Submitted:
06 July 2023
Posted:
07 July 2023
You are already at the latest version
Abstract
The purpose of this study was to elucidate patterns and mechanisms driving seasonal and inter-annual variations of carbon and nitrogen stable isotopes in the zooplankton crustacean commu-nity of Lake Maggiore (Italy), during the period 2009-2020. Different zooplankton taxa and groups revealed different ranges of d13C signatures, giving an insight into food sources and niche partition. In particular, cyclopoids had a restricted range with most negative d13C‰ values and increase in d13C fractionation with the establishment of water thermal vertical stratification, high-lighting the importance of vertical distribution as key factor for taxa coexistence in a vertically heterogenous environment. d13C of the zooplankton community and of Daphnia were positively related to water temperature (R2=0.58 p <0.0001 and R2=0.68 p <0.0001, respectively) and d13C Daphnia signature to chlorophyll-a (R2=0.32, p < 0.0001). Decomposition of time series data for zooplankton carbon and nitrogen signatures and environmental parameters identified increasing trends in water temperature, chlorophyll-a and water conductivity and decrease in nitrate, that matched changes in carbon isotopic signature trend in some zooplankton taxa (Bosmina, Daphnia and diaptomids). Overall, the observed patterns in zooplankton isotopic signatures were interpreted as integrations of climate warming effects in Lake Maggiore, affecting both food sources availability and environmental conditions.
Keywords:
freshwater zooplankton
; stable isotope analysis
; lake
; climate change
; food web
1. Introduction
This study represents a detailed seasonal analysis, along a decadal period (2009-2020), of freshwater zooplankton δ13C and δ15N signatures, in relation to patterns of environmental variables in Lake Maggiore (Italy). Because freshwater zooplankton represents a crucial link within lake food webs, their study can provide important information for understanding the impact of changes of environmental and climatic factors on pelagic food webs.
Lake ecosystems are very sensitive to climatic changes and are characterised by strong seasonal variability, determined by physical constraints in the temperature and light regime, controlling water column mixing, nutrient cycling and ultimately primary and secondary production within pelagic waters [1]. Lakes are also subject to long term changes determined by global climatic forcing, external inputs from catchments and by internal dynamics [2,3,4,5,6]. Effects of global climatic changes on terrestrial and aquatic ecosystems are increasingly evident during the last two decades [7,8,9].
Worldwide climatic change has the potential to affect many fundamental properties of lakes, with the increase in temperature modifying numerous chemical and physical parameters of aquatic environments and ultimately their biotic communities [10,11,12]. Great efforts are required to understand the effects on aquatic communities, especially on population structure (e.g. biodiversity, density, biomass; [13,14], ecosystem functionality (trophic relationships, redundancy, resilience; [15,16,17,18], and on variation in the trophic position of organisms. The ecological role of an organism in an aquatic ecosystem is subject to seasonal fluctuations, depending on ecological variables such as variations of the population structure, food sources availability and chemical and physical environmental variables [19,20,21,22]. It’s becoming clear that each taxon isn’t a fixed entity within an ecosystem, but its ecological "role" and its relative relationships with other taxa vary over time. Consequently, trophic webs are not rigid schemes of organisms trophic positions, but should be considered as networks of dynamic interactions among different organisms.
The use of carbon and nitrogen stable isotope analysis (SIA) represents a useful way to identify dynamic interactions among aquatic organisms in trophic networks, since it makes it possible to determine their relative trophic positions and variations over time. Stable isotope analysis has been increasingly used in ecological studies since it allows a functional approach to the investigation of trophic webs and ecosystem functioning [21,22,23,24,25]. Providing an integrated measure over time of C and N signatures in the different trophic levels, stable isotope analysis allows the tracing of the path of environmental variability through the trophic web. Analysing stable isotopes over plurennial timeframes along with environmental variables provides an insight into paths and mechanisms through which global changes, such as warming, are processed through the food web.
In this study, carbon and nitrogen isotope signatures (δ13C‰ and δ15N‰) of different zooplankton species (Daphnia sp., Bosmina, Diaphanosoma brachyurum, Bythotrephes longimanus, Leptodora kindtii) and groups (diaptomids, cyclopoids) were examined along with environmental variables during twelve annual production cycles (2009-2020) in Lake Maggiore, Italy. We aimed at detecting seasonal and inter-annual variability, and determining factors that might affect long-term trend evolution of zooplankton δ13C and δ15N. To our knowledge, there are no other studies on carbon and nitrogen isotopes of freshwater zooplankton communities for such a long time period. Through the use of isotopic signatures, we also aimed to trace habitat preferences, food sources and niche partition of zooplankton taxa.
Zooplankton is crucial for the transfer of matter and energy through the pelagic food web. The signature of δ13C in zooplankton primary consumers reflect changes in their diet, therefore following seasonal baseline fluctuations, which mainly depend on POM in winter and early spring and on phytoplankton in late spring and summer [26,27,28]. A detailed seasonal study of plankton δ13C and related variables in a dynamic lake system can provide important information for understanding the impact of environmental factors on the pelagic food web.
To date, most studies on the relationships between δ13C variability and environmental variables are incomplete and have primarily focused on marine systems [29,30,31,32,33,34,35]. [36] carried on an analysis of literature data revealing that carbon stable isotopes of POM were related to lake latitudinal location, water temperature and CO2 concentration, with no relationship with trophic state indices. [37] analysing δ13C and δ15N of zooplankton in eighteen reservoirs found correlations with Secchi depth, water temperature, nutrients and chlorophyll-a.
Lake Maggiore is a deep oligomictic lake belonging to the subalpine Lake District in Northern Italy. It is an ideal system to explore the relationships between carbon and nitrogen stable isotopic zooplankton signatures and environmental variables because its limnology has been studied over the long term. Lake Maggiore recovered from eutrophication in the 1970s and 1980s through the reduction of external phosphorus loads and reached oligotrophic status by the end of the 1990s [38,39]. Since about 2008 total phosphorous (TP) concentration in the epilimnion has remained stable, with low concentrations of 6 µg L–1. Long term studies have revealed a trend of decreasing concentrations in nitrate (N-NO3) and total nitrogen (TN) in the lake since 2009, when the depletion of N-NO3 in the epilimnion started to be particularly evident especially in the summer period, with concentrations below 500 µg L–1.
Chlorophyll-a concentration and phytoplankton biomass confirmed the oligotrophication process of the lake from the mid-1980s until about 2005, with annual mean chlorophyll-a concentration declining from 4.5-5.5 µg L–1 to 2.0- 3.0 µg L–1. However, since 2007, strong fluctuations of chlorophyll-a values and annual mean values around 4.0 µg L–1 started to reoccur. Analysis of the phytoplankton community has shown marked interannual changes in biovolume in the last decade, with increases in some years (2011 and 2016) due to algal blooms during the summer months [40,41].
Recent studies have highlighted how, after reaching a stable trophic status, climatic drivers became more prominent in Lake Maggiore in shaping the dynamics of phytoplankton species fluctuations and the pelagic food web as a whole [5,12,42,43]. Long term studies show how water temperature has increased significantly in Lake Maggiore [5,44,45]. The annual average values of the Schmidt stability increased in time (MKT p<0.001), confirming the aggrading resistance of the water column to mixing. The mixing depth and mixed volume of water at spring overturn in Lake Maggiore have been steadily declining in recent years due to the increasing thermal stability of the water column, an effect of climate change on deep lakes, and it is likely to continue in the near future [5,44,45].
From the evolution of the lake parameters described, it is understandable that during the last decade climate change represented a main driving factor of the lake dynamics, affecting lake hydrodynamics, nutrient status and biological dynamics [4,12]. Although nutrient status reached oligotrophy, climatic changes affected lake thermal dynamics, increasing water column stability, diminishing water mixing depth and thus affecting the primary production of the lake.
We therefore aimed at exploring the potential effects of (climatic) and environmental variables that could have driven inter-annual differences and long-term trends in stable isotopic zooplankton signatures. Among the environmental variables, we focused special attention on water temperature, which is expected to influence zooplankton isotopic signature [22,31] as it controls the solubility of CO2 and in turn influences the fractionation of carbon isotopes during phytoplankton uptake [30]. We also explored water column thermal stratification, known to influence cell activity, assimilation and competition for nutrients, light competition, the rate of CO2 and diffusion and ultimately phytoplankton species succession, abundance and community structure. As water thermal stratification has such a strong effect on the phytoplankton δ13C [46] it follows that it should also influence zooplankton isotopic signature. We explored also chlorophyll-a, found to increase in the most recent years in Lake Maggiore due to a “eutrophication like” effect of climate warming [47,48], as primary productivity and CO2 concentration are known to affect carbon isotopic signature. We investigated also changes in nitrogen concentration, as nutrient cycling, particularly of nitrogen, affects isotope ratios [49].
There are two main objectives of this paper:
1) to examine the seasonal and inter-annual variability of the isotopic signals of δ13C and δ15N in the different zooplankton groups and taxa
2) to investigate the long-term trends of zooplankton isotopic signals of δ13C and δ15N and their relationships with environmental variables
2. Materials and Methods
Zooplankton sampling was performed in Lake Maggiore at Ghiffa station (45°58’30’’N; 8°39’09’’E; Figure 1), where the lake reaches its maximum depth (370 m). Samples were collected using a 58-cm diameter zooplankton net (450-µm mesh size) hauled vertically in the water layer 0-50m several times, in order to obtain a sufficient amount of organisms needed to perform the analyses (min. = 2, max = 15). The total volume filtered was at least 26 m3 per sample. The large net mesh sizes facilitated high filtration efficiency, excluding clogging by large phytoplankton colonies, while ensuring collection of the most important taxa representative of the lake net zooplankton (i.e. the Cladocerans Daphnia longispina-galeata gr., Bosmina, Diaphanosoma brachyurum, Leptodora kindtii, Bythotrephes longimanus, as well as adults and last developmental stages of Diaptomids and Cyclopoids). Samples were collected seasonally (January, May, August, November) in duplicate from 2009 to 2020. In vivo specimens from one sample were concentrated in laboratory into ca. 1L of lake water, frozen at -20°C and subsequently used for carbon (13C) and nitrogen (15N) Stable Isotope Analysis (SIA). The other sample was fixed and preserved in 96% ethanol, for subsequent microscopic analysis and counting at 6.3X. SIA was performed on pooled zooplankton samples of the size fraction ≥450 µm and on the taxa/groups present. Under the dissecting microscope we pooled individuals of each taxon in order to reach a minimum dry weight (DW) of ca. 2 mg per sample (ca. 70-700 individuals depending on individual dry weight). Samples were oven-dried for 24 h at 60 °C, before homogenizing and transferal into tin capsules of 5 x 9 mm in size. The isotopic composition of organic carbon and nitrogen was determined by Ján Veizer Stable Isotope Laboratory (Ottawa University, Ontario, Canada) following the method already described in [50].
Many authors, including [51] suggested using the isotopic composition of Daphnia sp. as a pelagic baseline in lakes, as Daphnia sp. feeding behavior appeared to be fairly consistent and non-selective, both at different seasons and in different lakes. Because crustacean taxa might exhibit differing feeding behaviors and preferences [52], their isotope signatures were examined separately in this study. Potential modifications in their food sources and in their relative trophic positions during the survey were investigated by comparing their stable isotopic compositions to that of Daphnia sp.
The first assumption is that if a taxon relies solely on the pelagic baseline, then its δ13C value is similar to Daphnia’s, with a residual variance lower or equal to that expected for the δ13C enrichment per trophic level (S.D.= 1.3‰; [53] Post, 2002), as suggested by [51]. Otherwise, there might be multiple carbon sources fueling the zooplankton community. According to [54], a common carbon source is attributed when fractionation: F= δ13Cpredator - δ13Cprey ≤ 0.8 ‰ (±1.1 ‰ S.D.). Therefore, if individuals are specialised on one food type only, they will display isotopic values from this food type (adjusted for trophic fractionation), while individuals that feed on and incorporate many different food items with different isotopic values will have a mean isotopic value which reflects the different isotopic contributions in their diet (including fractionation).
In case of comparing zooplankton taxa with Daphnia as the pelagic baseline, fractionation is F= δ13Ctaxon - δ13CDaphnia.
Water temperature was measured at discrete depths with mercury-filled thermometers until 2010 [55]; later on, thermal profiles were recorded with a multi-parameter probe (Idronaut CTD 304); the two set of measures were used to calibrate the two series of measurements. Chemical analyses of lake samples were run at the water chemistry laboratory of the CNR Water Research Institute (CNR IRSA, previously CNR ISE) in Verbania Pallanza, using standard methods for freshwater samples [56,57]. Details on the analytical methods and the QA/QC procedures adopted in the laboratory can be found at http://www.idrolab.irsa.cnr.it/. Integrated samples for chlorophyll-a analysis were collected through the 0-20 m layer [42]. Chlorophyll-a was measured by spectrophotometric technique in the period 1984-2009 [57], while from 2010 measurements were taken using a vertical profiling instrument (FluoroProbe, BBE Moldaenke) after a careful check of the comparability of the two methods.
3. Results
The three filter feeder cladocerans Daphnia longispina-galeata gr., Bosmina and Diaphanosoma brachyurum had comparable values of δ15N‰ (mean values of 5.9 δ15N‰, 4.8 δ15N‰ and 4.2 δ15N‰, respectively) (Figure 1a). On some occasions, Bosmina δ15N‰ was very low, with values approaching zero. The other zooplankton components were placed at higher δ15N‰ values. The two predatory cladocerans (Bythotrephes longimanus and Leptodora kindtii) had similar mean and median values, however Bythotrephes had a wider range than Leptodora (range of δ15N‰=17.3 against a range of δ15N‰=8.3; median value of 8.6 δ15N‰ and 7.2 δ15N‰, respectively). The smallest range of variation for nitrogen isotopic signatures was found for Cyclopoids (range of δ15N‰=5.2), which had similarly high/enriched values as Diaptomids (median was 9.1 δ15N‰ for Diaptomids and 9.8 δ15N‰ for Cyclopoids).
The smallest ranges of δ13C (Figure 1b) were found for Diaptomids and Cyclopoids, both characterized by more negative values (mean of -34.2 δ13C‰ for Diaptomids and mean of -33.8 δ13C‰ for Cyclopoids) than the other zooplankton taxa. Diaphanosoma had the least negative δ13C signature (mean of -27.5 δ13C‰, median of -26.7 δ13C‰), likely due to its limited presence during the year, preferring warm months, and its tendency to occupy superficial waters. Daphnia and Bosmina had similar δ13C ranges and mean values (-31.8 δ13C‰ and -31.9 δ13C‰, respectively, Figure 1b).
The seasonality of carbon and nitrogen isotopic signatures of pooled zooplankton samples (≥450 µm), detected over the decadal time scale, is shown in Figure 2. Winter signatures are positioned in the top-left of the graph, and are opposed to summer signatures, which lie in the lower right. Spring signature values share with winter signals the left side of the graph but are characterized by lower δ15N‰ values. Summer signature values had similar δ15N‰ values as spring but differed substantially for the least depleted carbon signatures (δ13C‰ between -29 and -22 for summer and δ13C‰ -37 to -32 for spring). Autumn values, lying in the central part of the graph, have the largest between-year variability (δ13C‰ between -35 and -25), and tend to extend into the spring and summer areas.
The large variation in autumnal δ13C signature results from less 13C-depleted signatures of Diaphanosoma, Bosmina and the Diaptomids in October 2009 and of Bythotrephes and Leptodora in November 2010. Warm temperatures characterized the two above-mentioned months and water temperature stratification was still present in autumn 2009 and 2010, with warmer temperatures around 16°C down to 15 meters depth.
The four areas characterizing decadal seasonality of isotopic signatures are better separated when only Daphnia is considered (Figure 3). Between-year variability is still larger for autumn than for the other seasons, for both carbon and nitrogen isotopic signatures (δ13C‰ between -35 and -28; δ15N‰ between 3 and 10). The close correspondence between the isotopic values of pooled zooplankton and Daphnia reflects the major contribution of Daphnia to total zooplankton biomass throughout the year [60].
Carbon isotopic signatures of both pooled zooplankton and Daphnia were related to water temperature measured in the 0-50 meters layer δ13C pooled vs temperature 50m: R2=51% p ≤0.0001 n=42 and δ13C Daphnia vs temperature 50m: R2=61% p ≤0.0001 n=45), but this relationship improved when temperature in the 0-20 meters layer was used δ13C pooled vs temperature 20m: R2=58% p ≤0.0001 n=42 and δ13C Daphnia vs temperature 20m: R2=68.7% p ≤0.0001 n=45).
Warmer temperatures corresponded to less 13C-depleted zooplankton values, while colder temperatures were associated with most 13C-depleted signatures.
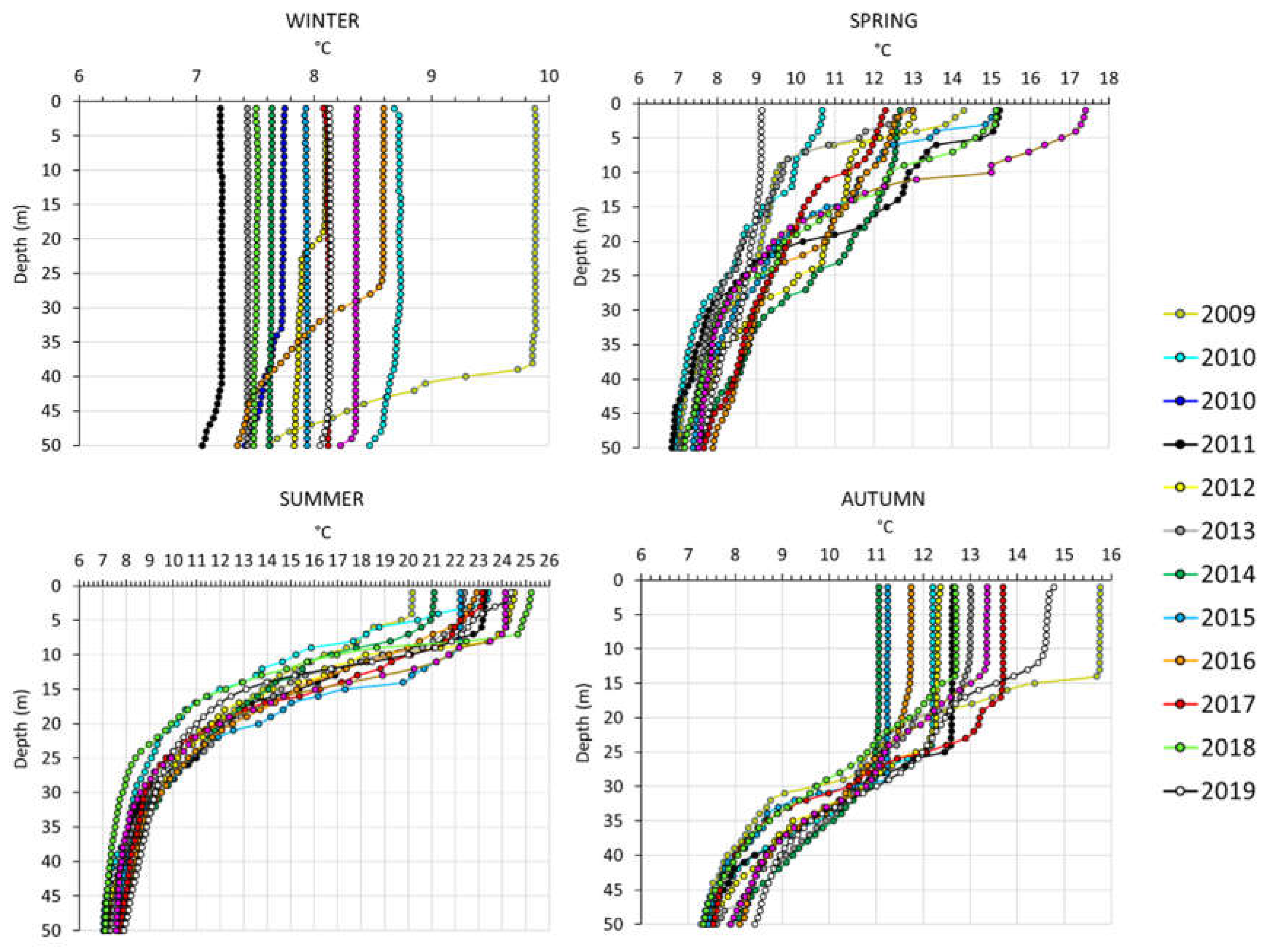
Between-year variability was large, and it reflected between year variations in temperature of the sampled layer, as well as differences among temperature vertical profiles. For example, in autumn relatively higher surface temperatures (15.8°C) down to 15 meters depth and stronger stratification (range temperature in the layer 0-50 m of 9 C°) recorded in 2009 and to a lesser extent in 2019 (Figure A4) corresponded to the least negative values of Daphnia δ13C (-28 to - 27 δ13C‰) in the graph of Figure 3. In contrast, lower autumn temperatures (11°C) and lighter thermal vertical stratification (range temperature in the layer 0-50 m of 3 to 4 C°) in 2014 and 2015 corresponded to the most negative autumn values (-34 δ13C‰) on the left side of the graph (Figure 3).
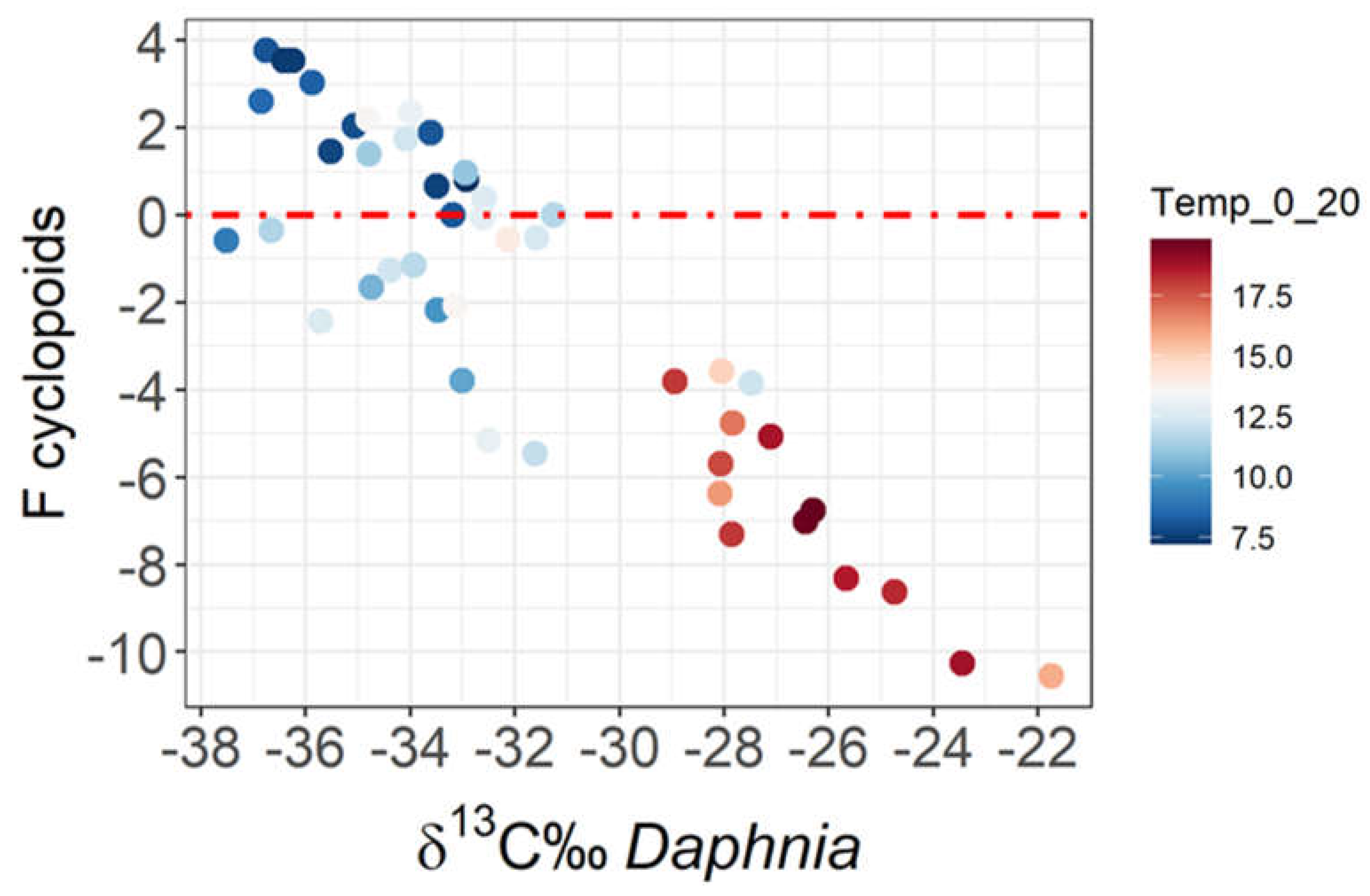
Cyclopoids shared the same carbon signature as Daphnia during the winter, having close to zero Fractionation (FCyclopoids = δ13C‰Cyclopoids - δ13C‰Daphnia; Figure 4). Fractionation gradually increased with warmer temperatures towards the summer, as also water thermal stratification proceeded.
The fractionation of cyclopoids increased substantially (from -4 to -10) at mean water temperatures ≥15 °C. The changes were quantitative and a regression between δ13CDaphnia and FCyclopoids was statistically significant (R2=78%, p<0.0001; n=44).
Decomposition of time series data [59] allowed the detection of seasonality and trends in the period of δ13C‰ and δ15N‰ data available (2010-2020) for the different zooplankton taxa and groups. The analyses were extended to the environmental parameters related to isotopic signatures.
The decomposition analysis for carbon isotopic signatures detected marked seasonality for the zooplankton taxa and groups considered. Variation of the carbon isotopic signature in the seasonally decomposed trends allowed the observation of patterns and changes over the decadal period.
Chlorophyll-a was related to pool zooplankton δ13C signature, indicating the influence of food availability (Figure A1). The linear relationship between chlorophyll-a concentrations and Daphnia δ13C signatures was statistically highly significant (R2 = 31.9%; p<0.0001, n = 41).
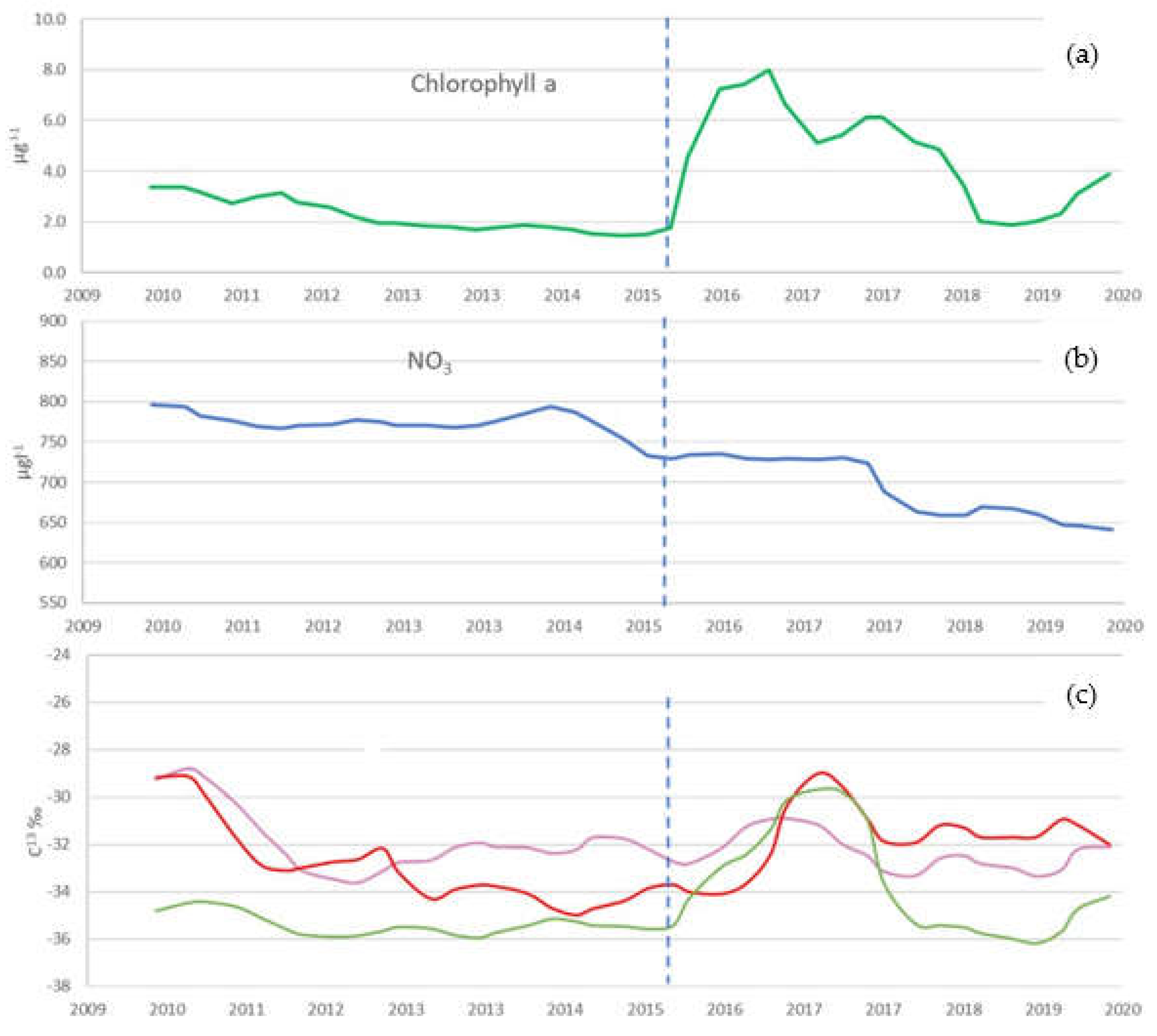
Seasonally decomposed trends clearly identified phases of changes in environmental variables (Figure A1); water temperature increased in the studied period, with a marked acceleration of the warming after 2015/2016. A gradual decreasing trend in nitrates (N-NO3) was detected up to 2015, followed by a sharper decline with values falling below 600 µg L-1 registered in the epilimnion during the summer periods. The trend in chlorophyll-a concentration initially showed a gradual decline until 2015, followed by a surge in concentration in 2016-2018, matching a phase of increase in the trend of water conductivity in the same period. Seasonally decomposed trends clearly identified phases of increased water temperature and a sharp acceleration of warming after 2015/2016 (Figure 1b and Figure A1).
The carbon signature trend of Daphnia varied little in the decadal period, with the exception of a decline from less negative values during the first two years (2010-2012) of the studied period (Figure 5c and Figure A2). The trend in carbon signature of Bosmina showed a gradual decrease from the beginning of the time period until 2015, followed by a steeper increase in 2016. The carbon signatures trend of diaptomids were quite stable at relatively depleted values (about -34‰) until 2016, when the signal started to increase towards less negative values (around -30‰ with extreme values up to -25‰) for a period of two years (2016-2018). Almost regular fluctuations in the carbon isotopic signature of the cyclopoids were observed, (Figure A2) with carbon signature varying in a relatively very small range, estimated between -38 and -32‰.
Figure 5.
Comparison of seasonally decomposed trends of chlorophyll-a (a) and nitrate (N-NO3) (b) with carbon isotopic signatures of selected zooplankton taxa ((c): red line= Bosmina; green line= diaptomids; purple line= Daphnia), during the period 2009-2020.arbon isotopic values vs. cyclopoids’ carbon fractionation (F).
Figure 5.
Comparison of seasonally decomposed trends of chlorophyll-a (a) and nitrate (N-NO3) (b) with carbon isotopic signatures of selected zooplankton taxa ((c): red line= Bosmina; green line= diaptomids; purple line= Daphnia), during the period 2009-2020.arbon isotopic values vs. cyclopoids’ carbon fractionation (F).
Our results for δ15N‰ Daphnia and δ15N‰ Bosmina time series decomposition (Figure A3) showed a common decreasing trend after 2014/2015, corresponding to the time of decrease in NO3 concentration in the epilimnion of the lake (while no decreasing or increasing trend in δ15N‰ was detected for the other taxa and groups).
4. Discussion
In lacustrine systems, δ13C% and δ15N% values of suspended particulate matter (SPM) vary over time [46,59]. This variability is related to external loadings, phytoplankton species composition and primary productivity, as well as sources and concentrations of dissolved inorganic C and N [62, 63). Such variability is transmitted along the food chain to higher trophic levels, like zooplankton organisms which are crucial in transferring matter and energy in pelagic food webs [51].
The results of this study showed that carbon and nitrogen zooplankton signatures are ruled by a strong seasonality, which is maintained across the twelve-year period considered. Tropho-chemical graphs (Figure 1 and Figure 2) show a similar pattern of seasonal partition when considering pooled zooplankton community and Daphnia. The close correspondence between the isotopic values of pooled zooplankton and Daphnia reflects the major contribution of Daphnia to total zooplankton biomass throughout the year. As proved in previous studies, the pooled zooplankton signature can be obtained from the sum of taxa specific isotope signatures weighted by taxa contribution to pooled zooplankton biomass obtained on the same date [60].
4.1. δ13 C and temperature
Carbon isotopic signature of both pooled zooplankton and Daphnia were significantly correlated with water temperature for both water layers of 0-50 m depth and 0-20 m depth. Warm temperatures corresponded to less 13C-depleted values, while colder temperatures were associated with the most 13C-depleted signatures. Temperature is an important determinant of seasonal changes in zooplankton δ13C‰ values [30,64,65]. Seasonal fluctuations of zooplankton δ13C‰ values generally reflect the phytoplankton 13C-fingerprint in lake ecosystems [63,66,67]. In monomictic lakes a positive correlation between δ13C‰ values and temperature is often observed [28,46,50,63].
Temperature controls the solubility of CO2 and in turn influences the fractionation of carbon isotopes during phytoplankton uptake [30,62,65]. Particularly in late spring and summer, the carbon baseline signature reflects the contribution by phytoplankton [27,28]. Changes in phytoplankton community structure and functional groups are also related to carbon isotopic signature [63], as temperature is also related to intrinsic changes of δ13C‰ values for zooplankters, due to the balance between carbon assimilation and respiration; experimental studies have reported variation of 13C-fingerprint in Daphnia reared at different temperatures regardless of the food sources [68,69].
In our study, values of δ13C‰ in zooplankters from -37 to -33.5 in winter corresponded to mean water temperatures (within the layer 0-50 m) below 8.5 °C; in summer δ13C‰ values ranged between -28 and -24 with mean water temperatures above 12.5°C.
4.2. Carbon isotopic signature in different taxa/groups as niche indicators
Carbon isotopic signature is related to habitat and food sources. Food selectivity also can play a role, particularly as phytoplankton size and shape seem to be important in the seasonality of carbon isotopic signature in general [63]. The relationship between temperature and carbon isotopic signature is indicative also of the vertical distribution of the zooplankton taxa.
Several studies have supported the concept that carbon isotopic signature is affected by water depth [46,50,61,70]. In deep lakes, the carbon isotopic signature is strongly influenced by depth, with organisms living in deeper layers characterized by more negative values than those living closer to the surface [69,72,73]. During the production year of a deep lake, SPM and zooplankton 13C-signature increases with water warming and the onset of thermal stratification [28,50,63].
In our study we observed differences in the 13C signature between Daphnia and adult Cyclopoids, likely due to the differences in food sources and habitat preferences. The changes were quantitative and the correlation between δ13C Daphnia and the delta fractionation of cyclopoids (Fcyclopoids) was statistically significant (Figure 4). Cyclopoids increasingly disengage from the water layer in which Daphnia feeds and lives when warming and vertical thermal stratification increase. However, the question remains whether food differentiation or habitat changes are more important in the resulting δ13C signal of the two zooplankton taxa during the warmer months characterised by thermal stratification. However, it seems evident that there is a distinct separation of the cyclopoids from the predatory cladocerans, which closely align with Daphnia’s isotopic carbon signature, likely inhabiting and moving within more superficial and warmer zones of the pelagic. Another study in a deep lake of the same Italian lake district [74] drew similar conclusions, with cyclopoids likely occupying a different and deeper habitat (>20 meters depth) than Daphnia. [75] conducted an experimental study in an oligotrophic lake in Japan, and found that the population of a cyclopoid copepods (Cyclops sp.) inhabited the hypolimnion during the summer stratification, while was scattered throughout the whole water column in the winter circulation period. Other studies [76,77] found spatial segregation during summer stratification, with daphnids resided above the thermocline whereas copepods at the lower edge of the thermocline.
Another explanation of the increasing variations between Daphnia and the Cyclopoids could result from their different life history strategies and metabolism. Some authors [78] have suggested that the δ13C‰ values of adult Cyclops respond more slowly to changes in the isotopic baseline because they have a lower carbon turnover rate compared with Daphnia that rapidly respond to changes in the δ13C‰ values of its phytoplankton food source. Such variation in the isotopic turnover between the herbivorous daphnids and predatory copepods, has been confirmed in experimental studies [72], and is attributed to distinct life history strategies [79].
The least negative δ13C signature of the filter feeding cladoceran Diaphanosoma brachyurum was likely due to its limited seasonal presence during the warmest months of the year (i.e. usually absent in winter and spring samples) and its tendency to occupy superficial water. This habitat preference of D. brachyurum has been demonstrated in previous studies in Lake Maggiore [80], with species biomass increasing during the establishment of thermal stratification in the warmer, upper ten meters of the lake.
Littoral carbon sources have less 13C-depleted signatures [50,81,82,83,84]. Seasonality has been demonstrated also in the littoral carbon signature, with the same pattern observed for the pelagic carbon signature, i.e. a transition from more (in winter) to less (in summer) negative δ13C‰ values. Previous studies on Lake Maggiore [60,63,83] demonstrated that ranges of isotopic signatures measured in the same season were quite distinct for littoral and pelagic habitat, allowing a correct interpretation of data when speculating on migration of zooplankters between different lake zones.
The two particle feeder cladocerans Daphnia and Bosmina shared the same δ13C‰ range and similar values, indicating they share of the same food sources and have similar mobility across lake habitats. The two taxa have, however, distinct feeding modes: Daphnia is a strict filter feeder, processing food particles through the filtering combs of the third and fourth thoracic limbs [85]. Whereas the foraging mode of Bosmina deviates markedly from filter feeding in a way that permits efficiency in the removal of highly edible flagellates, particularly when these items occur at low density [86]. In our twelve years’ study, on some occasions Bosmina’s δ13C-signature was less negative than Daphnia’s, likely reflecting a mobility through the water column of this small cladoceran and a tendency to feed in more superficial layers than Daphnia. The relatively low δ15N-signature of Bosmina with values approaching zero during the summer in few years, confirms the capability of this zooplankter to exploit nitrogen fixing cyanobacteria [74,83,86,87]. Cyanobacteria are usually lower in nitrogen isotopic signature than other phytoplankton groups, in particular nitrogen-fixing taxa [88,89,90], found in the phytoplankton community of Lake Maggiore [63]. The feeding mode of Bosmina avoids clogging during the growth of filamentous phytoplankton colonies, including those of cyanobacteria.
The range of variation of carbon isotopic signature (Figure 6a) was small for Cyclopoids and Diaptomids, indicating consistency of food sources throughout the studied period and that Cyclopoids and Diaptomids tend to be conservative in their habitat during the year. For Diaptomids, however, a more skewed distribution was observed toward less negative/depleted δ13C‰ values, suggesting an opportunistic feeding strategy with a discrete mobility across different zones or habitats of the lake. In particular, because less depleted values of δ13C‰ occurred during the summer periods, the migration of Diaptomids towards more superficial waters was considered [50].
Within the zooplankton community of Lake Maggiore, a narrower range of variation for nitrogen isotopic signatures (Figure A3) was found for Cyclopoids; which combined with their small range of carbon isotopic signatures suggests an overall niche specificity of this zooplankton group. Enrichment with respect to the pelagic baseline, however, cannot be calculated, being solidarity with carbon isotopic signature restricted to the unstratified thermal phase.
Figure 6.
Comparison of seasonally decomposed trends of chlorophyll-a (a) and nitrate (N-NO3) (b) with carbon isotopic signatures of selected zooplankton taxa ((c): red line= Bosmina; green line= diaptomids; purple line= Daphnia), during the period 2009-2020.arbon isotopic values vs. cyclopoids’ carbon fractionation (F).
Figure 6.
Comparison of seasonally decomposed trends of chlorophyll-a (a) and nitrate (N-NO3) (b) with carbon isotopic signatures of selected zooplankton taxa ((c): red line= Bosmina; green line= diaptomids; purple line= Daphnia), during the period 2009-2020.arbon isotopic values vs. cyclopoids’ carbon fractionation (F).
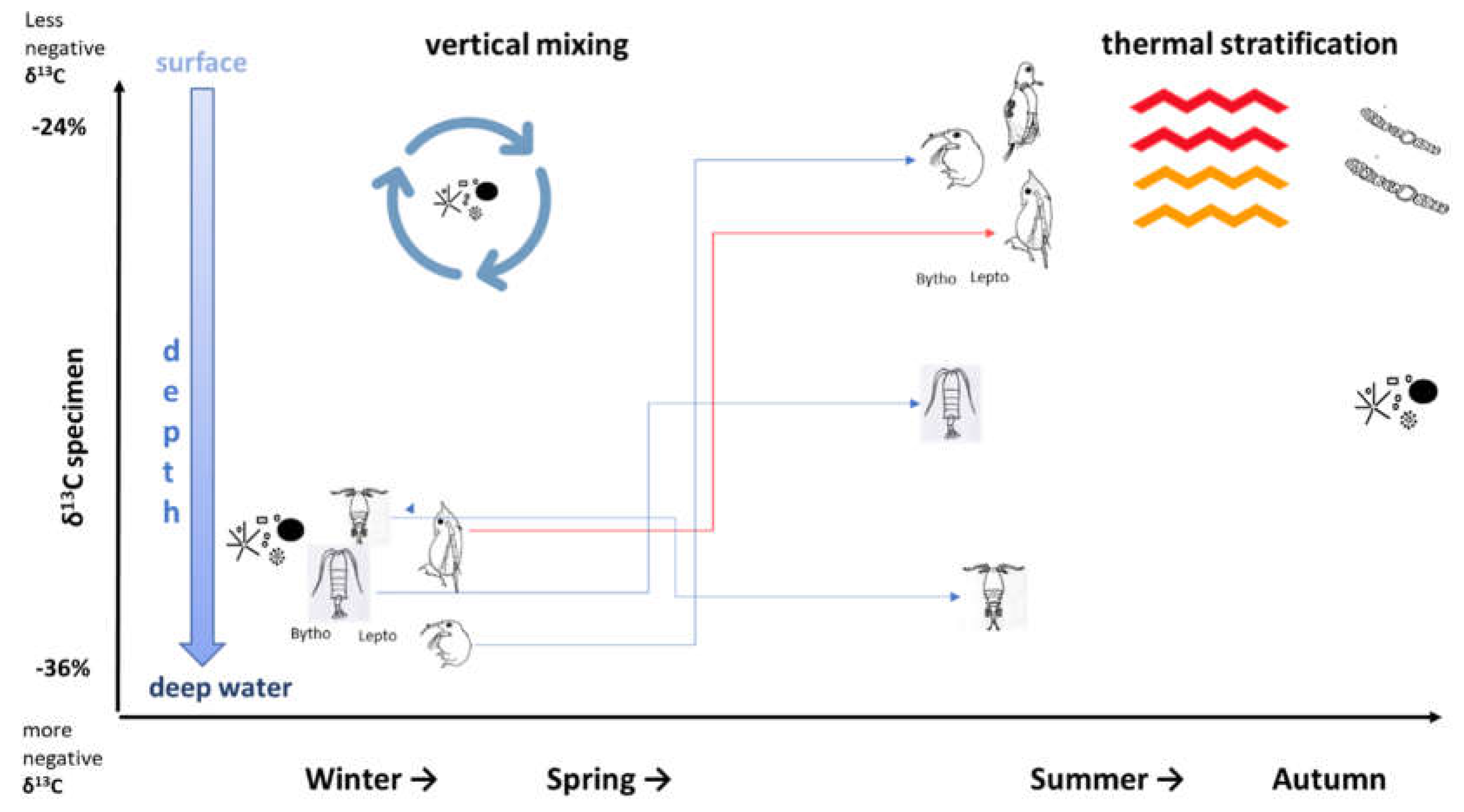
The conceptual diagram represents a summary of all the above-described variations in food sources, habitat preferences and seasonal variations of the zooplankton taxa considered in our study in Lake Maggiore. The diagram clearly illustrates how the relative δ13C‰ values of zooplankton taxa are collocated with respect to each other and to vertical lake depth in the different seasons. In particular, it illustrates the shift of δ13C‰ of different zooplankton taxa during two very different times of the year: late winter/springtime when water column circulation generally occurs in the lake, and summertime when stratification takes place (Figure A4).
In large deep lakes at median latitude, during late winter significant water mixing typically occurs, caused by a change in the air temperature and helped by windy conditions, thus increasing homogeneity in water conditions and environmental parameters along the water column (temperature, dissolved oxygen, pH, etc). Phytoplankton is consequently distributed in the water column and in Lake Maggiore the phytoplankton community is mainly represented by diatoms which require a high degree of turbulence to maintain in suspension [23] and having 13C-depleted values [46,91]. At this time of the year, we registered values of δ13C‰ for zooplankton taxa within in a narrow range, likely as a result of the homogeneity of phytoplankton type and of habitat.
With the advancing of warmer temperatures, water conditions change, establishing thermal stratification and creating a vertical gradient of the main environmental parameters. These conditions create heterogeneity in the lake water column, reflected also in more diverse plankton communities and vertical distribution of groups and biomass. Filamentous green algae, smaller diatoms, cyanobacteria, and other species capable of depth regulation are favored with intensified stratification [92]. In Lake Maggiore during the summer, cyanobacteria, green algae and diatoms are all relevant groups contributing to the phytoplankton community. A detailed study in Lake Maggiore [63] revealed a vertical gradient of phytoplankton density and biomass, in three different lake stations, during thermal stratification; recent phytoplankton monitoring with a fluorescent probe (FluoroProbe, BBE Moldaenke, GmbH, Germany) and with high frequency chlorophyll-a in situ sensors (Turner Design, USA) confirmed different chlorophyll-a concentrations and phytoplankton groups with water depth [personal comment, data unpublished). In this decadal study, δ13C‰ values for different zooplankton taxa in summer consistently underwent a clear separation from those of springtime when taxa δ13C‰ values were similar.
We hypnotize that it is likely because the conditions of thermal stratification during the summer create vertical heterogeneity in both food sources and habitats, allowing partial niche partitions for the different zooplankton taxa.
Although lakes are often considered as homogenous environments/habitats, this concept may be valid for the circulation period but not when vertical thermal stratification is established. Variation in vertical dimensions in temperature, light, dissolved oxygen and nutrients lead to vertical habitat diversification and creates various microhabitats, affecting the vertical distribution of zooplankton taxa with different food and temperature requirements and predation pressure [76,93,94,95,96,97,98,99] In our study (referring to our conceptual diagram) the stratification period corresponded to a phase of heterogeneity also for zooplankton taxa vertical distribution, as suggested by the distribution of isotopic signatures during the twelve year study period.
There is growing consensus that lakes, far from being homogeneous environments, offer a rich and dynamic heterogeneity at multiple spatial and temporal scales [100]. Natural and anthropogenic factors create spatial heterogeneity in lakes, leading to heterogeneity in environmental conditions and resource availability within-lake compartmentalization in food web structure [101].
4.3. Zooplankton (Daphnia) δ13C and chlorophyll-a: indication of food quality and availability
In our decadal study of Lake Maggiore, chlorophyll-a concentration was related to zooplankton δ13C signatures, indicating the importance of phytoplankton standing crop as food availability and utilization for zooplankton. Previous studies on Lake Maggiore reported significant correlations between chlorophyll-a and sestonic carbon concentration over the long term [102], suggesting that POM in the lake is mainly of autochthonous origin and likely made up of algal cells, with the seston δ13C-signature mainly related to phytoplankton. The correlation found in this study between zooplankton δ13C-signatures and chlorophyll-a concentration suggests that the phytoplankton biomass is important in determining zooplankton carbon signature, likely representing the main food source for zooplankton in Lake Maggiore. In other freshwater studies [62] it has been shown that zooplankton tend to utilize mainly algal derived autochthonous carbon during the more productive months. In addition, [65] found a correlation between the herbivorous zooplankter Daphnia and chlorophyll-a in their study of four annual production cycles in a subarctic lake.
Because chlorophyll-a variability is often a function of primary productivity in freshwaters [103,104], the δ13C seasonality can be interpreted as resulting from changing phytoplankton growth rates [65]. This finding is in good agreement with other field investigations [49,105,106,107] and culture experiments [106,108,109] which reveal a strong relationship between growth rate and δ13C‰ values of aquatic primary producers. Several models predict a rise in δ13C‰ values with increasing algal growth rate or carbon demand [31,108,109,110].
Plankton collected during periods of high biomass and primary productivity tends to exhibit the heaviest (i.e., less negative) δ13C-signatures, due to reduced isotopic fractionation at high cell densities or growth rates, or a switch to utilizing as CO2 concentration is depleted at high cell densities or high growth rates [111,112,113]. The relationships are also mediated by the difference in δ13C-signatures of the phytoplankton groups present in a certain season, since during the summer months Cyanobacteria tend to increase in Lake Maggiore [63] and this phytoplankton group is shown to possess relatively enriched δ13C values [114].
4.4. δ13C and δ15N time series and environmental variables
Decomposition of time series data allowed the detection of seasonality and trends of environmental parameters during the considered period (2010-2020). In particular, the seasonally decomposed trend of chlorophyll-a concentration increased, while the nitrate (N-NO3) trend decreased.
Recent long-term studies on Lake Maggiore [5] have shown that during the oligotrophication phase of the lake between 1988 and 2005, chlorophyll-a, and phytoplankton biovolume, significantly decreased down to low concentrations of 2.0-3.0 µg l-1. However, in the last decade despite nutrient concentrations remaining quite stable and low, chlorophyll-a concentration slightly increased, with fluctuations observed especially during summer periods. These data were recently confirmed by satellite observations, detecting since 2015 a change in chlorophyll-a seasonal pattern with less defined spring blooms and a tendency for annual maxima to occur in summer [115]. In this study, we detected those increases and fluctuations in chlorophyll-a especially between 2015 and 2020, matching an increasing trend of δ13C‰ values of zooplankton taxa such as Bosmina and Diaptomids. Explanation might be found in the new water conditions with increasing algal growth rates or carbon demand, leading to an increase in the δ13C-signature of phytoplankton as predicted by several models [31,108,109,110].
In our study, time series decomposition analysis also revealed that δ15N-signature of the cladocerans Daphnia and Bosmina had a decreasing trend during the last decade. A number of studies have demonstrated that the plankton δ15N-fingerprint is a good indicator of nitrogen sources, nitrogen cycling processes and consumer trophic position in aquatic systems [116]. Changes in phytoplankton species composition may influence the use of nitrogen sources (N2 fixation vs. uptake of dissolved combined nitrogen). Under nitrogen limitation, cyanobacterial N2 fixation is stimulated, resulting in depleted δ15N‰ values, which may then be transferred to zooplankton through feeding, resulting in a low zooplankton δ15N-signature [116]. Although daphnids cannot ingest the largest forms of cyanobacteria, they are capable of feeding on the smaller, decomposing pieces [118] and on the associated bacteria, so they will still reflect δ13C-signature of the baseline during cyanobacterial blooms [78]. The capability of Bosmina to exploit cyanobacteria has been already mentioned in this paper. Overall, the decreasing trend in δ15N-signature of both Daphnia and Bosmina during the last decade in our results match the conditions of nitrogen limitation and decreasing concentrations in nitrate and total nitrogen in the epilimnion of Lake Maggiore, as revealed by recent long-term studies [119].
Our twelve-year study of carbon and nitrogen stable isotopes revealed that zooplankton isotope signatures incorporate changes in lake environmental parameters, integrating the trends that have occurred during the last decade, thereby representing candidate sentinels capable of integrating the influence of climate change on the ecosystem of the lake.
Although the nutrient status in Lake Maggiore has reached oligotrophy, climatic change has affected lake thermal dynamics by increasing the water column stability, aggrading resistance of the water column to mixing and diminishing the water mixing depth [5], all factors which affect the primary production of the lake. The increase of fluctuations and of concentrations in chlorophyll-a experienced in Lake Maggiore during most recent times, could be the result of the recent changes in the lake which has modified conditions altering lake primary production.
Especially from work on shallow lakes, limnologists are realising that climate change is intensifying the symptoms of eutrophication in freshwaters [1,13], and results from direct experiments suggests that warming can increase the symptoms of eutrophication and changes in ecosystems structure. Our results, together with the most recent long-term studies on Lake Maggiore, could be interpreted as signs of eutrophication-like effects on a deep lake: because increased temperatures and water column stability promoted a prolonged period of water stratification and of epilimnion segregation during the warmest months, these new conditions could resemble those in the better studied shallow lakes, with similar final consequences in promoting lake primary production.
Figure 6 compares the seasonally decomposed trend of chlorophyll-a and nitrate with the carbon isotopic signature of selected zooplankton taxa. A clear-cut match is evident between a surge in chlorophyll-a concentration, the decrease in epilimnion nitrates since 2015 and the trend change in carbon isotopic signature of the considered zooplankters.
These conditions suggest that climate change represented the main driving factor of the lake dynamics during the last decade and that its effects have been integrated in the stable isotopes signatures of the zooplankton community, candidates to become sentinels of climatic changes in the lake.
Temperature as a stressor can impact freshwater ecosystems at lower trophic levels (e.g. primary producers) and can cause a cascading effect on other species through the food chain, which in turn affects ecosystem structure and function [120,121]. For aquatic ecosystems, the phytoplankton-zooplankton interaction determines the structure of food web, which represents the energy and materials flowing through the population in the system [122,123,124].
5. Conclusions
Tracing ecosystem functioning and the role of ecological variables under changing scenarios is a challenging goal, also in view of predicting the impact of climate change in freshwater environments. We used carbon and nitrogen SIA to identify the seasonal trophic positions of crustacean zooplankton taxa during twelve years in a deep subalpine lake. Along the seasons, water temperature together with the dynamics of thermal stratification and food availability were important factors determining the carbon isotopic signatures of different zooplankton taxa. Vertical heterogeneity of the environment, established during the summer with thermal stratification, promoted vertical spatial segregation of pelagic zooplankton taxa, suggesting that cyclopoid copepods occupy the deepest waters and cladocerans such as Diaphanosoma more superficial waters. Zooplankton stable isotopic signatures and their decadal temporal changes were related to temperature, increase of chlorophyll-a concentration and a decrease of N-NO3 in the epilimnion, observed during the last decade in Lake Maggiore. Such changes in environmental parameters occurred from direct and indirect effects of climate change on the lake ecosystem; the isotope composition of zooplankton taxa, reflecting their dietary assimilated carbon and nitrogen and their habitat choice, may therefore integrate these long-term environmental changes.
Our study, having considered the seasonal dynamics of carbon and nitrogen isotopes in the zooplankton community, with a detail of the main crustacean taxa and extended to a considerable period of twelve years, represents an important insight for interpreting similar investigations restricted to single years.
We overall further confirm that stable isotopic studies are a useful tool for tracing the ecological role of organisms in an aquatic ecosystem, subject to seasonal fluctuations and depending on ecological variables such as food sources availability and environmental conditions. Because zooplankton can integrate environmental variability in their isotopic signals, their analysis allows to detect patterns in a functional perspective.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, R.C., R.P., M.M.; formal analysis, R.C.; investigation, R.P., M.M.; writing—original draft preparation, R.C., R.P., M.M.; writing—review and editing, G.F.; funding acquisition, M.M., R.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Commissione Internazionale per la Protezione delle Acque Italo-Svizzere (CIPAIS).
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Figure A1.
Decomposition of additive time series of some environmental values: temperature (A; mean water temperature of the layer 0-20m), chlorophyll-a (B), NO3 nitrates (C) and conductivity (D).
Figure A1.
Decomposition of additive time series of some environmental values: temperature (A; mean water temperature of the layer 0-20m), chlorophyll-a (B), NO3 nitrates (C) and conductivity (D).
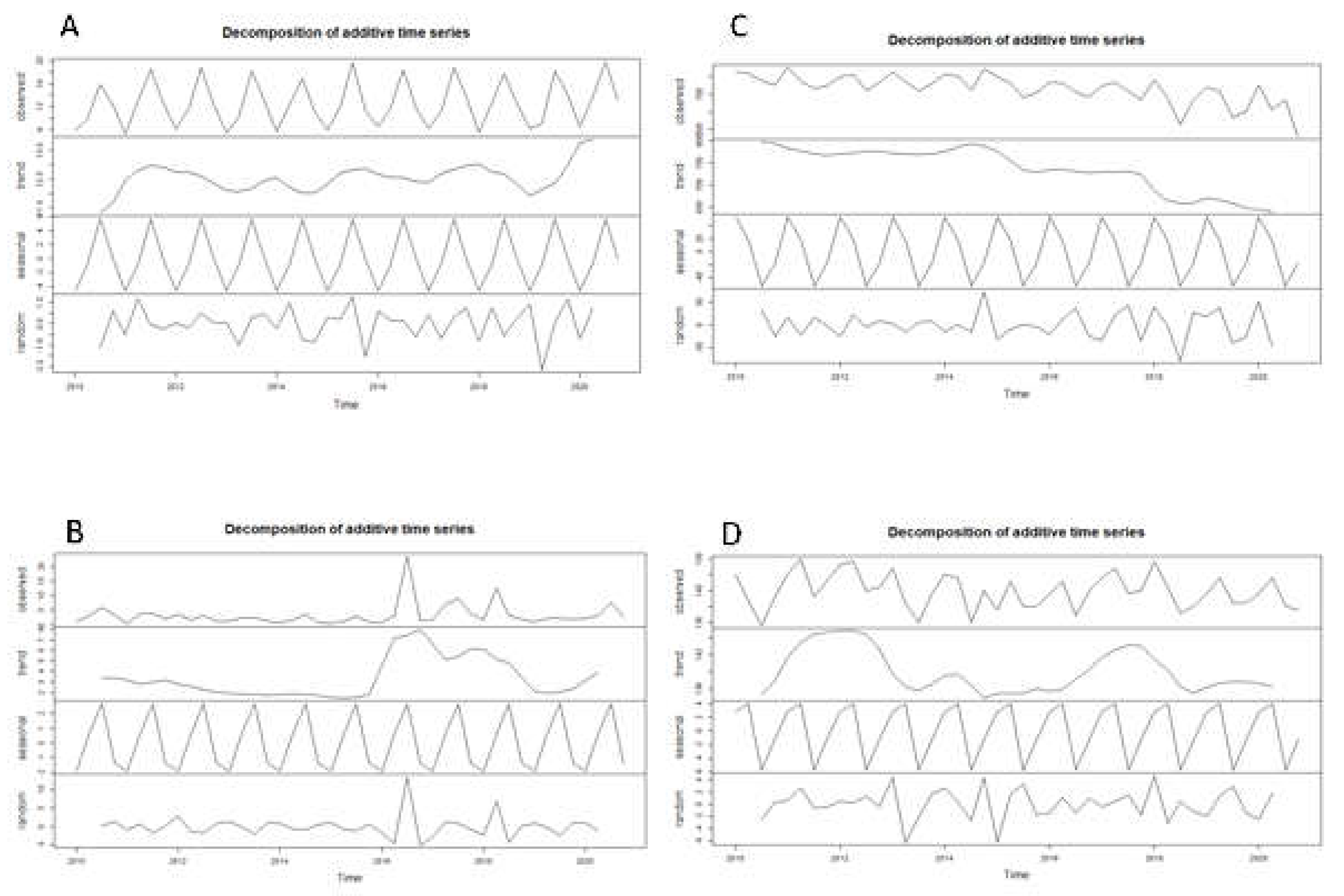
Figure A2.
Decomposition of additive time series of carbon isotopic values of Daphnia (A), Bosmina (B), Diaptomids (C) and Cyclopoids (D).
Figure A2.
Decomposition of additive time series of carbon isotopic values of Daphnia (A), Bosmina (B), Diaptomids (C) and Cyclopoids (D).
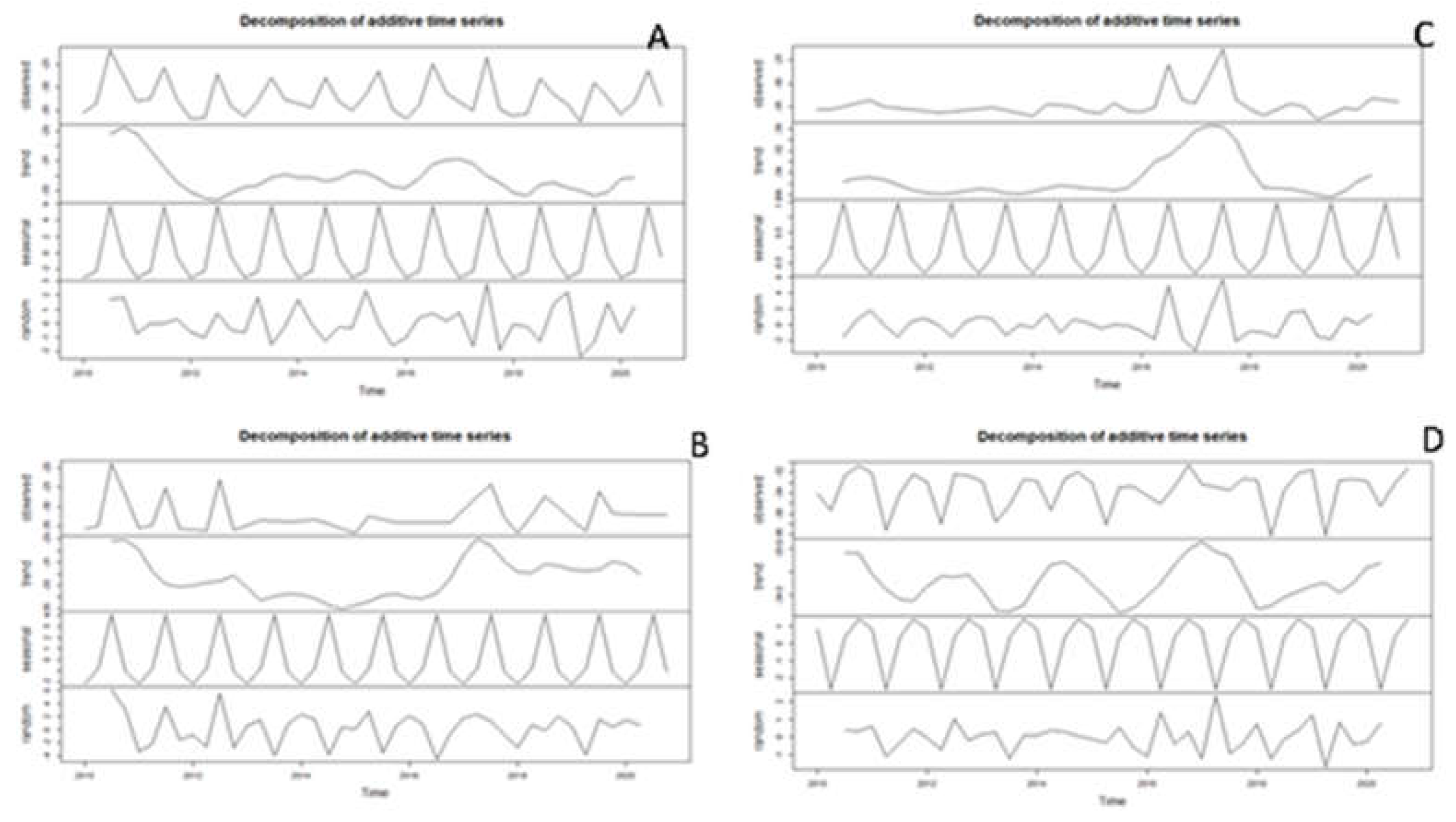
Figure A3.
Decomposition of additive time series of nitrogen isotopic values of Daphnia (A) and Bosmina (B).
Figure A3.
Decomposition of additive time series of nitrogen isotopic values of Daphnia (A) and Bosmina (B).
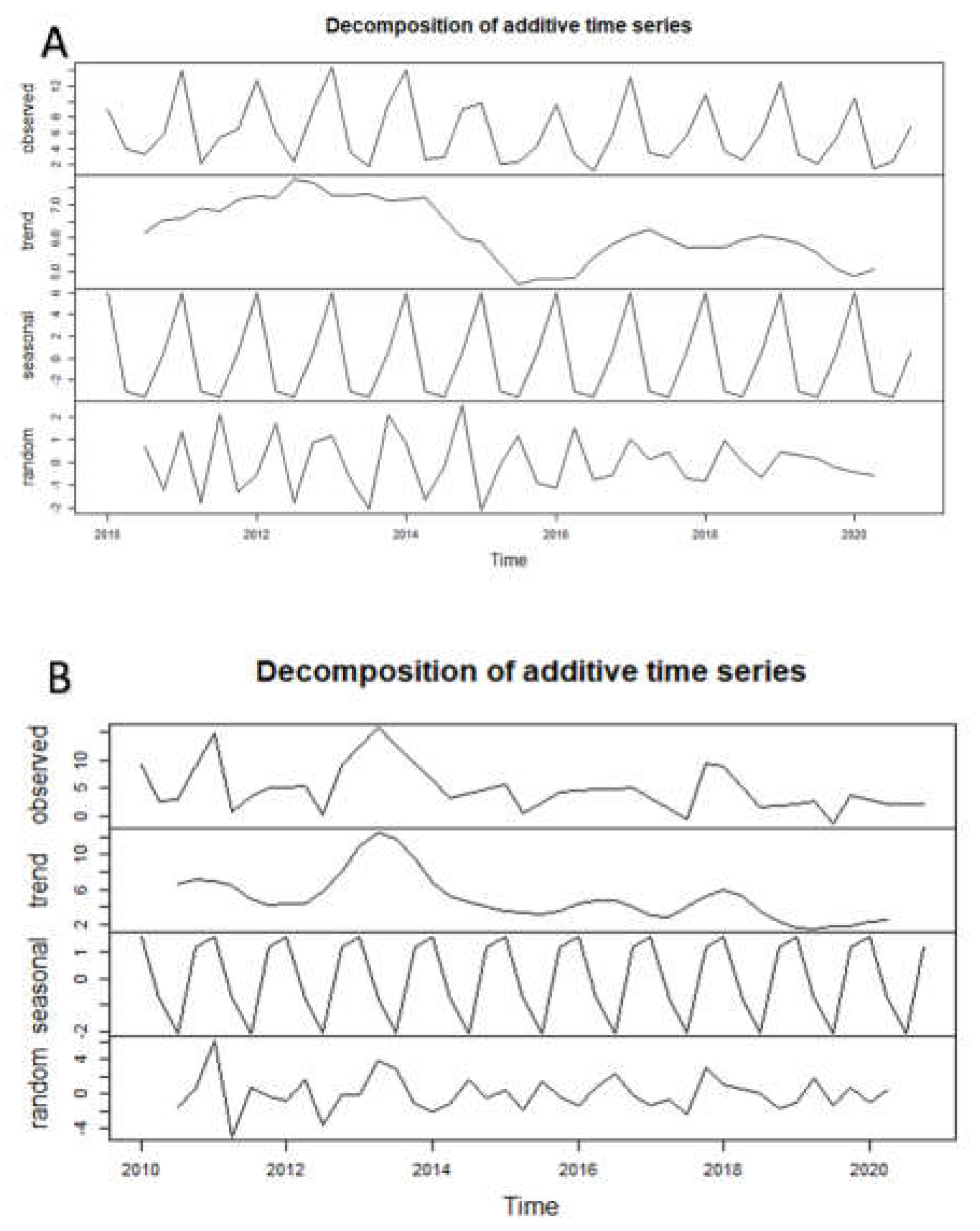
Figure A4.
Seasonal vertical profiles of water temperature of the upper 50 m in Lake Maggiore, from 2009 to 2020.
Figure A4.
Seasonal vertical profiles of water temperature of the upper 50 m in Lake Maggiore, from 2009 to 2020.
References
- Moss, B. Cogs in the endless machine: lakes, climate change and nutrient cycles: a review. STOTEN, 2012, 434, 130-142. [CrossRef]
- Kagalou, I., Papastergiadou, E., Leonardos, I. Long term changes in the eutrophication process in a shallow Mediterranean lake ecosystem of W. Greece: Response after the reduction of external load. J. Environ. Manage., 2008, 87(3), 497-506. [CrossRef]
- Foley, B., Jones, I.D., Maberly, S.C., Rippey, B. Long-term changes in oxygen depletion in a small temperate lake: effects of climate change and eutrophication. Freshwat. Biol., 2012, 57(2), 278-289. [CrossRef]
- Arfè, A., Quatto, P., Zambon, A., MacIsaac, H. J., Manca, M. Long-term changes in the zooplankton community of Lake Maggiore in response to multiple stressors: A functional principal components analysis. Water, 2019, 11(5), 962. [CrossRef]
- Rogora, M., Austoni, M., Caroni, R., Giacomotti, P., Kamburska, L., Marchetto, A., Mosello, R., Orrù, A., Tartari, G., Dresti, C. Temporal changes in nutrients in a deep oligomictic lake: the role of external loads versus climate change. J. Limnol., 2021, 80(3), 2051. [CrossRef]
- Chang, C.W., Miki, T., Ye, H., Souissi, S., Adrian, R., Anneville, O., Agasild, H., Ban, S., Be’eri-Shlevin, Y., Chiang, Y.R., Feuchtmayr, H., Gal, G., Ichise, S., Kagami, M., Kumagai, M., Liu, X., Matsuzaki, S.-I. S., Manca, M. M., Nõges, P., Piscia, R., Rogora, M., Shiah, F.K., Thackeray, S.J., Widdicombe, C. E., Wu, J.T., Zohary, T., Hsieh, C.-H. Causal networks of phytoplankton diversity and biomass are modulated by environmental context. Nat. Commun., 2022, 13(1), 1-11. [CrossRef]
- Poff, N.L., Brinson, M.M., Day, J.W. Aquatic ecosystems and global climate change. Pew Center on Global Climate Change, Arlington, VA 2002, 44, 1-36. https://www.c2es.org/wp-content/uploads/2002/01/aquatic.pdf.
- Mooij, W.M., Hülsmann, S., De Senerpont Domis, L.N., Nolet, B.A., Bodelier, P.L., Boers, P., Pires, L.M.D., Gons, H.J., Ibelings, B.W., Noordhuis, R., Portielje, R., Wolfstein, K., Lammens, E.H.R.R. The impact of climate change on lakes in the Netherlands: a review. Aquat. Ecol., 2005, 39(4), 381-400. [CrossRef]
- Tylianakis, J.M., Didham, R.K., Bascompte, J., Wardle, D.A. Global change and species interactions in terrestrial ecosystems. Ecol. Lett., 2008, 11(12), 1351-1363. [CrossRef]
- Williamson, C.E., Saros, J.E., Vincent, W.F., Smol, J.P. Lakes and reservoirs as sentinels, integrators, and regulators of climate change. Limnol. Oceanogr., 2009, 54(6), 2273-2282. [CrossRef]
- Fenocchi, A., Rogora, M., Sibilla, S., Dresti, C. Relevance of inflows on the thermodynamic structure and on the modeling of a deep subalpine lake (Lake Maggiore, Northern Italy/Southern Switzerland). Limnologica, 2017, 63, 42-56. [CrossRef]
- Tanentzap, A.J., Morabito, G., Volta, P., Rogora, M., Yan, N.D., Manca, M. Climate warming restructures an aquatic food web over 28 years. Glob. Change Biol., 2020, 26(12), 6852-6866. [CrossRef]
- Jeppesen, E., Meerhoff, M., Holmgren, K., González-Bergonzoni, I., Teixeira-de Mello, F., Declerck, S. A., De Meester, L., Søndergaard, M., Lauridsen, T.L., Bjerring, R., Conde-Porcuna, J.M., Mazzeo, N., Iglesias, C., Reizenstein, M., Malmquist, I.J., Liu, Z., Balayla, D., Lazzaro, X. Impacts of climate warming on lake fish community structure and potential effects on ecosystem function. Hydrobiologia, 2010, 646(1), 73-90. [CrossRef]
- Jeppesen, E., Brucet, S., Naselli-Flores, L., Papastergiadou, E., Stefanidis, K., Noges, T., Noges, P., Attayde, J.L., Zohary, T., Coppens, J., Bucak, T., Menezes, R.F., Freitas, F.R.S., Kernan, M., Søndergaard, M., Beklioğlu, M. Ecological impacts of global warming and water abstraction on lakes and reservoirs due to changes in water level and related changes in salinity. Hydrobiologia, 2015, 750(1), 201-227. [CrossRef]
- Lin, Q., Xu, L., Hou, J., Liu, Z., Jeppesen, E., Han, B.P. Responses of trophic structure and zooplankton community to salinity and temperature in Tibetan lakes: Implication for the effect of climate warming. Wat. Res., 2017, 124, 618-629. [CrossRef]
- Rantala, M.V., Luoto, T.P., Weckström, J., Rautio, M., Nevalainen, L. Climate drivers of diatom distribution in shallow subarctic lakes. Freshwat. Biol., 2017, 62(12), 1971-1985. [CrossRef]
- Woolway, R.I., Merchant, C.J. Worldwide alteration of lake mixing regimes in response to climate change. Nat. Geosci., 2019, 12(4), 271-276. [CrossRef]
- Woolway, R.I., Kraemer, B.M., Lenters, J.D., Merchant, C.J., O’Reilly, C.M., Sharma, S. Global lake responses to climate change. Nat. Rev. Earth Environ., 2020, 1(8), 388-403. [CrossRef]
- Sommer, U., Adrian, R., De Senerpont Domis, L., Elser, J.J., Gaedke, U., Ibelings, B., Jeppesen, E., Lurling, M., Molinero, J. C., Mooij, W.M., van Donk, E., Winder, M. Beyond the Plankton Ecology Group(PEG) Model: Mechanisms Driving Plankton Succession. Annu. Rev. Ecol. Evol. Syst., 2012, 43, 429-448. 2012. [CrossRef]
- Varpe, Ø. Fitness and phenology: annual routines and zooplankton adaptations to seasonal cycles. J. Plank. Res., 2012, 34(4), 267-276. [CrossRef]
- Kürten, B., Painting, S.J., Struck, U., Polunin, N.V., Middelburg, J.J. Tracking seasonal changes in North Sea zooplankton trophic dynamics using stable isotopes. Biogeochemistry, 2013, 113(1), 167-187. [CrossRef]
- Bănaru, D., Carlotti, F., Barani, A., Grégori, G., Neffati, N., Harmelin-Vivien, M. Seasonal variation of stable isotope ratios of size-fractionated zooplankton in the Bay of Marseille (NW Mediterranean Sea). J. Plank. Res., 2014, 36(1), 145-156. [CrossRef]
- Moss, B.R. Ecology of fresh waters: man and medium, past to future. John Wiley & Sons Pub., 2009, 482 pp ISBN-10:1405113324.
- Visconti, A., Caroni, R., Rawcliffe, R., Fadda, A., Piscia, R., Manca, M. Defining seasonal functional traits of a freshwater zooplankton community using δ13C and δ15N stable isotope analysis. Water, 2018, 10(2), 108. [CrossRef]
- Cicala, D., Polgar, G., Mor, J. R., Piscia, R., Brignone, S., Zaupa, S., Volta, P. Trophic niches, trophic positions, and niche overlaps between non-native and native fish species in a subalpine lake. Water, 2020, 12(12), 3475. [CrossRef]
- Pel, R., Hoogveld, H., Floris, V. Using the hidden isotopic heterogeneity in phyto-and zooplankton to unmask disparity in trophic carbon transfer. Limnol. Oceanogr., 2003, 48(6), 2200-2207. [CrossRef]
- Matthews, B., Mazumder, A. Temporal variation in body composition (C:N) helps explain seasonal patterns of zooplankton δ13C. Freshwat. Biol., 2005, 50, 502–515. [CrossRef]
- Perga M.E., Gerdaux, D. Seasonal variability in the δ13C and δ15N values of the zooplankton taxa in two alpine lakes. Acta Oecol., 2006, 30, 69-77. [CrossRef]
- Gearing, J.N., Gearing, P.J., Rudnick, D.T., Requejo, A.G., Hutchins, M.J. Isotopic variability of organic carbon in a phytoplankton-based, temperate estuary. Geochim. Cosmochim. Acta, 1984, 48(5), 1089-1098. [CrossRef]
- Rau, G.H., Takahashi, T., Marais, D.J.D. Latitudinal variations in plankton δ13C: implications for CO2 and productivity in past oceans. Nature, 1989, 341(6242), 516-518. [CrossRef]
- Goericke, R., Fry, B. (1994). Variations of marine plankton δ13C with latitude, temperature, and dissolved CO2 in the world ocean. Glob. Biogeochem. Cycles, 1994, 8(1), 85-90. [CrossRef]
- Miller, T.W., Brodeur, R.D., Rau, G.H. Carbon stable isotopes reveal relative contribution of shelf-slope production to the Northern California Current pelagic community. Limnol. Oceanogr., 2008, 53(4), 1493-1503. [CrossRef]
- Lara, R.J., Alder, V., Franzosi, C.A., Kattner, G. Characteristics of suspended particulate organic matter in the southwestern Atlantic: influence of temperature, nutrient and phytoplankton features on the stable isotope signature. J. Mar. Systems, 2010, 79(1-2), 199-209. [CrossRef]
- Papiol, V., Cartes, J.E., Fanelli, E., Rumolo, P. Food web structure and seasonality of slope megafauna in the NW Mediterranean elucidated by stable isotopes: relationship with available food sources. J. Sea Res., 2013, 77, 53-69. [CrossRef]
- Kürten, B., Al-Aidaroos, A.M., Struck, U., Khomayis, H.S., Gharbawi, W.Y., Sommer, U. Influence of environmental gradients on C and N stable isotope ratios in coral reef biota of the Red Sea, Saudi Arabia. J. Sea Res., 2014, 85, 379-394. [CrossRef]
- Gu, B., Schelske, C.L., Waters, M.N. Patterns and controls of seasonal variability of carbon stable isotopes of particulate organic matter in lakes. Oecologia, 2011, 165(4), 1083-1094. [CrossRef]
- Hou W., Gu B., Zhang, H., Gu, J, Han, B.P. The relationship between carbon and nitrogen stable isotopes of zooplankton and select environmental variables in low-latitude reservoirs. Limnology, 2013, 14(I), 97-104. [CrossRef]
- Ruggiu, D., Morabito, G., Panzani, P., Pugnetti, A. Trends and relations among basic phytoplankton characteristics in the course of the long-term oligotrophication of Lake Maggiore (Italy). Hydrobiologia, 1998, 369/370, 243–257. [CrossRef]
- Salmaso, N., Mosello, R. Limnological research in the deep southern subalpine lakes: synthesis, directions and perspectives. Adv. Oceanogr.Limnol., 2010, 1(1), 29-66. [CrossRef]
- Tapolczai, K., Anneville, O., Padisák, J., Salmaso, N., Morabito, G., Zohary, T., Tadonléké, R.D., Rimet, F. Occurrence and mass development of Mougeotia spp. (Zygnemataceae) in large, deep lakes. Hydrobiologia, 2015, 745(1), 17-29. [CrossRef]
- Bresciani, M., Cazzaniga, I., Austoni, M., Sforzi, T., Buzzi, F., Morabito, G., Giardino, C. Mapping phytoplankton blooms in deep subalpine lakes from Sentinel-2A and Landsat-8. Hydrobiologia, 2018, 824(1), 197-214. [CrossRef]
- Morabito, G., Oggioni, A., Austoni, M. Resource ratio and human impact: how diatom assemblages in Lake Maggiore responded to oligotrophication and climatic variability. In: Phytoplankton responses to human impacts at different scales. Salmaso, N., Naselli-Flores, L., Cerasino, L., Flaim, G., Tolotti, M., Padisák, J. (eds), Develop. Hydrobiol., 2012, 221, 47-60. [CrossRef]
- Tanentzap, A.J., Fitch, A., Orland, C., Emilson, E.J., Yakimovich, K.M., Osterholz, H., Dittmar, T. Chemical and microbial diversity covary in fresh water to influence ecosystem functioning. PNAS, 2019, 116(49), 24689-24695. [CrossRef]
- Fenocchi, A., Rogora, M., Sibilla, S., Ciampittiello, M., Dresti, C. Forecasting the evolution in the mixing regime of a deep subalpine lake under climate change scenarios through numerical modelling (Lake Maggiore, Northern Italy/Southern Switzerland). Clim. Dyn., 2018, 51(9), 3521-3536. [CrossRef]
- Rogora, M., Buzzi, F., Dresti, C., Leoni, B., Lepori, F., Mosello, R., Patelli, M., Salmaso, N. Climatic effects on vertical mixing and deep-water oxygen content in the subalpine lakes in Italy. Hydrobiologia, 2018, 824(1), 33-50. [CrossRef]
- Zohary T., Erez J., Gophen M., Berman-Frank I., Stiller M. Seasonality of stable carbon isotopes within the pelagic food web of Lake Kinneret. Limnol. Oceanogr., 1994, 39(5), 1030-1043. [CrossRef]
- Schindler, D.W. The cumulative effects of climate warming and other human stresses on Canadian freshwaters in the new millennium. Can. J. Fish. Aquat. Sci., 2001, 58, 18-29. [CrossRef]
- Visconti, A., Manca, M., de Bernardi, R. Eutrophication-like response to climate warming: an analysis of Lago Maggiore (N. Italy) zooplankton in contrasting years. J. Limnol., 2008, 67(2), 87-92. [CrossRef]
- Takahashi, K., Yoshioka, T., Wada, E., Sakamoto, M. Temporal variations in carbon isotope ratio of phytoplankton in a eutrophic lake. J. Plank. Res., 1990, 12(4), 799-808. [CrossRef]
- Visconti, A., Manca, M. Seasonal changes in the δ13C and δ15N signatures of the Lago Maggiore pelagic food web. J. Limnol., 2011, 70(2), 263–271. [CrossRef]
- Matthews, B., Mazumder, A. (2003). Compositional and interlake variability of zooplankton affect baseline stable isotope signatures. Limnol. Oceanogr., 2003, 48(5), 1977–1987. [CrossRef]
- Wetzel, R.G. Limnology: lake and river ecosystems. Academic Press, London, 2001, 1024 pp. eBook ISBN: 9780080574394.
- Post, D.M., Pace, M.L., Hairston Jr, N.G. Ecosystem size determines food-chain length in lakes. Nature, 2000, 405(6790), 1047-1049. [CrossRef]
- DeNiro, M. J., Epstein, S. Influence of diet on the distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta, 1978, 42, 495-506. [CrossRef]
- Ambrosetti, W., Barbanti, L., Carrara, E.A. (2010). Mechanisms of hypolimnion erosion in a deep lake (Lago Maggiore, N. Italy). J. Limnol., 2010, 69(1), 3-14. [CrossRef]
- APHA, AWWA and WEF. Standard Methods for the Examination of Water & Wastewater, 22nd Edition. 2012, Washington DC: American Public Health Association.
- P. A. T. IRSA-CNR. Metodi analitici per le acque. Manuali e Linee guida, 2003, 29, 1153. ISBN 88-448-0083-7.
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. Springer Cham Pubb., 2006, 260pp. [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing (version 3.6.1). Vienna, Austria: R Foundation for Statistical Computing, 2019. https://www.R-project.org/.
- Piscia, R., Mazzoni, M., Bettinetti, R., Caroni, R., Cicala, D., Manca, M.M. Stable Isotope Analysis and persistent organic pollutants in crustacean zooplankton: The role of size and seasonality. Water, 2019, 11(7), 1490. [CrossRef]
- Lehmann, M.F., Bernasconi, S.M., McKenzie, J.A. Seasonal variation of the δ13C and δ15N of particulate and dissolved carbon and nitrogen in Lake Lugano: Constraints on biogeochemical cycling in a eutrophic lake. Limnol. Oceanogr., 2004, 49(2), 415-429. [CrossRef]
- Grey, J., Jones R.I. Seasonal changes in the importance of the source of organic matter to the diet of zooplankton in Loch Ness, as indicated by stable isotope analysis. Limnol. Oceanogr., 2001, 46(3), 505–513. [CrossRef]
- Caroni, R., Free, G., Visconti, A., Manca, M. Phytoplankton functional traits and seston stable isotopes signature: a functional-based approach in a deep, subalpine lake, Lake Maggiore (N. Italy). J. Limnol., 2012, 71(1), 84-94. [CrossRef]
- Fadda, A., Rawcliffe, R., Padedda, B. M., Luglie, A., Sechi, N., Camin, F., Ziller, L., Manca, M. Spatiotemporal dynamics of C and N isotopic signature of zooplankton: a seasonal study on a man-made lake in the Mediterranean region. Ann. Limnol.-Int. J. Limnol., 2014; 50(4), 279-287. [CrossRef]
- Rau, G.H., Takahashi, T., Des Marais, D.J., Repeta, D.J., Martin, J.L. The relationship between δ13C of organic matter and [CO2 (aq)] in ocean surface water: data from a JGOFS site in the northeast Atlantic Ocean and a model. Geochim. Cosmochim. Acta, 1992, 56(3), 1413-1419. [CrossRef]
- Yoshioka, T., Wada, E., Hayashi, H. A stable isotope study on seasonal food web dynamics in a eutrophic lake. Ecology, 1994, 75(3), 835-846. [CrossRef]
- Gu, B., Alexander, V., Schell, D.M. Seasonal and interannual variability of plankton carbon isotope ratios in a subarctic lake. Freshwat. Biol., 1999, 42(3), 417-426. [CrossRef]
- Masclaux, H., Richoux, N.B. Effects of temperature and food quality on isotopic turnover and discrimination in a cladoceran. Aquat. Ecol., 2017, 51(1), 33-44. [CrossRef]
- Power, M., Guiguer, K.R.R.A., Barton, D.R. Effects of temperature on isotopic enrichment in Daphnia magna: implications for aquatic food-web studies. RCM, 2003, 17(14), 1619-1625. [CrossRef]
- Gu, B. Schelske, D.M. Temporal and spatial variations in phytoplankton carbon isotopes in a polymictic subtropical lake. J. Plank. Res., 1996, 18, 2081-2092. [CrossRef]
- Vander Zanden, M.J., Rasmussen, J.B. Primary consumer δ13C and δ15N and the trophic position of aquatic consumers. Ecology, 1999, 1395-1404. [CrossRef]
- Grey, J., Jones, R. I., Sleep, D. Stable isotope analysis of the origins of zooplankton carbon in lakes of differing trophic state. Oecologia, 2000, 123, 232-240. [CrossRef]
- Cattaneo, A., Manca, M., Rasmussen, J.B. Peculiarities in the stable isotope composition of organisms from an alpine lake. Aquat. Sci., 2004, 66(4), 440-445. [CrossRef]
- Leoni, B. Zooplankton predators and preys: Body size and stable isotope to investigate the pelagic food in a deep lake (Lake Iseo, Northern Italy). J. Limnol., 2017, 76(1), 85-93. [CrossRef]
- Makino, W., Yoshida, T., Sakano, H., Ban, S. Stay cool: habitat selection of a cyclopoid copepod in a north temperate oligotrophic lake. Freshwat. Biol., 2003, 48(9), 1551-1562. [CrossRef]
- Helland, I.P., Freyhof, J., Kasprzak, P., Mehner, T. Temperature sensitivity of vertical distributions of zooplankton and planktivorous fish in a stratified lake. Oecologia, 2007, 151(2), 322-330. [CrossRef]
- Thackeray, S.J., Elliott, J.A., Fielding, R.F., Swinburne, K. Effects of onset of thermal stratification on vertical distribution of phytoplankton and zooplankton species. Verh. Int. Ver. Theor. Angew. Limnol., 2005, 29(1), 555-559. [CrossRef]
- Smyntek, P.M., Maberly, S.C., Grey, J. Dissolved carbon dioxide concentration controls baseline stable carbon isotope signatures of a lake food web. Limnol. Oceanogr., 2012, 57(5), 1292-1302. [CrossRef]
- Smyntek, P.M., Teece, M.A., Schulz, K.L., Storch, A.J. Taxonomic differences in the essential fatty acid composition of groups of freshwater zooplankton relate to reproductive demands and generation time. Freshwat. Biol., 2008, 53(9), 1768-1782. [CrossRef]
- Manca, M., Cavicchioni, N., Morabito, G. First observations on the effect of a complete, exceptional overturn of Lake Maggiore on plankton and primary productivity. Int. Rev. Hydrobiol., 2000, 85(2-3), 209-222. [CrossRef]
- France, R.L. Differentiation between littoral and pelagic food webs in lakes using stable carbon isotopes. Limnol. Oceanogr., 1995, 40(7), 1310-1313. [CrossRef]
- Croteau, M.N., Luoma, S.N., Stewart, A.R. Trophic transfer of metals along freshwater food webs: evidence of cadmium biomagnification in nature. Limnol. Oceanogr., 2005, 50(5), 1511-1519. [CrossRef]
- Visconti, A., Volta, P., Fadda, A., Manca, M. Roach in Lake Maggiore: A Peaceful Invasion Detected with C, N Stable Isotope Analysis. GJSFR, 2013, 13, 1-8.
- Buhan, E., Kaymak, N., Akin, S., Turan, H. Trophic Pathways from Pelagic and Littoral Sources Supports Food Web in A Eutrophic Natural Lake (Lake Zinav, Turkey). Turk. J. Fish. Aquat. Sci., 2018, 19(2), 99-107. [CrossRef]
- Geller, W., Müller, H. The filtration apparatus of Cladocera: filter mesh-sizes and their implications on food selectivity. Oecologia, 1981, 49(3), 316-321. [CrossRef]
- DeMott, W.R., Kerfoot, C.W. Competition among cladocerans: nature of the interaction between Bosmina and Daphnia. Ecology, 1982, 63(6), 1949-1966. [CrossRef]
- Deng, D., Xie, P., Zhou, O., Yang, H., Guo, L., Geng, H. Field and experimental studies on the combined impacts of cyanobacterial blooms and small algae on crustacean zooplankton in a large, eutrophic, subtropical, Chinese lake. Limnology, 2008, 9(1), 1-11. [CrossRef]
- Carpenter, E.J., Montoya, J.P., Burns, J., Mulholland, M.R., Subramaniam, A., Capone, D.G. Extensive bloom of a N2-fixing diatom/cyanobacterial association in the tropical Atlantic Ocean. Mar. Ecol. Progr. Series, 1999, 185, 273-283. [CrossRef]
- Bauersachs, T., Schouten, S., Compaoré, J., Wollenzien, U., Stal, L.J., Damsteé, J.N.S. Nitrogen isotopic fractionation associated with growth on dinitrogen gas and nitrate by cyanobacteria. Limnol Oceanog., 2009, 54(4), 1403-1411. [CrossRef]
- Gu, B., Chapman, A.D., Schelske, C.L. Factors controlling seasonal variations in stable isotope composition of particulate organic matter in a softwater eutrophic lake. Limnol. Oceanogr., 2006, 51(6), 2837-2848. [CrossRef]
- Jones, R.I., Grey, J., Quarmby, C., Sleep, D. An assessment using stable isotopes of the importance of allochthonous organic carbon sources to the pelagic food web in Loch Ness. Proc. R. Soc. Lond. B, 1998, 265, 105–111. [CrossRef]
- Winder, M., Hunter, D.A. Temporal organization of phytoplankton communities linked to physical forcing. Oecologia, 2008, 156(1), 179-192. [CrossRef]
- Lampert, W. The adaptive significance of diel vertical migration of zooplankton. Func. Ecol., 1989, 3(1), 21-27. [CrossRef]
- Hanazato, T. Direct and indirect effects of low-oxygen layers on lake zooplankton communities. Ergeb. Limnol., 1992, 35, 87-98.
- Dini, M.L., Soranno, P.A., Scheuerell, M., Carpenter, S.R. Effects of predators and food supply on diel vertical migration of Daphnia. In: The trophic cascade in lakes; Carpenter, S.R., Kitchell, J.F. Eds., Cambridge University Press, 1993, 153-171. [CrossRef]
- Persaud, A.D., Williamson, C.E. Ultraviolet and temperature effects on planktonic rotifers and crustaceans in northern temperate lakes. Freshwat. Biol., 2005, 50(3), 467-476. doi:10.1111/j.1365-2427.2005.01334.x. [CrossRef]
- Thackeray, S.J., George, D.G., Jones, R.I., Winfield, I.J. Statistical quantification of the effect of thermal stratification on patterns of dispersion in a freshwater zooplankton community. Aquat. Ecol., 2006, 40(1), 23-32. [CrossRef]
- Gelinas, M., Pinel-Alloul, B. Summer depth selection in crustacean zooplankton in nutrient-poor boreal lakes is affected by recent residential development. Freshwat. Biol., 2008, 53(12), 2438-2454. [CrossRef]
- Williamson, C.E., Rose, K.C. Ultraviolet insights: Attempting to resolve enigmatic patterns in pelagic freshwaters–The historical context and a view to the future. Int. Rev. Hydrobiol., 2009, 94(2), 129-142. [CrossRef]
- Kratz, T.K., MacIntyre, S., Webster, K.E. Causes and consequences of spatial heterogeneity in lakes. In Ecosystem function in heterogeneous landscapes. Springer, New York, NY, 2005, 329-347 pp.
- Borrelli, J.J., Relyea, R.A. A review of spatial structure of freshwater food webs: Issues and opportunities modeling within-lake meta-ecosystems. Limnol. Oceanogr., 2022, 67, 1746–1759. [CrossRef]
- Bertoni, R., Callieri, C. Organic carbon trend during the oligotrophication of Lago Maggiore. In: Strategies for lake ecosystems beyond 2000; de Bernardi, R., Pagnotta, R., Pugnetti, A. Eds., Mem. Ist. Ital. Idrobiol., 1993, 52, 191-205.
- Smith, V.H. Nutrient dependence of primary productivity in lakes 1. Limnol. Oceanogr., 1979, 24(6), 1051-1064. [CrossRef]
- Beaver, J.R., Crisman, T.L. Temporal variability in algal biomass and primary productivity in Florida lakes relative to latitudinal gradients, organic color and trophic state. Hydrobiologia, 1991, 224(2), 89-97. [CrossRef]
- Cifuentes, L.A., Sharp, J.H., Fogel, M.L. Stable carbon and nitrogen isotope biogeochemistry in the Delaware estuary. Limnol. Oceanogr., 1998, 33(5), 1102-1115. [CrossRef]
- 105 Fry, B., Wainright, S.C. Diatom sources of 13C-rich carbon in marine food webs. Mar. Ecol. Progr. Series, 1991, 76(2), 149-157. [CrossRef]
- Nakatsuka, T., Handa, N., Wada, E., Wong, C.S. The dynamic changes of stable isotopic ratios of carbon and nitrogen in suspended and sedimented particulate organic matter during a phytoplankton bloom. J. Mar. Res., 1992, 50(2), 267-296. [CrossRef]
- Takahashi, A., Takeda, K., Ohnishi, T. Light-induced anthocyanin reduces the extent of damage to DNA in UV-irradiated Centaurea cyanus cells in culture. Plant Cell Physiol., 1991, 32(4), 541-547. [CrossRef]
- Laws, E.A., Popp, B.N., Bidigare, R.R., Kennicutt, M.C., Macko, S.A. Dependence of phytoplankton carbon isotopic composition on growth rate and [CO2) aq: theoretical considerations and experimental results. Geochim. Cosmochim. Acta, 1995, 59(6), 1131-1138. [CrossRef]
- Francois, R., Altabet, M.A., Goericke, R., McCorkle, D.C., Brunet, C., Poisson, A. Changes in the δ13C of surface water particulate organic matter across the subtropical convergence in the SW Indian Ocean. Glob. Biogeochem. Cycles, 1993, 7(3), 627-644. [CrossRef]
- France, R.L., Del Giorgio, P.A., Westcott, K.A. Productivity and heterotrophy influences on zooplankton delta13C in northern temperate lakes. Aquat. Microb. Ecol., 1997, 12(1), 85-93. [CrossRef]
- Leggett, M.F., Servos, M.R., Hesslein, R., Johannsson, O., Millard, E.S., Dixon, D.G. Biogeochemical influences on the carbon isotope signatures of Lake Ontario biota. Can. J. Fish. Aquat. Sci., 1999, 56, 2211-2218. [CrossRef]
- Leggett, M.F., Johannsson, O., Hesslein, R., Dixon, D.G., Taylor, W.D., Servos, M.R. Influence of inorganic nitrogen cycling on the δ15N of Lake Ontario biota. Can. J. Fish. Aquat. Sci., 2000, 57, 1489-1496. [CrossRef]
- Vuorio, K., Meili, M., Sarvala, J. Taxon-specific variation in the stable isotopic signatures (δ13C and δ15N) of lake phytoplankton. Freshwat. Biol., 2006, 51(5), 807-822. [CrossRef]
- Free, G., Bresciani, M., Pinardi, M., Ghirardi, N., Luciani, G., Caroni, R., Giardino, C. Detecting climate driven changes in chlorophyll-a in deep subalpine lakes using long term satellite data. Water, 2021, 13(6), 866. [CrossRef]
- Gu, B., Ma, L. Smoak, D., Ewe, S., Zhu, Y., Irick, D., Ross, M., Li, Y. Mercury and sulfur environmental assessment for the Everglades. South Florida Environmental Report. South Florida Water Management District, West Palm Beach, 2013 3B18–3B20.
- Gu, B., Schell, D.M., Alexander, V. Stable carbon and nitrogen isotopic analysis of the plankton food web in a subarctic lake. Can. J. Fish. Aquat. Sci., 1994, 51(6), 1338-1344. [CrossRef]
- Gulati, R.D., Bronkhorst, M., Van Donk, E. Feeding in Daphnia galeata on Oscillatoria limnetica and on detritus derived from it. J. Plank. Res., 2001, 23(7), 705-718. [CrossRef]
- Rogora, M., Steingruber, S., Marchetto, A., Mosello, R., Giacomotti, P., Orru, A., Tartari, G.A., Tiberti, R. Response of atmospheric deposition and surface water chemistry to the COVID-19 lockdown in an alpine area. Environ. Sci. Pollut. Res., 2022, 1-18. [CrossRef]
- Webb, B.W., Hannah, D.M., Moore, R.D., Brown, L.E., Nobilis, F. Recent advances in stream and river temperature research. Hydrol. Process., 2008, 22(7), 902-918. [CrossRef]
- Woodward, G, Perkins, D.M., Brown, L.E. Climate change and freshwater ecosystems: impacts across multiple levels of organization. Philos. Trans. R. Soc. Lond., B, Biol. Sci., 2010, 365(1549), 2093-2106. [CrossRef]
- McQueen, D.J., Johannes, M.R.S., Post, J.R., Stewart, T.J., Lean, D.R.S. Bottom-up and top-down impacts on freshwater pelagic community structure. Ecol. Monogr., 1989, 59(3), 289-309. [CrossRef]
- Brett, M.T., Goldman, C.R. A meta-analysis of the freshwater trophic cascade. PNAS, 1996, 93(15), 7723-7726. [CrossRef]
- Brett, M.T., Goldman, C.R. Consumer versus resource control in freshwater pelagic food webs. Science, 1997, 275(5298), 384-386. [CrossRef]
Figure 1.
Boxplot of values (circles refer to means, horizontal bars to medians, boxes to upper and lower quartiles, vertical lines to ranges and dots indicate outliers) of δ15N‰ (a) and δ13C‰ (b) of crustacean zooplankton taxa and groups during the decadal period.
Figure 1.
Boxplot of values (circles refer to means, horizontal bars to medians, boxes to upper and lower quartiles, vertical lines to ranges and dots indicate outliers) of δ15N‰ (a) and δ13C‰ (b) of crustacean zooplankton taxa and groups during the decadal period.
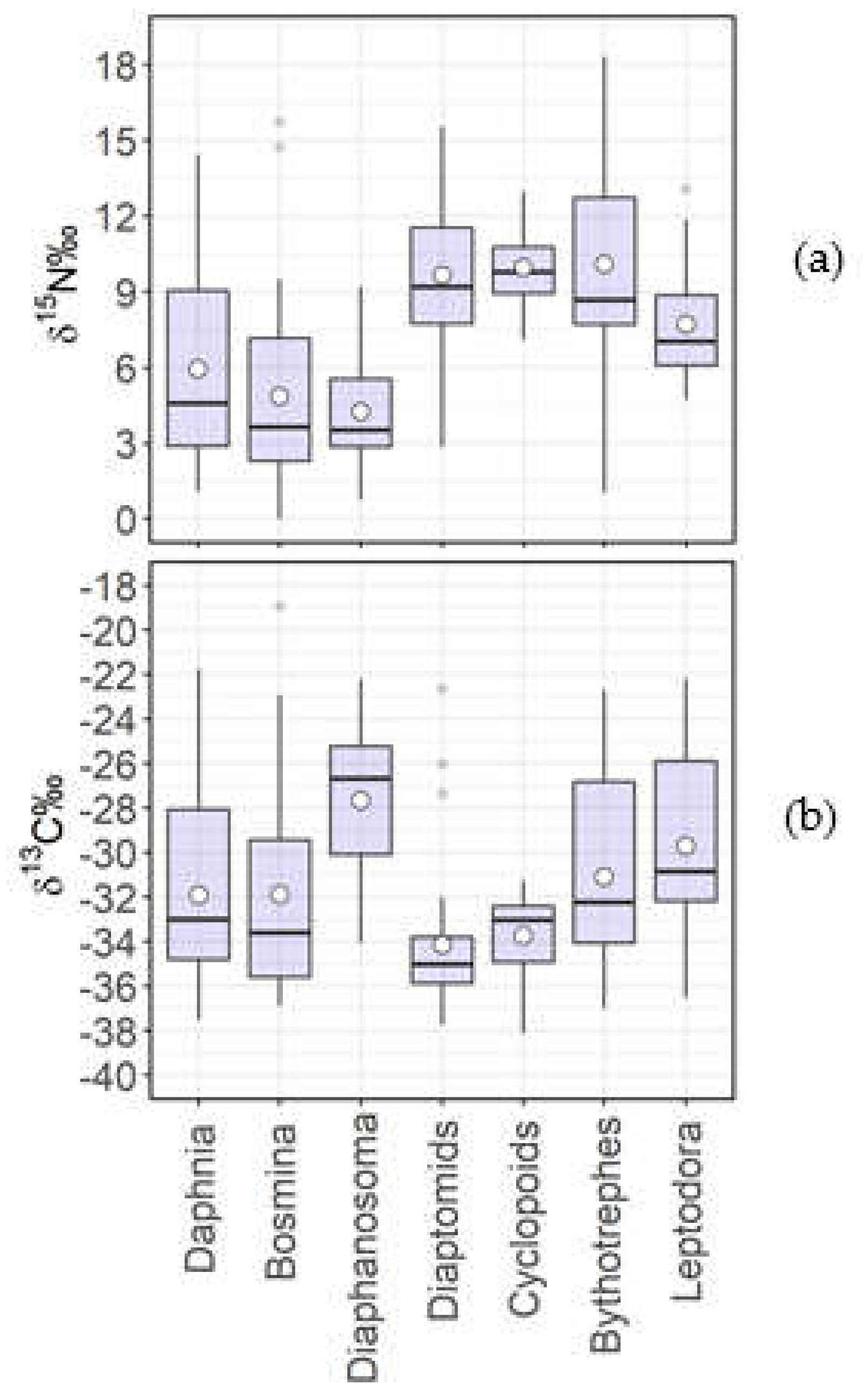
Figure 2.
Trophochemical graph of carbon and nitrogen stable isotope signatures of pooled zooplankton samples (≥450 µm) measured in four seasons of the decadal period.
Figure 2.
Trophochemical graph of carbon and nitrogen stable isotope signatures of pooled zooplankton samples (≥450 µm) measured in four seasons of the decadal period.
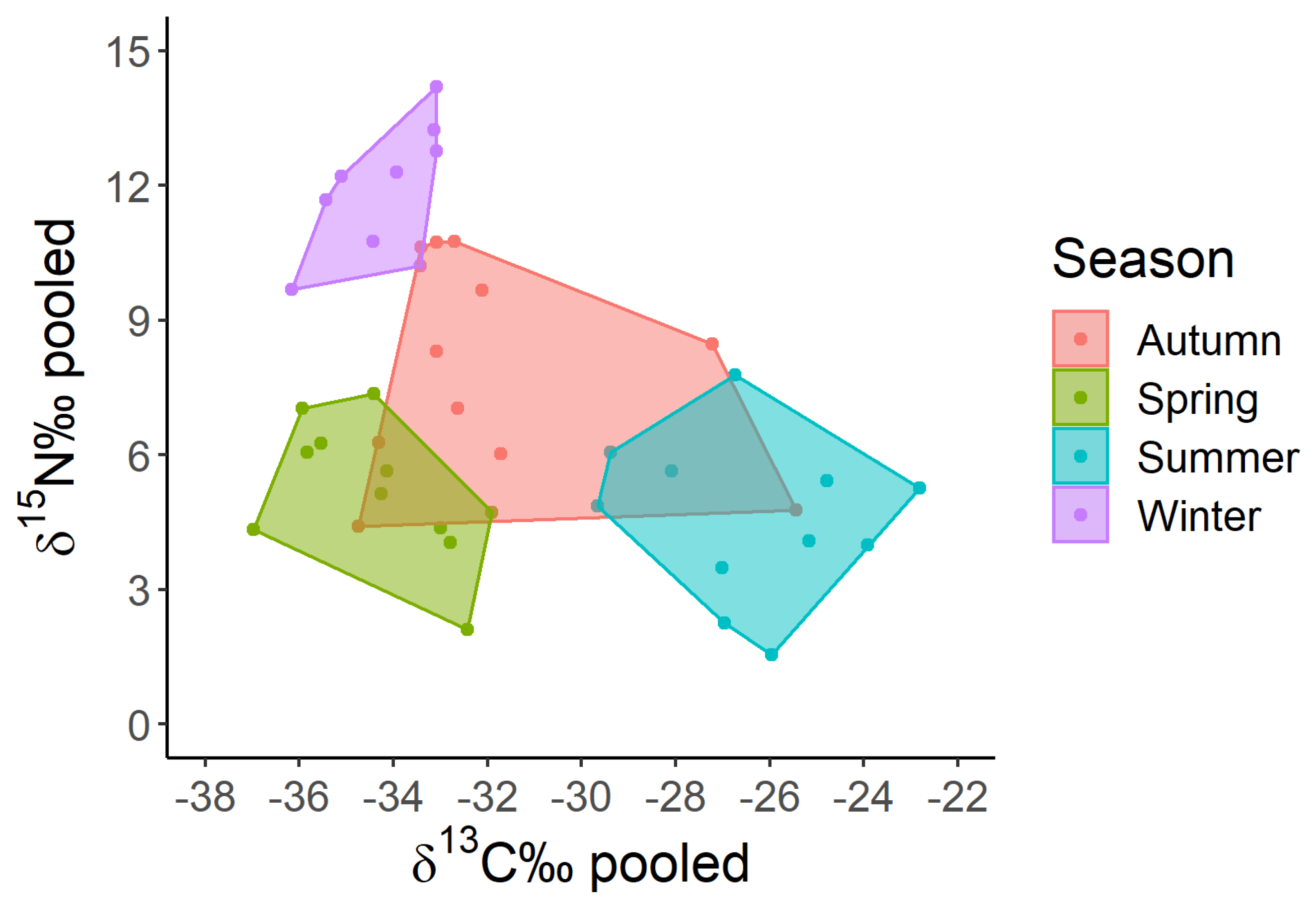
Figure 3.
Trophochemical graph of Daphnia stable isotope signatures measured seasonally during the decadal period.
Figure 3.
Trophochemical graph of Daphnia stable isotope signatures measured seasonally during the decadal period.
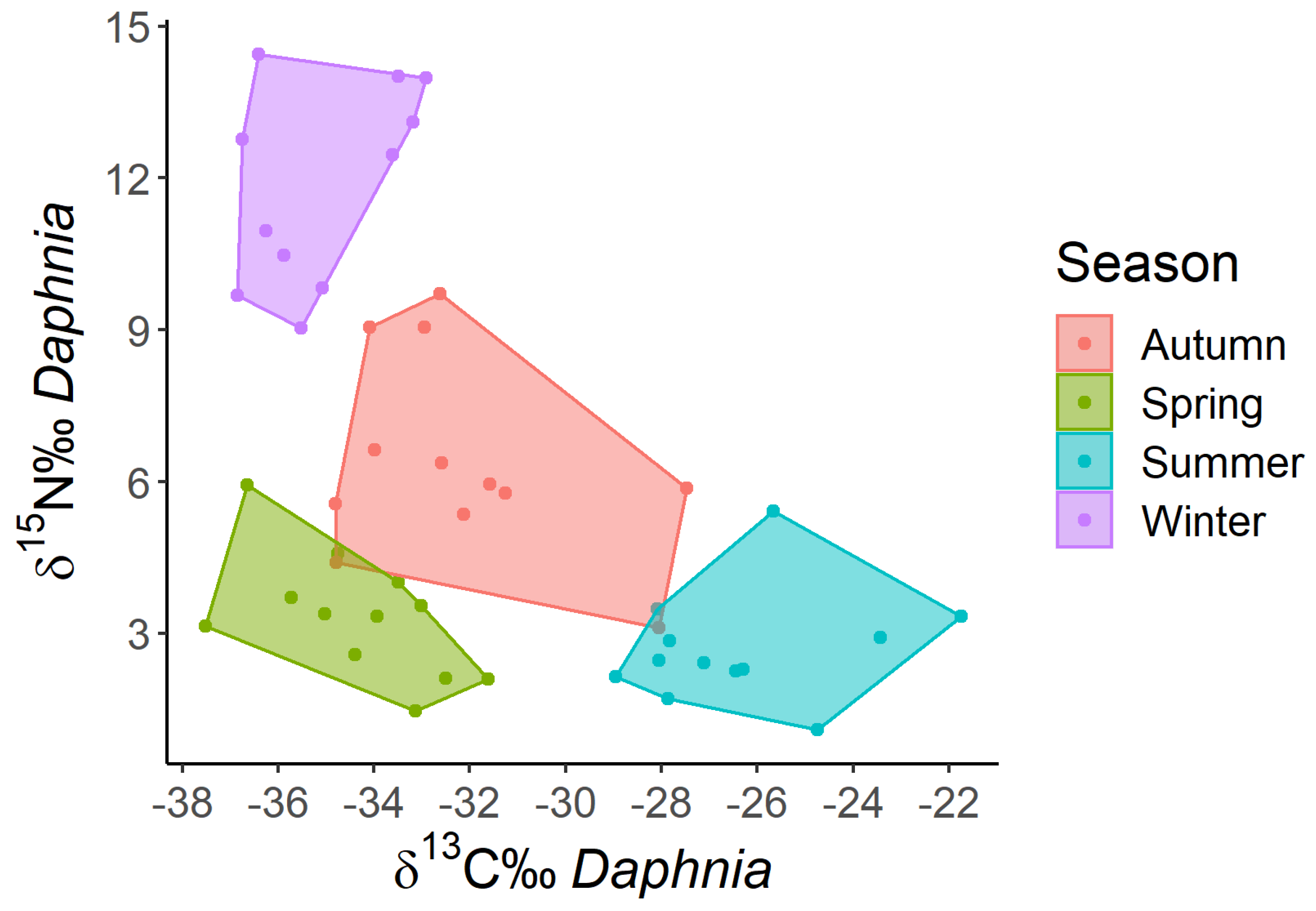
Figure 4.
Daphnia’s carbon isotopic values vs. cyclopoids’ carbon fractionation (F).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



