
Submitted:
05 July 2023
Posted:
07 July 2023
You are already at the latest version
Abstract
Plant roots host various microorganisms around and inside their roots, known as the root microbiome. To become healthy and productive, plants should keep under surveillance niches around the roots to recognize disease-causing microbes and similarly exploit the services of beneficial microorganisms in nutrient acquisition, stress mitigation and growth promotion. Here we presented the communication strategies between plant roots and root-associated microbes in improving plant growth and yield. Understanding how plant root and root-associated microbes communicate is vital in designing ecofriendly strategies for targeted disease suppression and improved plant growth that will help in sustainable agriculture.
Keywords:
plant-microbe interactions
; signaling molecule
; root exudate
; disease suppression
; crop production
1. Introduction
In time past microbial ecologists used to face high challenges in investigating multifaceted microbial societies. However, today the tide has changed thanks to the methodological advances like the high throughput deoxyribonucleic acid sequencing machinery that provides comprehensive information on the composition and structure of microbial groups [1,2,3]. Though, the bulk of the investigative practice emphasis a single property of the communities being examined. For instance, studies unfolding and relating the structure of microbial societies often concentrate on distinct lineages present or the total numbers of taxa in a given sample (alpha diversity) or between samples (beta diversity) [4,5]. The same attention has not been given when using sequence data to decipher indirect or direct collaboration among microbial taxa and plants [6,7,8]. With the availability of many sequence datasets from environmental samples, the focus now is to go beyond alpha and beta diversity and look more at the interactions between microbial taxa and their host [9,10]. Studying interactions between taxa across intricate and different inhabitants like plant microbiomes will aid in establishing useful roles or ecological niches engaged by beneficial microorganisms in promoting plant health [11,12,13]. Plants harbor diverse groups of microorganisms that live inside and outside their roots. The root microbiome is the active community of microbes connected with plant roots, be it inside or around the root, which is extensively involved in plant wellbeing and serves as a receptacle of extra genes that plants can acquire when required [14].
The concept and relevance of the root microbiome to plant health will improve our insight into the colossal power of these tiny giants in ecosystem function. The microbial communities and interaction networks inside the root, root surface and bulk soil are distinct, although the similarity in species structure can be seen [15,16]. With the use of bar-coded pyrosequencing of rRNA genes, NR Gottel, HF Castro, M Kerley, Z Yang, DA Pelletier, M Podar, T Karpinets, E Uberbacher, GA Tuskan, R Vilgalys, et al. [17] revealed that Populus deltoides root endophytic bacterial and fungal communities are discrete groups rather than opportunistic subgroups of the rhizosphere. Significant differences between the bacterial interaction networks and diversity of Agave lechuguilla rhizosphere and bulk soil were also reported by NE López-Lozano, A Echeverría Molinar, EA Ortiz Durán, M Hernández Rosales and V Souza [18] when they used the 16S rRNA gene sequence from Agave lechuguilla rhizospheric and bulk soil samples. Root microbiome is a significant driver for plant yield, health, and ecosystem functioning because it is the intersection point between a plant and the ecosphere. Furthermore, it is accountable for key functions such as nutrient acquisition and signals vital to plant development. Nevertheless, how plant roots and their associated microbes communicate to enhance plant development is incipient. Understanding the communication of plant roots with microbial communities will provide sustainable solutions in raising agronomic crop production. Thus, this review presented communications between root microbiome and plant root, the mechanism employed by root-associated microbes in promoting plant growth and applications of plant root collaborations and root microbiome in improving plant growth and yield through stress mitigation, disease suppression and nutrient acquisition.
2. Communication between root microbiome and plant root
The rhizosphere of plants has evolved into diverse and complex microbial groups with different information-processing systems involved in plant enlargement and growth, and plant defense response [19]. These information-processing systems and signals that are involved in the recognition of beneficial organisms in the rhizosphere and counteracting immune detection are still poorly understood [20]. As a result, recent research on immune signaling mediated by plant growth-promoting rhizobacteria is gaining considerable interest.
Plants induce substantial selection pressure on the development of the rhizobiome are, achieved by the emission of exudates from plant roots. The secreted root exudates contain compounds of different kinds that attract the development of specific plant microbiota. The attracted organisms utilize these exudates as sources of energy and multiply in the vicinity [20]. Plant and microorganisms' ability to communicate before initiating somatic contact is crucial. It helps each partner to take full advantage of the association and alleviate the threat of damage. It also lets plants regulate the microbe they interact with throughout their development and growth process [21]. The formation of these relations entails synchronized crosstalk among the plant and a well-suited microbe that occurs in signals from the interacting organisms [21].
Plants have developed a myriad of communication systems to integrate information from the environment and to actively respond to abiotic and biotic factors [22]. They have equally developed a means of communication between plants and their associated microbes through transkingdom signaling [22]. Communication in the rhizosphere is highly sophisticated and controlled by a wide range of specialized exudates and metabolites. These exudates and metabolites end up in the altered gene expression in one or both of the interacting partners [20]. These exudates, mostly nutrients, are used by the rhizosphere organisms to colonize plant roots. This sophisticated form of communication results in plant growth promotion, control of soil-borne pathogens, availability of nutrients, biofilm formation and accumulation of soil microbes [20]. Nevertheless, numerous compounds in the rhizosphere, produced by plants or microbes, could likewise act as signals for communication [23].
Signals are low molecular-weight diffusible compounds produced by one organism and recognized by another organism that elicit, at low concentration, a specific response in the latter through a signal-transducing cascade [23]. Upon signal recognition, the molecules can be up or down-regulating gene expression and alter the physiology and activities of the receiving organism [24,25]. In the rhizosphere, there is a numerous number of these molecules whose concentration varies according to the distance from the point of emission. These deposits are collectively known as rhizodeposits that include sloughed-off tissue and cells, H+ efflux, CO2 from cell respiration, mucilage, intact root border cells and proteins [26]. The low molecular weight organic compounds, also known as root exudates, contain amides, sugars, phenolic, aromatic and amino acids. They facilitate communications and function as chemical attractants and repugnant that drive the root microbiome [27]. The recognition of these signals is important in driving the plant-microbe networking system. Plant-microbe communication through signal molecules can influence the stimulation and suppression of gene expression in both partners. This communication protects plants against abiotic and biotic stresses, enhanced nutrient acquisition and availability, and promotes plant growth and health [25]. With these root exudates, plants can interact with microbes to stimulate succor in environmental adjustment to relieve stresses such as drought-restrictive nutrient procurement, metal toxicity, and pathogen spell [28]. This synergy leads to more intricate collaborations, inducing plant growth and resource competition. Similarly, signal molecules are crucial for root colonization by rhizospheric microbes and for establishing plant-microbe interactions. Pseudomonas fluorescens establishes interaction and root colonization with tomato by the o-antigenic side chain of bacterial lipopolysaccharide [29], while the initial host recognition in cereals is achieved by the major outer membrane proteins (MOMP) in Azospirillum brasilense [30]. Plants also employ the rhizophagy cycle/process in recruiting beneficial microbes by producing the exudates they feed on. The root-associated microbes employ direct or indirect strategies to impact plant health status and growth [31]. Direct strategies include nutrient acquirement, phytohormones production, and phosphate solubilization. Indirect strategies occur by eliciting plant immune responses, preventing plant pathogens from proliferating and competing with their resources (Figure 1) [32]. For example, IA Stringlis, K Yu, K Feussner, R de Jonge, S Van Bentum, MC Van Verk, RL Berendsen, PA Bakker, I Feussner and CM Pieterse [33] showed that the probiotic-plant-rhizobacteria-collaboration elicited the root-specific transcription factor MYB72 and further led to the production and emission of MYB72-controlled β-glucosidase BGLU42-reliant scopolin and scopoletin, respectively, resulting in a well-established niche for microbial consortiums and resistance profits for the host plant against Verticillium dahlia and Fusarium oxysporum (soil-borne fungal pathogens). Bacterial assemblages connected with plant roots contribute a vital role in subduing soil-borne pathogens, and multispecies probiotic associations could boost disease suppression efficiency. For example, J Hu, Z Wei, V-P Friman, S-h Gu, X-f Wang, N Eisenhauer, T-j Yang, J Ma, Q-r Shen, Y-c Xu, et al. [34] reported that the addition of Pseudomonas consortia in Solanum lycopersicum rhizosphere microbiome reduced Ralstonia solanacearum concentration and lessened the disease incidence because of the meddling and increased resource competition with the pathogen. Similarly, an increase in the Pseudomonas consortia richness resulted in increased plant biomass and effective absorption of nutrients in Solanum lycopersicum plants.
Root-related microbes likewise produce communicating compounds ranging from antibiotics, organic acids, volatile signals, phytohormones, extracellular enzymes and quorum sensing molecules (QSM) [35]. These compounds aid plant roots associated microbe's relationship that facilitates plant growth. For instance, N-acyl-L-homoserine lactones (quorum-sensing molecules) were reported by R Ortiz-Castro [36] to influence the lateral root formation, root system architecture, primary root growth, and root hair development of Arabidopsis thaliana in their post-embryonic stage. Bacterial strains such as Bacillus amyloliquefaciens L3 use-communicating molecules like volatile organic compounds to stimulate reactions in fungi and plants and recruit induced systemic resistance (ISR) in plants, consequently eliciting expression of defense genes that mitigate the negative effect of viruses, oomycetes, bacteria, and fungi on plants [37,38,39,40]. Root-resultant exudates, apart from aiding plant fitness and longevity, also profit microorganisms that use them as a resource (carbon-rich products with other nutrients) that support microbial multiplication [41,42,43].
Alongside different rhizodeposits produced in the rhizosphere, different hormones are also produced that aid plant-microbe communication cascades [25]. These hormones include abscisic acid, auxin, cytokinins, gibberellin and peptide hormones that regulate plant growth and development. Several plant growth-promoting rhizobacteria have been reported to produce indole acetic acid in chemically defined medium with tryptophan precursor [44,45]. Auxin production enhances seed germination, nutrient uptake and root growth and development [25,46]. For instance, cytokinin was found to stimulate cell division, inhibit root elongation and affect root hair development [47], while gibberellin was reported to alter many physiological and developmental processes in plants by promoting seed germination, stem elongation, flowering and fruit setting in plants [48]. Gibberellin also facilitates cell-to-cell communication [25].
3. Applications of plant root-associated microbes in improving plant growth and yield
The increase in farming productivity to meet the high food expectations of the human populace is a problem of great worry for all nations. Chemical fertilization to boost crop production has long alleviated these worries [49]. However, the prolonged misused of chemical fertilizers in farming has negatively imparted the ecosystem [50] and has led to the call for ecofriendly approach to boost crop production and to increase plant production is to make plant healthy [51,52]. To be healthy, plants must mitigate environmental disturbances. The plant root-associated microbes in mitigating abiotic and biotic stress, nutrient acquisition, and growth promotion need to be promoted to enhance sustainable agriculture [53,54].
3.1. Root microbiome role in abiotic stress tolerance
Crops are often exposed to physical stresses such as soil salinization, submergence, extreme temperatures, nutrient imbalances, drought, to mention but a few [55]. The fact that these stresses will rise in nearby days makes it a big concern as plant growth, yield, and productivity will be hindered. To overcome these abiotic stresses, plants must undergo adaptive modifications or solicit the help of beneficial microbes to live and will promote plant function [2,56]. Root-associated microbes can stimulate the growth and defend the host via many molecular pieces of machinery in abiotic stress circumstances (Table 1). CM Ribeiro and EJBN Cardoso [57] revealed that strains of Bacillaceae, Enterobacteriaceae, and Pseudomonadaceae isolated from the Araucaria angustifolia root were tremendous plant growth-promoting bacteria. Some of these bacterial strains are P-solubilizing microbes that help the crops tolerate drought, salt, and extreme temperature conditions through the production of numerous phytohormones, antioxidants, exopolysaccharide, 1-aminocyclopropane-1-carboxylate deaminase, enrichment of nutrient uptake, many volatile compounds, and initiation of the buildup of osmolytes. They also help in the regulation of stress-responsive genes [58,59]. Under drought settings, T Yuwono, D Handayani and J Soedarsono [60] found that osmotolerant rhizobacterial inoculated with rice increased root and shoot dry weight. It was also proven that under stress conditions, these isolates had what it took to produce betaine, signifying that the drought tolerance was because of the increase in osmolyte. They also revealed that the rhizobacteria-plant interaction led to indole acetic acid production. Under drought experiment conducted by JM Ruiz-Lozano, C Collados, JM Barea and R Azcón [61] showed that co-inoculation of Glomus mosseae and Bradyrhizobium japonicum in drought-stressed soybean plants resulted in increased leghemoglobin content, acetylene reductase activity and protein content by 25%, 112% and 15% respectively compared with well watered soybean plants and plants colonized by Bradyrhizobium alone.
Root associated microbes also use other strategies to retain ion homeostasis in plants facing salt stress. For instance, bacterial exopolysaccharides fix to Na+, and confine Na+ inflow into roots. They produced volatile organic compounds (VOCs) during stress conditions so that VOCs can activate high-affinity K+ transporter (HKT1) reduction in root and stimulate HKT1 in shoots, thereby restraining Na+ entrance into roots and easing shoot-to-root Na+ retransmission. The K+/Na+ ratio is increased by arbuscular mycorrhizal fungi by immensely improving Ca 2 + and K+ absorption and eluding the movement of toxic Na+ under salty circumstances. Furthermore, for effective water assimilation in saline-strained plants, root closely associated microbes control the processes of genes encrypting the plasma membrane integral proteins to aquaporin activity [62,63,64]. Boosting the antioxidative systems in plants for ROS (reactive oxygen species), scavenging, and production of polyamines and proline are also part of mechanisms employed by root-associated microbes for mitigating salt stress in plants. A Bano and M Fatima [65] induced salt stress conditions and co-applied Pseudomonas and Rhizobium at the seedling stage of maize. Their findings showed that under sodium chloride conditions alone, a harmful effect on maize growth and development was seen. Furthermore, improved sodium chloride tolerance of maize upon co-inoculation with Pseudomonas and Rhizobium is linked with reduced electrolyte leakage, increased proline production and conservation of leaf water contents. The improvement of nutrient uptake to boost plant survival under salt conditions is another mechanism employed by root-associated microbes. For instance, the introduction of Bacillus aquimaris to wheat plants resulted in a substantial rise in phosphorus, nitrogen and potassium in wheat leaves (Upadhyay and Singh 2015). The root-associated microbe can also aid plants to withstand high or low-temperature conditions, either by increasing or decreasing anthocyanin, proline and sugar contents. [66] reported that under low temperatures, Burkholderia phytofirmans strain PsJN bacterized with grapevine plant, increased physiological activity and grapevine growth through a substantial increase in proline, starch deposition, carbohydrates and phenol contents compared with the control.

Table 1.
Root microbiome and their mitigation on abiotic stress confronting plant.
| Stress type | Root associated microbes | Plant host | Inoculated with | Activities | The effect on plant | Reference |
|---|---|---|---|---|---|---|
| Drought | Enterobacter, Bacillus, Moraxella and Pseudomonas | Acacia arabica | Triticum aestivum L. | indole-3-carboxylic acid, Indole-3-lactic acid and indole-3-acetic acid production | Improvement in shoot length, tillers and number of spikelets and increases in spike length and seed weight of Triticum aestivum L. | [67] |
| Salt | Halomonas and one Bacillus |
Salicornia rubra, Sarcocornia utahensis, and Allenrolfea occidentalis |
Alfalfa | - | The total biomass of alfalfa increased, and root length were improved by 2.6 and 1.5 fold in Halomonas and Bacillus inoculated plants, respectively, compared with the uninoculated alfalfa | [68] |
| salt or drought | Bacillus amyloliquefaciens SB-9 | grapevine | Grapevine plantlet | melatonin secretion, 5-hydroxytryptophan, serotonin, and N-acetylserotonin | Lessen the antagonistic effects of salt- and drought-induced stress by decreasing the secretion of malondialdehyde, O2- and H2O2 (reactive oxygen species) in roots. | [69] |
| Heavy metal stress | Phialocephala fortinii, Rhizodermea veluwensis, and Rhizoscyphus sp | Clethra barbinervis | Clethra barbinervis seedling | Siderophores | Improved K absorption in shoots and decreased the concentrations of Cd, Zn, Pb, Cu, and Ni in roots. | [70] |
| Heavy metal | Penicillium ruqueforti Thom | Solanum surattense Burm | Wheat seedling | indole-3-acetic acid | It led to low concentrations of heavy metals in the root and shoot. In increase nutrient uptake and higher plant growth, | [71] |
| Heat | Thermomyces sp | Cullen plicata | cucumber | Increase in antioxidant enzyme activities, soluble proteins, flavonoids, saponins, and total sugars. | It maintains the optimal quantum efficiency of photosystem II, water use efficiency, and photosynthesis rate and increases the root length, induced accumulation of saponins, total sugars, soluble proteins, flavonoids, and antioxidant enzyme activities | [72] |
| High temperature, salinity, and glyphosate pollution | Ochrobactrum cytisi strain IPA7.2 | Solanum tuberosum L. | Solanum tuberosum L | .indole-3-acetic acid and type II 5-enolpyruvylshikimate-3-phosphate synthase | Improved the mitotic index of root meristem cells, the number of roots, the number of leaves and the length of shoots | [73] |
| Flood | Klebsiella variicola AY13 | soybean | soybean | Indole acetic acid production | Plants' growth improved with enriched chlorophyll content and quantum efficiency of chlorophyll fluorescence | [74] |
3.2. Root microbiome role in nutrient acquisition
Most micronutrients and macronutrients important for plant growth are available in the soil in insoluble forms. Plants devise several mechanisms for the acquisition of these nutrients in the soil. The plant root microbiome enhances the uptake of major micronutrients by mineralizing or solubilizing them and ensuring their bioavailability through acidification [75,76], secretion of hydrolytic enzymes such as phytase or phosphatase, excretion of proton and production of siderophore [77]. Endophytes, rhizospheric microbiomes and Arbuscular mycorrhizal fungi (AMF) help the plant in the acquisition of nutrients from the soil through the solubilization of nutrients such as sulphur (S), potassium (K), calcium (Ca), iron (Fe), zinc (Zn) and phosphorus (P) [78,79,80]. Some notable root microbiome genera associated with maize, wheat rice and legumes such as Streptomyces, Pantoea, Citrobacter, Azospirillum, Bacillus, Herbaspirillum, Achromobacter, Gluconacetobacter, Burkholderia, Chryseobacterium, Bacillus, Klebsiella, Azotobacter and Pantoea have been reported to enhance plant development and growth via the uptake of micronutrients and stimulate the development of plant roots [81,82,83,84]. Siderophores secreted by endophytes aid plant's iron uptake from the soil; this is because iron cannot directly penetrate the plant cell even through transporters [85]. Root endophytes, such as Azoarcus, Herbaspirillum, Acetobacter and diazotrophicus, have been reported to be active in nitrogen fixation. Some diazotrophic endophytic microbial communities, such as Bacillus, Gammaproteobacteria, and Actinobacteria have been largely reported as an atmospheric nitrogen fixer in rice [85,86,87]. Rhizobia, most importantly, Burkholderiales form root nodules with legumes, which convert atmospheric nitrogen into ammonia, which is readily available to the plant, while the plant, in return, produces carbon compounds [88,89]. Some root microbiomes such as Brevibacillus, Kineococcus, Microbacterium, Rhizobium, Burkholderia, Nocardia, Bacillus, Rhodococcus, Methylobacterium, Mesorhizobium, and Paenibacillus, associated with Eucalyptus plant have been reported to be involved in the fixing of nitrogen [85,90]. A summary of other studies on nutrient acquisition attributes of some plant root microbiomes is presented in Table 2.
3.3. Root microbiome role in disease suppression/biocontrol
Insect and pathogens attack plants and retard their yield, growth and health. However, plant root microbiomes have been reported to be a reservoir of many bioactive metabolites that can protect and enhance plant resistance against attacks from pathogens and pests [85]. Phyllospheric microorganisms isolated from different crop plants showed the abundance of Firmicutes that is capable of secreting volatile organic compounds active in the protection of crops from several fungal and bacterial pathogens/diseases [109]. Plant root microbiome protects the plant through induced systemic resistance (ISR) or antibiosis from insects, pathogens and herbivores. Siderophores, antibiotics, salicylic acid, N-acyl homoserine lactones, lipopolysaccharide, jasmonic acid and flagella secreted by endophytic bacteria have been reported to be capable of inducing systemic resistance in plants [110]. Also, endophytic fungi, majorly of the phyla Glomeromycota, Basidiomycota, Ascomycota, and Zygomycota are capable of secreting inhibitory compounds, some of which are terpenoids, polyketones, phenols, chlorinated compound, alkaloids, peptides, steroids and flavonoids, which aid the protection of plants from insects, pathogens and herbivores [111]. Actinomycetes have also been widely studied due to its ability to secrete notable antimicrobial compounds active against plant pathogens. Streptomyces spp. secretes many antimicrobial compounds such as indolo-sesquiterpene antimicrobial compounds, munumbicins, coronamycin and kakadumycins [85,112,113]. Studies have also revealed that siderophore can induce ISR in plants and enhance biocontrol activities. For example, strains of endophytic methylobacterium successfully suppressed Xylella fastidiosa (a pathogen responsible for chlorosis in citrus trees) via siderophore production [53]. Rhizobiomes such as Actinobacteria, Proteobacteria and Firmicutes have been linked with the inhibition of Rhizoctonia solani commonly attacking sugar beet [85], while the Gammaproteobacteria also successfully inhibit the disease via non-ribosomal peptide synthesis (NRPS) [114]. A high abundance of bacteria such as Streptomyces, Bacillus, Paenibacillus, and Rhizobium in the root microbial community of cucumber cultivated and monitored in suppressive soil [115]. A summary of similar studies on the biocontrol attributes of some plant root microbiomes is presented in Table 3.
4. Conclusion and future prospects
It is evident that plant-microbe signaling cascades are essential regulators of plant development and growth, and these signal molecules can alter the morphology and physiology of the host plant. Plants develop complex interactions and communicate with various microbes in their rhizosphere through different signals that affect plant growth and modulate plant-specific core root microbiome. These signals, secreted by micro- and macro-symbionts, can enhance root development, increase nutrient and water uptake, and promote tolerance to biotic and abiotic stresses. As a result of the roles plant growth-promoting organisms played in improving plant growth and yield, the role of plant-microbe signals in sustainable agriculture and the recovery of marginal lands cannot be overemphasized. It is, therefore, important to focus future research on the understanding of intra- and inter-communication that can lead to the identification of more signal molecules and similarly improve plant growth and development. Thus, the need to develop efficient technologies for isolating and identifying signal compounds useful for sustainable development.
Author Contributions
All authors contributed equally and also approved the final draft.
Funding
This research received no external funding.
Data Availability Statement
Not applicable
Acknowledgments
B.J.E.: FAE and AAS thanks South Africa’s National Research Foundation/The World Academy of Science African Renaissance grant. Work in O.O.B.’s lab is based on the support of the National Research Foundation of South Africa (grants ref: UID81192, UID105248, UID95111; OOB).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Enagbonma, B.J.; Aremu, B.R.; Babalola, O.O. Profiling the functional diversity of termite mound soil bacteria as revealed by shotgun sequencing. Genes 2019, 10, 637. [Google Scholar] [CrossRef] [PubMed]
- Enagbonma, B.J.; Babalola, O.O. Unveiling plant-beneficial function as seen in bacteria genes from termite mound soil. J. Soil Sci. Plant Nutr. 2020, 1–10. [Google Scholar] [CrossRef]
- Babalola, O.O.; Alawiye, T.T.; Lopez, C.R.; Ayangbenro, A.S. Shotgun metagenomic sequencing data of sunflower rhizosphere microbial community in South Africa. Data in Brief 2020, 31. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Babalola, O.O. Metagenomics methods for the study of plant-associated microbial communities: A review. J Microbiol Methods 2020, 170, 105860. [Google Scholar] [CrossRef] [PubMed]
- Regalado, J.; Lundberg, D.S.; Deusch, O.; Kersten, S.; Karasov, T.; Poersch, K.; Shirsekar, G.; Weigel, D. Combining whole-genome shotgun sequencing and rRNA gene amplicon analyses to improve detection of microbe–microbe interaction networks in plant leaves. ISME J. 2020, 14, 2116–2130. [Google Scholar] [CrossRef]
- Li, J.; Xie, S.; Wilson, G.W.; Cobb, A.B.; Tang, S.; Guo, L.; Wang, K.; Deng, B. Plant–microbial interactions facilitate grassland species coexistence at the community level. Oikos 2020, 129, 533–543. [Google Scholar] [CrossRef]
- Qi, G.; Ma, G.; Chen, S.; Lin, C.; Zhao, X. Microbial network and soil properties are changed in bacterial wilt-susceptible soil. Appl. Environ. Microbiol. 2019, 85, e00162–00119. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Muller, D.; Babalola, O.O. A metagenomic lens into endosphere microbial communities, promises, and discoveries. Lett. Appl. Microbiol. 2023, 76, ovac030. [Google Scholar]
- Khan, M.W.; Bohannan, B.J.; Nüsslein, K.; Tiedje, J.M.; Tringe, S.G.; Parlade, E.; Barberán, A.; Rodrigues, J.L. Deforestation impacts network co-occurrence patterns of microbial communities in Amazon soils. FEMS Microbiol. Ecol. 2019, 95, fiy230. [Google Scholar] [CrossRef]
- Wakung’oli, M.; Amoo, A.E.; Enagbonma, B.J.; Babalola, O.O. Termite societies promote the taxonomic and functional diversity of archaeal communities in mound soils. Biology 2020, 9, 136. [Google Scholar] [CrossRef]
- Simard, S.W. Mycorrhizal networks facilitate tree communication, learning, and memory. In: Memory and learning in plants. Springer; 2018, 191-213.
- Huang, X.-F.; Chaparro, J.M.; Reardon, K.F.; Zhang, R.; Shen, Q.; Vivanco, J.M. Rhizosphere interactions: Root exudates, microbes, and microbial communities. Botany 2014, 92, 267–275. [Google Scholar] [CrossRef]
- King, A.; Farrer, E.; Suding, K.; Schmidt, S. Co-occurrence patterns of plants and soil bacteria in the high-alpine subnival zone track environmental harshness. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Guo, L.; Bai, Y.; Liu, W.; Yan, J.; Bucher, M. Microbiomics and plant health: An interdisciplinary and international workshop on the plant microbiome. Mol. Plant 2019, 12, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.; Johnson, C.; Santos-Medellín, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc Nat Acad Sci 2015, 112, E911–E920. [Google Scholar] [CrossRef] [PubMed]
- Enebe, M.C.; Babalola, O.O. Effects of inorganic and organic treatments on the microbial community of maize rhizosphere by a shotgun metagenomics approach. Ann. Microbiol. 2020, 70, 1–10. [Google Scholar] [CrossRef]
- Gottel NR, Castro HF, Kerley M, Yang Z, Pelletier DA, Podar M, Karpinets T, Uberbacher E, Tuskan GA, Vilgalys R et al. Distinct microbial communities within the endosphere and rhizosphere of Populus deltoides roots across contrasting soil types. Appl. Environ. Microbiol. 2011, 77, 5934–5944. [Google Scholar] [CrossRef]
- López-Lozano, N.E.; Echeverría Molinar, A.; Ortiz Durán, E.A.; Hernández Rosales, M.; Souza, V. Bacterial diversity and interaction networks of Agave lechuguilla rhizosphere differ significantly from bulk soil in the oligotrophic basin of cuatro cienegas. Front. Plant Sci. 2020, 11, 1028–1028. [Google Scholar] [CrossRef]
- Babalola, O.O.; Fadiji, A.E.; Enagbonma, B.J.; Alori, E.T.; Ayilara, M.S.; Ayangbenro, A.S. The nexus between plant and plant microbiome: Revelation of the networking strategies. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Bukhat, S.; Imran, A.; Javaid, S.; Shahid, M.; Majeed, A.; Naqqash, T. Communication of plants with microbial world: Exploring the regulatory networks for PGPR mediated defense signaling. Microbiol Res 2020, 238, 126486. [Google Scholar] [CrossRef]
- Antar, M.; Gopal, P.; Msimbira, L.A.; Naamala, J.; Nazari, M.; Overbeek, W.; Backer, R.; Smith, D.L. Inter-organismal signaling in the rhizosphere. In: Rhizosphere Biology: Interactions Between Microbes and Plants. Edited by Gupta VVSR, Sharma AK: Springer Singapore; 2021, 255-293.
- Castro, R.O.; Bucio, J.L. Small molecules involved in transkingdom communication between plants and rhizobacteria. In: Molecular Microbial Ecology of the Rhizosphere. Edited by Bruijn FJd, vol. 1, John Wiley & Sons, Inc.; 2013, 295-307.
- Drogue, B.; Combes-Meynet, E.; Moënne-Loccoz, Y.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Control of the cooperation between plant growth-promoting rhizobacteria and crops by rhizosphere signals. In: Molecular Microbial Ecology of the Rhizosphere. Edited by Bruijn FJd, vol. 1: John Wiley & Sons, Inc.; 2013, 279-293.
- Atkinson, S.; Williams, P. Quorum sensing and social networking in the microbial world. J. R. Soc. Interface 2009, 6, 959–978. [Google Scholar] [CrossRef]
- Phour, M.; Sehrawat, A.; Sindhu, S.S.; Glick, B.R. Interkingdom signaling in plant-rhizomicrobiome interactions for sustainable agriculture. Microbiol Res 2020, 241, 126589. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Reverdy, A.; She, Q.; Sun, B.; Chai, Y. The role of rhizodeposits in shaping rhizomicrobiome. Env. Micro Rep. 2020, 12, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Vives-Peris, V.; de Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep 2020, 39, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Korenblum, E.; Dong, Y.; Szymanski, J.; Panda, S.; Jozwiak, A.; Massalha, H.; Meir, S.; Rogachev, I.; Aharoni, A. Rhizosphere microbiome mediates systemic root metabolite exudation by root-to-root signaling. Proc Nat Acad Sci 2020, 117, 3874–3883. [Google Scholar] [CrossRef]
- Dekkers, L.C.; Van Der Bij, A.J.; Mulders, I.H.M.; Phoelich, C.C.; Wentwoord, R.A.R.; Glandorf, D.C.M.; Wijffelman, C.A.; Lugtenberg, B.J.J. Role of the O-antigen of lipopolysaccharide, and possible roles of growth rate and of NADH:ubiquinone oxidoreductase (nuo) in competitive tomato toot-tip colonization by Pseudomonas fluorescens WCS365. Mol Plant-Microbe Interact 1998, 11, 763–771. [Google Scholar] [CrossRef]
- Burdman, S.; Dulguerova, G.; Okon, Y.; Jurkevitch, E. Purification of the major outer membrane protein of Azospirillum brasilense, its affinity to plant roots, and its involvement in cell aggregation. Mol. Plant-Microbe Interact. ® 2001, 14, 555–561. [Google Scholar] [CrossRef]
- Ryu, C.-M. Promoting plant protection by root-associated microbes. Plant Pathol J 2013, 29, 123–124. [Google Scholar]
- Caddell, D.F.; Deng, S.; Coleman-Derr, D. Role of the Plant Root Microbiome in Abiotic Stress Tolerance. In: Seed Endophytes: Biology and Biotechnology. Edited by Verma SK, White JJF. Cham: Springer International Publishing; 2019, 273-311.
- Stringlis IA, Yu K, Feussner K, de Jonge R, Van Bentum S, Van Verk MC, Berendsen RL, Bakker PA, Feussner I, Pieterse CM. MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health. Proc Nat Acad Sci 2018, 115, E5213–E5222. [Google Scholar]
- Hu J, Wei Z, Friman V-P, Gu S-h, Wang X-f, Eisenhauer N, Yang T-j, Ma J, Shen Q-r, Xu Y-c et al. Probiotic diversity enhances rhizosphere microbiome function and plant disease suppression. Am. Soc. Microbiol. 2016, 7, e01790–e01716. [Google Scholar]
- Joo, H.-S.; Deyrup, S.T.; Shim, S.H.J.P.R. Endophyte-produced antimicrobials: A review of potential lead compounds with a focus on quorum-sensing disruptors. 2020, 1-26.
- Ortiz-Castro, R. Martinez-Trujillo, M. Lopez-Bucio J. N-acyl-L-homoserine lactones: A class of bacterial quorum-sensing signals alter post-embryonic root development in Arabidopsis thaliana. Plant Cell Env. 2008, 31, 1497–1509. [Google Scholar] [CrossRef]
- Heil, M.; Bostock, R.M. Induced systemic resistance (ISR) against pathogens in the context of induced plant defences. Ann Bot 2002, 89, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Junker, R.R.; Tholl, D. Volatile organic compound mediated interactions at the plant-microbe interface. J Chem Ecol 2013, 39, 810–825. [Google Scholar] [CrossRef] [PubMed]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Wei, H.-X.; Paré, P.W.; Kloepper, J.W. Bacterial volatiles promote growth in Arabidopsis. Biol Control 2003, 100, 4927–4932. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, J.; Li, C.; Ma, Y. Antifungal and plant growth promotion activity of volatile organic compounds produced by Bacillus amyloliquefaciens. MicrobiologyOpen 2019, 8, e00813. [Google Scholar] [CrossRef]
- Bukhat, S.; Imran, A.; Javaid, S.; Shahid, M.; Majeed, A.; Naqqash, T. Communication of plants with microbial world: Exploring the regulatory networks for PGPR mediated defense signaling. Microbiological Research 2020, 126486. [Google Scholar] [CrossRef]
- Kawasaki, A.; Donn, S.; Ryan, P.R.; Mathesius, U.; Devilla, R.; Jones, A.; Watt, M. Microbiome and exudates of the root and rhizosphere of Brachypodium distachyon, a model for wheat. PLoS ONE 2016, 11, e0164533. [Google Scholar] [CrossRef]
- Korenblum, E.; Dong, Y.; Szymanski, J.; Panda, S.; Jozwiak, A.; Massalha, H.; Meir, S.; Rogachev, I.; Aharoni, A. Rhizosphere microbiome mediates systemic root metabolite exudation by root-to-root signaling. ProcNat Acad Sci 2020, 117, 3874–3883. [Google Scholar] [CrossRef]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant–microbe interactions. Antonie Van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Jangu, O.; Sindhu, S. Differential response of inoculation with indole acetic acid producing Pseudomonas sp. in green gram (Vigna radiata L.) and black gram (Vigna mungo L.). Microbiol. J. 2011, 1, 159–173. [Google Scholar] [CrossRef]
- Park J-M, Radhakrishnan R, Kang S-M, Lee I-J. IAA producing Enterobacter sp. I-3 as a potent bio-herbicide candidate for weed control: A special reference with lettuce growth inhibition. Indian J Microbiol 2015, 55, 207–212. [Google Scholar] [CrossRef]
- Amara, U.; Khalid, R.; Hayat, R. Soil bacteria and phytohormones for sustainable crop production. In: Bacterial Metabolites in Sustainable Agroecosystem. Edited by D. M, vol. 12. Cham: Springer International Publishing; 2015, 87-103.
- Gao, X.; Zhang, Y.; He, Z.; Fu, X. Gibberellins. In: Hormone Metabolism and Signaling in Plants. Edited by Li J, Li C, Smith SM: Academic Press; 2017, 107-160.
- Enagbonma, B.J.; Babalola, O.O. Potentials of termite mound soil bacteria in ecosystem engineering for sustainable agriculture. Ann Microbiol 2019, 69, 211–219. [Google Scholar] [CrossRef]
- Liu, E.; Yan, C.; Mei, X.; He, W.; Bing, S.H.; Ding, L.; Liu, Q.; Liu, S.; Fan, T. Long-term effect of chemical fertilizer, straw, and manure on soil chemical and biological properties in northwest China. Geoderma 2010, 158, 173–180. [Google Scholar] [CrossRef]
- Enagbonma, B.J.; Babalola, O.O. Environmental sustainability: A review of termite mound soil material and its bacteria. Sustainability 2019, 11, 3847. [Google Scholar] [CrossRef]
- Adedeji, A.A.; Häggblom, M.M.; Babalola, O.O. Sustainable agriculture in Africa: Plant growth-promoting rhizobacteria (PGPR) to the rescue. Sci. Afr. 2020, 9, e00492. [Google Scholar] [CrossRef]
- Araújo, W.L.; Lacava, P.T.; Andreote, F.D.; Azevedo, J.L. Interaction between endophytes and plant host: Biotechnological aspects. Plant-Microbe Interact. 2008, 1, 1–21. [Google Scholar]
- Olanrewaju OS, Babalola OO. Bacterial consortium for improved maize (Zea mays L.) production. Microorganisms 2019, 7, 519. [Google Scholar] [CrossRef]
- Rho, H.; Hsieh, M.; Kandel, S.L.; Cantillo, J.; Doty, S.L.; Kim, S.-H. Do Endophytes Promote Growth of Host Plants Under Stress? A Meta-Analysis on Plant Stress Mitigation by Endophytes. Microb. Ecol. 2018, 75, 407–418. [Google Scholar] [CrossRef]
- Xu, L.; Coleman-Derr, D. Causes and consequences of a conserved bacterial root microbiome response to drought stress. Curr. Opin. Microbiol. 2019, 49, 1–6. [Google Scholar] [CrossRef]
- Ribeiro, C.M.; Cardoso, E.J.B.N. Isolation, selection and characterization of root-associated growth promoting bacteria in Brazil Pine (Araucaria angustifolia). Microbiol. Res. 2012, 167, 69–78. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Yadav, A.N.; Yadav, N.; Kumar, V.; Kumar, A.; Sayyed, R.; Hesham, A.E.-L.; Dhaliwal, H.S.; Saxena, A.K. Drought-tolerant phosphorus-solubilizing microbes: Biodiversity and biotechnological applications for alleviation of drought stress in plants. In: Plant growth promoting rhizobacteria for sustainable stress management. Springer; 2019, 255-308.
- Kour, D.; Rana, K.L.; Kaur, T.; Sheikh, I.; Yadav, A.N.; Kumar, V.; Dhaliwal, H.S.; Saxena, A.K. Microbe-mediated alleviation of drought stress and acquisition of phosphorus in great millet (Sorghum bicolour L.) by drought-adaptive and phosphorus-solubilizing microbes. Biocatal. Agric. Biotechnol. 2020, 23, 101501. [Google Scholar] [CrossRef]
- Yuwono, T.; Handayani, D.; Soedarsono, J. The role of osmotolerant rhizobacteria in rice growth under different drought conditions. Aust. J Agric Res 2005, 56, 715–721. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Collados, C.; Barea, J.M.; Azcón, R. Arbuscular mycorrhizal symbiosis can alleviate drought-induced nodule senescence in soybean plants. 2001, 151, 493–502. 2001, 151, 493–502. [Google Scholar]
- Qin, Y.; Druzhinina, I.S.; Pan, X.; Yuan, Z. Microbially Mediated Plant Salt Tolerance and Microbiome-based Solutions for Saline Agriculture. Biotechnol. Adv. 2016, 34, 1245–1259. [Google Scholar] [CrossRef] [PubMed]
- del Carmen Orozco-Mosqueda M, Glick BR, Santoyo G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt stress in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef]
- Etesami, H.; Glick, B.R. Halotolerant plant growth–promoting bacteria: Prospects for alleviating salinity stress in plants. Environmental Experimental Botany 2020, 104124. [Google Scholar] [CrossRef]
- Bano, A.; Fatima, M. Salt tolerance in Zea mays (L). following inoculation with Rhizobium and Pseudomonas. Biol. Fertil. Soils 2009, 45, 405–413. [Google Scholar] [CrossRef]
- Barka, E.A.; Nowak, J.; Clément, C. Enhancement of chilling resistance of inoculated grapevine plantlets with a plant growth-promoting rhizobacterium, Burkholderia phytofirmans strain PsJN. Appl. Environ. Microbiol. 2006, 72, 7246–7252. [Google Scholar] [CrossRef]
- Raheem A, Shaposhnikov A, Belimov AA, Dodd IC, Ali B. Auxin production by rhizobacteria was associated with improved yield of wheat (Triticum aestivum L.) under drought stress. Arc Agron Soil Sci 2018, 64, 574–587. [Google Scholar] [CrossRef]
- Kearl, J.; McNary, C.; Lowman, J.S.; Mei, C.; Aanderud, Z.T.; Smith, S.T.; West, J.; Colton, E.; Hamson, M.; Nielsen, B.L. Salt-tolerant halophyte rhizosphere bacteria stimulate growth of alfalfa in salty soil. Frontier in Microbiology 2019, 10, 1849. [Google Scholar] [CrossRef]
- Jiao, J.; Ma, Y.; Chen, S.; Liu, C.; Song, Y.; Qin, Y.; Yuan, C.; Liu, Y. Melatonin-producing endophytic bacteria from grapevine roots promote the abiotic stress-induced production of endogenous melatonin in their hosts. Front Plant Sci 2016, 7, 1387. [Google Scholar] [CrossRef]
- Yamaji, K.; Watanabe, Y.; Masuya, H.; Shigeto, A.; Yui, H.; Haruma, T. Root fungal endophytes enhance heavy-metal stress tolerance of clethra barbinervis growing naturally at mining sites via growth enhancement, promotion of nutrient uptake and decrease of heavy-metal concentration. PLoS ONE 2016, 11, e0169089. [Google Scholar] [CrossRef]
- Ikram M, Ali N, Jan G, Jan FG, Rahman IU, Iqbal A, Hamayun M. IAA producing fungal endophyte Penicillium roqueforti Thom., enhances stress tolerance and nutrients uptake in wheat plants grown on heavy metal contaminated soils. PLoS ONE 2018, 13, e0208150. [Google Scholar]
- Ali, A.H.; Abdelrahman, M.; Radwan, U.; El-Zayat, S.; El-Sayed, M.A. Effect of Thermomyces fungal endophyte isolated from extreme hot desert-adapted plant on heat stress tolerance of cucumber. Appl. Soil Ecol. 2018, 124, 155–162. [Google Scholar] [CrossRef]
- Burygin, G.L.; Kargapolova, K.Y.; Kryuchkova, Y.V.; Avdeeva, E.S.; Gogoleva, N.E.; Ponomaryova, T.S.; Tkachenko, O.V.J.W.J.o.M. Biotechnology: Ochrobactrum cytisi IPA7. 2 promotes growth of potato microplants and is resistant to abiotic stress. 2019, 35, 55. [Google Scholar]
- Kim A-Y, Shahzad R, Kang S-M, Seo C-W, Park Y-G, Park H-J, Lee I-J. biotechnology: IAA-producing Klebsiella variicola AY13 reprograms soybean growth during flooding stress. J Crop Sci 2017, 20, 235–242. [Google Scholar]
- Jog, R.; Pandya, M.; Nareshkumar, G.; Rajkumar, S. Mechanism of phosphate solubilization and antifungal activity of Streptomyces spp. isolated from wheat roots and rhizosphere and their application in improving plant growth. Microbiology 2014, 160, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Otieno N, Lally RD, Kiwanuka S, Lloyd A, Ryan D, Germaine KJ, Dowling DN. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar]
- Li G, Kronzucker HJ, Shi W. The response of the root apex in plant adaptation to iron heterogeneity in soil. Front. Plant Sci. 2016, 7, 344. [Google Scholar]
- Gaiero JR, McCall CA, Thompson KA, Day NJ, Best AS, Dunfield KE. Inside the root microbiome: Bacterial root endophytes and plant growth promotion. Am. J. Bot. 2013, 100, 1738–1750. [Google Scholar] [CrossRef]
- Behie SW, Padilla-Guerrero IE, Bidochka MJ. Nutrient transfer to plants by phylogenetically diverse fungi suggests convergent evolutionary strategies in rhizospheric symbionts. Commun. Integr. Biol. 2013, 6, e22321. [Google Scholar] [CrossRef]
- Fadiji AE, Babalola OO. Elucidating mechanisms of endophytes used in plant protection and other bioactivities with multifunctional prospects. Front. Bioeng. Biotechnol. 2020, 8, 467. [Google Scholar] [CrossRef] [PubMed]
- Rascovan N, Carbonetto B, Perrig D, Díaz M, Canciani W, Abalo M, Alloati J, González-Anta G, Vazquez MP. Integrated analysis of root microbiomes of soybean and wheat from agricultural fields. Sci. Rep. 2016, 6, 28084. [Google Scholar] [CrossRef] [PubMed]
- Suman A, Yadav AN, Verma P. Endophytic microbes in crops: Diversity and beneficial impact for sustainable agriculture. In: Microbial inoculants in sustainable agricultural productivity, New Delhi. Springer pp 117-143; 2016, 117-143.
- Yadav, A.; Rana, K.; Kumar, V.; Dhaliwal, H. Phosphorus solubilizing endophytic microbes: Potential application for sustainable agriculture. EU Voice 2016, 2, 21–22. [Google Scholar]
- Yadav, A.N.; Verma, P.; Kour, D.; Rana, K.L.; Kumar, V.; Singh, B.; Chauahan, V.S.; Sugitha, T.; Saxena, A.K.; Dhaliwal, H.S. Plant microbiomes and its beneficial multifunctional plant growth promoting attributes. Int. J. Environ. Sci. Nat. Resour. 2017, 3, 1–8. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, M.; Verma, S.; Choudhary, P.; Chakdar, H. Plant Microbiome: Trends and Prospects for Sustainable Agriculture. In Plant Microbe Symbiosis; Varma, A., Tripathi, S., Prasad, R., Eds.; Springer: Cham, 2002; pp. 129–151. [Google Scholar] [CrossRef]
- Sengupta, S.; Ganguli, S.; Singh, P.K. Metagenome analysis of the root endophytic microbial community of Indian rice (O. sativa L.). Genom. Data 2017, 12, 41–43. [Google Scholar] [CrossRef]
- Ji SH, Gururani MA, Chun S-C. Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiol. Res. 2014, 169, 83–98. [Google Scholar] [CrossRef]
- Gyaneshwar P, Hirsch AM, Moulin L, Chen W-M, Elliott GN, Bontemps C, Estrada-de los Santos P, Gross E, dos Reis Jr FB, Sprent JI. Legume-nodulating betaproteobacteria: Diversity, host range, and future prospects. Mol. Plant-Microbe Interact. 2011, 24, 1276–1288. [Google Scholar] [CrossRef]
- Oldroyd GE, Murray JD, Poole PS, Downie JA. The rules of engagement in the legume-rhizobial symbiosis. Annu. Rev. Genet. 2011, 45, 119–144. [Google Scholar] [CrossRef]
- da Silva Fonseca E, Peixoto RS, Rosado AS, de Carvalho Balieiro F, Tiedje JM, da Costa Rachid CTC. The microbiome of Eucalyptus roots under different management conditions and its potential for biological nitrogen fixation. Microb. Ecol. 2018, 75, 183–191. [Google Scholar] [CrossRef]
- Verma P, Yadav AN, Khannam KS, Panjiar N, Kumar S, Saxena AK, Suman A. Assessment of genetic diversity and plant growth promoting attributes of psychrotolerant bacteria allied with wheat (Triticum aestivum) from the northern hills zone of India. Ann. Microbiol. 2015, 65, 1885–1899. [Google Scholar] [CrossRef]
- Singh D, Geat N, Rajawat MVS, Mahajan MM, Prasanna R, Singh S, Kaushik R, Singh RN, Kumar K, Saxena AK. Deciphering the mechanisms of endophyte-mediated biofortification of Fe and Zn in wheat. J. Plant Growth Regul. 2018, 37, 174–182. [Google Scholar] [CrossRef]
- Oliveira Ad, Urquiaga S, Döbereiner J, Baldani J. The effect of inoculating endophytic N2-fixing bacteria on micropropagated sugarcane plants. Plant Soil 2002, 242, 205–215. [Google Scholar] [CrossRef]
- Subramanian P, Kim K, Krishnamoorthy R, Sundaram S, Sa T. Endophytic bacteria improve nodule function and plant nitrogen in soybean on co-inoculation with Bradyrhizobium japonicum MN110. Plant Growth Regul. 2015, 76, 327–332. [Google Scholar] [CrossRef]
- Zhao L, Xu Y, Lai X. Antagonistic endophytic bacteria associated with nodules of soybean (Glycine max L.) and plant growth-promoting properties. Braz. J. Microbiol. 2018, 49, 269–278. [Google Scholar] [CrossRef]
- Perin L, Martínez-Aguilar L, Paredes-Valdez G, Baldani J, Estrada-De Los Santos P, Reis V, Caballero-Mellado J. Burkholderia silvatlantica sp. nov., a diazotrophic bacterium associated with sugar cane and maize. Int. J. Syst. Evol. Microbiol. 2006, 56, 1931–1937.
- Ramesh A, Sharma SK, Sharma MP, Yadav N, Joshi OP. Inoculation of zinc solubilizing Bacillus aryabhattai strains for improved growth, mobilization and biofortification of zinc in soybean and wheat cultivated in Vertisols of central India. Appl. Soil Ecol. 2014, 73, 87–96. [Google Scholar] [CrossRef]
- Puri A, Padda KP, Chanway CP. Nitrogen-fixation by endophytic bacteria in agricultural crops: Recent advances. Nitrogen in agriculture IntechOpen, London, GBR 2018, 73–94. [Google Scholar]
- Reis V, Estrada-De Los Santos P, Tenorio-Salgado S, Vogel J, Stoffels M, Guyon S, Mavingui P, Baldani V, Schmid M, Baldani J. Burkholderia tropica sp. nov., a novel nitrogen-fixing, plant-associated bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 2155–2162.
- Mbai F, Magiri E, Matiru V, Nganga J, Nyambati V. Isolation and characterisation of bacterial root endophytes with potential to enhance plant growth from kenyan basmati rice. Am. Int. J. Contemp. Res. 2013, 3, 25–40. [Google Scholar]
- Govindarajan M, Balandreau J, Kwon S-W, Weon H-Y, Lakshminarasimhan C. Effects of the inoculation of Burkholderia vietnamensis and related endophytic diazotrophic bacteria on grain yield of rice. Microb. Ecol. 2008, 55, 21–37. [Google Scholar] [CrossRef]
- Kumar P, Dubey R, Maheshwari D, Bajpai V. ACC deaminase producing Rhizobium leguminosarum rpn5 isolated from root nodules of Phaseolus vulgaris L. Bangladesh J. Bot. 2016, 45, 477–484. [Google Scholar]
- Joseph B, Ranjan Patra R, Lawrence R. Characterization of plant growth promoting rhizobacteria associated with chickpea (Cicer arietinum L.). Int. J. Plant Prod. 2012, 1, 141–152. [Google Scholar]
- Singh D, Rajawat MVS, Kaushik R, Prasanna R, Saxena AK. Beneficial role of endophytes in biofortification of Zn in wheat genotypes varying in nutrient use efficiency grown in soils sufficient and deficient in Zn. Plant Soil 2017, 416, 107–116. [Google Scholar] [CrossRef]
- Puri A, Padda KP, Chanway CP. Can a diazotrophic endophyte originally isolated from lodgepole pine colonize an agricultural crop (corn) and promote its growth? Soil Biol. Biochem. 2015, 89, 210–216. [Google Scholar] [CrossRef]
- Sandhya V, Shrivastava M, Ali SZ, Prasad VSSK. Endophytes from maize with plant growth promotion and biocontrol activity under drought stress. Russ. Agric. Sci. 2017, 43, 22–34. [Google Scholar] [CrossRef]
- Tanvir KLRDK, Yadav KRDAN, Anil NYHSD, Saxena K. Endophytic microbes: Biodiversity, plant growth-promoting mechanisms and potential applications for agricultural sustainability. 2020, 113, 1075–1107.
- Fritz, M.; Jakobsen, I.; Lyngkjær, M.F.; Thordal-Christensen, H.; Pons-Kühnemann, J. Arbuscular mycorrhiza reduces susceptibility of tomato to Alternaria solani. Mycorrhiza 2006, 16, 413–419. [Google Scholar] [CrossRef]
- Ortega RA, Mahnert A, Berg C, Müller H, Berg G. The plant is crucial: Specific composition and function of the phyllosphere microbiome of indoor ornamentals. FEMS Microbiology Ecology 2016, 92, fiw173. [Google Scholar] [CrossRef]
- Bordiec S, Paquis S, Lacroix H, Dhondt S, Ait Barka E, Kauffmann S, Jeandet P, Mazeyrat-Gourbeyre F, Clément C, Baillieul F. Comparative analysis of defence responses induced by the endophytic plant growth-promoting rhizobacterium Burkholderia phytofirmans strain PsJN and the non-host bacterium Pseudomonas syringae pv. pisi in grapevine cell suspensions. J. Exp. Bot. 2011, 62, 595–603. [Google Scholar] [CrossRef]
- Higginbotham SJ, Arnold AE, Ibañez A, Spadafora C, Coley PD, Kursar TA. Bioactivity of fungal endophytes as a function of endophyte taxonomy and the taxonomy and distribution of their host plants. PLoS ONE 2013, 8, e73192. [Google Scholar]
- Ding L, Maier A, Fiebig H-H, Lin W-H, Hertweck C. A family of multicyclic indolosesquiterpenes from a bacterial endophyte. Org. Biomol. Chem. 2011, 9, 4029–4031. [Google Scholar] [CrossRef] [PubMed]
- Ezra D, Castillo UF, Strobel GA, Hess WM, Porter H, Jensen JB, Condron MA, Teplow DB, Sears J, Maranta M. Coronamycins, peptide antibiotics produced by a verticillate Streptomyces sp.(MSU-2110) endophytic on Monstera sp. Microbiology 2004, 150, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Mendes R, Kruijt M, De Bruijn I, Dekkers E, van der Voort M, Schneider JH, Piceno YM, DeSantis TZ, Andersen GL, Bakker PA. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Klein E, Ofek M, Katan J, Minz D, Gamliel A. Soil suppressiveness to Fusarium disease: Shifts in root microbiome associated with reduction of pathogen root colonization. Phytopathology 2013, 103, 23–33. [Google Scholar] [CrossRef]
- Asghari, S.; Harighi, B.; Mozafari, A.A.; Esmaeel, Q.; Barka, E.A. Screening of endophytic bacteria isolated from domesticated and wild growing grapevines as potential biological control agents against crown gall disease. BioControl 2019, 64, 723–735. [Google Scholar] [CrossRef]
- Dalal J, Kulkarni N, Bodhankar M. Antagonistic and plant growth promoting potentials of indigenous endophytic fungi of soybean (Glycine max (L) Merril). Indian J. Adv. Plant Res. 2014, 1, 9–16. [Google Scholar]
- Kumar, V.; Jain, L.; Jain, S.K.; Chaturvedi, S.; Kaushal, P. Bacterial endophytes of rice (Oryza sativa L.) and their potential for plant growth promotion and antagonistic activities. South African Journal of Botany 2020. [Google Scholar] [CrossRef]
- Shehata H, Lyons E, Jordan K, Raizada M. Bacterial endophytes from wild and ancient maize are able to suppress the fungal pathogen Sclerotinia homoeocarpa. J. Appl. Microbiol. 2016, 120, 756–769. [Google Scholar] [CrossRef]
- Ardebili ZO, Ardebili NO, Mahdi Hamdi SM. Physiological effects of'Pseudomonas fluorescens' CHA0 on tomato ('Lycopersicon esculentum'Mill.) plants and its possible impact on Fusarium oxysporum f. sp.'Lycopersici'. Aust. J. Crop Sci. 2011, 5, 1631. [Google Scholar]
- Falahian F, Ardebili ZO, Fahimi F, Khavarinejad R. Effect of mycorrhizal fungi on some defense enzymes against Gaeumannomyces gaminis in wheat. Pak. J. Biol. Sci. 2007, 10, 2418–2422. [Google Scholar] [CrossRef]
- Lee, J.; Seo, M.; Kim, H. Isolation and characterization of an antagonistic endophytic bacterium Bacillus velezensis CB3 the control of citrus green mold pathogen Penicillium digitatum. Korean J. Mycol. 2012, 40, 118–123. [Google Scholar] [CrossRef]
- Omomowo IO, Fadiji AE, Omomowo OI. Assessment of bio-efficacy of Glomus versiforme and Trichoderma harzianum in inhibiting powdery mildew disease and enhancing the growth of cowpea. Ann. Agric. Sci. 2018, 63, 9–17. [Google Scholar] [CrossRef]
- Yang F, Zhang R, Wu X, Xu T, Ahmad S, Zhang X, Zhao J, Liu YJMP. An endophytic strain of the genus Bacillus isolated from the seeds of maize (Zea mays L.) has antagonistic activity against maize pathogenic strains. Microb. Pathog. 2020, 142, 104074. [Google Scholar] [CrossRef] [PubMed]
- Irabor A, Mmbaga M. Evaluation of Selected Bacterial Endophytes for Biocontrol Potential against Phytophthora Blight of Bell Pepper (Capsicum annuum L.) J. Plant Pathol. Microbiol. 2017, 8, 31–34.
- Kushwaha, P.; Kashyap, P.L.; Srivastava, A.K.; Tiwari, R.K. Plant growth promoting and antifungal activity in endophytic Bacillus strains from pearl millet (Pennisetum glaucum). Braz. J. Microbiol. 2020, 51, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Sha, Y.; Wang, Q.; Li, Y. Suppression of Magnaporthe oryzae and interaction between Bacillus subtilis and rice plants in the control of rice blast. SpringerPlus 2016, 5, 1238. [Google Scholar] [CrossRef]
- 128. Nourozian J, Etebarian HR, Khodakaramian G. Biological control of Fusarium graminearum on wheat by antagonistic bacteria. Songklanakarin Journal of Science and Technology 2006, 28 Suppl. 1, 29–38.
- Wang, S.; Hu, T.; Jiao, Y.; Wei, J.; Cao, K. Isolation and characterization of Bacillus subtilis EB-28, an endophytic bacterium strain displaying biocontrol activity against Botrytis cinerea Pers. Front. Agric. China 2009, 3, 247–252. [Google Scholar] [CrossRef]
- Fiorilli V, Vannini C, Ortolani F, Garcia-Seco D, Chiapello M, Novero M, Domingo G, Terzi V, Morcia C, Bagnaresi P. Omics approaches revealed how arbuscular mycorrhizal symbiosis enhances yield and resistance to leaf pathogen in wheat. Sci. Rep. 2018, 8, 1–18. [Google Scholar]
- Mustafa, G.; Khong, N.G.; Tisserant, B.; Randoux, B.; Fontaine, J.; Magnin-Robert, M.; Reignault, P.; Sahraoui, A.L.-H. Defence mechanisms associated with mycorrhiza-induced resistance in wheat against powdery mildew. Funct. Plant Biol. 2017, 44, 443–454. [Google Scholar] [CrossRef]
- Behn, O. Influence of Pseudomonas fluorescens and arbuscular mycorrhiza on the growth, yield, quality and resistance of wheat infected with Gaeumannomyces graminis. J. Plant Dis. Prot. 2008, 115, 4–8. [Google Scholar] [CrossRef]
Figure 1.
mechanisms employed by plant root-associated microbes in improving plant health.
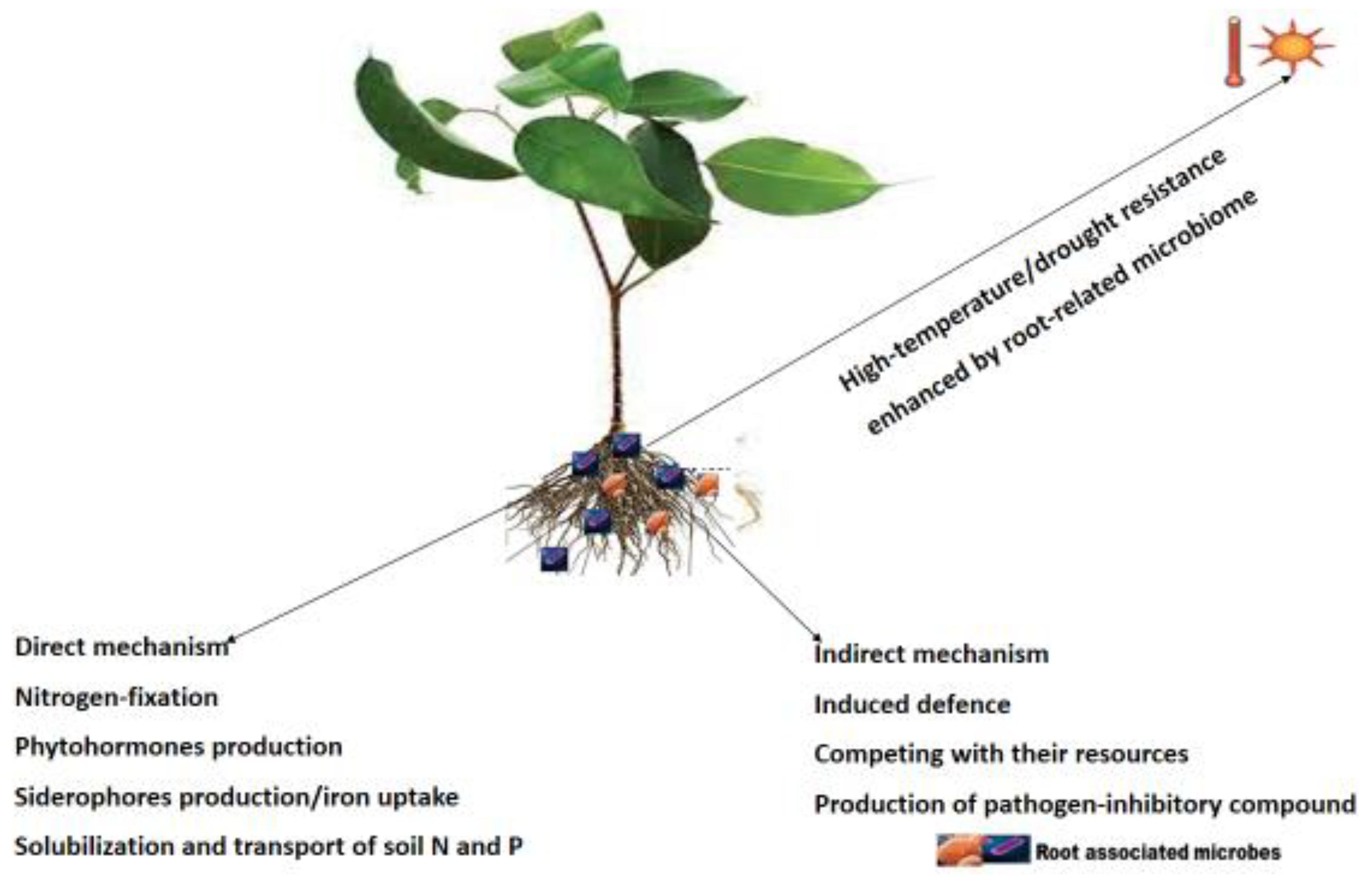
Figure 2.
Plant root-root associated microbe communication.
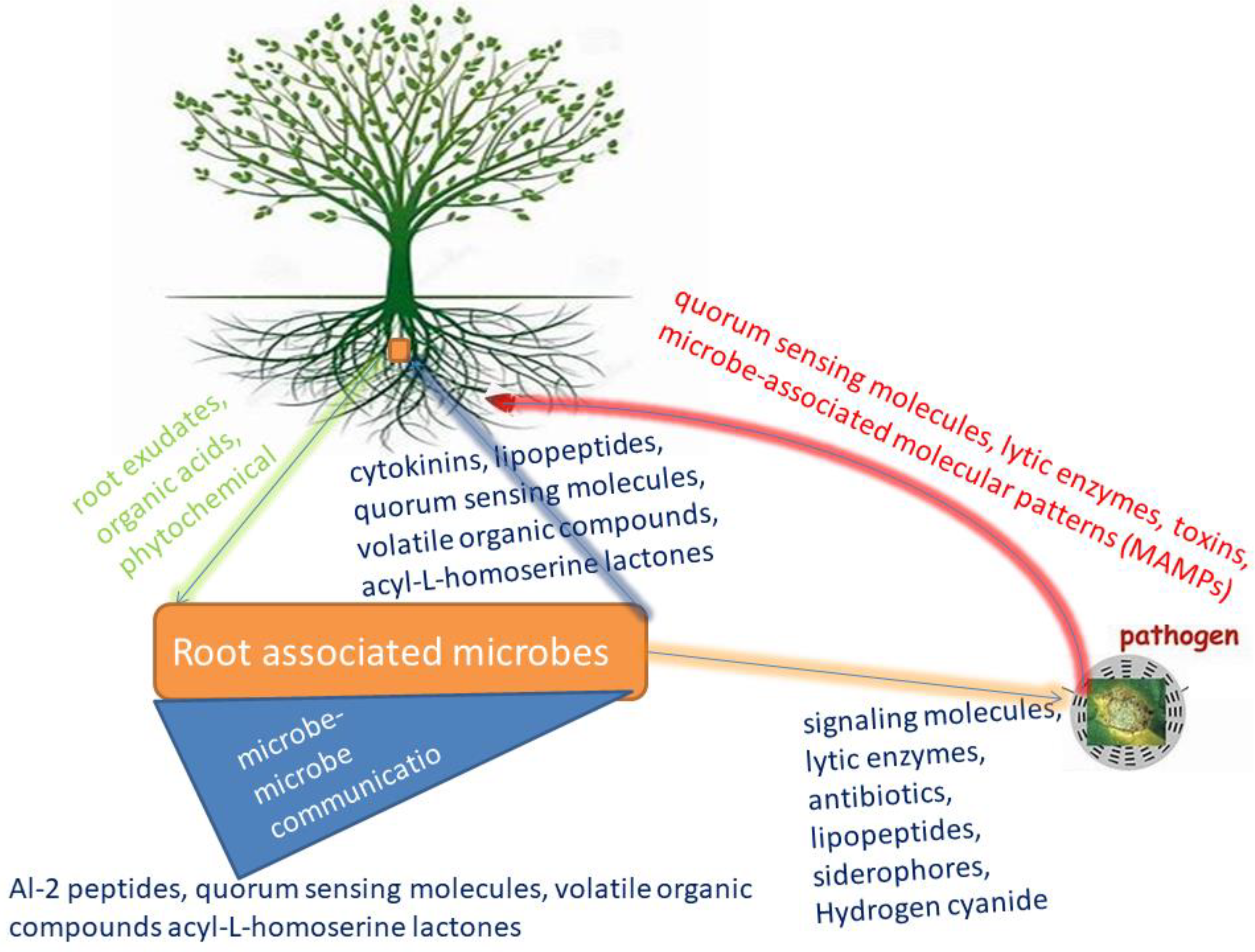
Table 2.
Nutrient acquisition attributes of notable plant root microbiomes.
| Root Microbiomes | Host plant | Phosphorus (P) | Potassium (K) | Nitrogen fixers (N2F) | Siderophore (Sid) | Zinc (Zn) | References |
|---|---|---|---|---|---|---|---|
| B. amyloliquefacien | Rice | + | + | + | + | + | [91] |
| A. sulfonivorans | Wheat | - | - | - | + | + | [92] |
| A. amazonense | Sugarcane | - | - | + | - | - | [93] |
| B. megaterium | Soybean | + | - | + | + | - | [94] |
| P. agglomerans | Rice | + | - | + | - | - | [91] |
| P. putida | Soybean | - | - | + | + | - | [95] |
| B. silvatlantica | Sugarcane | - | - | + | - | - | [96] |
| B. aryabhattai | Soybean | - | - | - | - | + | [97] |
| K. pneumoniae | Rice | - | - | - | + | - | [98] |
| B. tropica | Sugarcane | - | - | + | - | - | [99] |
| P. putida | Rice | + | - | - | - | - | [100] |
| P. dispersa | Wheat | - | - | - | + | + | [91] |
| B. vietnamiensis | Rice | - | - | + | - | - | [101] |
| R. leguminosarum | Beans | + | - | - | + | + | [102] |
| B. licheniformis | Chickpea | + | - | - | - | - | [103] |
| B. subtilis | Soybean | - | - | + | + | - | [104] |
| P. polymyxa | Maize | - | - | + | - | - | [105] |
| P. thivervalensis | Maize | - | - | - | + | - | [106] |
| E. asburiae | Maize | - | - | - | + | - | [106] |
| R. endophyticum | Beans | + | - | - | - | - | [107] |
| R. irregularis | Tomato | + | - | - | - | - | [108] |
+ =Active, - = Inactive.
Table 3.
Biocontrol activities of some plant root microbiomes.
| Root Microbiomes | Host plant | Pathogens active against | Activities and metabolites secreted/Induced | References |
|---|---|---|---|---|
| Pseudomonas sp., Pantoea sp. | Grapevine | A. tumefaciens, A. vitis | - | [116] |
| A. calcoaceticus | Soybean | P. sojae 01 | Siderophore and indole acetic acid | [95] |
| Bacillus sp. | Soybean |
C. truncatum, R. solani, F oxysporum, S. rolfsii, A. alternata, and M. phaseolina |
Siderophore and Hydrogen cyanide. |
[117] |
| B. subtilis | Rice |
R. solani, F. verticelloides, and S. rolfsii |
Lipopeptides | [118] |
| B. gladioli 3A12 | Maize | S. homoeocarpa | - | [119] |
| P. fluorescens 63-28 | Pea |
P. ultimum and F. oxysporum f. sp. pisi |
Induced peroxidase, polyphenoloxisae, Superoxide dismutase and phenylalanine amonialyase. | [120] |
| P. aeruginosa FTR | Maize |
F. oxysporium, P. aphanidermatum, Alternaria sp., R solani, M. phaseolina, Alternaria sp. and S. rolfii, |
- | [106] |
| Glomus etunicatum | Wheat | G. graminis | Isozyme | [121] |
| B. velezensis CB3 | Citrus | P. digitatum | - | [122] |
| G. versiforme and T harzianum | Cowpea | E. flexuosa | - | [123] |
| B. velezensis | Maize |
T. funiculosus, P. oxalicum, and F. verticillioides |
Lipopeptide | [124] |
| R. leguminosarum RPN5 | Beans |
M. phaseolina, F. oxysporum, S. sclerotiorum and F. solani. |
- | [102] |
| Serratia (B17B), Enterobacter (E), and Bacillus (IMC8, Y, Ps, Psl, and Prt) | Papaya and Bean | P. capsici | - | [125] |
| Acremonium sp., Leptosphaeria sp., T. flavus, and P. simplicissimum. | Cotton | V. dahliae strain Vd080 | - | [107] |
| Bacillus sp. | Millet |
R. solani, S. rolfsii, and F. solani |
Antimicrobial peptides | [126] |
| B. subtilis | Rice | M. oryzae | Enhanced activity of peroxidase, polyphenol oxidase and superoxide dismutase | [127] |
| Pseudomonas sp. | Wheat | F. graminearum | - | [128] |
| B. subtilis EB-28 | Tomato | B. cinerea | - | [129] |
| F. mosseae | Wheat | X. translucens | - | [130] |
| R. irregularis | Tomato | A. solani | - | [108] |
| F. mosseae | Wheat | B. graminis | - | [131] |
| F. mosseae and P. fluorescens | Wheat | G. graminis | - | [132] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



