
Submitted:
10 July 2023
Posted:
11 July 2023
You are already at the latest version
Abstract
During the last years, the increasing evidence of dietary antioxidant compounds and reducing chronic diseases and the relationship between diet and health have promoted an important innovation on the baked product sector, aiming at healthier formulations. This study aims to develop a tool based on mathematical models to predict baked goods total antioxidant capacity (TAC). The high variability of antioxidant properties of flours based on the aspects related to type of grains, varieties, proximal composition and processing, among others, makes very difficult to innovate on food product development without specific analysis. Using Total phenol content (TP), Oxygen radical absorbance capacity (ORAC) and Ferric reducing antioxidant power assay (FRAP) as proxies of antioxidant capacity. Three Bayesian-type models are proposed based on a double exponential parameterised curve that reflects the initial decrease and subsequent increase as a consequence of the observed processes of degradation and generation, respectively, of the antioxidant compounds. Once the values of the main parameters of each curve have been determined, support vector machines (SVR) with exponential kernel were used to predict, based on the temperature and duration of baking as well as the values of proteins and fibers of each native grain, the values of TAC during the baking time.
Keywords:
pulses
; antioxidant capacity
; prediction
; thermal processing
; flour
; Bayesian model
; cereals
; support vector machines (SVR)
1. Introduction
Cereals and legumes are common staple food. These products represent an important source of nutrients from diet and have an irreplaceable role in nutrition [1]. Cereals are frequently consumed as baked goods, such as bread or biscuits, among other products, with legumes gaining importance as total or partial substitutes of cereal flours in those types of products. In the 90´s decade, different aspects such as industrialization and changes in lifestyles, among other factors, favoured the production of under-nourishing baked goods based on refined flours, rich in sugars and saturated fatty acids, with poor fiber and naturally present antioxidants.
Traditionally, fruits and vegetables have been identified as major source of dietary antioxidants, but grains have gained interest due to the proved inverse association between whole grain consumption and risk of chronic diseases, such as cardiovascular diseases, diabetes and cancer [2,3,4,5], due to their high content in fiber and free and bound phenolic compounds [6,7].
During the last years, the increasing awareness by consumers of the relationship between diet and health, and the industrial innovation to satisfy consumer demands, have driven the baked product sector to work on more nutritional and healthier formulations. The market of natural antioxidants, with high demand not only in the food sector but with other applications such as cosmetics and plastic, is expected to grow from $321.4 million in 2022 to $409.7 million in 2032, at a CAGR of 2.4% [8].
Our body is naturally equipped with antioxidant mechanisms in order to control damage from ROS and RNS (Reactive Oxygen or Nitrogen Species); however, dietary antioxidants from food or nutritional supplements are believed to contribute to oxidative balance [9]. Moreover, these compounds affect food quality by avoiding lipid and protein oxidation.
Food bioactive molecules are characterised by their chemical diversity, which complicates the measurement of single compound antioxidant capacity. The concept of total antioxidant capacity (TAC), including the synergic and redox interactions between the different molecules present in the food, was introduced in order to avoid these limitations. The appropriate use of TAC measurements provides support for the interpretation of complex phenomena and can be a tool for screening studies previous to in-depth research investigations [10].
The broad types of chemical structures of antioxidant compounds in food complex matrices makes necessary the use of more than one measurement to assess their antioxidant capacity. One of the main aspects to select antioxidant assays is the reaction mechanism. There are two main mechanisms, single electron transfer (SET) and hydrogen atom transfer (HAT). In SET antioxidant reactions, the free radical loses its condition by "pairing" its unpaired electron. In HAT reactions, the free radical is electronically stabilised through a mechanism that involves the direct transfer of a hydrogen atom [11].
Different assays can be used to estimate the total capacity of foods to scavenge reactive oxygen and nitrogen species, free radicals, or to assess the content of reducing compounds. Among the most popular assays, ABTS+• (2,2′-azinobis-(3-ethylbenzothiazoline-6-sulfonic acid) and DPPH• (2,2-diphenyl-1-picrylhydrazyl) radicals reduction methods can be found; both can be reported with Trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid) as standard. ABTS+• assay is effective to estimate hydrophilic and lipophilic antioxidants, while DPPH• assay has important limitation for lipophilic compounds. Although they are used widely in screening studies, both have limitations associated to oxygen, pH or uncertain reaction kinetic endpoint, which can reduce their reproducibility. Also, both assays use synthetic radicals, which are not present in biological systems.
Oxygen radical absorbance capacity (ORAC) is another widely adopted method; in this case, although the radical initiator is a synthetic compound, the peroxyl radicals produced, against which antioxidant capacity is evaluated, are of relevance for food oxidation mechanisms [12]. Lipid and protein peroxidation have been linked to the pathogenesis of various diseases through different mechanisms, such as disturbance of membrane organization, functional loss of proteins and DNA, or accumulation of modified proteins in cells [13].
The reducing capacity is also important in order to assess the TAC; the ferric reducing antioxidant power assay (FRAP) is based on the reduction of the 2,4,6-tripyridyl- -triazine (TPTZ)–Fe3+ to TPTZ-Fe2+ complex.
Wholegrain cereals and pulses contain a wide range of bioactive components with antioxidant effect such as proteins, dietary fiber (TDF), and phenolic compounds present in free and bound forms [14,15,16], which contribute to product antioxidant capacity. Folin-Ciocalteau assay is generally used to measure total phenol content (TPC); it is not a specific method for phenolic compounds, although it is a quick and effective method to evaluate them. It is important to highlight that this method can also respond to amines and reducing sugars, which can interfere in the real value, but also it can be interesting in the case of processed foods, where Maillard reactions occur, as this method can evaluate the antioxidant capacity of Maillard Reaction Products (MRP) [17].
During the last years (2010-2022), more than 8,000 articles have been published on the antioxidant capacity in foods area [18]. However, there is a high variability of data published, which comes from different aspects such as complexity of food matrices, raw material variability, lack of standards in the assays of the antioxidant capacity, and extraction methodologies, among others.
One strategy to overcome the difficulties of the variability in antioxidant capacity evaluation is relying on extensive databases [19] which can be used for comparison and reference. For example, USDA has developed two databases, one of reducing power values of more than 1100 food raw and processed products [20] and a second of ORAC values, with 326 different food items [21]. Another example would be the attempt for estimation of the daily intake of hydrophilic antioxidants of the Japanese population by Takebayashi and col. [22].
The use of mathematical tools for assessing and predicting antioxidant properties of compounds can also be used as strategy for food antioxidant estimation. QSAR models (Quantitative Structure-Activity Relationships) are used successfully to identify patterns in chemical structure data and correlate and predict with antioxidant capacity of compounds [23]. However, these studies have limited applications to food matrices, where complex mixtures of different compounds are found.
For this reason, the objective of this paper was to develop a mathematical modelling tool based on proximal composition to predict total antioxidant capacity, as measured with three widely adopted methods integrating different aspects of antioxidant capacity assays (TP, ORAC, and FRAP). This work was intended to model the redox status of thermally-processed flours from different types of cereals and pulses, and provide an estimation of TAC of baked cereal and legume-based products to the food industry.
2. Materials and Methods
2.1. Raw material
A total of 136 varieties of 10 different grain types (cereals and pulses) were provided by Experimental Agronomic Programme (PEA) of ITACyL or obtained from local markets. The varieties provided by PEA were cultivated and harvested according to uniform protocols which reduce variability associated to agronomic aspects. The varieties and grains were the following:
Wheat (Chambo, LGWF 16 1321, RW72009, LGWF17 118, LGWE 18 8199, LGWF 17 5114, LGWF 5114, CF 14255, RW 21837, RW 72010, RW 72003, RW 72003, Marcopolo, RW 21846, Filón, Camargo, FDN 18 WW 0240, Nogal, Berdun, LG WE 178300, Boticelli, HAW 18-001, RW21968, LGW 178302, RW 72006, RW71804, Candeal PANE 247, Candeal and 12 commercial flours provided from local markets). Rye (Teodor, Igor, Loretto, Vinetto, Su Promotor, Petkus, Poseido, Arvid, Stannos and two commercial flours provided from local markets. Corn (LG3490, PO937, LG31545, SY Caarioca, Urbanix, 47M, Berlioz, KWS Selecto, RGT Huxxo and 4 commercial flours provided from local markets). Triticale (Kitsurf, Saleroso, Randam, Rivolt, RGTsuliac, Vivacio, RGT/Zaragozac., Trimour, Amarillo/105, LG Plutón, BELOTAC Cerraton (BU), RGT/KADJAC, RGT/COPLAC Cerraton, Bondadoso Cerraton). Oats (9 commercial flours provided from local markets). Rice (6 commercial flours provided from local markets). Sorghum (White: KSH9635W, KSH8G26, KWS Octavius, KSH9G37W, KSH9G32W, P1288Y20 and Brown: KWS Nemesis, PR88P68, Shamal, Foehm, Es Alize). Chickpea (Tauriton, Sultano, Turi, Villaturiel, Urbel, Ituci, Krema, Maragia L., Reale L, Vulcano, Pirón, Kasin and 3 commercial flours provided from local markets). Lentil (Guareña, Paula and 3 commercial flours provided from local markets). Soybean (Sirocca+, Sirocca-, SB8, ES Pallador, Luna, Pepita, Es Isidor, Es Mentor, S/19/12, Primus, SB07, SB44, Proteix, Panoramix, Casleis).
2.2. Sample preparation
Grain samples were reduced to uniform powder using a mill (Model Cyclotec 1093, Foss, Hilleroed, Denmark) fitted with a 0.5 mm screen, and then stored in sealed polyethylene/plastic bags in dark conditions to ensure stability until analysis.
After, wheat and rye flours were used as models for evaluation of thermal processing (temperature and time) on the antioxidant capacity (Figure 1). Biscuit was used as model food. Biscuits were formulated with 9.5 g flour, with 3.6 g sunflower oil and 70 g 100 g-1 of water. Each biscuit had a weight of 15 grams. After kneading, the doughs were cut using a round mould of Ø 9.5 cm and 0.5 cm height. Biscuits were baked at temperatures 180, 200 and 220 °C for a time range of 0-1500 s. The biscuits were frozen and lyophilised in order to stabilise the samples. After that, biscuits were milled to a particle size below 0.5 mm, and stored in sealed polyethylene/plastic bags in dark conditions to ensure stability until analysis.
2.3. Proximal composition
Moisture content was measured gravimetrically by drying samples at 100 °C for 24 h. Total protein content was determined by the Dumas method (AOAC method 990.03) [24]. A conversion factor of 5.7 was used to calculate protein content from nitrogen values. Total fat content was determined using dried samples extracted with petroleum ether (BP 40–60 °C) during 4 h in a Soxtec fat extracting unit (AOAC 2005, method 2003.05) [24]. Ash content was determined by sample incineration in a muffle furnace at 550 °C for 5 h (AOAC 2005, method 923.03) [24]. Carbohydrates were estimated by difference. Total dietary fiber (TDF) content was evaluated using a kit provided by Sigma (TDF100A-1KT, St. Louis, MO, USA), in accordance with manufacturer’s instructions, based on AOAC method 985.29 [24]. All parameters were evaluated in duplicate. Proximal composition analysis was expressed in g 100 g−1 dry matter (d.m.).
2.4. Colorimetric analysis
Colour parameters lightness (L*), redness (a*) and yellowness (b*) were measured using a colorimeter (CM-2600d, Konica Minolta Osaka, Japan) adjusted as D65 standard illuminate, 45/0 sensor and 10 ° standard observer. The colorimeter was standardised using a light trap and a white calibration plate. Measurements were taken directly on the samples.
2.5. Extract preparation
One gram of ground (mesh size 0.5 mm) sample was extracted with 8 mL of methanol: water (1:1, v/v; acidified to pH=2 with 0.1M HCl) in an orbital shaker (250 rpm, 30 min) using magnetic agitation. After centrifugation (2,057 xg, 10 min), the supernatant was collected, filtered (Filter lab paper n. 1249). The extraction was repeated three times. The combined methanol extracts were adjusted to 25 mL. All analyses were performed by duplicate. Extract aliquots were stored at -80 °C until further analysis.
2.6. Total phenol (TP) content
TP were measured using the Folin-Ciocalteu method as described by Slinkard and Singleton, with modifications [25]. The absorbance was measured at 765 nm with a microplate reader (Fluostar Omega, BMG Ortenberg, Germany). Results were calculated using a calibration curve with Gallic acid as standard (9.8-70 mM) and expressed as mg Gallic Acid Equivalents (GAE) 100 g-1 sample (d.m.).
2.7. Oxygen radical absorbance capacity (ORAC)
The procedure was based on a previously reported method by Ou et al. [26], with slight modifications. Standard curve of Trolox (7,5-180 mM) and samples were diluted in phosphate buffer (10 mM, pH 7.4). Fluorescence was monitored between 100- 120 min with a microplate reader (Fluostar Omega, BMG, Ortenberg, Germany), using 485 nm excitation and 520 nm emission filters. Results calculated using the areas under the fluorescein decay curves, between the blank and the sample, and expressed as µmol Trolox Equivalents (TE) 100 g-1 sample (d.m.).
2.8. Ferric reducing antioxidant power (FRAP)
The procedure was based on a previously reported method by Benzie and Strain [27], with slight modifications. A 300 mM acetate buffer, pH 3.6 (mixing a solutions of 300 mM sodium acetate and 300 mM glacial acetic acid until pH 3.6), a 10 mM TPTZ (2,4,6-tripyridyl-triazine) solution in 40 mM HCL, and a 20 mM FeCL3.6H2O solution were prepared. The FRAP working solution was prepared by mixing the acetate buffer, TPTZ solution and FeCl3.6H2O solution in a volume ratio of 10:1:1. The absorbance was measured at 593 nm with a microplate reader (Fluostar Omega, BMG Ortenberg, Germany) using a calibration curve with FeSO4.7H2O (100-1000 µM). The results were expressed as mmol Fe Equivalents (FeE) 100 g-1 sample (d.m.).
2.9. Statistical analysis and mathematical modelling
Chemical composition and antioxidant capacity were shown using average and standard deviation, ANOVA one-way analyses were carried out to find differences between groups, and results displayed in stacked bars and box-plots graphs. In addition, prior centred and standardised data, Principal Component Analysis (PCA) and Distributed Stochastic Neighbour Embedding (t-SNE) were used to visualise the chemical composition profiles of the grains, and Pearson correlation coefficients were performed to elucidate the relationships among chemical variables and antioxidant biomarkers. All statistical analyses were performed with both R (R: The R Project for Statistical Computing (r-project.org) and Python (Welcome to Python.org) software packages.
Modelling of the antioxidant capacity of the grains based on their proximal composition was performed with the RJAGS package (R Project, rjags: Bayesian Graphical Models using MCMC, 2023). Models were tested using scikit-learn packages from Python and lme4, nlme and e1071 libraries from R.
3. Results and Discussion
3.1. Compositional characteristics of the native flours
Proximal analysis (protein, fat and fiber) was evaluated in all samples as it has been suggested that compositional variations can affect the antioxidant capacity of grains [28,29] (Figure 2 and Figure 3).
As it was expected grain showed a wide variation in protein content; values ranged from 6 to 40 g 100 g-1 (Figure 2 and Figure 3I). The lowest values corresponded to cereals, being corn and rice where lowest protein content appeared and pulses where highest values were observed. These results are within the range of those reported by USDA food database [30], which reports cereal protein values in the range of 6.69-7.5 g 100 g-1 for rice, 3.0-16.7 g 100 g-1 for wheat, 10.1-15.1 g 100 g-1 for oat or 5.43-6.67 g 100 g-1 for corn. Pulses had higher protein content than cereals ranging 14.50-23.97 g 100 g-1 for chickpea or 22.19-25.13 g 100 g-1 for lentil. As it was expected soy was the grain with highest protein content (34.06-44.38 g 100 g-1) (Figure 2 and Figure 3I). Cereals had higher differences in protein content associated to variety factor, as compared to pulses. Rice was the grain where major variability due to variety was observed.
Protein content and profile differ with species, variety, genotype, and environmental and fertilization conditions, among others factors [31,32]. The amount of protein is quite important from a nutritional point of view, but, the protein profile and the ratio of secondary structure are relevant regarding thermal stability after process and in their later digestibility [33]. For example, grains with higher β-sheets content are easy-to-cook versus normal grains [34]. The reason is that temperature alters protein structure affecting disulphide and hydrogen linkages resulting in reorganization of protein structure and becoming more or less accessible during protein digestibility. These changes can affect to their antioxidant properties, since many of the protein fragments produced during digestion exhibit antioxidant ability, as previously demonstrated in oats [35], rye [36], wheat [37], buckwheat, rice, corn [38], soybean, lentil and chickpeas [39,40,41] among other grains.
Fat was also evaluated in all the grains (Figure 2 and Figure 3II), as variations in this fraction can affect the antioxidant status of cereals and pulses [42,43]. The content ranged from 2 to 5 g 100 g-1 in cereal, similar to previous results by other authors, which reported 1.9 g 100 g-1 for rice, 2.0 g 100 g-1 for wheat, 3.9 for g 100 g-1 corn and 3.3 for g 100 g-1 sorghum [44]. Pulses as lentil, pea and chickpea showed content from 1-4 g 100 g-1 and soy showed, as it occurred also in protein, the highest values ranging from 31 to 38 g 100 g-1. Oatmeal, chickpea and soy were the grains where the effect of variety produced a higher dispersion of results in fat content.
Total dietary fiber (TDF) has been strongly related to food antioxidant capacity [45,46]. TDF according to European Food Safety Authority (EFSA), contains non-digestible carbohydrates and lignin, non-starch polysaccharides (cellulose, hemicelluloses, pectins and hydrocolloids such as β-glucans), resistant oligosaccharides (fructo-oligosaccharides and galacto-oligosaccharides) other resistant oligosaccharides and resistant starch (RS). The lowest values were observed in corn, where ranged from 3.49 to 10.85 g 100 g-1, and the highest in soy with values from 31.60 to 37 g 100 g-1, the results agreed with TDF reported for grains [47]. The content in TDF was strongly affected by the variety studied, especially in the case of corn, oatmeal and rice where the variability associated was higher.
3.2. Colorimeter analysis characteristics of the native flours
Colour of the grain samples was evaluated and CIELAB parameters (L*, a* and b*, Figure 4 I, II and III) obtained. The colour was measured since it is a quick assessment that can show a certain significant (p≤0.05) association to antioxidant activity [48]. The type of grain was statistically significant (p≤0.05) on L values; the darkest values were observed in sorghum, meanwhile the highest lightness was observed in corn grains. The highest a* values (intensity of red colour) appeared in corn and sorghum samples, while the grains with lowest a* values (lowest intensity of red) was observed in lentil, rye and soy. Significant differences (p≤0.05) were observed when considering the b* parameter (blueness-yellowness). Positive b* values were observed in all samples indicating a yellow component in all the cases, the highest values were observed for corn, chickpea and soy, and the lowest b* values (lowest yellowness) were obtained in the case of rye, rice and sorghum. The parameter a* was highly dispersed depending on the variety in the case of corn and sorghum, meanwhile in b* value, corn and rice were the two grains that showed a higher variability in yellowness as related to variety.
3.3. Total phenol (TP) content of the native flours
Phenolic compounds are secondary metabolites linked with the colour and antioxidant activities of grains. The redox potentials of the phenolic compounds are the key factors for their antioxidant properties. Total phenol (TP) content was evaluated in all the samples (Figure 5). The values ranged from 25 mg GAE 100 g-1 in rice to 250 mg GAE 100 g-1 in lentil, where highest values were observed.
Lentil is a legume with a high level of phenol compounds and high antioxidant activity, as it has been described previously by different authors [49,50]. Soy was the second grain with higher TP content (120 mg GAE 100 g-1), significantly higher than sorghum (90 mg GAE 100 g-1), which was the cereal with highest TP content. The lowest TP values appeared in three of the cereal studied (rice, corn and wheat) and chickpea. The effect of variety on TP was more significant in the case of sorghum, lentil, pea and oatmeal. In the case of triticale, rye, wheat, corn and chickpea, the TP content showed a lower dispersion associated to variety.
3.4. Oxygen radical absorbance capacity (ORAC) of the native flours
The relevance of ORAC methodology for antioxidant activity to in vivo conditions is based on the free radical source used (peroxyl), which shows the highest prevalence in human biology [51]. ORAC assay was used to evaluate the ability of the 136 varieties of cereal and pulse grains studied to scavenge peroxyl free radicals; the values ranged from 437 to 7962 µmol Eq. Trolox 100 g-1 dry weight (Figure 6I). The pulses, lentil and soy, showed the highest ORAC values as compared to the rest of grains analysed. A significant effect of the variety was observed for the different grains, which can be associated to genetic and environmental factors as it has been observed in different studies [52]. The values observed agreed with hydrophilic ORAC values reported by the USDA [53] for these types of grains.
3.5. Ferric reducing antioxidant power assay (FRAP) of the native flours
Reducing power was also evaluated in all the samples (Figure 6II), FRAP ranged from 0.24 to 22.6 (µmol reduced iron 100 g-1). Pea was the grain where higher reducing power was observed, 15-20-fold the values of the other grains analysed. Lentil also showed high FRAP capacity, after pea, and presenting the highest differences due to variety. In contrast, a very low variability due to variety was observed in most of the grains.
3.6. Parameter correlations and distribution of parameters based on principal components analysis (PCA) and T-distributed Stochastic Neighbour Embedding (T-SNE) of native flours
The degree of correlation between antioxidant (TP, ORAC and FRAP), proximal parameters (proteins, fiber and fat) and colour parameters (L* a* and b*) was analysed. (Supplementary Figure 1). Among the methods used to evaluate the antioxidant capacity of the samples, the higher correlation was found between ORAC and total phenol content (R2 0.75), followed by protein (R2 0.72), fiber (R2 0.65) and the lowest correlation was found with fat (R2 0.51). According to the results, ORAC was a feature that explained the TAC associated not only with total phenol, but also to other compounds involved directly or indirectly with antioxidant activity, such as protein (peptides) and fiber.
A projection of the proximal composition (protein, fiber, fat) and type of grain on the principal component plane, with PC1 and PC2 on the X and Y axes, respectively, are presented in Figure 7I. PC1 explained 91% of variance, meanwhile PC2 explained 6%. PC1 and PC2 distinct two main clusters, cereals and pulses. Protein/fat and fiber showed an orthogonal spatial disposition, contributing to differentiate the grains. Fat resulted in redundancy with protein to identify the different types of grains. Protein appeared as the most useful parameter to segregate the different samples. Soy, pea, chickpea and lentil appeared clearly separated from cereals, with the exception of oatmeal. Higher fat content was the proximal composition variable that clustered oatmeal within the pulses.
Figure 7II shows the PCA for colour parameters L*a*b* and type of grains. PC1 explained 51% of the variance, while PC2 only 38%. This level of variance explanation was lower than that found in the PCA of compositional parameters. A good separation of different samples was not achieved with the colour parameters, as it can be observed in the intermeshing of the two groups of grains (cereals and pulses) and the species studied.
With the objective to find non-linear similarities in the antioxidant capacity between the different grains, t-distributed Stochastic Neighbour Embedding (T-SNE) analysis was used, with the aim of reducing initial spatial dimension and represent the observed data based on the three antioxidant parameters evaluated. Different perplexity levels were used to find the highest separation between samples, being 20 the level of perplexity where better stratification was obtained (Figure 8). Legumes lentil and soy were the two grains showing single clusters of points, appearing with relative similarity between them and clearly separated from the rest of samples. The rest of the legumes (chickpea, pea) and all cereals were distributed in groups with different levels of fragmentation. Wheat showed a wide, round area of main distribution, with some small groups of samples scattered along axis T-SNE1; wheat was clearly separated from the other cereals, except from corn. Rye also presented a homogeneous distribution; these two cereals, wheat and rye, showed similar variation in relation to axis T-SNE1 and T-SNE2. The rest of cereals showed fragmented dispersions, with projections in separated in equivalent groups, as it occurred in rice, corn, oatmeal and sorghum, or highly dependent on the T-SNE1 axis, as in the case of triticale.
3.7. Effect of thermal treatment on total antioxidant capacity (TAC)
Baked goods require a thermal process prior to consumption; the thermal treatment conditions (time and temperature) will determine the final antioxidant capacity of the product, through the degradation of thermally unstable polyphenol compounds and formation of the novo antioxidants from the polymerization of Maillard reaction products [54,55,56]. Baked good models (biscuits) were used for the kinetic study to evaluate the effect of temperature on the TAC. The types of grains wheat and rye were selected for model development and training from their homogenous projection in the Stochastic Neighbour Embedding (T-SNE) distribution analysis; regarding varieties, Berdun (wheat) and Loretto, Igor and Teodor (rye) were selected due to their distribution of antioxidant data closer to a normal distribution, as compared with the rest of varieties.
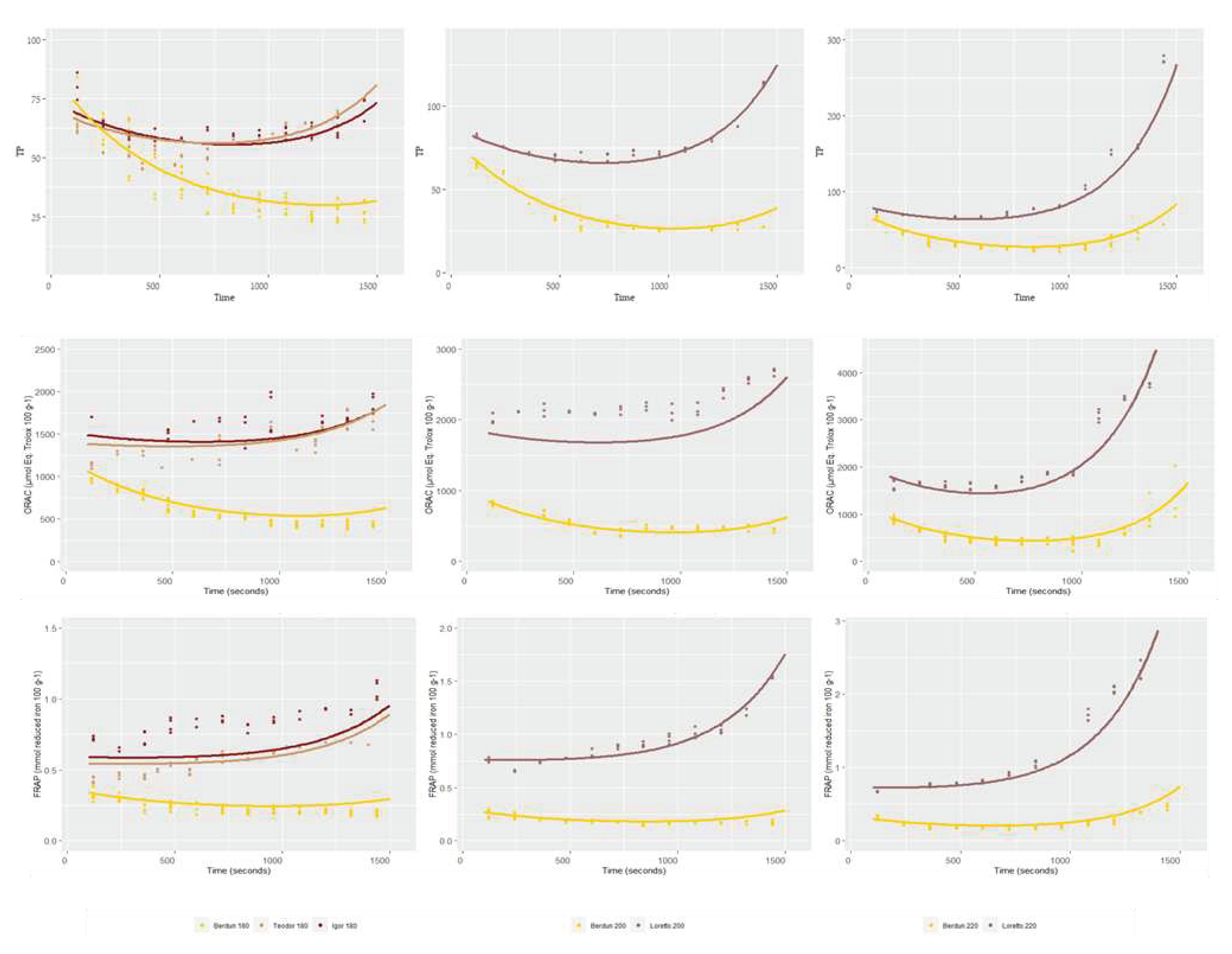
Figure 9 shows the total antioxidant capacity of model baked products (biscuits) prepared using wheat and rye flours. TP content was affected by the baking thermal process (Figure 9I). It is observed an initial decrease in the TP content, regardless of the type of flour used (wheat/rye), with increasing temperatures, until approximately 800 s.
After this time, an increase in TP is observed in wheat biscuits baked at 220 °C, and in rye biscuits 180, 200 and 220 °C. At the end of baking process (1500 s), wheat TP values were similar or lower the initial values, while significant increases after thermal processing were observed in the case of rye samples treated at 200 and 220 °C. Rye samples treated al 180 °C did not show significant variation after the baking process. TP methodology based on Folin Ciocalteu reagent can reflect not only phenolic content but also the presence of other antioxidants [57]. Rye TP content showed lower sensitivity to temperature, as compared to wheat. This behaviour could be explained by a faster degradation of thermolabile antioxidants, in the case of wheat, or a significantly higher formation of Maillard reaction products (MRPs), which in the case of rye, would be favoured by its higher content in lysine, as compared to wheat [58]. Previously, it has been suggested a correlation of acrylamide formation an increment in antioxidant capacity in wheat and rye bread models [59], suggesting health-related benefits with low temperature-long-time treatments.
On the other hand, degradation of acrylamide may also occur in high temperature treatments [60]. On the other hand, final MRPs, melanoidins have shown biological activities and could be considered as antioxidant fiber [60,61].
A similar behaviour of the antioxidant capacity measure against peroxyl radicals (ORAC) was observed, as compared to TP results (Figure 9II). The antioxidant capacity of wheat samples decreased in the initial stages of the thermal treatment. After 1000 seconds of treatment, the antioxidant capacity of all samples (wheat and rye) treated at higher temperature (200 °C and 220 °C) increased, and in the case of the rye samples, the levels reached (2600-3800 µmol Eq. Trolox 100 g-1) were significantly higher than the initial levels (1500-2000 µmol Eq. Trolox 100 g-1). The gradual formation during thermal process of high molecular weight melanoidins and binding of low molecular weight MRPs to melanoidin skeletons, can contribute to the final antioxidant activity of the samples. These compounds react with the peroxyl radicals and are measured by the ORAC assay, as it happened in the case of Folin Ciocalteu reagent in the TP method [62].
The ferric reducing ability (FRAP values) of the rye was significantly higher than the wheat (Figure 9III). As it was observed in TP and ORAC assays, a decrease in FRAP values was observed for wheat samples up to 800 s of treatment. After this period, the FRAP values increased in a temperature dependent manner. Samples treated at 220 °C showed the highest final ferric reducing values (FRAP) values. This behaviour again could be associated to the presence of melanoidins, which have been reported among their biological activities significant ferric reducing capacity [63].
3.8. Modelling of the effect of thermal treatment on total antioxidant capacity (TAC).
Previous to model building, experimental data (Figure 9) were assumed to follow a function with two exponential parts: a negative exponential simulating antioxidant compound degradation and a positive exponential simulating de novo synthesis of antioxidant compounds (Equation 1, ‘t’ represents time, ‘T’ temperature and antioxidant capacity as TP, ORAC or FRAP).
As a first exploratory search of space of hyperparameters (θ, α1, α2, β1, β2), iterative optimization tools were used (R packages “nlme” and “nls”). Parameters β1 and β2 were fixed at 0.08 and 0.12 respectively, due to their low influence on curve shape. Number of hyperparameters was reduced to 3 (ϴ, α1 and α2), simplifying the solution search space and facilitating convergence in the search for an optimal solution (Equation 2).
First level of modelling
As each of the 7 available curves could be clustered based on factors type of grain and temperature, a hierarchical model was selected. A Bayesian hierarchical model was used to determine the values of ϴ, α1 and α2 parameters for each seven combination of temperature and variety, which uses the baking temperature, time and type of grain as predictors. After that, the median value of MonteCarlo distributions obtained in the Bayesian training, for each configuration (variety, temperature), was obtained (Table 1).
With the use of the Bayesian hierarchical model, the parameters (ϴ, α1 and α2) of the double exponential curve were obtained; best fitting curves obtained are shown in Figure 10.
Second level of modelling
Using as reference the optimal parameters ('ϴ, 'α1' and 'α2') fitted with the Bayesian hierarchical model, a training procedure was carried out using as input data the compositional (protein and fiber), type of grain (wheat or rye) and temperature (180, 200 or 220 °C). Independent training steps using Support Vector Regression (SVR) for each combination of parameters ('ϴ, 'α1' and 'α2') and antioxidant value (TP, ORAC and FRAP) were carried out. Datasets used for training of the different models are provided as Supplementary Tables S1, S2 and S3. Corresponding cost parameter and R2 obtained for SVR model training can be found in Supplementary Table Supplementary 4.
Figure 11.
Curves of the level of TP, ORAC and FRAP predicted with the second level models for the series according to baking temperature.
Figure 11.
Curves of the level of TP, ORAC and FRAP predicted with the second level models for the series according to baking temperature.
Using the obtained SVR models, it is possible to predict the antioxidant capacity (TP, ORAC and FRAP) at any given thermal processing temperature and time, protein and fiber as proximal parameters, and wheat or rye as type of grain (Figure 12).
Therefore, relying solely on the secondary models to determine the shape of the curve to be predicted, that is, using baking temperature, grain type, protein, and fiber as predictors, the fit shown below was achieved (Table 2, Figure 11), where it should be noted that they are accompanied by graphs in which the values of the residuals are represented and that, except for the predictions of Teodor and Igor at 180ºC, all the models manage to explain more than 80% of the variance present in the data.
Sensitivity of modelling
In order to estimate model stability, the variability of the predictions after different entry values was analysed. Temperature and proximal composition (protein, fiber) independent variations (maintaining the rest of variables fixed) were used as inputs for the model and compared with model outcome using available values of temperature and composition as reference. Figure 13 shows predicted behaviour of rye and wheat at a range of temperatures from 175 to 225 °C, at 5 °C intervals. As it can be observed, the model is more sensitive for temperature changes at advanced treatment times, with a linear relationship of temperature and curve slope, with lower variation at the initial stages of the thermal process; there was a higher variability of the final TP values in the case of rye product, as compared with wheat (Supplementary Figures 3-5), revealing a more critical importance in the first case of the temperature, as compared with the second, when selecting a treatment temperature with the objective of obtaining a certain TP final value.
3. Conclusions
The total antioxidant capacity profiles of a range of cereal and legume flours, as evaluated through three different methods (TP, ORAC and FRAP), and its distribution variability related to proximal composition and colour was reported. The distribution of the variability of the antioxidant data was significantly affected by the protein and fiber composition of the samples. The variability b Legume flours showed in general higher values of antioxidant capacity, as compared to cereals, with special relevance of lentil flour. Reduced dimensionality techniques (PCA and T-SNE) applied to the data showed wheat and rye as two samples with homogeneous distribution and well separated.
The behaviour of the antioxidant capacity profiles of the baked products suffered a decrease over the first half of the treatment times, with a subsequent increase afterwards. A predictive model for estimation of the TAC of baked products, using Bayesian and SVR modelling tools, and considering protein and fiber content was developed. A double exponential parameterised curve that reflects the initial decrease and subsequent increase as a consequence of the processes of degradation and generation of the antioxidant compounds is proposed. An estimation of the antioxidant capacity of a baked products during its thermal process has been achieved; this model is intended to be improved with further types of grains and varieties in order to create a tool which may result of interest to the food industry; this tool would estimate optimal processing conditions in order to maximise antioxidant capacity of baked products, using as input values type of grain, protein and fiber content, and temperature and time of process.
Acknowledgments
The authors greatly acknowledge to the PEA team (Gabriel Villamayor, Rosa Fernandez, Jose Ramon Valles, Jose Clementino Prieto and Eduardo Aguado) for providing all plant material with trazability in harvest and postharvest for species and varieties for the study and EU-FEDER project FOODTOOL, PEP: 2020/712 for the financial support.
References
- Sajdakowska, M.; Gębski, J.; Jeżewska-Zychowicz, M.; Królak, M. Consumer Choices in the Bread Market: The Importance of Fiber in Consumer Decisions. Nutrients 2021, 13, 132. [Google Scholar] [CrossRef]
- Anderson, J; Hanna, T. Whole grains and protection against coronary heart disease: what are the active components and mechanisms? American Journal of Clinical Nutrition. 1999, 70, 307–308. [Google Scholar] [CrossRef]
- Anderson, JW; Hanna, TJ; Peng, X; Kryscio, RJ. Whole grain foods and heart disease risk. Journal of the American College of Nutrition. 2000, 19 Suppl. 3, 291S–299S. [Google Scholar] [CrossRef]
- Bourdon, I; Yokoyama, W; Davis, P; Hudson, C; Backus, R; Richter, D; Knuckles, B; Schneeman, BO. Postprandial lipid, glucose, insulin and cholecystokinin responses in men fed barley pasta enriched with beta-glucan. American Journal of Clinical Nutrition. 1999, 69, 55–63. [Google Scholar] [CrossRef]
- Slavin, J; Jacobs, D; Marquart, L. Whole-grain consumption and chronic disease: protective mechanisms. Nutrition in Cancer. 1997, 27, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Martín-Diana, AB; García-Casas, MJ; Martínez-Villaluenga, C; Frías, J; Peñas, E; Rico, D. Wheat and Oat Brans as Sources of Polyphenol Compounds for Development of Antioxidant Nutraceutical Ingredients. Foods. 2021, 10, 115. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Vodnar, D.C. Whole Grains and Phenolic Acids: A Review on Bioactivity, Functionality, Health Benefits and Bioavailability. Nutrients 2018, 10, 1615. [Google Scholar] [CrossRef]
- Available online: https://www.sphericalinsights.com/reports/europe-phenolic-antioxidant-market.
- Pruteanu, L. L.; Bailey, D. S.; Grădinaru, A. C.; Jäntschi, L. The Biochemistry and Effectiveness of Antioxidants in Food, Fruits, and Marine Algae. Antioxidants. 2023, 12, 860. [Google Scholar] [CrossRef]
- Pellegrini, N.; Vitaglione, P.; Granato, D.; Fogliano, V. Twenty-five years of total antioxidant capacity measurement of foods and biological fluids: merits and limitations. Journal of the science of food and agriculture. 2020, 100, 5064–5078. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R. L. The chemistry behind antioxidant capacity assays. Journal of agricultural and food chemistry. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J. A.; Deemer, E. K. Development and validation of oxygen radical absorbance capacity assay for lipophilic antioxidants using randomly methylated cyclodextrin as the solubility enhancer. J. Agric. Food Chem. 2002, 50, 1815–1821. [Google Scholar] [CrossRef] [PubMed]
- Niki, E. Lipid peroxidation: physiological levels and dual biological effects. Free radical biology & medicine. 2009, 47, 469–484. [Google Scholar] [CrossRef]
- Vanegas, S.M.; Meydani, M.; Barnett, J.B.; Goldin, B.; Kane, A.; Rasmussen, H.; Brown, C.; Vangay, P.; Knights, D.; Jonnalagadda, S.; et al. Substituting whole grains for refined grains in a 6-wk randomised trial has a modest effect on gut microbiota and immune and inflammatory markers of healthy adults. Am. J. Clin. Nutr. 2017, 105, 635–650. [Google Scholar] [CrossRef]
- Björck, I.; Östman, E.; Kristensen, M.; Mateo Anson, N.; Price, R.K.; Haenen, G.R.M.M.; Havenaar, R.; Bach Knudsen, K.E.; Frid, A.; Mykkänen, H.; et al. Cereal grains for nutrition and health benefits: Overview of results from in vitro, animal and human studies in the HEALTHGRAIN project. Trends Food Sci. Technol. 2012, 25, 87–100. [Google Scholar] [CrossRef]
- García-Castro, A.; Román-Gutiérrez, A.D.; Castañeda-Ovando, A.; Cariño-Cortés, R.; Acevedo-Sandoval, O.A.; López-Perea, P.; Guzmán-Ortiz, F.A. Cereals as a Source of Bioactive Compounds with Anti-Hypertensive Activity and Their Intake in Times of COVID-19. Foods 2022, 11, 3231. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Tsiaka, T; Kritsi, E; Tsiantas, K; Christodoulou, P; Sinanoglou, VJ; Zoumpoulakis, P. Design and Development of Novel Nutraceuticals: Current Trends and Methodologies. Nutraceuticals. 2022, 2, 71–90. [Google Scholar] [CrossRef]
- Halvorsen, B.L.; Holte, K.; Myhrstad, M.C.; Barikmo, I.; Hvattum, E.; Remberg, S.F.; Wold, A.B.; Haffner, K.; Baugerød, H.; Andersen, L.F.; et al. A systematic screening of total antioxidants in dietary plants. J. Nutr. 2002, 132, 461–471. [Google Scholar] [CrossRef]
- Halvorsen, B.L.; Carlsen, M.H.; Phillips, K.M.; Bøhn, S.K.; Holte, K.; Jacobs, D.R., Jr.; Blomhoff, R. Content of redox-active compounds (ie, antioxidants) in foods consumed in the United States. Am. J. Clin. Nutr. 2006, 84, 95–135. [Google Scholar] [CrossRef]
- Haytowitz, DB.; Bhagwat, S. USDA Database for the Oxygen Radical Absorbance Capacity (ORAC) of Selected Foods, Release 2. United States Department of Agriculture 2010. Available online: http://www.orac-info-portal.de/download/ORAC_R2.pdf.
- Takebayashi, J.; Oki, T.; Watanabe, J.; Yamasaki, K.; Chen, J.; Sato-Furukawa, M.; Tsubota-Utsugi, M.; Taku, K.; Goto, K.; Matsumoto, T.; et al. Hydrophilic antioxidant capacities of vegetables and fruits commonly consumed in Japan and estimated average daily intake of hydrophilic antioxidants from these foods. J. Food Comp. Anal. 2013, 29, 25–31. [Google Scholar] [CrossRef]
- Zhiming, Li; Kaiying, Nie; Zhaojing, Wang. Quantitative Structure Activity Relationship Models for the Antioxidant Activity of Polysaccharides; Department of Bioengineering and Biotechnology, Huaqiao University: China, 2016. [Google Scholar]
- AOAC. Methods 990.03, 2003.05, 985.29 & 923.03. In Official Methods of Analysis of AOAC International, 18th ed.; AOAC: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Slinkard, K.; Singleton, V.L. Total phenol analyses: Automation and comparison with manual methods. Am. J. Enol. Viticult. 1977, 28, 49–55. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Badarinath, A.V.; Rao, K.M.; Chetty, C.M.S.; Ramkanth, S.; Rajan, T.V.S. Review on In-vitro Antioxidant Methods: Comparisions, Correlations and Considerations. Inter. J. PharmTech Res. 2010, 2, 1276–1285. [Google Scholar]
- Nile, S.H.; Khobragade, C.N.; Park, S.W. Optimized and Comparative Antioxidant Assays and Its Applications in Herbal and Synthetic Drug Analysis as an Antioxidants. Mini-Rev. Med. Chem. 2012, 12, 1007–1014. [Google Scholar] [CrossRef]
- Available online: https://fdc.nal.usda.gov/.
- Singh, N. Pulses: an overview. J Food Sci Technol. 2017, 54, 853–857. [Google Scholar] [CrossRef]
- Singh, B; Singh, JP; Shevkani, K; Singh, N; Kaur, A. Bioactive constituents in pulses and their health benefits. J Food Sci Technol. 2017, 54, 858–870. [Google Scholar] [CrossRef] [PubMed]
- Gulati, P; Li, A; Holding, D; Santra, D; Zhang, Y; Rose, DJ. Heating Reduces Proso Millet Protein Digestibility via Formation of Hydrophobic Aggregates. J Agric Food Chem. 2017, 65, 1952–1959. [Google Scholar] [CrossRef] [PubMed]
- Parmar, N; Singh, N; Kaur, A; Virdi, AS; Shevkani, K. Protein and microstructure evaluation of harder-to-cook and easy-to-cook grains from different kidney bean accessions. LWT-Food Sci Technol. 2017, 79, 487–495. [Google Scholar] [CrossRef]
- Du, Y.; Esfandi, R.; Willmore, W.G.; Tsopmo, A. Antioxidant activity of oat proteins derived peptides in stressed hepatic HepG2 cells. Antioxidants. 2016, 5, 39. [Google Scholar] [CrossRef] [PubMed]
- Leung, R.; Venus, C.; Zeng, T.; Tsopmo, A. Structure-function relationships of hydroxyl radical scavenging and chromium-VI reducing cysteine-tripeptides derived from rye secalin. Food Chem. 2018, 254, 165–169. [Google Scholar] [CrossRef]
- Chen, S.; Lin, D.; Gao, Y.; Cao, X.; Shen, X. A novel antioxidant peptide derived from wheat germ prevents high glucose-induced oxidative stress in vascular smooth muscle cells in vitro. Food Funct. 2017, 8, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Raychaudhuri, U.; Chakraborty, R. Cereal based functional food of Indian subcontinent: a review. J Food Sci Technol. 2012, 49, 665–72. [Google Scholar] [CrossRef] [PubMed]
- Torres-Fuentes, C.; Contreras, M.; Recio, I.; Alaiz, M.; Vioque, J. Identification and characterization of antioxidant peptides from chickpea protein hydrolysates. Food Chem. 2015, 180, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Du, M.; Shen, M.; Wu, T.; Lin, L. Physico-chemical properties, antioxidant activities and angiotensin-I converting enzyme inhibitory of protein hydrolysates from Mung bean (Vigna radiate). Food Chem. 2019, 270, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; He, H.; Xiang, J.; Li, B.; Zhao, M.; Hou, T. Selenium-containing soybean antioxidant peptides: Preparation and comprehensive comparison of different selenium supplements. Food Chem. 2021, 358, 129888. [Google Scholar] [CrossRef]
- Konopka, I.; Grabiński, J.; Skrajda, M.; Dąbrowski, G.; Tańska, M.; Podolska, G. Variation of wheat grain lipid fraction and its antioxidative status under the impact of delayed sowing. Journal of Cereal Science 2017, 76, 56–63. [Google Scholar] [CrossRef]
- Ciudad-Mulero, M.; Matallana-González, M. C.; Cámara, M.; Fernández-Ruiz, V.; Morales, P. Antioxidant Phytochemicals in Pulses and their Relation to Human Health: A Review. Current pharmaceutical design. 2020, 26, 1880–1897. [Google Scholar] [CrossRef]
- Das, A.; Raychaudhuri, U.; Chakraborty, R. Cereal based functional food of Indian subcontinent: a review. J Food Sci Technol. 2012, 49, 665–72. [Google Scholar] [CrossRef]
- Núñez-Gómez, V.; González-Barrio, R.; Periago, M.J. Interaction between Dietary Fiber and Bioactive Compounds in Plant By-Products: Impact on Bioaccessibility and Bioavailability. Antioxidants. 2023, 12, 976. [Google Scholar] [CrossRef] [PubMed]
- Călinoiu, LF.; Vodnar, DC. Whole Grains and Phenolic Acids: A Review on Bioactivity, Functionality, Health Benefits and Bioavailability. Nutrients. 2018, 10, 1615. [Google Scholar] [CrossRef]
- Tosh, SM.; Farnworth, ER.; Brummer, Y.; Duncan, AM.; Wright, AJ.; Boye, JI.; Marcotte, M.; Benali, M. Nutritional Profile and Carbohydrate Characterization of Spray-Dried Lentil, Pea and Chickpea Ingredients. Foods. 2013, 2, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Muzolf-Panek, M.; Wa´skiewicz, A. Relationship between Phenolic Compounds, Antioxidant Activity and Color Parameters of Red Table Grape Skins Using Linear Ordering Analysis. Appl. Sci. 2022, 12, 6146. [Google Scholar] [CrossRef]
- Rico, D.; Peñas, E.; Del Carmen García, M.; Rai, D.K.; Martínez-Villaluenga, C.; Frias, J.; Martín-Diana, A.B. Development of Antioxidant and Nutritious Lentil (Lens culinaris) Flour Using Controlled Optimized Germination as a Bioprocess. Foods 2021, 10, 2924. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, Y.; Dueñas, M.; Estrella, I.; Hernández, T.; Benitez, V.; Esteban, R.M.; Martín-Cabrejas, M.A. Evaluation of Phenolic Profile and Antioxidant Properties of Pardina Lentil as Affected by Industrial Dehydration. Journal of Agricultural and Food Chemistry 2010, 58, 10101–10108. [Google Scholar] [CrossRef]
- Prior, R.L. Oxygen radical absorbance capacity (ORAC): New horizons in relating dietary antioxidants/bioactives and health benefits. Journal of Functional Foods. 2015, 18, 797–810. [Google Scholar] [CrossRef]
- Mpofu, A.; Sapirstein, H.D.; Beta, T. Genotype and environmental variation in phenolic content, phenolic acid composition, and antioxidant activity of hard spring wheat. J. Agric. Food Chem. 2006, 54, 1265–1270. [Google Scholar] [CrossRef]
- Available online: http://www.ars.usda.gov/nutrientdata.
- Sablani, S.S.; Marcotte, M.; Baik, O.D.; Castaigne, F. Modeling of simultaneous heat and water transport in the baking process. Lebensm Wiss Technol. 1998, 31, 201–209. [Google Scholar] [CrossRef]
- Blanch, G.P.; Ruiz del Castillo, M.L. Effect of Baking Temperature on the Phenolic Content and Antioxidant Activity of Black Corn (Zea mays L.) Bread. Foods. 2021, 10, 1202. [Google Scholar] [CrossRef]
- Delgado-Andrade, C.; Rufián-Henares, J.A.; Morales, F.J. Assessing the Antioxidant Activity of Melanoidins from Coffee Brews by Different Antioxidant Methods. J. Agric. Food Chem. 2005, 53, 7832–7836. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, RL. The chemistry behind antioxidant capacity assays. J Agric Food Chem. 2005, 53, 1841–56. [Google Scholar] [CrossRef] [PubMed]
- Rani, M.; Singh, G.; Siddiqi, R.A.; Gill, B.S.; Sogi, D.S.; Bhat, M.A. Comparative Quality Evaluation of Physicochemical, Technological, and Protein Profiling of Wheat, Rye, and Barley Cereals. Frontiers in Nutrition. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Przygodzka, M.; Piskula, M.K.; Kukurová, K.; Ciesarová, Z.; Bednarikova, A.; Zieliński, H. Factors influencing acrylamide formation in rye, wheat and spelt breads. Journal of Cereal Science 2015, 65, 96–102. [Google Scholar] [CrossRef]
- Çelik, E.E.; Gökmen, V. Formation of Maillard reaction products in bread crust-like model system made of different whole cereal flours. European Food Research and Technology. 2020, 246, 1207–1218. [Google Scholar] [CrossRef]
- Saura-Calixto, F. Antioxidant dietary fiber product: a new concept and a potential food ingredient. J Agric Food Chem. 1998, 46, 4303–4306. [Google Scholar] [CrossRef]
- Liu, J.; Gan, J.; Yu, Y.; Zhu, S.; Yin, L.; Cheng, Y. Effect of laboratory-scale decoction on the antioxidative activity of Zhenjiang Aromatic Vinegar: The contribution of melanoidins. Journal of Functional Foods. 2016, 21, 75–86. [Google Scholar] [CrossRef]
- Delgado-Andrade, C.; Rufián-Henares, JA.; Morales, FJ. Assessing the antioxidant activity of melanoidins from coffee brews by different antioxidant methods. J Agric Food Chem. 2005, 53, 7832–6. [Google Scholar] [CrossRef] [PubMed]
- Froufe, H.J.C.; Abreu, R.M.V.; Ferreira, I.C.F.R. A QCAR model for predicting antioxidant activity of wild mushrooms. Escola Superior Agrária Instituto Politécnico de Bragança, Portugal 2009. [Google Scholar] [CrossRef]
Figure 1.
Biscuits prepared before and after thermal treatment (baking process) at different temperatures (180, 200 and 220 °C and times (0-1500 s).
Figure 1.
Biscuits prepared before and after thermal treatment (baking process) at different temperatures (180, 200 and 220 °C and times (0-1500 s).

Figure 2.
Stacked bar graph representing the proximal profile according to grain type. Letters of each colour denotes statistical differences between means (one-way ANOVA, post hoc Duncan’s test, p ≤ 0.05). Data was express in percentage (g 100 g-1 d.m.).
Figure 2.
Stacked bar graph representing the proximal profile according to grain type. Letters of each colour denotes statistical differences between means (one-way ANOVA, post hoc Duncan’s test, p ≤ 0.05). Data was express in percentage (g 100 g-1 d.m.).
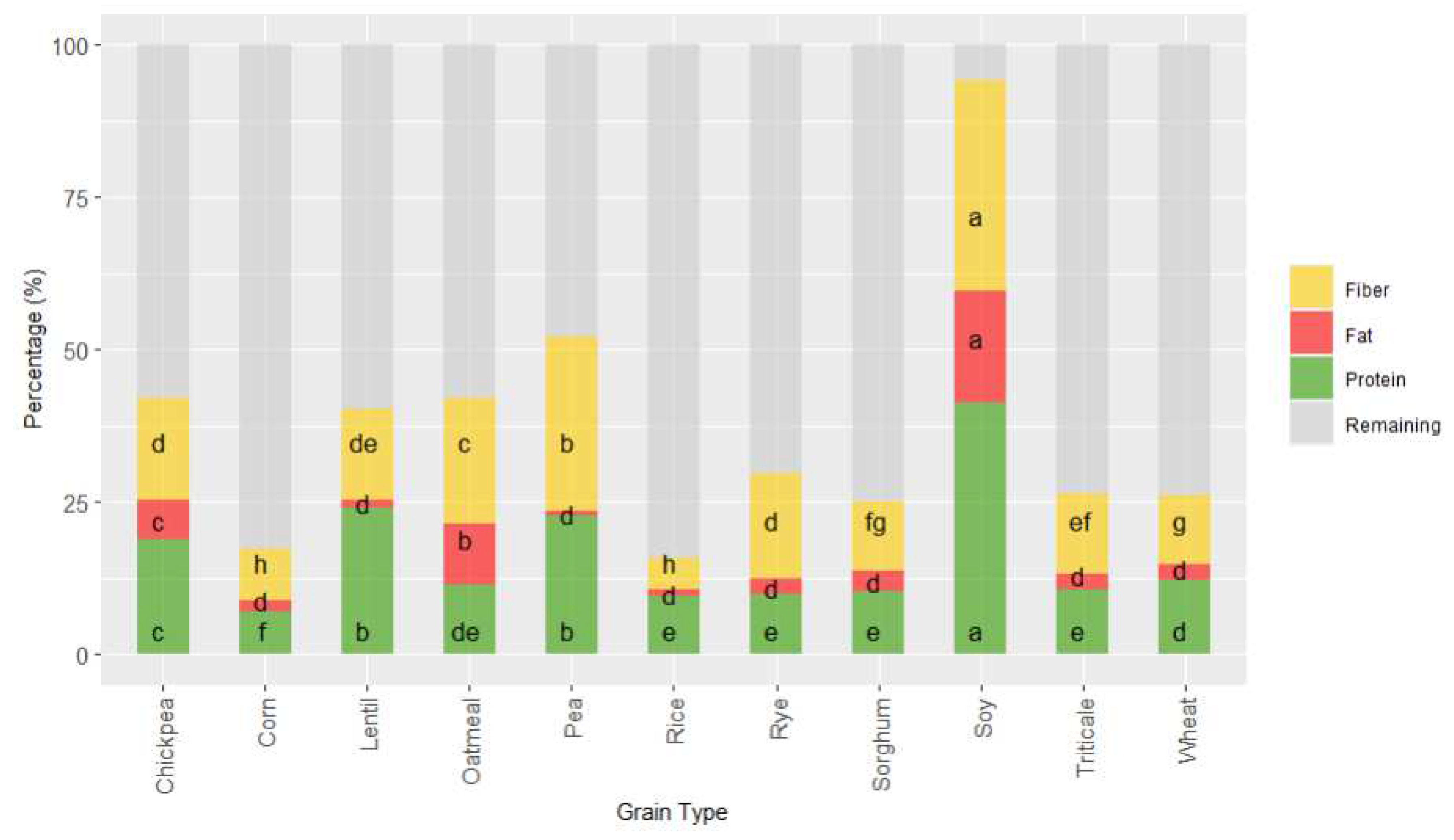
Figure 3.
Box-Plot distribution for (I) Protein, (II) Fat and (III) Fiber (Total Dietary Fiber) according to the grain. Data was express in percentage (g 100 g-1 d.m.). Letters denotes statistical differences between means (one-way ANOVA, post hoc Duncan’s test, p ≤ 0.05).
Figure 3.
Box-Plot distribution for (I) Protein, (II) Fat and (III) Fiber (Total Dietary Fiber) according to the grain. Data was express in percentage (g 100 g-1 d.m.). Letters denotes statistical differences between means (one-way ANOVA, post hoc Duncan’s test, p ≤ 0.05).
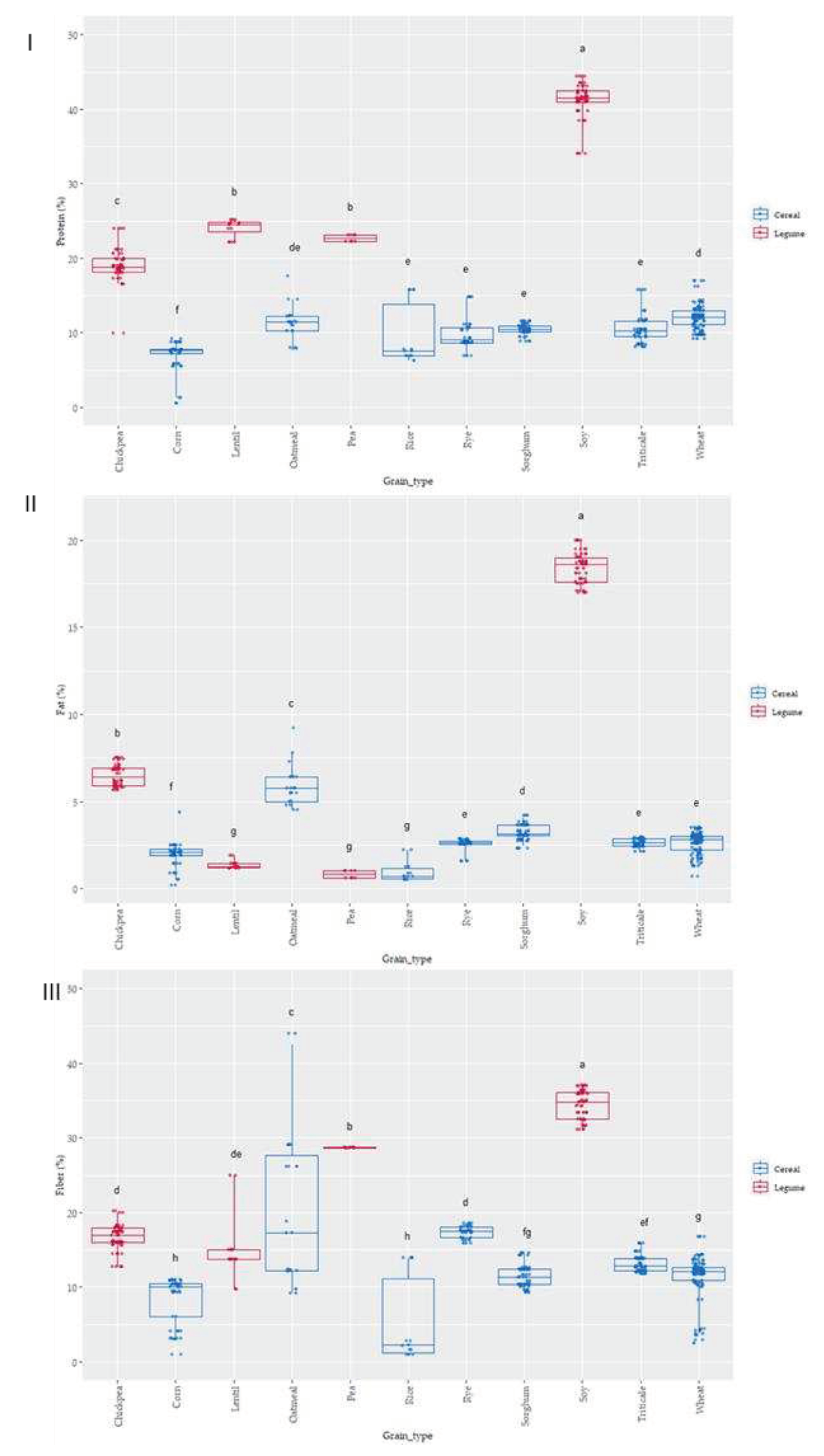
Figure 4.
Box-Plot distribution for (I) Luminosity, (II) a* and (III) b* according to the grain. Letters denotes statistical differences between means (one-way ANOVA, post hoc Duncan’s test, p ≤ 0.05).
Figure 4.
Box-Plot distribution for (I) Luminosity, (II) a* and (III) b* according to the grain. Letters denotes statistical differences between means (one-way ANOVA, post hoc Duncan’s test, p ≤ 0.05).
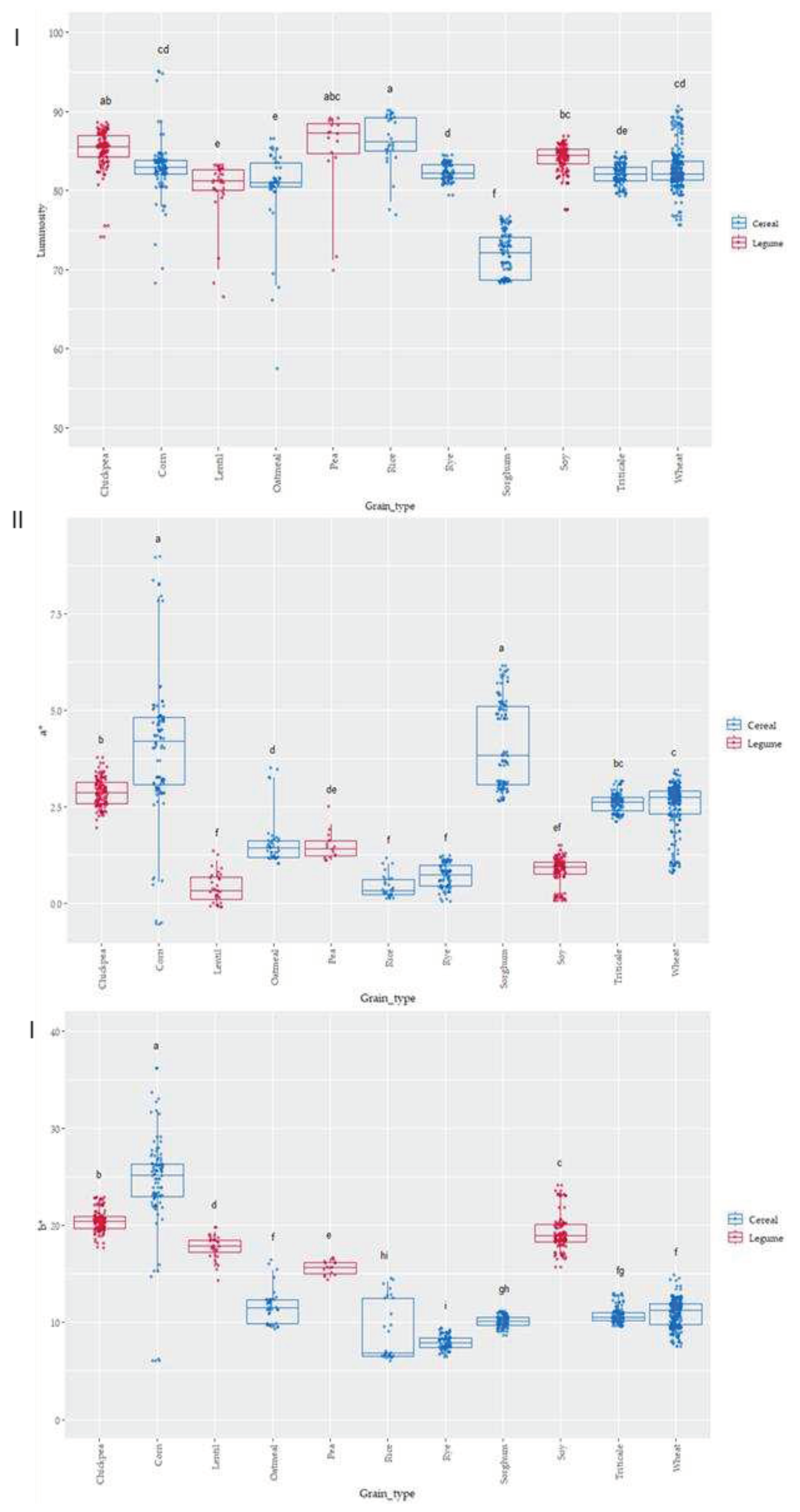
Figure 5.
Box-Plot distribution for TP (mg GAE 100 g-1) according to the grain. Data are mean values. Letters denotes statistical differences between means (one-way ANOVA, post hoc Duncan’s test, p ≤ 0.05).
Figure 5.
Box-Plot distribution for TP (mg GAE 100 g-1) according to the grain. Data are mean values. Letters denotes statistical differences between means (one-way ANOVA, post hoc Duncan’s test, p ≤ 0.05).
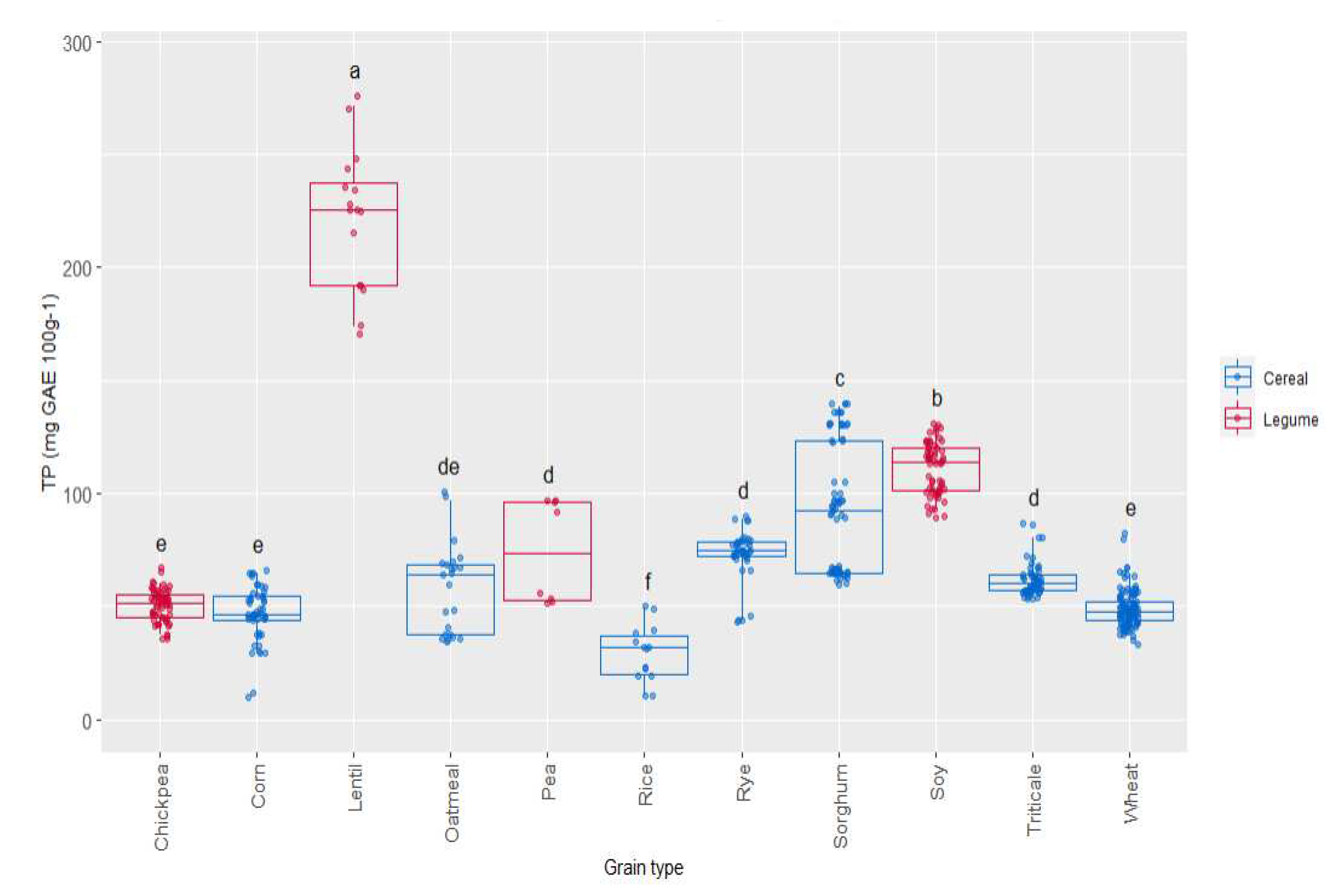
Figure 6.
Box-Plot distribution for (I) ORAC (µmol Eq. Trolox 100 g-1) and (II) FRAP (µmol reduced iron 100 g-1) according to the grain. Data are mean values. Letters denotes statistical differences between means (one-way ANOVA, post hoc Duncan’s test, p ≤ 0.05).
Figure 6.
Box-Plot distribution for (I) ORAC (µmol Eq. Trolox 100 g-1) and (II) FRAP (µmol reduced iron 100 g-1) according to the grain. Data are mean values. Letters denotes statistical differences between means (one-way ANOVA, post hoc Duncan’s test, p ≤ 0.05).
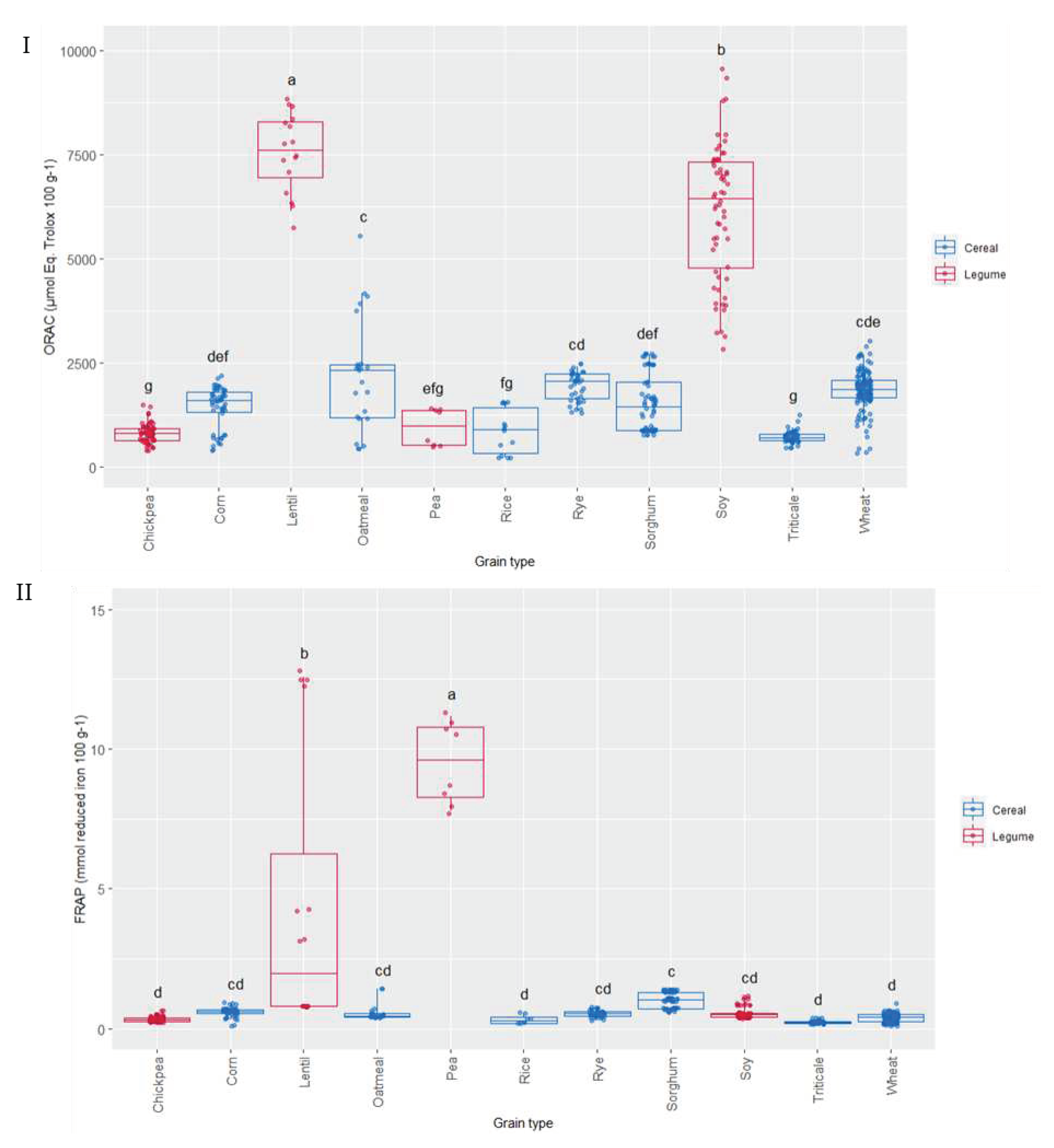
Figure 7.
Representation of the proximal profile (I) and colourimeter parameters (II) of the grain types based on principal components analysis (PCA).
Figure 7.
Representation of the proximal profile (I) and colourimeter parameters (II) of the grain types based on principal components analysis (PCA).
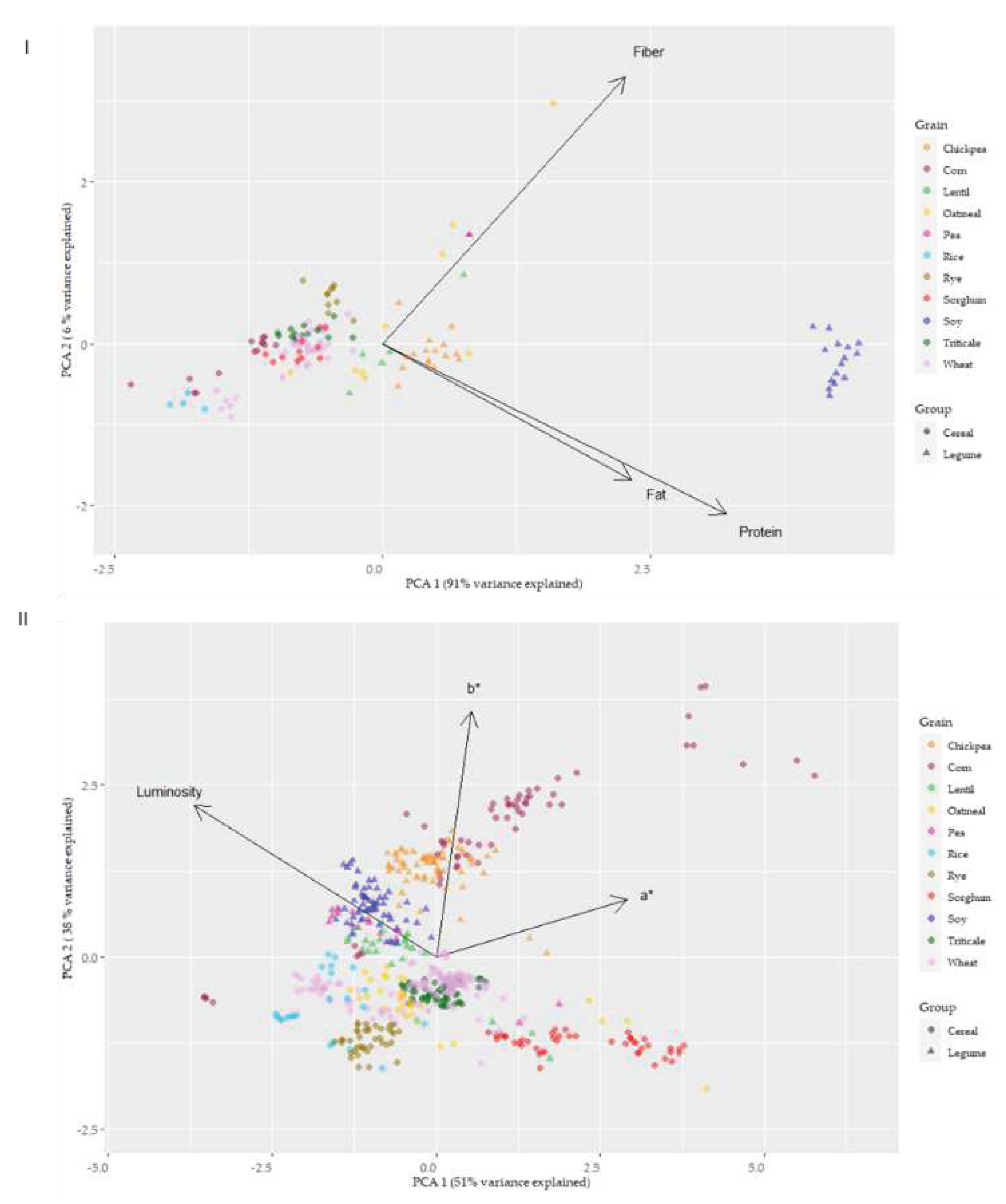
Figure 8.
Distributed Stochastic Neighbour Embedding (T-SNE) distribution of type of grains based on their antioxidant parameters (TP, ORAC and FRAP).
Figure 8.
Distributed Stochastic Neighbour Embedding (T-SNE) distribution of type of grains based on their antioxidant parameters (TP, ORAC and FRAP).
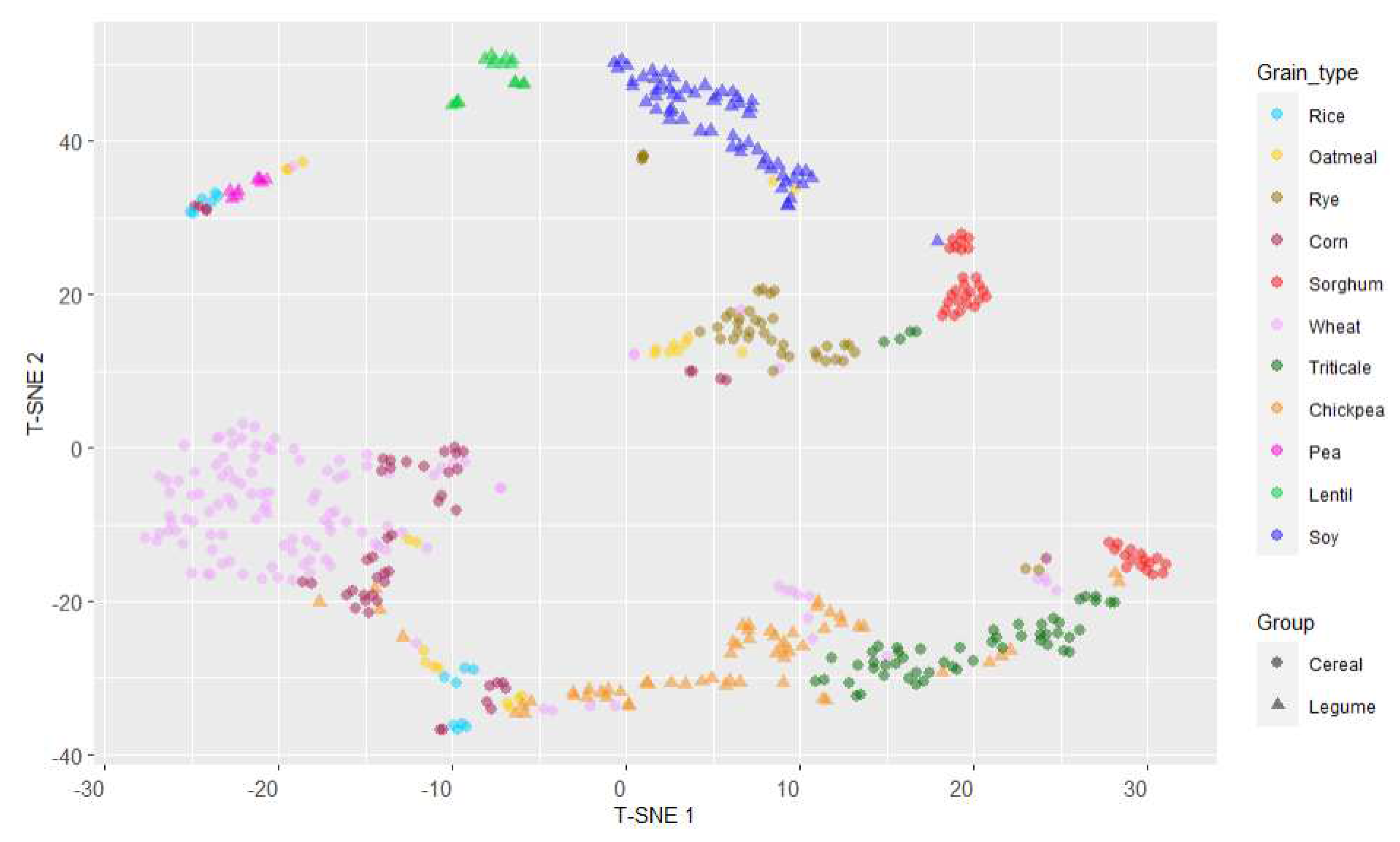
Figure 9.
Effect of temperature and time in TP (mg GAE 100 g-1), ORAC (µmol Eq. Trolox 100 g-1) and FRAP (µmol reduced iron 100 g-1) for wheat (Berdun) and rye (Teodor, Loreto e Igor).
Figure 9.
Effect of temperature and time in TP (mg GAE 100 g-1), ORAC (µmol Eq. Trolox 100 g-1) and FRAP (µmol reduced iron 100 g-1) for wheat (Berdun) and rye (Teodor, Loreto e Igor).
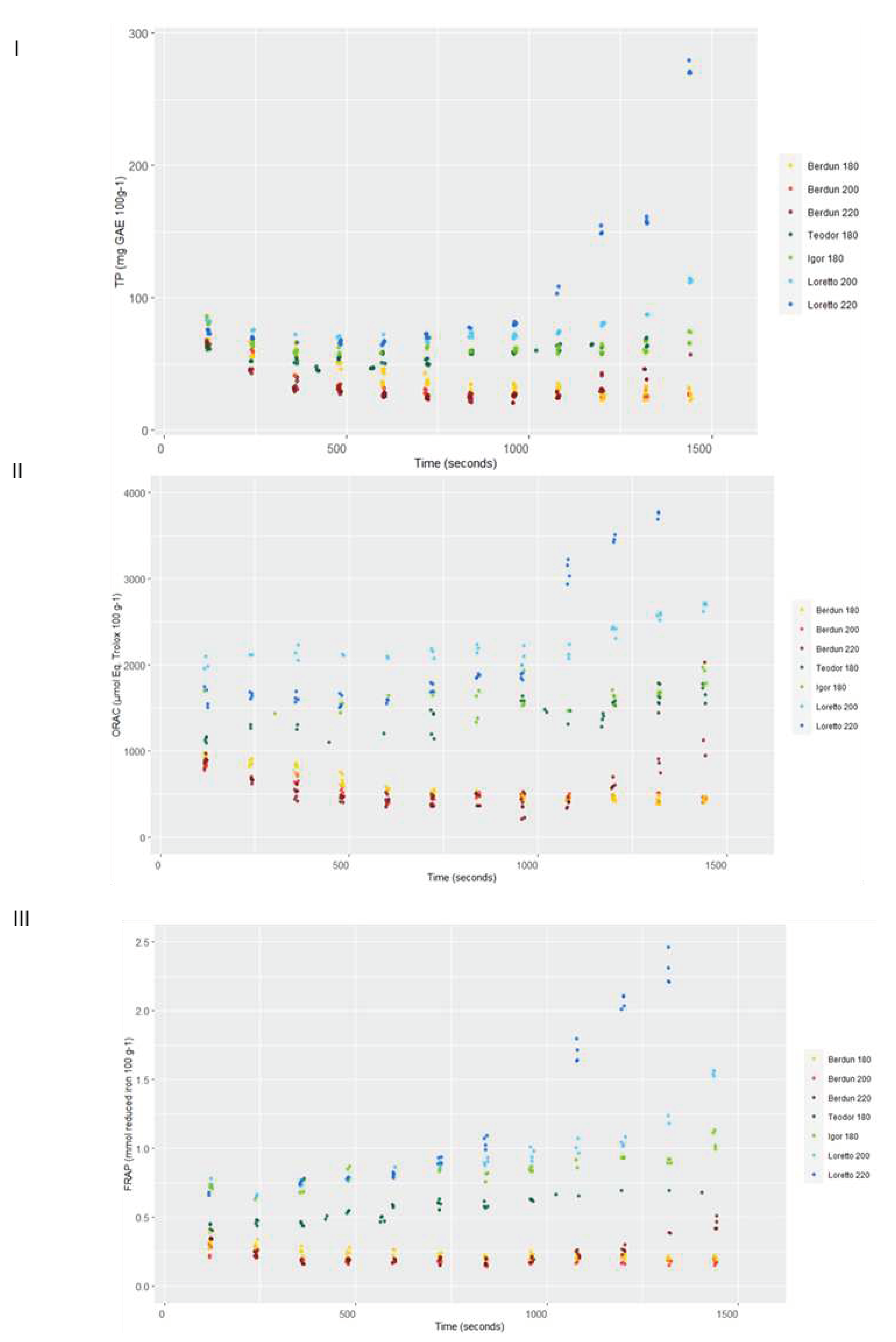
Figure 10.
Level curves of Total Phenols predicted with the first level model for the series according to baking temperature
Figure 10.
Level curves of Total Phenols predicted with the first level model for the series according to baking temperature
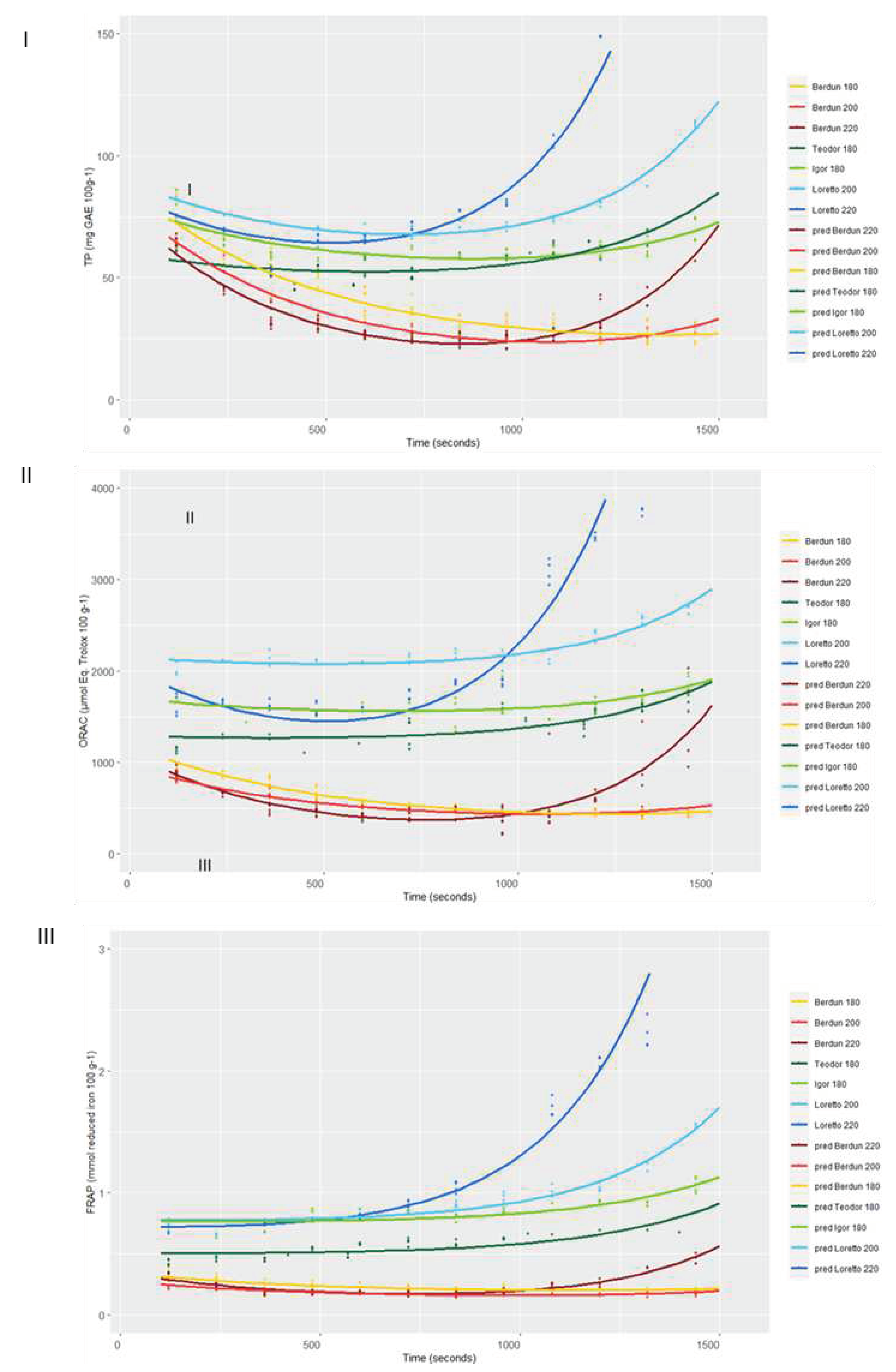
Figure 12.
Schematic representation of the global model.
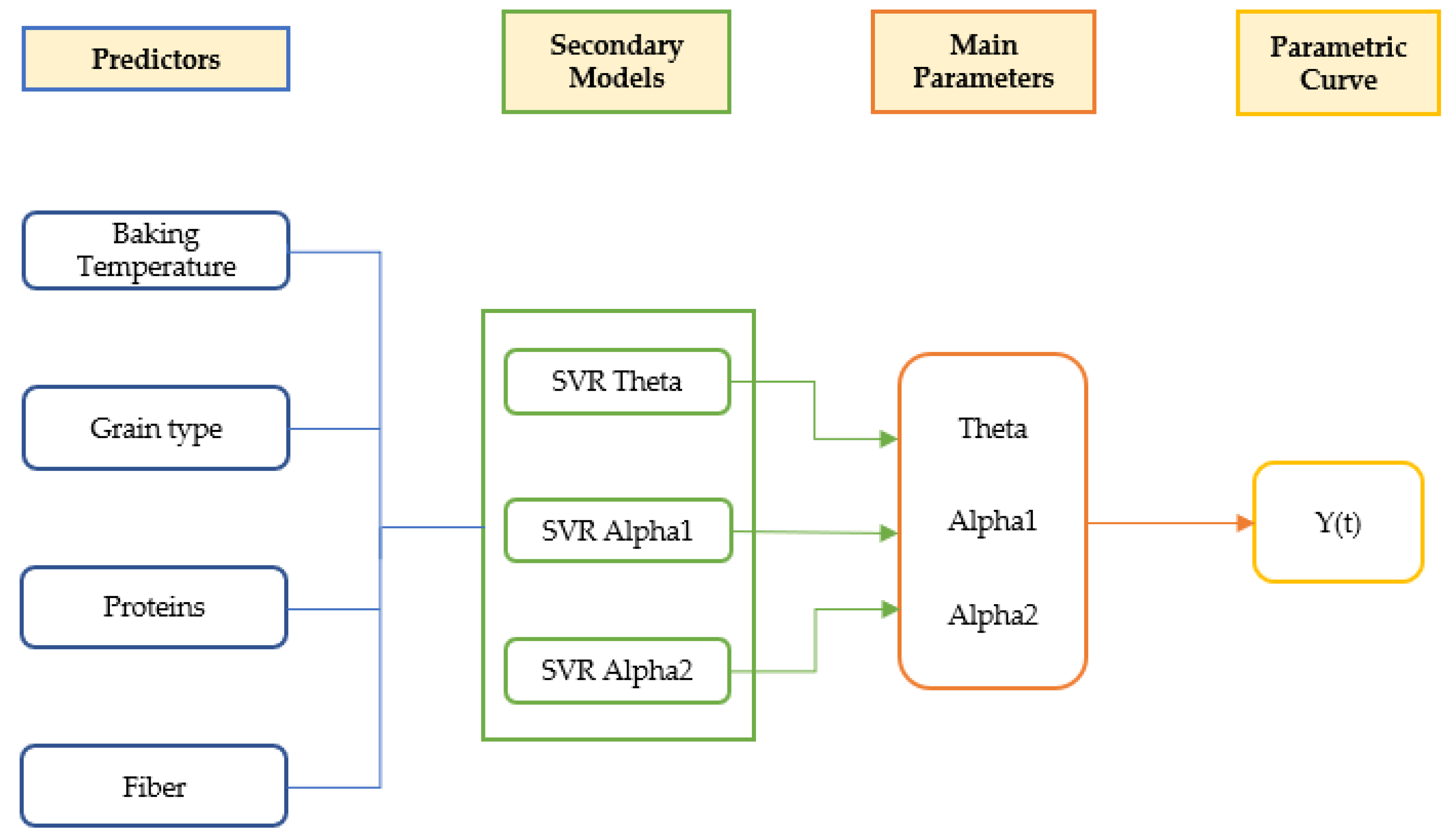

Table 1.
Values corresponding to the parameters 'ϴ, 'α1' and 'α2' of TP, ORAC and FRAP models.
| Setting | TP | ORAC | FRAP | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ϴ | Alpha1 | Alpha2 | ϴ | Alpha1 | Alpha2 | ϴ | Alpha1 | Alpha2 | |
| Berdun 180 °C | 0.605 | 1.985 | 0.100 | 0.291 | 0.826 | 0.117 | 0.296 | 0.297 | 0.059 |
| Igor 180 °C | 1.497 | 0.850 | 0.573 | 0.317 | 0.588 | 0.106 | 0.231 | 0.246 | 0.061 |
| Teodor 180 °C | 1.391 | 0.375 | 1.013 | 0.112 | 0.926 | 0.566 | 0.186 | 0.409 | 0.292 |
| Berdun 200 °C | 0.431 | 1.960 | 0.265 | 1.875 | 0.134 | 0.564 | 1.285 | 0.045 | 1.020 |
| Loretto 200 °C | 1.717 | 0.938 | 1.022 | 0.873 | 1.016 | 3.034 | 1.131 | 0.090 | 2.658 |
| Berdun 220 °C | 0.309 | 1.976 | 0.566 | 1.371 | 0.220 | 0.450 | 1.278 | 0.034 | 0.680 |
| Loretto 220 °C | 1.462 | 1.020 | 2.516 | 1.140 | 0.062 | 0.686 | 0.836 | 0.025 | 0.755 |
Table 2.
R2 of SRV models.
| Temperature | Variety | R2_TP | R2_ORAC | R2_FRAP |
|---|---|---|---|---|
| 180 °C | Berdun | 0.807 | 0.655 | -0.372 |
| 180 °C | Teodor | 0.021 | 0.500 | 0.387 |
| 180 °C | Igor | 0.396 | -0.404 | -2.081 |
| 200 °C | Berdun | 0.893 | 0.716 | -0.266 |
| 200 °C | Loretto | 0.952 | -2.051 | 0.935 |
| 220 °C | Berdun | 0.632 | 0.712 | 0.423 |
| 220 °C | Loretto | 0.919 | 0.844 | 0.881 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



