
Submitted:
10 July 2023
Posted:
11 July 2023
You are already at the latest version
Abstract
A new pterygosomatid mite species, Neopterygosoma robertmertensi sp. n. (Acariformes: Pterygosomatidae) was collected from two specimens of Liolaemus robertmertensi (Liolaemidae) from Argentina. This new species is described based on active stages: adults (female and male) and juveniles (deutonymphs, protonymphs, larvae), and quiescent stages (nymphchrysalis, deutochrysalis and imagochrysalis). The changes in morphological characters during the ontogeny of N. robertmertensi have been analyzed in detail. A difference in larval sex morphology was observed for the first time in the family Pterygosomatidae (female larvae differ from male larvae by the shape and size of the idiosoma and position of the genital area). This new mite species is most similar to N. cyanogasteri but can be distinguished by the presence of different leg chaetotaxy patterns of genua IV and femora IV, four to six genital setae, three to five dorsomedial setae and two to three ventromedial setae. Phylogenetic analysis was conducted based on 120 morphological characters of all Neopterygosoma spp. and four outgroup species using the maximum parsimony approach. The results indicated that this species is nested within mites of the chilensis group of Neopterygosoma associated with host species of the section chiliensis of Liolaemus s. str. An updated diagnosis of the chilensis group of Neopterygosoma and an identification key for all species of this genus has been provided.
Keywords:
scale-mites
; Acari
; phylogeny
; ontogeny
; Liolaemus.
1. Introduction
Mites of the genus Neopterygosoma are permanent ectoparasites associated with endemic South American iguanian lizards of the genus Liolaemus (Sauria: Liolaemidae), and until recently, they were placed in the genus Pterygosoma [1,2]. The first species of this genus was described by Dittmar de la Cruz et al. [2] from tree lizards (Liolaemidae) in Argentina, exceeding the geographical range of the genus Pterygosoma. Later on, Fajfer and González-Acuña [1] described six new species from Chilean tree lizards and established a group ligare for mites associated with liolaemids. Nevertheless, the phylogenetic trees constructed by Fajfer [3] clearly showed that the genus Pterygosoma was paraphyletic; therefore, a new genus Neopterygosoma was erected for mites associated with liolaemids lizards [3]. Since then, only one new species, N. schroederi Fajfer, 2020, was described [4].
Currently, mites of the genus Neopterygosoma are represented by eight species associated with the lizards of the genus Liolaemus. They are divided into two groups: chilensis, represented by monoxenous species associated with lizards from Chile and patagonica, represented by a single oligoxenous species, N. patagonica (Dittmar de la Cruz, Morando & Avila, 2004), recorded on several Liolaemus spp. from Argentina [2,5].
Although eight species have been described in the genus Neopterygosoma so far, most of the descriptions were based only on a few adult females. This was dictated by the fact that most of the described mite material was accidentally collected by herpetologists during the investigation of lizards or from museum-preserved specimens, which were often washed before being fixed in formalin or alcohol. However, it should be emphasized that to gain a complete understanding of the mite taxonomy, phylogeny, ecology, and biology, it is essential to study both immature instars and males. In Pterygosomatidae, as in other mites, the description of juvenile stages enabled finding homologous features and establishing the nomenclature used during species description [6,7]. So far, only immatures of one species, N. schroederi, and a male of N. patagonica have been described [2,4]. However, the original description of the male was insufficient, as it only presented the idiosoma’s width and length, chaetotaxy of trochanter-tibiae I−IV, and a vague figure of the idiosoma dorsum without any details. Moreover, the type series (syntypes) comprises five males, all of which have been designated as holotypes (!), and five females. However, exact locality data was not provided, only the provinces and host species were listed separately.
In this paper, we describe a new species, Neopterygosoma robertmertensi sp. n., from Liolaemus robertmertensi from Argentina, including a first comprehensive description of the male within the genus. We extensively examine the post-embryonic stages using scanning electron microscopy, and we note differences between larval males and females for the first time within the family. Additionally, we infer the phylogenetic position of N. robertmertensi based on morphological data. Considering both morphology and phylogeny, this new species belongs to the chilensis group (the first record of Argentinian host species within the group) and is a sister taxon to Chilenian mite species associated with Liolaemus pictus and L. chiliensis. Additionally, we have revised the diagnosis of the chilensis group and provided an updated identification key for the genus (based on females).
2. Materials and Methods
Mite Sampling
The mite specimens were collected from the geckos housed in the herpetological collection of HUJ (abbreviations of the institutions are presented below). All lizards kept in separate jars with 75% ethyl alcohol were examined for mites, which were then removed from the lizards under a stereomicroscope (Nikon SMZ745). Then, the mites were placed in small vials (2 ml) containing 75% ethyl alcohol.
Morphological Analysis
Before mounting in Hoyer’s medium, mite specimens were cleared and softened in Nesbitt’s solution at +45°C for 8–48 h. All specimens were mounted as vouchers using Hoyer’s medium on a glass slide using the standard method [8].
Specimens destined for scanning electron microscopy (SEM) were dehydrated in ethanol, covered with gold, and examined using a Carl Zeiss AG–EVO®40 electron microscope at the Institute of Plant Protection of the National Research Institute in Poznan (IPP NRI), Poland. Additionally, the mites were studied and measured using a Leica DMD108 microscope. All measurements, including scale bars, are given in micrometers (μm). In species descriptions, measurements (ranges) of paratypes are given in parentheses, following the data of the holotype.
Terminology
In the species descriptions, names of the leg and idiosomal setae followed Grandjean [9,10], as described by Norton [6], whereas those of the palpal setae followed Grandjean [11]. Grandjean’s nomenclature [9,10] has been applied to the family Pterygosomatidae by Bochkov and OConnor [7]. The scientific names of the lizards are followed by the Reptile Database [12]. All specimens were deposited in the arachnid collections of HUJ and CSWU. The type material of the Neopterygosoma spp. was loaned from the AMU.
Abbreviations for Museums and Collections
AMU – Department of Animal Morphology, Adam Mickiewicz University, Poznan, Poland;
CSWU – Department of Molecular Biology and Genetics, Institute of Biological Sciences, Cardinal Stefan Wyszynski University in Warsaw, Poland;
HUJ – National Natural History Collections of the Hebrew University of Jerusalem, Israel;
NHM – Natural History Museum, London, the United Kingdom;
ZSM – Bavarian State Collection of Zoology, Munich, Germany.
Phylogeny Reconstructions Methods
For the analysis of phylogenetic relationships between Neopterygosoma species, all species from the genus were used. The outgroup taxa were selected based on the analyses of [3]. As a distant outgroup, Pimeliaphilus podapolipophagus Trägårdh, 1905 was designated, and as a close outgroup the representatives of the genus Geckobia (3 spp.) of the family Pterygosomatidae. We chose G. nitidus because it was a sister taxon to Neopterygosoma in the analyses of Fajfer [3], and G. gerrhopygus and G. hirsti because they were grouped separately in the analysis (see Figure 6 and Figure 7 in [3]).
Cladistic Analysis
All characters were unordered and unweighted. In total, 13 species and 120 morphological characteristics of adult females were included in the analysis (List S1, Table S2). Preparing and editing the data matix were done using NEXUS Data Editor 0.5.0 [13]. The missing states were designated as “?” and inapplicable characters as “-“. The reconstruction of phylogenetic relationships was performed in PAUP 4.0.a 147 for Microsoft Windows [14]. The branch-and-bound option was used for maximum parsimony analysis. Nodal support was evaluated using the Bremer indices calculated using PRAP2 [15]. Analysis of character distributions and drawing and editing of the trees were performed using FigTree v1.4.3 [16], and the final illustrations were made in Adobe Illustrator CS6.
3. Results
3.1. Systematics
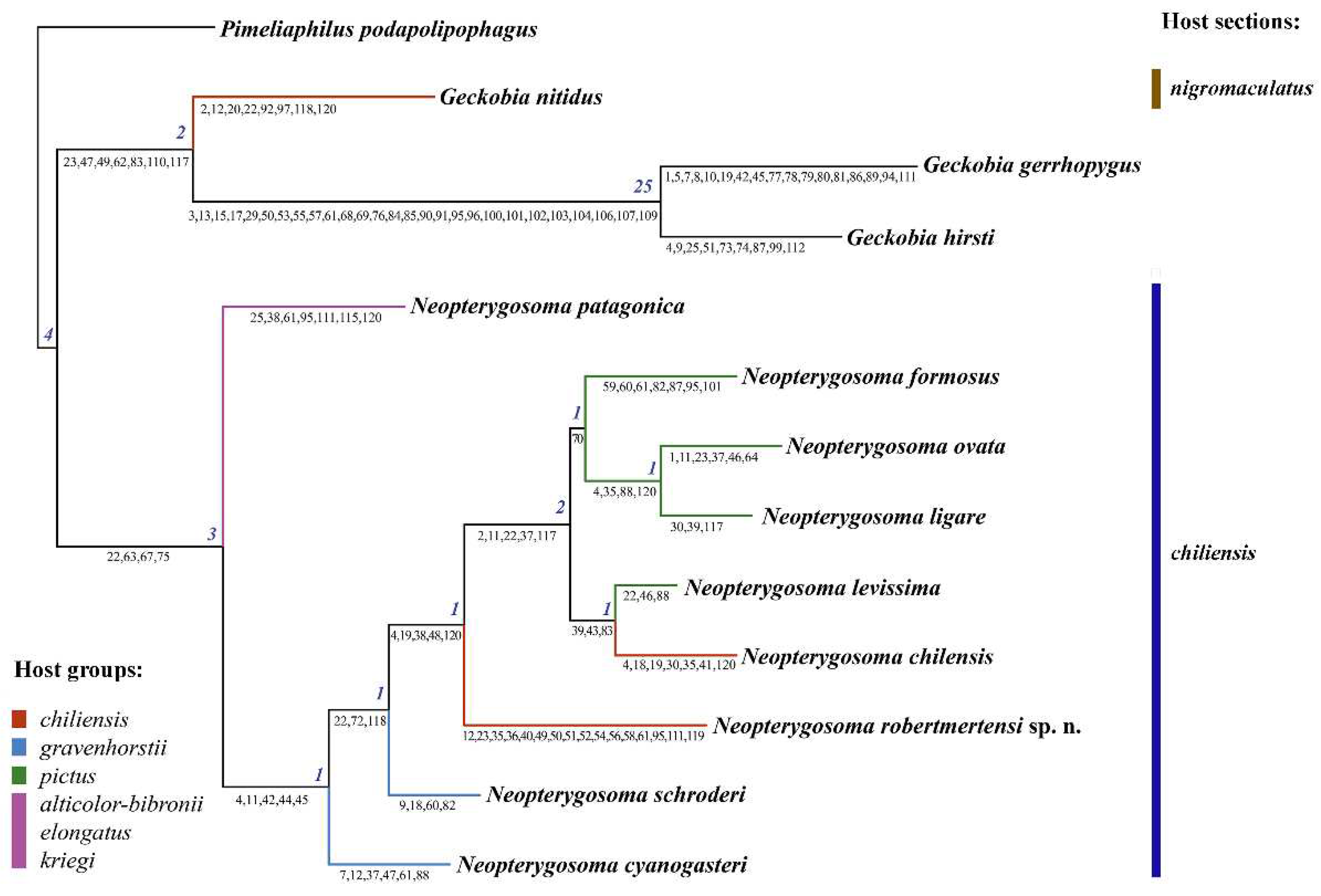
The new species described here was assigned to the chilensis group of the genus Neopterygosoma Fajfer, 2019 of the family Pterygosomatidae Oudemans, 1910, based on morphological and phylogenetic evidence. It possesses the diagnostic morphological features of the chilensis group (see below) and is phylogenetically nested within the chilensis group of Neopterygosoma, but with weak support (Bremer=1, Fig. 15).
3.1.1. Description
Species group chilensis
Diagnosis
Body much wider (1.5–1.8 times) than long. Posteromedial part of idiosomal dorsum and venter with 3–22 pairs of dorsomedial setae or 2–21 pairs of ventromedial setae, respectively. Peripheral setae numerous and much longer than dorsal and ventral setae situated anteriorly, medially and laterally. Setae tc’ and tc” of legs II–IV serrate.
Microhabitat
Under the scales of the whole body.
Distribution and host range
This group is associated with tree lizards of the genus Liolaemus (Sauria: Liolaemidae) from Chile and Argentina.
Species included
Neopterygosoma chilensis (Fajfer & González-Acuña, 2013), N. cyanogasteri (Fajfer & González-Acuña, 2013), N. formosus (Fajfer & González-Acuña, 2013), N. levissima (Fajfer & González-Acuña, 2013), N. ligare (Fajfer & González-Acuña, 2013), N. ovata (Fajfer & González-Acuña, 2013), N. schroederi Fajfer, 2020, Neopterygosoma robertmertensi sp. n
Neopterygosoma robertmertensi sp. n. (Figure 1, Figure 1,Figure 2,Figure 3,Figure 4,Figure 5,Figure 6,Figure 7,Figure 8,Figure 9,Figure 10,Figure 11,Figure 12,Figure 13,Figure 14).
Female (holotype, range for 14 paratypes). Gnathosoma. Chelicerae 145 (145–150) long. Swollen, proximal part of cheliceral base and slender distal half subequal in length, about 75 (70–75) long. Fixed cheliceral digit spinous, about 10 (10) long. Palpal femur and genu with serrate dorsal seta dF and dG, 75 (75–80) and 55 (45–60) long, respectively. Palpal tibia with slightly serrate lateral setae l’Ti and l’’Ti, and with barely serrate ventral seta vTi. Palpal tarsi with 5 setae and solenidion (Figure 4b). Hypostome with rounded apex. Peritremes with clearly visible chambers, about 85 (85–90) long. Subcapitular seta n smooth or with barely discernible serration, 70–75 (75– 85) long. Idiosoma 535 (405–550) long and 973 (715–975) wide. Dorsum (Figure 1) with antero-mid cluster of 56 (53–60) plumose setae (20–30 long) that slightly increase in length from anterior to posterior part of this cluster. These setae situated on smooth weakly sclerotized propodonotal shield (Figure 2a). Laterally to this cluster about 100 (98–110) setae, 30–40 long, on each side present. About 25 (20–25) of these setae inserted ventrally, and among them small eyes present. Medio-lateral and postero-lateral parts of idiosoma with 48 (45–50) pairs of setae that increase in length from anterior to posterior part, 40–135 long. Dorsomedial part with 3 (3–5) pairs of serrate dorsomedial setae (dm). Setae dm1 75 (75–80) long and situated most anteriorly, setae dm2–dm5 90–125 (110–125) long and situated antero-laterally to genital area. Peripheral part of body with about 30 pairs of serrate setae, 10–155 long, inserted dorsally (10–12 pairs) or ventrally (18–23 pairs). Venter (Figure 3) with 2 or 3 (2–3) pairs of serrate setae vm, about 80–95 long, situated laterally to genital area (Figure 2b). Genital series represented by 5 pairs of serrate setae g1–g5, 55–60 (55–60) long, 50–55 (60–65) long, 35–45 (55), 85–90 (75–95) and 70–75 long, respectively. Setae g1–g4 densely serrate and situated dorsally, setae g5 slightly serrate and situated terminally. In 3 paratypes unpaired setae g3 present and in 5 paratypes 6 genital setae present (seta g3 is doubled). Pseudanal setal series represented by 5 pairs of setae ps1–ps5, 75–120 long. Setae ps1–ps3 situated terminally and ps4–ps5 ventrally. Legs. Coxal setation 1a, 1b, 2b, 3a, 3b, 3c and 3d arranged in formula 2–1–4–0. Setae 1a, 3a, 3b situated outside coxal plates. All coxal setae smooth except for slightly serrate setae 3d. Setae of trochanters I–IV: 1–1–1–1, femora I–IV: 5–4–3–2, genua I–IV: 5–4–3–2 and tibiae I–IV: 5–5–5–5. Setae vTrI–IV, vFI–FIII, v”FI–II filiform and smooth, v'GI, v”GI–II, v’TiI–IV, v”TiI–IV, v’FIV, vGIV with barely discernible serration, d'FI–IV, d”FI–III, l’FI, d'GI–IV, d”GI–III, l’GI, dTiI, l’Ti–IV, l”TiI–IV serr ate. Setation of tarsi: I 14 setae (ft, tc’, tc”, p’, p”, it’, it”, a’, a”, u’, u”, vs’, vs”, pl’) and solenidion ω1 (Figure 4a); II 10 setae (tc’, tc”, p’, p”, a’, a”, u’, u”, vs’, vs”) and ω1; III and IV with 10 setae each (tc’, tc”, p’, p”, a’, a”, u’, u”, vs’, vs”). Setae tc’, tc”, it’ and it” of leg I represented by eupathidia; all setae p’ and p” fan-like; setae a’, a”, u’, u”of legs I−IV and tc’, tc” of leg II with barely discernible serration; setae tc’ and tc” of legs III–IV serrate.
Male (range for 13 paratypes). Gnathosoma. Chelicerae 95 long, swollen cheliceral part 40–50 long, slender distal part 45 long. Setae dF filiform and smooth, 50–65 long, setae dG filiform with barely discernible serration, 30–45 long. Supcapitular seta n filiform and smooth, 35–50 long. Each branch of peritremes about 50–70 long. Idiosoma 255–320 long and 435–480 wide. Dorsum (Figure 5) with barely visible propodonotal shield bearing plumose setae grouped in anterior mid-dorsal cluster (34–38 setae); these setae, 10–20 long, progressively elongate from anterior to posterior parts of this cluster. Numerous slightly longer plumose setae, 25–40 long, situated laterally to this cluster. Between them small inconspicuous eyes present most laterally near one long seta, about 80 long. In medial part of the dorsum 3 pairs of serrate setae present: dm1–dm3 about 30–40 long, 45–65 and 60–90 long, respectively. In lateral and posterior parts of idiosoma about 12 pairs of longer slightly serrate setae, 50–100 long, present; most of them situated dorsally, 2–4 pairs situated ventrally in posterior part of idiosoma. Aedeagus 130–140 long. Genital area with 3 pairs of setae, 5–10 long, situated on anal valve and 3 pairs of genital papillae, 10–25 long, situated laterally to anal valve (as in Figure 7). Venter with two pairs of ventromedial setae vm1 and vm2. Setae vm1 40–65 long and setae vm2 70–75 long. Legs. Coxae in formula: 2–1-4–0 and all setae filiform and smooth. Setae 1a, 3a, 3b outside coxal plates. Chaetotaxy oof legs I–IV as in female except for lack of setae on tarsi IV. Setae dTiI–IV, l’TiI–IV, l"TiI–IV, v’TiI–IV, v”TiI–IV, dGI, l’GI, l”GI, v’GI, v”GI, dGII, vGII, l’FII–IV, vFIII–IV, lTrI–IV smooth; setae l’GII, l”GII, l”FII and l’FIII with barely discernible serration; setae l'FII, l"FI, l’FIII–IV, dFI–III and vFI–II serrate.
Imagochrysalis (tritonymph, based on 1 female and 1 male paratype). Gnathosoma reduced, peritremes barely visible (Figure 8B). Legs absent, only coxae I–IV visible. Idiosoma of female chrysalis (inside deutonymphal exoskeleton) 625 long and 690 wide (inside imagochrysalis fully developed coiled female with idiosoma 615 long and 685 wide present). Idiosoma of male imagochrysalis (inside larval integument) 320 long and 425 wide (inside imagochrysalis coiled fully developed male with idiosoma 295 long and 395 wide present).
Deutonymph (range for 9 paratypes). Gnathosoma as in female. Chelicerae about 90–95 long. Slender cheliceral part and swollen distal part subequal in length, about 45–50 long. Setae dF and dG slightly serrate, 55 and 40, respectively. Subcapitular setae n slightly serrate and 50 long. Peritremes 55 long. Idiosoma 305–330 long and 530–560 wide. Dorsum (Figure 9) with smooth propodonotal shield covered with antero-mid cluster of 26–34 setae, about 25 long. Laterally to this cluster about 26 shorter antero-lateral setae, 25–30 long, situated more anteriorly; about 30 longer antero-lateral setae, 45–60 long, situated more posteriorly; and about 10 antero-lateral short setae inserted ventrally (among them one pair of small eyes present). Dorsomedial setae represented by 3 pairs: dm1 about 35 long, dm2 about 50 long and dm3 65 long. Peripheral setae situated dorsally (7−8 pairs) and ventrally (11−12 pairs) and about 105 long. Venter (Figure 10) with 2 ventromedial setae vm1 and vm2. Genital region with 3 setae g1–g3. Setae g1 and g2 20–25 long, setae g3 35–45 long. Pseudanal setal series represented by 3 pairs of setae ps1–ps3, 70−75 long. Legs as in female, except for lack of setae vTrIV.
Deutochrysalis (based on 2 paratypes in exoskeleton of protonymph). Gnathosoma reduced, with barely discernible peritremes. Idiosoma 415–360 long and 620–650 wide. Legs absent, only coxae I–IV present. Inside deutochrysalis fully developed deutonymph present.
Protonymph (range for 5 paratypes). Gnathosoma. Chelicerae 95 long; slender cheliceral part and swollen distal part subequal in length, 45–50 long. Hypostome with rounded apex. Setae dF and dG slightly serrate, 40–60 and 40–45 long, respectively. Subcapitular seta n filiform and smooth, about 50 long. Each branch of peritremes about 60 long. Idiosoma 315–345 long and 535–550 wide. Dorsum (Figure 11) with weakly sclerotized propodonotal shield with densely plumose setae grouped in anterior mid-dorsal cluster (27–42 setae). These setae subequal in length, 20–25 long. Numerous (about 63–67 pairs) of slightly longer plumose setae, 25–40 long, situated laterally to this cluster. Between them small inconspicuous eyes present. In medial part 3 pairs of setae dm1 (30 long), dm2 (55−65) and dm3 (60−70) present, and about 20–28 pairs of postero-lateral setae, 40–95 long. Venter (Figure 12) with setae vm1, 55 long, and about 29 pairs of serrate peripheral setae in postero-lateral part of idiosoma, 60–70 long. These setae situated: ventrally (12 pairs), terminally (7−8 pairs) and dorsally (10−11 pairs). Genital area with 3 pairs of genital setae g1–g3 30, 15 and 25 long, respectively; and with 3 pairs of densely serrate pseudanal setae ps1–3, 70−80 long. Legs. Coxal setae 1a, 1b, 2b, 3a, 3b, 3c filiform and smooth, setae 3d slightly serrate. Setae 1a and 3a situated outside coxal plates. Chaetotaxy pattern of legs I–IV as in female, except for lack of setae vTrIV.
Nymphchrysalis (based on 2 specimens in larval exoskeleton). Gnathosoma reduced, with barely discernible peritremes. Idiosoma 225–240 long and 350–360 wide with completely developed protonymph inside, about 205 long and 330 wide. Legs absent, only coxae I–IV visible.
Larva (range for 8 larval male paratypes and 11 larval female paratypes). Gnathosoma. Chelicerae about 50 long; swollen cheliceral part 20–25 long and slender distal part about 30 long. Fixed cheliceral digit absent. Tarsi with 5 setae and solenidion (Figure 14b). Each part of peritremal branch 35–40 long. Setae dG 20–40 long, setae dF 40–50 long. Subcapitular setae n absent. Idiosoma wider (290–360 wide) than long (170– 250) in female larvae and almost as long as wide in male larvae (155–200 long and 170–215 wide). Dorsum with out propodonotal shield (Figure 14a) and with 11 plumose setae situated as in Figure 13a,c. Five setae situated in anterior part thicker and shorter (15–30 long) than narrower and longer (35–50 long) setae situated in posterior half of idiosoma. Eyes present on lateral margins of idiosoma. Venter (Figure 13b,d) devoid of any setation. Genital area (Figure 14d) with 3 genital setae g1–g3, 10–15 long and 2 pseudanal setae ps1 and ps2. Setae ps1 40−50 long and ps2 30–50 long. Legs. Coxae in formula: 2–0–1; setae 1a, 1b, 3a filiform and smooth. Chaetotaxy of legs I–IV as follows: (5–5–5) (2–2–1) (4–4–3) (0–0–0). Setae dTiI–III, l’TiI–III, l”TiI–III, vTiI–III, dl’GI–III, dl”GI–II, dl’FI–III, dl”FI–III filiform and slightly serrate.
Setae vFI–II with barely discernible serration and setae dFI–III serrate. Setation of tarsi: I 11 setae (ft, p’, p”, it’, a’, a”, u’, u”, vs’, vs”, pl’) and solenidion ω1; II 10 setae (tc’, tc”, p’, p”, a’, a”, u’, u”, vs’, vs”) and ω1; III and IV with 10 setae each (tc’, tc”, p’, p”, a’, a”, u’, u”, vs’, vs”). Setae vs’, vs”, a’, a”, pl’ smooth or with barely discernible serration, setae p’ and p” fan-like, setae tc’, tc” of legs II–III slightly serrate (tc’ longer than tc”), setae ft smooth, setae it’ in form of eupathidion (Figure 12c).
Eggs 170–180 long 150–160 wide.
Type material
Holotype and 8 female, 12 male, 9 deutonymph, 4 protonymph, 2 imagochrysalis, 1 deutochrysalis, 1 nymphchrysalis, 8 male larvae and 10 female larvae paratypes from Liolaemus robertmertensi Hellmich, 1964 (HUJ no. 17923) (Iguania: Liolaemidae), Argentina, Catamarca, 30 km south of Andalgalá, September 1987, coll. O. Pagaburo and Yehudah L. Werner; 7 female, 1 male, 1 deutonymph, 1 nymph chrysalis, 1 protonymph chrysalis, 1 dutonymph chrysalis, and 1 female larva paratypes from same host (HUJ no. 18091) and location, September 1987, coll. O. Pagaburo and Yehudah L. Werner.
Type Material Deposition
Female holotype and most paratypes were deposited in the HUJ (reg. HUJINV-Acari_Pte00003.1−38 and HUJINV-Acari_Pte00004.1−11), except for six female, 3 male, 3 deutonymph and 2 protonymph and 4 larvae paratypes in the CSWU (reg. no. CSWU-Pte20.1.1−16 and Pte20.2.1−2).
Etymology
The species name is derived from the species name of the host.
Differential diagnosis
This species is most similar to Neopterygosoma cyanogasteri from Liolaemus cyanogaster (Duméril and Bibron) from Chile [1]. In females of both species, setation of tarsi I–IV, tibiae I–IV, genua I–III, femora I and III, and trochanters I–IV is the same, fixed cheliceral digit is spinous, palp seta dF is longer than dG, subcapitular seta n is smooth or with barely discernible serration, the antero-mid cluster of dorsal setae is represented by about 60 setae, and five pseudanal setae ps are present. In Neopterygosoma robertmertensi setae lv’GIV, lv’GII and ld’FIV are absent, coxal setae 3a are smooth, 4–6 pairs of serrate genital setae are present, 3–5 pairs of dorsomedial setae and 2 or 3 pairs of ventromedial setae are present. In N. cyanogasteri setae lv’GIV, lv’GII and ld’FIV are present, coxal setae 3a are serrate, one smooth genital seta, 17–21 dorsomedial setae, and 14–18 ventromedial setae are present.
Remarks
Our research used scanning electron microscopy to enhance taxonomic descriptions of the new Neopterygosoma species. As a result, we noticed that in the original description of Neopterygosoma spp. [1], some inaccuracies are mentioned. The detailed photographs revealed that a smooth and weakly sclerotized propodonotal shield is present in all Neopterygosoma mites (Figure 4B) (it appears in protonymph).
3.1.2. Key to Species of Neopterygosoma (Females) (Based on the Key of FAJFER [4])
- 1.
- Body much wider than long (1.5–1.8 times). Setae tc’ and tc” of legs II–IV serrate. Peripheral setae much longer than dorsal and ventral setae situated anteriorly, medially and laterally … chilensis group 2
- -
- Body circular, only slightly wider than long (1.1–1.3 times). Setae tc’ and tc” of legs II–IV smooth. Peripheral setae subequal with anterior, medial and lateral setae on idiosomal dorsum and venter … patagonica group ... N. patagonica (Dittmar de la Cruz, Morando & Avila, 2004)
- 2.
- Five setae on genu I and 5 pseudanal setae ps… 3
- -
- Four setae on genu I and 3 pseudanal setae ps … N. formosus (Fajfer & González–Acuña, 2013)
- 3.
- Four setae on femur II … 4
- -
- Five setae on femur II … 5
- 4.
- Five pseudanal setae present. Setae vTrI–IV densely serrate. Swollen cheliceral part of chelicerae shorter than slender distal part. Subcapitular setae n short (45–65 long) ... N. chilensis (Fajfer & González–Acuña, 2013)
- -
- Four pseudanal setae present. Setae vTrI–IV smooth. Swollen cheliceral part of chelicerae longer than slender distal part. Subcapitular setae n long (about 125 long) ... N. schroederi Fajfer, 2019
- 5.
- Three setae on femur IV. One pair of genital setae g1. Dorsomedial setae dm represented by 15–21 pairs of setae. Ventro–medial setae vm represented by 10–18 pairs ...6
- -
- Two setae on femur IV. Four or five pairs of genital setae. Dorsomedial setae dm represented by 3–5 pairs of setae. Ventromedial setae vm represented by 1–3 pairs … N. robertmertensi sp. n.
- 6.
- Genital setae smooth. Fixed cheliceral digit spinous, palp setae dF serrate only distally, subcapitular setae n serrate … N. cyanogasteri (Fajfer & González-Acuña, 2013)
- -
- Genital setae serrate. Fixed cheliceral digit reduced to rounded structure, palp setae dF serrate on all length, subcapitular setae n smooth … 7
- 7.
- Coxal fields I with 2 setae. Gnathosoma situated apically. Free peritremal branch present. Setae dG serrate on all length … 8
- -
- Coxal fields I with 3 setae. Gnathosoma displaced on dorsal side. Free peritremal branch absent. Setae dG serrate only at distal tip … N. ovata (Fajfer & González-Acuña, 2013)
- 8.
- Antero-medial setae increase in length from anterior to posterior part of setal cluster. Setae a’ and a” of tarsi I slightly serrate. Setae v’TrI–IV serrate. Setae 3a smooth and situated outside coxal plates … N. levissima (Fajfer & González–Acuña, 2013)
- -
- Antero-medial setae subequal in length. Setae a’ and a” of tarsi I smooth. Setae v’TrI–IV with barely discernible serration. Setae 3a slightly serrate and situated on coxal plates … N. ligare (Fajfer & González-Acuña, 2013).
3.2. Phylogeny
3.2.1. Unweighted Parsimony Analysis
The analysis of the data matrix (Table S2) showed that out of 120 characters (List S1), 85 were informative. The analysis with all characters treated as unordered and unweighted was performed with Paup and produced one parsimonious tree (Figure 15). The tree is 219 steps long and has a consistency index (CI) of 0.64; retention index (RI) of 0.56, and rescaled consistency index (RC) of 0.36.
The monophyly of the genus Neopterygosoma is supported by four synapomorphies (Bremer index 3), of which two are unique (length of coxae I, absence of coxal setae 2a and 4a). As expected, the resulting topology in this analysis is very similar to that in Fajfer [3]; in that hypothesis the P. patagonica was the sister taxon to three species of Chilean species (P. chilensis, P. ligare and P. formosus) included in the analysis. Our analysis confirms that N. patagonica from Argentina, considered less specialized (it has a circular body shape that is unable to hide under the host’s scales), is the sister group to all the other species of the genus from Chile, considered more specialized (their idiosoma is wider than long, therefore, they live completely hidden beneath the scales). Its position is also supported by five common synapomorphies (Bremer index 2), of which three are unique (e.g. the presence of much longer setae in the postero-lateral part and peripheral part of idiosoma than in the anterior half of the dorsum).
The new species, L. robertmertensi, is a sister taxon to all species collected from Liolaemus pictus (N. formosus, N. ovata, N. ligare and N. levissima) and L. chiliensis (N. chilensis) and is supported by the presence of five non-unique synapomorphies (Bremer index 1). The node uniting all the above-mentioned mite species collected from the two host species is supported by five non-unique synapomorphies (Bremer index 2). Within the clade, the relationship within the species is weakly supported: N. formosus is a sister taxon to N. ovata + N. ligare (Bremer index 1), and the three species are a sister group to N. levissima + N. chilensis (Bremer index 1). Notably, the positions of both N. schroederi and N. cyanogasteri, are weakly supported by several non-unique synapomorphies (Bremer index of 1).
The only differences between the tree presented in [3] and this tree lay in the position of the outgroup species. In the analysis [3], the genus Geckobia was paraphyletic with G. nitidus as a sister taxon to representatives of species of the genus Neopterygosoma, while Geckobia gerrhopygus + G. hirsti were as a sister taxon to the genera: Gerrhosaurobia + Zanurobia + Ixodiderma + Scaphotrix + Pterygosoma. In our analysis, all the outgroup Geckobia spp. are grouped in a common clade.
Discussion
The genus Liolaemus is the most ecologically diverse and species-rich genus distributed in South America from the high Andes of central Perú to the shores of Tierra del Fuego, and it spans an altitudinal range from sea level to over 5000 m [17]. The liolaemid lizards cover various climatic regimes and inhabit a great diversity of habitats (e.g. arid Atacama desert or humid rainforests). Moreover, the lizards exhibit a wide range of reproductive modes, types of diets, coloration patterns, and body sizes [18]. They also have a long evolutionary history dating back to 18–22 million years ago [19,20].
Currently, the genus includes over 280 species [12], but new species are being discovered at a rapid rate every year (e.g. [21,22]) therefore, it is estimated that the actual number of the species may be much higher. The genus is subdivided into two subgenera: Liolaemus (sensu stricto) and Eulaemus [23], which appear to have separated at least 12.6 million years ago and are currently each divided into several groups. The presence of Neopterygosoma mites has been detected in 12 different species of hosts belonging to Liolaemus s. str. living on both sides of the Andes at different elevations, having different types of scales, coloration patterns, etc. [18,24].
As a rule, mites from different pterygosomatid genera are strictly specific with respect to lizard's hosts and the cospeciation has a strong influence on the architecture of host-parasite relationships within the family Pterygosomatidae [3]. All representatives of the genus Neopterygosoma are monoxenous parasites (the chilensis group) except for N. patagonica collected from several Liolaemus spp. (oligoxenous parasite). Since host species from the same communities (these host taxa distributions partially overlap [17]) do not carry the same set of parasite species we can expect to observe at least partially parallel evolution of Neopterygosoma mites of the chilensis group and Liolaemus hosts.
Nonetheless, the co-phylogenetic studies require phylogenetic hypotheses or data matrices for both lineages involved in the coevolutionary process. So far, the relationships between Liolaemus lizards at the species level are still questionable (e.g. [19,25]). Recently, Troncoso-Palacios et al. [26] conducted a phylogenetic study of the relationship of species of Liolaemus s. str. (based on three fragments of the mitochondrial genome); as a result, the species were divided into two main clades named: chiliensis and nigromaculatus sections. These findings were congruent with the phylogenetical tree (Figure 3 in [17]) based on previous works [19,25,27]. Until now, all Neopterygosoma spp. are associated with closely-related hosts belonging to the chiliensis section, whereas representatives of another pterygosomatid genus, i.e. Geckobia nitidus and G. zapallarensis were collected from lizards of the nigromaculatus section [28] (marked on Figure 15).
However, not all the host species groups were recovered monophyletic in Troncoso-Palacios et al. [26] work, therefore Parenza et al. [29] infer a robust phylogeny (based on 541 ultra-conserved elements and 44 protein-coding genes) for a Chilenian clade of Liolaemus s. str. using representatives of all thirteen groups. As a result, only the relationship among the major Chilean clade of Liolaemus was resolved, as in previous studies [26] (Figure 15). All mites of the chilensis group (i.e. monoxenous 'more specialized' mite species ) have been associated with closely-related hosts belonging to three host groups of [26] i.e.: robertmertensi, gravenhorsti, and pictus. The pterygosomatids have been found on all representatives within the two former groups except for L. sanjuanensis (robertmertensi group) and L. gravenhorsti (gravenhorsti group), which suggest that checking numerous host specimens of the two species for mites might lead to new species descriptions.
The highest number of Neopterygosoma spp. was described from a single host species − L. pictus (4 spp.) belonging to the pictus group including 11 host species. However, the number of species in this group is debatable because a few species have been treated as subspecies of L. pictus [30,31] or synonymized with L. pictus [32]. This host species has a wide distribution and forms a local population at low elevations (0-1600 a.s.l.) on both sides of the Andes, whereas the remaining Liolaemus spp. live either in the eastern or western slopes of the mountains [19]. It is unknown if the mite species occupy the full geographical range of their main host because so far, they have been found only in Isla Mocha (Arauco Province, Chile), although attempts to collect the mites from different localities were made (by M. Fajfer in ZSM and NHM). This could be interpreted as a consequence of recent evolution of new mite species which are competing on the same host, therefore, further studies may prove that this group of parasites undergoes rapid adaptive radiation.
Our phylogenetic analysis shows that N. patagonica is a sister taxon to all monoxenous mites of the chilensis group. It agrees with the findings of Fajfer [3]. P. patagonica inhabits various host species of three different groups (see Figure 15) [17, 26], which might suggest that this mite species association is not fully recovered, and we can expect even more multi-host associations. P. patagonica, due to its circular shape of idiosoma, is morphologically unable to take shelter under the scales, therefore, most of its idiosoma protrudes beyond the scales. It probably allows the mite, by virtue of their effective dispersal abilities, to switch off quickly from a host when the opportunity arises, locate and colonize another host. It is probable especially if the host species, as in this case, share the same diet and occur at least partially in the same habitat [17].
The phylogenetical analysis indicates that the newly described species, N. robertmertensi, is nested within the mites of the chilensis group of Neopterygosoma associated with species of the section chiliensis of Liolaemus s. str. Its placement is also confirmed by a set of morphological features (see Figure 15), although the Bremmer support is only 1. The reason for this may be that N. robertmertensi has many unique features (e.g. the number of dorsomedial, ventromedial, or genital setae i.e. characters 36, 40, and 49–52 in Figure 15, respectively), which do not match the description of the chilensis group provided in [4]. Therefore, a revised description of the species group is presented here.
For the first time, we collected all mites from the host specimens that were preserved directly after collection. As a result, we collected hundreds of mites which were completely hidden beneath the lizard's scales. We found 1−12 specimens under a single scale, and the mites inhabited each body part of the host specimens. This lack of topical (habitat) specificity is quite surprising because in pterygosomatids living under the scales (such as Pterygosoma or Geckobia), a high preference towards the microhabitat on the host body is observed [33,34].
This large number of mites allowed us to observe morphological diversity among juveniles and adults and to illustrate for the first time the complete morphological ontogeny of these mites. For the first time in the family Pterygosomatidae, we were able to determine differences between the sexes of larvae. In male larvae of N. robertmertensi, the idiosoma is smaller and almost as long as wide (155–200 long and 170–215 wide), the genital region is situated ventrally, and the male develops directly in chrysalis inside the larval integument. In female larvae, the idiosoma is bigger and wider than long (170– 250 long and 290–360 wide), the genital region is situated terminally and the life cycle of the larva consists of both: active stages that feed on blood (protonymph, deutonymph, adult) and legless inactive stages (nymphchrysalis, deutochrysalis and imagochrysalis).
Our study shows that a female larva forms a chrysalis that resembles those found in other pterygosomatids (e.g. see Figure 8C in [35]). Inside the chrysalis, a coiled protonymph develops. After molting, the newly emerged protonymph is larger than larva, and we observe the appearance of: four pair of legs with the full set of setae on femora–tarsi IV, numerous idiosomal setae arranged similarly to subsequent stages, subcapitular setae n, weakly sclerotized small propodonotal shield, additional setae ps3 in the genital region, leg setae on coxae II–III (2b, 3b, 3c, 3d), genua-trochanter I–III (v'GI–III, v"G–III, l'GI, vFI, vTrI–III), and tarsi I, i.e. it"(ζ), tc' (ζ) and tc"(ζ).
In the protonymph integument, we observed a deutochrysalis with a completely formed coiled deutonymph. This stage differs from a protonymph by the presence of much smaller gnathosoma and longer palpal setae (dF, dG), fewer setae on the mid-dorsal cluster, and the arrangement of setae (fewer in number) that resembles that in females. In the imagochrysalis (tritonymph), develops an adult female. It differs from a deutonymph by the size of the idiosoma, the presence of additional two or three genital setae (g4–g6) and pseudanal setae (ps4–ps5), and ventral setae on trochanter IV. The males develop directly in the chrysalis inside the exoskeleton of larvae.
At this point, it is unclear if the presence of both male and female larvae is unique for the genus Neopterygosoma. In Pterygosomatidae, as a rule, the description of juvenile morphology is often neglected. It can be caused by several reasons such as: (i) the difficulty in associating juveniles with an adult if the adults are missing in the sample, (ii) a small number of specimens found on hosts in museum collections (the mites might fall off the host during its preservation), and finally (iii) the presence of only female mites on hosts, which may be explained by the short duration of their juvenile stages or (iv) small size and transparency of the juvenile stages which make them difficult to notice on the hosts.
It is interesting to note that the larvae of Neopterygosoma differ from those of other genera, such as Pterygosoma or Geckobia, due to the absence of setae on tarsi I, specifically it"(ζ), tc' (ζ) and tc"(ζ). In other pterygosomatid larvae, only one fan-like proral setae p', one simple tectal seta tc', and paired iterals it' and it" in the form of eupathidia are present. Additionally, Norton's description of leg chaetotaxy [6], based on Grandjean's work [10,11], referred to the iterals as "post-larval setae" that are added in the protonymph stage. Yet, in Neopterygosoma spp. larvae, there is only one euphatidial setae it' while in contrast, the larvae of Pterygosoma have a pair of iterals (it’ and it”).
Conclusions
In this research, we meticulously described and illustrated the morphology of the new species of pterygosomatid mite, Neopterygosoma robertmertensi, using scanning electron microphotography. As a result, we found new morphological features which were not recognized in previous studies of Neopterygosoma spp. such as the presence of a weakly sclerotized propodonotal shield. We observed the species morphological ontogeny and analyzed the main morphological differences between juvenile stages. For the first time in Pterygosomatidae, we observed both male and female larvae that differ mainly by the size and shape of idiosoma and from other pterygosomatid larvae by chaetotaxy of tarsi I. Additionally, the phylogenetic analysis showed that this species is nested within the chilensis group of Neopterygosoma which was consistent with the morphological analysis. Neopterygosoma mites occur only on hosts belonging to three groups of the chiliensis section of the subgenus Liolaemus s. str. whose distributions partially overlap. Nonetheless, the hosts do not carry the same sets of parasite species. It suggests that mites of the chilensis group might be a good fit for cophylogenetic studies, especially if we take into account the fact that some studies conducted on pterygosomatid mites revealed a cophylogenetic pattern [3].
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, List S1 List of morphological characters and character states used in the analyses. Table S2: Matrix of morphological characters used in the phylogenetical analysis.
Author Contributions
Conceptualization, M.F. and M.S.; methodology, M.F. and M.S; investigation, M.F.; resources and material collection, M.F.; writing—original draft preparation, M.F.; writing—review and editing, M.F. and M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a grant from the European Commission’s (FP6) Integrated Infrastructure Initiative programme SYNTHESYS+ IL-TAF (2021).
Informed Consent Statement
Not applicable.
Data Availability Statement
The material is stored in Cardinal Stefan Wyszynski University (Warsaw, Poland) and will be shared upon reasonable request to Monika Fajfer.
Acknowledgments
I want to express my gratitude to Prof. Dror Hawlena and Dr. Boaz Shacham from HUJI for their invaluable assistance during the mite collection process, as well as to Efrat Gavish-Regev from HUJI for lending us the mites. Additionally, I am extremely appreciative of Magdalena Gawlak from IPP NRI for capturing SEM photos of the mites.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fajfer, M.; González-Acuña, D. Pterygosomatid mites of a new species group ligare (Acariformes: Pterygosomatidae: Pterygosoma) parasitizing tree iguanas (Squamata: Liolaemidae: Liolaemus). Zootaxa 2013, 3693, 301–319. [Google Scholar] [CrossRef] [PubMed]
- Dittmar de la Cruz, K.; Morando, M.; Avila, L. Description of a new pterygosomatid mite (Acari: Actinedida: Pterygosomatidae) parasitic on Liolaemus spp. (Iguania: Liolaemini) from Argentina. Zootaxa 2004, 521, 1–6. [Google Scholar] [CrossRef]
- Fajfer, M. Systematics of reptile-associated scale mites of the genus Pterygosoma (Acariformes: Pterygosomatidae) derived from external morphology. Zootaxa 2019, 4603. [Google Scholar] [CrossRef] [PubMed]
- Fajfer, M. A systematic revision of the genus Neopterygosoma Fajfer, 2019 (Acariformes: Pterygosomatidae) with the description of a new species. Syst. Parasitol. 2020, 97, 535–551. [Google Scholar] [CrossRef] [PubMed]
- Fajfer, M. Redescription of Pterygosoma patagonica (Acariformes: Pterygosomatidae) with new host and distribution data. Int. J. Acarol. 2014, 40, 160–164. [Google Scholar] [CrossRef]
- Norton, R.A. A. A review of F. Grandjean's system of leg chaetotaxy in the Oribatei and its application to the Damaeidae. In Biology of Oribatid Mites, 1st ed.; Dindal, D.L., Ed.; S.U.N.Y College of Environmental Science and Forestry: Syracuse, NY, USA, 1977; pp. 33–61. [Google Scholar]
- Bochkov, A.V.; OConnor, B.M. A review of the external morphology of the family Pterygosomatidae and its systematic position within the Prostigmata (Acari: Acariformes). Parasitologiya 2006, 40, 201–214. [Google Scholar]
- Krantz, G.W.; Walter, D.E. A Manual of Acarology; Texas Tech University Press: Lubbock, TX, USA, 2009. [Google Scholar]
- Grandjean, F. Les Segments Post-Larvaires de L’hystérosoma Chez Les Oribates (Acariens). Bull. Soc. Zool. Fr. 1939, 64, 273–284. [Google Scholar]
- Grandjean, F. Observations sur les Acariens de la famille des Stigmaeidae. Arch. Sci. Phys. Nat. 1944, 26, 103–1131. [Google Scholar]
- Grandjean, F. Au sujet de l'organe de Claparede, des eupathides multiples et des taenidies mandiubulaires chez les Acariens actinochitineux. Arch. Sci. Phys. Nat. 1946, 28, 63–87. [Google Scholar]
- The Reptile Database. Available online: http://www.reptile-database.org (accessed on 1 25 June 2023).
- Page, R.D.M. NDE, NEXUS Data Editor 0.5.0; University of Glasgow: Glasgow, Scotland, 2001. [Google Scholar]
- Swofford, D.L. PAUP*. In Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4; Sinauer Associates: Sunderland, MA, USA, 2002; p. 144. [Google Scholar]
- Müller, K. PRAP—Computation of Bremer support for large data sets. Mol. Phylogenet. Evol. 2004, 31, 780–782. [Google Scholar] [CrossRef]
- Rambaut, A.; Institute of Evolutionary Biology, University of Edinburgh, Edinburgh. FigTree v1.3.1. 2010. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 10 June 2023).
- Pincheira-Donoso, D.; Scolaro, J.; Sura, P. A monographic catalogue on the systematics and phylogeny of the South American iguanian lizard family Liolaemidae (Squamata, Iguania). Zootaxa 2008, 1800, 1–85. [Google Scholar] [CrossRef]
- Pincheira-Donoso, D.; Tregenza, T.; Hodgson, D.J. Body size evolution in South American Liolaemus lizards of the boulengeri clade: A contrasting reassessment. J. Evol. Biol. 2007, 20, 2067–2071. [Google Scholar] [CrossRef] [PubMed]
- Schulte, J.A.; Macey, J.R.; Espinoza, R.E.; Larson, A. Phylogenetic relationships in the iguanid lizard genus Liolaemus: Multiple origins of viviparous reproduction and evidence for recurring Andean vicariance and dispersal. Zool. J. Linn. Soc. 2000, 69, 75–102. [Google Scholar] [CrossRef]
- Fontanella, F.M.; Olave, M.; Avila, L.J.; Morando, M. Molecular dating and diversification of the South American lizard genus Liolaemus (subgenus Eulaemus) based on nuclear and mitochondrial DNA sequences. Zool. J. Linn. Soc. 2012, 164, 825–835. [Google Scholar] [CrossRef]
- Avila, L.J.; Perez, C.H.F.; Minoli, I.; Medina, C.D.; Sites, J.W., Jr.; Morando, M. New species of Liolaemus (Reptilia, Squamata, Liolaemini) of the Liolaemus donosobarrosi clade from northwestern Patagonia, Neuquén province, Argentina. Zootaxa, 2017, 4362, 535–563. [Google Scholar] [CrossRef]
- Sánchez, K.I.; Morando, M.; Avila, L.J. A new lizard species of the Liolaemus kingii group (Squamata: Liolaemidae) from northwestern Chubut province (Argentina). Zootaxa 2023, 235–255. [Google Scholar] [CrossRef]
- Laurent, R.F. Contribución al conocimiento de la estructura taxonómica del género Liolaemus Wiegmann (Iguanidae). Bol. Asoc. Herp. Arg. 1983, 1, 16–18. [Google Scholar]
- Fernández, M.G.; Abdala, C.S.; Ruiz-Monachesi, M.R.; Semham, R.V.; Quinteros, A.S. Redescription of Liolaemus robertmertensi, Hellmich 1964 (Iguania: Liolaemidae) with description of a new species. Cuad. herpetol. 2021, 35, 65–78. [Google Scholar] [CrossRef]
- Espinoza, R.E.; Wiens, J.J.; Tracy, C.R. Recurrent evolution of herbivory in small, cold-climate lizards: Breaking the ecophysiological rules or reptilian herbivory. Proc. Natl. Acad. Sci. USA 2004, 101, 16819–16824. [Google Scholar] [CrossRef]
- Troncoso-Palacios, J.; Schulte, J.A.; Marambio-Alfaro, Y.; Hiriart, D. Phenotypic variation, phylogenetic position and new distributional records for the poorly known Liolaemus silvai Ortiz, 1989 (Iguania: Iguanidae: Liolaemini). S. Am. J. Herpetol. 2015, 10, 71–81. [Google Scholar] [CrossRef]
- Schulte, J.A.; Losos, J.B.; Cruz, F.B.; Núñez, H. The relationship between morphology, escape behaviour and microhabitat occupation in the lizard clade Liolaemus (Iguanidae: Tropidurinae: Liolaemini). J. Evol. Biol. 2004, 17, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Fajfer, M. Mites of the new species group nitidus (Acariformes: Pterygosomatidae: Geckobia), parasites of lizards in South America. Syst. Parasitol. 2015, 90, 213–222. [Google Scholar] [CrossRef]
- Panzera, A.; Leaché, A.D.; D’Elia, G.; Victoriano, P.F. Phylogenomic analysis of the Chilean clade of Liolaemus lizards (Squamata: Liolaemidae) based on sequence capture data. PeerJ 2017, 5, e3941. [Google Scholar] [CrossRef]
- Pincheira-Donoso, D.; Núñez, H. Las especies chilenas del género Liolaemus Wiegmann, 1834 (Iguania Tropiduridae: Liolaeminae). Taxonomía, sistemática y evolución. Mus. Nac. Hist. Nat. Chile, Publ. Occ. 2005, 59, 7–486. [Google Scholar]
- Ruiz de Gamboa, M. Lista actualizada de los reptiles de Chile. Bol. Chil. Herp. 2016, 3, 7–12. [Google Scholar]
- Lobo, F.; Espinoza, R.E.; Quinteros, S. A critical review and systematic discussion of recent classification proposals for liolaemid lizards. Zootaxa 2010, 2549, 1–30. [Google Scholar] [CrossRef]
- Fajfer, M.; Karanth, P. New morphological and molecular data reveal an underestimation of species diversity of mites of the genus Geckobia (Acariformes: Pterygosomatidae) in India. Diversity 2022, 14, 1064. [Google Scholar] [CrossRef]
- Fajfer, M. Three new species of scale mites (Acari: Pterygosomatidae) parasitizing Agama sankaranica (Sauria: Agamidae). Zootaxa 2013, 3700, 271–272. [Google Scholar] [CrossRef] [PubMed]
- Fajfer, M. Two new species of the genus Pterygosoma (Acariformes: Pterygosomatidae) parasitizing agamid lizards (Sauria: Agamidae) from the Indian subcontinent. Acta Parasitol. 2016, 61, 343–354. [Google Scholar] [CrossRef]
Figure 1.
Neopterygosoma robertmertensi sp. n., female in dorsal view.
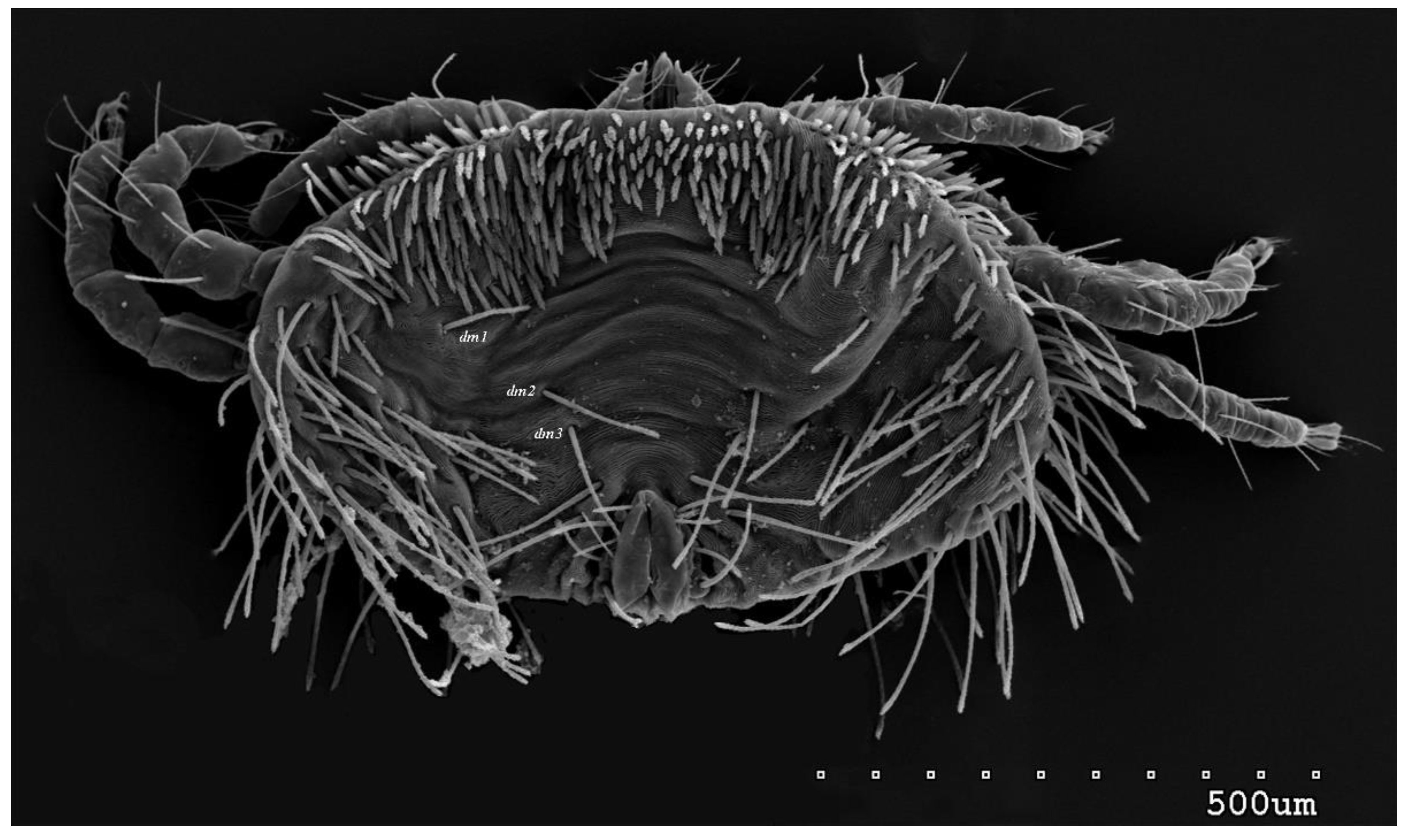
Figure 2.
Neopterygosoma robertmertensi sp. n., female details: (a) propodonotal shield (b) genital region.
Figure 2.
Neopterygosoma robertmertensi sp. n., female details: (a) propodonotal shield (b) genital region.

Figure 3.
Neopterygosoma robertmertensi sp. n., female in ventral view.
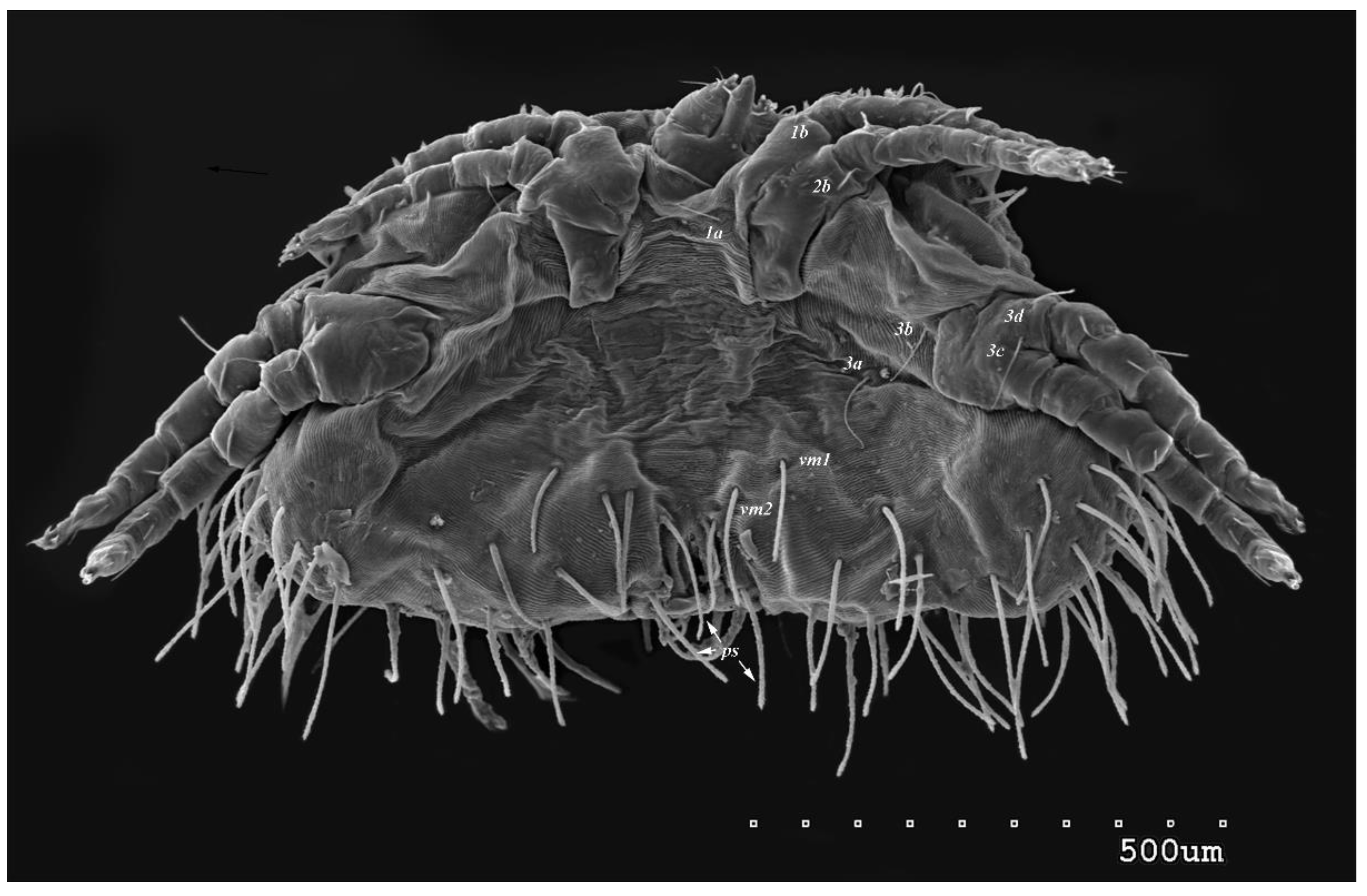
Figure 4.
Neopterygosoma robertmertensi sp. n., female details: (a) tarsi I in dorsal view; (b) palps in ventral view.
Figure 4.
Neopterygosoma robertmertensi sp. n., female details: (a) tarsi I in dorsal view; (b) palps in ventral view.
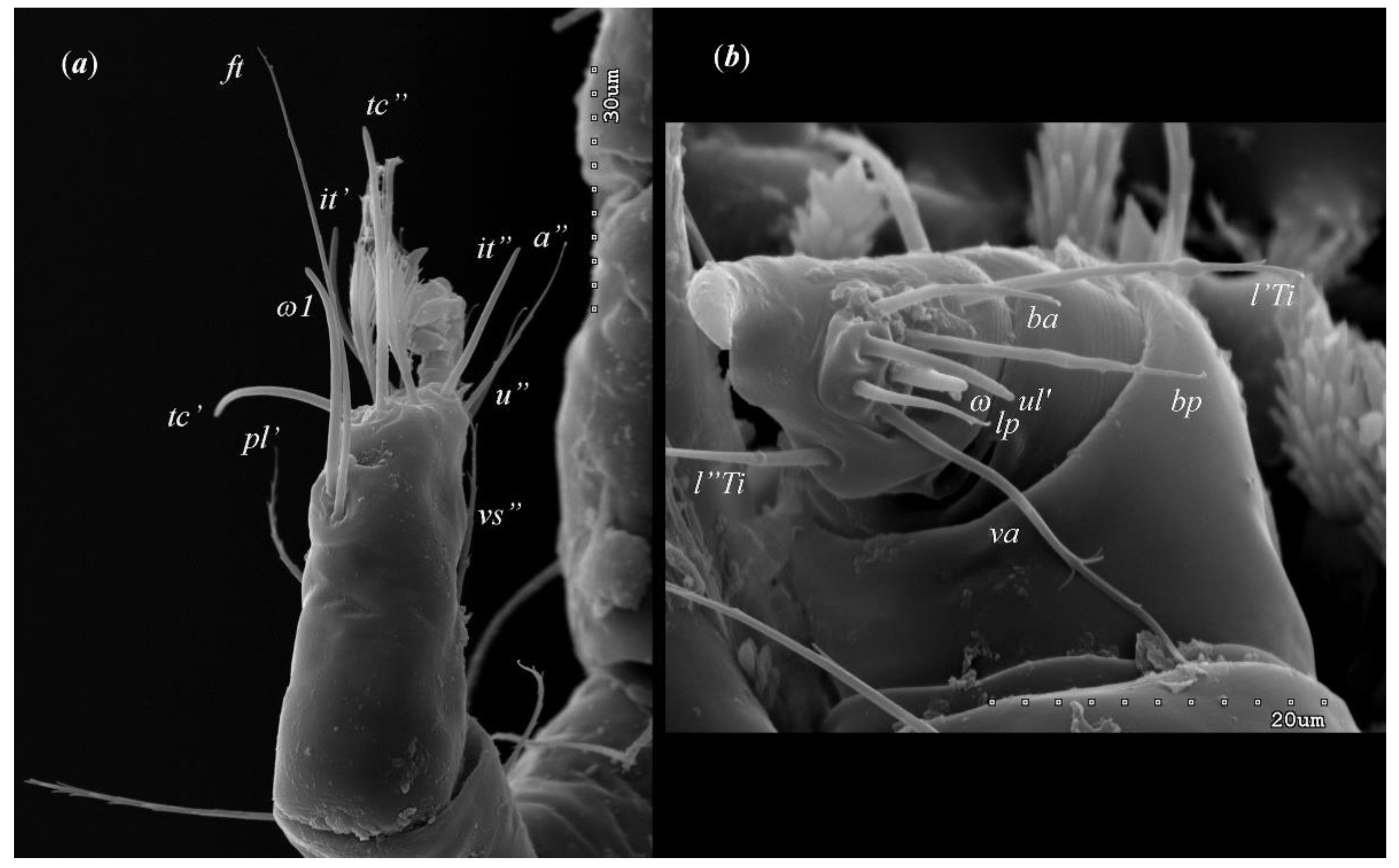
Figure 5.
Neopterygosoma robertmertensi sp. n., male in dorsal view.
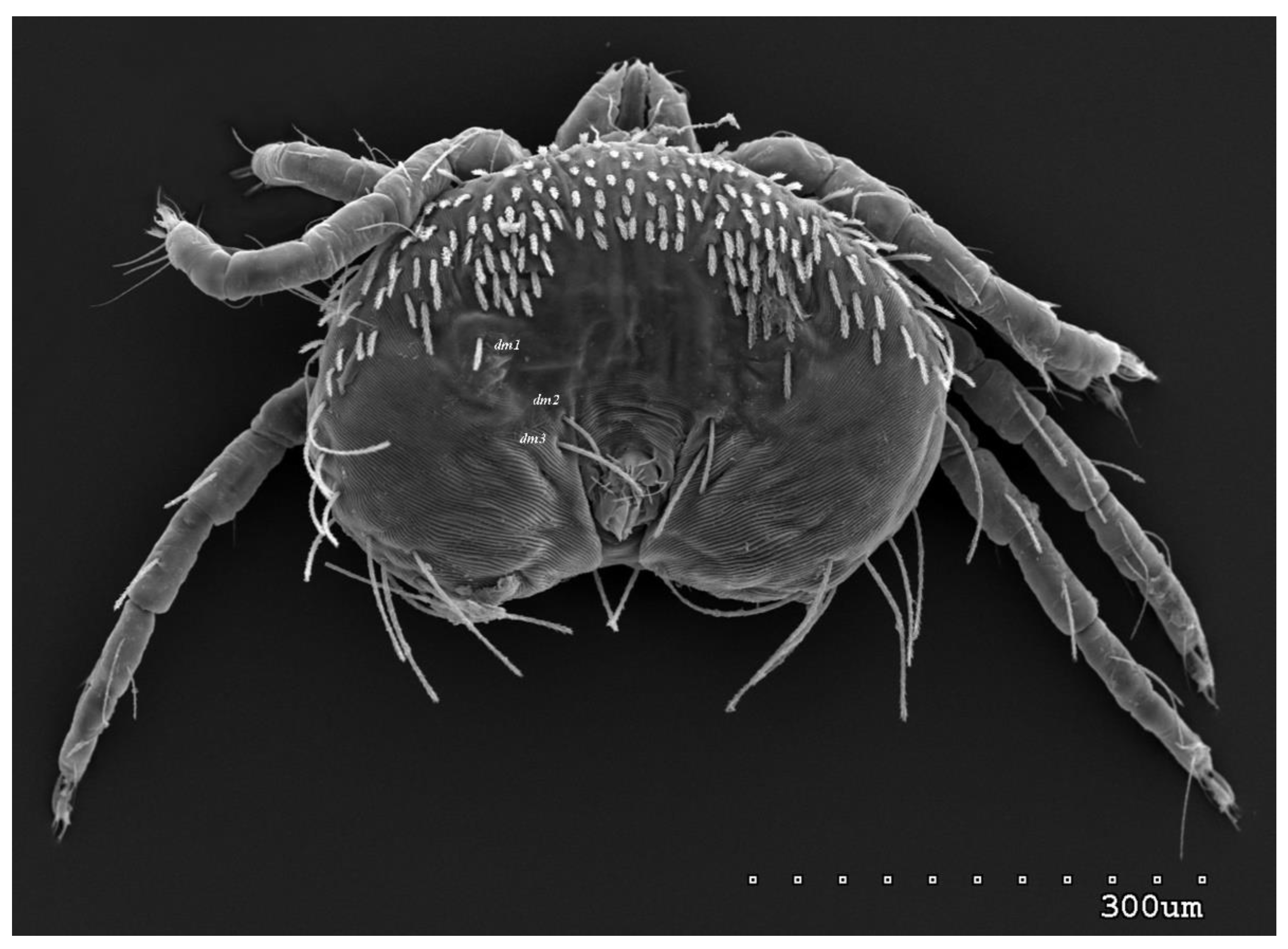
Figure 6.
Neopterygosoma robertmertensi sp. n., male in ventral view.

Figure 7.
Neopterygosoma robertmertensi sp. n., male, genital area, enlarged.

Figure 8.
Neopterygosoma robertmertensi sp. n. (a) imagochrysalis in the exoskeleton of deutonymph, dorsal view; (b) reduced gnathosoma, peritremes and coxae I–II of imagochrysalis, enlarged.
Figure 8.
Neopterygosoma robertmertensi sp. n. (a) imagochrysalis in the exoskeleton of deutonymph, dorsal view; (b) reduced gnathosoma, peritremes and coxae I–II of imagochrysalis, enlarged.
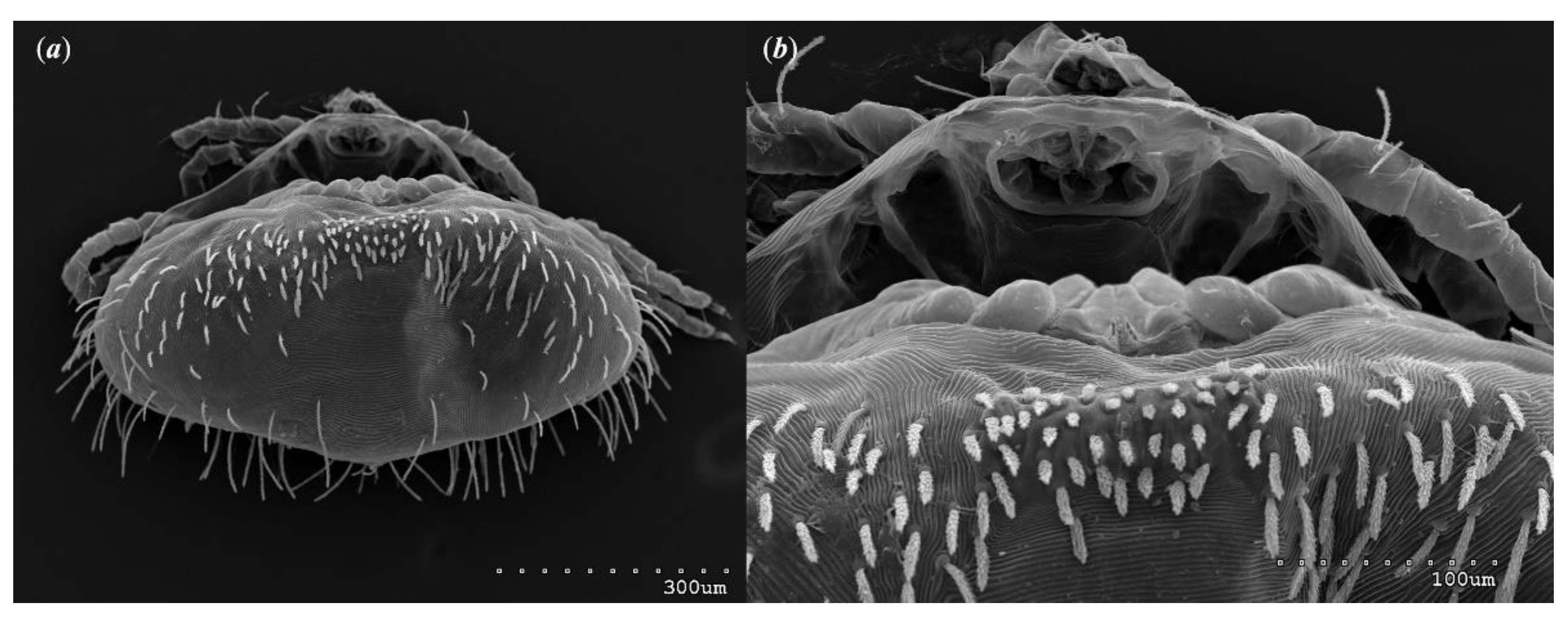
Figure 9.
Neopterygosoma robertmertensi sp. n., deutonymph in dorsal view.
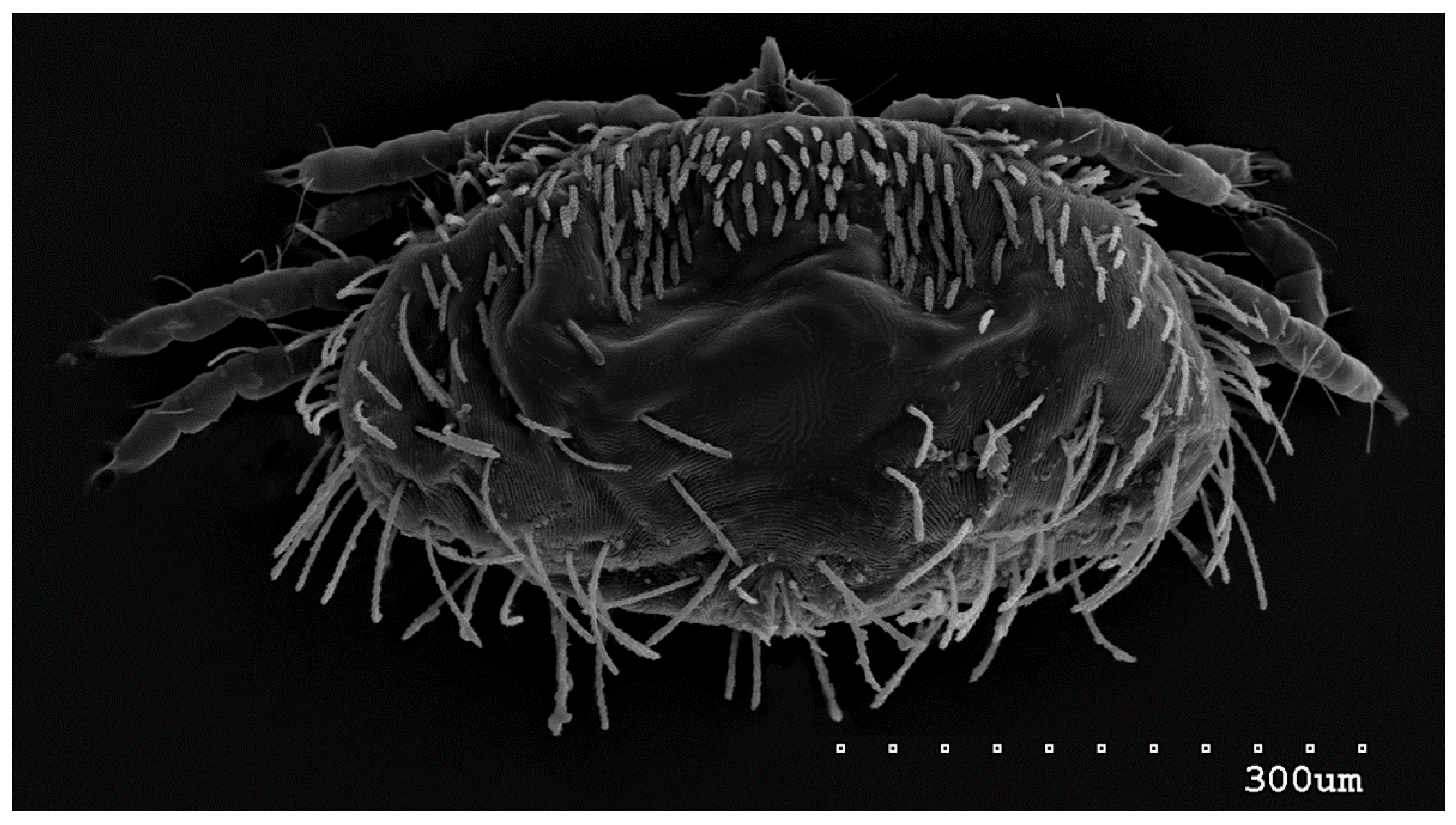
Figure 10.
Neopterygosoma robertmertensi sp. n., deutonymph in ventral view.
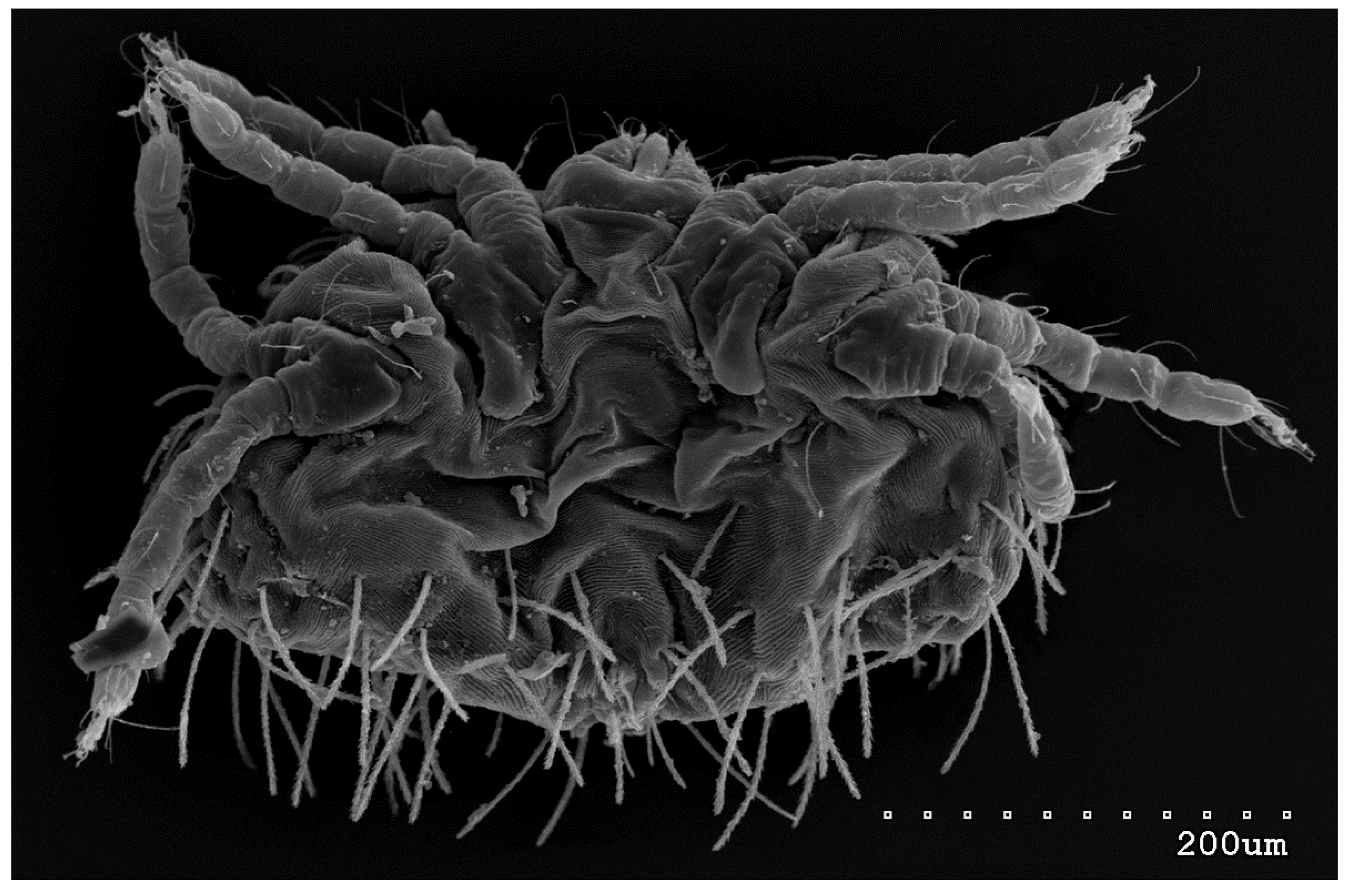
Figure 11.
Neopterygosoma robertmertensi sp. n., protonymph in dorsal view.
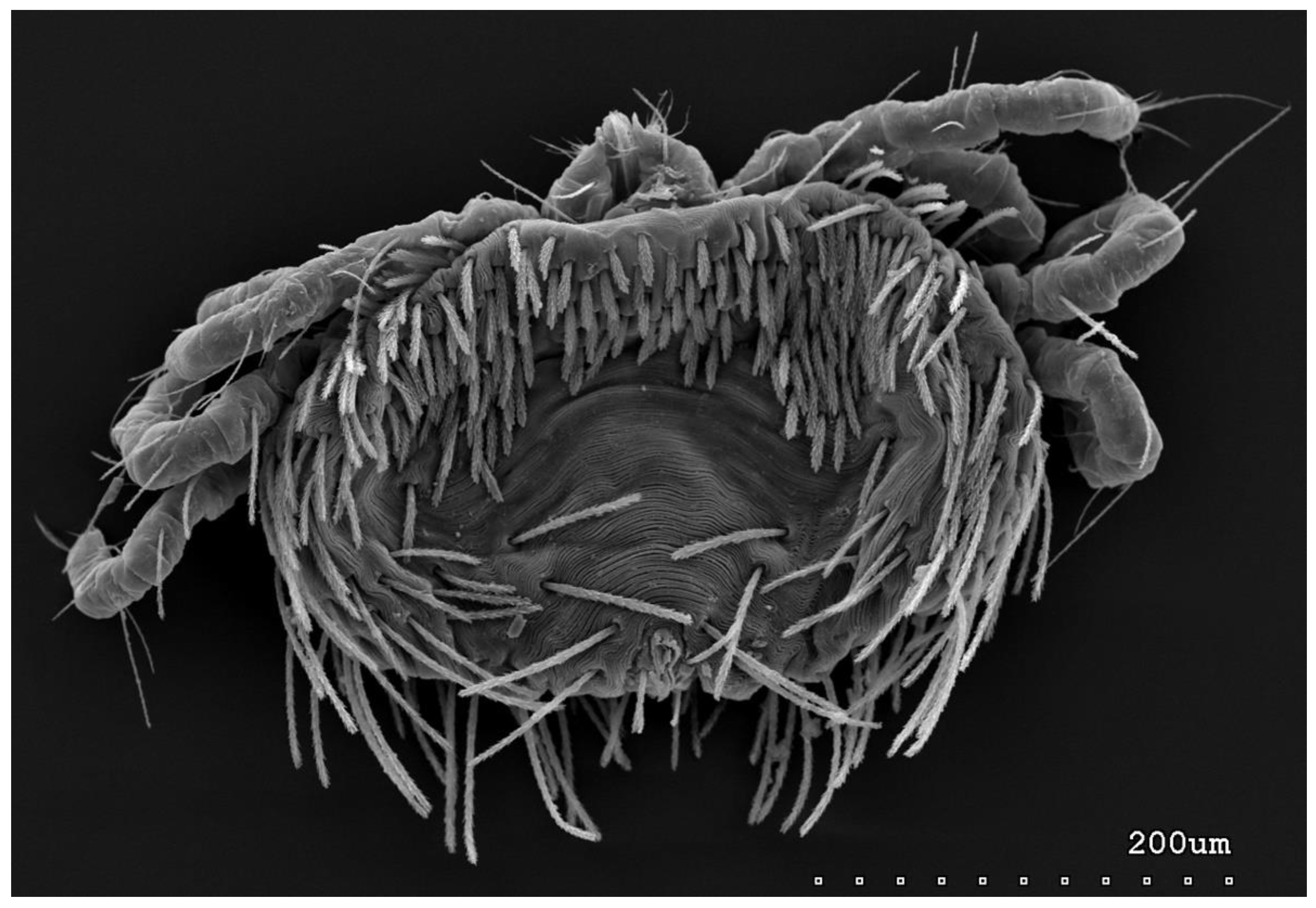
Figure 12.
Neopterygosoma robertmertensi sp. n., protonymph in ventral view.
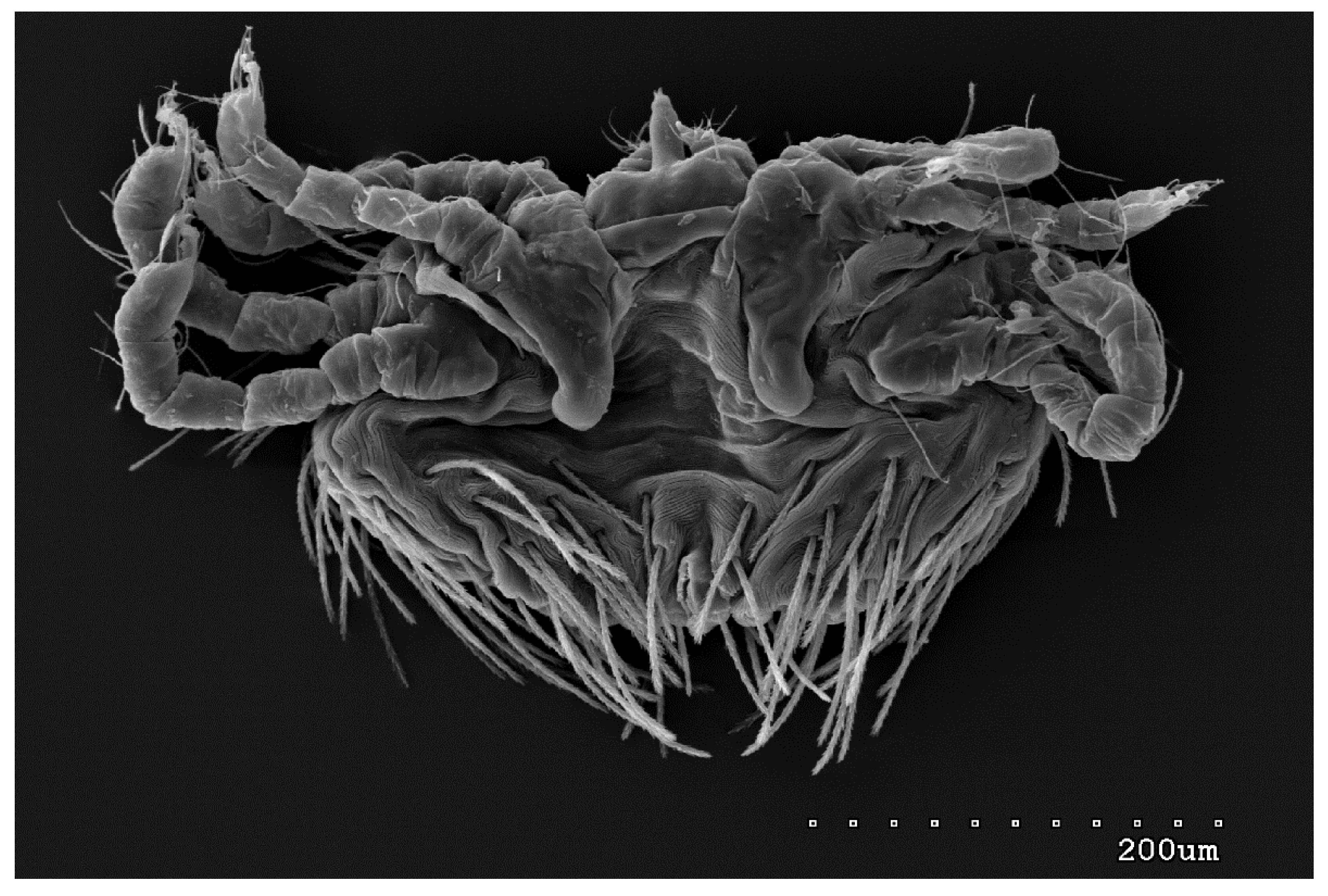
Figure 13.
Neopterygosoma robertmertensi sp. n. (a) female larva in dorsal view; (b) female larva in ventral view; (c) male larva in dorsal view; (d) male larva in ventral view.
Figure 13.
Neopterygosoma robertmertensi sp. n. (a) female larva in dorsal view; (b) female larva in ventral view; (c) male larva in dorsal view; (d) male larva in ventral view.

Figure 14.
Neopterygosoma robertmertensi sp. n. larva, details (a) dorsal setae; (b) part of gnathosoma in ventral view; (c) tarsi I in dorso-lateral view; (d) genital region.
Figure 14.
Neopterygosoma robertmertensi sp. n. larva, details (a) dorsal setae; (b) part of gnathosoma in ventral view; (c) tarsi I in dorso-lateral view; (d) genital region.

Figure 15.
The most parsimonious tree (tree length 219, CI of 0.64, RI of 0.56, RC of 0.36) found using the branch-and-bound search option for the unordered and unweighted dataset. Numbers at nodes are Bremer indices. Numbers below branches are common synapomorphies (character numbers refer to ListS1). Distribution of the mite species within host groups and section are marked in different colours.
Figure 15.
The most parsimonious tree (tree length 219, CI of 0.64, RI of 0.56, RC of 0.36) found using the branch-and-bound search option for the unordered and unweighted dataset. Numbers at nodes are Bremer indices. Numbers below branches are common synapomorphies (character numbers refer to ListS1). Distribution of the mite species within host groups and section are marked in different colours.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



