
Submitted:
05 July 2023
Posted:
07 July 2023
You are already at the latest version
Abstract
Embryo and fetus grow in a hypoxic environment. Intrauterine oxygen levels fluctuate throughout the pregnancy allowing the oxygen to modulate apparently contradictory functions, such as the expansion of stemness but also differentiation. We have recently demonstrated that in the last weeks of pregnancy oxygenation progressively increases, but the trend of oxygen levels during the previous weeks remains to be clarified. In the present study, umbilical venous and arterial oxygen levels, fetal oxygen extraction, oxygen content, CO2, and lactate were evaluated in a cohort of healthy newborns with gestational age < 37 weeks. A progressive decrease in pO2 levels associated with a concomitant increase in pCO2 and reduction of pH has been observed starting from the 23rd week until approximately the 33-34th week of gestation. Over this period, despite the increased hypoxemia, oxygen content remains stable thanks to increasing hemoglobin concentration, which allows the fetus to becoming more hypoxemic, but not more hypoxic. Starting from the 33-34th week, fetal oxygenation increases and ideally continues following the trend recently described in term fetuses. The present study confirms that oxygenation during intrauterine life continues to vary even after placenta development, showing a clear biphasic trend. Fetuses, in fact, from mid-gestation to near-term become progressively more hypoxemic. However, starting from the 33-34th week, oxygenation progressively increases until birth. In this regard, our data suggest that the placenta is the hub that ensures this variable oxygen availability to the fetus, and we speculate that this biphasic trend is functional to the promotion, in specific tissues and at specific timing, of stemness and intrauterine differentiation.
Keywords:
newborn
; intrauterine hypoxia
; fetal hypoxia
; differentiation
1. Introduction
The embryo and the fetus live in a physiologically hypoxic environment (the so-called Everest in utero) [1], although the intrauterine environment is characterized by a variable level of hypoxia throughout pregnancy.
During the first stages of pregnancy, the concentrations of oxygen are very low, similar to those measurable within the non-pregnant uterus [2,3]. Over the first weeks of pregnancy, the placental development favors the increase of oxygen availability to the fetoplacental unit and, at the beginning of the second trimester of gestation, placenta oxygenation triples the oxygen availability reaching a maximum around the 16th week of gestation [3]. From this week onwards, a slow gradual reduction in placental oxygen levels is observed, and this trend appears to be consistent with a progressive reduction in fetal oxygenation status [3]. In fact, a series of studies carried out by analyzing human umbilical-cord venous and arterial blood samples obtained via cordocentesis, demonstrates decreasing fetal partial pressure of oxygen (pO2) and saturation of oxygen (SaO2), which starts from the 16-18th week of gestation and progresses with advancing gestation [4,5,6,7].
However, some considerations led us to imagine a reversion of the trend of oxygenation during the final stages of pregnancy. This hypothesis got inspiration from the observation that in many animals, vasculature is still immature at birth and that its maturation occurs after the birth. This is particularly evident in the brain of rodents, whose appropriate glial-vascular interaction is instituted after birth [8,9] and in the retina of mice, which is avascular at birth but begins to vascularize during the first week after birth [10] or even later [11], demonstrating, at least chronologically, a strict relationship between vessel maturation and increase in oxygen exposure. If vascular maturity requires an increase in oxygen levels, it was legitimate to imagine that in humans, whose vascularization is completed during the last weeks of intrauterine life [12], there was a progressive increase in oxygen tension in the more advanced stages of pregnancy.
To verify if oxygenation increased in the last weeks of pregnancy, we have recently performed an observational study that assessed the umbilical gas analysis collected from a cohort of healthy newborns with gestational age ≥ 37 weeks. Results demonstrated a progressive increase in fetal oxygenation from the 37th to the 41st weeks of gestation (approximately an increase of about 1 mmHg per week) [13]. These findings are in accordance with a previous study that reported a slight increase in cord venous pO2 between near-term and term newborns [14]. The pooled analysis of studies evaluating fetal oxygen levels suggests that oxygenation during intrauterine life continues to vary also after placenta development, taking on a biphasic trend. Fetuses, in fact, from mid-gestation to near-term grow in an environment progressively more hypoxic, while, approaching the term of pregnancy, oxygenation reverse its trend. The knowledge of this dynamic assumes a particular importance because it helps us to understand the physiological mechanisms that guarantee the maturation of the vascularization, whether it takes place after birth, as in rodents, or in utero, as in humans.
The main objective of this study is to reconstruct, with the same methodology used to analyze the blood gas variations in term infants, the fetal oxygenation status from the 23rd to the 36th weeks of gestation. Our objectives are to verify that healthy fetuses become more hypoxemic over the weeks, to confirm that this trend at a certain point is reverted, and to identify the time when this reversal occurs.
2. Results
Out of 12544 newborns born between 2016 and 2022, 11375 newborns with GA ≥37 weeks were excluded from the analysis. 1169 preterm newborns were eligible for this study. Umbilical-cord gas analyses of 560 newborns were not performed, incomplete, or unreliable. Out of 609 samples, 5 with acidosis at birth were excluded (Figure 1).
Of 604 newborns enrolled, males were prevalent (310/604, 51.2%). Neonates born by vaginal delivery were a minority (148/604, 24.5%). Table 1 shows the cumulative demographic and gas analytical parameters of all newborns.
According to what was recently observed in term newborns [13], also in preterm neonates the modality of delivery influences the blood gas analysis. Neonates born by vaginal delivery showed higher values of pO2 and lactate both in venous and arterial cord samples. Lower levels of pH, partial pressure of carbon dioxide (pCO2), and bicarbonate were evident only in venous cord samples. Neonates born by cesarean section displayed an oxygen extraction significantly higher than neonates born by spontaneous delivery, in line with what was demonstrated in term neonates [13]. To verify whether the incidence of vaginal or caesarean delivery substantially changed as gestation progressed, data of blood gas analysis were arbitrarily grouped in six newborn groups: a group of preterm newborns between 22 to 26+6 weeks of gestation, 27-28+6, 29-30+6, 31-33+6, 34-35+6, and 36-36+6 (Figure 1). The percentage of vaginal or cesarean delivery was not statistically different among the groups, allowing us to evaluate the blood samples as a whole, regardless of the type of delivery, without further stratification.
To evaluate whether oxygen tension varied during the pregnancy, data of venous and arterial pO2, as well as oxygen extraction, were assessed in function to the progression of pregnancy (Figure 2). Values from umbilical venous samples revealed a progressive decrease in pO2 levels until approximately 230-240 days of gestation (approximately the 33rd-34th week of gestation) when pO2 appears to reach its lower point; then, starting from the 34th week of pregnancy, pO2 seems to increase (Figure 2A), to then continue, ideally, with the positive progression observed from the 37th to the 41st weeks of gestation [13]. This tendency is better highlighted in Supplementary Figure 1, where data were stratified according to groups of weeks of pregnancy. The arterial pO2 level also appears reduced with the progression of gestation (Figure 2B), with a minimal increase in the weeks near term (Supplementary Figure 1). Overall, oxygen extraction didn’t show significant change throughout the period considered (Figure 2C), even though a modest increase was evident comparing the group of 27-28+6 and 29-30+6 towards the more advanced stages of pregnancy (Supplementary Figure 1).
The levels of venous and arterial hemoglobin (Hb) progressively increased from the 23rd weeks onwards, reaching a plateau approximately at 230-240 days of gestation (around week 33-34 of gestation) (Figure 3A,B). The Hb rise appears to follow a trend opposite to that followed by pO2, with an increase of around 1 g/dL per groups of gestational age, as shown in Supplementary Figure 2. This increase explains why, despite the reduced intake of oxygen from the placenta to the fetus, the venous content of oxygen remained stable throughout the period examined (Figure 3C), excluding a minimal increase during the weeks near term (Supplementary Figure 2).
Simultaneously with the decrease in pO2, a linear reduction in venous and arterial pH was observed as pregnancy proceeded (Figure 4A,B), coupled with a mild reduction in venous base excess (Figure 4C), but not in arterial base excess (Figure 4D), and a mild increase in arterial bicarbonate (Figure 4F), but not in venous (Figure 4E). This tendence is emphasized in Supplementary Figure 3, where data were stratified according to groups of weeks of pregnancy.
In analogy to what observed in term fetuses, also in preterm fetuses, blood gas analysis displayed for the umbilical-cord content of CO2 an opposing tendency if compared with the trend of oxygen, with a progressive surge both in venous (Figure 5A) and arterial samples (Figure 5B). This increase appears to be progressive at least until the group with gestational age 31-33+6 weeks (Supplementary Figure 4). Starting from the 34th week of pregnancy, pCO2 seems to decrease and then to continue, ideally, with the negative progression recently described in term fetuses [13]. Overall, CO2 production didn’t seem to change significantly (Figure 5C), even though, during the more advanced weeks of gestation, we observe an apparent increase of CO2 production (Supplementary Figure 4) that then continues, ideally, with the progression recently described from the 37th to the 41st week of gestation [13].
Between the 23rd to the 36th weeks of gestation, no significant change in lactate levels were observed either in venous samples (Figure 6A) or in arterial samples (Figure 6B). A mild increase in fetal lactate production seemed evident (Figure 6C) as a consequence to a mild increase detected in newborns belonging to more advanced gestational age (Supplementary Figure 5).
3. Discussion
Conception, embryogenesis, and fetus growth occur in the female reproductive tract where the oxygen concentration is very low. A physiological hypoxia has been indicated as the key regulator of the harmonious processes of placental and embryonal development [15,16]. The importance of this peculiar environmental oxygenation is witnessed by the improved live-birth rate of preimplantation embryos cultured under hypoxic conditions compared with normoxic culture [17]. However, the embryo and the following fetoplacental unit grow and differentiate in a milieu where the levels of hypoxia undergo evident and recurring variations throughout the pregnancy.
Placental and embryonal cells react to a hypoxic environment with a series of adaptive adjustments of genic expression. The upregulation of Hypoxia Inducible Factors (HIFs) represents the hub through whom hypoxia promotes placental development [18]. Under hypoxia, HIF-1α translocates into the nucleus and dimerizes with HIF-1β leading to the transcription of several hundred specific target genes [19], favoring cell survival in a hypoxic environment (induction of specific enzymes involved in energy metabolism, erythropoiesis, and angiogenesis). [20]. Therefore, HIF modulation guarantees at the same time proliferation and metabolic adaptation of embryonal cells, placental development as well as trophoblast differentiation [21,22].
HIF regulation during pregnancy is complex. Although HIF is regulated also through oxygen-independent mechanisms such as hormones (Angiotensin II), cytokines (interleukin-1β, tumour necrosis factor α, and NF-kβ) or growth factors (transforming growth factor- β1 and insulin-like growth factor), all significantly upregulated during pregnancy [23], however oxygen levels (and its fluctuations) represent the main regulator of placental HIF levels, placenta development, as well as normal mammalian embryo morphogenesis [24]. The pivotal role of HIF for embryo/fetus wellness is indirectly confirmed by the high embryonic lethality, dysmorphogenesis or severe placental defects observed in HIF-1α-/- mice embryos [18].
Although hypoxia and HIF are indispensable for the harmonious development of the fetus-placental unit, from the first moments of pregnancy it is evident that the levels of oxygen and HIF change following a precise dynamic.
Conception occurs when a sperm cell successfully fertilizes an egg cell in the fallopian tube, at an oxygen concentration of approximately 5-7% [25]. By this time, the morula reaches the uterine cavity, where the oxygen concentration is significantly reduced to a level of 2% [26,27]. The transition to this more hypoxic environment is essential in the first 2-3 weeks of life to allow the proliferation, implantation, anchoring and invasion of the blastocyst into the maternal uterus [28]. In fact, on one side, the hypoxia (and the up-regulation of HIFs) is crucial to maintain the pluripotent attitude of the embryonic stem cells [29], as well as of cytotrophoblast cells, which are usually considered to be trophoblast stem cells [3], and to promote their proliferation [30]. On the other side, the exposure to a decreasing oxygen gradient during the travel from the oviduct to the uterus promotes a metabolic shift of cytotrophoblast cells from oxidative phosphorylation towards a glycolytic metabolism (Warburg effect) [31,32,33]. This metabolic adaptation confers a proliferative advantage, since it promotes the production of many intermediates of the pentose phosphate pathway such as ribose sugars necessary for nucleic acid synthesis and induces the production of elevated lactic acid levels that are useful to promote nesting in surrounding tissues [34]. In fact, the increased production and extrusion of lactate facilitates disaggregation of uterine tissues and promotes trophoblast invasion [35].
Increasing hypoxia and high levels of HIF during the first weeks of life are necessary also to assure cytotrophoblasts transmigration across the uterine epithelium and their differentiation into extravillous trophoblasts [18,36], which in turn migrate along the lumen of the spiral arterioles. These cells progressively remodel the spiral arteries replacing the smooth muscle and the elastic lamina of the vessel. In this way, the low-flow, high-resistance original spiral arteries become high-flow, low-resistance vessels and assure in the following weeks of pregnancy a gradual increase of oxygen delivery [28].
In summary, the intrauterine low-oxygen tension promotes embryonic cytotrophoblast cells proliferation, their intrauterine invasion, and interconnections with the uterine spiral arterioles [37], thus laying the foundations for a subsequent increase in oxygen levels. In the meantime, HIF upregulation promotes the concomitant activation of a series of angiogenic factors including vascular endothelial growth factor (VEGF), basic fibroblast growth factor, platelet-derived growth factor, angiopoietin-1, angiopoietin-2, Tie2, and monocyte chemoattractant protein-1 [38]. These proangiogenic factors in turn raise the foundation for embryonal vascular and placenta development and a further increase in oxygen availability [3]. In turn, such an increase in microenvironmental oxygen promotes the differentiation of cytotrophoblasts into multinucleated syncytiotrophoblasts, necessary to secrete several hormones for pregnancy maintenance [39].
Essentially, what happens to the embryo in the first weeks of life represents a plastic demonstration that hypoxia is indispensable; however, within this hypoxic environment, we observe a fine regulation of oxygen levels, of HIF, and consequently of the expression of its target genes aimed at modulating apparently contradictory functions, such as the expansion of stemness and differentiation.
Placentation explains why placental oxygen tension, which is less than 20 mmHg (more or less 2% O2) during the first weeks of human gestation (approximately until the 10th week) [2,3], increases to around 60 mmHg (roughly 8% O2) at the beginning of the second trimester of gestation [2]. The increased oxygen availability from the first weeks to the early weeks of the second trimester justifies why the fetal habitat can be considered a dynamic environment where hypoxia initially attenuates.
While recent studies performed on healthy fetal animals reported no significant variations in blood gas and acid-base status during gestation [40,41,42,43,44,45,46,47], the available data for human fetuses obtained via cordocentesis, although limited, indicate a progressive reduction in fetal oxygen levels starting from the 16-18th weeks of gestation toward its end [4,5,6,7]. The reduced oxygen supply to the fetuses is explained by the increased placental oxygen extraction with advancing gestation, as demonstrated by samples of maternal blood drawn from the subchorial lake under the chorionic plate [4]. However, these studies were performed on pregnancies complicated by fetal anomalies, a variety of prenatal pathologies, or maternal infections [4,5,6,7]; thus, such results would need to be confirmed in a healthy population. The fluctuations of oxygen levels are further confirmed by the analysis of a large amount of umbilical cord blood gas analysis in which a clear trend reversal of oxygenation in the last weeks of gestation was demonstrated [13,14]. Although the investigation of the mechanisms underlying the increased oxygenation was beyond the scope of these studies, this phenomenon has been attributed to an increased oxygen diffusion from the placenta to the fetus, probably correlated with the physiological aging of the placenta. In fact, the thinning of the placental barrier surface area due to a reduction of the distance between fetal vessels and the trophoblastic membrane over time favors an increase in placental oxygen transport [48]. In conclusion, the literature shows data apparently contradictory; while, from mid-gestation to near term, several historical studies report a progressive reduction of fetal oxygenation [4,5,6,7], other more recent studies describe increased oxygenation during the last weeks of gestation [13,14]. This contradiction can only be explained by imagining a biphasic trend of fetal oxygenation, with a first phase characterized by a progressive reduction in fetal oxygenation, followed by a phase of increased oxygenation.
The results of the present study confirm that fetuses grow in a dynamic low-oxygen environment and demonstrate for the first time that the concentration of oxygen follows a biphasic trend. In fact, at least from the 22nd week onward, we observed a progressive and linear reduction of venous pO2, that slows and then reverses at around 33-34 weeks of gestation. A similar trend is observed also regarding the arterial samples, and therefore the oxygen extraction values do not change substantially during this time. These data suggest that the fetus is subject to a slow and progressive reduction of oxygen supply from the placenta, at least from the 22nd until the 33-34th week which seems to represent the inflection point between the two phases. This observation is in line with what has been observed from the second to the third trimester of pregnancy in the intervillous space of the human placentas, where mean oxygen tension measured gradually declines [3,4,49].
If we consider that the pO2 level in mothers remains stable throughout pregnancy [50] in concomitance with the uterine blood flow [51], the gradual reduction of venous umbilical pO2 suggests that placental growth and its increasing metabolic activity progressively makes less oxygen available to the fetus. It is well known that placental oxygen consumption accounts for a large percentage of the collective fetal and placental oxygen consumption, and that this percentage increases over the weeks [15,51]. Therefore, our data suggest that the increasing consumption of oxygen by the placenta causes a progressive accentuation of fetal hypoxemia until the 33-34th week of pregnancy. In the face of a continuous reduction in the placental oxygen supply, the venous pCO2 progressively increases until the 33-34th week of gestation, confirming the hypothesis that these changes in the supplies to the fetus are consequent to an increased metabolic activity of the placenta. The combination of the reduction in oxygenation with the increase in pCO2 explains why the venous pH gradually decreases, at least up to the 33-34th week of pregnancy.
Increasing fetal hypoxemia is confirmed by the progressive increase in Hb concentration, the synthesis of which is known to be induced by low levels of oxygen, through the activation of the HIF/erythropoietin axis [52]. The present study confirms that the Hb level gradually and progressively increases as pregnancy progresses, starting from the 22nd week of gestation, showing a trend opposite to that of pO2, as already reported by several authors [53]. Despite the progressive decrease in pO2, the increased Hb explains why the umbilical venous blood oxygen content remains unchanged. This strategy allows the fetus, at least from the 22nd to the 33-34th weeks of pregnancy, to obtain simultaneously two results: on the one hand it guarantees a constant oxygen content and therefore does not deprive the peripheral tissues of the necessary oxygen, but at the same time the concentration of oxygen dissolved in the blood is reduced. This explains why the fetus becomes progressively more hypoxemic despite the constant oxygen content maintaining the fetus in a status of stable hypoxia. The maintenance, from the 22nd to the 33-34th week of gestation, of this efficient balance is evidenced by the absolute stability of lactate values.
However, from the 33-34th week forward, this balance gets broken and oxygenation starts to increase as recently described in full-term newborns [13]: this gestational age represents the watershed beyond which umbilical venous oxygenation increases, probably due to a physiologic aging of the placenta that favors an increased diffusion of oxygen [48], and consequently, the increase in fetal Hb levels stops. The oxygen availability to fetuses during the last weeks of gestation is probably even more significant because in late gestation the percentage of fetal Hb decreases as adult Hb (which has a lower affinity for oxygen) begins to increase [54]. A comparable reversal is observed also for venous pCO2, which increases until the 34th week, after that pCO2 stops increasing, ideally continuing to decrease until the end of pregnancy [13]. The arterial pCO2 instead seems to continue to increase also after the 34th week, suggesting a possible increase in fetal pCO2 production and therefore a possible metabolic shift toward a mitochondrial activation, which will be more evident after 37 weeks [13].
Therefore, the biological role of oxygen fluctuations observed in the more advanced stages of pregnancy, also after placenta development, although difficult to interpret, may serve for tissue-specific stem cell recruitment [55] and cell differentiation [56]. In some anatomic districts, for instance, hypoxemia is essential in preserving naïve stemness potential [57,58], or modulating cell differentiation [56]; this is particularly evident in the nervous central system, where hypoxia stimulates several processes including cell survival, proliferation, catecholaminergic differentiation of isolated neural crest stem cells [59] or mesencephalic precursor cells into neurons [60], or undifferentiated astrocytes to differentiate [61,62]. In other regions, instead, it is the increased oxygenation that drives cell differentiation as in neural retinal tissue [63], pancreatic cells [64], keratinocytes [65], hepatocytes cell lines [66], or endothelial cells [67]. Therefore, it is realistic to speculate that the intrauterine environment, which physiologically becomes more hypoxic from mid-gestation to near-term and then less hypoxic until term, guarantees a tissue-specific cellular differentiation depending on oxygen tension.
As shown here, the 34th week of gestation would represent the watershed between a hypoxic first phase, characterized by an increased stem cells proliferation and by a differentiation of specific cell populations, to a subsequent phase in which increasing oxygenation promotes the reduction of stem cell pool and cell differentiation. This relationship between stemness and gestational age is well evident for endothelial precursor cells (EPCs), which are significantly more expressed in the umbilical cord blood of preterm newborns if compared with term neonates [68,69]. Interestingly, cord blood of preterm infants contains a number of EPCs that grows as gestational age progresses: very low at 24-28 weeks, and then much more expressed at 28-35 weeks [70] or until 34 weeks [71]. Considering that the differentiation of endothelial cells is favored by the increase in oxygen levels [67], we hypothesize that the expansion of EPCs is synchronous with increasing hypoxemia and stops around 33-34th when oxygen increases promoting the completion of the vasculature.
The topic of vasculogenesis is particularly intriguing, considering that in rodents, at variance with humans, the vascularization of many organs is immature at birth. This is particularly evident in the brain, where vascularization completes after birth [8,9], and even more in the retina, which is avascular during intrauterine life, but vascularizes after birth [10,11].
Although vasculature development depends on hypoxia-drive HIF and VEGF increase [38], however, in the retina vessel appearance becomes evident a few days after birth, when oxygenation increases in concomitance with lowered levels of HIF and VEGF [72]. This observation suggests that vascularization occurs in two phases. In the first intrauterine phase, hypoxia induces astrocyte maturation and the production of VEGF [73] favoring, in turn, the recruitment of EPCs: then, over the second phase characterized by increased oxygenation and reduction of HIF and VEGF, EPCs can differentiate in endothelial cells. A similar immaturity is present in very preterm neonates, whose retina at birth is only partially vascularized, and whose brain is incompletely vascularized, especially in the germinal matrix [74]. This immaturity is believed to predispose preterm infants to an elevated risk to develop intraventricular hemorrhage. However, this risk decreases for neonates born after 33-34 weeks of pregnancy or, in highly preterm newborns, after the first days of life, suggesting that exposure environment more oxygenated induces endothelial cell maturation [74].
Considering that oxygen plays an important regulation in the modulation of the stem population and the processes of differentiation, the knowledge of its fluctuations and the consequent biological effects opens new perspectives for the studies that in the future will try to artificially reproduce the benefits of intrauterine life. In this regard, studies aimed at the development of artificial placenta technology [75] cannot ignore the physiological levels of oxygenation which must be respected. But even the attempts to mimic the biological effects of intrauterine hypoxemia through the stimulation of β3-adrenoceptor agonists [76] will have to take into consideration that fetal hypoxemia increases only up to the 33-34th week.
Limitations
1. Umbilical-cord samples represent a safe, non-invasive method to obtain valuable information regarding intrauterine conditions [77]. However, the type of delivery may affect the gas analytic values: fetal oxygenation of neonates born by vaginal delivery in fact may be altered by the engagement of the fetus through the birth canal or by the reduction of blood flow during uterine contractions [78], as well as cesarean section can affect fetal well-being due to the cardiovascular effects of anesthesia, maternal position, and maternal ventilation [79,80]. Although the impact of delivery type on the gas test result is well known [13], in this study we didn’t find a different incidence of vaginal or cesarean delivery at different gestational ages, and therefore the blood samples were analyzed as a whole, without further stratification.
2. In this study, the umbilical-cord blood samples have been immediately collected at birth. The merging of data obtained from preterm and term newborns would have provided a better interpretation of the fluctuations of fetal oxygenation. However, the pooling of such data would not have been methodologically suitable, as umbilical-cord blood samples in neonates born at term are usually performed by blood collected approximately 60 sec after birth [13].
Conclusions
The present study confirms that oxygenation during fetal life undergoes important fluctuations and integrates the recently published study on oxygenation status during the last weeks of pregnancy [13]. The combination of these studies suggests that from the 22nd week onwards, the fetus becomes progressively more hypoxemic, but this condition reverses around the 33-34th week when oxygen levels progressively increase until the end of the pregnancy.
4. Materials and Methods
The study was performed in the University Hospital of Pisa, Italy, with around 1700 births per year. All the newborns preterm born between January 1st, 2016, and December 31st, 2022 were eligible. Their umbilical-cord blood samples were collected immediately after birth (early cord clamping). Samples collected from term newborns (≥ 37 weeks) or gestations with fetal or maternal intrapartum complications (i.e., fetuses with an operative vaginal delivery involving application of forceps or a vacuum extractor, abnormal intrapartum cardiotocography requiring emergency cesarean section, meconium-stained amniotic fluid, placental abruption, cord prolapse, chorioamnionitis, maternal sepsis, convulsions, hemorrhages, uterus rupture, snapped cord) were excluded from the study. Umbilical-cord blood samples for which pH, base excess (BE), or both were not available and with acidosis at birth (pH ≤ 7.00 and/or BE ≤ -12 mmol/L) [81,82] were excluded from the analysis.
Values of umbilical (venous and arterial) parameters < or > 3 SD from their respective means were individually evaluated and i) rectified if probably mids-entered, ii) maintained unchanged if considered plausible, and iii) excluded if considered not plausible [14]. Results of cord blood gas that did not fulfill the following criteria: i. arterial pH < the venous pH (by at least a difference of 0.022) and ii. arterial pCO2 > the venous pCO2 (by at least a difference of 5.3 mmHg) were considered unreliable and excluded from the analysis [83].
Cord blood was collected as recently published [13]. The samples after their labeling and identification as venous or arterial, were analyzed as soon as possible, using an automatic blood gas analyzer (GEM® Premier 4000, Instrumentation Laboratory, Lexington, MA, USA). The pH, pCO2, pO2, SaO2, lactate, and Hb were measured, whereas bicarbonate and base excess were calculated, respectively, from measured pH and pCO2 using the Henderson–Hasselbalch equation pH=6.1+log([HCO3−]/pCO2×0.03) [84] and the formula described by Siggaard- Anderson: (1 - 0.014 x Hb) x [HCO3- - 24.8 + (1.43 x Hb + 7.7) x (pH - 7.4)] [85]. Oxygen content (ml/dl) was calculated using the formula [(1.36 x Hb (g/dl) x SaO2 (%)/100) + (0.0031 x pO2 (mmHg)], where 1.36 is O2 (ml) bound by a gram of Hb, 0.0031 the solubility coefficient of O2 in blood that represents the volume of O2 (ml) dissolved in 100 ml plasma for each mmHg of O2 partial pressure. Fetal oxygen extraction was determined as the difference between umbilical venous and arterial blood oxygen contents divided by umbilical venous oxygen content. Fetal CO2 and lactate productions were calculated, respectively, as the difference between arterial blood CO2 or lactate and venous contents, divided by the respective venous content.
Statistical Analysis
Categorical data were described with the absolute and relative (%) frequency, and continuous data were summarized with the mean and standard deviation. Gestational age was compared with continuous clinical factors using regression models to identify the best model between the linear and the quadratic one. Furthermore, the curves and the equations of the best models were also calculated. Significance was set at 0.05 and all analyzes were performed by SPSS v.28 technology.
Ethical Approval
The present study was conducted in accordance with the ethical principles for medical research involving human subjects adopted by the Word Medical Association General Assembly (Declaration of Helsinki) and its later amendments. The present study was approved by the Pediatric Ethical Committee for Clinical Research of Tuscany region (number 291/2022).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplementary Figure 1. Umbilical-cord oxygenation status (venous pO2, panel A; arterial pO2, panel B; fetal oxygen extraction, panel C) stratified by groups of gestational age. * = p <0.05; ** = p < 0.01; *** = p < 0.001. Data are expressed as median and IQR. Supplementary Figure 2. Umbilical-cord hemoglobin levels (venous hemoglobin, panel A; arterial hemoglobin, panel B) and fetal venous oxygen content (panel C) stratified by groups of gestational age. * = p <0.05; ** = p < 0.01; *** = p < 0.001. Data are expressed as median and IQR. Supplementary Figure 3. Umbilical-cord pH (venous pH, panel A; arterial pH, panel B), BE levels (venous BE, panel C; arterial BE, panel D), and bicarbonate (venous bicarbonate, panel E; arterial bicarbonate, panel F) stratified by groups of gestational age. * = p <0.05; ** = p < 0.01; *** = p < 0.001. Data are expressed as median and IQR. Supplementary Figure 4. Umbilical-cord carbon dioxide levels (venous pCO2, panel A; arterial pCO2, panel B; fetal CO2 production, panel C) stratified by groups of gestational age. * = p <0.05; ** = p < 0.01; *** = p < 0.001. Data are expressed as median and IQR. Supplementary Figure 5. Umbilical-cord lactate levels (venous lactate, panel A; arterial lactate, panel B; fetal lactate production, panel C) stratified by groups of gestational age. * = p <0.05; ** = p < 0.01; *** = p < 0.001. Data are expressed as median and IQR.
Author Contributions
Conceptualization, L.F.; methodology, L.F..; formal analysis, L.F., F.P., F.I., N.C., A.M., and R.M.; investigation, R.T.S.; data curation, L.F., F.P., F.I., N.C., A.M.; writing-original draft preparation, L.F.; writing-review and editing, A.P., M.C., and P.B.; supervision, L.F., A.P., M.C., and P.B. All authors have read and agreed to the published version of the manuscript.”
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board (or Ethics Committee) of the Pediatric Ethical Committee for Clinical Research of Tuscany region (protocol code 291/2022, date of approval 21 december 2021).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
We are most grateful to Jean-Luc Baroni and Cristina Ranzato for their support in conducting this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Eastman, N.J. Mount Everest in utero. Am. J. Obstet. Gynecol. 1954, 67, 701–11. [Google Scholar] [CrossRef] [PubMed]
- Jauniaux, E.; Watson, A.; Burton, G. Evaluation of respiratory gases and acid-base gradients in human fetal fluids and uteroplacental tissue between 7 and 16 weeks’ gestation. Am. J. Obstet. Gynecol. 2001, 184, 998–1003. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Cindrova-Davies, T.; Yung, H.W.; Jauniaux, E. Hypoxia and reproductive health: Oxygen and development of the human placenta. Reproduction 2021, 161, F53–F65. [Google Scholar] [CrossRef]
- Soothill, P.W.; Nicolaides, K.H.; Rodeck, C.H.; Campbell, S. Effect of gestational age on fetal and intervillous blood gas and acid-base values in human pregnancy. Fetal Ther. 1986, 1, 168–75. [Google Scholar] [CrossRef]
- Weiner, C.P.; Sipes, S.L.; Wenstrom, K. The effect of fetal age upon normal fetal laboratory values and venous pressure. Obstet. Gynecol. 1992, 79, 713–8. [Google Scholar]
- Pardi, G.; Cetin, I.; Marconi, A.M.; Lanfranchi, A.; Bozzetti, P.; Ferrazzi, E.; Buscaglia, M.; Battaglia, F.C. Diagnostic value of blood sampling in fetuses with growth retardation. N. Engl. J. Med. 1993, 328, 692–696. [Google Scholar] [CrossRef] [PubMed]
- Nava, S.; Bocconi, L.; Zuliani, G.; Kustermann, A.; Nicolini, U. Aspects of fetal physiology from 18 to 37 weeks’ gestation as assessed by blood sampling. Obstet. Gynecol. 1996, 87, 975–980. [Google Scholar] [CrossRef]
- Semple, B.D.; Blomgren, K.; Gimlin, K.; Ferriero, D.M.; Noble-Haeusslein, L.J. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol 2013, 106-107, 1–16. [Google Scholar] [CrossRef]
- Coelho-Santos, V.; Shih, A.Y. Postnatal development of cerebrovascular structure and the neurogliovascular unit. Wiley Interdiscip. Rev. Dev. Biol. 2020, 9, e363. [Google Scholar] [CrossRef]
- Stahl, A.; Connor, K.M.; Sapieha, P.; Chen, J.; Dennison, R.J.; Krah, N.M.; Seaward, M.R.; Willett, K.L.; Aderman, C.M.; Guerin, K.I.; Hua, J.; Löfqvist, C.; Hellström, A.; Smith, L.E. The mouse retina as an angiogenesis model. Invest. Ophthalmol. Vis. Sci. 2010, 51, 2813–2826. [Google Scholar] [CrossRef]
- Aguilar, E.; Dorrell, M.I.; Friedlander, D.; Jacobson, R.A.; Johnson, A.; Marchetti, V.; Moreno, S.K.; Ritter, M.R.; Friedlander, M. Chapter 6. Ocular models of angiogenesis. Methods Enzymol. 2008, 444, 115–158. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.W.; Strube, Y.N. Retinopathy of Prematurity. In Pediatric Ophthalmology and Strabismus; Wright, K.W., Strube, Y.N., Eds.; Oxford University Press: New York, NY, 2012; pp. 957–992. [Google Scholar]
- Filippi, L.; Scaramuzzo, R.T.; Pascarella, F.; Pini, A.; Morganti, R.; Cammalleri, M.; Bagnoli, P.; Ciantelli, M. Fetal oxygenation in the last weeks of pregnancy evaluated through the umbilical cord blood gas analysis. Front. Pediatr. 2023, 11, 1140021. [Google Scholar] [CrossRef] [PubMed]
- Richardson, B.S.; de Vrijer, B.; Brown, H.K.; Stitt, L.; Choo, S.; Regnault, T.R.H. Gestational age impacts birth to placental weight ratio and umbilical cord oxygen values with implications for the fetal oxygen margin of safety. Early Hum. Dev. 2022, 164, 105511. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.M. Placental Gas Exchange and the Oxygen Supply to the Fetus. Compr. Physiol. 2015, 5, 1381–1403. [Google Scholar] [CrossRef]
- Fathollahipour, S.; Patil, P.S.; Leipzig, N.D. Oxygen Regulation in Development: Lessons from Embryogenesis towards Tissue Engineering. Cells Tissues Organs 2018, 205, 350–371. [Google Scholar] [CrossRef]
- Houghton, F.D. HYPOXIA AND REPRODUCTIVE HEALTH: Hypoxic regulation of preimplantation embryos: lessons from human embryonic stem cells. Reproduction 2021, 161, F41–F51. [Google Scholar] [CrossRef]
- Cowden Dahl, K.D.; Fryer, B.H.; Mack, F.A.; Compernolle, V.; Maltepe, E.; Adelman, D.M.; Carmeliet, P.; Simon, M.C. Hypoxia-inducible factors 1alpha and 2alpha regulate trophoblast differentiation. Mol. Cell. Biol. 2005, 25, 10479–10491. [Google Scholar] [CrossRef]
- Pringle, K.G.; Kind, K.L.; Sferruzzi-Perri, A.N.; Thompson, J.G.; Roberts, C.T. Beyond oxygen: complex regulation and activity of hypoxia inducible factors in pregnancy. Hum. Reprod. Update. 2010, 16, 415–31. [Google Scholar] [CrossRef]
- Dengler, V.L.; Galbraith, M.; Espinosa, J.M. Transcriptional regulation by hypoxia inducible factors. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 1–15. [Google Scholar] [CrossRef]
- Genbacev, O.; Zhou, Y.; Ludlow, J.W.; Fisher, S.J. Regulation of human placental development by oxygen tension. Science 1997, 277, 1669–1672. [Google Scholar] [CrossRef]
- Aplin, J.D. Hypoxia and human placental development. J. Clin. Investig. 2000, 105, 559–560. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.; Landers, K.; Mortimer, R.H.; Richard, K. Regulation of hypoxia inducible factors (HIF) in hypoxia and normoxia during placental development. Placenta. 2010, 31, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Dunwoodie, S.L. The role of hypoxia in development of the Mammalian embryo. Dev. Cell. 2009, 17, 755–773. [Google Scholar] [CrossRef]
- Fischer, B.; Bavister, B.D. Oxygen tension in the oviduct and uterus of rhesus monkeys, hamsters and rabbits. J. Reprod. Fertil. 1993, 99, 673–679. [Google Scholar] [CrossRef]
- Yedwab, G.A.; Paz, G.; Homonnai, T.Z.; David, M.P.; Kraicer, P.F. The temperature, pH, and partial pressure of oxygen in the cervix and uterus of woman and uterus of rats during the cycle. Fertil. Steril. 1976, 27, 304–309. [Google Scholar] [CrossRef]
- Ottosen, L.D.; Hindkaer, J.; Husth, M.; Petersen, D.E.; Kirk, J.; Ingerslev, H.J. Observations on intrauterine oxygen tension measured by fibre-optic microsensors. Reprod. Biomed. Online 2006, 13, 380–385. [Google Scholar] [CrossRef]
- Zhao, H.; Wong, R.J.; Stevenson, D.K. The Impact of Hypoxia in Early Pregnancy on Placental Cells. Int. J. Mol. Sci. 2021, 22, 9675. [Google Scholar] [CrossRef] [PubMed]
- Forristal, C.E.; Wright, K.L.; Hanley, N.A.; Oreffo, R.O.; Houghton, F.D. Hypoxia inducible factors regulate pluripotency and proliferation in human embryonic stem cells cultured at reduced oxygen tensions. Reproduction 2010, 139, 85–97. [Google Scholar] [CrossRef]
- Abdollahi, H.; Harris, L.J.; Zhang, P.; McIlhenny, S.; Srinivas, V.; Tulenko, T.; DiMuzio, P.J. The role of hypoxia in stem cell differentiation and therapeutics. J. Surg. Res. 2011, 165, 112–7. [Google Scholar] [CrossRef]
- Leese, H.J. Metabolic control during preimplantation mammalian development. Hum. Reprod. Update 1995, 1, 63–72. [Google Scholar] [CrossRef]
- Thompson, J.G.; Partridge, R.J.; Houghton, F.D.; Cox, C.I.; Leese, H.J. Oxygen uptake and carbohydrate metabolism by in vitro derived bovine embryos. J. Reprod. Fertil. 1996, 106, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Vaupel, P.; Multhoff, G. Revisiting the Warburg effect: historical dogma versus current understanding. J. Physiol. 2021, 599, 1745–1757. [Google Scholar] [CrossRef] [PubMed]
- Krisher, R.L.; Prather, R.S. A role for the Warburg effect in preimplantation embryo development: metabolic modification to support rapid cell proliferation. Mol. Reprod. Dev. 2012, 79, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.N.; Huang, X.B.; Muyayalo, K.P.; Mor, G.; Liao, A.H. Lactic Acid: A Novel Signaling Molecule in Early Pregnancy? . Front. Immunol. 2020, 11, :279. [Google Scholar] [CrossRef] [PubMed]
- Adelman, D.M.; Gertsenstein, M.; Nagy, A.; Simon, M.C.; Maltepe, E. Placental cell fates are regulated in vivo by HIF-mediated hypoxia responses. Genes Dev. 2000, 14, 3191–3203. [Google Scholar] [CrossRef] [PubMed]
- Robins, J.C.; Heizer, A.; Hardiman, A.; Hubert, M.; Handwerger, S. Oxygen tension directs the differentiation pathway of human cytotrophoblast cells. Placenta 2007, 28, 1141–1146. [Google Scholar] [CrossRef]
- Krock, B.L.; Skuli, N.; Simon, M.C. Hypoxia-induced angiogenesis: good and evil. Genes Cancer 2011, 2, 1117–1133. [Google Scholar] [CrossRef]
- Chang, C.W.; Wakeland, A.K.; Parast, M.M. Trophoblast lineage specification, differentiation and their regulation by oxygen tension. J. Endocrinol. 2018, 236, R43–R56. [Google Scholar] [CrossRef]
- Meschia, G.; Cotter, J.R.; Breathnach, C.S.; Barron, D.H. The hemoglobin, oxygen, carbon dioxide and hydrogen ion concentrations in the umbilical bloods of sheep and goats as sampled via indwelling plastic catheters. Q. J. Exp. Physiol. Cogn. Med. Sci. 1965, 50, 185–195. [Google Scholar] [CrossRef]
- Comline, R.S.; Silver, M. Daily changes in foetal and maternal blood of conscious pregnant ewes, with catheters in umbilical and uterine vessels. J. Physiol. 1970, 209, 567–586. [Google Scholar] [CrossRef]
- Dawes, G.S.; Fox, H.E.; Leduc, B.M.; Liggins, G.C.; Richards, R.T. Respiratory movements and rapid eye movement sleep in the foetal lamb. J. Physiol. 1972, 220, 119–143. [Google Scholar] [CrossRef] [PubMed]
- Clewlow, F.; Dawes, G.S.; Johnston, B.M.; Walker, D.W. Changes in breathing, electrocortical and muscle activity in unanaesthetized fetal lambs with age. J. Physiol. 1983, 341, 463–476. [Google Scholar] [CrossRef]
- Fowden, A.L.; Taylor, P.M.; White, K.L.; Forhead, A.J. Ontogenic and nutritionally induced changes in fetal metabolism in the horse. J. Physiol. 2000, 528, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Giussani, D.A.; Forhead, A.J.; Fowden, A.L. Development of cardiovascular function in the horse fetus. J. Physiol. 2005, 565, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Yamaleyeva, L.M.; Sun, Y.; Bledsoe, T.; Hoke, A.; Gurley, S.B.; Brosnihan, K.B. Photoacoustic imaging for in vivo quantification of placental oxygenation in mice. FASEB J. 2017, 31, 5520–5529. [Google Scholar] [CrossRef] [PubMed]
- Basak, K.; Luís Deán-Ben, X.; Gottschalk, S.; Reiss, M.; Razansky, D. Non-invasive determination of murine placental and foetal functional parameters with multispectral optoacoustic tomography. Light Sci. Appl. 2019, 8, 71. [Google Scholar] [CrossRef]
- Saini, B.; Morrison, J.; Seed, M. Gas Exchange across the Placenta. In Respiratory Disease in Pregnancy; Lapinsky, S., Plante, L., Eds.; Cambridge University Press: Cambridge, United Kingdom, 2020; pp. 34–56. [Google Scholar]
- Schneider, H. Oxygenation of the placental-fetal unit in humans. Respir. Physiol. Neurobiol. 2011, 178, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Templeton, A.; Kelman, G.R. Maternal blood-gases, (PAo2-Pao2), physiological shunt and VD/VT in normal pregnancy. Br J. Anaesth. 1976, 48, 1001–1004. [Google Scholar] [CrossRef]
- Assali, N.S.; Rauramo, L.; Peltonen, T. Measurement of uterine blood flow and uterine metabolism. VIII. Uterine and fetal blood flow and oxygen consumption in early human pregnancy. Am. J. Obstet. Gynecol. 1960, 79, 86–98. [Google Scholar] [CrossRef]
- Varma, S.; Cohen, H.J. Co-transactivation of the 3’ erythropoietin hypoxia inducible enhancer by the HIF-1 protein. Blood Cells Mol. Dis. 1997, 23, 169–176. [Google Scholar] [CrossRef]
- Jopling, J.; Henry, E.; Wiedmeier, S.E.; Christensen, R.D. Reference ranges for hematocrit and blood hemoglobin concentration during the neonatal period: data from a multihospital health care system. Pediatrics. 2009, 123, e333–e337. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, V.G.; Xu, J.; Orkin, S.H. Advances in the understanding of haemoglobin switching. Br. J. Haematol. 2010, 149, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, M.N.; Jedrusik, A.; Vuoristo, S.; Recher, G.; Hupalowska, A.; Bolton, V.; Fogarty, N.N.M.; Campbell, A.; Devito, L.; Ilic, D.; Khalaf, Y.; Niakan, K.K.; Fishel, S.; Zernicka-Goetz, M. Self-organization of the human embryo in the absence of maternal tissues. Nat. Cell Biol. 2016, 18, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.C.; Keith, B. The role of oxygen availability in embryonic development and stem cell function. Nat. Rev. Mol. Cell. Biol. 2008, 9, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Di Mattia, M.; Mauro, A.; Citeroni, M.R.; Dufrusine, B.; Peserico, A.; Russo, V.; Berardinelli, P.; Dainese, E.; Cimini, A.; Barboni, B. Insight into Hypoxia Stemness Control. Cells 2021, 10, 2161. [Google Scholar] [CrossRef]
- Semenza, G.L. Dynamic regulation of stem cell specification and maintenance by hypoxia-inducible factors. Mol. Aspects Med. 2016, 47-48, 15–23. [Google Scholar] [CrossRef]
- Morrison, S.J.; Csete, M.; Groves, A.K.; Melega, W.; Wold, B.; Anderson, D.J. Culture in reduced levels of oxygen promotes clonogenic sympathoadrenal differentiation by isolated neural crest stem cells. J. Neurosci. 2000, 20, 7370–7376. [Google Scholar] [CrossRef]
- Studer, L.; Csete, M.; Lee, S.H.; Kabbani, N.; Walikonis, J.; Wold, B.; McKay, R. Enhanced proliferation, survival, and dopaminergic differentiation of CNS precursors in lowered oxygen. J. Neurosci. 2000, 20, 7377–7383. [Google Scholar] [CrossRef]
- Zhang, Y.; Porat, R.M.; Alon, T.; Keshet, E.; Stone, J. Tissue oxygen levels control astrocyte movement and differentiation in developing retina. Brain Res. Dev. Brain Res. 1999, 118, 135–145. [Google Scholar] [CrossRef]
- Nakamura-Ishizu, A.; Kurihara, T.; Okuno, Y.; Ozawa, Y.; Kishi, K.; Goda, N.; Tsubota, K.; Okano, H.; Suda, T.; Kubota, Y. The formation of an angiogenic astrocyte template is regulated by the neuroretina in a HIF-1-dependent manner. Dev. Biol. 2012, 363, 106–114. [Google Scholar] [CrossRef]
- Gao, L.; Chen, X.; Zeng, Y.; Li, Q.; Zou, T.; Chen, S.; Wu, Q.; Fu, C.; Xu, H.; Yin, Z. Q. Intermittent high oxygen influences the formation of neural retinal tissue from human embryonic stem cells. Sci. Rep. 2016, 6, 29944. [Google Scholar] [CrossRef]
- Heinis, M.; Simon, M.T.; Ilc, K.; Mazure, N.M.; Pouysségur, J.; Scharfmann, R.; Duvillié, B. Oxygen tension regulates pancreatic beta-cell differentiation through hypoxia-inducible factor 1alpha. Diabetes. 2010, 59, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Ngo, M.A.; Sinitsyna, N.N.; Qin, Q.; Rice, R.H. Oxygen-dependent differentiation of human keratinocytes. J. Invest. Dermatol. 2007, 127, 354–361. [Google Scholar] [CrossRef] [PubMed]
- van Wenum, M.; Adam, A.A.A.; van der Mark, V.A.; Chang, J.C.; Wildenberg, M.E.; Hendriks, E.J.; Jongejan, A.; Moerland, P.D.; van Gulik, T.M.; Oude Elferink, R.P.; Chamuleau, R.A.F.M.; Hoekstra, R. Oxygen drives hepatocyte differentiation and phenotype stability in liver cell lines. J. Cell. Commun. Signal. 2018, 12, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Berthelemy, N.; Kerdjoudj, H.; Schaaf, P.; Prin-Mathieu, C.; Lacolley, P.; Stoltz, J.F.; Voegel, J.C.; Menu, P. O2 level controls hematopoietic circulating progenitor cells differentiation into endothelial or smooth muscle cells. PLoS One 2009, 4, e5514. [Google Scholar] [CrossRef]
- Baker, C.D.; Ryan, S.L.; Ingram, D.A.; Seedorf, G.J.; Abman, S.H.; Balasubramaniam, V. Endothelial colony-forming cells from preterm infants are increased and more susceptible to hyperoxia. Am. J. Respir. Crit. Care Med. 2009, 180, 454–461. [Google Scholar] [CrossRef]
- Wisgrill, L.; Schüller, S.; Bammer, M.; Berger, A.; Pollak, A.; Radke, T.F.; Kögler, G.; Spittler, A.; Helmer, H.; Husslein, P.; Gortner, L. Hematopoietic stem cells in neonates: any differences between very preterm and term neonates? PLoS One. 2014, 9, e106717. [Google Scholar] [CrossRef]
- Javed, M.J.; Mead, L.E.; Prater, D.; Bessler, W.K.; Foster, D.; Case, J.; Goebel, W.S.; Yoder, M.C.; Haneline, L.S.; Ingram, D.A. Endothelial colony forming cells and mesenchymal stem cells are enriched at different gestational ages in human umbilical cord blood. Pediatr. Res. 2008, 64, 68–73. [Google Scholar] [CrossRef]
- Borghesi, A.; Massa, M.; Campanelli, R.; Bollani, L.; Tzialla, C.; Figar, T.A.; Ferrari, G.; Bonetti, E.; Chiesa, G.; de Silvestri, A.; Spinillo, A.; Rosti, V.; Stronati, M. Circulating endothelial progenitor cells in preterm infants with bronchopulmonary dysplasia. Am. J. Respir. Crit. Care Med. 2009, 180, 540–546. [Google Scholar] [CrossRef]
- Cammalleri, M.; Amato, R.; Dal Monte, M.; Filippi, L.; Bagnoli, P. The β3 adrenoceptor in proliferative retinopathies: “Cinderella” steps out of its family shadow. Pharmacol. Res. 2023, 190, 106713. [Google Scholar] [CrossRef]
- Puebla, M.; Tapia, P.J.; Espinoza, H. Key Role of Astrocytes in Postnatal Brain and Retinal Angiogenesis. Int. J. Mol. Sci. 2022, 23, 2646. [Google Scholar] [CrossRef]
- Ballabh, P. Pathogenesis and prevention of intraventricular hemorrhage. Clin Perinatol. 2014, 41, 47–67. [Google Scholar] [CrossRef]
- Spencer, B.L.; Mychaliska, G.B. Milestones for clinical translation of the artificial placenta. Semin. Fetal Neonatal Med. 2022, 27, 101408. [Google Scholar] [CrossRef] [PubMed]
- Filippi, L.; Pini, A.; Cammalleri, M.; Bagnoli, P.; Dal Monte, M. β3-Adrenoceptor, a novel player in the round-trip from neonatal diseases to cancer: Suggestive clues from embryo. Med. Res. Rev. 2022, 42, 1179–1201. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, L.; Stenson, B.J. Use of umbilical cord blood gas analysis in the assessment of the newborn. Arch. Dis. Child. Fetal Neonatal Ed. 2007, 92, F430–F434. [Google Scholar] [CrossRef]
- Brotanek, V.; Hendricks, C.H.; Yoshida, T. Changes in uterine blood flow during uterine contractions. Am. J. Obstet. Gynecol. 1969, 103, 1108–1116. [Google Scholar] [CrossRef]
- Szymanowski, P.; Szepieniec, W.K.; Zarawski, M.; Gruszecki, P.; Szweda, H.; Jóźwik, M. The impact of birth anesthesia on the parameters of oxygenation and acid-base balance in umbilical cord blood. J. Matern. Fetal Neonatal Med. 2020, 33, 3445–3452. [Google Scholar] [CrossRef]
- Raghuraman, N.; Temming, L.A.; Doering, M.M.; Stoll, C.R.; Palanisamy, A.; Stout, M.J.; Colditz, G.A.; Cahill, A.G.; Tuuli, M.G. Maternal Oxygen Supplementation Compared With Room Air for Intrauterine Resuscitation: A Systematic Review and Meta-analysis. JAMA Pediatr. 2021, 175, 368–376. [Google Scholar] [CrossRef]
- Ayres-de-Campos, D.; Arulkumaran, S.; FIGO Intrapartum Fetal Monitoring Expert Consensus Panel. FIGO consensus guidelines on intrapartum fetal monitoring: Physiology of fetal oxygenation and the main goals of intrapartum fetal monitoring. Int. J. Gynaecol. Obstet. 2015, 131, 5–8. [Google Scholar] [CrossRef]
- Executive summary: Neonatal encephalopathy and neurologic outcome, second edition. Report of the American College of Obstetricians and Gynecologists’ Task Force on Neonatal Encephalopathy. Obstet. Gynecol. 2014, 123, 896–901. [CrossRef]
- Westgate, J.; Garibaldi, J.M.; Greene, K.R. Umbilical cord blood gas analysis at delivery: a time for quality data. Br. J. Obstet. Gynaecol. 1994, 101, 1054–1063. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, A.G. Clinical application of the Henderson-Hasselbalch equation. Appl. Ther. 1965, 7, 730–6. [Google Scholar] [PubMed]
- Siggaard-Andersen, O.; Engel, K. A new acid-base nomogram. An improved method for the calculation of the relevant blood acid-base data. Scand. J. Clin. Lab. Invest. 1960, 12, 177–186. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Flow chart illustrating patient enrollment of this retrospective observational cohort study.
Figure 1.
Flow chart illustrating patient enrollment of this retrospective observational cohort study.
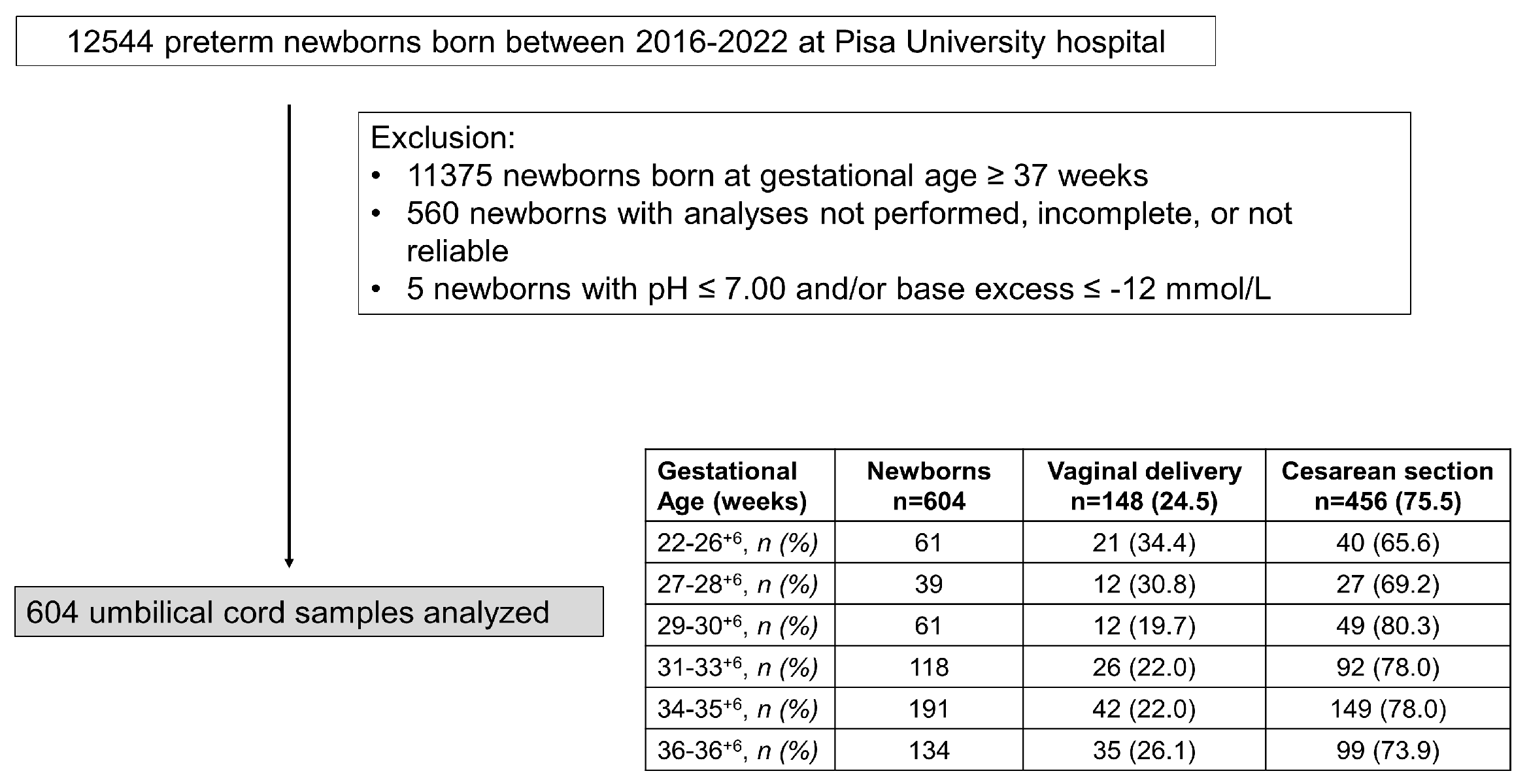
Figure 2.
Scatter plots with regression lines representing umbilical-cord oxygenation status [venous pO2, panel A (n=532); arterial pO2, panel B (n=500); fetal oxygen extraction, panel C (n=399)] of the whole study population.
Figure 2.
Scatter plots with regression lines representing umbilical-cord oxygenation status [venous pO2, panel A (n=532); arterial pO2, panel B (n=500); fetal oxygen extraction, panel C (n=399)] of the whole study population.
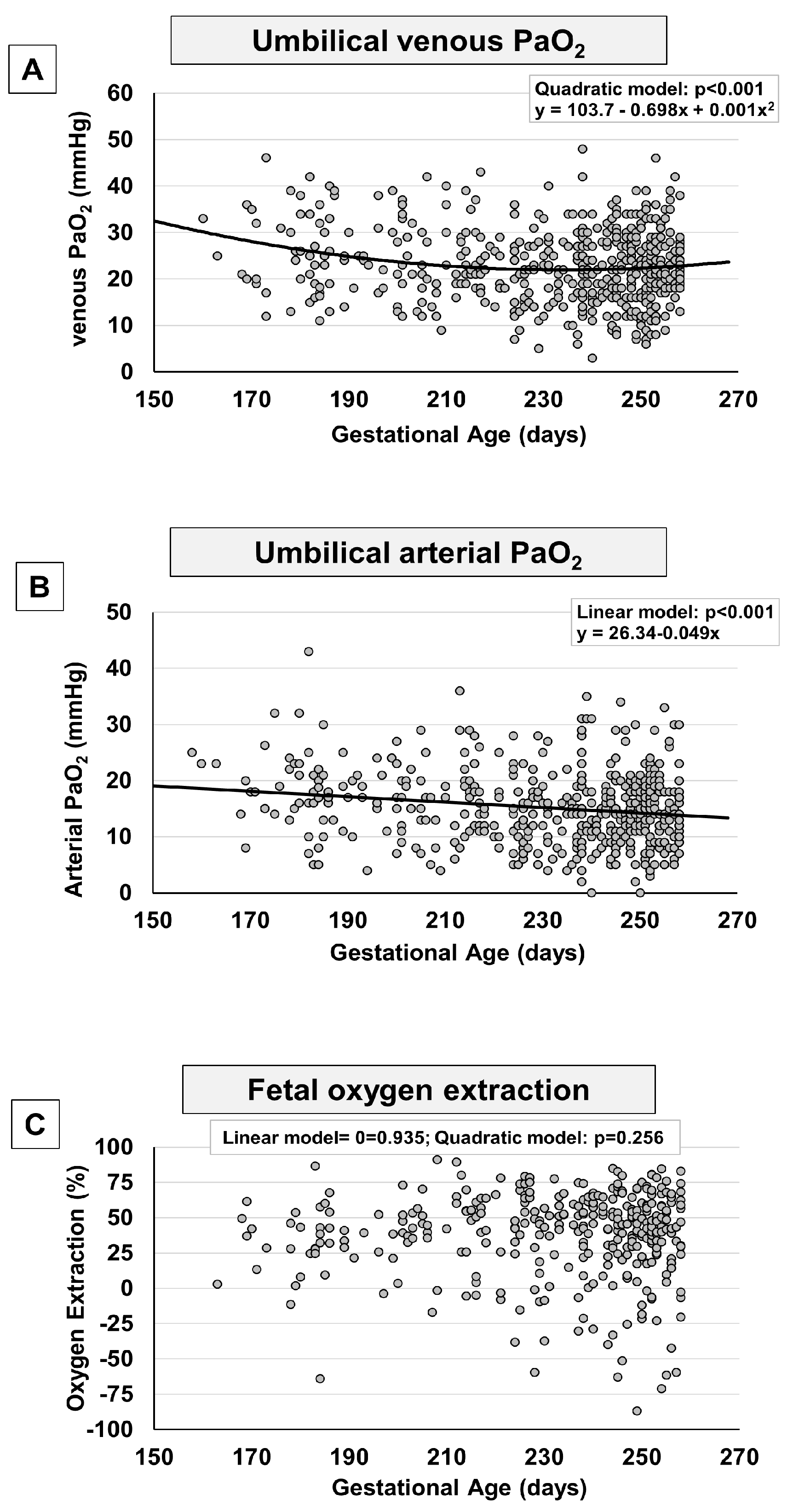
Figure 3.
Scatter plots with regression lines representing umbilical-cord Hb and oxygen content [venous Hb, panel A (n=566); arterial Hb, panel B (n=534); fetal venous oxygen content, panel C (n=427)] of the whole study population.
Figure 3.
Scatter plots with regression lines representing umbilical-cord Hb and oxygen content [venous Hb, panel A (n=566); arterial Hb, panel B (n=534); fetal venous oxygen content, panel C (n=427)] of the whole study population.
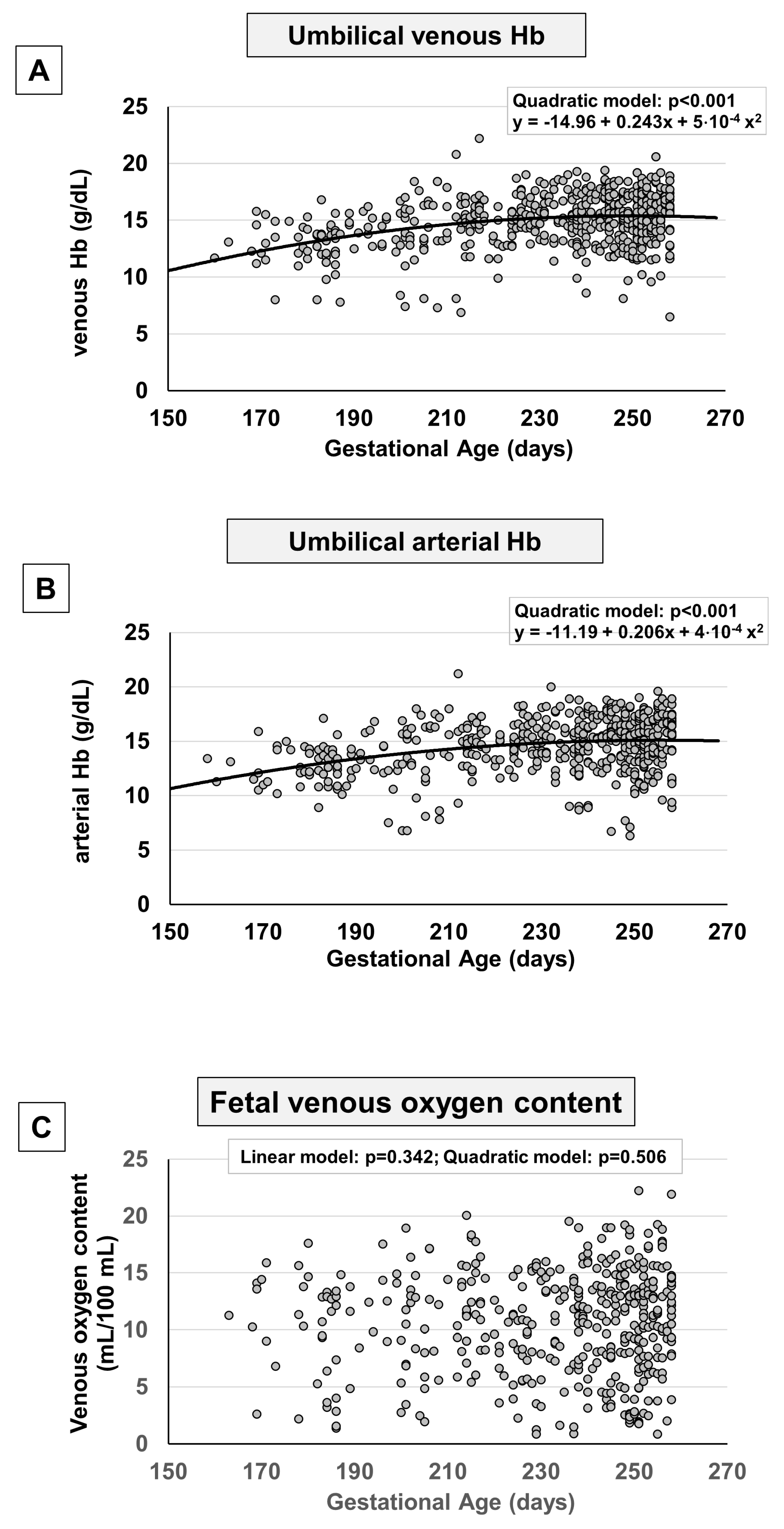
Figure 4.
Scatter plots with regression lines representing umbilical-cord pH, Base excess, and Bicarbonate [venous pH, panel A (n=584); arterial pH, panel B (n=552); venous Base excess, panel C (n=582); arterial Base excess, panel D (n=554); venous Bicarbonate, panel E (n=573); arterial Bicarbonate, panel F (n=541)] of the whole study population.
Figure 4.
Scatter plots with regression lines representing umbilical-cord pH, Base excess, and Bicarbonate [venous pH, panel A (n=584); arterial pH, panel B (n=552); venous Base excess, panel C (n=582); arterial Base excess, panel D (n=554); venous Bicarbonate, panel E (n=573); arterial Bicarbonate, panel F (n=541)] of the whole study population.
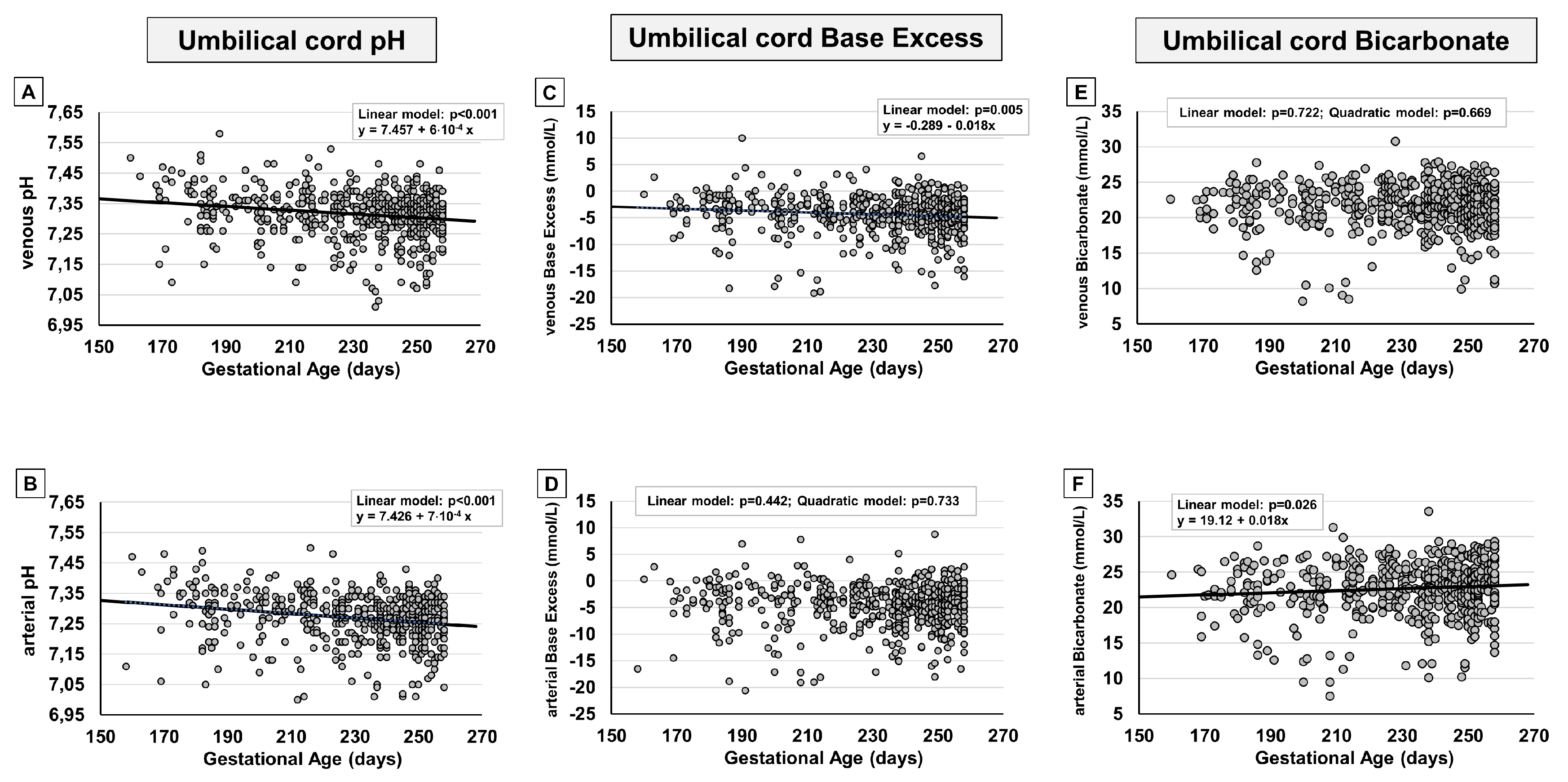
Figure 5.
Scatter plots with regression lines representing umbilical-cord carbon dioxide levels [venous pCO2, panel A (n=580); arterial pCO2, panel B (n=548); fetal CO2 production, panel C (n=527)].
Figure 5.
Scatter plots with regression lines representing umbilical-cord carbon dioxide levels [venous pCO2, panel A (n=580); arterial pCO2, panel B (n=548); fetal CO2 production, panel C (n=527)].
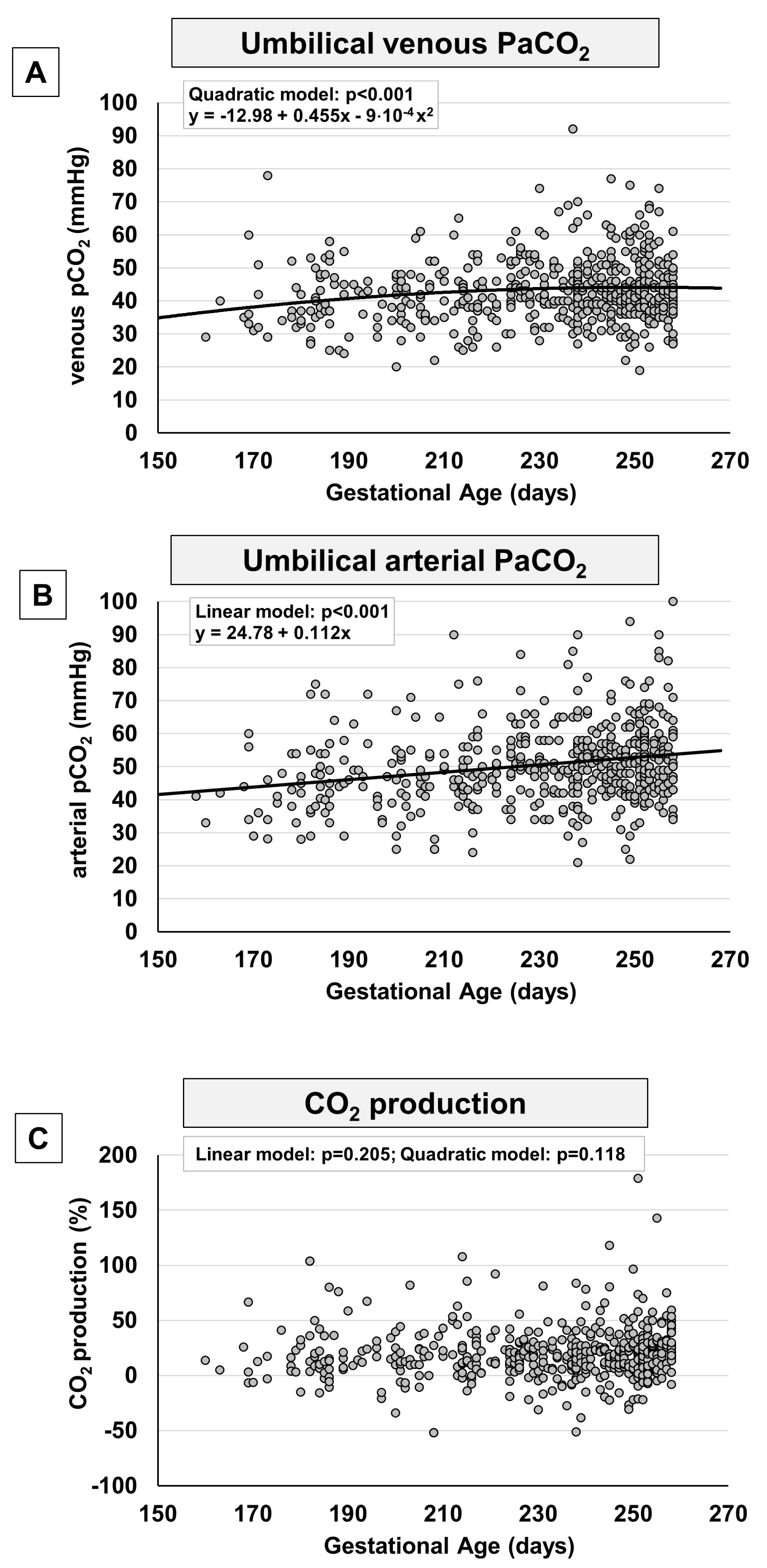
Figure 6.
Scatter plots with regression lines representing umbilical-cord lactate levels [venous Lactate, panel A (n=513); arterial Lactate, panel B (n=481); fetal Lactate production, panel C (n=465)].
Figure 6.
Scatter plots with regression lines representing umbilical-cord lactate levels [venous Lactate, panel A (n=513); arterial Lactate, panel B (n=481); fetal Lactate production, panel C (n=465)].
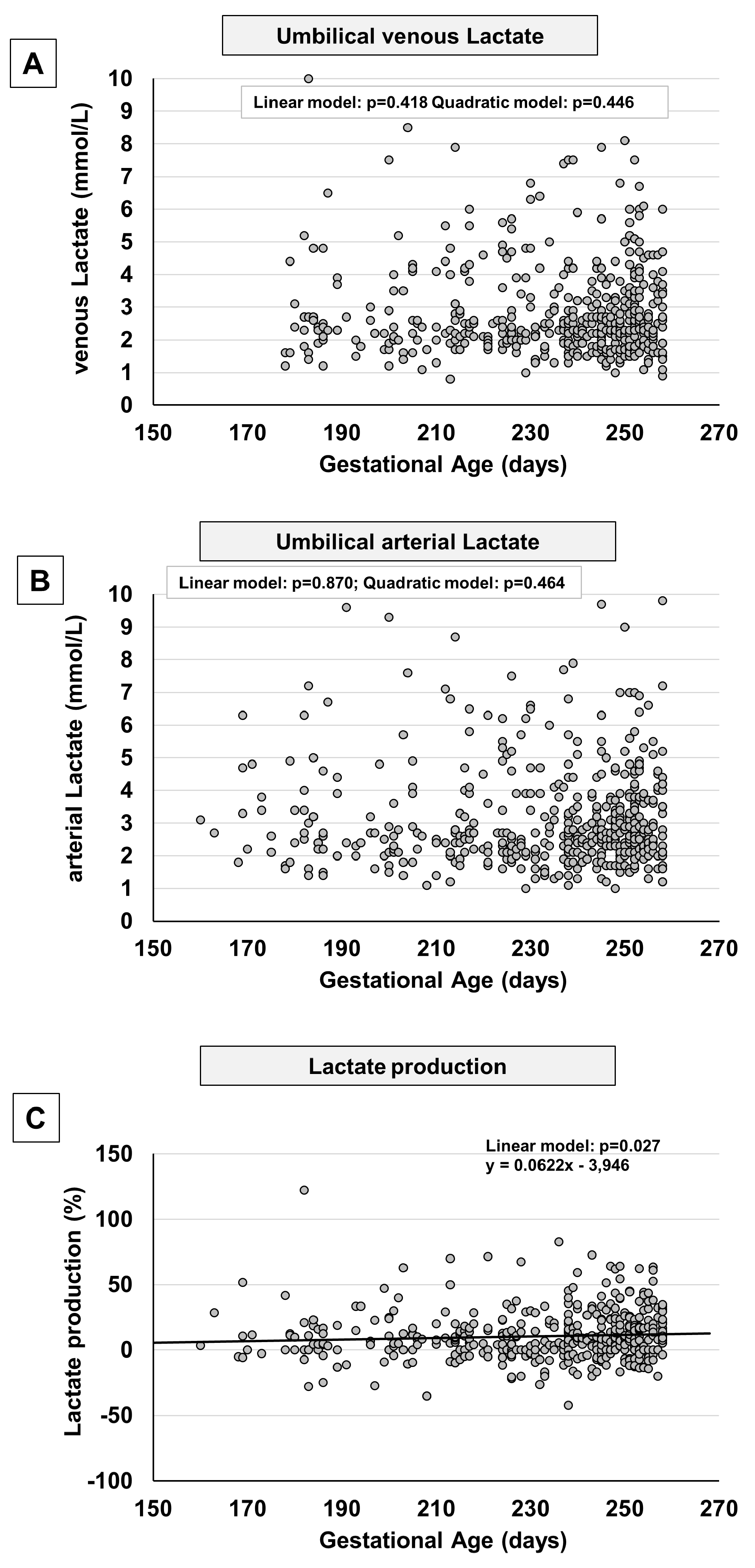

Table 1.
Umbilical-cord blood gas analysis in all enrolled preterm newborns and separately analyzed by the type of delivery.
Table 1.
Umbilical-cord blood gas analysis in all enrolled preterm newborns and separately analyzed by the type of delivery.
| All preterm newborns n=604 |
Vaginal delivery n=148 |
Cesarean section n=456 |
p value | |
|---|---|---|---|---|
| GA, days, mean (SD) | 230 (24) | 228 (28) | 231 (23) | 0.127 |
| Birth weight, g, mean (SD) | 1855 (687) | 1926 (728) | 1832 (672) | 0.155 |
| Male, n (%) | 310 (51.2) | 80 (54.0) | 230 (50.4) | 0.474 |
| Apgar Score, mean (SD) | 7.6 (1.2) | 7.6 (1.4) | 7.7 (1.1) | 0.781 |
| Umbilical venous cord sampling | ||||
| pH, mean (SD) | 7.315 (0.08) | 7.334 (0.08) | 7.309 (0.08) | < 0.001 |
| pCO2, mmHg, mean (SD) | 43.2 (9.4) | 39.9 (9.5) | 44.3 (9.2) | < 0.001 |
| pO2, mmHg, mean (SD) | 22.8 (7.9) | 25.3 (8.3) | 22.0 (7.6) | < 0.001 |
| Bicarbonate, mmol/L, mean (SD) | 21.7 (3.1) | 20.9 (2.7) | 22.0 (3.1) | < 0.001 |
| BE(B), mmol/L, mean (SD) | -4.3 (3.6) | -4.6 (3.5) | -4.3 (3.6) | 0.250 |
| Lactate, mmol/L, mean (SD) | 2.8 (1.4) | 3.3 (1.4) | 2.7 (1.3) | < 0.001 |
| Hemoglobin, g/dL, mean (SD) | 14.96 (2.3) | 15.5 (1.9) | 14.8 (2.4) | 0.001 |
| SaO2, %, mean (SD) | 52.9 (22.1) | 57.4 (21.4) | 51.3 (22.2) | 0.011 |
| Umbilical arterial cord sampling | ||||
| pH, mean (SD) | 7.267 (0.08) | 7.274 (0.09) | 7.278 (0.08) | 0.327 |
| pCO2, mmHg, mean (SD) | 50.7 (11.4) | 49.7 (11.8) | 51.0 (11.3) | 0.243 |
| pO2, mmHg, mean (SD) | 15.2 (6.9) | 17.6 (6.7) | 14.4 (6.8) | < 0.001 |
| Bicarbonate, mmol/L, mean (SD) | 22.7 (3.8) | 22.3 (3.2) | 22.8 (3.9) | 0.227 |
| BE(B), mmol/L, mean (SD) | -4.6 (4.1) | -4.8 (3.7) | -4.5 (4.3) | 0.414 |
| Lactate, mmol/L, mean (SD) | 3.1 (1.5) | 3.7 (1.6) | 2.9 (1.5) | < 0.001 |
| Hemoglobin, g/dL, mean (SD) | 14.96 (2.3) | 15.3 (2.1) | 14.4 (2.5) | < 0.001 |
| SaO2, %, mean (SD) | 33.0 (19.2) | 40.4 (20.0) | 30.0 (18.2) | < 0.001 |
| Veno-arterial O2 difference, mmHg, mean (SD) | 7.7 (6.4) | 7.5 (7.1) | 7.8 (6.2) | 0.665 |
| Fetal oxygen extraction, %, mean (SD) | 35.7 (35.8) | 27.3 (32.0) | 38.6 (36.8) | 0.007 |
| GA = gestational age; pO2=partial pressure of oxygen; pCO2 = partial pressure of carbon dioxide; BE=base excess | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



