
Submitted:
10 July 2023
Posted:
12 July 2023
You are already at the latest version
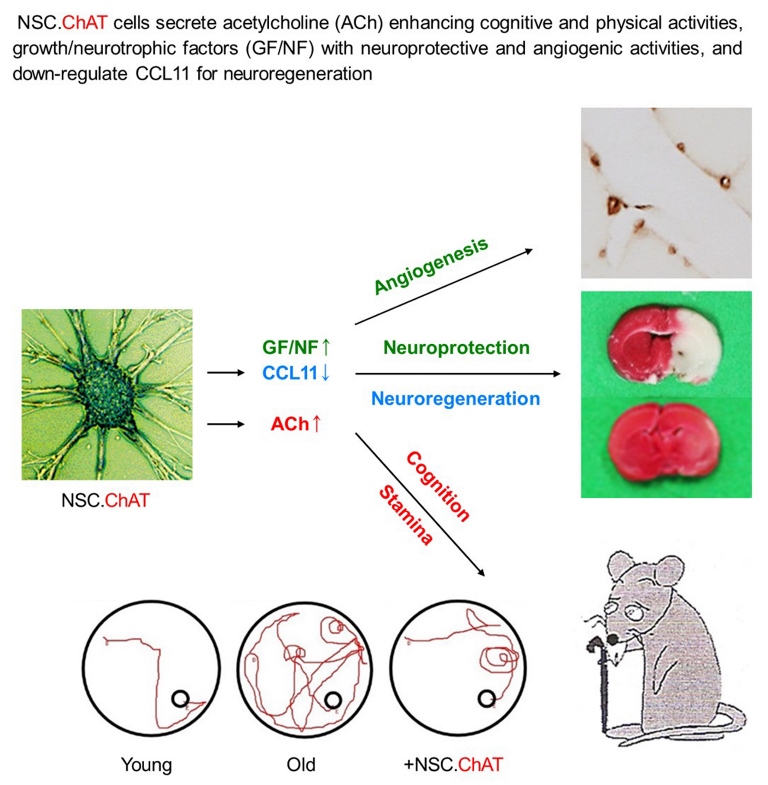
Abstract
Ageing may be characterized by progressive loss of cognitive function as well as decrease in physical activities, in addition to change in appearance. Previously, we reported that intracerebroventricular transplantation of a human neural stem cell (NSC) line (F3) over-expressing choline acetyltransferase (F3.ChAT) gene, an enzyme responsible for acetylcholine synthesis, markedly improved cognitive function and physical activity of aged animals by enhancing acetylcholine and growth factors (GFs) and neurotrophic factors (NFs). In the present study, 10-month-old F344 rats were intravenously transplanted with F3 or F3.ChAT NSCs (1 x 106 cells) once a month to 21st month of age when the cumulative mortality reached 20%, and physical activity and learning/memory function were investigated, in addition to the analyses of brain acetylcholine concentration, and cholinergic and dopaminergic system restoration. Neuroprotective and neuroregenerative activities of stem cells were also confirmed by analyzing oxidative damages, neuronal skeletal protein, angiogenesis, brain and muscle weights, and proliferating host stem cells. Stem cells markedly improved both the cognitive (passive avoidance and water-maze performances) and physical (locomotor and rota-rod activities) functions, in parallel with the elevation of acetylcholine levels in cerebrospinal fluid and in muscles, in which F3.ChAT cells were superior to F3 parental cells. Stem cell transplantation down-regulated CCL11, leading to restoration of microtubule-associated protein 2 as well as functional markers of cholinergic and dopaminergic systems, and preserved brain and muscular capillaries, recovering muscle mass. In addition, stem cells enhanced antioxidative capacity, attenuating oxidative damage of multiple organs including muscles and the brain. The results indicate that NSCs over-expressing ChAT gene improve cognitive function and physical activity of ageing animals by protecting and recovering functions of multiple organs including cholinergic and dopaminergic systems as well as muscles from oxidative injuries through secretion of acetylcholine and GFs/NFs, increased antioxidant elements, and enhanced blood flow.
Keywords:
ageing
; cognitive function
; physical activity
; human neural stem cell
; choline acetyltransferase
; acetylcholine
; growth factor
; neurotrophic factor
1. Introduction
As a natural process of ageing, various changes in the body appear. Brain ageing of animals results in precipitous decreases in neural stem cells (NSCs) and neurogenesis along with impairments of physical activity and cognitive function including learning and memory performances [1,2]. Brain stem cell population is declined during ageing process which may be due to decreased proliferation and differentiation. Accordingly, neuronal production may not enough for replenishment of cell death, and nerve cell network is changed insufficiently for proper signal transmission. These changes lead to cognitive impairment focusing on learning and memory functions [2,3].
It has been demonstrated that cholinergic system dysfunction leads to cognitive impairment. Concentration of acetylcholine (ACh), a major neurotransmitter responsible for memory acquisition, is diminished in aged animals as a result of cholinergic nerve degeneration [4,5,6]. ACh synthesis is regulated by choline transporter (ChT) which transports choline to presynaptic terminals wherein choline acetyltransferase (ChAT) catalyzes the production of ACh from choline and acetyl-CoA. Vesicular acetylcholine transporter (VaChT) delivers the synthesized ACh to synaptic vesicles. Therefore, degenerative changes in cholinergic nerves in aged persons and Alzheimer’s disease (AD) patients results in learning and memory loss following disintegration of the cholinergic factors including ChAT, ChT and VaChT [5,6,7,8,9].
On the other hand, various growth factors (GFs) and neurotrophic factors (NFs) including brain-derived neurotrophic factor (BDNF), nerve growth factor (NGF), and vascular endothelial growth factor (VEGF) plays roles in neuronal proliferation, protection, and regeneration, which decrease in aged, damaged, and diseased brains [10,11]. Thus, these alterations in GFs/NFs reduce neurogenesis and inactivate cholinergic nervous system, and thereby accelerate cognitive deficits. BDNF controls NSC survival and proliferation via tyrosine kinase receptor B (TrkB) signaling [12,13]. BDNF also relates to brain hippocampal synaptic plasticity and activates cholinergic nervous system by up-regulating ChAT expression [14]. Simultaneously, NGF facilitates neuronal proliferation and extends survival period of neurons in the hippocampus via TrkA [15]. In addition, NGF regulates innervation and nerve terminal signaling [16,17]. Notably, ageing affects angiogenesis in the brain and muscles. Reduced angiogenic activity in the aged brain is mostly related to the reduction of VEGF expression in response to degenerative change [18]. As a consequence, impaired VEGF production hinders neurogenesis, resulting in a decrease in neuronal population [18,19].
As another phenomenon due to ageing, physical activity is declined prominently. Aged animals show slow movement and physical inactivation compared to young animals. Such aggravations in physical functions are in part mediated by reduced dopaminergic regulation [20,21]. It is well known that physical activity is significantly reduced in Parkinson’s disease (PD) patients and PD model animals with dopaminergic system impairment. Dopaminergic nerve functions mainly via D2 receptors (D2R) distributed in the cortical region, and cortex is also major site for motor neuron networking [22]. Moreover, cholinergic system degeneration by ageing lowers dopaminergic activity via its regulation procedures. In was confirmed that cholinergic nerve impairment accompanying decreased ACh synthesis negatively affects dopaminergic system, diminishing physical activity and movement [23]. Among neurotrophins, glial cell-derived neurotrophic factor (GDNF) controls dopaminergic nerves by raising monoamine production through TrkB receptors. Collectively, physical activity is mainly controlled by the regulation of dopaminergic and cholinergic nervous systems [24].
Besides of the control by central nervous system (CNS), physical stamina depends on the muscular strength and endurance to fatigue. In aged rats, muscle mass and strength were declined along with a decreased distribution of blood vessels in the muscles [25]. Notably, BDNF and insulin-like growth factor-1 (IGF-1) are engaged in muscle growth and myofiber differentiation [25,26]. GDNF regulates nerve growth and neuromuscular junction innervation, and VEGF enhances muscular angiogenesis for sufficient glucose and oxygen supply [27,28,29]. During muscular degeneration and ageing, it was found that GFs and NGs including BDNF, IGF-1, VEGF, and GDNF were diminished [27].
Although cognitive function and physical activity are regulated by nervous system and muscular integrity, such regulatory systems are impaired following tissue injuries from oxidative stress, a typical result in ageing process. By oxidative stress, damaged mitochondrial DNA induces neuronal degeneration with glial cell proliferation [30]. Interestingly, GFs/NFs, including BDNF, NGF, VEGF, and IGF possessing antioxidative activities, decreased in the aged brain and muscles [31,32], indicative of the multiple roles of GFs/NFs in protecting tissues and organs related to the antioxidative defense system. Even though ageing includes various deteriorations by multiple reasons, most representative ageing signs might be focused on cognitive dysfunction, physical inactivation, and muscle degeneration.
Although there are many research reports on the palliative improvement of cognitive and physical functions using small molecule drugs and natural products, results showing long-term salient effects of stem cells are limited. Previously, we established human neural stem cells (NSCs) encoding ChAT gene (named F3.ChAT cells), an enzyme responsible for ACh synthesis, by inserting ChAT gene into F3 NSC line, and demonstrated that the F3.ChAT cells increased brain ACh level, and thereby fully restored the learning and memory functions of Alzheimer’s disease (AD) model animals [33,34].
In addition, single intracerebroventricular (ICV) transplantation of F3.ChAT NSCs markedly recovered both the cognitive function and physical activity of aged mice by restoring the levels of ACh and various GFs/NFs [35]. Based on the previous results, anti-ageing effects of F3 and F3.ChAT NSCs focusing on cognitive dysfunction and physical inactivation as well as underlying action mechanisms were investigated in naturally-ageing rats following repeated life-long intravenous (IV) injections from 10 months of age.
2. Materials and Methods
2.1. Human NSC Line
An immortalized human NSC lines, HB1.F3 (named F3) and F3.ChAT [33,34,36,37], from the University of British Columbia, BC, Canada (provided by Prof. SU Kim), were cultivated in Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin (Invitrogen, Carlsbad, CA, USA). Cultures were maintained under 5% CO2 at 37°C in tissue culture flasks. Media were changed every 2–3 days.
2.2. Ageing Animals and NSC Transplantation
Ten-month-old Fisher 344 (F344) rats were purchased from Daehan-Biolink (Eumseong, Korea). The animals (n = 20/group) were maintained at a constant temperature (23 ± 2°C), relative humidity of 55 ± 10%, and 12-hour light/dark cycle, and fed standard rodent chow and purified water ad libitum. Animals were transplanted with NSCs (1 × 106 cells/rat) in 100 μL saline via intravenous route. The rats received the cells once a month repeatedly for 11 months (from 11 to 21 months of age).
2.3. Measurement of Physical Activity
2.3.1. Locomotor Activity
Spontaneous activities and exploratory behaviors were evaluated using a video tracking system (Smart v2.5; Panlab Technology, Barcelona, Spain), connected to a CCTV monitor (Samsung, Changwon, Korea) after a week of cell transplantation. Rats were placed in a quiet chamber with a dim light. The types of movement, i.e., resting, slow-moving and fast-moving times, were recorded for 5 min, and the ratio was analyzed.
2.3.2. Rota-rod Performance
Motor balance and coordination were evaluated using a rota-rod test system (Panlab Technology) after 1 week of stem cell transplantation. Rats were placed on a rotating rod at a constant speed of 12 rpm, and the time for the rats to fall off the rod was recorded. The average latency was calculated from 3 consecutive measurements.
2.4. Measurement of Learning and Memory Functions
2.4.1. Passive Avoidance Performance
Passive avoidance performance was assessed by Shuttle box (ENV-010MD; Med Associates Inc., St. Albans, VT, USA) to evaluate memory acquisition and retention when the survival rate of animals was 80% (21 months old). The Shuttle box apparatus consists of light and dark compartments: i.e., light chamber equipped with a lamp and dark chamber with a steel-grid floor for electric shock. On the trials, electric shock was delivered when animals entered the dark compartment from the light room through a guillotine door. The latency time of staying in a room with the light on was recorded following electric shock (1 mA for 2 sec) in a dark compartment. Seven consecutive trials, once a day, were performed with electric shock when animals entered the dark compartment. The end-point was set to 300 sec, denoting full acquisition of memory.
2.4.2. Water-maze Performance
Morris water-maze performance was assessed in a round water bath (180 cm in diameter) filled with water (27 cm in depth) maintained at 22 ± 2°C to evaluate spatial memory. The bath was divided into four quadrants, and a hidden escape platform (10 cm in diameter, 25 cm in height) was submerged in the center of one quadrant, 2 cm below the surface of water. The animals were subjected to 7 trials once a day to find the platform hidden by white styrofoam granules (5 cm in diameter) on the surface of water, based on several cues external to the maze. The position of the cues was maintained unchanged throughout the experiments. Escape latency time, which were spent to escape onto the platform during trials, was recorded. The end-point was set to 300 sec.
2.5. Analysis of ACh Concentration
The animals were sacrificed 24 hours after the final cognitive function test. Cerebrospinal fluid (CSF) was collected, and gastrocnemius muscles were obtained and homogenized to analyze ACh concentration. ACh concentrations in CSF and muscle homogenate were measured with an Amplex Red acetylcholine/acetylcholinesterase assay kit (Molecular Probes, Eugene, OR, USA) according to the manufacturer’s instructions. In this assay, ACh is hydrolyzed by AChE to release choline, which is then oxidized by choline oxidase to betaine and H2O2. H2O2 interacts with Amplex Red (7-dihydroxyphenoxazine) in the presence of horse radish peroxidase to produce highly fluorescent resorufin. The resulting fluorescence was measured in a fluorescence microplate reader using excitation in the range of 530–560 nm and emission at 590 nm.
2.6. Stem Cell Distribution
The brains of animals were perfusion-fixed with 10% paraformaldehyde solution and post-fixed in the same fixation solution for 2 days, followed by cryoprotection in 30% sucrose for 3 days to confirm the distribution of transplanted F3 and F3.ChAT cells. Coronal cryosections of 30-μm thickness were prepared and processed for immunostaining for human mitochondria (hMito) using an antibody specific for hMito (1:200; mouse monoclonal, Chemicon, Temecula, CA, USA). In order to identify proliferating neural stem cells, brain sections were double immunostained with antibodies of nestin (1:200; mouse polyclonal, Chemicon) or Ki-67 (1:200; rabbit polyclonal, Chemicon). Brain sections were incubated with primary antibodies overnight at 4°C and with secondary antibodies conjugated with Alexa Fluor-488 or -594 (1:500; rabbit polyclonal, Molecular Probes) for 2 hours at room temperature. All samples were examined immediately after staining and photographed with a laser-scanning confocal microscope (LSM710; Zeiss, Oberkochen, Germany).
2.7. RT-PCR Analysis
Total RNA was extracted from the brain tissue using TRIzol (Invitrogen). Complimentary DNA templates were prepared from 1 μg of total RNA primed with oligodT primers using 40 U of Moloney Murine Leukemia Virus reverse transcriptase (Promega, Madison, WI, USA) followed by 40 PCR cycles. RT-PCR products were separated electrophoretically on an 1.2% agarose gel containing ethidium bromide. The primers used for RT-PCR (ChAT, VAChT, ChT1, m1-AChR, nAChR α5, nAChR 2β, AChE, TH, VMAT2, DAT, D1R, D2R, CCL11, and GAPDH) were described in Supplementary Table S1 (Bioneer, Daejeon, Korea).
2.8. Western Blot Analysis
Whole brains and gastrocnemius muscles of animals were homogenized in RIPA buffer (Sigma-Aldrich, St. Louis, MO, USA). Proteins were obtained by centrifugation at 15,000 rpm at 4°C for 15 min and quantified using the BCA Protein Assay kit (Pierce, Rockford, IL, USA). Proteins were denatured by heating for 5 min at 95°C in 0.5 M Tris-HCl buffer (pH 6.8) containing 10% sodium dodecyl sulfate (SDS) and 10% ammonium persulfate, separated by electrophoresis on 7.5% SDS-polyacrylamide gels, and transferred to a polyvinylidene difluoride membrane in 25 mM Tris buffer containing 15% methanol, 1% SDS, and 192 mM glycine. After blocking for 2 hours with 5% skim milk in Tris-buffered saline-Tween (TBS-T; 20 mM Tris pH 7.6, 137 mM NaCl, and 0.1% Tween 20), the membrane was incubated with antibodies specific for microtubule-associated protein 2 (MAP2) (1:500; rabbit polyclonal, Santa Cruz Biotechnology, Santa Cruz, CA, USA) overnight at 4°C. After washing with TBS-T, the membrane was incubated with a secondary goat anti-rabbit IgG conjugated with horseradish peroxidase (1:2,000; Santa Cruz Biotechnology) for 2 hours at room temperature. The membrane was then developed using an enhanced chemiluminescence solution (Pierce, Rockford, IL, USA).
2.9. Identification of Capillary
Microvessel density was analyzed by immunohistochemical staining for von Willebrand Factor (vWF) to identify endothelial cells of blood vessels. The brain and gastrocnemius muscles were fixed in 10% neutral formalin, and paraffin-embedded sections were pretreated with citrate buffer (pH 6.0), blocked with serum, and incubated with primary antibody specific for vWF (1:200; rabbit polyclonal, Chemicon) overnight at 4°C. The sections were incubated with biotinylated secondary antibody for 1 hour at room temperature, followed by avidin-biotin complex kit (Vector Laboratory, Burlingame, CA, USA), and developed with diaminobenzidine (Sigma-Aldrich). Sections were observed under the field of x200 of a light microscope.
2.10. Antioxidative Activity of Stem Cells
To assess oxidative tissue injury, brain and gastrocnemius muscles were homogenized in 9 volumes of 10 mM sodium phosphate-buffered saline (PBS, pH 7.4) to make 10% homogenate at 4°C. An aliquot (500 μL) of the homogenate was acidified by adding 500 μL sodium dodecyl sulphate (SDS, 8.1% solution) and 1 mL of 20% acetic acid (adjusted to pH 3.5). After adding 500 μL of 2-thiobarbituric acid (TBA, 0.75% solution), the mixture was boiled in a glass tube capped for 30 min at 95°C. Samples were cooled on ice, centrifuged at 13,000 g for 10 min, and absorbance of the supernatant was read at 532 nm for the quantification of thiobarbituric acid-reactive substances (TBARS).
2.11. Statistical Analysis
Data are presented as mean ± SEM. The statistical significance between group comparisons for behavioral data was determined by one-way analysis of variance (ANOVA), followed by post-hoc Tukey's multiple comparison test. P-values<0.05 were considered to be statistically significant.
3. Results
3.1. Improvement of Physical Activity
Old animals (11-20 months old) exhibited severely decreased movement as measured by locomotor activity (Figure 1A). The resting time of aged rats markedly increased to 65-80%, while young animals displayed 70-95% moving (slow- and fast-moving) activity. However, transplantation of F3 or F3.ChAT cells significantly improved the activity of old animals, showing increased slow-moving time, up to 20 months of age when 20% of the rats died, although fast-moving time increased at 11 months following F3.ChAT treatment.
The latency time in rota-rod performance of aged animals also markedly decreased to 10-40% of that of young animals, and especially, the impairment of performance further increased according to the progress of ageing (Figure 1B). Notably, repeated transplantation of F3 or F3.ChAT cells greatly improved the motor coordination of old rats, in which F3.ChAT was more effective than F3 cells.
3.2. Recovery of Cognitive Function and ACh Concentration
In addition to the loss of physical activity, 21-month-old rats showed severe impairment of cognitive function (Figure 2A,B). The aged rats showed a delayed increase in the latency time in spite of repeated trials in the passive avoidance performance, by comparison with the full memory acquisition at the 5th trial in young animals (Figure 2A). In water-maze performance (Figure 2B), the old animals did not show memory-acquiring capacity during 7 repeated trials, in contrast to the rapid decrease in the escape latency in young animals. However, repeated transplantation of F3 cells significantly improved the cognitive function in both performances. Notably, higher cognition-enhancing effects were achieved with transplantation of F3.ChAT cells, than F3 cells, in both passive avoidance and water-maze performances.
The ACh concentration in CSF (4.3 μM) of aged (21-month-old) rats was much lower than that of young (7-week-old) animals (7.3 μM) (Figure 2C). Such decreased ACh level in CSF was significantly restored to 5.1 and 6.1 μM following transplantation of F3 and F3.ChAT cells, respectively.
Muscular ACh concentration in old rats (1.6 μmole/g tissue) was significantly lower than that of young animals (2.4 μmole/g tissue), too (Figure 2D). Muscular ACh level was also recovered to 2.0 and 2.5 μmole/g tissue by transplantation of F3 and F3.ChAT cells, respectively.
3.4. Distribution of Transplanted Cells
hMito-immunoreactivity was detected in the brain after 12-times transplantation of F3 cells (1 × 106 cells/rat) into 10-month-old animals (Figure 3A). The transplanted cells distributed predominantly in the hippocampus (89.1 cells/mm2), although a part of the cells were found in the cortex (34.2 cells/mm2) (Figure 3B). Transplanted F3.ChAT cells were also detected in the hippocampus (103.7 cells/mm2) and cortex (48.0 cells/mm2) of rats (Figure 3A,C), indicative of higher penetration of F3.ChAT cells into the brain than F3 cells.
3.5. Cholinergic and Dopaminergic Activation in the Host Brain
Expressions of functional genes of cholinergic system were markedly changed according to ageing. That is, genes for synthesis, secretion, and reception of ACh including ChAT, VAChT, ChT1, m1-AChR, nAChR α5, and nAChR β2 decreased, while gene of AChE, an ACh-degrading enzyme, increased in aged animals (Figure 4A). Such alterations in the gene expression of cholinergic markers were remarkably restored after transplantation of F3 or F3.ChAT cells, in which F3. ChAT cells were superior to F3 cells.
Expressions of genes of dopaminergic system such as TH, VMAT2, DAT, D1R, and D2R also decreased in aged animals. However, the decreased expression of genes associated with dopamine synthesis, transportation, and reception were markedly recovered following transplantation of F3 or F3.ChAT cells (Figure 4B).
3.6. Angiogenic Effects
In old rats, the vWF-positive microvessel density in the brain was reduced to 27.8% of young animals (Figure 5A,B), and the relative brain weight decreased meaning brain atrophy (Figure 5C). Interestingly, transplantation of F3 or F3.ChAT cells led to increase in the number of blood vessels, indicative of preservation or neovascularization (Figure 5B), in which F3.ChAT was much more effective than F3 cells. Such effects may be related to the attenuated brain atrophy (Figure 5C).
In gastrocnemius muscles of aged animals, the vessel density also decreased to 66.7% of young animals (Figure 5D,E), resulting in the significant decrease in muscle weight. Transplantation of F3 or F3.ChAT cells increased the number of capillaries and the muscle mass, although the effects were more prominent in F3.ChAT cells than F3 cells (Figure 5E,F).
3.8. Antioxidative Effects
The concentration of TBARS, by-products of lipid peroxidation, significantly increased in the brain and muscles of old rats compared with that of young animals (Figure 6). Such increased oxidative damage was near-fully attenuated by transplantation of F3 or F3.ChAT cells.
3.9. Neuroprotection and Regeneration
The number of host neural (nestin-positive) stem cells in old rats markedly reduced to 26.9% of young animals (Figure 7A,B). Most of the nestin-positive cells also exhibited immunoreactivity to Ki-67, a proliferating cell marker. Interestingly, transplantation of F3 and F3.ChAT cells significantly increased the number of host proliferating (Ki-67-positive) cells, in which F3.ChAT cells were superior to F3 cells (Figure 7B).
On the other hand, the expression of CCL11, an inhibitor of neurogenesis, increased during ageing, in parallel with the marked decrease in the content of MAP2, a neuronal skeletal protein (Figure 7C). The changes in CCL11 expression and MAP2 production were fully reversed following transplantation of F3 or F3.ChAT cells.
4. Discussion
In this research, it was confirmed that F3 and F3.ChAT NSCs possess anti-ageing effects against spontaneous systemic ageing. As a result of anti-ageing effect of transplanted cells, cognitive function was improved compared to aged animals, in which both passive avoidance and water-maze performances were markedly enhanced by F3 NSCs. Interestingly, the cognition-enhancing effect of F3.ChAT cells were much higher than that of F3 parental cells, exerting full memory acquisition within 5-7th trials comparable to young animals.
It is well known that acquisition of memory is mainly regulated by ACh of cholinergic nervous system. According to the degenerative change of cholinergic system during ageing, learning and memory functions are gradually inactivated [4,38]. In PCR analysis, the expressions of genes for ACh synthesis and release decreased in aged animals, whereas AChE, an ACh-degrading enzyme increased. However, F3 and F3.ChAT NSCs reversed the changes in the gene expression related to both ACh synthesis and degradation, normalizing cholinergic function, as confirmed by recovery of ACh concentration. In addition, the gene expression of muscarinic and nicotinic receptors of ACh, responsible for memory acquisition, was recovered similarly. Notably, such effects on the reactivation of cholinergic system, i.e., gene expression of cholinergic system markers as well as ACh concentration, were outstanding in the rats transplanted with F3.ChAT cells. In previous reports, F3.ChAT cells were also superior to their parental F3 cells, in the recovery of cognitive function of AD model rats and aged mice, indicative of the major role of ChAT gene [33,34,35].
With immunohistochemical analysis of the brain, it was confirmed that many NSCs entered the ageing brain after repeated IV injection. Transplanted cells were found predominantly in the hippocampus and cortex. Increased permeability of the blood-brain barrier (BBB) with ageing which might be mediated by oxidative damage has been confirmed [39]. Thus, it is assumed that intravenously transplanted cells migrated into the aged brain, and contributed the increased ACh concentration in CSF, as also confirmed in AD model and ageing animals [33,34,35]. Muscarinic and nicotinic receptors are widely distributed in the hippocampus and cortex, and play key roles in spatial and working memories [40,41]. ChAT is also one of the well-known markers of motor neurons [42]. Repeated transplantation of NSCs markedly recovered the decreased levels of MAP2 and cholinergic and dopaminergic nervous system markers in aged rats, indicative of a structural restoration of brain integrity. The improvements of cognitive and motor functions by F3.ChAT cell transplantation might be due to the increased ACh levels in the brain and muscles originated not only from the transplanted stem cells, but also from restored host cholinergic neurons. In addition, the restored dopaminergic nervous system might led to additional improvement of physical activity.
Furthermore, neurogenesis plays a key role for the retention of learning and memory. Neurogenesis is promoted by neural progenitor cells’ maturation, and cognitive function is improved by increase in neural cell population [38,43,44]. It was found that declined cognitive function by ageing was recovered with neurogenesis promotion near the hippocampus which is the main region associated to cognitive function. Previous researches demonstrated that NSC transplantation enhanced learning and memory function in aged animals [45]. In the present study, F3 and F3.ChAT NSCs also promoted neurogenesis and inhibited neuronal degeneration: i.e., stem cell transplantation induced proliferation of host nestin-positive cells mainly in CA1 and CA2 regions of hippocampus as confirmed by immunoreaction to Ki-67, wherein F3.ChAT cells were more effective than F3 parental cells. Such a neurogenic effect of stem cells might be mediated by GFs/NFs such as BDNF and NGF [46,47]. It is well known that BDNF and NGF promote neurogenesis, proliferation, and differentiation into neurons in the hippocampus [12,13]. In our previous reports, it was well confirmed that F3.ChAT cells expressed and produced diverse neurotrophins [48], and transplantation of the cells restored GFs and NFs [35,47,48,49]. Therefore, it is suggested that the GFs/NFs might contributed the neurogenesis of the ageing animals.
Brain angiogenesis is an important factor in neurogenesis. It was reported that in stroke or ischemic injury, angiogenesis was promoted which might be mediated by GFs including VEGF [50,51]. Interestingly, the microvessel density decreased in aged rat brain was remarkably restored by F3.ChAT cell transplantation. F3.ChAT cells highly expressed VEGF and increased the capillary density in middle cerebral artery occlusion (MCAO) stroke model animals [48]. Accordingly, the attenuation of brain atrophy and restoration of muscle mass may be mediated in part by the microvessel preservation and possible neovascularization. Notably, it was found that the expression of CCL11, a chemokine inhibiting neuroregeneration, significantly increased in aged animals [52]. However, the CCL11 expression was fully suppressed by transplantation of F3 or F3.ChAT cells. As a consequence of such neurogenic effects of F3 and F3.ChAT cells, it is believed that host neural (nestin-positive) stem cells underwent proliferation (Ki-67-positive), and neuronal skeletal protein MAP2 was recovered following NSCs transplantation [48,49,53].
In addition to the enhancement of ACh synthesis by cholinergic system activation, dopaminergic system was also activated by F3 and F3.ChAT NSCs. That is, F3 or F3.ChAT cell transplantation recovered the expression of several genes related to dopamine synthesis and release including TH, VMAT2, and DAT. Ideally, expression of dopaminergic receptors such as dopamine receptors 1 and 2 were recovered. Such effects were higher in F3.ChAT-transplanted rats brain than in F3-treated animals. It was reported that dopaminergic activation was regulated by cholinergic nervous system with secretion of NFs such as GDNF [54], and that dopaminergic activation enhanced physical activity and extended moving distance in F344 rats [20,55]. Considering the high expression of GDNF in F3.ChAT cells [48], it is explained that the dopaminergic system was stimulated via cholinergic activation and recovered GDNF expression by F3 or F3.ChAT cell transplantation. As results of dopaminergic activation, physical activities (moving times in locomotor activity and rota-rod performance) increased following F3.ChAT cell transplantation. In comparison, F3 cells also improved physical activity, although the effect did not reach that of F3.ChAT cells.
Related to muscular differentiation and innervation, physical activities are regulated by GFs/NFs. In particular, IGFs facilitate muscle growth and myofiber differentiation via PI3-Akt signaling [56,57]. BDNF and GDNF enhance muscle innervation and synaptic plasticity [58,59]. BDNF also promotes axonal sprout and myofiber differentiation via TrkB signaling. Moreover, GDNF controls muscle innervation and ACh receptor expression in neuromuscular junction. As observed our study demonstrating a major role of BDNF in the stamina-enhancing effect of adipose-derived stem cells (ADSCs) [60], GFs and NFs increased by stem cell transplantation might led to the muscular innervation and activation of cholinergic and dopaminergic nerves. Notably, F3.ChAT transplantation recovered the decreased microvessel density in muscles of aged rats, leading to an increase in muscle mass as well as attenuation of brain atrophy. Moreover, F3 and F3.ChAT cells protected against oxidative injury of muscles and brain tissues. Therefore, it is explained that the increased physical activity of aged rats transplanted with stem cells resulted not only from enhanced cholinergic and dopaminergic nervous control, but also from improved muscular integrity.
Taken together, it is believed that the anti-ageing effects of F3 and F3.ChAT cells were mediated by combinational activities on ACh production, neuroprotection and neuroregeneration, cholinergic and dopaminergic system activation, antioxidation and angiogenesis, and muscular regeneration accompanying GFs/NFs secretion. The anti-ageing effects obtained with F3.ChAT cells were superior to those of F3 cells, especially as observed in the improvement of learning/memory performance and moving behavior. Therefore, it is suggested that human NSCs over-expressing ChAT gene could be a promising candidate to delay ageing process characterized as cognitive dysfunction and physical activity impairments.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Y.-B.K.; investigation, J.K., D.K. and D.P.; analysis, K.S.; data curation, S.-C.H. and T.M.K.; writing—original draft preparation, E.-K.C. and Y.-B.K.; writing—review and editing, Y.-B.K.; supervision, T.M.K.; funding acquisition, S.-C.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by “Regional Innovation Strategy (RIS)” through the National Research Foundation of Korea (NRF), funded by the Ministry of Education (MOE) (2021RIS-001) and a grant of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI) funded by the Ministry of Health and Welfare, Republic of Korea (grant number: HI22C1650).
Institutional Review Board Statement
All animal experimental procedures were approved and carried out in accordance with the Institutional Animal Care and Use Committee of Laboratory Animal Research Center at Chungbuk National University, Republic of Korea (Approval No. CBNUA-752-14-01).
Informed Consent Statement
Not applicable.
Acknowledgments
This work was conducted during the research year of Chungbuk National University in 2022.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vanguilder, H.D.; Bixler, G.V.; Sonntag, W.E.; Freeman, W.M. Hippocampal expression of myelin-associated inhibitors is induced with age-related cognitive decline and correlates with deficits of spatial learning and memory. J. Neurochem. 2012, 121, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Sahay, A.; Scobie, K.N.; Hill, A.S.; O'Carroll, C.M.; Kheirbek, M.A.; Burghardt, N.S.; Fenton, A.A.; Dranovsky, A.; Hen, R. Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature 2011, 472, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Artegiani, B.; Calegari, F. Age-related cognitive decline: Can neural stem cells help us? Aging 2012, 4, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Itou, Y.; Nochi, R.; Kuribayashi, H.; Saito, Y.; Hisatsune, T. Cholinergic activation of hippocampal neural stem cells in aged dentate gyrus. Hippocampus 2011, 21, 446–459. [Google Scholar] [CrossRef]
- Abdulla, F.A.; Abu-Bakra, M.A.; Calaminici, M.R.; Stephenson, J.D.; Sinden, J.D. Importance of forebrain cholinergic and GABAergic systems to the age-related deficits in water maze performance of rats. Neurobiol. Aging 1995, 16, 41–52. [Google Scholar] [CrossRef]
- Watanabe, T.; Yamagata, N.; Takasaki, K.; Sano, K.; Hayakawa, K.; Katsurabayashi, S.; Egashira, N.; Mishima, K.; Iwasaki, K.; Fujiwara, M. Decreased acetylcholine release is correlated to memory impairment in the Tg2576 transgenic mouse model of Alzheimer's disease. Brain Res. 2009, 1249, 222–228. [Google Scholar] [CrossRef]
- Li, J.Y.; Dahlström, A.M.; Hersh, L.B.; Dahlström, A. Fast axonal transport of the vesicular acetylcholine transporter (VAChT) in cholinergic neurons in the rat sciatic nerve. Neurochem. Int. 1998, 32, 457–467. [Google Scholar] [CrossRef]
- Hernandez, C. M/; Høifødt, H.; Terry, AV., Jr. Spontaneously hypertensive rats: further evaluation of age-related memory performance and cholinergic marker expression. J. Psychiatry Neurosci. 2003, 28, 197–209. [Google Scholar]
- Zurkovsky, L.; Bychkov, E.; Tsakem, E.L; Siedlecki, C.; Blakely, R.D.; Gurevich, E.V. Cognitive effects of dopamine depletion in the context of diminished acetylcholine signaling capacity in mice. Dis. Model Mech. 2013, 6, 171–183. [Google Scholar] [CrossRef]
- Pirondi, S.; D'Intino, G.; Gusciglio, M.; Massella, A.; Giardino, L.; Kuteeva, E.; Ogren, S.O.; Hökfelt, T.; Calzà, L. Changes in brain cholinergic markers and spatial learning in old galanin-overexpressing mice. Brain Res. 2007, 1138, 10–20. [Google Scholar] [CrossRef]
- Katoh-Semba, R.; Semba, R.; Takeuchi, I.K.; Kato, K. Age-related changes in levels of brain-derived neurotrophic factor in selected brain regions of rats, normal mice and senescence-accelerated mice: a comparison to those of nerve growth factor and neurotrophin-3. Neurosci. Res. 1998, 31, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.Q.; Zheng, J.L.; Karihaloo, M. Neurotrophin-4/5 (NT-4/5) and brain-derived neurotrophic factor (BDNF) act at Later stages of cerebellar granule cell Differentiation. J. Neurosci. 1995, 15, 2656–2667. [Google Scholar] [CrossRef] [PubMed]
- Bartkowska, K.; Turlejski, K.; Djavadian, RL. Neurotrophins and their receptors in early development of the mammalian nervous system. Acta Neurobiol. Exp. 2010, 70, 454–467. [Google Scholar]
- Aguiar, A.S., Jr.; Castro, A.A.; Moreira, E.L.; Glaser, V.; Santos, A.R.; Tasca, C.I.; Latini, A.; Prediger, R.D. Short bouts of mild-intensity physical exercise improve spatial learning and memory in aging rats: Involvement of hippocampal plasticity via AKT, CREB and BDNF signaling. Mech. Ageing Dev. 2011, 132, 560–567. [Google Scholar] [CrossRef]
- Chae, C.H.; Lee, H.C.; Jung, S.L.; Kim, T.W.; Kim, J.H.; Kim, N.J.; Kim, H.T. Swimming exercise increases the level of nerve growth factor and stimulates neurogenesis in adult rat hippocampus. Neuroscience 2012, 212, 30–37. [Google Scholar] [CrossRef]
- Campenot, R.B. Local control of neurite development by nerve growth factor. Proc. Natl. Acad. Sci. USA 1977, 74, 4516–4519. [Google Scholar] [CrossRef] [PubMed]
- Kuruvilla, R.; Zweifel, L.S.; Glebova, N.O.; Lonze, B.E.; Valdez, G.; Ye, H.; Ginty, D.D. A neurotrophin signaling cascade coordinates sympathetic neuron development through differential control of TrkA trafficking and retrograde signaling. Cell 2004, 118, 243–255. [Google Scholar] [CrossRef]
- Gao, P.; Shen, F.; Gabriel, R.A.; Law, D.; Yang, E.Y.; Yang, G.Y.; Young, W.L.; Su, H. Attenuation of brain response to VEGF-mediated angiogenesis and neurogenesis in aged mice. Stroke 2009, 40, 3596–3600. [Google Scholar] [CrossRef]
- Rachamimov, A.; Darland, T.; D'Amore, P.A. Vascular endothelial growth factor (VEGF) isoform regulation of early forebrain development. Dev. Biol. 2011, 358, 9–22. [Google Scholar]
- Sanchez, H.L.; Silva, L.B.; Portiansky, E.L.; Herenu, C.B.; Goya, R.G.; Zuccolilli, G.O. Dopaminergic mesencephalic systems and behavioral performance in very old rats. Neuroscience 2008, 154, 1598–1606. [Google Scholar] [CrossRef]
- Del Arco, A.; Segovia, G.; de Blas, M.; Garrido, P.; Acuña-Castroviejo, D.; Pamplona, R.; Mora, F. Prefrontal cortex, caloric restriction and stress during aging: Studies on dopamine and acetylcholine release, BDNF and working memory. Behav. Brain Res. 2011, 216, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Parr-Brownlie, L.C.; Hyland, B.I. Bradykinesia induced by dopamine D2 receptor blockade is associated with reduced motor cortex activity in the rat. J. Neurosci. Res. 2005, 25, 5700–5709. [Google Scholar] [CrossRef] [PubMed]
- Lapchak, P.A.; Miller, P.J.; Jiao, S. Glial cell line-derived neurotrophic factor induces the dopaminergic and cholinergic phenotype and increases locomotor activity in aged Fischer 344 rats. Neuroscience 1997, 77, 745–752. [Google Scholar] [CrossRef]
- Love, S.; Plaha, P.; Patel, N.K.; Hotton, G.R.; Brooks, D.J.; Gill, S.S. Glial cell line–derived neurotrophic factor induces neuronal sprouting in human brain. Nat. Med. 2005, 11, 703–704. [Google Scholar] [CrossRef] [PubMed]
- Horner, A.M.; Russ, D.W.; Biknevicius, A.R. Effects of early-stage aging on locomotor dynamics and hind limb muscle force production in the rat. J. Exp. Biol. 2011, 214, 3588–3595. [Google Scholar] [CrossRef] [PubMed]
- Audet, G.N.; Meek, T.H.; Garland, T., Jr.; Olfert, I.M. Expression of angiogenic regulators and skeletal muscle capillarity in selectively bred high aerobic capacity mice. Exp. Physiol. 1996, 11, 1138–1150. [Google Scholar] [CrossRef]
- Ming, Y.; Bergman, E.; Edström, E.; Ulfhake, B. Reciprocal changes in the expression of neurotrophin mRNAs in target tissues and peripheral nerves of aged rats. Neurosci. Lett. 1999, 273, 187–190. [Google Scholar] [CrossRef]
- Andreassen, C.S.; Jacobsen, J.; Flyvbjerg, A.; Andersen, H. Expression of neurotrophic factors in diabetic muscle—relation to neuropathy and muscle strength. Brain 2009, 132, 2724–2733. [Google Scholar] [CrossRef]
- McCullough, M.J.; Peplinski, N.G.; Kinnell, K.R.; Spitsbergen, J.M. Glial cell line-derived neurotrophic factor (GDNF) protein content in rat skeletal muscle is altered by increased physical activity in vivo and in vitro. Neuroscience 2011, 174, 234–244. [Google Scholar] [CrossRef]
- Wang, W.; Esbensen, Y.; Kunke, D.; Suganthan, R.; Rachek, L.; Bjørås, M.; Eide, L. Mitochondrial DNA damage level determines neural stem cell differentiation fate. J. Neurosci. 2011, 31, 9746–9751. [Google Scholar] [CrossRef]
- Bimonte-Nelson, H.A.; Granholm, A.C.; Nelson, M.E.; Moore, A.B. Patterns of neurotrophin protein levels in male and female Fischer 344 rats from adulthood to senescence: how young is “young” and how old is "old”? Exp. Aging Res. 2008, 34, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.H.; Mattson, M.P. Neurotrophic factors protect cortical synaptic terminals against amyloid- and oxidative stress-induced impairment of glucose transport, glutamate transport and mitochondrial function. Cereb. Cortex 2000, 10, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Joo, S.S.; Kim, T.K.; Lee, S.H.; Kang, H.; Lee, H.J.; Lim, I.; Matsuo, A.; Tooyama, I.; Kim, Y.-B.; Kim, S.U. Human neural stem cells overexpressing choline acetyltransferase restore cognitive function of kainic acid-induced learning and memory deficit animals. Cell Transplant. 2012, 21, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Lee, H.J.; Joo, S.S.; Bae, D.-K.; Yang, G.; Yang, Y.-H.; Lim, I.; Matuo, A.; Tooyama, I.; Kim, Y.-B.; Kim, S.U. Human neural stem cells over-expressing choline acetyltransferase restore cognition in rat model of cognitive dysfunction. Exp. Neurol. 2012, 234, 521–526. [Google Scholar] [CrossRef]
- Park, D.; Yang, Y.-H.; Bae, D.K.; Lee, S.H.; Yang, G.; Kyung, J.; Kim, D.; Choi, E.-K.; Lee, S.W.; Kim, G.H.; Hong, J.T.; Choi, K.-C.; Kim, S.U.; Kim, Y.-B. Improvement of cognitive function and physical activity of ageing mice by human neural stem cells over-expressing choline acetyltransferase. Neurobiol. Aging 2013, 34, 2639–2646. [Google Scholar] [CrossRef]
- Lee HJ; Kim KS; Kim EJ; Choi HB; Lee KH; Park IH; Ko Y; Jeong SW; Kim SU. Brain transplantation of immortalized human neural stem cells promotes functional recovery in mouse intracerebral hemorrhage stroke model. Stem Cells 2007, 25, 1204–1212. [Google Scholar] [CrossRef]
- Kim, S.U.; Nagai, A.; Nakagawa, E.; Choi, H.B.; Bang, J.H.; Lee, H.J.; Lee, M.A.; Lee, Y.B.; Park, I.H. Production and characterization of immortal human neural stem cell line with multipotent differentiation property. Methods Mol. Biol. 2008, 438, 103–121. [Google Scholar]
- Schliebs, R.; Arendt, T. The cholinergic system in aging and neuronal degeneration. Behav. Brain Res. 2011, 221, 555–563. [Google Scholar] [CrossRef]
- Popescu, B.O.; Toescu, E.C.; Popescu, L.M.; Bajenaru, O.; Muresanu, D.F.; Schultzberg, M.; Bogdanovic, N. Blood-brain barrier alterations in ageing and dementia. J. Neurol. Sci. 2009, 283, 99–106. [Google Scholar] [CrossRef]
- Langmead, C.J.; Watson, J.; Reavill, C. Muscarinic acetylcholine receptors as CNS drug targets. Pharmacol. Ther. 2008, 117, 232–243. [Google Scholar] [CrossRef]
- Wess, J. Muscarinic acetylcholine receptor knockout mice: Novel phenotypes and clinical implications. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 423–450. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chigurupati, S.; Holloway, H.W.; Mughal, M.; Tweedie, D.; Bruestle, D.A.; Mattson, M.P.; Wang, Y.; Harvey, B.K.; Ray, B.; Lahiri, D.K.; Greig, N.H. Exendin-4 ameliorates motor neuron degeneration in cellular and animal models of amyotrophic lateral sclerosis. PLoS One 2012, 7, e32008. [Google Scholar] [CrossRef] [PubMed]
- Couillard-Despres, S.; Iglseder, B.; Aigner, L. Neurogenesis, cellular plasticity and cognition: The impact of stem cells in the adult and aging brain—a mini-review. Gerontology 2011, 57, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Encinas, J.M.; Sierra, A. Neural stem cell deforestation as the main force driving the age-related decline in adult hippocampal neurogenesis. Behav. Brain Res. 2012, 227, 433–439. [Google Scholar] [CrossRef]
- Waldau, B. Stem cell transplantation for enhancement of learning and memory in adult neurocognitive disorders. Aging Dis. 2010, 1, 60–71. [Google Scholar] [PubMed]
- Blurton-Jones, M.; Kitazawa, M.; Martinez-Coria, H.; Castello, N.A.; Müller, F.J.; Loring, J.F.; Yamasaki, T.R.; Poon, W.W.; Green, K.N.; LaFerla, F.M. Neural stem cells improve cognition via BDNF in a transgenic model of Alzheimer disease. Proc. Natl. Acad. Sci. USA 2009, 106, 13594–13599. [Google Scholar] [CrossRef]
- Lee, H.J.; Lim, I.; Park, S.W.; Kim, Y.-B.; Ko, Y.; Kim, S.U. Human neural stem cells genetically modified to express human nerve growth factor (NGF) gene restore cognition in the mouse with ibotenic acid-induced cognitive dysfunction. Cell Transplant. 2012, 21, 2487–2496. [Google Scholar] [CrossRef]
- Kim, J.; Shin, K.; Cha, Y.; Ban, Y.-H.; Park, S.K.; Jeong, H.S.; Park, D.; Choi, E.-K.; Kim, Y.-B. Neuroprotective effects of human neural stem cells over-expressing choline acetyltransferase in a middle cerebral artery occlusion model. J. Chem. Neuroanat. 2020, 103, 101730. [Google Scholar] [CrossRef]
- Park, D.; Choi, E.-K.; Cho, T.-H.; Joo, S.S.; Kim, Y.-B. Human neural stem cells encoding ChAT gene restore cognitive function via acetylcholine synthesis, Aβ elimination, and neuroregeneration in APPswe/PS1dE9 mice. Int. J. Mol. Sci. 2020, 21, 3958. [Google Scholar] [CrossRef]
- Uemura, M.; Kasahara, Y.; Nagatsuka, K.; Taguchi, A. Cell-based therapy to promote angiogenesis in the brain following ischemic damage. Curr. Vasc. Pharmacol. 2012, 10, 285–288. [Google Scholar] [CrossRef]
- Wei, L.; Fraser, J.L.; Lu, Z.Y.; Hu, X.; Yu, SP. Transplantation of hypoxia preconditioned bone marrow mesenchymal stem cells enhances angiogenesis and neurogenesis after cerebral ischemia in rats. Neurobiol. Dis. 2012, 46, 635–645. [Google Scholar] [CrossRef]
- Villeda, S.A.; Luo, J.; Mosher, K.I.; Zou, B.; Britschgi, M.; Bieri, G.; Stan, TM.; Fainberg, N.; Ding, Z.; Eggel, A.; Lucin, K.M.; Czirr, E.; Park, J.S.; Couillard-Després, S.; Aigner, L.; Li, G.; Peskind, E.R.; Kaye, J.A.; Quinn, J.F.; Galasko, D.R.; Xie, X.S.; Rando, T.A.; Wyss-Coray, T. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature 2011, 477, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Ban, Y.-H.; Park, D.; Choi, E.-K.; Kim, T.M.; Joo, S.S.; Kim, Y.-B. Effectiveness of a combinational treatment for Alzheimer’s disease with human neural stem cells and microglial cells over-expressing functional genes. Int. J. Mol. Sci. 2023, 24, 9561. [Google Scholar] [CrossRef] [PubMed]
- Pertusa, M.; García-Matas, S.; Mammeri, H.; Adell, A.; Rodrigo, T.; Mallet, J.; Cristòfol, R.; Sarkis, C.; Sanfeliu, C. Expression of GDNF transgene in astrocytes improves cognitive function deficits in aged rats. Neurobiol. Aging 2008, 29, 1366–1379. [Google Scholar] [CrossRef] [PubMed]
- Lester, D.B.; Rogers, T.D.; Blaha, C.D. Acetylcholine–dopamine interactions in the pathophysiology and treatment of CNS disorders. CNS Neurosci. Ther. 2010, 16, 137–162. [Google Scholar]
- Jiao, S.; Ren, H.; Li, Y.; Zhou, J.; Duan, C.; Lu, L. Differential regulation of IGF-I and IGF-II gene expression in skeletal muscle cells. Mol. Cell. Biochem. 2013, 373, 107–113. [Google Scholar] [CrossRef]
- Aguiar, A.F.; Vechetti-Júnior, I.J.; Alves de Souza, R.W.; Castan, E.P.; Milanezi-Aguiar, R.C.; Padovani, C.R.; Carvalho, R.F.; Silva, M.D. Myogenin, MyoD and IGF-I Regulate muscle mass but not fiber-type conversion during resistance training in rats. Int. J. Sports Med. 2013, 34, 293–301. [Google Scholar] [CrossRef]
- Yang, L.X.; Nelson, P.G. Glia cell line-derived neurotrophic factor regulates the distribution of acetylcholine receptors in mouse primary skeletal muscle cells. Neuroscience 2004, 128, 497–509. [Google Scholar] [CrossRef]
- Miura, P.; Amirouche, A.; Clow, C.; Bélanger, G.; Jasmin, B.J. Brain-derived neurotrophic factor expression is repressed during myogenic differentiation by miR-206. J. Neurochem. 2012, 120, 230–238. [Google Scholar] [CrossRef]
- Yoon, E.-J.; Seong, H.R.; Kyung, J.; Kim, D.; Park, S.; Choi, E.-K.; Kim, Y.-B.; Park, D. Stamina-enhancing effect of human adipose-derived stem cells. Cell Transplant. 2021, 30, 1–10. [Google Scholar] [CrossRef]
Figure 1.
Locomotor activity (resting, slow-moving, and fast-moving times) (A) and rota-rod performance (B) of rats at 11-20 months of age. Black: young animals (7 weeks old), red: aged animals, green: aged animals transplanted with F3 cells (1 × 106 cells every month), blue: aged animals transplanted with F3.ChAT cells (1 × 106 cells every month). *Significantly different from young animals (P<0.05). #Significantly different from aged animals (P<0.05).
Figure 1.
Locomotor activity (resting, slow-moving, and fast-moving times) (A) and rota-rod performance (B) of rats at 11-20 months of age. Black: young animals (7 weeks old), red: aged animals, green: aged animals transplanted with F3 cells (1 × 106 cells every month), blue: aged animals transplanted with F3.ChAT cells (1 × 106 cells every month). *Significantly different from young animals (P<0.05). #Significantly different from aged animals (P<0.05).
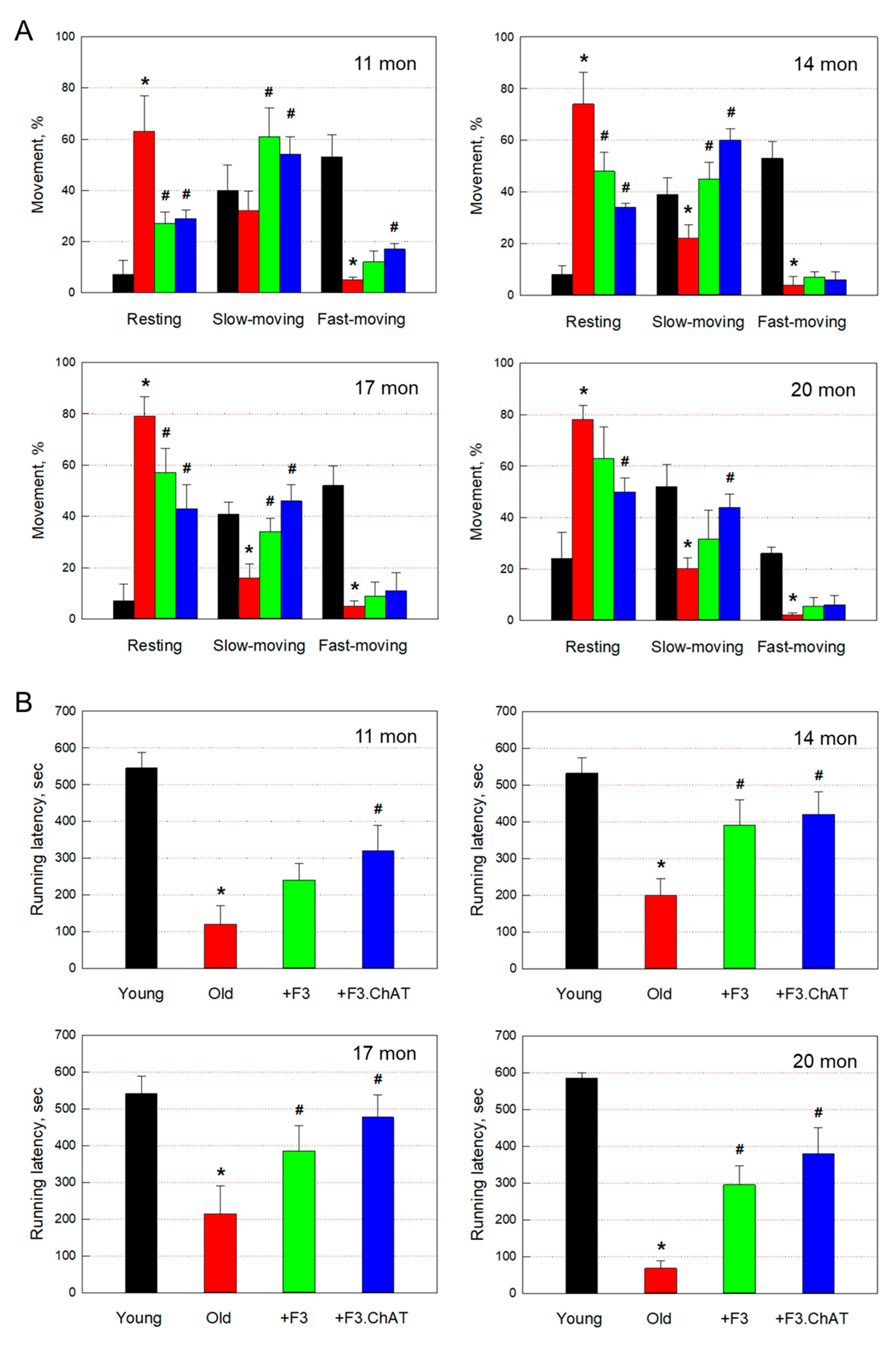
Figure 2.
Passive avoidance (A) and Morris water-maze (B) performances and acetylcholine levels in the brain (C) and muscles (D) at 21 months of age. Black: young animals (7 weeks old), red: aged animals, green: aged animals transplanted with F3 cells (1 × 106 cells every month), blue: aged animals transplanted with F3.ChAT cells (1 × 106 cells every month). *Significantly different from young animals (P<0.05). #Significantly different from aged animals (P<0.05).
Figure 2.
Passive avoidance (A) and Morris water-maze (B) performances and acetylcholine levels in the brain (C) and muscles (D) at 21 months of age. Black: young animals (7 weeks old), red: aged animals, green: aged animals transplanted with F3 cells (1 × 106 cells every month), blue: aged animals transplanted with F3.ChAT cells (1 × 106 cells every month). *Significantly different from young animals (P<0.05). #Significantly different from aged animals (P<0.05).
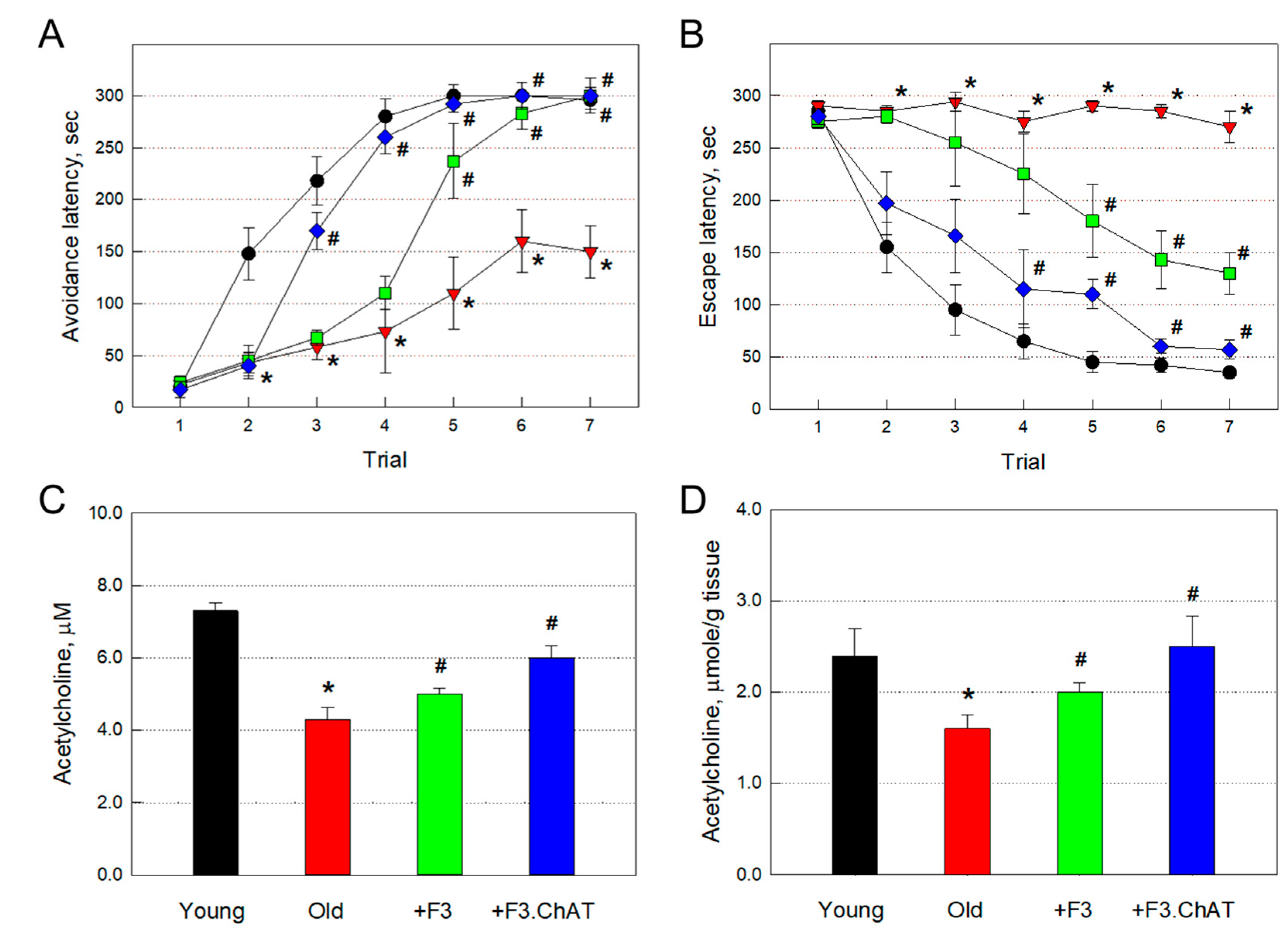
Figure 3.
Distribution of the transplanted human (hMito-positive) F3 (A & B) and F3.ChAT (A & C) cells.
Figure 3.
Distribution of the transplanted human (hMito-positive) F3 (A & B) and F3.ChAT (A & C) cells.
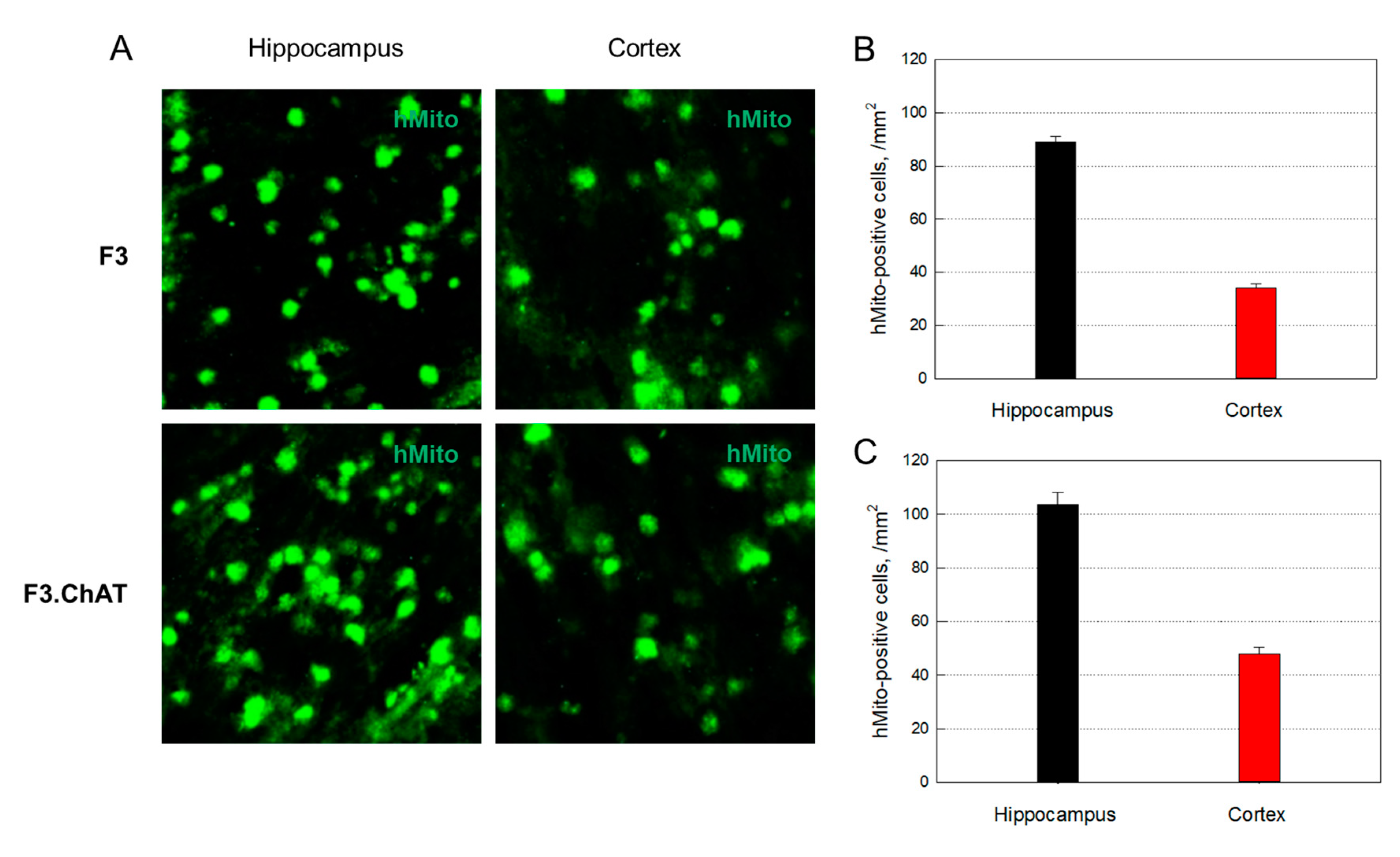
Figure 4.
Restoration of cholinergic (A) and dopaminergic (B) nervous systems following F3 or F3.ChAT cell transplantation. ChAT: choline acetyltransferase, VAChT: vesicular acetylcholine transporter, ChT1: choline transporter 1, m1-AChR: muscarinic 1 acetylcholine receptor, nAChR: nicotinic acetylcholine receptor, AChE: acetylcholinesterase, GAPDH: glyceraldehyde-3-phosphate dehydrogenase, TH: tyrosine hydroxylase, VMAT2: vesicular monoamine transporter 2, DAT: dopamine transporter, DR: dopamine receptor.
Figure 4.
Restoration of cholinergic (A) and dopaminergic (B) nervous systems following F3 or F3.ChAT cell transplantation. ChAT: choline acetyltransferase, VAChT: vesicular acetylcholine transporter, ChT1: choline transporter 1, m1-AChR: muscarinic 1 acetylcholine receptor, nAChR: nicotinic acetylcholine receptor, AChE: acetylcholinesterase, GAPDH: glyceraldehyde-3-phosphate dehydrogenase, TH: tyrosine hydroxylase, VMAT2: vesicular monoamine transporter 2, DAT: dopamine transporter, DR: dopamine receptor.
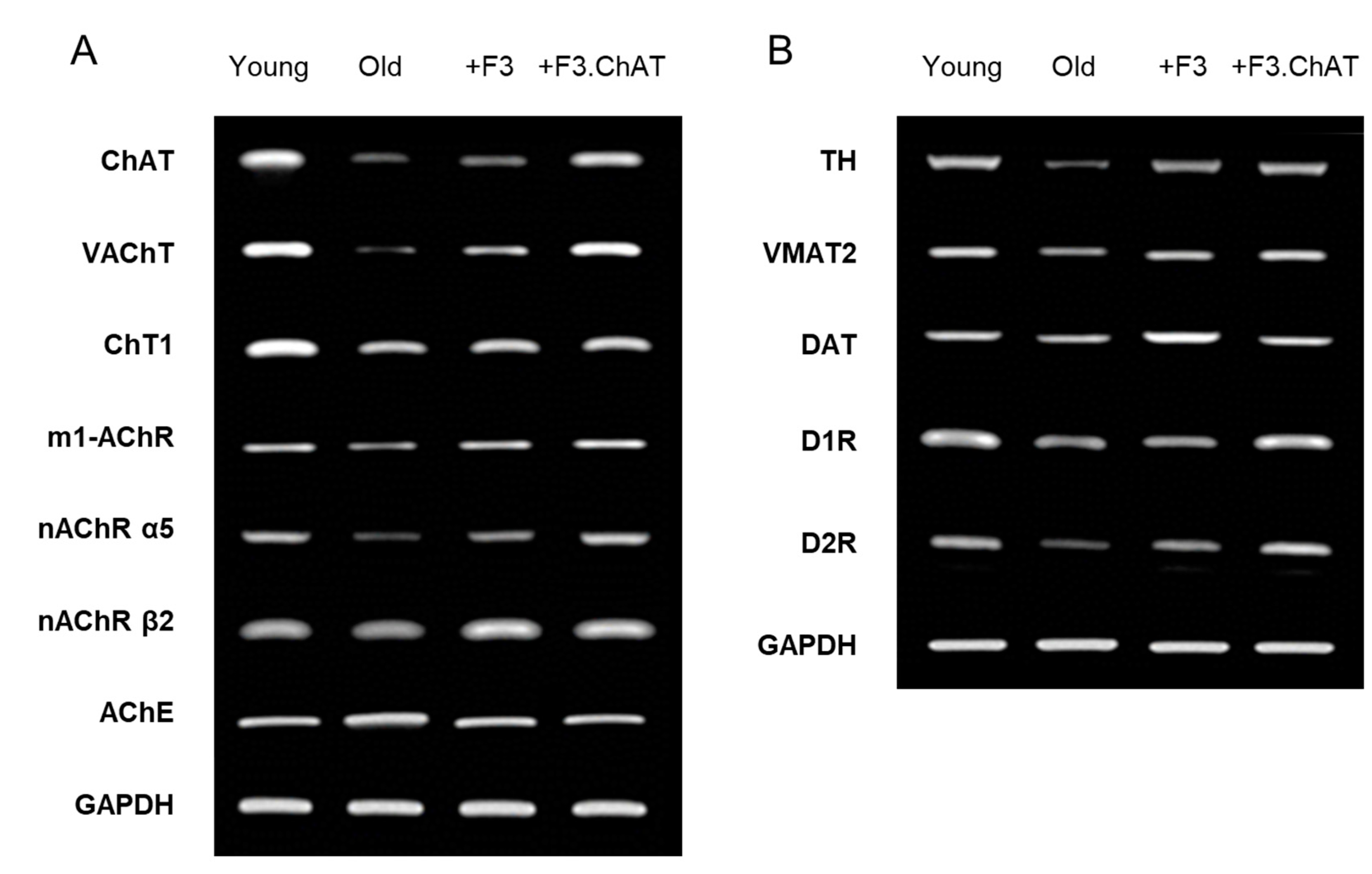
Figure 5.
Microvessel (vWF-positive) density in the brain (A & B) and muscles (D & E) and relative brain (C) and muscle (D) weights. *Significantly different from young rats (P < 0.05). #Significantly different from aged rats (P < 0.05).
Figure 5.
Microvessel (vWF-positive) density in the brain (A & B) and muscles (D & E) and relative brain (C) and muscle (D) weights. *Significantly different from young rats (P < 0.05). #Significantly different from aged rats (P < 0.05).
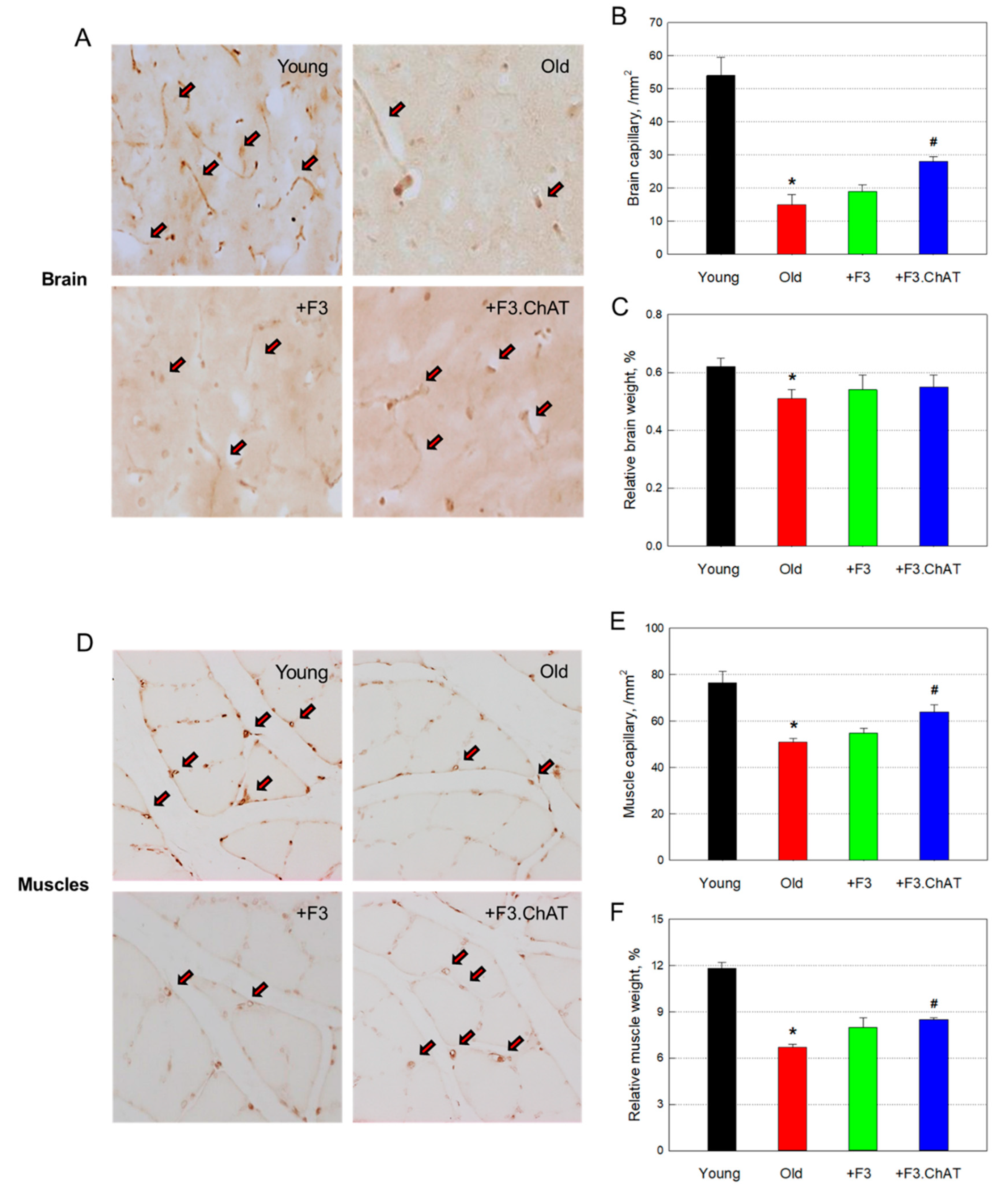
Figure 6.
Concentration of thiobarbituric acid-reactive substances (TBARS) in the brain (A) and muscles (B). *Significantly different from young rats (P < 0.05). #Significantly different from aged rats (P < 0.05).
Figure 6.
Concentration of thiobarbituric acid-reactive substances (TBARS) in the brain (A) and muscles (B). *Significantly different from young rats (P < 0.05). #Significantly different from aged rats (P < 0.05).
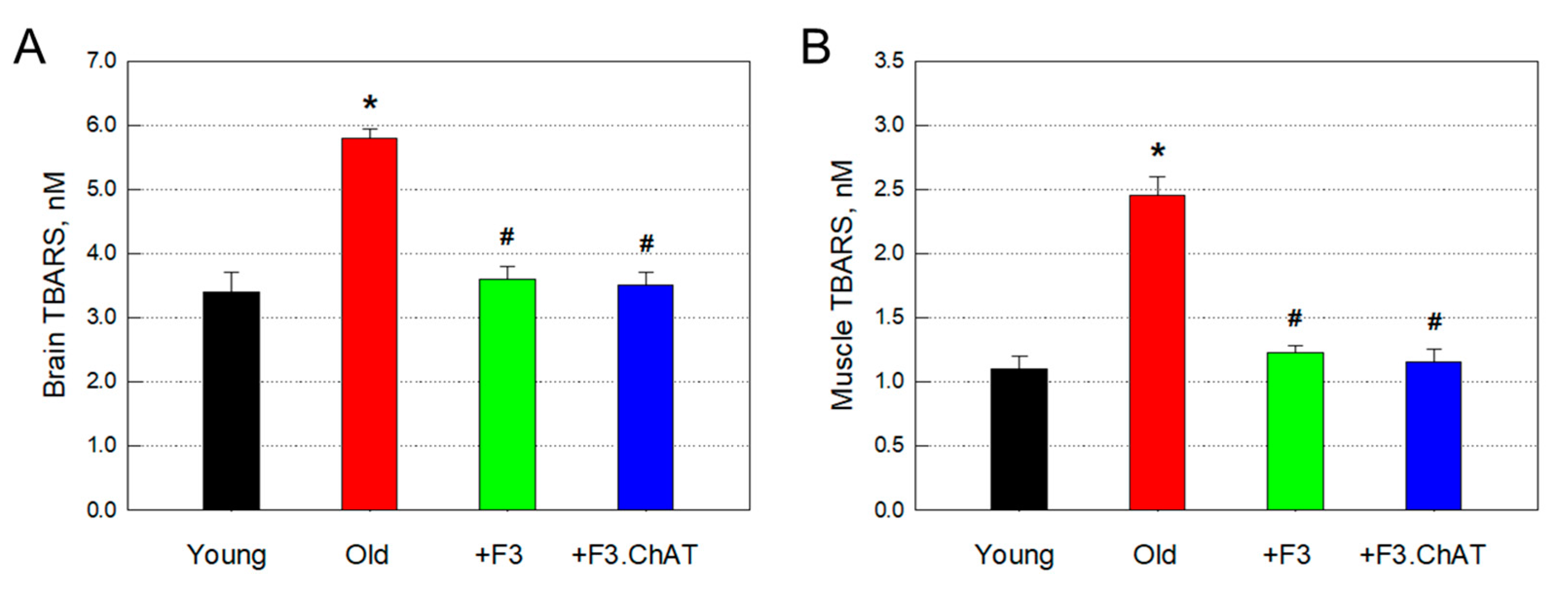
Figure 7.
Host neural stem cell (nestin-positive) regeneration and proliferation (Ki-67-positive) (A & B), CCL11 mRNA down-regulation (C), and neuronal integrity (MAP2) restoration (C). *Significantly different from young rats (P < 0.05). #Significantly different from aged rats (P < 0.05).
Figure 7.
Host neural stem cell (nestin-positive) regeneration and proliferation (Ki-67-positive) (A & B), CCL11 mRNA down-regulation (C), and neuronal integrity (MAP2) restoration (C). *Significantly different from young rats (P < 0.05). #Significantly different from aged rats (P < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



