., Furuse, M. Oral administration of chicken breast extract increases brain carnosine and anserine concentrations in rats. Nutr. Neurosci. 2007, 10, 181–186. [CrossRef]
- Hoffman, J.R., Rathmacher, J.A., Robinson, J., Gepner, Y., Cohen, H. Effect of β-alanine supplementation on carnosine and histidine content in the hippocampus of 14-month-old rats. Appl. Physiol. Nutr. Metab. 2019, 44, 1112–1115. [CrossRef] [PubMed]
- Araminia, B., Shalbafan, M., Mortezaei, A., Shirazi, E., Ghaffari, S., Sahebolzamani, E., Mortazavi, S.H., Shariati, B., Ardebili, M.E., Aqamolaei, A., et al. L-Carnosine combination therapy for major depressive disorder: a randomized, double-blind, placebo-controlled trial. J. Affect. Disord. 2020, 267, 131–136.
- Hisatsune, T., Kaneko, J., Kurashige, H., Cao, Y., Satsu, H., Totsuka, M., Katakura, Y., Imabayashi, E., Matsuda, H. Effect of anserine/carnosine supplementation on verbal episodic memory in elderly people. J. Alzheimers Dis. 2016, 50, 149–159. [CrossRef]
- Masuoka, N., Yoshimine, C., Hori, M., Tanaka, M., Asada, T., Abe, K., Hisatsune, T. Effects of anserine/carnosine supplementation on mild cognitive impairment with APOE4. Nutrients 2019, 11, 1626. [CrossRef] [PubMed]
- Oppermann, H., Heinrich, M., Birkemeyer, C., Meixensberger, J., Gaunitz, F. The proton-coupled oligopeptide transporters PEPT2, PHT1 and PHT2 mediate the uptake of carnosine in glioblastoma cells. Amino Acids 2019, 51, 999–1008. [CrossRef]
- Hata, J., Ohara, T., Katakura, Y., Shimizu, K., Yamashita, S., Yoshida, D., Honda, T., Hirakawa, Y., Shibata, M., Sakata, S., et al. Association between serum β-alanine and risk of dementia. Am. J. Epidemiol. 2019, 188, 1637–1645.
- Stuerenburg, H.J. The roles of carnosine in aging of skeletal muscle and in neuromuscular diseases. Biochemistry 2000, 65, 862–865. [Google Scholar] [PubMed]
- J Anderson, E., Vistoli, G., A Katunga, L., Funai, K., Regazzoni, L., Monroe, T.B., Gilardoni, E., Cannizzaro, L., Colzani, M., De Maddis, D., Rossoni, G., Canevotti, R., Gagliardi, S., Carini, M., Aldini G. A carnosine analog mitigates metabolic disorders of obesity by reducing carbonyl stress. J. Clin. Invest. 2018, 128, 5280–5293.
- de Jager, S., Vermeulen, A., De Baere, S., Van der Stede, T., Lievens, E., Croubels, S., Jäger, R., Purpura, M., Bourgois, JG., Derave, W. Acute balenine supplementation in humans as a natural carnosinase-resistant alternative to carnosine. Sci Rep. 2023, 13, 6484. [CrossRef]
- Kawahara, M., Konoha, K. Drugs for prevention or treatment of vascular dementia [Translated from Japanese]. JP5382633, 11 October 2013. 11 October.
- Kawahara, M., Konoha, K. Drugs for prevention or treatment of vascular dementia [Translated from Japanese]. JP5294194, 21 June 2013.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



