
Submitted:
10 July 2023
Posted:
11 July 2023
You are already at the latest version
Abstract
Bioactive plant compounds, also known as phytochemicals, are essential for plant resistance, growth, and development. These chemicals often possess therapeutic properties that have been used in traditional medicine for centuries as herbal remedies for a variety of diseases. As a member of the Lamiaceae family, Rydingia (Syn: Otostegia) is found in Asia and northeastern Africa. R. michauxii, is a unique shrub that grows in the Southern region of Iran, Fars province. Rydingia species have been used as traditional herbal medicine in Iran for many years. It seems that R. michauxii offers great potential for in-depth research on its diverse biological properties, especially its antioxidant and lipid-lowering effects. Nevertheless, no clinical investigation has been carried out on the phytochemical characteristics of this plant to date. Therefore, this study aims to contribute to the scientific knowledge on the pharmacological properties of R. michauxii and its phytochemical compounds and encourage researchers to evaluate its biological properties in animal models. In this study, all available scientific literature was compiled and all phytochemical and pharmacological properties of the R. michauxii plant were investigated, including antibacterial, antifungal, antiviral, antioxidant, antimalarial, cytotoxic, and lipid-lowering effects, which may prove useful in developing novel formulations with greater therapeutic benefits in clinical applications.
Keywords:
Bioactive chemicals
; Pharmacology
; Phytochemistry
; Rydingia michauxii
; Traditional Medicine
1. Introduction
The therapeutic use of medicinal plants dates back to ancient times [1]. Plants with medicinal properties produce secondary metabolites with a wide range of biological functions [2]. Lamiaceae is a family of flowering plants with about 236 genera and 6900-7200 species [3]. Plants belonging to the Lamiaceae family are annual or perennial, herbaceous or shrub, and are usually strongly glandular and aromatic [4]. Rydingia (Syn: Otostegia) is a Lamiaceae genus found in Asia and Northeastern Africa [5].
Four species of Rydingia (R. integrifolia, R. persica, R. limbata, and R. michauxii) have been described [6]. As an endemic shrub, R. michauxii grows in the areas of Fars province (Iran), where it occurs in shrub populations near land along or on slopes facing a river [7,8]. The plant is morphologically characterized by spines in the leaf axils, spinose, yellow flowers, bracteoles, and few-flowered verticillasters [9,10]. The chromosome number of R. michauxii is 2n=82 [11]. A variety of biological effects are associated with Rydingia species that are used in herbal medicine [9,12,13].
Traditionally, the aerial parts of Rydingia species are used in the forms of decoction and infusion to treat diabetes, palpitation, gastric discomfort, rheumatism, blood pressure, cardiac distress, cough, headaches, toothache, and arthritis. The plant also has sedative, antispasmodic, antidepressant, anti-hyperlipidemia and analgesic, anti-nociceptive, anti-glycation, antimicrobial, antimalarial, anti-inflammatory, antianxiety, morphine withdrawal, antihypertensive, anti-diabetic, healing of burn-wound, mosquito repellent, laxative, antipyretic, hepatoprotective, carminative, and antioxidant properties [13-19]. R. michauxii has shown to have numerous beneficial effects, including antibacterial, antifungal, antiviral, antimalarial, cytotoxic, and lipid-lowering properties (Figure 1). Research conducted between 2000 and 2022 demonstrate an upward trend in the development of natural compounds in all fields (Figure 2).
However, there is currently no comprehensive report available on R. michauxii that describes its promising natural compounds and pharmacological activities. In this review, we have summarized the biological and pharmacological potential of R. michauxii and its major compounds, which could serve as an overture for new studies and drug development.
2. Phytochemical reports
2.1. Essential oil
A total of forty-five chemical components were found in the essential oil of R. michauxii. Among the major constituents of the essential oil included dillapiole (23.9%), 2-methylbenzofuran (12.9%), and α-pinene (8.1%) [8]. Two phenylpropanoid compounds, namely dillapiole and 2-methylbenzofuran, were the dominant constituents, accounting for 36.8% of the total essential oil [8].
Another study [9] found that R. michauxii essential oil contained eighty compounds, including humulene epoxide II (4.6%), linalool (5%), trans-verbenol (10.2%), and caryophyllene oxide (20.1%). We have previously determined the essential oil composition of R. michauxii during various growth stages, i.e., dormant, vegetative, and flowering phases. Our findings indicate that the different growth stages were associated with the distinct phytochemical profiles of the essential oils. Specifically, the essential oil consisted of fifty-four, sixty-five, and fifty-nine components in the respective growth stages. [20].
The major chemical compounds of R. michauxii during dormancy were carvacrol (9.35%), eugenol acetate (21.02%), as well as eugenol (36.81%). At the vegetative growth stage, linalool (8.28%), cumin aldehyde (9.63%), eugenol (13.23%), and carvacrol (16.08%) were the major chemical constituents. The main volatile components at the flowering stage were carvacrol (14.20%), eugenol (8.98%), -cadinene (8.90%), caryophyllene oxide (8.43%), and n-hexadecanoic acid (7.07%). Compared to other stages, the highest levels of bornyl acetate (1.58%), (E)-caryophyllene (2.52%), eugenol acetate (21.02%), and eugenol (36.81%) were detected during plant dormancy. During vegetative growth, the highest concentrations were found for thuja-2,4(10)-diene (1.37%), methyl eugenol (1.4%), thymol (1.63%), trans-α-bergamotene (2.88%), epi-α-cadinol (4.33%), linalool (8.28%), cumin aldehyde (9.63%), and carvacrol (16.08%). While, the highest concentrations of γ-cadinene (2.54%), (Z)-4-decen-1-ol (3.02%), (Z)-4-decen-1-ol (3.02%), caryophyllene oxide (8.43%), and -cadinene (8.9%) were found during the flowering stage [20]. The structure of the major compounds found in R. michauxii essential oils is depicted in Figure 3. The chemical composition of R. michauxii essential oil exhibited significant variation based on geographical locations and developmental stages (Table 1).
2.2. Pharmacological activities of volatile constituents of R. michauxii
The GC-MS analysis identified various volatile compounds in R. michauxii essential oil. Dillapiole, caryophyllene oxide, eugenol, eugenol acetate, and carvacrol were the major compounds of the essential oil of R. michauxii. The biological activities of these compounds have been investigated in the previous studies.
2.3. Dillapiole
Dillapiole is a phenylpropanoid derivative mainly found in dill with a wide number of biological activities including anti-microbial properties, the ability to lower uric acid levels, cytotoxic, anti-inflammatory, insecticidal, anti-proliferative, anti-leishmanial, and anti-cancer effects. Dillapiole has also been reported to synergistically increase the insecticidal activity of some insecticides and a number of essential oil volatiles [21-29]. Moreover, it has been shown that when dillapiole is combined with myristicin, the synergistic effect of these compounds enhanced their ability to effectively eliminate bacteria [30]. In another study, dillapiole inhibited the production of aflatoxin G1 in Aspergillus parasiticus with the IC50 value of 0.15 µM [31]. Also, dillapiole demonstrated potent antifungal activity against Crinipellis perniciosa, dermatophytes, and non-dermatophyte fungi [32,33]. In addition, the larvicidal and insecticidal activities of dillapiole were found to control Anopheles marajoara and Aedes aegypti [32]. The synergism between dillapiole and gedunin (a tetranortriterpenoid compound) decreases malarial parasitemia caused by Plasmodium berghei. Dillapiole was also found to inhibit enzyme activity in drug metabolism [34]. Moreover, the cytotoxic effect of dillapiole is attributed to apoptosis induction [25,35]. In this regard, dillapiole up-regulated pro-apoptotic caspase-8 protein (CASP8) and down-regulated anti-apoptotic B-cell lymphoma 2 (BCL2) protein expression in RPMI 2650 cells that the involvement of CASP8 and BCL2 signaling pathways could activate apoptosis [36].
2.4. Caryophyllene oxide
Caryophyllene oxide is a sesquiterpenoid oxide found in lemon balm, eucalyptus, prickly paperbark [37], with a wide-spectrum antifungal activity in plant defense and insecticidal/antifeedant properties [38,39]. Antifungal, antibacterial, and antiplatelet aggregation activities of caryophyllene oxide have also been proven [40,41]. It is believed that caryophyllene oxide contributes to the potent cytotoxic effects observed in the essential oil of Heteropyxis dehniae leaves [42]. Caryophyllene oxide showed toxicity to Helicoverpa virescens and synergizes the growth-inhibiting effect of gossypol [43]. In addition, β-caryophyllene oxide has demonstrated anti-inflammatory, antioxidant, anticancer, antiviral, anticarcinogenic, and analgesic properties [44-49]. β-caryophyllene oxide alters various functional pathways such as mitogen-activated protein kinase, STAT3 and PI3K/AKT/mTOR/S6K1 and reduces the expression of procancer genes/proteins, while increases the levels of proteins involved in proapoptotic properties [49]. β-caryophyllene oxide could suppress the activation of NLRP3 inflammasome that could alleviate rheumatoid arthritis symptoms [50]. Caryophyllene oxide can suppress anti-oxidation in cells and induce ferritinophagy by regulating the NCOA4/FTH1/LC3 pathway in hepatocellular carcinoma. NCOA4, FTH1, and LC3Ⅱ are the targeted regulators of caryophyllene oxide that could promote ferritinophagy. This compound could also decrease tumor volume, improve LC3 and NCOA4 protein levels in tumor tissue, and increase Fe2+ and malondialdehyde in serum. Caryophyllene oxide reduces FTH1, GPX4, NRF2, and HO-1 expression that could lead to the inhibition of hydroxyl radical’s inhibitory activity and GSH in serum, the promotion of iron deposition in tumor tissue and the inhibition of tumor growth [51]. Moreover, prostaglandins, NO, and sulfhydryl groups all play a role in the gastroprotective mechanism of caryophyllene oxide [52].
2.5. Eugenol and eugenol acetate
Eugenol is a volatile bioactive phenolic monoterpenoid compound that belongs to phenylpropanoids group [53]. This compound is frequently found in a variety of aromatic plants such as clove, tulsi, cinnamon, nutmeg, and pepper [53]. The antimicrobial, anticancer, antipyretic, antidiabetic, anti hypercholesterolemic, antiparasitic, antioxidant, antiatherogenic, anti-inflammatory, and analgesic properties of eugenol have been previously reported in pharmacological studies [53-55]. The insecticidal activity of eugenol has also been exhibited against Sitophilus zeamais [56]. It was also found to be toxic and repellent to the beetle Dinoderus bifloveatus and tick (Ixodes ricinus) [56]. Moreover, eugenol contained fumigant property to Callosobruchus maculatus [56]. Eugenol acetate is an acetylated derivative of eugenol and exhibits antibacterial, anti-carcinogen, acaricide, anticandidal, antioxidant, and anti-virulence activities [57-59].
Also, eugenol shows a preventive effect on lipid peroxidation and dopamine depression through the reduction of lipid peroxidation and stimulation of reduced glutathione [60]. Anti-inflammatory activity of eugenol is also attributed to the suppression of cyclooxygenase II enzyme expression, inhibition of cytokines in macrophages, and suppression of NF-Kappa B (NF-kB). Eugenol increased lipid layer breaking in the colon cancer cells, induced apoptosis via mitochondrial membrane destruction and reactive oxygen species production [60]. In addition, eugenol could inhibit free radicals, decrease lipid peroxidation, suppress antioxidant enzymes depletion, hinder α-glucosidases, and enhance enzymes activities involved in carbohydrate metabolism. Antibacterial mechanism of eugenol is related to the induction of cell lysis via lipid and protein leakage in the cell membrane [60], disruption of bacterial cell wall and membrane, permeability and expansion of cell membrane, leakage of intracellular fluids, release of lipids and proteins, inhibition of respiration, change in bacterial ion transport patterns, disturbance of membrane proteins, and cell death [55]. Also, the antiviral mechanism of eugenol is related to the disintegration of Herpes simplex virus (HSV) envelope, induction of glutathione S-transferase expression, upregulation of glycoprotein expression of HSV that controls and blocks HSV replication. Additionally, eugenol shows antifungal activity through arrestation of cell cycle, induction of ROS accumulation in fungal cells, disruption of cell membrane integrity, alteration of fungal membrane enzymes, and changes in morphology, permeability, and membrane fluidity [55]. Moreover, eugenol significantly reduces neutrophils and TNF-α and cyclooxygenase-2 (COX-2) levels that could result in inflammatory activity of this compound. It has also been reported that prostaglandin synthesis and macrophage chemotaxis are involved in anti-inflammatory property of eugenol [55].
2.6. Carvacrol
Carvacrol is a monoterpenoid phenol compound found in the essential oil of a wide number of plants, such as oregano, thyme, pepperwort, sweet basil, black cumin and savory [61-64]. The plants with the high values of carvacrol are frequently used in traditional medicine around the globe [65,66]. Numerous studies have explored the diverse potential applications of carvacrol; these include antibacterial effects, antioxidant properties, antiseptic properties, antispasmodic activity, promotion of growth, antifungal properties, antiviral activity, anti-inflammatory effects, expectorant properties, antitussive effects, immunomodulatory effects, chemo-preventive activity, and the ability to modify rumen microbial fermentation and reduce methane emissions[67-70]. Many diverse clinical applications of carvacrol such as antimicrobial, acaricidal, antitumor, antihypertensive, antimutagenic, antigenotoxic, antiplatelet, analgesic, anti-obesity, antispasmodic, anti-inflammatory, antidepressant/anxiolytic, angiogenic, antiparasitic, AChe inhibitory, ant elastase, antinociceptive, insecticidal, immunomodulatory, spasmolytic, antihepatotoxic, vasorelaxant, and hepatoprotective activities have also been shown [71-73]. Interaction of carvacrol with Bacillus cereus membranes leads to changes in permeability and H+ and K+ ions gradients, impairment in cell processes, and cell death [74]. It has also been shown that carvacrol reduced the levels of alanine aminotransferase, lactic acid dehydrogenase, aspartate aminotransferase, pancreatitis-induced 8-hydroxydeoxyguanosine and malondialdehyde, increased enzyme activities of glutathione peroxidase, superoxide dismutase, and catalase, and improved coagulation, necrosis, and inflammation in the liver [75]. Carvacrol has anti-inflammatory activity through activation of TRPA1 receptor, decrease in inflammatory-related receptors, reduction of pro-inflammatory cytokines production, and decrease in oxygen reactive species levels. Furthermore, carvacrol could increase anti-oxidant enzymes and inhibit inducible nitric oxide synthase and pro-inflammatory mediators [76]. Additionally, anticancer mechanism of carvacrol is related to decrease in matrix metalloprotease 2 and 9, cyclin B1 expression, and cell-proliferation and also arrestation of cell cycle G2 via increase in apoptosis, caspase-3 activation, PARP cleavage, and decrease in Bcl-2 gene expression [76].
2.7. Non-volatile compounds
Extracts of R. michauxii were examined during the three growth stages to characterize its constituents [10]. Many different polyphenols were found, including phenylethanoid glycosides, labdane diterpenes, O- and C-glycosylated flavonoids, and iridoids. The labdane diterpenes were the dominant compounds during the vegetative and flowering stages, making up 20% of the total extract.
The phytochemicals identified in the extracts contained chemically heterogeneous compounds, including iridoid, phenolics, labdane derivatives, lignan medioresinol, and antirrhinoside. The findings indicated the existence of flavonol glycosides, both C and O derivatives, and phenols, particularly derivatives of caffeic acid. The extracts of the three stages yielded several polyphenols, including hydroxycinnamic and phenylethanoid derivatives, gallic acid derivatives, and O-glycosylated as well as C-glycosylated flavonoids. Only during the vegetative and flowering stages, terpene derivatives cloud be detected.
A number of phenylethanoid glycosides were found in the extracts, including echinacoside, isoverbascoside, O-glycosylated/C-glycosylated flavonoids, verbascoside, apigenin, luteolin derivatives, kaempferol. O-glycosylated and C-glycosylated flavonoids, mainly apigenin and luteolin derivatives, could not be detected during the dormant stage. More phytochemicals were present during the vegetative and flowering stages compared with the dormant plants. Among the extracts, the flowering extract contained the highest levels of flavonoids and labdane derivatives [10].
3. Pharmacological activities
R. michauxii has been demonstrated to have antibacterial, antifungal, antiviral, antioxidant, antimalarial, cytotoxic, and lipid-lowering properties.
3.1. Antimicrobial activity
In our previous study, we conducted an evaluation of the antimicrobial properties of R. michauxii methanolic extract during the dormant, vegetative, and flowering stages. We tested the extract against viruses [Tobacco mosaic virus (TMV) and Cucumber mosaic virus (CMV)], Gram-positive bacteria (Bacillus subtilis, Clavibacter michiganensis, Staphylococcus aureus, and Curtobacterium flaccumfaciens), Gram-negative bacteria (Xanthomonas citri, Pseudomonas syringae pv. syringae, Agrobacterium tumefaciens, and Escherichia coli), and fungi (Bipolaris micropus, Fusarium oxysporum, F. solani, and Rhizoctonia solani). The MICs of the extracts ranged from 8 to 64 mg/mL and MBCs between 32 to more than 128 mg/mL [77].
Gram-positive bacteria, i.e., C. flaccumfaciens and C. michiganensis, had the lowest MBC value (32 mg/ml). B. subtilis exhibited the highest sensitivity and the lowest average MIC (13.33 mg/mL), while E. coli and S. aureus showed greater resistance: the highest average MIC (53.33 mg/mL). During dormant, vegetative, and flowering stages, methanolic extracts of R. michauxii were evaluated for antifungal activity against Bipolaris micropus, Rhizoctonia solani, Fusarium oxysporum, and Fusarium solani. Based on the average MICs (R. michauxii extracts), F. oxysporum as well as R. solani showed the greatest resistance (24 mg/mL) and sensitivity (18.67 mg/mL) to the extracts [78].
A half-leaf bioassay demonstrated that methanolic extracts of R. michauxii inhibited tobacco mosaic virus (TMV) infection based on the number of local lesions during the dormant (51.88%), vegetative (34.07%), and flowering stages (36.52%). In addition, plant extracts inhibited cucumber mosaic virus (CMV) infection at all three growth stages by 44.84%, 29.24%, and 20.92%. After treatment with the plant extract (dormant stage), the average number of local lesions caused by CMV infection was 11.8 ± 3.19, in comparison with 23 ± 3.7 in the control samples. After applying the plant extract at the dormant stage, there were 11.8 ± 4.04 mean local lesions in response to TMV infection, while that of the control was 21.6 ± 2.08. The plant extracts showed no inhibitory effects on CMV and TMV infections according to leaf disc method [77]. Based on these results, TMV and CMV could be inactivated before entering host cells by the plant extract. The plant extract had the strongest antibacterial and antiviral effects at the dormant stage.
Pathogens are inhibited by secondary metabolites produced by antagonistic bacteria [79,80]. A number of previous studies indicate that medicinal plants' antimicrobial activity is influenced by antagonistic endophytes [81]. Consequently, the strong antibacterial properties of the R. michauxii root could be attributed to endophytic microorganisms colonizing plant roots during the dormant stage.
3.2. Antimicrobial activity
3.3. Antioxidant activity
Methanolic extracts of R. michauxii were found to have ferric reducing antioxidant activity of 26.7 ± 1.3 μM/g DW and an IC50 2,2-diphenyl-1-picrylhydrazyl (DPPH) scavenging activity of 465.4 ± 76.6 μg DW/mL [84]. For our study, antioxidant activity was determined through a radical scavenging assay. The vegetative and flowering stages of R. michauxii exhibited the highest antioxidant activity: 91.7% and 91.52% (DPPH scavenging activity), respectively. In the dormant stage, this parameter was 22.15% (unpublished data).
3.4. Cytotoxic activity
The methanolic extracts of R. michauxii were found to have the lowest IC50 levels: 1.24 mg/mL and 1.23 mg/mL during the vegetative and flowering stages. Thus, the dormant stage had the lowest cytotoxic action (IC50 of 3.2 mg/mL) on MCF-7 breast adenocarcinoma cells, while the flowering/vegetative stages had the highest cytotoxic effects on tumor cells [77].
Cell viability decreased dose-dependently due to the cytotoxic activity of the plant extracts. Compared to the flowering and vegetative stages, the dormant stage had higher levels of cell viability. This extract exhibited the highest level of cytotoxicity during the flowering stage at 3 mg/mL with an inhibition of cell growth of 97.2%. On the other hand, cytotoxicity was lowest during the dormant stage at 2 mg/mL with a 25.3% inhibition of cell growth [77].
In MCF-7 cells treated with plant extracts, microscopic examination revealed that the vegetative and flowering stages had the most cytotoxic effects and cell death [77]. In another study, dichloromethane extract of R. michauxii showed IC50 of 0.73, 0.67 and 0.68 mg/mL on oral squamous cell carcinoma cell line after 24, 48, and 72 hours [85]. The cytotoxic activity of the methanolic extract of R. michauxii was evaluated against WEHI-164, MCF-7, MDBK, A-549, and HepG-2 cell lines. The results showed IC50 > 50 (μg/ mL) against the cell lines [86].
3.5. Lipid lowering activity
In an in vitro model, isolated compounds from R. michauxii extract were evaluated for their ability to lower cholesterol levels. The separated compounds were tested on the low-density lipoprotein receptors (LDLR) and the proprotein convertase subtilisin/kexin type 9 (PCSK9) in the human hepatocarcinoma cell line Huh7. The aim was to investigate any potential hypocholesterolemic effects of these compounds. After applying a number of promising compounds, cholesterol levels in the same cell lines were assessed. Some compounds identified in R. michauxii extracts significantly lowered intracellular cholesterol. Among them, antirrhinoside exhibited the strongest induction of LDLR and PCSK9 at a concentration of 50μM. According to the in vitro data, antirrhinoside appears to lower cholesterol through a mechanism similar to statins. Our findings highlight antirrhinoside as an effective inducer of low-density lipoprotein receptors (LDLR), resulting in a substantial decrease in intracellular cholesterol levels within hepatic cells. This suggests new pathways through which iridoids can potentially reduce cholesterol. [10].
Previous literature has highlighted the beneficial role of iridoid compounds in lipid metabolism and the formation of atherosclerotic plaques. Building upon this knowledge, our in vitro findings have identified another compound within the same chemical category that may possess hypocholesterolemic activity. This compound appears to operate through a mechanism similar to statins, potentially inhibiting the mevalonate pathway (as depicted in Figure 4). It is plausible that this inhibition is responsible for the observed induction of both proprotein convertase subtilisin/kexin type 9 (PCSK9) and LDL receptors, similar to the effects of the HMG-CoA reductase inhibitor simvastatin. However, further studies are necessary to determine the precise mode of action in an in vivo model [10].
4. Pharmacological activities of non-volatile constituents of R. michauxii
The LC-MS analysis detected several compounds in R. michauxii methanolic extract [10]. Some of these compounds have been extensively studied in various articles and have been the subject of several review articles. Notable examples include Verbascoside [87,88], Echinacoside [89,90], Chlorogenic acid [91], Hydroxycinnamic acid [92], Luteolin [93,94], Apigenin [95], and Kaempferol [96]. Therefore, other detected constituents such as Medioresinol (1), Martynoside (2), Antirrhinoside (3), Isoverbascoside (4), and Apigenin-7-O-glucoside (5) (Figure 5) were discussed for their biological activities here.
4.1. Medioresinol (MD)
Medioresinol is a furofuran-type lignin, which is identified in some plants such as Sambucus williamsi [97], Cinnamomi cortex Spissus [98], Dirca occidentalis [99], etc.
MD has been investigated for its antimicrobial activity. The results of antibacterial susceptibility testing demonstrated its remarkable effectiveness against Enterococcus faecium, Staphylococcus aureus, Cutibacterium acnes, Escherichia coli O157:H7, Escherichia coli, and Pseudomonas aeruginosa with MIC values ranging from 2.5 to 20.0 μg/mL compared with Ampicillin, cefotaxime, and chloramphenicol antibiotics. In the broth microdilution checkerboard procedure, the combination of MD and conventional antibiotics specially cefotaxime exhibited synergistic interactions against all strains even antibiotics-resistant P. aeruginosa, whereas allcombinations showed nointeraction against E. faecium [100].
MD has also shown significant antifungal activity. The results of the MTT assay demonstrated its effectiveness against Candida albicans, Candida parapsilopsis, Trichosporon beigelii, and Malassezia furfur with MIC value of 3.125 mg/mL. MD induces cell cycle arrest at the G0/G1 phase in C. albicans cells. This arrest is accompanied by the accumulation of intracellular reactive oxygen species (ROS). MD was found to induce apoptosis in C. albicans cells by mitochondrial membrane depolarization, the release of cytochrome c, and plasma membrane phosphatidylserine externalization. In the late stages of MD-induced apoptosis, apoptotic changes were observed in the nucleus and DNA [97].
MD, which is previously isolated from Doliocarpus dentatus, displayed a leishmanicidal effect against amastigotes of the Guyanan strain H142 of Leishmania amazonemis with the index of survival of the amastigotes 40% at 60 μg/mL [101].
Pyroptosis is a type of cell death with the release of inflammatory factors. It’s revealed that MD inhibits pyroptosis, and consequently ischemic injury in brain microvascular endothelial cells due to enhancement of phenylalanine and inhibition of mtROS production through PGC-1α/PPARα/GOT1 axis pathway [102].
4.2. Martynoside (MR)
Martynoside, a reported phenyl propanoid glycoside from R. michauxii, was previously isolated from different plant species, such as Martynia louisiana [103], Boscia albitrunca [104], Ballota nigra [105], and Pedicularis alashanica [106].
One study confirmed MR scavenging capability on various radicals such as N3∙, Br2∙-, and superoxide radical anion O2∙-. The results of this study showed that MR had better singlet oxygen quenching capacity than extensively studied antioxidants such as ferulic acid, quercetin, and kaempferol. Moreover, electron reduction ability of MR radical was comparable with those of antioxidants such as hydroxycinnamic acid derivative radicals [107]. Intense contraction of skeletal muscle is associated with ROS formation leading to fatigue. MR which hasantioxidant and ROS scavenging abilities, could alleviate fatigue by retarding the contractility of the Bufo gastrocnemius muscle [108].
Furthermore, the results of the experiments on animals undergoing exercise, have suggested that MR had the potential to antagonize sports anemia. The basic hematological parameters such as red blood cells, hemoglobin, and hematocrit levels increased in the groups treated with MR in comparison to the untreated group, while the mean corpuscular hemoglobin concentration, the mean corpuscular hemoglobin plasma, the plasma malonyl dialdehyde levels, and percentage of the abnormally shaped red blood cells in the MR-treated group were lower than the untreated samples [109].
Prescribing 5-fluorouracil (5-FU), a chemotherapeutic drug, is associated with myelotoxicity as the primary dose-limiting cytotoxicity. The hematopoietic activity of MR was found against 5-FU-induced bone marrow cytotoxicity ex vivo and in vivo. The data obtained from m-RNA sequencing and transcriptome-based data analysis showed that the molecular mechanisms of Matynoside against 5-FU-induced bone marrow cytotoxicity could be increased number and survival of bone marrow nucleated cells, as well as the improvement of the bone marrow microenvironment [110].
Information is available on the estrogen receptor modulation of Matynoside. Its estrogenic/antiestrogenic activity via estrogen receptor alpha (ERα) and beta (ERβ) isoforms was assessed in different cells, including breast cancer cells (MCF7), endometrial cancer cells (Ishikawa), and osteoblasts (KS483). In breast cancer, Matynoside revealed an antiestrogenic effect, while the induction of nodule mineralization in osteoblasts was associated with its esterogenic effect. Furthermore, it showed an antiproliferative effect in endometrial cells [111].
4.3. Antirrhinoside
Antirrhinoside is an iridoid glycoside, known in the Scrophulariaceae and has been reported from various plants for example Asarina scandens [112], Antirrhinum majus L. [113], Kickxia spuria subsp. integrifolia [114], and etc.
Antirrinoside displayed significant inflammation inhibition in different edema models such as Carrageenan-induced hind paw, PGE2-induced hind paw, and TPA-induced mouse ear edema. Moreover, the anti-nociceptive effect of Antirrinoside was found in the p-benzoquinone-induced abdominal constriction test [115].
The findings of the antifungal activity assay showed that Antirrhinoside drastically inhibited aflatoxin B1 production in Aspergillus flavus Link, so may be used as a preservative in food products [116].
4.4. Isoverbascoside (IVer)
IVer is a phenylpropanoid glycoside derivative, identified as a positional isomer of Verbascoside, which is the most abundant compound of numerous plants [117].
Anti-oxidant activity of IVer was determined in vitro using two methods: 1,1-diphenyl-2-picrylhydrazyl (DPPH) and hydrogen peroxide (H2O2). Its free radical scavenging property was comparable to that of ascorbic acid, a strong antioxidant agent, in the DPPH assay. In addition, IVer exhibited higher potency than ascorbic acid in scavenging H2O2 [118].
Transforming growth factor-β1 (TGF-β1) is a cytokine that induces fibrotic activity through Smad and non-Smad pathways. Pharmaceutical research has suggested that IVer exhibits anti-fibrosis potential by inhibiting the TGF-β1 signaling pathway and collagen I production in murine (MLg 2908) and human primary lung cells [119].
IVer has been shown to enhance glucose uptake and subsequently ameliorate insulin resistance in L6 muscle cells by affecting signaling pathways such as protein kinase B (AKT), AMP-activated protein kinase (AMPK), and mammalian target of rapamycin (mTOR) regulating energy homeostasis, and is involved in insulin receptor activation. Furthermore, IVer mitigates ROS-mediated oxidative damage, which is a key process in insulin resistance, through the downregulation of p38 mitogen-activated protein kinase (p38 MAPK)/ c-Jun N-terminal kinase (JNK) pathway [120].
The results of semi-soft agar colony assay, a method which estimate tumorigenicity of cancer cells, revealed that IVer is capable of inducing differentiation in human hepatocarcinoma cells (SMMC-7721). Vervascoside-induced differentiation was associated with several phenomena, including arresting G0/G1 phase, suppressing γ-glutamyltransferase activity (a marker enzyme of hepatocarcinoma cells), and elevating tyrosine aminotransferase activity (an indicator of differentiated hepatic cells) [121]. In addition, in human gastric cancer cell line MGC 803, IVer inhibited cell proliferation, induced differentiation and G0/G1 arrest, as well as suppressed alkaline phosphatase (ALP), and lactate dehydrogenase (LDH) [122]. The differentiation potential of IVer is also evidenced in HL-60 cells, as observed through morphological changes, increasing phagocytosis, and reducing tumorigenesis [123].
Neutrophils play an important role in inflammation-related pathological conditions. A study investigated the effect of IVer on neutrophil function and found that it exerted significant effects. IVer inhibited TLR2 and TLR4-mediated apoptosis, as well as the expression of the adhesion molecule CD11b, chemokine receptor CXCR2, and p38 phosphorylation. Furthermore, IVer increased the production of Tumor Necrosis Factor (TNF-α) in purified murine bone marrow neutrophils. Importantly, all these effects were dependent on the activation level and functional state of neutrophils, highlighting the therapeutic potential of IVer in inflammatory conditions [124].
4.5. Apigenin-7-O-glucoside (AG)
Apigenin-7-O-glucoside is flvonoid compound predominantly found in Asteraceae plants [125]. It has been shown to possess many therapeutic potentials, including antioxidant, anti-inflammatory, anti-spasmodic, anti-viral, anti-thrombotic, and anti-angiogenic properties, primarily attributed to the aglycone portion of AG [126].
Anti-oxidant and anti-inflammatory effects of AG have been investigated in different experimental systems and compared with those of Trolox, a vitamin E analogue. These two compounds exerted the same inhibition effect on H2O2-induced ROS production in RAW264.7 cells, while AG showed higher inhibitory effects on LPS-induced inflammation in RAW246.7 cells than Trolox [127].
The anti-inflammatory effects of AG in alleviating colitis were investigated in mice with dextran sulfate sodium (DSS)-induced colitis. Administration of AG led to an increase in colon length, improvement in colonic histopathology, and modulation of gut microbiota. Furthermore, the suppression of p38, ERK, and JNK phosphorylation levels indicated the involvement of the Mitogen-activated protein kinases (MAPKs) pathway in AG's protective effect against DSS-induced colitis [110].
A study investigated the effect of AG on human cervical cancer (HeLa) cells. AG demonstrated anti-cancer activity by inducing G0/G1 phase arrest, promoting apoptosis through the inhibition of the PTEN/PI3K/AKT pathway, and inhibiting cell migration through the matrix metalloproteinase 2 and 9 pathway [128].
The evaluation of AG on 14 different species of Candida spp. yielded significant results, demonstrating its substantial antifungal potential. The suggested underlying mechanism for this effect was the inhibition of both extracellular and intracellular ROS production [129].
Antiparasitic activity of AG alone or in combination with pyrimethamine, a dihydrofolate inhibitor, against Toxoplasma gondii growth was examined using high throughput antiparasitic drug screening cell-based assay. AG plus pyrimethamine showed additive parasitic inhibition, as well as highly selective inhibition against the parasite versus human foreskin fibroblast cell [130].
The yeast DEL assay demonstrated the potent anti-mutagenic activity of AG against mutagens (ethyl methanesulfonate, acridine, 9-aminoacridine, and sodium azide) in Salmonella typhimurium TA1535 and TA1537 strains and Saccharomyces cerevisiae RS112 strain [131].
The crystal violet staining method revealed the strong inhibitory activity of AG against bacterial biofilm formation and bacterial activity in Staphylococcus aureus and Escherichia coli. AG inhibited biofilm formation by targeting factors such as exopolysaccharide production, quorum sensing, and cell surface hydrophobicity, which are crucial for the formation of bacterial biofilms [132].
5. Conclusion
Our review presents compelling evidence for the diverse biological and pharmacological properties of R. michauxii. Specifically, we discussed antimicrobial, antioxidant, antimalarial, cytotoxic, and lipid lowering properties of the species. The plant's abundance of phytochemicals with various pharmacological effects holds great promise for the food and pharmaceutical industries. Given the current trend of embracing natural compounds, R. michauxii exhibits potential applications in the fields of food and human health. Additionally, exploring the interactions among major bioactive compounds in pharmacological applications is crucial, as synergistic effects may enhance therapeutic benefits. Synthesizing analogues of promising natural products could also improve the drug-like properties of these compounds. However, considering the wide variety of compounds and pharmacological properties found in this plant, further research is necessary to investigate its toxicology, elucidate unknown properties, demonstrate its clinical efficacy, delve into its detailed spectrum of activity, and unravel the underlying signaling pathways, associated genes, and mechanisms of action.
Author Contributions
Conceptualization, A.T., and C.W.L.; methodology, A.T., B.J. and V.A.; writing—original draft preparation, A.T., B.J. and V.A.; writing—review and editing, A.T., C.W.L. and H.J.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture and Forestry (IPET) through Agricultural Machinery/Equipment Localization Technology Development Program, the Ministry of Agriculture, Food and Rural Affairs (MAFRA-321057052HD030 to C.W.L.).
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Halberstein, R.A. Medicinal plants: historical and cross-cultural usage patterns. Annals of epidemiology 2005, 15, 686–699. [Google Scholar] [CrossRef] [PubMed]
- Katz, L.; Baltz, R.H. Natural product discovery: past, present, and future. Journal of Industrial Microbiology and Biotechnology 2016, 43, 155–176. [Google Scholar] [PubMed]
- Tamokou, J.; Mbaveng, A.; Kuete, V. Antimicrobial activities of African medicinal spices and vegetables. In Medicinal spices and vegetables from Africa; Elsevier: 2017; pp. 207-237.
- Kokkini, S.; Karousou, R.; Hanlidou, E. HERBS| Herbs of the Labiatae. 2003.
- Mabberley, D.J. The plant-book: a portable dictionary of the vascular plants; Cambridge university press: 1997.
- Scheen, A.-C.; Albert, V.A. Nomenclatural and taxonomic changes within the Leucas clade (Lamioideae; Lamiaceae). Systematics and Geography of Plants 2007, 229–238. [Google Scholar]
- Ghahremaninejad, F.; Hoseini, E. Identification of Medicinal and Aromatic Plants of Iran, Valiollah Mozaffarian. Farhang Moaser Publishers, Tehran (2012). 1444 pp., 2470 colored images (Language: Mainly Persian with English preface and several indexes). Hardback, ISBN: 978-600-1050-31-2. Format: 23.5× 15.5 cm. Price: 900000 IR-Rials. 2015.
- Javidnia, K.; Miri, R.; Soltani, M.; Khosravi, A. Chemical constituents of the essential oil of Otostegia michauxii Briq. from Iran. Journal of Essential Oil Research 2010, 22, 1–2. [Google Scholar] [CrossRef]
- Karami, A.; Esmaeili, H.; Shahbazian, D. Essential oil composition of Rydingia michauxii (Briq.) Scheen & VA Albert endemic of Iran. Natural product research 2018, 32, 229–233. [Google Scholar]
- Sut, S.; Tahmasebi, A.; Ferri, N.; Ferrarese, I.; Rossi, I.; Panighel, G.; Lupo, M.G.; Maggi, F.; Karami, A.; Dall’Acqua, S. NMR, LC-MS Characterization of Rydingia michauxii Extracts, Identification of Natural Products Acting as Modulators of LDLR and PCSK9. Molecules 2022, 27, 2256. [Google Scholar] [CrossRef]
- Sadeghian, S.; Hatami, A.; Hamzeh’ee, B. CHROMOSOME COUNTS OF SIX TAXA OF LAMIACEAE FROM IRAN. The Iranian Journal of Botany 2021, 27, 58–61. [Google Scholar]
- Rosselli, S.; Fontana, G.; Bruno, M. A review of the phytochemistry, traditional uses, and biological activities of the Genus Ballota and Otostegia. Planta medica 2019, 85, 869–910. [Google Scholar] [CrossRef]
- Sadeghi, Z.; Akaberi, M.; Valizadeh, J. Otostegia persica (Lamiaceae): A review on its ethnopharmacology, phytochemistry, and pharmacology. Avicenna journal of phytomedicine 2014, 4, 79. [Google Scholar]
- Khan, S.; Syed, F. Bioactive constituents from genus Otostegia. SARJ of Physical Sci 2013, 1, 15–25. [Google Scholar]
- Safa, O.; Soltanipoor, M.A.; Rastegar, S.; Kazemi, M.; Dehkordi, K.N.; Ghannadi, A. An ethnobotanical survey on hormozgan province, Iran. Avicenna journal of phytomedicine 2013, 3, 64. [Google Scholar] [PubMed]
- Toori, M.A.; Joodi, B.; Sadeghi, H.; Sadeghi, H.; Jafari, M.; Talebianpoor, M.S.; Mehraban, F.; Mostafazadeh, M.; Ghavamizadeh, M. Hepatoprotective activity of aerial parts of Otostegia persica against carbon tetrachloride-induced liver damage in rats. Avicenna journal of phytomedicine 2015, 5, 238. [Google Scholar]
- Tofighi, Z.; Ostad, S.; Khezrrahdoost, S.; Salehizadeh, H.; Yassa, N. Potent anti-nociceptive and anti-inflammatory effects of methanol fraction of Otostegia persica extract and its components. Research Journal of Pharmacognosy 2017, 4, 23–29. [Google Scholar]
- Ganjali, A.; Sotoudeh, A.; Jahanshahi, A.; Takhtfooladi, M.A.; Bazzazan, A.; Roodbari, N.; Harati, M.P. Otostegia persica extraction on healing process of burn wounds. Acta Cirurgica Brasileira 2013, 28, 407–411. [Google Scholar] [CrossRef]
- Safaeian, L.; Ghasemi-Dehkordi, N.; Javanmard, S.H.; Namvar, H. Antihypertensive and antioxidant effects of a hydroalcoholic extract obtained from aerial parts of Otostegia persica (Burm.) Boiss. Research in Pharmaceutical Sciences 2015, 10, 192. [Google Scholar]
- Tahmasebi, A.; Hosseini, S.M.; Karami, A.; Afsharifar, A.; Sharifi Olounabadi, A.R. Variation in essential oil composition of Rydingia michauxii at the three developmental stages. Natural product research 2021, 35, 342–345. [Google Scholar] [CrossRef]
- Handa, S.; Dewan, R. Evaluation of dillapiole and dihydrodillapiole as synergists for pyrethrins in dust formulations. Pyrethrum post 1975. [Google Scholar]
- Tomar, S.S.; Maheshwari, M.L.; Mukerjee, S.K. Syntheses and synergistic activity of some pyrethrum synergists from dillapiole. Agricultural and Biological Chemistry 1979, 43, 1479–1483. [Google Scholar]
- Tomar, S.; Maheshwari, M.; Mukerjee, S. Synthesis and synergistic activity of dillapiole based pyrethrum synergists. Journal of Agricultural and Food chemistry 1979, 27, 547–550. [Google Scholar] [CrossRef]
- Bernard, C.B.; Arnason, J.; Philogene, B.; Lam, J.; Waddell, T. In vivo effect of mixtures of allelochemicals on the life cycle of the European corn borer, Ostrinia nubilalis. Entomologia experimentalis et applicata 1990, 57, 17–22. [Google Scholar] [CrossRef]
- Chan, W.-H. Cytotoxic effects of dillapiole on embryonic development of mouse blastocysts in vitro and in vivo. International Journal of Molecular Sciences 2014, 15, 10751–10765. [Google Scholar] [CrossRef] [PubMed]
- Parise-Filho, R.; Pastrello, M.; Pereira Camerlingo, C.E.; Silva, G.J.; Agostinho, L.A.; de Souza, T.; Motter Magri, F.M.; Ribeiro, R.R.; Brandt, C.A.; Polli, M.C. The anti-inflammatory activity of dillapiole and some semisynthetic analogues. Pharmaceutical Biology 2011, 49, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Musdja, M.Y.; Asmara, D.; Musir, A. Effect of ethanol extract of Foeniculum Vulgare Mill on inhibition of uric acid crystals formation in male rats. 2018.
- Parise-Filho, R.; Pasqualoto, K.F.M.; Magri, F.M.M.; Ferreira, A.K.; da Silva, B.A.V.G.; Damião, M.C.F.C.B.; Tavares, M.T.; Azevedo, R.A.; Auada, A.V.V.; Polli, M.C. Dillapiole as antileishmanial agent: discovery, cytotoxic activity and preliminary SAR studies of dillapiole analogues. Archiv der Pharmazie 2012, 345, 934–944. [Google Scholar] [CrossRef] [PubMed]
- AMIN, I.M.; RUSLAN, N.B.; ZULKIPLI, Z.A.; ZAKARIA, N.A.; JALIL, M.T.M.; ARIS, F. CYTOTOXIC EFFECT OF DILLAPIOLE ON HUMAN BREAST CANCER MCF-7 CELLS. Malaysian Applied Biology 2022, 51, 29–35. [Google Scholar] [CrossRef]
- Brazao, M.A.B.; Brazao, F.V.; Guilherme, J.; Monteiro, M.C. Antibacterial activity of the Piper aduncum oil and dillapiole, its main constituent, against multidrug-resistant strains. Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas 2014, 13, 517–526. [Google Scholar]
- Razzaghi-Abyaneh, M.; Yoshinari, T.; Shams-Ghahfarokhi, M.; Rezaee, M.-B.; Nagasawa, H.; Sakuda, S. Dillapiol and apiol as specific inhibitors of the biosynthesis of aflatoxin G1 in Aspergillus parasiticus. Bioscience, biotechnology, and biochemistry 2007, 71, 2329–2332. [Google Scholar] [CrossRef]
- de Almeida, R.R.; Souto, R.N.; Bastos, C.N.; da Silva, M.H.; Maia, J.G. Chemical variation in Piper aduncum and biological properties of its dillapiole-rich essential oil. Chemistry & biodiversity 2009, 6, 1427–1434. [Google Scholar]
- Ferreira, R.; Monteiro, M.; Silva, J.; Maia, J. Antifungal action of the dillapiole-rich oil of Piper aduncum against dermatomycoses caused by filamentous fungi. British Journal of Medicine and Medical Research 2016, 15, 1–10. [Google Scholar] [CrossRef]
- Morais, V.P.; Cabral, F.V.; Fernandes, C.C.; Miranda, M.L.D. Brief Review on Piper aduncum L., its Bioactive Metabolites and its Potential to Develop Bioproducts. Brazilian Archives of Biology and Technology 2023, 66. [Google Scholar] [CrossRef]
- Ferreira, A.K.; de-Sá-Júnior, P.L.; Pasqualoto, K.F.M.; de Azevedo, R.A.; Câmara, D.A.D.; Costa, A.S.; Figueiredo, C.R.; Matsuo, A.L.; Massaoka, M.H.; Auada, A.V.V. Cytotoxic effects of dillapiole on MDA-MB-231 cells involve the induction of apoptosis through the mitochondrial pathway by inducing an oxidative stress while altering the cytoskeleton network. Biochimie 2014, 99, 195–207. [Google Scholar] [CrossRef]
- Ruslan, N.B. Dillapiole effects on apoptosis in human nasal epithelial carcinoma, RPMI 2650 cells involves BCL-2 and caspase-8 signalling pathway. Universiti Teknologi MARA, 2021.
- Farag, R.; Shalaby, A.; El-Baroty, G.; Ibrahim, N.; Ali, M.; Hassan, E. Chemical and biological evaluation of the essential oils of different Melaleuca species. Phytotherapy Research: An International Journal Devoted to Pharmacological and Toxicological Evaluation of Natural Product Derivatives 2004, 18, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Bettarini, F.; Borgonovi, G.; Fiorani, T.; Gagliardi, I.; Caprioli, V.; Massardo, P.; Ogoche, J.; Hassanali, A.; Nyandat, E.; Chapya, A. Antiparasitic compounds from East African plants: Isolation and biological activity of anonaine, matricarianol, canthin-6-one and caryophyllene oxide. International Journal of Tropical Insect Science 1993, 14, 93–99. [Google Scholar] [CrossRef]
- Langenheim, J.H. Higher plant terpenoids: a phytocentric overview of their ecological roles. Journal of chemical ecology 1994, 20, 1223–1280. [Google Scholar] [CrossRef]
- Russo, E.B.; Marcu, J. Cannabis pharmacology: the usual suspects and a few promising leads. Advances in pharmacology 2017, 80, 67–134. [Google Scholar] [PubMed]
- Schmidt, E.; Bail, S.; Friedl, S.M.; Jirovetz, L.; Buchbauer, G.; Wanner, J.; Denkova, Z.; Slavchev, A.; Stoyanova, A.; Geissler, M. Antimicrobial activities of single aroma compounds. Natural product communications 2010, 5, 1934578X1000500906. [Google Scholar] [CrossRef]
- Bodiba, D.; Lall, N. Heteropyxis dehniae. In Underexplored Medicinal Plants from Sub-Saharan Africa; Elsevier: 2020; pp. 167-171.
- Morgan, E.D.; Wilson, I.D. Insect hormones and insect chemical ecology. ChemInform 2001, 32, no. [Google Scholar]
- Tung, Y.-T.; Chua, M.-T.; Wang, S.-Y.; Chang, S.-T. Anti-inflammation activities of essential oil and its constituents from indigenous cinnamon (Cinnamomum osmophloeum) twigs. Bioresource technology 2008, 99, 3908–3913. [Google Scholar] [CrossRef]
- Hammami, S.; Jmii, H.; El Mokni, R.; Khmiri, A.; Faidi, K.; Dhaouadi, H.; El Aouni, M.H.; Aouni, M.; Joshi, R.K. Essential oil composition, antioxidant, cytotoxic and antiviral activities of Teucrium pseudochamaepitys growing spontaneously in Tunisia. Molecules 2015, 20, 20426–20433. [Google Scholar] [CrossRef]
- Zheng, G.-Q.; Kenney, P.M.; Lam, L.K. Sesquiterpenes from clove (Eugenia caryophyllata) as potential anticarcinogenic agents. Journal of natural products 1992, 55, 999–1003. [Google Scholar] [CrossRef]
- Singh, T.P.; Singh, R.K.; Malik, P. Analgesic and anti-inflammatory activities of Annona squamosa Linn bark. J. Scientific and Innov. Res 2014, 3, 60–64. [Google Scholar] [CrossRef]
- Jun, N.J.; Mosaddik, A.; Moon, J.Y.; Ki-Chang, J.; Dong-Sun, L.; Ahn, K.S.; Cho, S.K. Cytotoxic activity of [beta]-Caryophyllene oxide isolated from jeju guava (Psidium cattleianum sabine) leaf. Records of Natural Products 2011, 5, 242. [Google Scholar]
- Fidyt, K.; Fiedorowicz, A.; Strządała, L.; Szumny, A. β-caryophyllene and β-caryophyllene oxide—natural compounds of anticancer and analgesic properties. Cancer medicine 2016, 5, 3007–3017. [Google Scholar] [CrossRef]
- Li, W.-x.; Qian, P.; Guo, Y.-t.; Gu, L.; Jurat, J.; Bai, Y.; Zhang, D.-f. Myrtenal and β-caryophyllene oxide screened from Liquidambaris Fructus suppress NLRP3 inflammasome components in rheumatoid arthritis. BMC Complementary Medicine and Therapies 2021, 21, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Z.; Zhu, Y.; Han, J.; Li, Y.; Yang, X.; Yang, G.; Song, G.; Li, S.; Li, Y.; Cheng, C. Caryophyllene oxide induces ferritinophagy by regulating the NCOA4/FTH1/LC3 pathway in hepatocellular carcinoma. Frontiers in Pharmacology 2022, 13, 930958. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Mendoza, M.E.; Cruz-Antonio, L.; GuadalupeCupido-Sánchez, M.; GarcíaCastillo, G.; Arrieta, J. Gastroprotective activity of caryophyllene oxide: the role of nitric oxide, prostaglandins and sulfhydryls. Journal of Applied Pharmaceutical Science 2014, 4, 001–005. [Google Scholar]
- Sharma, A.; Bhardwaj, G.; Sohal, H.S.; Gohain, A. Eugenol. In Nutraceuticals and health care; Elsevier: 2022; pp. 177-198.
- Abdou, A.; Elmakssoudi, A.; El Amrani, A.; JamalEddine, J.; Dakir, M. Recent advances in chemical reactivity and biological activities of eugenol derivatives. Medicinal Chemistry Research 2021, 30, 1011–1030. [Google Scholar] [CrossRef]
- Nisar, M.F.; Khadim, M.; Rafiq, M.; Chen, J.; Yang, Y.; Wan, C.C. Pharmacological properties and health benefits of eugenol: A comprehensive review. Oxidative Medicine and Cellular Longevity 2021, 2021. [Google Scholar]
- Tripathi, A.K.; Mishra, S. Plant monoterpenoids (prospective pesticides). In Ecofriendly pest management for food security; Elsevier: 2016; pp. 507-524.
- Musthafa, K.S.; Hmoteh, J.; Thamjarungwong, B.; Voravuthikunchai, S.P. Antifungal potential of eugenyl acetate against clinical isolates of Candida species. Microbial pathogenesis 2016, 99, 19–29. [Google Scholar] [CrossRef]
- Carrasco A, H.; Espinoza C, L.; Cardile, V.; Gallardo, C.; Cardona, W.; Lombardo, L.; Catalán M, K.; Cuellar F, M.; Russo, A. Eugenol and its synthetic analogues inhibit cell growth of human cancer cells (Part I). Journal of the Brazilian Chemical Society 2008, 19, 543–548. [Google Scholar] [CrossRef]
- Pasay, C.; Mounsey, K.; Stevenson, G.; Davis, R.; Arlian, L.; Morgan, M.; Vyszenski-Moher, D.; Andrews, K.; McCarthy, J. Acaricidal activity of eugenol based compounds against scabies mites. PloS one 2010, 5, e12079. [Google Scholar] [CrossRef] [PubMed]
- Nejad, S.M.; Özgüneş, H.; Başaran, N. Pharmacological and toxicological properties of eugenol. Turkish journal of pharmaceutical sciences 2017, 14, 201. [Google Scholar] [CrossRef] [PubMed]
- Alagawany, M.; Farag, M.R.; Abdelnour, S.A.; Elnesr, S.S. A review on the beneficial effect of thymol on health and production of fish. Reviews in Aquaculture 2021, 13, 632–641. [Google Scholar] [CrossRef]
- Bayir, A.G.; Kiziltan, H.S.; Kocyigit, A. Plant family, carvacrol, and putative protection in gastric cancer. In Dietary interventions in gastrointestinal diseases; Elsevier: 2019; pp. 3-18.
- Salehi, B.; Mishra, A.P.; Shukla, I.; Sharifi-Rad, M.; Contreras, M.d.M.; Segura-Carretero, A.; Fathi, H.; Nasrabadi, N.N.; Kobarfard, F.; Sharifi-Rad, J. Thymol, thyme, and other plant sources: Health and potential uses. Phytotherapy research 2018, 32, 1688–1706. [Google Scholar]
- Sharifi-Rad, M.; Varoni, E.M.; Iriti, M.; Martorell, M.; Setzer, W.N.; del Mar Contreras, M.; Salehi, B.; Soltani-Nejad, A.; Rajabi, S.; Tajbakhsh, M. Carvacrol and human health: A comprehensive review. Phytotherapy Research 2018, 32, 1675–1687. [Google Scholar]
- Han, F.; Ma, G.-q.; Yang, M.; Yan, L.; Xiong, W.; Shu, J.-c.; Zhao, Z.-d.; Xu, H.-l. Chemical composition and antioxidant activities of essential oils from different parts of the oregano. Journal of Zhejiang University. Science. B 2017, 18, 79. [Google Scholar] [CrossRef]
- Miara, M.D.; Bendif, H.; Ouabed, A.; Rebbas, K.; Hammou, M.A.; Amirat, M.; Greene, A.; Teixidor-Toneu, I. Ethnoveterinary remedies used in the Algerian steppe: Exploring the relationship with traditional human herbal medicine. Journal of ethnopharmacology 2019, 244, 112164. [Google Scholar] [CrossRef]
- Luna, A.; Labaque, M.; Zygadlo, J.; Marin, R. Effects of thymol and carvacrol feed supplementation on lipid oxidation in broiler meat. Poultry Science 2010, 89, 366–370. [Google Scholar] [CrossRef]
- Soltan, Y.A.; Morsy, A.S.; Araujo, R.C.; Elziat, H.; Sallam, S.M.A.; Louvandini, H.; Abdalla, A.L. Carvacrol and eugenol as modifiers of rumen microbial fermentation, and methane production in vitro. In Proceedings of the Proc. Of 4th Animal Wealth Research Conference in the middle East and North Africa pp; 2011; pp. 354–364. [Google Scholar]
- Hashemipour, H.; Kermanshahi, H.; Golian, A.; Veldkamp, T. Effect of thymol and carvacrol feed supplementation on performance, antioxidant enzyme activities, fatty acid composition, digestive enzyme activities, and immune response in broiler chickens. Poultry science 2013, 92, 2059–2069. [Google Scholar] [CrossRef]
- Bravo, D.; Pirgozliev, V.; Rose, S. A mixture of carvacrol, cinnamaldehyde, and capsicum oleoresin improves energy utilization and growth performance of broiler chickens fed maize-based diet. Journal of Animal Science 2014, 92, 1531–1536. [Google Scholar] [CrossRef] [PubMed]
- Can Baser, K. Biological and pharmacological activities of carvacrol and carvacrol bearing essential oils. Current pharmaceutical design 2008, 14, 3106–3119. [Google Scholar] [CrossRef]
- Suntres, Z.E.; Coccimiglio, J.; Alipour, M. The bioactivity and toxicological actions of carvacrol. Critical reviews in food science and nutrition 2015, 55, 304–318. [Google Scholar] [CrossRef] [PubMed]
- Rathod, N.B.; Kulawik, P.; Ozogul, F.; Regenstein, J.M.; Ozogul, Y. Biological activity of plant-based carvacrol and thymol and their impact on human health and food quality. Trends in Food Science & Technology 2021, 116, 733–748. [Google Scholar]
- Ultee, A.; Kets, E.; Smid, E. Mechanisms of action of carvacrol on the food-borne pathogen Bacillus cereus. Applied and environmental microbiology 1999, 65, 4606–4610. [Google Scholar] [CrossRef]
- Bakır, M.; Geyikoglu, F.; Colak, S.; Turkez, H.; Bakır, T.O.; Hosseinigouzdagani, M. The carvacrol ameliorates acute pancreatitis-induced liver injury via antioxidant response. Cytotechnology 2016, 68, 1131–1146. [Google Scholar] [CrossRef]
- Imran, M.; Aslam, M.; Alsagaby, S.A.; Saeed, F.; Ahmad, I.; Afzaal, M.; Arshad, M.U.; Abdelgawad, M.A.; El-Ghorab, A.H.; Khames, A. Therapeutic application of carvacrol: A comprehensive review. Food Science & Nutrition 2022, 10, 3544–3561. [Google Scholar]
- Tahmasebi, A.; Karami, A.; Hosseini, S.M.; Afsharifar, A.; Moghadam, A.; Biniaz, Y. Cytotoxic and antimicrobial activities of Rydingia michauxii methanolic extracts during various growth stages. Cleaner Engineering and Technology 2021, 4, 100225. [Google Scholar] [CrossRef]
- Tahmasebi, A.; Karami, A.; Hosseini, S.M. Evaluation of antifungal activity of Rydingia michauxii extracts at the three different growth stages. In Proceedings of the 21th National & 9th International Congress on Biology, Semnan, Iran; 2021. [Google Scholar]
- Bhore, S.J.; Ravichantar, N.; Loh, C.Y. Screening of endophytic bacteria isolated from leaves of Sambung Nyawa [Gynura procumbens (Lour.) Merr.] for cytokinin-like compounds. Bioinformation 2010, 5, 191. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: principles, mechanisms of action, and future prospects. Applied and environmental microbiology 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Wirth, S.; Behrendt, U.; Ahmad, P.; Berg, G. Antimicrobial activity of medicinal plants correlates with the proportion of antagonistic endophytes. Frontiers in microbiology 2017, 8, 199. [Google Scholar] [CrossRef]
- Esmaeili, S.; Naghibi, F.; Mosaddegh, M. Indigenous knowledge of traditionally used plants from Iran for fever/malaria treatment. Planta Medica 2010, 76, P509. [Google Scholar] [CrossRef]
- Irani, M.; Esmaeili, S.; Pirani, A.; Naghibi, F.; Mosaddegh, M. Evaluation of antiplasmodial activity of some plants ethnopharmacologically used to treat malaria in Iran. 2015.
- Firuzi, O.; Javidnia, K.; Gholami, M.; Soltani, M.; Miri, R. Antioxidant activity and total phenolic content of 24 Lamiaceae species growing in Iran. Natural product communications 2010, 5, 1934578X1000500219. [Google Scholar] [CrossRef]
- Dehghani Nazhvani, A.; Razmkhah, M.; Jassbi, A.; Khademalizadeh, M.; Mahmoodi, A. Evaluation of anticancer effect of Hibiscus Sabdarifa, Otostegia Persica Otostegia Aucheri, and Otostegia Michauxii on oral squamous cell carcinoma cell line. Journal of Dental Medicine 2020, 32, 208–215. [Google Scholar]
- Esmaeili, S.; Hamzeloo-Moghadam, M.; Ghaffari, S.; Mosaddegh, M. Cytotoxic activity screening of some medicinal plants from south of Iran. Research Journal of Pharmacognosy 2014, 1, 19–25. [Google Scholar]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside - A review of its occurrence, (bio)synthesis and pharmacological significance. Biotechnology Advances 2014, 32, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.G.; Wang, P.; Yang, Z.Q.; Li, M.X.; Tian, X.Y. Pharmacological effects of verbascoside: research advances. Journal of International Pharmaceutical Research 2020, 47, 1078–1087. [Google Scholar] [CrossRef]
- He, W.; Fang, T.; Tu, P. Research progress on pharmacological activities of echinacoside. Zhongguo Zhongyao Zazhi 2009, 34, 476–479. [Google Scholar]
- Liu, J.; Yang, L.; Dong, Y.; Zhang, B.; Ma, X. Echinacoside, an Inestimable Natural Product in Treatment of Neurological and other Disorders. Molecules 2018, 23. [Google Scholar] [CrossRef]
- Singh, A.K.; Singla, R.K.; Pandey, A.K. Chlorogenic Acid: A Dietary Phenolic Acid with Promising Pharma-cotherapeutic Potential. Current Medicinal Chemistry 2023, 30, 3905–3926. [Google Scholar] [CrossRef]
- Sova, M.; Saso, L. Natural sources, pharmacokinetics, biological activities and health benefits of hydroxycinnamic acids and their metabolites. Nutrients 2020, 12, 1–30. [Google Scholar] [CrossRef]
- Çetinkaya, M.; Baran, Y. Therapeutic Potential of Luteolin on Cancer. Vaccines 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Punia Bangar, S.; Kajla, P.; Chaudhary, V.; Sharma, N.; Ozogul, F. Luteolin: A flavone with myriads of bioactivities and food applications. Food Bioscience 2023, 52. [Google Scholar] [CrossRef]
- Bhardawaj, H.; Vasudeva, N.; Sharma, S. Phytochemistry and Pharmacological Aspects of Apigenin: A Review. Natural Products Journal 2023, 13, 64–82. [Google Scholar] [CrossRef]
- Shahbaz, M.; Imran, M.; Alsagaby, S.A.; Naeem, H.; Al Abdulmonem, W.; Hussain, M.; Abdelgawad, M.A.; El-Ghorab, A.H.; Ghoneim, M.M.; El-Sherbiny, M.; et al. Anticancer, antioxidant, ameliorative and therapeutic properties of kaempferol. International Journal of Food Properties 2023, 26, 1140–1166. [Google Scholar] [CrossRef]
- Hwang, J.H.; Hwang, I.-s.; Liu, Q.-H.; Woo, E.-R.; Lee, D.G. (+)-Medioresinol leads to intracellular ROS accumulation and mitochondria-mediated apoptotic cell death in Candida albicans. Biochimie 2012, 94, 1784–1793. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.J.; Seo, D.-G.; Park, S.-Y. Phenylpropanoids from cinnamon bark reduced β-amyloid production by the inhibition of β-secretase in Chinese hamster ovarian cells stably expressing amyloid precursor protein. Nutrition Research 2016, 36, 1277–1284. [Google Scholar] [CrossRef]
- Badawi, M.M.; Handa, S.S.; Kinghorn, A.D.; Cordell, G.A.; Farnsworth, N.R. Plant anticancer agents XXVII: antileukemic and cytotoxic constituents of Dirca occidentalis (Thymelaeaceae). Journal of pharmaceutical sciences 1983, 72, 1285–1287. [Google Scholar] [CrossRef]
- Hwang, J.H.; Choi, H.; Hwang, I.-s.; Kim, A.R.; Woo, E.-R.; Lee, D.G. Synergistic antibacterial and antibiofilm effect between (+)-medioresinol and antibiotics in vitro. Applied biochemistry and biotechnology 2013, 170, 1934–1941. [Google Scholar] [CrossRef]
- Sauvain, M.; Kunesch, N.; Poisson, J.; Gantier, J.C.; Gayral, P.; Dedet, J.P. Isolation of leishmanicidal triterpenes and lignans from the Amazonian liana Doliocarpus dentatus (Dilleniaceae). Phytotherapy research 1996, 10, 1–4. [Google Scholar] [CrossRef]
- Wang, Y.; Guan, X.; Gao, C.-L.; Ruan, W.; Zhao, S.; Kai, G.; Li, F.; Pang, T. Medioresinol as a novel PGC-1α activator prevents pyroptosis of endothelial cells in ischemic stroke through PPARα-GOT1 axis. Pharmacological Research 2021, 169, 105640. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Taguchi, H.; ENDo, T.; Yosioka, I.; Higashiyama, K.; OTOMASU, H. The glycosides of Martynia louisiana Mill. A new phenylpropanoid glycoside, martynoside. Chemical and Pharmaceutical Bulletin 1978, 26, 2111–2121. [Google Scholar] [CrossRef]
- Pendota, S.; Aderogba, M.; Van Staden, J. In vitro antimicrobial activity of extracts and an isolated compound from Boscia albitrunca leaves. South African Journal of Botany 2015, 96, 91–93. [Google Scholar] [CrossRef]
- Tóth, E.; Tóth, G.; Máthé, I.; Blunden, G. Martynoside, forsythoside B, ladanein and 7a-acetoxyroyleanone from Ballota nigra L. Biochemical Systematics and Ecology 2007, 12, 894–897. [Google Scholar] [CrossRef]
- Jianjun, G.; Rongzhao, L.; Guiqiu, H.; Zhongjian, J. Studies on the topoisomerase inhibitors of phenylpropanoid glycosides from Pedicularis alaschanica Maxim. Beijing yi ke da xue xue bao= Journal of Beijing Medical University 1996, 28, 50–51. [Google Scholar]
- Miao, J.; Wang, W.; Yao, S.; Navaratnam, S.; Parsons, B. Antioxidative properties of Martynoside: pulse radiolysis and laser photolysis study. Free radical research 2003, 37, 829–833. [Google Scholar] [CrossRef]
- Liao, F.; Zheng, R.; Gao, J.; Jia, Z. Retardation of skeletal muscle fatigue by the two phenylpropanoid glycosides: verbascoside and martynoside from Pedicularis plicata Maxim. Phytotherapy Research: An International Journal Devoted to Pharmacological and Toxicological Evaluation of Natural Product Derivatives 1999, 13, 621–623. [Google Scholar] [CrossRef]
- Zhu, M.; Tan, N.; Zhu, H.; Zeng, G.; He, W.; Yu, B.; Chen, X. Anti-sports anaemia effects of verbascoside and martynoside in mice. International journal of sports medicine 2010, 31, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Guan, X.; He, Z.; Xie, Y.; Niu, Z.; Zhang, W.; Wang, A.; Zhang, J.; Si, C.; Li, F. Apigenin-7-O-glucoside alleviates DSS-induced colitis by improving intestinal barrier function and modulating gut microbiota. Journal of Functional Foods 2023, 104, 105499. [Google Scholar]
- Papoutsi, Z.; Kassi, E.; Mitakou, S.; Aligiannis, N.; Tsiapara, A.; Chrousos, G.P.; Moutsatsou, P. Acteoside and martynoside exhibit estrogenic/antiestrogenic properties. The Journal of Steroid Biochemistry and Molecular Biology 2006, 98, 63–71. [Google Scholar] [PubMed]
- Gowan, E.; Lewis, B.A.; Turgeon, R. Phloem transport of antirrhinoside, an iridoid glycoside, in Asarina scandens (Scrophulariaceae). Journal of chemical ecology 1995, 21, 1781–1788. [Google Scholar] [CrossRef]
- Beninger, C.W.; Cloutier, R.R.; Grodzinski, B. The iridoid glucoside, antirrhinoside, from Antirrhinum majus L. has differential effects on two generalist insect herbivores. Journal of Chemical Ecology 2008, 34, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Venditti, A.; Frezza, C.; Serafini, I.; Ciccòla, A.; Sciubba, F.; Serafini, M.; Bianco, A. Iridoids of chemotaxonomy relevance, a new antirrhinoside ester and other constituents from Kickxia spuria subsp. integrifolia (Brot.) R. Fern. Chemistry & Biodiversity 2018, 15, e1700473. [Google Scholar]
- Akkol, E.K.; Ercil, D. Antinociceptive and anti-inflammatory activities of some Linaria species from Turkey. Pharmaceutical Biology 2009, 47, 188–194. [Google Scholar] [CrossRef]
- Frezza, C.; Venditti, A.; Marcucci, E.; Parroni, A.; Reverberi, M.; Serafini, M.; Bianco, A. Phytochemical analysis of Linaria purpurea (L.) Mill. and inhibitory activity on the production of aflatoxin B1 (AFB1) in Aspergillus flavus Link. of one of its metabolites, antirrhinoside. Industrial crops and products 2019, 139, 111554. [Google Scholar] [CrossRef]
- Quirantes-Piné, R.; Herranz-López, M.; Funes, L.; Borrás-Linares, I.; Micol, V.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Phenylpropanoids and their metabolites are the major compounds responsible for blood-cell protection against oxidative stress after administration of Lippia citriodora in rats. Phytomedicine 2013, 20, 1112–1118. [Google Scholar] [CrossRef] [PubMed]
- Luhata, L.P.; Usuki, T. Free radical scavenging activities of verbascoside and isoverbascoside from the leaves of Odontonema strictum (Acanthaceae). Bioorganic & Medicinal Chemistry Letters 2022, 59, 128528. [Google Scholar]
- Chen, C.-Y.; Tung, H.-Y.; Tseng, Y.-F.; Huang, J.-S.; Shi, L.-S.; Ye, Y.-L. Verbascoside and isoverbascoside ameliorate transforming growth factor β1-induced collagen expression by lung fibroblasts through Smad/non-Smad signaling pathways. Life Sciences 2022, 308, 120950. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, B.; Barge, S.R.; Bharadwaj, S.; Deka, B.; Rahman, S.; Ghosh, A.; Manna, P.; Dutta, P.P.; Sheikh, Y.; Kandimalla, R. Evaluation of therapeutic effect of Premna herbacea in diabetic rat and isoverbascoside against insulin resistance in L6 muscle cells through bioenergetics and stimulation of JNK and AKT/mTOR signaling cascade. Phytomedicine 2021, 93, 153761. [Google Scholar] [CrossRef] [PubMed]
- Rui-Chuan, C.; Jin-Hua, S.; Gao-Liang, O.; Ke-Xia, C.; Jin-Quan, L.; Xiao-Guang, X. Induction of differentiation in human hepatocarcinoma cells by isoverbascoside. Planta medica 2002, 68, 370–372. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.-C.; Su, J.-H.; Yang, S.-M.; Li, J.; Wang, T.-J.; Zhou, H. Effect of isoverbascoside, a phenylpropanoid glycoside antioxidant, on proliferation and differentiation of human gastric cancer cell. Acta Pharmacol Sin 2002, 23, 997–1001. [Google Scholar]
- Wang, T.; Yang, S.; Huang, Z.; Li, Q.; Li, Q.; Zhou, H.; Zheng, R.; Jia, Z. Induced-differentiation and cytotoxicity of isoverbascoside on HL-60 cells. Shi yan Sheng wu xue bao 1999, 32, 321–327. [Google Scholar]
- Dimitrova, P.; Alipieva, K.; Stojanov, K.; Milanova, V.; Georgiev, M.I. Plant-derived verbascoside and isoverbascoside regulate Toll-like receptor 2 and 4-driven neutrophils priming and activation. Phytomedicine 2019, 55, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Z.; Huang, Y.; Wen, X.; Wu, Y.; Zhao, Y.; Ni, Y. Extraction, purification, and hydrolysis behavior of apigenin-7-O-Glucoside from Chrysanthemum morifolium tea. Molecules 2018, 23, 2933. [Google Scholar] [CrossRef] [PubMed]
- Minda, D.; Avram, S.; Pavel, I.Z.; Kis, B.; Ghitu, A.; Zupkó, I.; Dehelean, C.; Buda, V.; Diaconeasa, Z.; Scurtu, A. An in vitro evaluation of apigenin and apigenin-7-o-glucoside against hela human cervical cancer cell line. Revista de Chimie 2020, 71, 140–144. [Google Scholar] [CrossRef]
- Wang, W.; Yue, R.-F.; Jin, Z.; He, L.-M.; Shen, R.; Du, D.; Tang, Y.-Z. Efficiency comparison of apigenin-7-O-glucoside and trolox in antioxidative stress and anti-inflammatory properties. Journal of Pharmacy and Pharmacology 2020, 72, 1645–1656. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-M.; Ma, R.-H.; Ni, Z.-J.; Thakur, K.; Cespedes-Acuña, C.L.; Jiang, L.; Wei, Z.-J. Apigenin 7-O-glucoside promotes cell apoptosis through the PTEN/PI3K/AKT pathway and inhibits cell migration in cervical cancer HeLa cells. Food and Chemical Toxicology 2020, 146, 111843. [Google Scholar] [CrossRef]
- Smiljkovic, M.; Stanisavljevic, D.; Stojkovic, D.; Petrovic, I.; Vicentic, J.M.; Popovic, J.; Grdadolnik, S.G.; Markovic, D.; Sankovic-Babice, S.; Glamoclija, J. Apigenin-7-O-glucoside versus apigenin: Insight into the modes of anticandidal and cytotoxic actions. EXCLI journal 2017, 16, 795. [Google Scholar]
- Abugri, D.A.; Witola, W.H. Interaction of apigenin-7-O-glucoside with pyrimethamine against Toxoplasma gondii growth. Journal of Parasitic Diseases 2020, 44, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Gulluce, M.; Orhan, F.; Yanmis, D.; Arasoglu, T.; Guvenalp, Z.; Demirezer, L.O. Isolation of a flavonoid, apigenin 7-O-glucoside, from Mentha longifolia (L.) Hudson subspecies longifolia and its genotoxic potency. Toxicology and industrial health 2015, 31, 831–840. [Google Scholar] [CrossRef]
- Pei, Z.J.; Li, C.; Dai, W.; Lou, Z.; Sun, X.; Wang, H.; Khan, A.A.; Wan, C. The Anti-Biofilm Activity and Mechanism of Apigenin-7-O-Glucoside Against Staphylococcus aureus and Escherichia coli. Infection and Drug Resistance 2023, 16, 2129–2140. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of pharmacological properties of Rydingia michauxii.
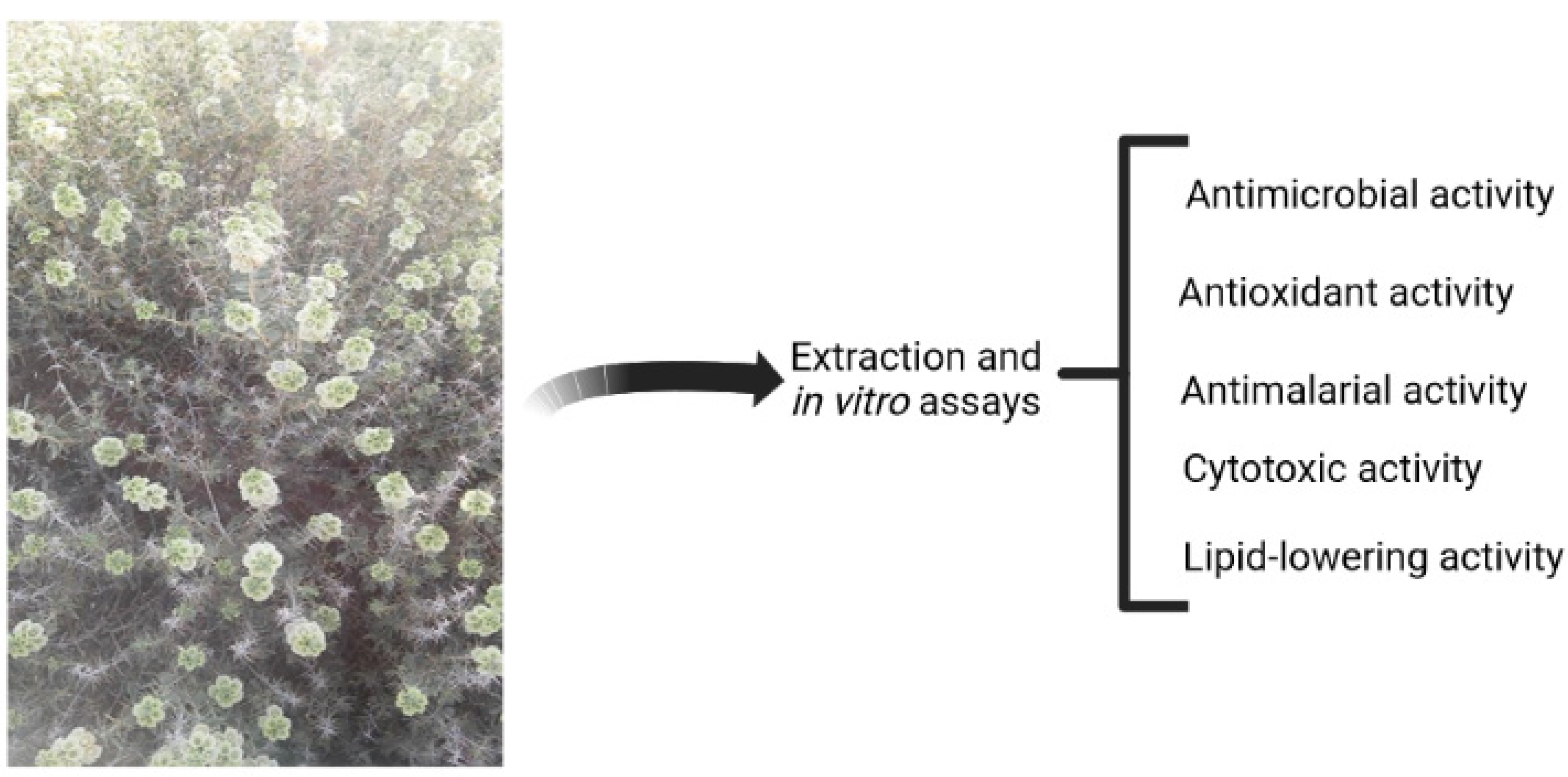
Figure 2.
Number of publications in each year including the keyword “natural and compounds” (generated using the Scopus online database).
Figure 2.
Number of publications in each year including the keyword “natural and compounds” (generated using the Scopus online database).
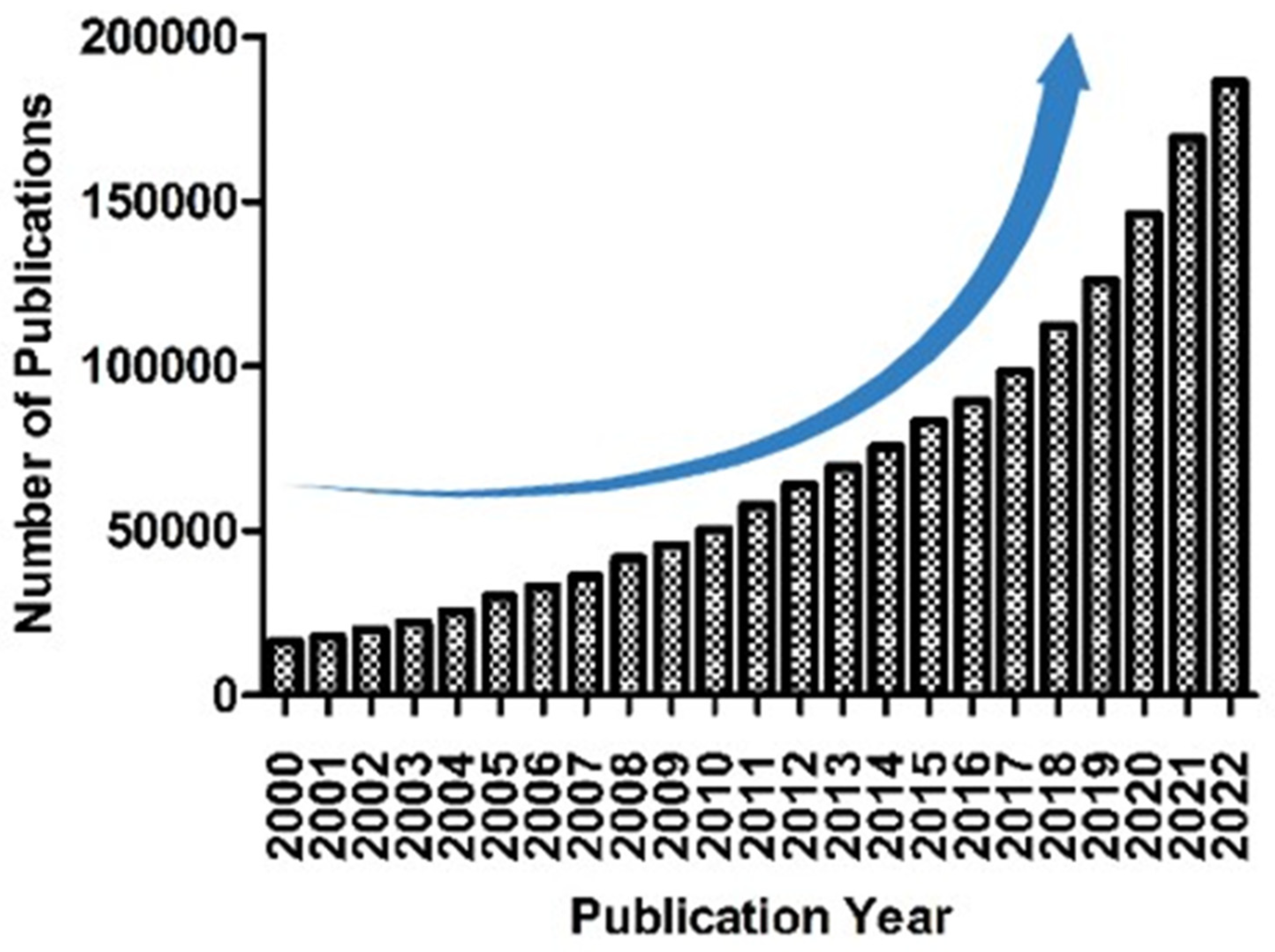
Figure 3.
The chemical structures of Dillapiole (1), Caryophyllene oxide (2), Eugenol (3). Eugenol acetate (4) and Carvacrol (5).
Figure 3.
The chemical structures of Dillapiole (1), Caryophyllene oxide (2), Eugenol (3). Eugenol acetate (4) and Carvacrol (5).
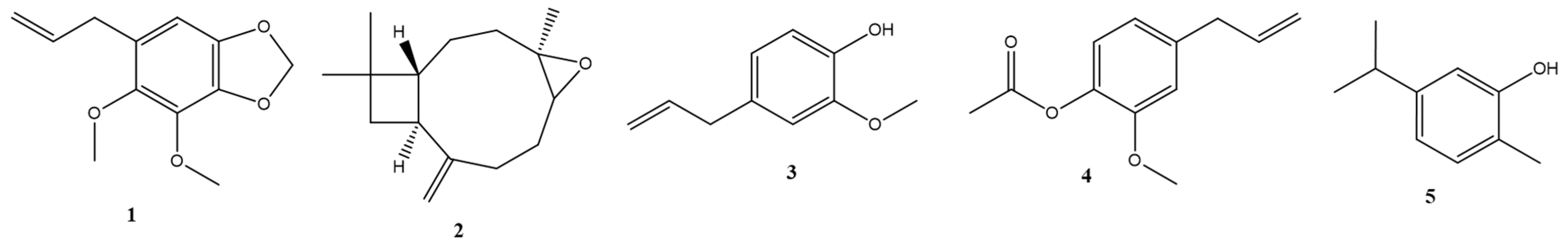
Figure 4.
Mechanism of action of statins in lipid lowering activity.
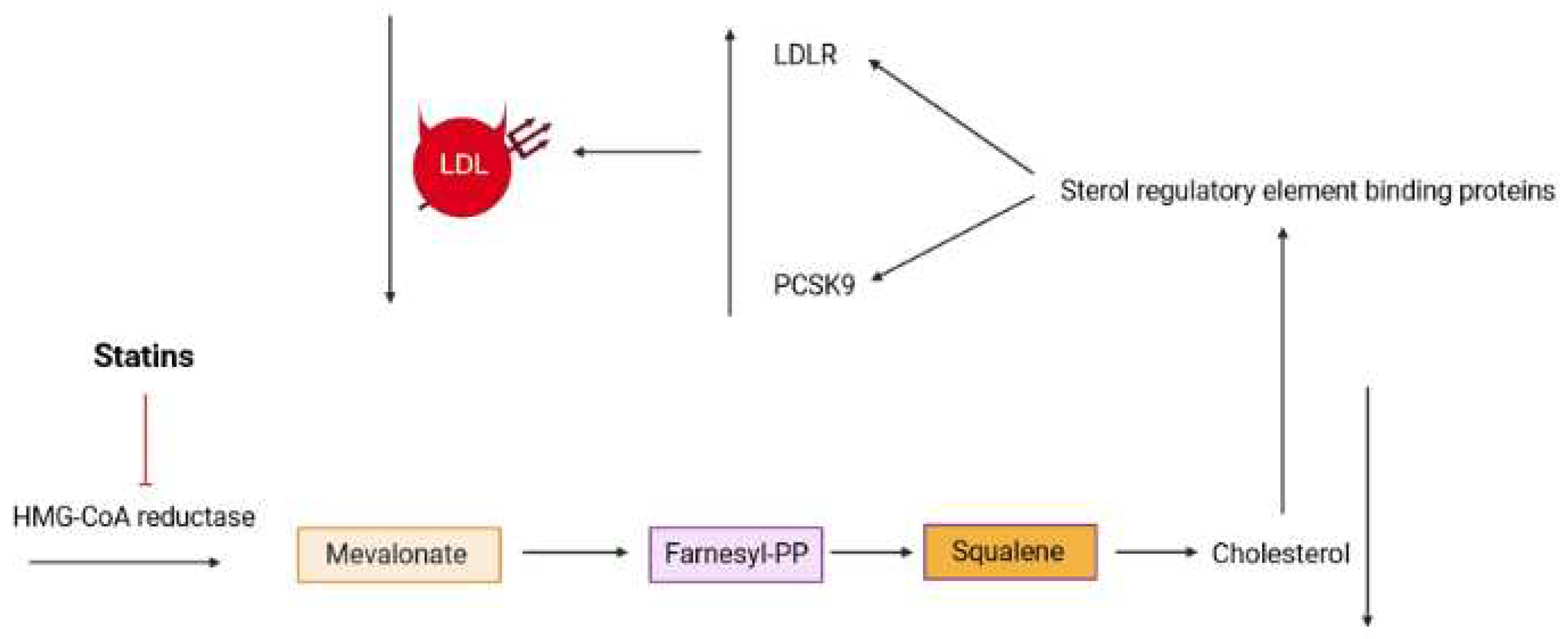
Figure 5.
The chemical structures of Medioresinol (1), Martynoside (2), Antirrhinoside (3), Isoverbascoside (4) and Apigenin-7-O-glucoside (5).
Figure 5.
The chemical structures of Medioresinol (1), Martynoside (2), Antirrhinoside (3), Isoverbascoside (4) and Apigenin-7-O-glucoside (5).
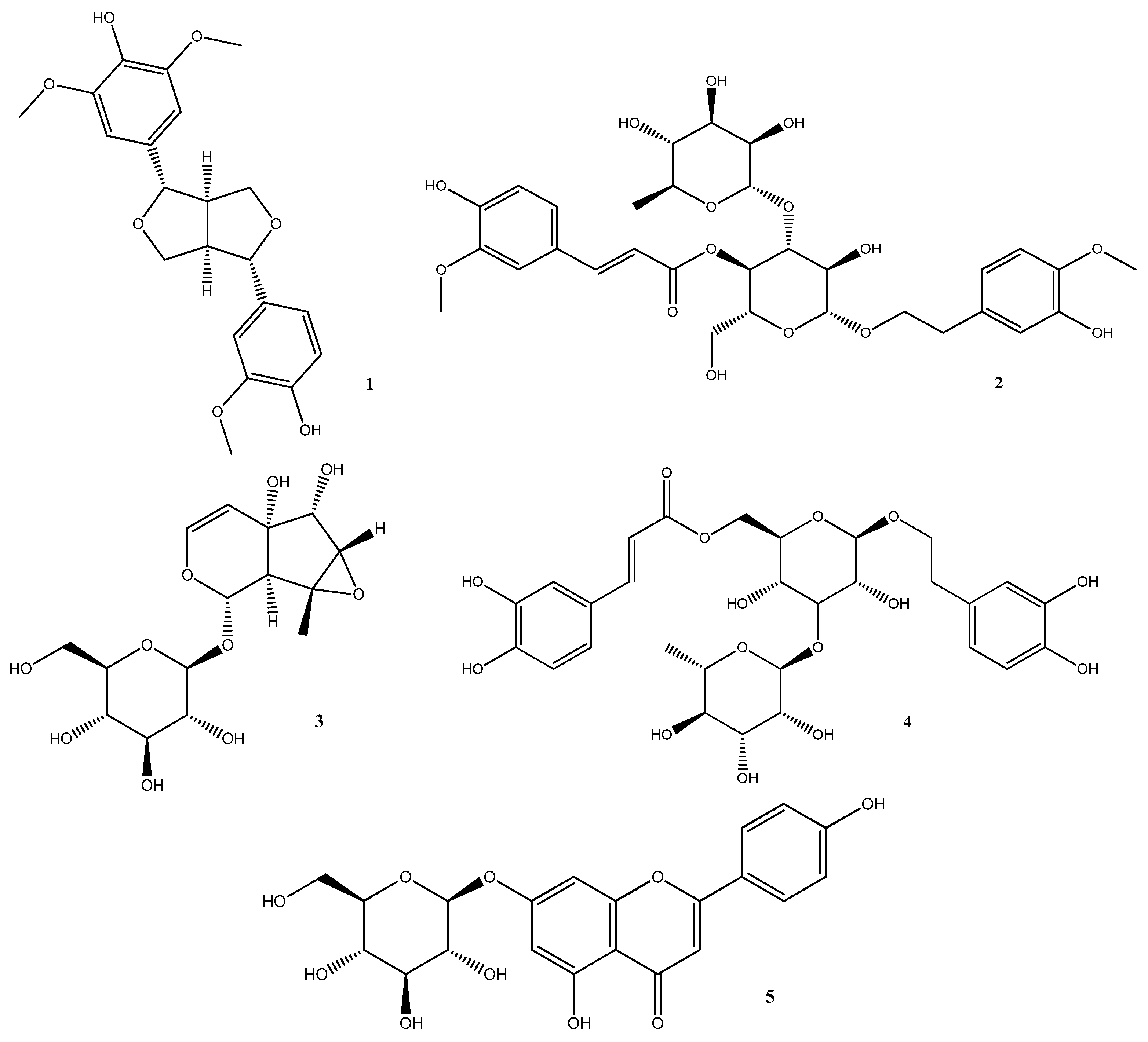

Table 1.
Chemical composition of the major constituents of the essential oils of Rydingia michauxii at different geographical locations and developmental stages.
Table 1.
Chemical composition of the major constituents of the essential oils of Rydingia michauxii at different geographical locations and developmental stages.
| Dominant constituents of the essential oils | Percentage | Developmental stage | Location | Reference |
|---|---|---|---|---|
| Dillapiole | 23.9 | Flowering | Chahzangooleh, Fars province, Iran | [8] |
| 2-methylbenzofuran | 12.9 | Flowering | Chahzangooleh, Fars province, Iran | [8] |
| α-pinene | 8.1 | Flowering | Chahzangooleh, Fars province, Iran | [8] |
| Caryophyllene oxide | 20.1 | Flowering | Kazeroon, Fars province, Iran | [9] |
| Trans-verbenol | 10.2 | Flowering | Kazeroon, Fars province, Iran | [9] |
| Linalool | 5.3 | Flowering | Kazeroon, Fars province, Iran | [9] |
| Humulene epoxide II | 4.6 | Flowering | Kazeroon, Fars province, Iran | [9] |
| Eugenol | 36.81 | Dormant | Kazeroon, Fars province, Iran | [20] |
| Eugenol acetate | 21.02 | Dormant | Kazeroon, Fars province, Iran | [20] |
| Carvacrol | 9.35 | Dormant | Kazeroon, Fars province, Iran | [20] |
| Carvacrol | 16.08 | Vegetative | Kazeroon, Fars province, Iran | [20] |
| Eugenol | 13.23 | Vegetative | Kazeroon, Fars province, Iran | [20] |
| Cumin aldehyde | 9.63 | Vegetative | Kazeroon, Fars province, Iran | [20] |
| Linalool | 8.28 | Vegetative | Kazeroon, Fars province, Iran | [20] |
| Carvacrol | 14.20 | Flowering | Kazeroon, Fars province, Iran | [20] |
| Eugenol | 8.98 | Flowering | Kazeroon, Fars province, Iran | [20] |
| δ-cadinene | 8.90 | Flowering | Kazeroon, Fars province, Iran | [20] |
| Caryophyllene oxide | 8.43 | Flowering | Kazeroon, Fars province, Iran | [20] |
| n-hexadecanoic acid | 7.07 | Flowering | Kazeroon, Fars province, Iran | [20] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



