
Submitted:
08 July 2023
Posted:
10 July 2023
You are already at the latest version
Abstract
In the 21st century, the effects of HIV-associated neurocognitive disorders (HAND) have been significantly reduced in individuals due to the development of antiretroviral therapies (ARTs). However, the growing epidemic of polysubstance use (PSU) has led to concern for the effects of PSU on HIV-seropositive individuals. To effectively treat individuals affected by HAND, it is critical to understand the biological mechanisms affected by PSU including identification of novel markers. To fill this important knowledge gap, we used an in vivo HIV-Transgenic (HIV-Tg) animal model, to investigate the effects of the combined use of chronic methamphetamine (METH) and oxycodone (oxy). RNA-Seq analysis on the striatum - a brain region that is primarily targeted by both HIV and drugs of abuse - identified key differentially expressed markers post-METH and oxy exposure. Furthermore, ClueGO analysis and Ingenuity Pathway Analysis (IPA) revealed crucial molecular and biological functions associated with ATP-activated adenosine receptor, neuropeptide hormone activity, and the oxytocin signaling pathway to be altered between the different treatment groups. The current study further reveals the harmful effects of chronic PSU and HIV infection that can subsequently impact neurological outcomes in polysubstance users with HAND.
Keywords:
Polysubstance use
; methamphetamine
; oxycodone
; HIV
; transgenic rat
; RNA-sequencing
; bioinformatics
1. Introduction
Polysubstance use (PSU) is a rapidly growing form of substance use disorder (SUD), involving the combined use of more than one type of drug to elicit the user’s desired effect [1]. Commonly, polysubstance users will start off with abusing one drug often enough to be categorized as having SUD. Eventually, the user will develop a tolerance towards it or become tired of the repeated effects, prompting them to explore the combined effects of multiple drugs [2]. Using drugs from different classifications together, such as stimulants (i.e., methamphetamine and heroin) and depressants (i.e., alcohol and opioids), can help individuals feel new and addictive sensations leading to long-term PSU. This form of PSU is also known as “speedballing.” However, using multiple drugs at once makes it difficult for individuals to track the amount of each drug in their system. Furthermore, it is not possible to predict how each individual’s body will react to the combined usage of more than one drug. This can lead to an increased chance of overdosing on drugs. In 2019, it was reported that one-third of drug overdose deaths in the United States involved the co-use of stimulants and opioids [3]. Many studies have examined the impacts of singular drug use, allowing scientists to develop treatment methods for SUD. Unfortunately, in the context of PSU, these treatments are ineffective due to the lack of research focused primarily on multiple drug use.
Therefore, the present study investigates the combinatorial effects of methamphetamine (METH), a notorious stimulant, and oxycodone (oxy), a commonly prescribed opioid. Mixing drugs with opposing effects, like stimulants which increase activity in the central nervous system (CNS), and depressants which slow down activity in the CNS, aids chronic drug users in dealing with the negative consequences of the drugs associated with individually using them [4]. These drug classifications are known to permeate the blood-brain barrier (BBB) and reach the CNS, encompassing the brain and spine [5]. Thus, chronic drug use can cause permanent changes in an individual’s behaviors and cognitive function.
PSU can lead to increased impulsivity due to delayed processing and poor judgment skills. This puts substance abusers in danger of contracting incurable, life-altering diseases like human immunodeficiency virus (HIV). While under the unpredictable influence of multiple substances, individuals may partake in high-risk behaviors such as unprotected sex, violence, and suicide [6,7] . Polysubstance users are also more likely to use intravenous injections to absorb drugs into their bloodstream directly. This allows them to feel the effects of the drugs faster but puts them at a higher risk of HIV infection. Intravenous injections can lead to substance abusers using a needle more than once or sharing a needle with others; both can increase the risk of spreading the virus [8]. HIV can infect the CNS, allowing it to damage neurons and cause inflammation in the brain. Furthermore, HIV introduces viral proteins to the CNS, making the BBB more permeable and susceptible to other harmful infections [9].
Currently, very little is known about the consequences of chronic PSU in HIV-seropositive individuals. This study utilized RNA sequencing and bioinformatic analysis to identify key regulated genes affected by HIV and PSU. The RNA was isolated from striatal tissue in the brains of HIV-Transgenic (HIV-Tg) rats. An animal model featuring HIV-Tg rats was used to model the harmful effects of PSU on individuals with HIV at the transcriptional level. Previous studies have utilized these HIV-1 Tg animals, making them widely accepted in the realm of neuroHIV [10-12]. The HIV-1 Tg rats express viral proteins in their CNS and blood, causing neurocognitive impairments and behavioral changes similar to those seen in HIV-seropositive humans on CART and drug abuse [10-12]. Additionally, we used high throughput ‘omics’ to identify molecular processes and pathways associated with the adverse effects of chronic PSU and HIV. Overall, this study’s findings point to further directions for developing therapeutic targets to improve the health and quality of life of HIV-seropositive individuals with a history of PSU.
2. Materials and Methods
2.1. Animals
Male and female Fischer 344 wildtype (WT) and F344 Hsd: HIV-1 Transgenic (Tg) rats aged 6-7 months and bred under licensure were used in the study. All animals were housed under constant conditions in a 12h light-dark cycle and had access to food and water ad libitum. All procedures and protocols were approved by the Institutional Animal Care and Use Committee (IACUC) of the University of Nebraska Medical Center (IACUC protocol: 20-104-01-FC date 08/19/2021) and conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
2.2. Polysubstance administration
To mimic the start of a chronic PSU regimen, we first subjected the animals to a single drug. METH administration was performed in the first seven days with an escalation paradigm (Figure 1). From the 8th day onwards, the animals received a 10mg/kg METH+15mg/kg oxy co-treatment until day 14. The concentration of oxy used in our animal model mimics the high dose range equivalent in the induction of chronic analgesia in humans [13]. Control animals received isovolumetric saline during the treatment regimen. The treatment regimen lasted 14 days, and animals were sacrificed one h after the last dose, after which the brain and blood plasma was harvested.
2.3. Total RNA extraction, quality control, library preparation, and RNA-Sequencing
Total RNA was extracted from the brain cortex of 16 rats from both sexes and different treatment groups using the Direct-Zol RNA kit (Zymo Research, Irvine, CA, USA) following the manufacturer's protocol. RNA samples were sent on dry ice to LC Sciences (Houston, TX, USA) for RNA sequencing.
Library preparation was done by trimming fastq format files with fqtrim tool (https://ccb.jhu.edu/software/fqtrim) to remove adapters, unknown terminal bases (Ns) and low-quality 3’ regions (Phred score < 30). The trimmed fastq files were processed by FastQC [14]. FastQC: a quality control tool for high throughput sequence data; available online at http://www.bioinformatics.babraham.ac.uk/projects/fastqc for quality control. Rat mRatBN7.2 (https://www.ncbi.nlm.nih.gov/data-hub/genome/GCF_015227675.2/) and HIV-1 GCF_000864765.1 (https://www.ncbi.nlm.nih.gov/assembly/GCF_000864765.1) reference genome and annotation files were downloaded from NCBI RefSeq. The downloaded rat and HIV-1 ref genome and annotation files were combined as one rat_HIV-1 ref genome file and one rat_HIV-1 annotation file. The trimmed fastq files were mapped to rat_HIV-1 by CLC Genomics Workbench 22 (QIAGEN, Aarhus, Denmark) for RNAseq analyses.
Once the transcriptome library was prepared, TPM values (Transcripts Per Kilobase Million) were calculated by dividing the read counts by the length of each gene in kilobases. Genes expression were normalized using read per kilobase (RPK) in R (version 4.1.2), package DESeq2 [15].
2.4. Bioinformatic data analysis
After normalization, a student t-test was done for RNA analysis to identify RNAs showing significant differences between groups (WT vs. HIV; WT-PSU vs HIV-PSU; and WT vs. WT-PSU). RNAs were filtered using False Discovery Rates (FDR) ≤ 5% and p < 0.05 were considered significant. Cytoscape plug-in ClueGO was used to perform gene ontology (GO) analysis on the differentially expressed genes (DEGs) [16]. Canonical pathway analysis was performed using the Ingenuity Pathway Analysis (IPA) software (Ingenuity® Systems, Redwood City, CA, USA, www.ingenuity.com, accessed on 17 January 2023) by comparing the differentially expressed proteins against known canonical pathways (signaling and metabolic) within the IPA database [17].
3. Results
3.1. HIV and Chronic PSU Leads to Changes in Striatum Transcriptome
To corroborate the effects of polysubstance use on HIV transcriptome, we subjected striatal RNA from the wildtype (WT), HIV, wildtype with polysubstance use (WT+PSU), and HIV with polysubstance use (HIV+PSU) groups to high throughput quantitative RNA sequencing. Post normalization, three comparisons were made between the control and experimental groups to examine the differentially expressed genes (DEGs) that resulted from PSU and HIV independently and together. At a criterion of absolute fold-change greater than 2 and FDR adjusted p-value of 0.05, WT vs. WT+PSU had a total of 62 DEGs, HIV vs. HIV+PSU had 232 genes, and WT+PSU vs. HIV+PSU had 193 DEGs. Noteworthy, the majority of the DEGs are protein-coding genes, followed by the long non-coding RNA. Furthermore, principal component analysis (PCA) revealed good reproducibility of the biological replicates and overall separation between the groups (Figure 2).
Since high throughput ‘omics’ studies generate many potential hits, it is imperative to illustrate them visually. Therefore, we generated heatmaps on the top 50 DEGs for each comparison (Figure 3).
3.2. ClueGO and IPA Analysis Demonstrates Molecular Activities and Pathways Associated with HIV and Chronic PSU
Next, using the bioinformatics tool ClueGO, we analyzed the biological processes (BP) and molecular functions (MF) enriched with these DEGs. In Figure 4A, the most abundant BP associated with DEGs in the WT vs. WT+PSU group is the hormone activity, accounting for nearly two-thirds of the gene ontology (GO) terms (62.67%). Other GO terms associated with WT vs. WT+PSU are related to cellular responses to progesterone, alkaline, or fatty acid, which account for 29.33%, 4.0%, and 1.33%, respectively. Similar to the BP of the WT vs. WT+PSU group, the HIV vs. HIV+PSU and WT+PSU vs. HIV+PSU groups also had DEGs that are associated with hormone activity such as response to corticotropin-releasing hormones (18.18% - HIV vs. HIV+PSU), and neurohypophyseal hormone activity (17.24% - WT+PSU vs. HIV+PSU) (Figure 4B and Figure 4C). Interestingly, for HIV vs. HIV+PSU and WT+PSU vs. HIV+PSU, there are relatively many DEGs that have similar BP GO-terms, such as regulation of muscle filaments sliding speed (18.18% for HIV vs. HIV+PSU and 13.79% for WT+PSU vs. HIV+PSU), and skeletal muscle activity (9.09% for HIV vs. HIV+PSU and 24.14% for WT+PSU vs. HIV+PSU) (Figure 4B and Figure 4C).
For MF, the top abundant GO terms associated with WT vs. WT+PSU are ATP-activated adenosine receptor activity (14.29%), and neuropeptide activity (14.29%) (Figure 5A); with HIV vs. HIV+PSU are regulation of ATPase activity (42.86%), and acidic amino acid transmembrane transporter activity (23.81%) (Figure 5B); with WT+PSU vs. HIV+PSU, those top GO terms are associate with voltage-gated calcium channel activity (18.18%), and neurohypophyseal hormone activity (18.18%) (Figure 5C). Noticeably, the GO terms in MF between WT vs. WT+PSU and HIV vs. HIV+PSU comparisons both consist of RNA polymerase II sequence-specific DNA binding (8.16% in WT vs. WT+PSU and 4.76% in HIV vs. HIV+PSU), while in WT vs. WT+PSU and WT+PSU vs. HIV+PSU comparisons, both consist of protein kinase inhibitor activity (2.04% and 4.55%) and protein homodimerization activity (2.04% and 4.55%). On the other hand, the only GO terms that have commonly been seen between HIV vs. HIV+PSU and WT+PSU vs. HIV+PSU comparisons is cytochrome-c oxidase activity (4.76% and 4.55%).
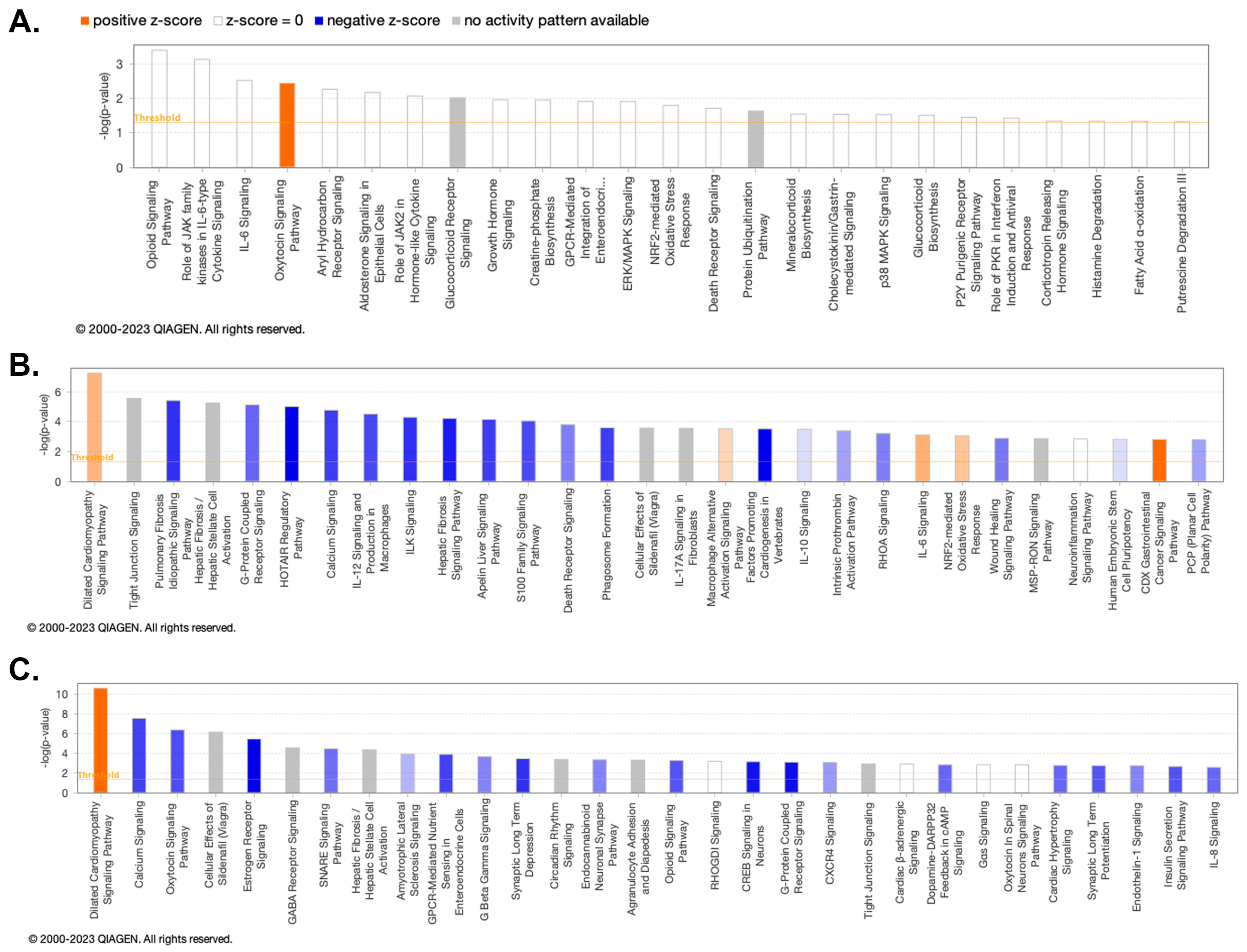
We further investigated potential enriched pathways associated with the DEGs using ingenuity pathways analysis (IPA). As seen in Figure 6A, the pathway associated with the oxytocin signaling pathway was enriched and highly activated in WT vs. WT+PSU. Figure 6B, pathways associated with HIV vs. HIV+PSU were highly enriched in several signaling pathways, such as dilated cardiomyopathy (DCM), macrophage alternative activation, and IL-6. In contrast, pulmonary fibrosis idiopathy, G-protein coupled receptors, calcium signaling, ILK, and many other signaling pathways were downregulated. Figure 6C showed that the DCM signaling pathway was highly enriched, similar to 6B. Also, in Figure 6C, many signaling pathways, such as calcium, oxytocin, estrogen receptor, GPCR-mediated nutrient sensing, and opioid signaling pathways, had significant negative z-score, indicating high deactivation. Gene-to-disease associations are provided in Table S3.
4. Discussion
Substance use disorder (SUD) is a chronic and relapsing disease that is characterized by a heightened sensitivity to drug-related cues and an increased motivation to maintain drug consumption [18]. With 35 million people globally and 19 million in the United States diagnosed with SUD, it has become a major public health issue that incurs both social and economic costs. The simultaneous use of multiple drugs is common among drug users, which complicates the study of pathological outcomes and limits the available treatment options. Polysubstance use (PSU) also results in an increased frequency of drug relapse and higher mortality rates compared to single-substance abuse [19-21]. While there is little research examining the impacts of PSU, specifically METH and oxy, on the CNS, there is even less research focusing on the effect of PSU in individuals with HIV. As of 2020, an estimated 1.1 million people in the United States are diagnosed and living with HIV [22]. Before combined antiretroviral therapy (cART) became a standard treatment for managing HIV, severe neurocognitive deficits due to the body’s immune response to HIV infection were prevalent [23]. More advanced cART forms have helped physicians limit cognitive impairment in HIV-seropositive cases, but HIV-associated neurocognitive disorders (HAND) remain persistent. Characteristics of HAND include memory impairment, difficulty concentrating, and poor coordination [24]. In 2020, about 43% of individuals with HIV worldwide were affected by HAND, indicating that HIV infection’s impact on the CNS is still a concerning and prevalent issue [25]. With the increased risk of contracting HIV that PSUs have, it is imperative to comprehensively understand how multiple non-prescribed substances and HIV interact in the body and affect brain function. The unknown consequences of PSU in HIV-infected individuals may render cART ineffective or limit its benefits when trying to decrease the severity of HAND. HIV-seropositive individuals using substances may benefit from a more specialized, alternative treatment plan than individuals without the virus. This study uses RNA sequencing to generate a comprehensive transcriptomic landscape of the brain in a rodent model of HAND to discover novel targets and develop therapeutic strategies for those suffering from HIV and PSU.
Our ClueGO analysis for both BP and MF found very interesting results. First, for the comparison between WT individuals and WT+PSU, we observed that DEGs are majorly involved in ATP-activated adenosine receptors (e.g., P2ry1) and neuropeptide hormone activity (e.g., Avp and Oxt). The P2ry1 gene is a protein-coding gene that codes for G-protein coupled P2Y receptors. Previous studies about microglia in culture showed that P2RY1 is responsible for rapid microglial membrane disturbance and whole-cell migration via ATP signaling [26,27]. Importantly, human and animal studies have shown that chronic use of most drugs of abuse causes a significant increase in peripheral and brain inflammation signals which trigger microglia and astrocyte activation in the brain. Microglia and/or astrocyte activation has been demonstrated in animal models of every drug studied, including amphetamines, cocaine, ethanol, opioids, cannabinoids, and nicotine [28]. In this study, we identified that the P2ry1 gene is upregulated upon exposure to METH+oxy. This highly suggests that the induction of METH+oxy could trigger P2ry1 as a responding mechanism for potential inflammation. On the other hand, Avp and Oxt genes are downregulated (Table S1). Arginine vasopressin (Avp) gene is encoded for neuropeptide vasopressin, which contributes to balancing the body’s osmotic, balances blood pressure, maintains sodium homeostasis, and regulates kidney’s function [29]. In the past several decades, there has been an increase in research into the roles of Avp in the brain and periphery. In anatomical, pharmacological, and transgenic animal studies, including "knockout" studies, mounting evidence implies that Avp is associated with regulating various social behaviors across species. Avp is also important in facilitating or promoting aggression [30]. Memory, one of the first Avp brain functions studied, is particularly important in social recognition. Stress-related corticotropin-releasing factor (CRF) and vasopressin (AVP) peptides are strongly associated with METH addiction-related psychostimulant-induced behaviors [31].
Additionally, we found oxytocin (OXT), a 9-amino acid neuropeptide, played a pivotal role in regulating social bonding, reproduction, and childbirth [32] and was downregulated upon exposure to METH+oxy. OXT is synthesized in the hypothalamus and released into the bloodstream by nerve endings in the posterior pituitary. In behavioral endocrinology, mounting evidence has shown that OXT is a key molecule that promotes anxiolytics [33] and enhances multiple aspects of social cognition such as emotion recognition, social perception, empathetic ability, and trust [34-36]. Notably, a study by Uhrig and colleagues [35] found evidence to support that the downregulation of OXT expression and receptors in brain regions involved in social cognition may lead to a dysfunction of oxytocin signaling, which is positively correlated to the development of schizophrenia. Particularly, many drugs such as opiates, methamphetamine, and cocaine cause long-term changes in oxytocin function markers, which may be linked to social behavior deficits seen in laboratory animals repeatedly exposed to these drugs [37]. Our study specifically showed that the combined use of METH and oxy also reduced oxytocin expression. However, the mechanism of how METH and/or oxy use leads to the reduction of oxytocin synthesis is unclear. Whether using METH and oxy together would have a significant summative effect on the oxytocin level and signaling remains unknown. The rising investigation on the effect of oxytocin treatment on various drug-seeking and drug-induced behaviors has shown the potential of oxytocin in diminishing drug use [38-41]. However, these studies mostly pertain to single drug addiction, while in the real world, people could be exposed to multiple drug use simultaneously. Hence, combining the results of our study and the potential therapeutic effect of oxytocin, it could be a promising investigation topic for determining whether oxytocin treatment is a powerful tool to alleviate PSU effects.
In both the HIV vs. HIV+PSU and WT+PSU vs. HIV+PSU comparisons, Tnnc1, Tnnt2, and Actc1 were found to be majorly involved in the calcium signaling and dilated cardiomyopathy (DCM) signaling pathways. The Tnnc1 and Tnnt2 genes are part of the troponin complex found on thin filaments of striated muscle [42]. These genes were found to be downregulated in the HIV+PSU group in both comparisons (Table S2). Troponin activates and regulates muscle contraction by transmitting information through structural changes in the filaments [43]. The Actc1 gene belongs to the actin family and is responsible for various cell functions, including cell motility and transcription regulation [44]. In our study, Actc1 was found to be downregulated in the HIV+PSU group. A published study showed that Actc1 is crucial to brain tumor cell survival. Actc1 is highly expressed in sonic hedgehog (SHH) medulloblastoma, a malignant brain tumor, and promotes cell migration and formation and leads to apoptosis resistance [45]. Altogether, the downregulation of the Tnnc1, Tnnt2, and Actc1 potentially suggests dysregulation in the calcium signaling pathway. This pathway significantly contributes to synaptic activity and neurotransmission [46]. Any dysregulation of the calcium signaling pathway has been linked to the development of brain-associated disorders, such as Alzheimer’s disease and schizophrenia [46]. HIV proteins have also been shown to cause calcium dysregulation in infected neurons, which can induce neuronal death or loss of function [47]. Thus, HIV-seropositive individuals that are chronic substance abusers may experience significantly more brain damage despite receiving modern cARTs.
Furthermore, alterations in the troponin genes, Tnnc1 and Tnnt2, and Actc1 can cause dysregulation in the DCM pathway. DCM is a condition that decreases the heart’s ability to pump blood to the rest of the body. This can lead to heart failure, which reduces oxygen levels in the brain, leading to neurological-associated symptoms like dizziness and nausea. Heart failure has also been shown to affect the brain structure acutely [48]. Drugs of abuse, like psychostimulants (e.g., METH) and opioids (e.g., oxy), have been associated with left-ventricular systolic desynchrony (LVSD), a relevant prognostic marker for identifying DCM in patients [49,50]. The increase in METH use has specifically led to increased methamphetamine-associated cardiomyopathy. Individuals with methamphetamine-associated cardiomyopathy experience more severe symptoms than non-METH users with DCM [51]. This implies that polysubstance use can increase the risk of developing and experiencing severe DCM. Importantly, the development of DCM is also highly correlated with HIV infection. HIV-associated cardiomyopathy is a growing problem in countries without cART readily available for HIV-seropositive patients [52,53]. Consequently, HIV-seropositive patients who are polysubstance users are prone to experiencing a severe form of DCM with less effective treatments. The downregulation of Tnnc1, Tnnt2, and Actc1 potentially provides insights into the underlying mechanisms of HIV that PSU affects.
In summary, this study for the first time elucidates the effect of chronic HIV infection along with combined use of METH and oxy affects in a model of HAND. The novel gene signatures including biological processes and molecular functions associated could further be developed to inform therapeutic treatments and in mitigating CNS dysfunction in these individuals.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
All authors have read and agreed to the published version of the manuscript. Pranavi Athota: Writing – Original Draft; Investigation; Visualization. Nghi M. Nguyen: Writing – Original Draft; Visualization; Data Curation; Formal Analysis. Victoria L. Schaal: Investigation. Sankarasubramanian Jagadesan: Data curation; Visualization; Formal Analysis. Chittibabu Guda: Data curation; Visualization; Formal Analysis. Sowmya Yelamanchili: Funding acquisition Supervision; Conceptualization; Writing – Review and Editing. Gurudutt Pendyala: Funding acquisition; Conceptualization; Project administration; Supervision; Writing – Review and Editing.
Funding
This work was supported by NIH grants, DA046284 (GP), DA046852 (GP and SY); departmental startup funds (GP and SY), and the Lieberman Endowment (GP). The funding entities played no role in the study design or its conclusion.
Institutional Review Board Statement
The study was conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and approved by the Institutional Animal Care and Use Committee (IACUC) of the University of Nebraska Medical Center (20-104-01-FC | approved: 08/19/2021).
Informed Consent Statement
Not Applicable.
Data Availability Statement
The data discussed in this publication have been deposited in NCBI's Gene Expression Omnibus (Athota et al., 2023) and are accessible through GEO Series accession number GSE236466 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc= GSE236466).
Acknowledgments
We would like to acknowledge Dr. Peng Xiao from UNMC Bioinformatics Core for providing support in parsing the initial raw data files.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cicero, T.J.; Ellis, M.S.; Kasper, Z.A. Polysubstance Use: A Broader Understanding of Substance Use During the Opioid Crisis. Am J Public Health 2020, 110, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Crummy, E.A.; O'Neal, T.J.; Baskin, B.M.; Ferguson, S.M. One Is Not Enough: Understanding and Modeling Polysubstance Use. Front Neurosci 2020, 14, 569. [Google Scholar] [CrossRef] [PubMed]
- O'Donnell, J.; Gladden, R.M.; Mattson, C.L.; Hunter, C.T.; Davis, N.L. Vital Signs: Characteristics of Drug Overdose Deaths Involving Opioids and Stimulants - 24 States and the District of Columbia, January-June 2019. MMWR Morb Mortal Wkly Rep 2020, 69, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Compton, W.M.; Valentino, R.J.; DuPont, R.L. Polysubstance use in the U.S. opioid crisis. Molecular Psychiatry 2021, 26, 41–50. [Google Scholar] [CrossRef]
- Pimentel, E.; Sivalingam, K.; Doke, M.; Samikkannu, T. Effects of Drugs of Abuse on the Blood-Brain Barrier: A Brief Overview. Frontiers in Neuroscience 2020, 14. [Google Scholar] [CrossRef]
- CDC. Substance Use and Sexual Risk Behaviors. Available online: https://www.cdc.gov/healthyyouth/substance-use/dash-substance-use-fact-sheet.htm.
- Baskin-Sommers, A.; Sommers, I. The co-occurrence of substance use and high-risk behaviors. Journal of Adolescent Health 2006, 38, 609–611. [Google Scholar] [CrossRef]
- Wang, S.-C.; Maher, B. Substance Use Disorder, Intravenous Injection, and HIV Infection: A Review. Cell Transplantation 2019, 28, 1465–1471. [Google Scholar] [CrossRef] [PubMed]
- Scutari, R.; Alteri, C.; Perno, C.F.; Svicher, V.; Aquaro, S. The Role of HIV Infection in Neurologic Injury. Brain Sci 2017, 7. [Google Scholar] [CrossRef]
- Lashomb, A.L.; Vigorito, M.; Chang, S.L. Further characterization of the spatial learning deficit in the human immunodeficiency virus-1 transgenic rat. Journal of NeuroVirology 2009, 15, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Kass, M.D.; Liu, X.; Vigorito, M.; Chang, L.; Chang, S.L. Methamphetamine-induced behavioral and physiological effects in adolescent and adult HIV-1 transgenic rats. Journal of neuroimmune pharmacology : the official journal of the Society on NeuroImmune Pharmacology 2010, 5, 566–573. [Google Scholar] [CrossRef]
- Vigorito, M.; Cao, J.; Li, M.D.; Chang, S.L. Acquisition and long-term retention of spatial learning in the human immunodeficiency virus-1 transgenic rat: effects of repeated nicotine treatment. J Neurovirol 2013, 19, 157–165. [Google Scholar] [CrossRef]
- Fan, R.; Schrott, L.M.; Arnold, T.; Snelling, S.; Rao, M.; Graham, D.; Cornelius, A.; Korneeva, N.L. Chronic oxycodone induces axonal degeneration in rat brain. BMC Neuroscience 2018, 19, 15. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: a quality control tool for high throughput sequence data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol 2010, 11, R106. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: a Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J., Jr; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2013, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Volkow, N.D. Neurobiology of addiction: a neurocircuitry analysis. Lancet Psychiatry 2016, 3, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Williamson, A.; Darke, S.; Ross, J.; Teesson, M. The effect of persistence of cocaine use on 12-month outcomes for the treatment of heroin dependence. Drug Alcohol Depend 2006, 81, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Staiger, P.K.; Richardson, B.; Long, C.M.; Carr, V.; Marlatt, G.A. Overlooked and underestimated? Problematic alcohol use in clients recovering from drug dependence. Addiction 2013, 108, 1188–1193. [Google Scholar] [CrossRef]
- de la Fuente, L.; Molist, G.; Espelt, A.; Barrio, G.; Guitart, A.; Bravo, M.J.; Brugal, M.T. Mortality risk factors and excess mortality in a cohort of cocaine users admitted to drug treatment in Spain. J Subst Abuse Treat 2014, 46, 219–226. [Google Scholar] [CrossRef]
- CDC. HIV Surveillance Report. Available online: https://www.cdc.gov/hiv/library/reports/hiv-surveillance/vol-33/index.html(accessed on Feb 20).
- Nweke, M.; Mshunqane, N.; Govender, N.; Akinpelu, A.O.; Ukwuoma, M. Impact of HIV-associated cognitive impairment on functional independence, frailty and quality of life in the modern era: a meta-analysis. Scientific Reports 2022, 12, 6470. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Sarkar, A.; Mitsuya, H. HIV-Associated Neurocognitive Disorder (HAND) and the Prospect of Brain-Penetrating Protease Inhibitors for Antiretroviral Treatment. Med Res Arch 2017, 5. [Google Scholar]
- Wang, Y.; Liu, M.; Lu, Q.; Farrell, M.; Lappin, J.M.; Shi, J.; Lu, L.; Bao, Y. Global prevalence and burden of HIV-associated neurocognitive disorder. A meta-analysis 2020, 95, e2610–e2621. [Google Scholar] [CrossRef] [PubMed]
- Honda, S.; Sasaki, Y.; Ohsawa, K.; Imai, Y.; Nakamura, Y.; Inoue, K.; Kohsaka, S. Extracellular ATP or ADP Induce Chemotaxis of Cultured Microglia through G<sub>i/o</sub>-Coupled P2Y Receptors. The Journal of Neuroscience 2001, 21, 1975–1982. [Google Scholar] [CrossRef]
- Davalos, D.; Grutzendler, J.; Yang, G.; Kim, J.V.; Zuo, Y.; Jung, S.; Littman, D.R.; Dustin, M.L.; Gan, W.-B. ATP mediates rapid microglial response to local brain injury in vivo. Nature Neuroscience 2005, 8, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Berríos-Cárcamo, P.; Quezada, M.; Quintanilla, M.E.; Morales, P.; Ezquer, M.; Herrera-Marschitz, M.; Israel, Y.; Ezquer, F. Oxidative Stress and Neuroinflammation as a Pivot in Drug Abuse. A Focus on the Therapeutic Potential of Antioxidant and Anti-Inflammatory Agents and Biomolecules. Antioxidants (Basel) 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Cuzzo, B.; Padala, S.A.; Lappin, S.L. Physiology, Vasopressin. In StatPearls; StatPearls Publishing Copyright © 2022, StatPearls Publishing LLC.: Treasure Island (FL), 2022. [Google Scholar]
- Caldwell, H.K.; Lee, H.J.; Macbeth, A.H.; Young, W.S. , 3rd. Vasopressin: behavioral roles of an "original" neuropeptide. Prog Neurobiol 2008, 84, 1–24. [Google Scholar] [CrossRef]
- Logrip, M.L.; Koob, G.F.; Zorrilla, E.P. Role of corticotropin-releasing factor in drug addiction: potential for pharmacological intervention. CNS Drugs 2011, 25, 271–287. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Macbeth, A.H.; Pagani, J.H.; Young, W.S. , 3rd. Oxytocin: the great facilitator of life. Prog Neurobiol 2009, 88, 127–151. [Google Scholar] [CrossRef]
- Neumann, I.D.; Slattery, D.A. Oxytocin in General Anxiety and Social Fear: A Translational Approach. Biological Psychiatry 2016, 79, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Lindenberg, A.; Domes, G.; Kirsch, P.; Heinrichs, M. Oxytocin and vasopressin in the human brain: social neuropeptides for translational medicine. Nature Reviews Neuroscience 2011, 12, 524–538. [Google Scholar] [CrossRef]
- Uhrig, S.; Hirth, N.; Broccoli, L.; von Wilmsdorff, M.; Bauer, M.; Sommer, C.; Zink, M.; Steiner, J.; Frodl, T.; Malchow, B.; et al. Reduced oxytocin receptor gene expression and binding sites in different brain regions in schizophrenia: A post-mortem study. Schizophrenia Research 2016, 177, 59–66. [Google Scholar] [CrossRef]
- Bartholomeusz, C.F.; Ganella, E.P.; Labuschagne, I.; Bousman, C.; Pantelis, C. Effects of oxytocin and genetic variants on brain and behaviour: Implications for treatment in schizophrenia. Schizophrenia Research 2015, 168, 614–627. [Google Scholar] [CrossRef] [PubMed]
- McGregor, I.S.; Bowen, M.T. Breaking the loop: oxytocin as a potential treatment for drug addiction. Horm Behav 2012, 61, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Sundar, M.; Patel, D.; Young, Z.; Leong, K.C. Oxytocin and Addiction: Potential Glutamatergic Mechanisms. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Leong, K.C.; Zhou, L.; Ghee, S.M.; See, R.E.; Reichel, C.M. Oxytocin decreases cocaine taking, cocaine seeking, and locomotor activity in female rats. Exp Clin Psychopharmacol 2016, 24, 55–64. [Google Scholar] [CrossRef]
- Edinoff, A.N.; Thompson, E.; Merriman, C.E.; Alvarez, M.R.; Alpaugh, E.S.; Cornett, E.M.; Murnane, K.S.; Kozinn, R.L.; Shah-Bruce, M.; Kaye, A.M.; et al. Oxytocin, a Novel Treatment for Methamphetamine Use Disorder. Neurol Int 2022, 14, 186–198. [Google Scholar] [CrossRef] [PubMed]
- King, C.E.; Gano, A.; Becker, H.C. The role of oxytocin in alcohol and drug abuse. Brain Res 2020, 1736, 146761. [Google Scholar] [CrossRef] [PubMed]
- Katrukha, I.A. Human cardiac troponin complex. Structure and functions. Biochemistry (Mosc) 2013, 78, 1447–1465. [Google Scholar] [CrossRef]
- Gomes, A.V.; Potter, J.D.; Szczesna-Cordary, D. The role of troponins in muscle contraction. IUBMB Life 2002, 54, 323–333. [Google Scholar] [CrossRef]
- Dominguez, R.; Holmes, K.C. Actin structure and function. Annual review of biophysics 2011, 40, 169–186. [Google Scholar] [CrossRef] [PubMed]
- Suresh, R.; Picard, D.; Lo, R.; Beaulieu, J.; Remke, M.; Diaz, R.J. Expression of cell type incongruent alpha-cardiac actin 1 subunit in medulloblastoma reveals a novel mechanism for cancer cell survival and control of migration. Neuro-Oncology Advances 2021, 3. [Google Scholar] [CrossRef]
- Berridge, M.J. Dysregulation of neural calcium signaling in Alzheimer disease, bipolar disorder and schizophrenia. Prion 2013, 7, 2–13. [Google Scholar] [CrossRef]
- Hu, X.T. HIV-1 Tat-Mediated Calcium Dysregulation and Neuronal Dysfunction in Vulnerable Brain Regions. Curr Drug Targets 2016, 17, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Mueller, K.; Thiel, F.; Beutner, F.; Teren, A.; Frisch, S.; Ballarini, T.; Möller, H.E.; Ihle, K.; Thiery, J.; Schuler, G.; et al. Brain Damage With Heart Failure: Cardiac Biomarker Alterations and Gray Matter Decline. Circ Res 2020, 126, 750–764. [Google Scholar] [CrossRef]
- Rangel, I.; Amorim, M.; Gonçalves, A.; Sousa, C.; Bettencourt, P.; Maciel, M.J. Toxic dilated cardiomyopathy: recognizing a potentially reversible disease. Arq Bras Cardiol 2014, 102, e37. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Tang, H.; Zhu, F.; Jiang, Z.; Shi, J.; Zhou, Y.; Garcia, E.V.; Li, D.; Zhou, W. Prognostic value of left-ventricular systolic and diastolic dyssynchrony measured from gated SPECT MPI in patients with dilated cardiomyopathy. Journal of Nuclear Cardiology 2020, 27, 1582–1591. [Google Scholar] [CrossRef] [PubMed]
- Zamanian, R.T.; Hedlin, H.; Greuenwald, P.; Wilson, D.M.; Segal, J.I.; Jorden, M.; Kudelko, K.; Liu, J.; Hsi, A.; Rupp, A.; et al. Features and Outcomes of Methamphetamine-associated Pulmonary Arterial Hypertension. Am J Respir Crit Care Med 2018, 197, 788–800. [Google Scholar] [CrossRef] [PubMed]
- Lumsden, R.H.; Bloomfield, G.S. The Causes of HIV-Associated Cardiomyopathy: A Tale of Two Worlds. BioMed Research International 2016, 2016, 8196560. [Google Scholar] [CrossRef] [PubMed]
- Barbaro, G. HIV-associated cardiomyopathy etiopathogenesis and clinical aspects. Herz 2005, 30, 486–492. [Google Scholar] [CrossRef]
Figure 1.
Preclinical rodent model mimicking METH+oxy use.
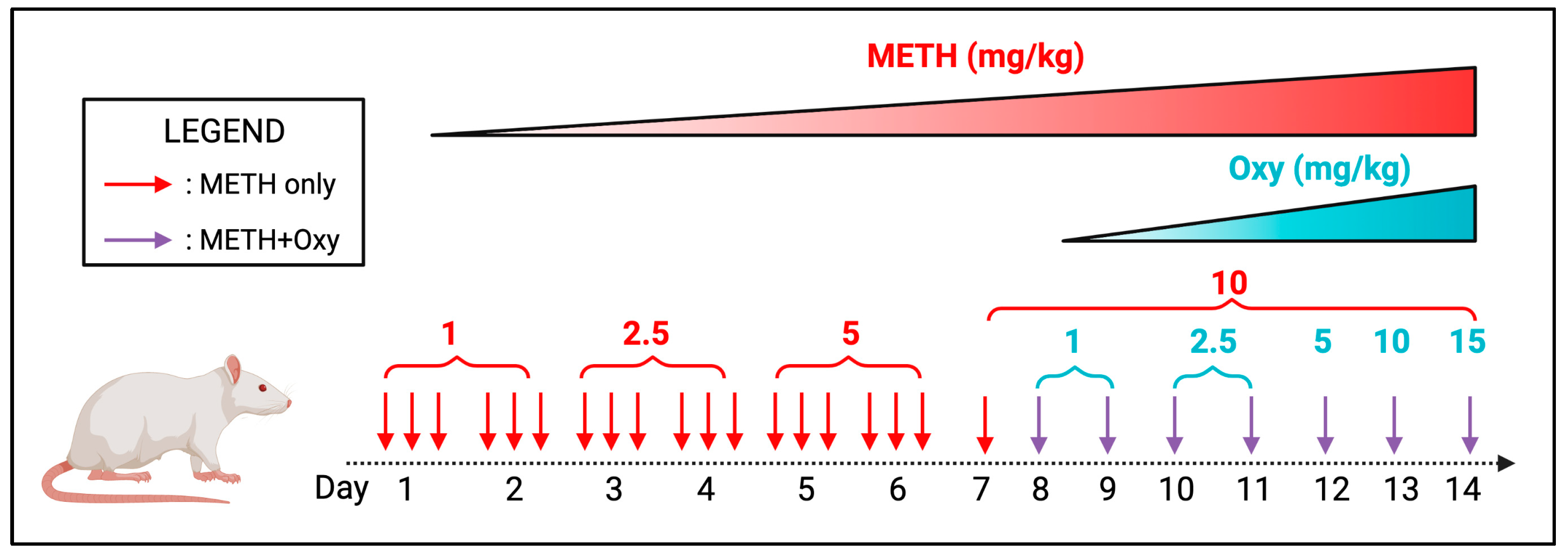
Figure 2.
Demographic of DEGs from RNA-Seq data. (A-C) Percentage of the different types of RNA (those that are differentially expressed) identified by RNA-seq data and principal component analysis (PCA) between each comparison: (A.) WT vs. WT+PSU (B.) HIV vs. HIV+PSU, and (C.) WT+PSU vs. HIV+PSU. (D.) Venn diagram showing total differentially expressed genes found in between samples (n=4/group).
Figure 2.
Demographic of DEGs from RNA-Seq data. (A-C) Percentage of the different types of RNA (those that are differentially expressed) identified by RNA-seq data and principal component analysis (PCA) between each comparison: (A.) WT vs. WT+PSU (B.) HIV vs. HIV+PSU, and (C.) WT+PSU vs. HIV+PSU. (D.) Venn diagram showing total differentially expressed genes found in between samples (n=4/group).
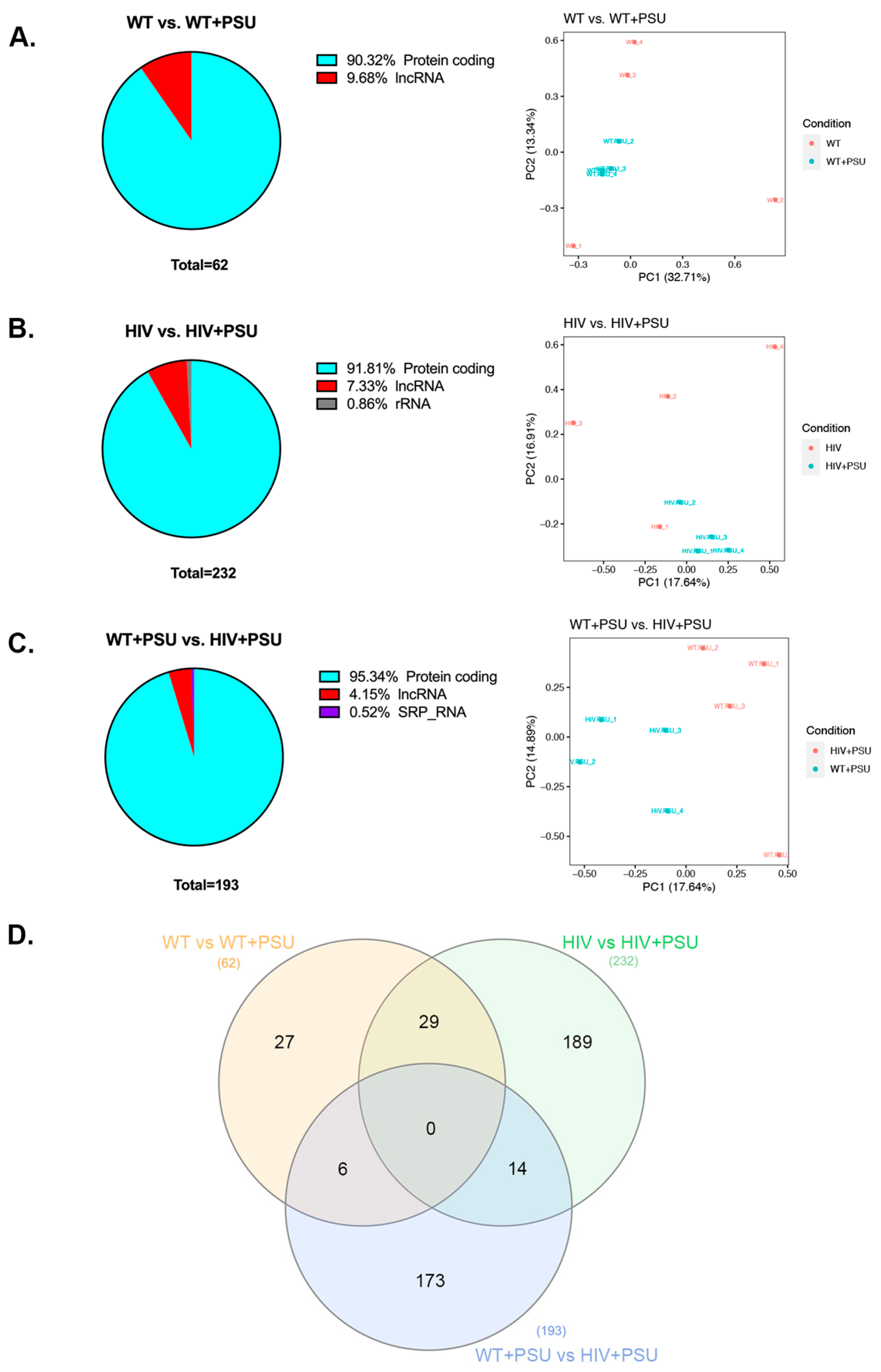
Figure 3.
Heatmaps of DEGs. Heatmap visualization of top 50 differentially regulated genes associated with each comparison: WT vs. WT+PSU, HIV vs. HIV+PSU, and WT+PSU vs. HIV+PSU (n=4/group).
Figure 3.
Heatmaps of DEGs. Heatmap visualization of top 50 differentially regulated genes associated with each comparison: WT vs. WT+PSU, HIV vs. HIV+PSU, and WT+PSU vs. HIV+PSU (n=4/group).
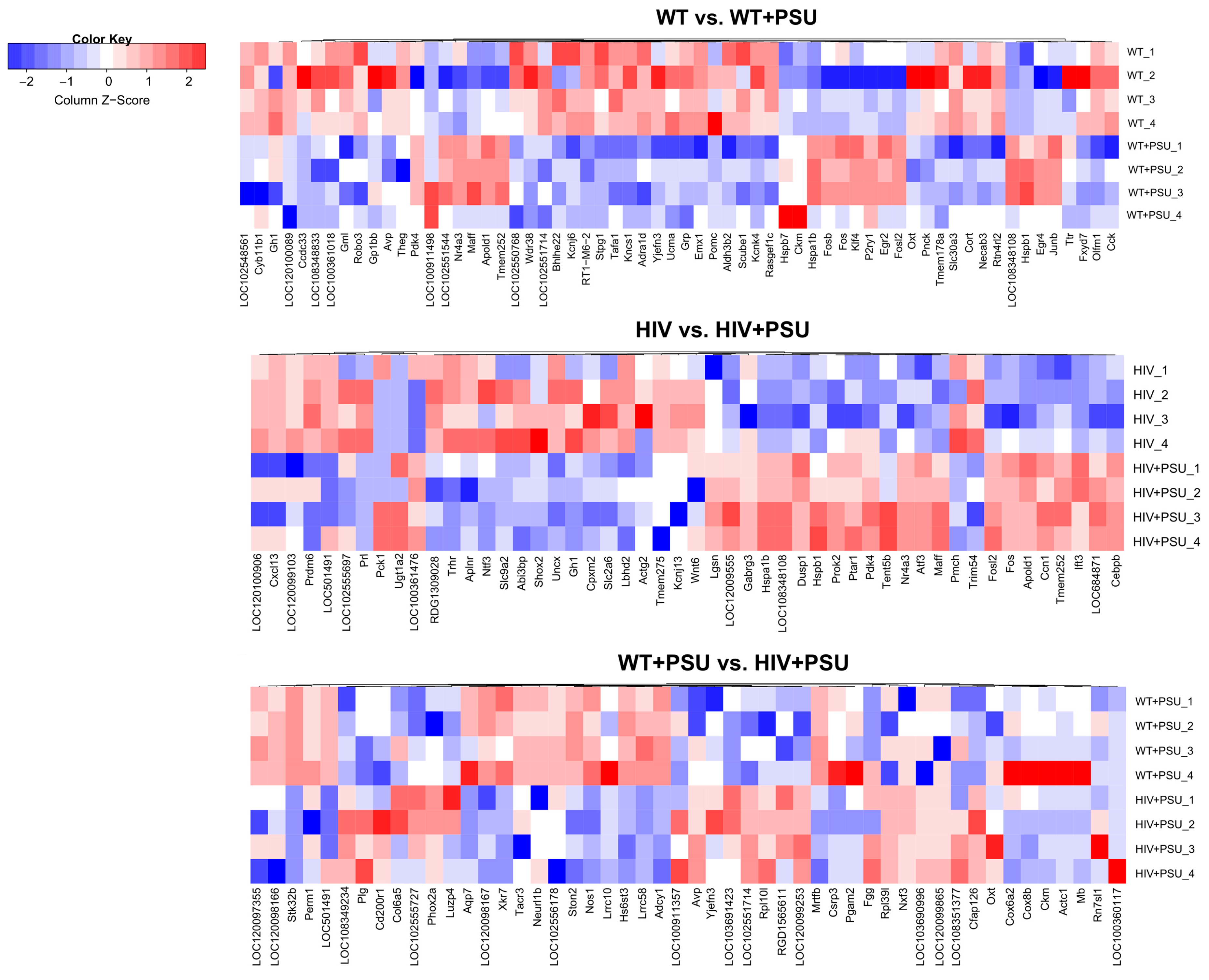
Figure 4.
ClueGO analysis of predicted biological processes. Pie charts representing groups of DEGs associated with targeted biological processes in each comparison (A.) WT vs. WT+PSU (B.) HIV vs. HIV+PSU, and (C.) WT+PSU vs. HIV+PSU.
Figure 4.
ClueGO analysis of predicted biological processes. Pie charts representing groups of DEGs associated with targeted biological processes in each comparison (A.) WT vs. WT+PSU (B.) HIV vs. HIV+PSU, and (C.) WT+PSU vs. HIV+PSU.
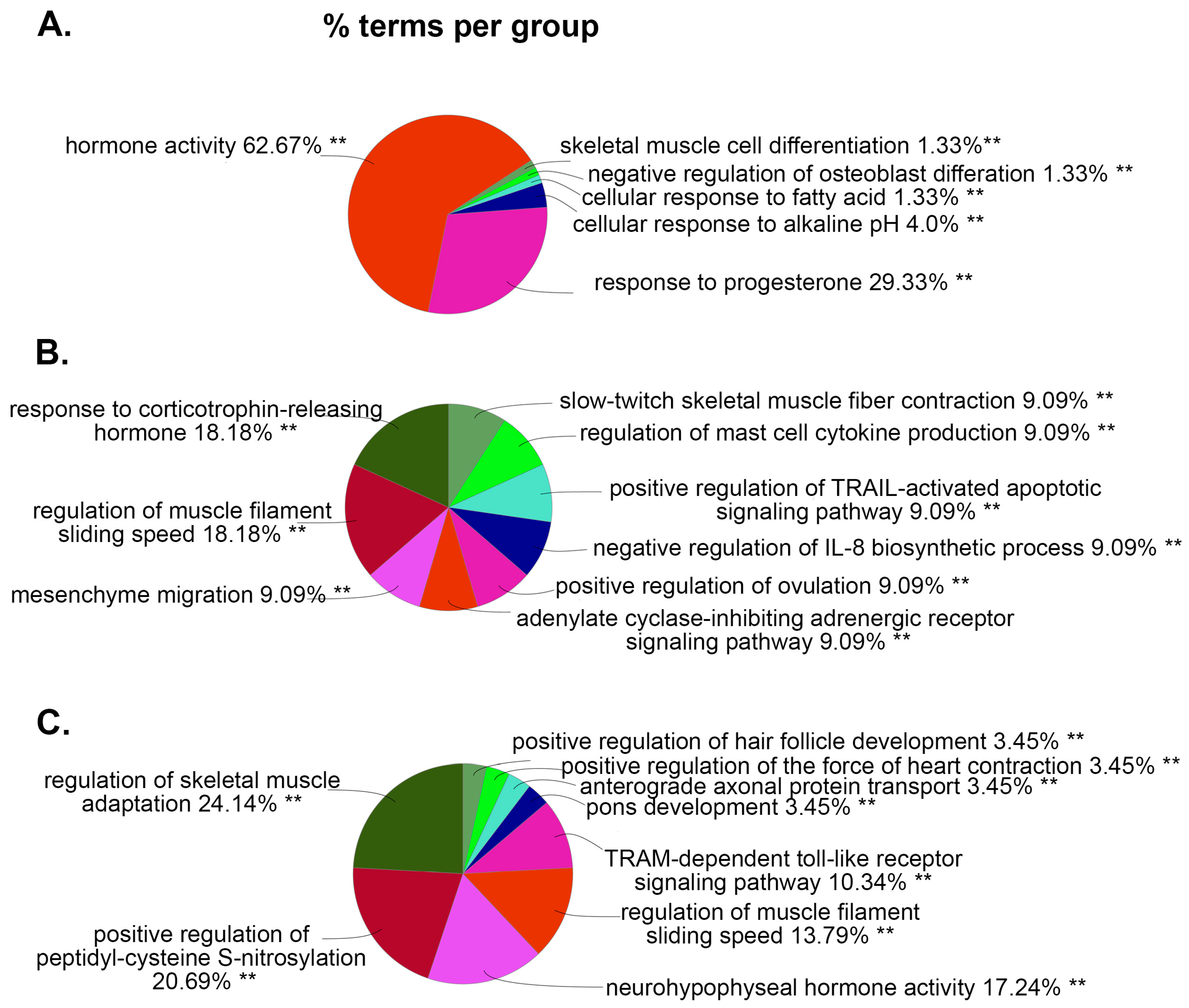
Figure 5.
ClueGO analysis of predicted molecular functions. Pie charts representing groups of DEGs associated with targeted molecular functions in each comparison (A.) WT vs. WT+PSU (B.) HIV vs. HIV+PSU, and (C.) WT+PSU vs. HIV+PSU.
Figure 5.
ClueGO analysis of predicted molecular functions. Pie charts representing groups of DEGs associated with targeted molecular functions in each comparison (A.) WT vs. WT+PSU (B.) HIV vs. HIV+PSU, and (C.) WT+PSU vs. HIV+PSU.
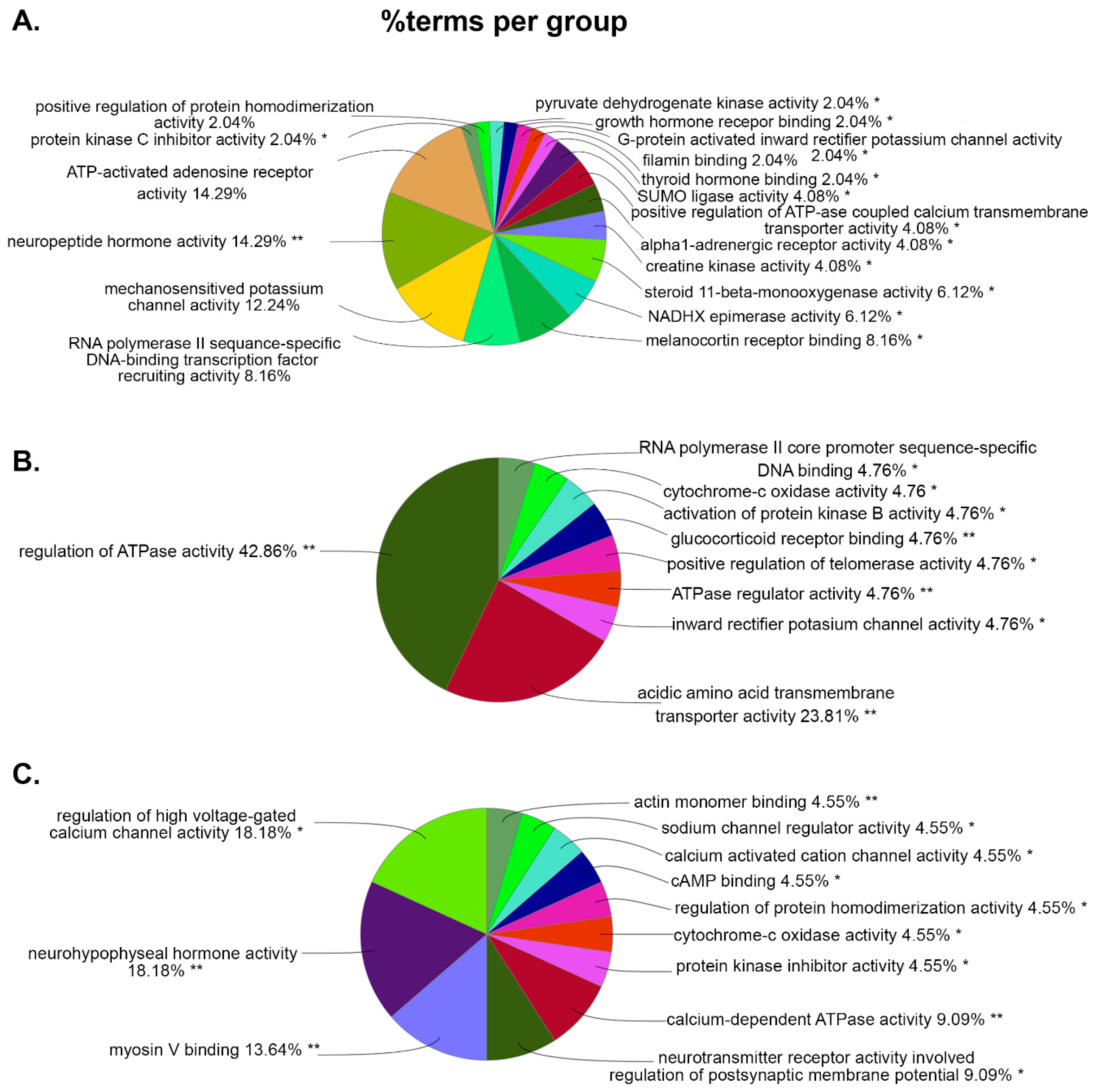
Figure 6.
DEGs Ingenuity pathway analysis (IPA). Enriched pathways associated with core pathways were analyzed by IPA, which identified relationships, mechanisms, functions, and pathways relevant to a dataset where the orange bar represent a positive z-score (activated pathway) and blue bar represents a negative z-score (inactivated pathway). (A.) WT vs. WT+PSU (B.) HIV vs. HIV+PSU, and (C.) WT+PSU vs. HIV+PSU.
Figure 6.
DEGs Ingenuity pathway analysis (IPA). Enriched pathways associated with core pathways were analyzed by IPA, which identified relationships, mechanisms, functions, and pathways relevant to a dataset where the orange bar represent a positive z-score (activated pathway) and blue bar represents a negative z-score (inactivated pathway). (A.) WT vs. WT+PSU (B.) HIV vs. HIV+PSU, and (C.) WT+PSU vs. HIV+PSU.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



