Submitted:
10 July 2023
Posted:
11 July 2023
You are already at the latest version
Abstract
High salinity in soil affects the normal growth and development of crop. MicroRNA171 (miR171) plays a momentous role in plant resistance to adversity stress. The functions of Sly-miR171d and Sly-miR171e on growth of tomato and fruit nutritional quality under salt stress were studied. The results showed that the seed germination and seedling growth of tomato that silencing of Sly-miR171d/e by short tandem target mimic (STTM) were better than those of wild-type (WT). Silencing of Sly-miR171d/e seeds maintained a high germination rate, while WT tomato seeds barely germinated under 50 mM NaCl stress. Under 100 mM NaCl stress, silencing of Sly-miR171d/e increased the expression of their target gene SlGRAS24 with increasing duration of salt stress, while they enhanced the antioxidant activity, reduced reactive oxygen species (ROS) accumulation, and enhanced the salt tolerance of tomato plant by regulating gibberellin (GA) level. Moreover, the preharvest and postharvest fruit nutritional quality under salt stress was also studied, and the results showed that silencing of Sly-miR171d/e increased soluble solids, lycopene, total carotenoids, Vitamin C, organic acids and phenolic substances in tomato fruit. The present study demonstrated that silencing of Sly-miR171d/e enhanced plant salt tolerance through the GA pathway, and improved fruit nutritional quality under salt stress.
Keywords:
tomato
; Sly-miR171d/e
; short tandem target mimic (STTM)
; salt stress
; fruit quality
1. Introduction
High salinity is one of the major abiotic stresses that causes deleterious consequences in plant, such as inhibits seed germination and plant growth and decreases fruit productivity and nutrition [1]. Excessive soil salinity hinders the absorption of water by plant root and reduces turgor pressure due to water flowing out of vacuole, thereby resulting in an insufficient osmotic adjustment [2]. Tomato is an important vegetable crop that is rich in carotenoids, vitamins, organic acids, minerals and phenolic compounds [3]. Most tomato cultivars are salt-sensitive at all stages of life, whether in seed germination stage, seedling growth stage or fruit stage [4]. The salt stress is detrimental to the growth of tomato and fruit quality and yield.
The microRNAs (miRNAs) are 18 to 25 nucleotides long small RNAs and derived from non-coding RNA genes [5]. They play important roles in repressing gene expression at the post-transcriptional level through target complementary mRNAs degradation and translational inhibition in all eukaryotes [6]. Increasing amounts of research has revealed they are involved in various biological processes, inclusive of development, hormone response and stress response [7]. In cassava, the target genes MesNAC was regulated by miR164 and the MesARF8 was regulated by miR167 under drought stress, resulted in stomatal closure and reduced leaf expansion [8]. In tomato, the expression of GAMYB-like1 altered heat, cold and reactive oxygen species (ROS) signaling, while Sha-miR319d repressed its expression and by which regulating tomato cold tolerance [9]. Knockdown of Sly-miR164a increased the expression of NAC and elevated abscisic acid (ABA) levels during cold storage, resulted in improved cold tolerance of postharvest tomato fruit [10]. The miR171 family was one of the first miRNAs to be characterized and the most conserved in plant [11].
The miR171 family regulates GRAS genes (GRAS was derived from GIBBERELLIN-INSENSITIVE, REPRESSOR of ga1-3 and SCARECROW), which is essential for growth, development and stress response in plant [12]. Growing evidence demonstrates that GRAS genes play a pivotal role in the response to salt stress [13]. In cucumber, CsGRAS2 and CsGRAS26 are key genes involved in environmental stress, they were induced by gibberellin (GA/GA3) and ABA and were responsive to salt stress [14]. Silencing of HhGRAS14 in Hibiscus hamabo reduced the tolerance to salt stress, while overexpression of HhGRAS14 in Arabidopsis thaliana significantly increased salt tolerance and decreased the sensitivity to ABA [15]. In tomato, overexpression of SlGRAS40 enhanced plant tolerance to salt stresses, and influenced auxin (IAA) and GA signaling [13]. In addition, exogenous application of ABA, IAA and GA induced transcriptional accumulation of SlGRAS26, which was significantly inhibited under salinity stress [16].
Numerous studies have shown that GA has various associations with GRAS proteins in plant, phytohormones had been involved in the growth and development of shoot apical meristem (SAM) and root apical meristem (RAM) [17]. Important roles in SAM and RAM have been attributed to GA signaling, and the hairy meristem (HAM) gene has been shown to be relied on the maintenance of SAM and RAM [17,18]. In tomato, the HAM subfamily contains six SlGRAS genes, among which SlGRAS24 possesses a conserved MIR-binding sequence that perfectly matched Sly-miR171 and have been shown to be targeted by miR171 to cleave mRNA [19]. It has been documented that SlGRAS24 is a key transcription factor coordinating the regulation of GA signaling pathways in tomato, and the HAM genes may be regulators of endogenous GA balance in SAM, RAM, and axillary meristem to control the maintenance of meristem and generation of organs [20]. In previous studies, it was found that silencing of miR171d and miR171e enhanced the cold tolerance of tomato fruit by targeting SlGRAS24 to regulate GA signaling [21,22].
Abiotic stress is detrimental to the growth of tomato plant and also severely affects tomato fruit quality and yield [23]. Genetic improvement of tomato through molecular genetic approaches is critical for maintaining yield and improving tomato fruit quality [24]. In this study, the tomato plant and fruit of silencing of Sly-miR171d/e by short tandem target mimic (STTM) were successfully achieved, and the salt tolerance of transgenic tomato plant and fruit quality under salt stress was evaluated.
2. Results
2.1. Seed germination under salt stress
The salt tolerance of seed germination was tested on seed harvested from wild-type (WT), STTM-miR171d and STTM-miR171e plant (Figure 1). When the salt concentration was above 150 mM NaCl, all plant stopped growing (data not shown). In the absence of salt treatment, the growth of STTM plant showed no difference from that of WT plant (Figure 1A). However, the growth of WT plant was impaired after 50 mM NaCl treatment, although the transgenic lines also exhibited reduced root growth, but to a lesser extent. The root elongation of all seedlings was delayed after salt treatment, among which STTM-miR171e#40-1-3 root had the longest root length (Figure 1B).
Under normal growth conditions, the germination rate, root length and fresh weight of 14-day-old transgenic seedling and WT seedling were not significantly different (Figure 1C-E). However, under stress conditions, the germination rates of the genotypes decreased at 50 mM NaCl, and the germination rate of STTM-miR171d was higher than that of STTM-miR171e, which was 63.33% and 53.33%, respectively. Importantly, WT seed did not germinate after salt treatment. These results indicated that STTM-miR171d/e seed and seedling were better resistant to salinity than WT coercion.
2.2. Plant growth under salt stress
Compared with WT tomato plant, STTM-miR171d and STTM-miR171e tomato plants grew faster at seedling stage under salt stress (Figure 2A). On the 24th day of salt treatment, the transgenic plants had borne fruit, while WT tomato plants were still in the flowering stage, and the plant height and stem diameter of transgenic plants were higher than those of WT plants (Figure 2B-C). Especially STTM-miR171e#40-1-3 tomato plants grew well under salt stress.
2.3. Expression level of miR171d/e and SlGRAS24
After salt treatment, the relative expression levels of Sly-miR171d and Sly-miR171e in STTM-miR171d and STTM-miR171e tomato root were lower than those in WT root. After salt treatment for 24 hours, the relative expression of Sly-miR171d in STTM-miR171d#3-2-3, STTM-miR171d#11-5-3 and STTM-miR171d#15-2-2 were 0.16, 0.16 and 0.11 (Figure 2D), the relative expression of Sly-miR171e in STTM-miR171e#14-2-3, STTM-miR171e#26-2-3 and STTM-miR171e#40-1-3 were 0.05, 0.04 and 0.05 (Figure 2E), and the relative expression levels of Sly-miR171d and Sly-miR171e in WT root were 0.5 and 0.57, respectively.
The relative expression of SlGRAS24 in STTM-miR171d and STTM-miR171e were higher than that of WT root (Figure 2F). After 24 hours of salt treatment, the relative expression of SlGRAS24 in STTM-miR171d and STTM-miR171e was 87.26 and 89.45, which was 46.91 and 48.09 times higher than that of SlGRAS24 in WT root.
2.4. Histochemical detection and ROS accumulation
From the Figure 3A, H2O2 and O2•– accumulation in leaf of STTM-miR171d/e lines were lower than that of WT under salt stress, with brown (DAB staining) and blue (NBT staining) pigments were evident. Compared with WT root, the H2O2 contents in the root of STTM-miR171d/e lines after salt treatment were consistently lower than that of WT root.
After 14 days of salt treatment, the average H2O2 contents in the root of STTM-miR171d and STTM-miR171e were 2.301 and 2.058 μmol g−1, respectively, and the H2O2 content of WT tomato root was 2.992 μmol g−1 (Figure 3B). The O2•– content increased gradually in the root of STTM-miR171d/e lines after salt treatment, while the WT root accumulated more O2•–, the O2•– contents in the root of STTM-miR171d and STTM-miR171e were 8.792 and 7.978 μmol g−1, respectively, and the O2•– content of WT tomato root was 7.062 μmol g−1 (Figure 3C).
2.5. The content of proline, malondialdehyde (MDA), GA3 and total chlorophyll
From the Figure 3D, the proline content of all tomato root was higher than that of the control after salt treatment, the proline content of STTM-miR171d/e tomato root was higher than that of WT tomato root after 21 days of salt stress treatment. After salt treatment, the MDA content in WT tomato root was higher than that in STTM-miR171d and STTM-miR171e tomato root (Figure 3E). The average MDA content in WT tomato root increased steadily from 1.657 to 4.216 mmol kg−1 from the start of stress to the 21st day after stress. However, the MDA contents in the STTM-miR171d/e tomato were always kept at a low level, STTM-miR171d#3-2-3 and STTM-miR171e#26-2-3 were only 3.002 and 3.074 mmol kg−1 at the 21st day, which was 0.71 and 0.73 times lower than that of the WT tomato root.
Under salt stress, the GA3 content in tomato root of WT, STTM-miR171d and STTM-miR171e increased first and then decreased, and STTM-miR171d/e tomato root was always lower than WT root (Figure 3F). After 21 days of salt stress, the average contents of GA3 in root of STTM-miR171d and STTM-miR171e were 14.14 and 13.95 pmol L−1, respectively. The average content of GA3 in WT root was 16.10 pmol L−1, which was 0.88 and 0.87 times that of root of STTM-miR171d and STTM-miR171e.
Total chlorophyll content of the tomato leaf in WT and STTM-miR171d/e plant increased first and then decreased during stress treatments, but the levels in STTM-miR171d/e were much higher than in WT (Figure 3G).
2.6. Color evaluation
There is a large difference in the ripening of preharvest WT and STTM-miR171d/e tomato fruit under salt stress (Figure 4A). At 32 days post-anthesis (dpa), when the fruit of STTM-miR171d/e entered the discoloration stage, the WT tomato fruit was still in the immature stage. At 36 dpa, most fruit of STTM-miR171e had entered the red ripening stage, STTM-miR171d was in the yellow fruiting stage, and the WT tomato fruit was still in the color breaking stage. With the change of fruit ripening, the L* value gradually decreases, the a* value gradually increases, and the b* value increases first and then decreases (Figure 4B-D). Compared with the WT fruit, the a* value of the preharvest STTM-miR171d/e fruit is generally higher than that of the WT fruit during the color breaking process (Figure 4C).
It showed that the color changes during the postharvest ripening process of WT and transgenic fruit were observed under salt stress, compared with WT, the process of transgenic fruit color conversion was a little faster, especially for STTM-miR171e (Figure 5A). The L* value of transgenic and WT tomato fruit decreased gradually with the increase of fruit maturity, and the L* value of WT tomato fruits was always at a very low level (Figure 5B). During storage time of broken color (Br) to Br+7 day, the a* value of WT, STTM-miR171d and STTM-miR171e fruit increased from -4.82 to 29.44, 4.57 to 32.19 and 6.43 to 32.47 (Figure 5C). At Br+3 day, we found significant differences in the b* values between transgenic and WT tomato fruit. b* values were 47.02, 52.65, and 53.13 for WT, STTM-miR171d and STTM-miR171e fruit, respectively (Figure 5D).
2.7. Fruit quality evaluation
Under salt stress, in both of preharvest and postharvest fruit, the lycopene and total carotenoid contents in the peel of transgenic tomato were higher than those of WT tomato (Figure 6A-D). Among them, both the lycopene of STTM-miR171e#40-1-3 preharvest and postharvest fruit was at a higher level (Figure 6A,B). The total carotenoids content in the peel of postharvest fruit of STTM-miR171d#3-2-3 was the highest, which was 7.926 mg mL−1 (Figure 6D).
After salt treatment, the firmness of STTM-miR171d and STTM-miR171e preharvest tomato was lower than that of WT fruit (Figure 6E). From mature green (MG) to Br+7 day, the firmness of STTM-miR171d and STTM-miR171e postharvest fruit decreased from 9.91 to 6.43 N and 9.35 to 5.88 N, while the WT fruit decreased from 11.6 to 9.04 N (Figure 6F). The soluble solids content (SSC) of STTM preharvest and postharvest tomato fruit was higher than that of WT under salt stress (Figure 6G,H). At Br+7 day, the SSC of STTM-miR171d#11-5-3 was 1.06 times than that of WT, and STTM-miR171e#14-2-3 was 1.1 times than that of WT, which were 4.67% and 4.82% (Figure 6H).
After salt treatment, the Vitamin C content of preharvest tomato fruit of STTM-miR171d#3-2-3 was 1.16 times that of WT fruit, and STTM-miR171e#14-2-3 was 1.15 times that of WT fruit, which were 33.09 and 33.98 mg 100mg−1 (Figure 6I). During postharvest storage, the average Vitamin C content of WT, STTM-miR171d and STTM-miR171e fruit was 29.713, 33.045 and 32.727 mg 100mg−1 at Br+7 day, respectively (Figure 6J). The average contents of total phenolic of preharvest tomato fruit of STTM-miR171d and STTM-miR171e were 0.933 and 0.925 mg mL−1, respectively, while WT fruit was 0.73 mg mL−1 (Figure 6K). The total phenolic content of WT postharvest fruit was 0.421 mg mL−1 at MG stage, increased to 0.611 mg mL−1 at Br+7 day, and the total phenolic contents of STTM-miR171d#15-2-2 and STTM-miR171e#40-1-3 were 0.707 and 0.756 mg mL−1 at Br+7 day, respectively (Figure 6L).
The contents of six organic acids in preharvest and postharvest transgenic tomato and WT tomato fruit under salt stress were detected. In general, the organic acids content of the tomato fruit after salt treatment was increased compared with that in the control condition. The average citric and malic acid contents of STTM-miR171d preharvest red-ripe (RR) tomato after salt treatment were 0.1994 and 0.0935 mg g−1, respectively, which were 0.0875 and 0.0554 mg g−1 higher than those of STTM-miR171d fruit under control conditions. From Br+4 to Br+7 day, the average citric acid content of salt treated STTM-miR171e tomato increased from 0.1367 to 0.2452 mg g−1 (Table 1).
3. Discussion
Salinity adversely affects nearly every aspect of plant growth and development, including seed germination, seedling growth, plant growth, and fruit yield [25]. Tomato plant is particularly sensitive to salt throughout the seedling growth period [26]. To assess the role of Sly-miR171d and Sly-miR171e in salt tolerance, STTM-miR171d, STTM-miR171e, and WT tomato seeds were placed in 50 mM NaCl ½ × MS solid medium to observe germination, and STTM tomato seeds were found to germinate at a higher rate than WT tomato seeds under salt stress (Figure 1). Tomato seedlings were watered with 100 mM NaCl solution every 48 hours as salt stress treatment for up to 28 days. All STTM plants grew better than WT plants under salt treatment (Figure 2A). These results suggest that silencing of Sly-miR171d/e can lead to greater salt tolerance and better germination performance of tomato. Under salt stress, when STTM plants emerged with fruits, WT plants had just flowered, and when STTM tomato fruit was completely red ripe, WT fruit was still in the color-changing stage (Figure 4A), and the rate of red ripening of postharvest STTM fruit was faster than that of WT fruit (Figure 5A). These results may be related to the regulation of fruit ripening and development by Sly-miR171d/e. Silencing of Sly-miR171d/e accelerates plant growth and development, promotes fruit ripening, and makes STTM fruit break color earlier and accumulates more pigment than WT fruit.
The typical major signal caused by salt stress was the ionic or ionic-toxicity effects on plant cell, and often caused complex secondary effects including oxidative stress and damage to cell components [27]. Plant undergoes oxidative stress under abiotic stress, resulting in the accumulation of large amounts of ROS [13]. There is increasing evidence that ROS-scavenging systems against H2O2 accumulation and toxicity are associated with plant salt tolerance [28]. Excessive salt accumulation not only affects water uptake by seeds but also causes severe damage to seed tissues [29]. Under salt stress, overexpression of SlCOMT1 induced excessive accumulation of endogenous melatonin in tomato seeds, provided ABA and GA metabolism to attenuate the inhibitory effects on seed germination, and activated tolerance-related signals to improve osmoregulation and ROS scavenging to prevent stress-induced damage so that tomato seeds acquired higher salt tolerance [30]. The present study deliberated accumulation of H2O2 and O2•− in WT, STTM-miR171d and STTM-miR171e plant under salt stress. Results showed that salt treatment induced WT plant to produce more H2O2 and O2•−, while the accumulation was significantly reduced in STTM plant (Figure 3A-C). MDA is a product of lipid peroxidation by ROS and is commonly used to assess ROS-mediated plant damage [31]. The MDA and proline contents in salt-treated plant were measured (Figure 3D,E), as their accumulation was characteristic of physiological stress. The MDA content of WT plant was significantly increased under salt stress, and the MDA content of STTM plant was slightly increased, but not as much as WT plant. Drought and salt stress often increase proline levels in plant for osmotic regulation [2]. Both WT and STTM plant increased proline content after salt stress, but STTM plant accumulated more proline under salt stress. The possible reason for these results is that silencing of Sly-miR171d/e reduced the accumulation of ROS and improved the salt tolerance of tomato.
Plant contains substantial genes with numerous biological functions and plenty genes with unknown roles that are activated and integrated through complex and diverse transcriptional mechanisms in response to abiotic stress [32]. Phytohormones regulate plant responses to salt stress [33]. GA is known to play an important role in regulating plant responses to the environment and controlling stress induced genes [34]. GA facilitates the growth of tomato plant under salt stress [35]. GA accumulation induces seed germination, and after germination reduction of GA levels or GA signaling to improve salt tolerance [36]. The DELLA proteins are derived from the GRAS family and play a central role in GA signaling. GA promotes plant growth by triggering DELLA degradation through the ubiquitin-proteasome pathway [37]. DELLAs are participated in ROS responses and developmental coordination when subjected to abiotic stress [38]. Salt stress can enhance the accumulation of DELLAs while upregulating genes encoding antioxidant systems and then reducing ROS abundance [39,40]. In cucumber, spermidine significantly increased the salt tolerance of cucumber under salt stress, enhanced the expression of GT-3b, and promoted the accumulation of GA3. When plants were treated with GA3 biosynthesis inhibitors, the salt tolerance induced by spermidine was blocked [41]. Plant of overexpressing SlGRAS24 had pleiotropic phenotypes associated with multiple agronomic traits, and GA-related genes were observed to be downregulated in seedling and altered in response to exogenous GA3 application [20]. In this study, the expression of SlGRAS24 in STTM-miR171d/e tomato root was significantly increased under salt stress (Figure 2F), and the GA3 level was lower than that of WT during the process (Figure 3F), so it was speculated that STTM-miR171d/e might be involved in salt resistance by regulating GA level.
Salt stress often has important adverse impacts on fruit yield and favorable impacts on fruit quality, which is a major problem facing modern agriculture, so that breeding salt-tolerant crops is essential for the development of sustainable agriculture [42]. Moderate soil salinity can improve the physiological indexes of fruit, such as the concentration of total soluble solids, Vitamin C, lycopene or some phenolic compounds, and which is very beneficial to improve organoleptic and antioxidant properties of the fruit [43]. Numerous studies have demonstrated that the positive effects of salinity can be used to improve the nutritional quality of fruit. In strawberry, lower levels of salt treatment favor photosynthesis, promote plant vegetative growth, and increase anthocyanin and sucrose content in fruit. Higher levels of salt treatment led to root growth and accumulation of more phenolic compounds, and the fruit quality of strawberry is improved by mild salt stress [44]. In cucumber, the soluble sugar, titratable acidity, and Vitamin C content of the fruit under NaCl stress were increased by grafting, so that the fruit quality was comprehensively improved [45]. Studies have shown that saltwater irrigation increased the content of sugar and organic acid and improved the taste of tomato [46]. Under salt stress, ions and organic molecules produced by plants will increase the content of soluble solids in fruit due to concentration effect and active accumulation of solutes [47], the increase in fruit acidity is related to organic acids content [48]. Saline irrigation and application of different nitrogen fertilizers significantly improved fruit quality in tomato [49. De Pascale et al. [49] investigated the combined effects of increasing NaCl concentration to irrigate and using fertilizers with divergent nitrogen sources on the chemical components of tomato fruit, showed that the carotenoid contents and antioxidant activity were increased while reduced acceptable yields by irrigating with brine containing 0.25% (w/v) NaCl. Concentration effect caused by reduced water content in plant under salt stress led to increased carotenoid content in salinized tomato plant [50]. SlGRAS24 has been proved to play a key role in fruit ripening, and GRAS gene is involved in mediating GA response during flower fruit transformation [19]. Sly-miR171d/e may promote fruit ripening by regulating SlGRAS24. STTM miR171d/e enters the mature stage faster than WT tomato fruit, and accumulates more nutrients. The present result is consistent with previous studies that STTM-miR171d/e preharvest fruit accumulated more nutrients than that of WT fruit after salt treatment, and the content of soluble solids, lycopene, total carotenoids, Vitamin C, phenolics and organic acids were significantly increased, which improved the quality of tomato fruit after salt treatment (Figure 6). This may be related to the fact that silencing of Sly-miR171d/e improved fruit salt tolerance.
The effects of salt stress can directly alter the quality of the fruit, especially the content of bioactive substances in the postharvest fruit, of which antioxidants are important for human health [51]. Antioxidant activity is a momentous indicator for estimating the nutritional value of fruit and vegetable [52]. Lycopene is the most abundant carotenoid in tomato fruits and it is responsible for fruit coloration during ripening and is also one of the main antioxidants [53]. During postharvest storage, STTM-miR171d/e transgenic tomato underwent a faster color change than WT tomatoes and accumulated more lycopene and total carotenoids than WT tomato fruits (Figure 6B,D). Total phenols in plants are also a natural class of antioxidants that can function as cellular signals under abiotic stress conditions through a series of secondary metabolites synthesized by the shikimic acid or malonic acid pathways [51]. These phenolic compounds improve the antioxidant capacity of fruits by reducing free radical levels and decreasing oxidative changes in cells [53]. Vitamin C is one of the most important bioactive compounds and potent antioxidants in tomato [54]. Under salt stress, the Vitamin C content of STTM-miR171d/e transgenic tomato was significantly increased compared to WT tomato fruits (Figure 6J), indicating that salt stress significantly reduced the Vitamin C content of postharvest tomato fruits and that STTM silencing miR171d/e may be involved in inducing Vitamin C production, which is related to the antioxidant response of the plant [55]. The antioxidant activity is supposed to result from the action of all the antioxidants in the fruit [56]. These bioactive substances are very important in the stress tolerance of plant, and their accumulation is a good response of plant [51].
4. Materials and Methods
4.1. Transformation of tomato plant
Silencing of Sly-miR171d or Sly-miR171e with STTM was performed according to Yan et al. [63]. Primary transgenic plant (T0) was generated by Agrobacterium tumefaciens-mediated transformation according to Zhao et al. [22]. Three STTM-miR171d and STTM-miR171e lines were selected from the T1 and T2. The PCR amplification in the presence of Hygromycin (STTM-miR171d/e lines) assay and agarose gel electrophoresis analysis proved that the three independent transgenic lines were all inserted with small target genes (Supplementary 1). Subsequently, three independent T3 generation transgenic lines of STTM-miR171d and STTM-miR171e were planted.
4.2. Plant materials and growth conditions
Tomato (Solanum lycopersicum cv. Micro-Tom) were grown on soil in a growth chamber with a 16 hours light (25 oC)/8 hours dark (18 oC) cycles. The seed germination was according to the modified methodology [13]. After disinfection, seeds of WT, STTM-miR171d and STTM-miR171e were placed on ½ × MS solid medium containing 50 mM NaCl and cultured in a cycle of 16 hours of light (25 oC)/ 8 hours of darkness (18 oC). Seed germination rate was counted after 7 days. Primary root length and fresh weight were determined after 14 days. At least 30 seeds were treated in each treatment, and all treatments were performed 3 times independently.
The stress treatment of seedings was based on the modified methodology [13]. The 28-day-old WT, STTM-miR171d and STTM-miR171e plant grown under the same conditions were selected and were watered with 100 mM NaCl every 3 days, and the normal water was used as the control. Plant height and stem diameter were recorded every 4 days, with representative seedling photographed. After 28 days of salt treatment, the root and leaf at the same stage of development were harvested and sampled independently for 3 times for each strain.
After growing under the same conditions until tomatoes reached flowering, healthy, uniformly growing WT and STTM-miR171d/e tomato plants were selected and watered with 100 mM NaCl aqueous solution every 4 days, and treated with water as a control. Flowering time and fruit color break were counted. Healthy and simultaneously flowering MG tomatoes were selected to record tomato color change phenotypes at 28, 32, 36 and 40 dpa, and healthy RR tomatoes were picked at 50 dpa, and fresh fruit were rapidly frozen. In addition, healthy MG tomato fruits with the same flowering time were selected for storage. The harvested tomato fruits were sterilized by immersion in 2% NaClO solution for a few seconds, removed and dried, and stored at room temperature and protected from light. Representative tomato fruits of Br day were selected for monitoring the color transfer process, and phenotypes were recorded by daily photographs. The fruit of MG, Br+4 d and Br+7 d was also sampled, frozen in liquid nitrogen and stored at -80 oC.
4.3. RNA extraction and quantitative real-time PCR analysis
The expressions levels of SlGRAS24, Sly-miR171d and Sly-miR171e were determined by an improved method [58]. Root samples were collected from 1 month old WT, STTM-miR171d and STTM-miR171e plant with at least 6 plants each line. Samples from different plants were mixed and immediately frozen in liquid nitrogen. Sampling was performed at 3 times independently. Total RNA of root samples on 0, 3, 6, 12 and 24 hours in salt treated tomato was extracted by RNA prep pure Plant Kit (Tiangen Biotechnology Co., Ltd., Ltd. Nanjing, China). Calculated the relative expression of SlGRAS24, Sly-miR171d and Sly-miR171e used equation 2-ΔΔCT. Supplementary 2 contained all the primers used in Real-time PCR.
4.4. ROS accumulation
Transgenic and WT tomato seedling grown to 28th day were subjected to salt stress for 14 days, and the 5th or 6th true leaf of the seedling was picked for histochemical staining. Histochemical detection of hydrogen peroxide (H2O2) and superoxide anion radical (O2•−) was performed according to Hu et al. [59]. Leaf samples were subjected to the 3,3ʹ-diaminobenzidine (DAB) and nitro-blue tetrazolium (NBT) stained and photographed. Used liquid nitrogen to thoroughly grind the tomato root samples to determine the H2O2 and O2•− by analysis kit (Solarbio Inc. Beijing, China). The H2O2 content and O2•− content were determined at 530 and 415 nm, respectively, and were expressed as μmol g−1.
4.5. Determination of proline, MDA, GA3 and chlorophyll
Following salt treatment, root and leaf (The 5th or 6th leaf from true leaf) at similar developmental stages from transgenic and WT plant were collected at predetermined times (During 28 days after salt treatment). The sample was fully ground into powder with liquid nitrogen. Tomato root samples were thoroughly ground with liquid nitrogen. Proline content and MDA content were measured by Duan et al. [60] improved method. Units of mmol kg−1 were used to indicate MDA content in fresh weight. The GA3 content was determined by assay kit (Meibiao Biotechnology Co., Ltd. Jiangsu, China). The leaf samples are used for chlorophyll determination using a method of Zhu et al. [61].
4.6. Fruit color measurement
Fruit was monitored from Br day until the 8th day (Br+7 day), with representative fruit photographed. Fruit color was determined using a Minolta Chroma Meter CR-400 (Minolta Camera Co. Ltd., Osaka, Japan). Three tomato per strain were randomly selected for color measurements, with two readings on two opposite sides of the equatorial region. The results were denoted by L*, a* and b*.
4.7. Fruit quality evaluation
Fruit of MG and RR was harvested. The lycopene assay was performed according to the method of Fish et al. [62]. Total carotenoids content was extracted using a modified protocol from Zhu et al. [61]. The firmness of each tomato fruit (three readings per treatment) was determined using a texture analyser (TA. XT Plus, Stable Micro Systems, Godalming, UK) at a rate of 50 mm s−1 with an insertion depth of 5 mm. Firmness was expressed in newtons (N). The SSC of samples was assayed using a digital refractometer (PAL−1, Atago, Japan).
The Vitamin C content was determined following the protocol proposed by Wan et al. [63]. The concentration of Vitamin C in the sample was calculated from a calibration line prepared with a standard solution of Vitamin C. The total phenolic content was determined following the protocol proposed by Wang et al. [64]. Calculation of standard curve made from gallic acid solution. Unit of g kg−1 was used to indicate total phenolic content in fresh weight. The organic acid content was detected by high performance liquid chromatography (HPLC, Agilent 1260, CA, United States).
4.8. Fruit quality evaluation
Data were analyzed by Two-way analysis of variance (ANOVA) using GraphPad Prism 9 (GraphPad, San Diego, CA, United States). When P < 0.05 (*) and P < 0.01 (**), there was a significant difference between the two values. The data of organic acid was subjected to one-way ANOVA using IBM SPSS Statistics 26 (SPSS in Chicago, IL, United States). Tukey test showed statistically significant difference (P < 0.05). All values were expressed as the average value ± standard deviation (SD).
5. Conclusions
The present study demonstrated that silencing of Sly-miR171d or Sly-miR171e increased the expression of target gene SlGRAS24 in tomato plant, reduced ROS accumulation, enhanced tomato salt tolerance through the GA pathway and improved fruit nutritional quality.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Supplementary 1: Positive verification of STTM-miR171d and STTM-miR171e; Supplementary 2: All Primers used in Real-time-PCR; Supplementary 3: Sequence of STTM-miR171d and STTM-miR171e; Supplementary 4: Sequence of pri-miR171d and pri-miR171e.
Author Contributions
Conceptualization, T.H. and L.L.; methodology, T.H. and X.X.; software, Z.X., S.W. and W.D.; validation, T.H., Z.X. and S.W.; formal analysis, Q.W. and H.S.; investigation, W.D. and L.L.; resources, X.X.; data curation, Q.W. and L.M.; writing—original draft preparation, T.H.; writing—review and editing, T.H. and X.X.; visualization, X.X.; supervision, Q.W. and L.M.; project administration, H.S. and X.X.; funding acquisition, H.S. and X.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Hainan Provincial Natural Science Foundation of China (321RC1025, 322RC565 and 323RC404).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Meng, X.; Cai, J.; Deng, L.; Li, G.; Sun, J.; Han, Y.; Dong, T.; Liu, Y.; Xu, T.; Liu, S.; Li, Z.; Zhu, M. SlSTE1 promotes abscisic acid-dependent salt stress-responsive pathways via improving ion homeostasis and reactive oxygen species scavenging in tomato. J. Integr. Plant Biol. 2020, 62, 1942–1966. [Google Scholar] [CrossRef] [PubMed]
- Devkar, V.; Thirumalaikumar, V.P.; Xue, G.P.; Vallarino, J.G.; Tureckova, V.; Strnad, M.; Fernie, A.R.; Hoefgen, R.; Mueller-Roeber, B.; Balazadeh, S. Multifaceted regulatory function of tomato SlTAF1 in the response to salinity stress. New Phytol. 2020, 225, 1681–1698. [Google Scholar] [CrossRef]
- Schwarz, D.; Thompson, A.J.; Klaering, H.P. Guidelines to use tomato in experiments with a controlled environment. Front Plant Sci. 2014, 5, 625. [Google Scholar] [CrossRef] [PubMed]
- Foolad, M.R. Recent advances in genetics of salt tolerance in tomato. Plant Cell Tiss. Org. Cult. 2004, 76, 101–119. [Google Scholar] [CrossRef]
- Cherian, S.; Figueroa, C.R.; Nair, H. 'Movers and shakers' in the regulation of fruit ripening: a cross-dissection of climacteric versus non-climacteric fruit. J. Exp. Bot. 2014, 65, 4705–4722. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef]
- Xu, X.; Ma, X.; Lei, H.; Yin, L.; Shi, X.; Song, H. MicroRNAs play an important role in the regulation of strawberry fruit senescence in low temperature. Postharvest Biol. Technol. 2015, 108, 39–47. [Google Scholar] [CrossRef]
- Phookaew, P.; Netrphan, S.; Sojikul, P.; Narangajavana, J. Involvement of miR164- and miR167-mediated target gene expressions in responses to water deficit in cassava. Biol. Plantarum. 2014, 58, 469–478. [Google Scholar] [CrossRef]
- Shi, X.; Jiang, F.; Wen, J.; Wu, Z. Overexpression of Solanum habrochaites microRNA319d (sha-miR319d) confers chilling and heat stress tolerance in tomato (S. lycopersicum). BMC Plant Biol. 2019, 19, 214. [Google Scholar] [CrossRef]
- Zhao, K.; Song, H.; Wang, Z.; Xing, Z.; Tian, J.; Wang, Q.; Meng, L.; Xu, X. Knockdown of Sly-miR164a by short tandem target mimic (STTM) enhanced postharvest chilling tolerance of tomato fruit under low temperature storage. Postharvest Biol. Technol. 2022, 187, 111872. [Google Scholar] [CrossRef]
- Axtell, M.J.; Bowman, J.L. Evolution of plant microRNAs and their targets. Trends Plant Sci. 2008, 13, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Schulze, S.; Schaefer, B.N.; Parizotto, E.A.; Voinnet, O.; Theres, K. LOST MERISTEMS genes regulate cell differentiation of central zone descendants in Arabidopsis shoot meristems. Plant J. 2010, 64, 668–678. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, W.; Xian, Z.; Hu, N.; Lin, D.; Ren, H.; Chen, J.; Su, D.; Li, Z. Overexpression of SIGRAS40 in tomato enhances tolerance to abiotic stresses and influences auxin and gibberellin signaling. Front Plant Sci. 2017, 8, 1659. [Google Scholar] [CrossRef]
- Li, C.; Dong, S.; Liu, X.; Bo, K.; Miao, H.; Beckles, D.M.; Zhang, S.; Gu, X. Genome-Wide characterization of cucumber (Cucumis sativus L.) GRAS genes and their response to various abiotic stresses. Horticulturae 2020, 6, 110. [Google Scholar] [CrossRef]
- Ni, L.; Wang, Z.; Liu, X.; Wu, S.; Hua, J.; Liu, L.; Yin, Y.; Li, H.; Gu, C. Genome-wide study of the GRAS gene family in Hibiscus hamabo Sieb. et Zucc and analysis of HhGRAS14-induced drought and salt stress tolerance in Arabidopsis. Plant Sci. 2022, 319, 111260. [Google Scholar] [CrossRef]
- Zhou, S.; Hu, Z.; Li, F.; Yu, X.; Naeem, M.; Zhang, Y.; Chen, G. Manipulation of plant architecture and flowering time by down-regulation of the GRAS transcription factor SlGRAS26 in Solanum lycopersicum. Plant Sci. 2018, 271, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Engstrom, E.M.; Andersen, C.M.; Gumulak-Smith, J.; Hu, J.; Orlova, E.; Sozzani, R.; Bowman, J.L. Arabidopsis homologs of the petunia HAIRY MERISTEM gene are required for maintenance of shoot and root indeterminacy. Plant Physiol. 2011, 155, 735–750. [Google Scholar] [CrossRef]
- Weiss, D.; Ori, N. Mechanisms of cross talk between gibberellin and other hormones. Plant Physiol. 2007, 144, 1240–1246. [Google Scholar] [CrossRef]
- Huang, W.; Xian, Z.; Kang, X.; Tang, N.; Li, Z. Genome-wide identification, phylogeny and expression analysis of GRAS gene family in tomato. BMC Plant Biol. 2015, 15, 209. [Google Scholar] [CrossRef]
- Huang, W.; Peng, S.; Xian, Z.; Lin, D.; Hu, G.; Yang, L.; Ren, M.; Li, Z. Overexpression of a tomato miR171 target gene SlGRAS24 impacts multiple agronomical traits via regulating gibberellin and auxin homeostasis. Plant Biotechnol. J. 2017, 15, 472–488. [Google Scholar] [CrossRef]
- Xing, Z.T.; Huang, T.S.; Zhao, K.Y.; Meng, L.H.; Song, H.M.; Zhang, Z.K.; Xu, X.B.; Liu, S.B. Silencing of Sly-miR171d increased the expression of GRAS24 and enhanced postharvest chilling tolerance of tomato fruit. Front Plant Sci, 2022, 13, 1006940. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Chen, R.; Duan, W.; Meng, L.; Song, H.; Wang, Q.; Li, J.; Xu, X. Chilling injury of tomato fruit was alleviated under low-temperature storage by silencing Sly-miR171e with short tandem target mimic technology. Front Nutr. 2022, 9, 906227. [Google Scholar] [CrossRef] [PubMed]
- Krishna, R.; Karkute, S.G.; Ansari, W.A.; Jaiswal, D.K.; Verma, J.P.; Singh, M. Transgenic tomatoes for abiotic stress tolerance: status and way ahead. 3 Biotech. 2019, 9, 143. [Google Scholar] [CrossRef]
- Wai, A.H.; Naing, A.H.; Lee, D.J.; Chang, K.K.; Chung, M.Y. Molecular genetic approaches for enhancing stress tolerance and fruit quality of tomato. Plant Biotechnol. Rep. 2020, 14, 515–537. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef]
- Al-Harbi, A.R.; Wahb-Allah, M.A.; Abu-Muriefah, S.S. Salinity and nitrogen level affects germination, emergence, and seedling growth of tomato. Int. J. Veg. Sci. 2008, 14, 380–392. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Xu, N.; Chu, Y.; Chen, H.; Li, X.; Wu, Q.; Jin, L.; Wang, G.; Huang, J. Rice transcription factor OsMADS25 modulates root growth and confers salinity tolerance via the ABA-mediated regulatory pathway and ROS scavenging. PLoS Genet. 2018, 14, e1007662. [Google Scholar] [CrossRef]
- Llanes, A.; Andrade, A.; Masciarelli, O.; Alemano, S.; Luna, V. Drought and salinity alter endogenous hormonal profiles at the seed germination phase. Seed Sci. Res. 2016, 26, 1–13. [Google Scholar] [CrossRef]
- Ge, L.; Yang, X.; Liu, Y.; Tang, H.; Wang, Q.; Chu, S.; Hu, J.; Zhang, N.; Shi, Q. Improvement of Seed Germination under Salt Stress via Overexpressing Caffeic Acid O-methyltransferase 1 (SlCOMT1) in Solanum lycopersicum L. Int. J. Mol. Sci. 2023, 24, 734. [Google Scholar] [CrossRef]
- Moore, K.; Roberts, L.J. Measurement of lipid peroxidation. Free Radical Res. 1998, 28, 659–671. [Google Scholar] [CrossRef]
- Kant, P.; Kant, S.; Gordon, M.; Shaked, R.; Barak, S. STRESS RESPONSE SUPPRESSOR1 and STRESS RESPONSE SUPPRESSOR2, two DEAD-box RNA helicases that attenuate Arabidopsis responses to multiple abiotic stresses. Plant Physiol. 2007, 145, 814–830. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; Faiq, M.; Khan, M.R.; Tareen, A.K.; Khan, A.; Ullah, A.; Ullah, N.; Huang, J.L. Phytohormones and plant responses to salinity stress: a review. Plant Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The role of gibberellin signalling in plant responses to abiotic stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Miceli, A.; Vetrano, F.; Moncada, A. Effects of foliar application of gibberellic acid on the salt tolerance of tomato and sweet pepper transplants. Horticulturae 2020, 6, 93. [Google Scholar] [CrossRef]
- Yu, Z.; Duan, X.; Luo, L.; Dai, S.; Ding, Z.; Xia, G. How plant hormones mediate salt stress responses. Trends Plant Sci. 2020, 25, 11. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Ito, T.; Fukazawa, J.; Takahashi, Y. Gibberellin induces an increase in cytosolic Ca2+ via a DELLA-independent signaling pathway. Plant Physiol. 2017, 175, 1536–1542. [Google Scholar] [CrossRef]
- Achard, P.; Renou, J.P.; Berthome, R.; Harberd, N.P.; Genschik, P. Plant DELLAs restrain growth and promote survival of adversity by reducing the levels of reactive oxygen species. Curr. Biol. 2008, 18, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Cheng, H.; De Grauwe, L.; Decat, J.; Schoutteten, H.; Moritz, T.; Van Der Straeten, D.; Peng, J.; Harberd, N.P. Integration of plant responses to environmentally activated phytohormonal signals. Science 2006, 311, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gong, X.; Liu, W.; Kong, L.; Si, X.; Guo, S.; Sun, J. Gibberellin mediates spermidine-induced salt tolerance and the expression of GT-3b in cucumber. Plant Physiol. Bioch. 2020, 152, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Massaretto, I.L.; Albaladejo, I.; Purgatto, E.; Flores, F.B.; Plasencia, F.; Egea-Fernandez, J.M.; Bolarin, M.C.; Egea, I. Recovering tomato landraces to simultaneously improve fruit yield and nutritional quality against salt stress. Front Plant Sci. 2018, 9, 1778. [Google Scholar] [CrossRef]
- Koleška, I.; Hasanagić, D.; Todorović, V.; Murtić, S.; Maksimović, I. Grafting influence on the weight and quality of tomato fruit under salt stress. Ann. Appl. Biol. 2018, 172, 187–196. [Google Scholar] [CrossRef]
- Galli, V.; Messias, R.D.S.; Perin, E.C.; Borowski, J.M.; Bamberg, A.L.; Rombaldi, C.V. Mild salt stress improves strawberry fruit quality. LWT-Food Sci. Technol. 2016, 73, 693–699. [Google Scholar] [CrossRef]
- Huang, Y.; Tang, R.; Cao, Q.; Bie, Z. Improving the fruit yield and quality of cucumber by grafting onto the salt tolerant rootstock under NaCl stress. Sci. Hortic. 2009, 122, 26–31. [Google Scholar] [CrossRef]
- Gough, C.; Hobson, G.E. A comparison of the productivity, quality, shelf-life characteristics and consumer reaction to the crop from cherry tomato plants grown at different levels of salinity. J. Hortic. Sci. 1990, 65, 431–439. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Ann. Rev. Plant Phys. 2000, 51, 463–499. [Google Scholar] [CrossRef]
- Grattan, S.R.; Grieve, C.M. Salinity-mineral nutrient relations in horticultural crops. Sci. Hortic. 1998, 78, 127–157. [Google Scholar] [CrossRef]
- De Pascale, S.; Maggio, A.; Fogliano, V.; Ambrosino, P.; Ritieni, A. Irrigation with saline water improves carotenoids content and antioxidant activity of tomato. J. Hortic. Sci. Biotechnol. 2001, 7, 447–453. [Google Scholar] [CrossRef]
- Petersen, K.K.; Willumsen, J.; Kaack, K. Composition and taste of tomatoes as affected by increased salinity and different salinity sources. J. Hortic. Sci. Biotechnol. 1998, 73, 205–15. [Google Scholar] [CrossRef]
- Hernández-Fuentes, A.D.; López-Vargas, E.R.; Pinedo-Espinoza, J.M.; Campos-Montiel, R.G.; Valdés-Reyna, J.; Juárez-Maldonado, A. Postharvest behavior of bioactive compounds in tomato fruits treated with Cu nanoparticles and NaCl stress. Appl. Sci. 2017, 7, 980. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radical Bio. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef] [PubMed]
- Klunklin, W.; Savage, G. Effect on quality characteristics of tomatoes grown under well-watered and drought stress conditions. Foods 2017, 6, 56. [Google Scholar] [CrossRef]
- Martí, R.; Leiva-Brondo, M.; Lahoz, I.; Campillo, C.; Cebolla-Cornejo, J.; Roselló, S. Polyphenol and L-ascorbic acid content in tomato as influenced by high lycopene genotypes and organic farming at different environments. Food Chem. 2018, 239, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Pinedo-Guerrero, Z.H.; Hernández-Fuentes, A.D.; Ortega-Ortiz, H.; Benavides-Mendoza, A.; Cadenas-Pliego, G.; Juárez-Maldonado, A. Cu nanoparticles in hydrogels of chitosan-PVA affects the characteristics of post-harvest and bioactive compounds of jalapeño pepper. Molecules 2017, 22, 926. [Google Scholar] [CrossRef]
- Kedare, S.B.; Singh, R.P. Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Tech. 2011, 48, 412–22. [Google Scholar] [CrossRef]
- Yan, J.; Gu, Y.; Jia, X.; Kang, W.; Pan, S.; Tang, X.; Chen, X.; Tang, G. Effective small RNA destruction by the expression of a short tandem target mimic in Arabidopsis. Plant Cell 2012, 24, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.X.; Xie, S.Y.; Zhang, P.; Wang, Q.; Li, J.K.; Xu, X.B. Attenuation of postharvest peel browning and chilling injury of banana fruit by Astragalus polysaccharides. Postharvest Biol. Technol. 2022, 184, 111783. [Google Scholar] [CrossRef]
- Hu, W.; Huang, C.; Deng, X.; Zhou, S.; Chen, L.; Li, Y.; Wang, C.; Ma, Z.; Yuan, Q.; Wang, Y.; Cai, R.; Liang, X.; Yang, G.; He, G. TaASR1, a transcription factor gene in wheat, confers drought stress tolerance in transgenic tobacco. Plant Cell Environ. 2013, 36, 1449–1464. [Google Scholar] [CrossRef]
- Duan, W.; Ngaffo Mekontso, F.; Li, W.; Tian, J.; Li, J.; Wang, Q.; Xu, X. Alleviation of postharvest rib-edge darkening and chilling injury of carambola fruit by brassinolide under low temperature storage. Sci. Hortic. 2022, 299, 110015. [Google Scholar] [CrossRef]
- Zhu, M.; Chen, G.; Zhou, S.; Tu, Y.; Wang, Y.; Dong, T.; Hu, Z. A new tomato NAC (NAM/ATAF1/2/CUC2) transcription factor, SlNAC4, functions as a positive regulator of fruit ripening and carotenoid accumulation. Plant Cell Physiol. 2014, 55, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Fish, W.W.; Perkins-Veazie, P.; Collins, J.K.A. A quantitative assay for lycopene that utilizes reduced volumes of organic solvents. J. Food Compos. Anal. 2002, 15, 309–317. [Google Scholar] [CrossRef]
- Wan, X.; Wang, Z.Q.; Duan, W.H.; Huang, T.S.; Song, H.M.; Xu, X.B. Knockdown of Sly-miR164a enhanced plant salt tolerance and improved preharvest and postharvest fruit nutrition of tomato. Int J Mol Sci, 2023, 24, 4639. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Pu, H.L.; Shan, S.S.; Zhang, P.; Li, J.K.; Song, H.M.; Xu, X.B. Melatonin enhanced chilling tolerance and alleviated peel browning of banana fruit under low temperature storage. Postharvest Biol. Technol. 2021, 179, 111571. [Google Scholar] [CrossRef]
Figure 1.
Comparative analysis of wild-type (WT) and silencing of Sly-miR171d/e by short tandem target mimic (STTM-miR171d/e) tomato seed germination under salt stress. (A,B) Germination of tomato seed under 50 mM NaCl stress. (C) Germination rate. (D) Primary root length. (E) Fresh weight. Vertical bars represent standard deviations of the means, n = 3. Asterisks indicate statistical difference of the values at P < 0.05 (*) or P < 0.01 (**). All statistical differences were compared with WT, and there was no comparison between transgenic plant groups.
Figure 1.
Comparative analysis of wild-type (WT) and silencing of Sly-miR171d/e by short tandem target mimic (STTM-miR171d/e) tomato seed germination under salt stress. (A,B) Germination of tomato seed under 50 mM NaCl stress. (C) Germination rate. (D) Primary root length. (E) Fresh weight. Vertical bars represent standard deviations of the means, n = 3. Asterisks indicate statistical difference of the values at P < 0.05 (*) or P < 0.01 (**). All statistical differences were compared with WT, and there was no comparison between transgenic plant groups.
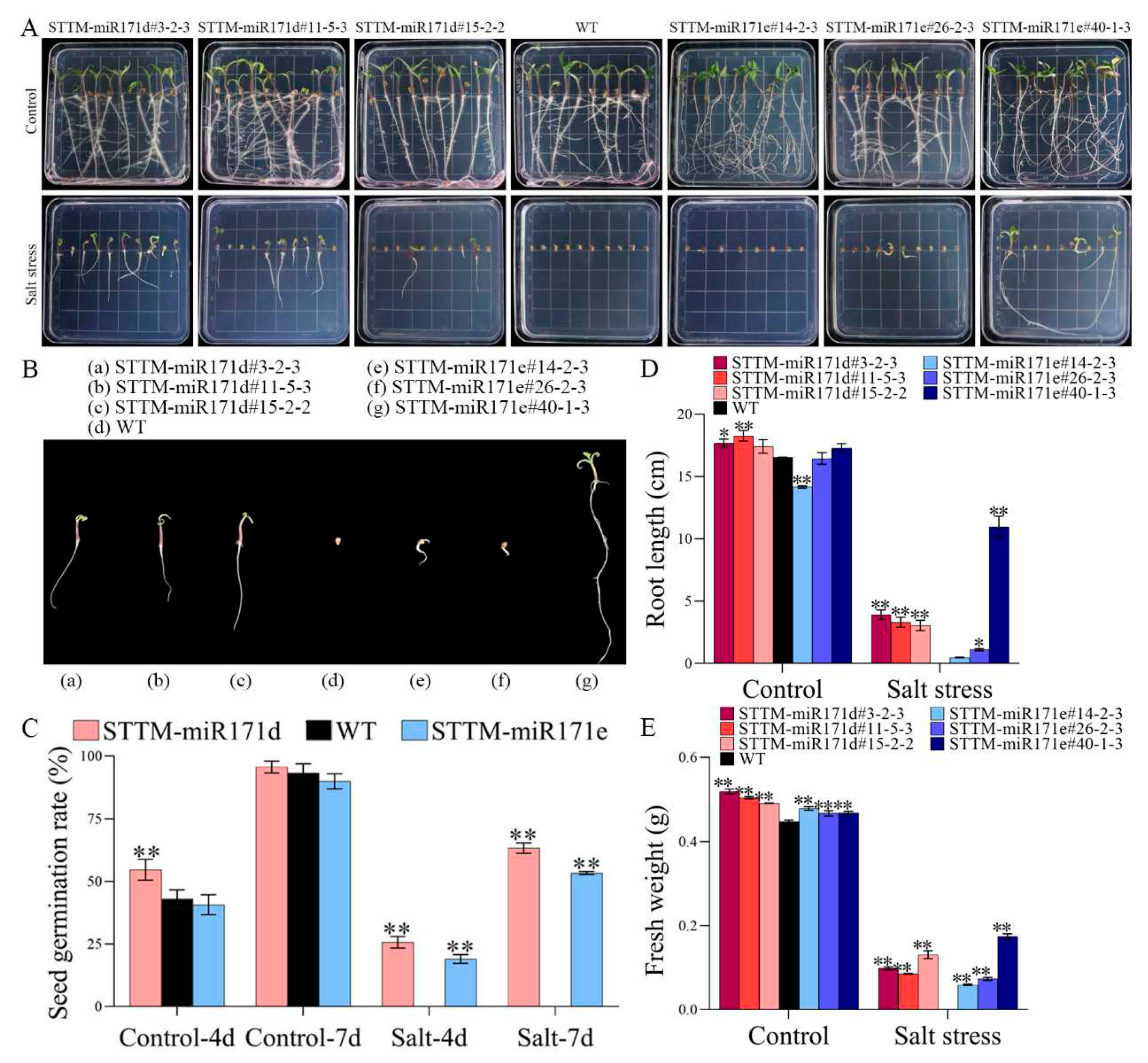
Figure 2.
The growth of WT and STTM-miR171d/e tomato seedlings under salt stress. (A) Growth of tomato seedling under 100 mM NaCl stress. The arrows indicate the fruit on the plant. (B) Plant height. (C) Stem diameter. The expression level of SlGRAS24 (D), Sly-miR171d (E) and Sly-miR171e (F) in tomato seedling root. Vertical bars represent standard deviations of the means, n = 3. Asterisks indicate statistical difference of the values at P < 0.05 (*) or P < 0.01 (**). All statistical differences were compared with WT, and there was no comparison between transgenic plant groups.
Figure 2.
The growth of WT and STTM-miR171d/e tomato seedlings under salt stress. (A) Growth of tomato seedling under 100 mM NaCl stress. The arrows indicate the fruit on the plant. (B) Plant height. (C) Stem diameter. The expression level of SlGRAS24 (D), Sly-miR171d (E) and Sly-miR171e (F) in tomato seedling root. Vertical bars represent standard deviations of the means, n = 3. Asterisks indicate statistical difference of the values at P < 0.05 (*) or P < 0.01 (**). All statistical differences were compared with WT, and there was no comparison between transgenic plant groups.
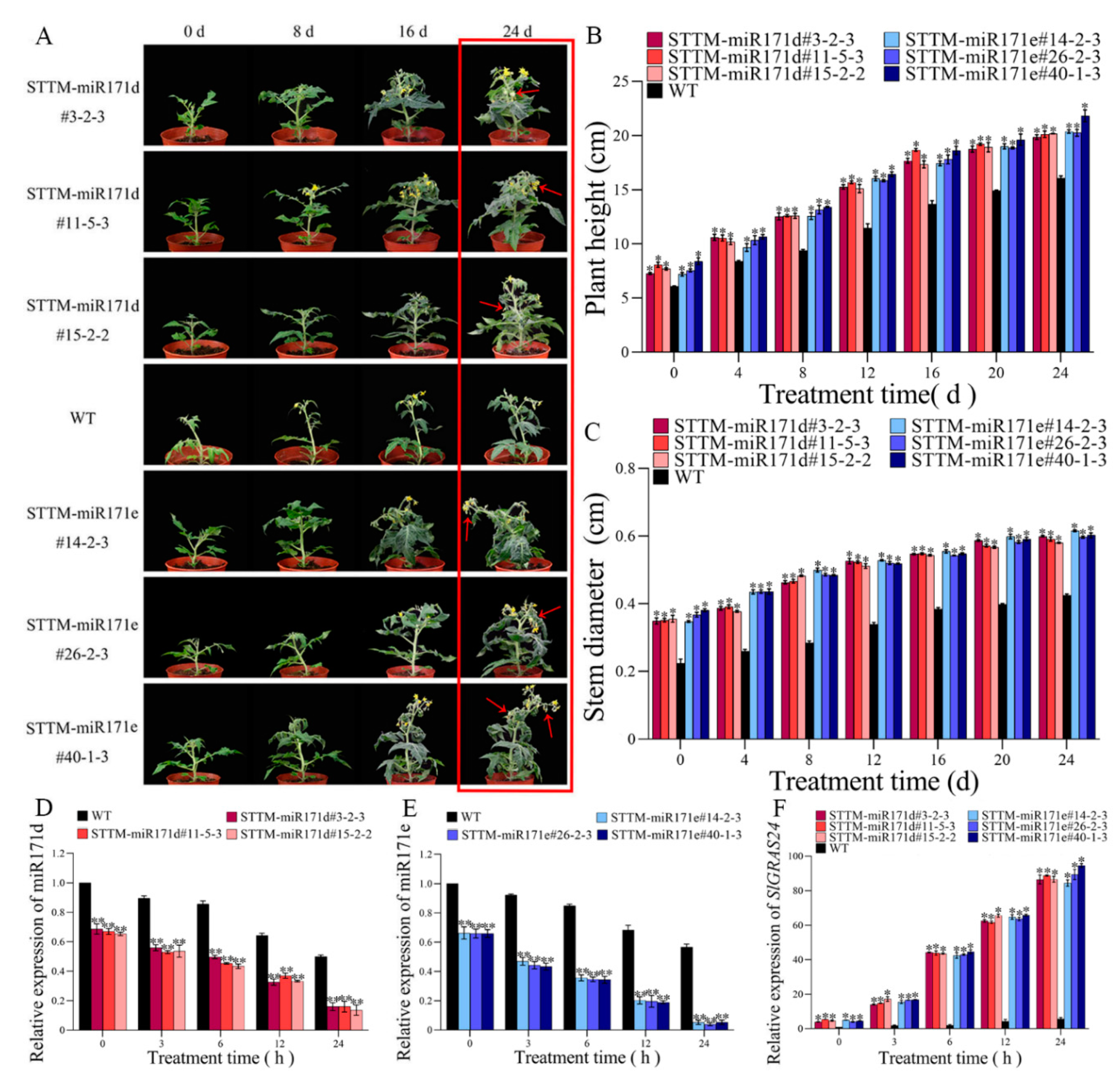
Figure 3.
Analysis of antioxidant capacity in WT and STTM-miR171d/e tomato seedling under salt stress. (A) Comparison of DAB staining and NBT staining of tomato leaf after 14 days of salt treatment. The H2O2 content (B), O2•– content (C), proline content (D), MDA content (E) and GA3 content (F) in tomato seedling root. Changes of chlorophyll content (G) in tomato leaf. Vertical bars represent standard deviations of the means, n = 3. Asterisks indicate statistical difference of the values at P < 0.05 (*) or P < 0.01 (**). All statistical differences were compared with WT, and there was no comparison between transgenic plant groups.
Figure 3.
Analysis of antioxidant capacity in WT and STTM-miR171d/e tomato seedling under salt stress. (A) Comparison of DAB staining and NBT staining of tomato leaf after 14 days of salt treatment. The H2O2 content (B), O2•– content (C), proline content (D), MDA content (E) and GA3 content (F) in tomato seedling root. Changes of chlorophyll content (G) in tomato leaf. Vertical bars represent standard deviations of the means, n = 3. Asterisks indicate statistical difference of the values at P < 0.05 (*) or P < 0.01 (**). All statistical differences were compared with WT, and there was no comparison between transgenic plant groups.
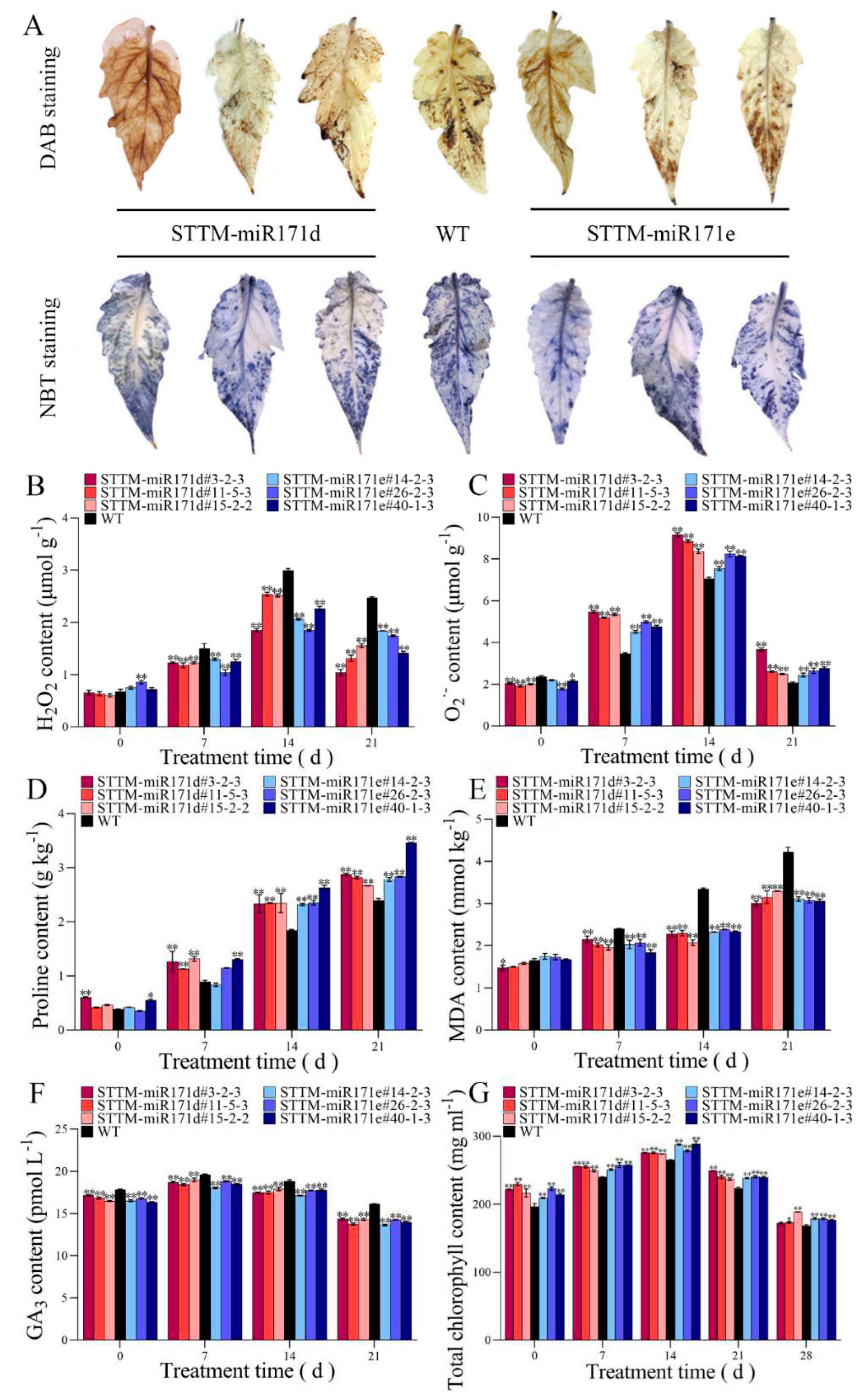
Figure 4.
Color assessment of preharvest fruit of WT and STTM-miR171d/e tomato under salt stress. (A) Comparison of preharvest fruit development at 28 days post-anthesis (dpa) to 40 dpa. Changes of L* value (B), a* value (C), and b* value (D) of preharvest fruit at broken color (Br) day to Br+7 day. Vertical bars represent standard deviations of the means, n = 3. Asterisks indicate statistical difference of the values at P < 0.05 (*). All statistical differences were compared with WT, and there was no comparison between transgenic plant groups.
Figure 4.
Color assessment of preharvest fruit of WT and STTM-miR171d/e tomato under salt stress. (A) Comparison of preharvest fruit development at 28 days post-anthesis (dpa) to 40 dpa. Changes of L* value (B), a* value (C), and b* value (D) of preharvest fruit at broken color (Br) day to Br+7 day. Vertical bars represent standard deviations of the means, n = 3. Asterisks indicate statistical difference of the values at P < 0.05 (*). All statistical differences were compared with WT, and there was no comparison between transgenic plant groups.
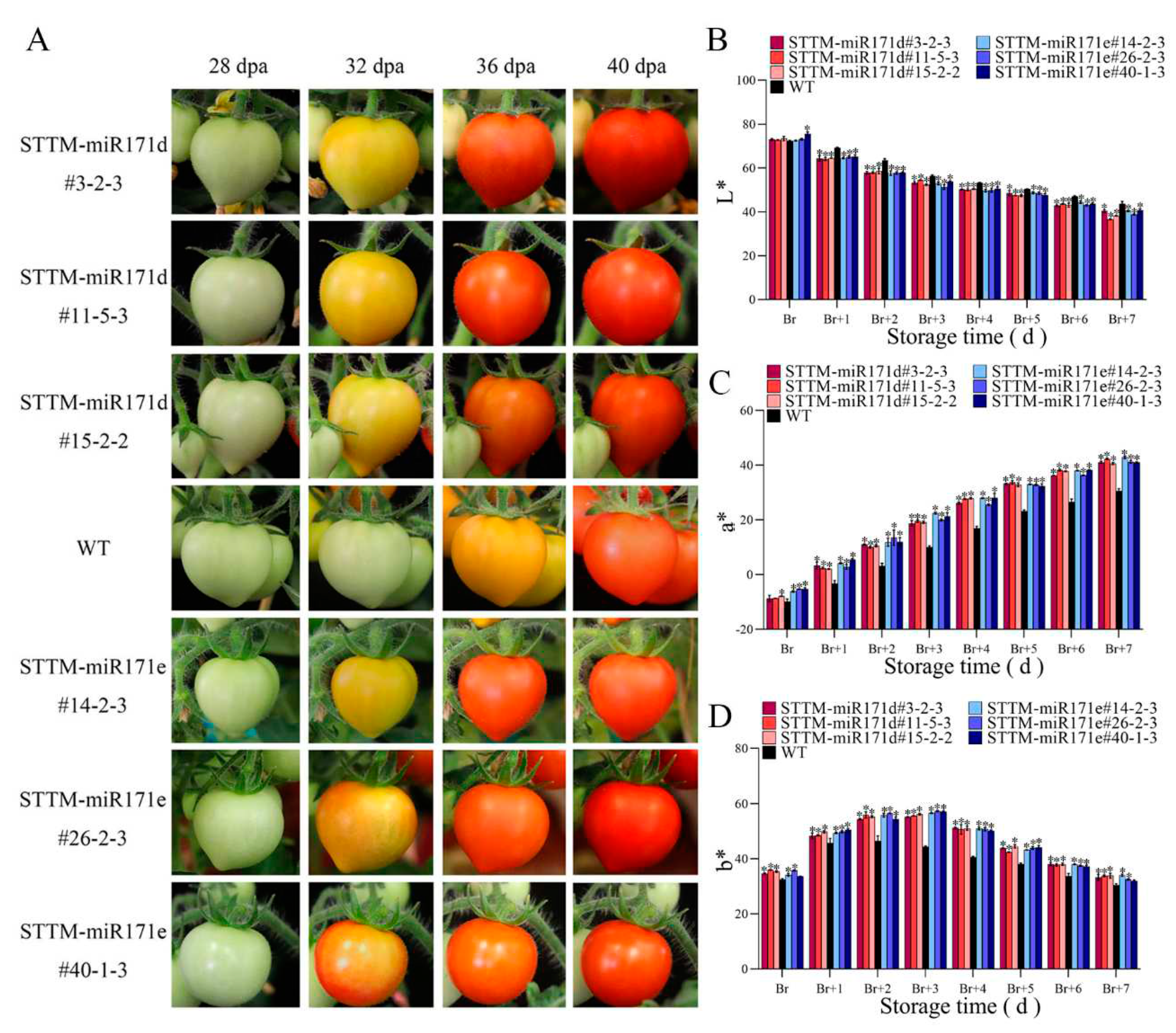
Figure 5.
Color assessment of postharvest fruit of WT and STTM-miR171d/e tomato under salt stress. (A) Comparison of postharvest fruit development at mature green (MG) to Br+7 day. Changes of L* value (B), a* value (C), and b* value (D) of postharvest fruit at Br to Br+7 day. Vertical bars represent standard deviations of the means, n = 3. Asterisks indicate statistical difference of the values at P < 0.05 (*). All statistical differences were compared with WT, and there was no comparison between transgenic plant groups.
Figure 5.
Color assessment of postharvest fruit of WT and STTM-miR171d/e tomato under salt stress. (A) Comparison of postharvest fruit development at mature green (MG) to Br+7 day. Changes of L* value (B), a* value (C), and b* value (D) of postharvest fruit at Br to Br+7 day. Vertical bars represent standard deviations of the means, n = 3. Asterisks indicate statistical difference of the values at P < 0.05 (*). All statistical differences were compared with WT, and there was no comparison between transgenic plant groups.
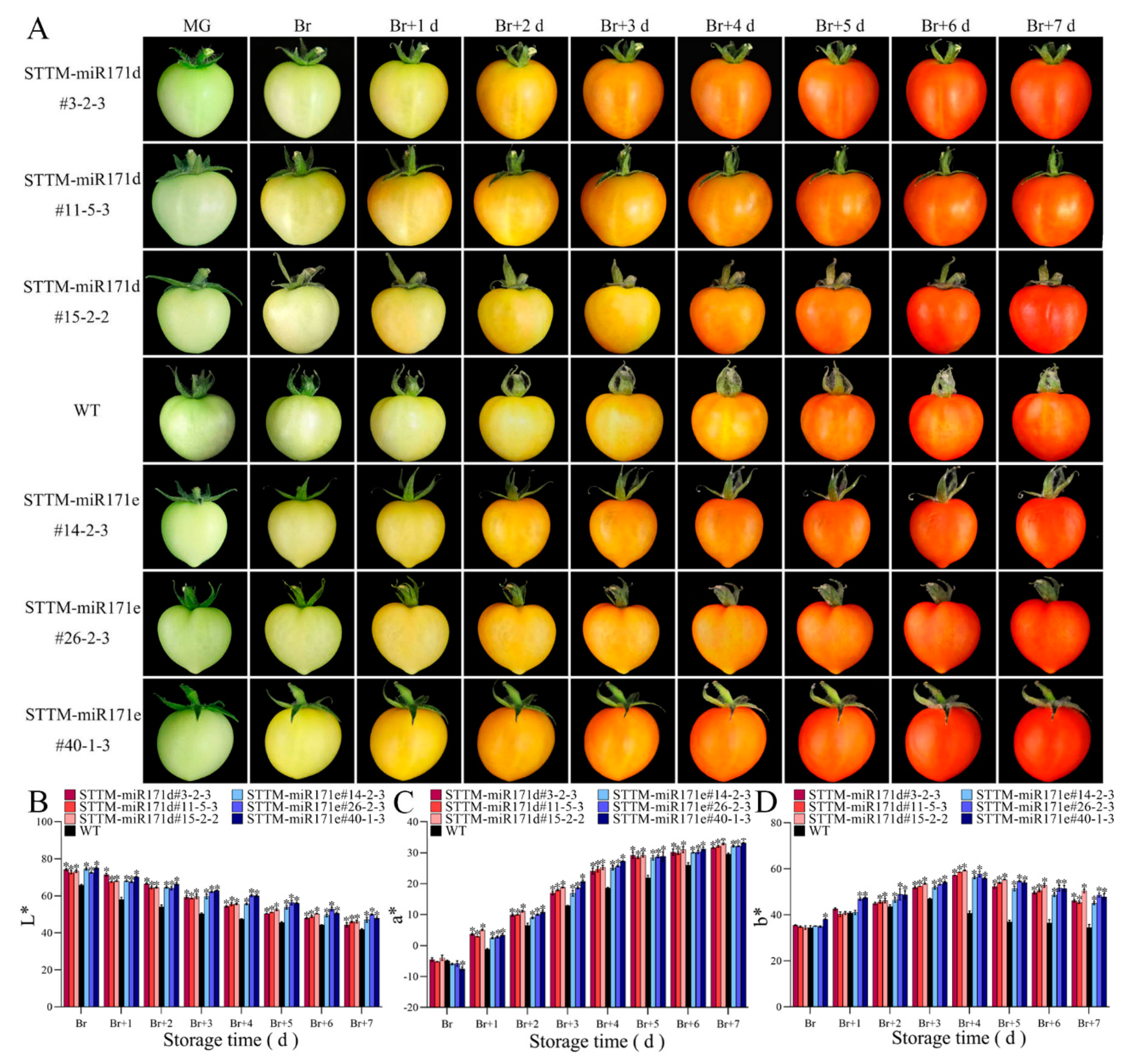
Figure 6.
Comparison of nutritional quality of preharvest fruit and changes of nutritional quality of postharvest fruit in WT and STTM-miR171d/e tomato under salt stress. The lycopene content (A,B), total carotenoids content (C,D), firmness (E,F), soluble solids content (G,H), Vitamin C content (I,J) and total phenolic content (K,L). Vertical bars represent standard deviations of the means, n = 3. Asterisks indicate statistical difference of the values at P < 0.05 (*) or P < 0.01 (**). All statistical differences were compared with WT, and there was no comparison between transgenic plant groups.
Figure 6.
Comparison of nutritional quality of preharvest fruit and changes of nutritional quality of postharvest fruit in WT and STTM-miR171d/e tomato under salt stress. The lycopene content (A,B), total carotenoids content (C,D), firmness (E,F), soluble solids content (G,H), Vitamin C content (I,J) and total phenolic content (K,L). Vertical bars represent standard deviations of the means, n = 3. Asterisks indicate statistical difference of the values at P < 0.05 (*) or P < 0.01 (**). All statistical differences were compared with WT, and there was no comparison between transgenic plant groups.
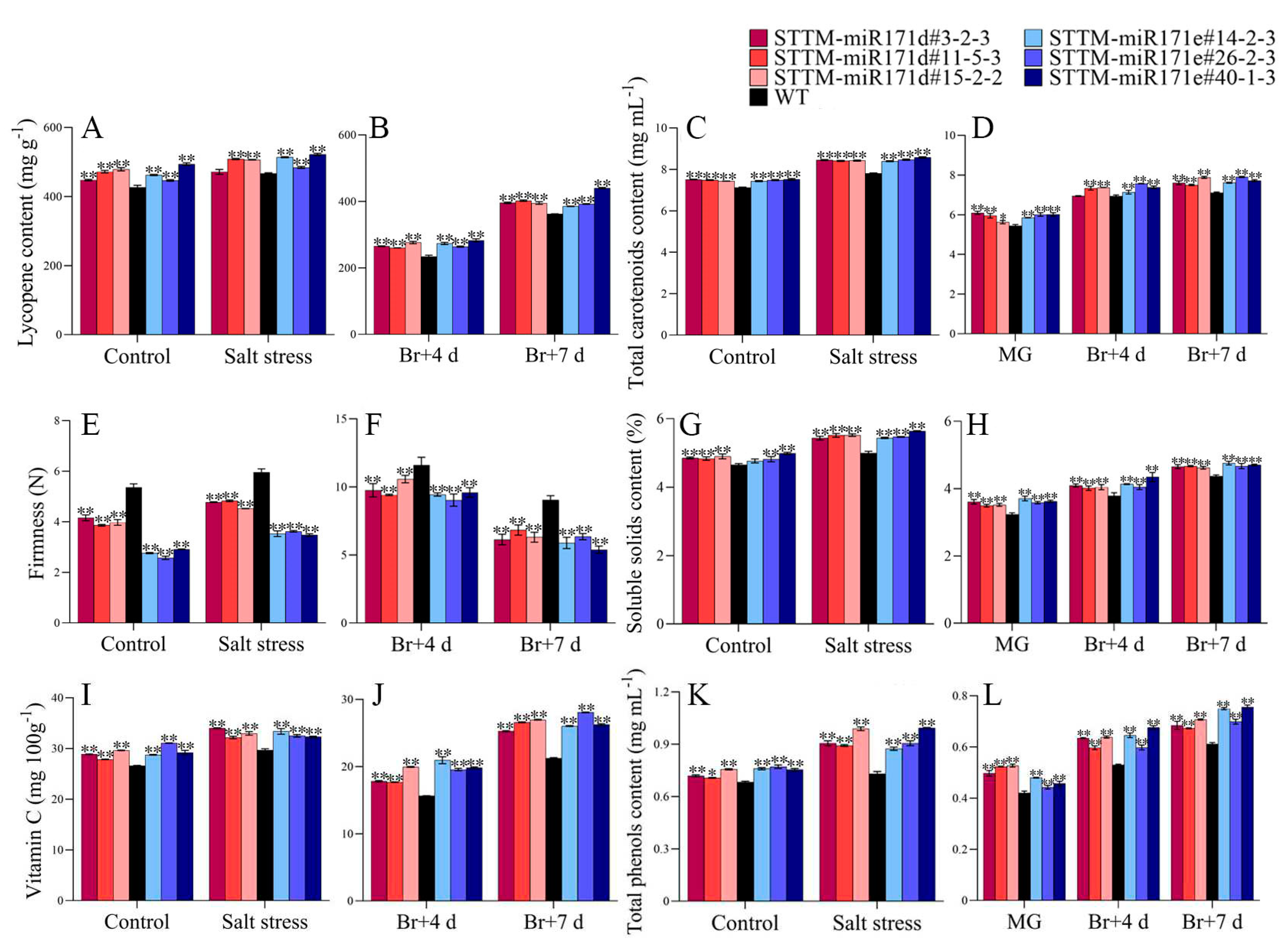

Table 1.
The organic acid content of preharvest and postharvest fruit in WT and STTM-miR171d/e tomato under salt stress. The different letters indicate significant differences (P < 0.05) based on Tukey’s multiple comparison test.
Table 1.
The organic acid content of preharvest and postharvest fruit in WT and STTM-miR171d/e tomato under salt stress. The different letters indicate significant differences (P < 0.05) based on Tukey’s multiple comparison test.
| Individual organic acid (mg g-1 FW) | |||||||
|---|---|---|---|---|---|---|---|
| Quinic acid | Shikimic acid | Pyruvate | Acetic acid | Citric acid | Malic acid | ||
| Control | STTM-miR171d | 0.0657±0.0043a | 0.0076±0.0003b | 0.1375±0.0005 a | 0.0334±0.0018a | 0.1119±0.0106b | 0.0381±0.0093b |
| WT | 0.0324±0.0018c | 0.0081±0.0009b | 0.1055±0.0108b | 0.023±0.0025b | 0.1486±0.0289ab | 0.035±0.0093b | |
| STTM-miR171e | 0.0585±0.0057b | 0.0112±0.0005a | 0.0897±0.0102c | 0.0347±0.0013a | 0.1534±0.0325a | 0.0657±0.0015a | |
| Salt stress | STTM-miR171d | 0.0369±0.0011b | 0.014±0.0002a | 0.0503±0.0006c | 0.0503±0.0017a | 0.1994±0.004a | 0.0935±0.0029a |
| WT | 0.0555±0.0066a | 0.0082±0.0003c | 0.1486±0.0012a | 0.0335±0.0028b | 0.1293±0.0107b | 0.0488±0.0019c | |
| STTM-miR171e | 0.0377±0.0065b | 0.0111±0.0023b | 0.0665±0.0105b | 0.0469±0.0065a | 0.1969±0.0313a | 0.0698±0.0173b | |
| Br+4 d | STTM-miR171d | 0.0476±0.0036b | 0.0026±0.0003b | 0.1063±0.0039b | 0.035±0.0016a | 0.1299±0.006b | 0.0445±0.0025ab |
| WT | 0.0253±0.0047c | 0.0048±0.0011a | 0.0789±0.0098c | 0.0236±0.0026b | 0.0629±0.0283c | 0.0516±0.0231a | |
| STTM-miR171e | 0.0579±0.0059a | 0.0035±0.0004ab | 0.1374±0.0042a | 0.0267±0.0008b | 0.1367±0.0103a | 0.032±0.0038b | |
| Br+7 d | STTM-miR171d | 0.0769±0.0302a | 0.0036±0.0006b | 0.1269±0.0073a | 0.0242±0.0014b | 0.1537±0.0132b | 0.0512±0.0083ab |
| WT | 0.0358±0.0114c | 0.0082±0.0003a | 0.1236±0.0002a | 0.0325±0.0015a | 0.154±0.0045b | 0.06±0.0117a | |
| STTM-miR171e | 0.0521±0.0007b | 0.0086±0.0011a | 0.0873±0.0094b | 0.0351±0.0009a | 0.2452±0.0374a | 0.0445±0.0052b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



