
Submitted:
10 July 2023
Posted:
12 July 2023
You are already at the latest version
Abstract
A novel group of conjugative plasmids of Pseudomonas is characterized. The prototype plasmid pPPUT-Tik1-1 (153663 bp), isolated from a permafrost strain of P. putida Tik1, carries a defective mercury transposon Tn501 and a streptomycin resistance transposon Tn5393. Ten plasmids and 34 contigs with backbone regions closely related to pPPUT-Tik1-1 have been found in Gene Bank. Two of these plasmids isolated from clinical strains of P. putida and P. fulva are almost identical to the ancient plasmid. A characteristic feature of this group of plasmids is the presence of two genes encoding the initiators of replication (repA1 and repA2). None of these genes has high similarity with plasmid replication genes belonging to known incompatibility groups. It has been demonstrated that while pPPUT-Tik1-1-like plasmids have homologous backbone regions they significantly differ by the molecular structure and the functional burden of their accessory regions. Some of the pPPUT-Tik1-1-related plasmids carry determinants of antibiotic resistance and/or heavy metal salts. Some plasmids are characterized by the ability to degrade xenobiotics. Plasmids related to pPPUT-Tik1-1 are characterized by a narrow host range and are found in various species of Pseudomonas genus. A group of shortened plasmids containing the same replication module, but without conjugation genes, were also found. These plasmids also contain other structural changes that strongly distinguish them from plasmids related to pPPUT-Tik1-1. The data obtained indicate the need for a complex approach to the classification of plasmids.
Keywords:
novel group of plasmids
; plasmid backbone
; accessory region
; antibiotic resistance
; heavy metals resistance
; conjugation
1. Introduction
Strains of the Pseudomonas genus are found in most soil and aquatic environments and some are implicated in diseases of humans, animals and plants [1]. Many Pseudomonas isolates harbour plasmids, which contribute to the adaptability of Pseudomonas species in a variety of natural habitats [2,3,4].
The number of sequenced plasmids has increased significantly in recent years, which has caused an urgent need to improve their classification. Initially, the classification of plasmids was based on their ability (inability) to coexist in a bacterial cell. According to this criterion, 14 groups of incompatibility were identified and described in Pseudomonas [5,6], and the role of replication initiation genes in this ability was shown [6,7]. Further studies of molecular structure of plasmids, in particular their replication genes, and analysis of newly sequenced plasmids, required the development of novel approaches to improve their classification [6]. Solving this problem is difficult for a number of reasons, since some plasmids could not be classified on the basis of their incompatibility [8]. In particular, for plasmids of plant pathogens from the group of P. syringae the ability to propagate together in the same cell is a characteristic feature [2]. The nucleotide sequences of the replication systems of plasmids belonging to IncP-5, IncP-8, IncP-10, IncP-11, IncP-12, IncP-13, and IncP-14 were not determined [6,9]. Furthermore, many plasmids remain outside of distinct incompatibility groups. For example, novel incompatibility plasmid group, IncpRBL16, have been recently described [10].
To improve the plasmid classification two main methods have been developed later, i.e., typing for the replication initiation genes and typing for the relaxation (MOB genes, encoding the conjugative complex) genes [6,11,12,13,14,15]. While replicon typing aims at plasmid replication regions with non-degenerate primers, MOB typing classifies plasmids into relaxase subfamilies using degenerate primers. As a result, MOB typing provides a deeper phylogenetic depth than replicon-typing [12]. The use of both methods resulted in a significant expansion of the number of detected of new groups of plasmids [16,17].
Currently, there is no generally accepted classification system for Pseudomonas plasmids. In most cases, the criteria used for classification include the molecular structure of the backbone region (replication, partition, mobilization and conjugation modules) and the properties determined by the accessory region (resistance to heavy metals and antibiotics, ability to degrade xenobiotics, pathogenicity in relation to plants, animals and humans), as well as plasmid host range. At the same time, it is obvious that development of a satisfactory classification of plasmids is still far away [14,15].
In the present work we describe a novel group of Pseudomonas plasmids. Its prototype is a large iteron-containing conjugative plasmid found in a P. putida strain isolated from a permafrost sample aged 220-390 thousand years. Plasmids of this group, containing the common backbone region, are characterized by various accessory regions and are spread among both environmental and clinical strains of the Pseudomonas genus.
2. Results
2.1. The prototype plasmid pPPUT-Tik1-1
Bacterial strain Tik1 was isolated from a permafrost sample collected in the area of the Laptev Sea Coast. According to the phenotypic features, it was preliminarily assigned to the species P. putida. The strain was resistant to inorganic mercury compounds and to streptomycin [18]. It was shown by conjugation crosses that both mercury resistance and streptomycin resistance determinants are located on a large conjugative plasmid, which could be successfully transferred into strains P. fluorescens P22-1-2 and P. putida BSA394-2 but not into strains E. coli C600 and Acinetobacter baylii BD413 [18]. For a more detailed study of the ancient plasmid and its host strain, we performed whole-genome sequencing of the Tik1 strain. The analysis of the chromosomal sequence of Tik1 showed that this strain really belongs to the P. putida species (99.27% and 99.4% similarity with the P. putida strains JCM 18798 (BBDC00000000) and KF715 (NC_021505.1), correspondingly).
The plasmid size, determined by its sequencing, is 153663 bp; a schematic map of pPPUT-Tik1-1 is presented on Figure 1. The backbone region of pPPUT-Tik1-1 contains genes repA1 and repA2 encoding replication initiator proteins, operon parAB encoding the partition complex for segregation of plasmid copies [19] and mob-tra1, tra2 and trb modules encoding functions of plasmid conjugal transfer. The accessory region including transposons ΔTn501 and Tn5393, containing resistance determinants to mercury compounds and streptomycin, respectively, is located between the repA1 and repA2 genes. Another group of accessory genes can be found between trb and tra2 (Figure 1). It should be noted that pPPUT-Tik1-1 contains more insertions of various mobile elements, as well as their remnants, compared to the previously described related plasmid1269-2 [20].
Interestingly, we revealed in the GenBank two plasmids isolated in China, which were highly similar to pPPUT-Tik1-1. The first of them, p140352-strA (MT074087), found in a P. putida strain 15140352 isolated in 2015, was an almost identical copy of pPPUT-Tik1-1. It differed from pPPUT-Tik1-1 by only few features: an insertion of 15 bp in the gene merA and three single-nucleotide changes in the genes repA1, mobA and in the gene encoding flavodoxin reductase. The second almost identical plasmid (unnamed, CP064947.1), found in a clinical strain of P. fulva ZDHY14 (150273 bp) isolated in 2019 from the pleural fluid in a respiratory ICU. In comparison with pPPUT-Tik1-1, it contained a 3401 bp deletion eliminating three genes encoding xanthine-uracil permease, nucleoside-binding outer membrane protein and 2-oxobutyrate oxidase.
The discovery of slightly different variants of the same plasmid in ancient and modern Pseudomonas strains belonging to different species indicates the success of this plasmid and suggests its adaptability to various habitats.
2.2. Genome of pPPUT-Tik1-1 and related plasmids
In addition to the two mentioned plasmids we found in the GenBank fourteen plasmids containing the same replication and maintenance genes (repA1, repA2, parA-parB) as pPPUT-Tik1-1 and therefore related to it (Figures 2 and S1 and Tables 1 and S2). Ten of them contained conjugative genes and their backbone regions that were overall similar to pPPUT-Tik1-1 (Figures 2 and S1). However, most of them differed significantly from pPPUT-Tik1-1 and from each other in the size and the structure of their accessory regions (Figure 2).
The remaining four plasmids completely lacked conjugation genes, but other replication and maintenance genes present in their backbone regions were similar. We also found another plasmid (pGYL5) encoding the RepA1 protein that was only 56% similar to the RepA1 proteins encoded by the other 14 plasmids (Table S1). Therefore, according to the existing classification it does not belong to the group of plasmids related to pPPUT-Tik1-1. Cluster analysis of all genes encoded by the 15 plasmids using the cd-hit program with a minimum global sequence identity of 50% by amino acids showed that conjugative and non-conjugative plasmids form two separate clusters, the members of each of which are similar in their structure as a whole (Figure S1). Based on this analysis the pGYL5 plasmid is a member of the subgroup of non-conjugative plasmids, although it contains a different allele of the repA1 gene.
The conjugative plasmids from the group of pPPUT-Tik1-1 have many more genes in common than non-conjugative ones. All these genes are found in at least nine plasmids from eleven (Figure 2). At the same time, some of them are most likely not directly related to the maintenance of plasmids, for example, the arabinose efflux permease gene and an orphan zeta gene belonging to the Zeta-Epsilon family [21]. The common backbone region of plasmids related to pPPUT-Tik1-1 is about 70 kb in length (72289 bp in pPPUT-Tik1-1). The size of the smallest plasmid of this group, pS7-1M, which almost entirely consists of genes common to all plasmids of this group, is 68717 bp. The pS7-1M plasmid contains only one gene that is not included in the backbone region. The other plasmids have many more additional genes. It should be noted that in most plasmids of this group, the common backbone regions include insertions of individual unique genes or small gene clusters (Figure 2). For example, the common backbone region of pPPUT-Tik1-1 includes eight such genes. The acquisition of all these genes is not associated with insertions of any mobile elements (Figure 1).
It is important to emphasize that a characteristic feature of pPPUT-Tik1-1-related plasmids is the presence of two genes encoding two different replication initiation proteins RepA1 and RepA2. Both genes are widely distributed in plasmids of various Pseudomonas species. Many plasmids from other groups (for example, pB13-200A from P. syringae pv. tomato B13-200 [CP019872.1]; pPHE20 from P. fluorescens PC20 [KY503036]; pOXA-198 from P. aeruginosa PA41437 [MG958650.1] contain only the larger repA1 gene, while we did not find plasmids with a single repA2 gene.Therefore, it can be suggested that the product of the repA1 gene plays the main role in the process of replication initiation.
This assumption was supported by analysis of the structure of plasmid regions adjacent to the repA1 genes. Earlier Xu et al. [20] revealed in the plasmid p1269-2 of P. putida the eight 15-bp imperfect direct repeats of TCGTAtaTCaCCGAt, designated R1 to R8, upstream of repA1 gene, which were assigned as putative iterons.
In the present study, we found that p1269-2 is a member of the group of plasmids related to pPPUT-Tik1-1. Therefore we have studied the structure of similar regions in pPPUT-Tik1-1 and in all related plasmids. In all plasmids containing conjugation transfer genes, the structure of this region was the same and coincided with that described by Xu et al. [20] (Figure 3a). The only exception in this group was the smallest plasmid pS7-1M, which lacked the third iteron, although the distance between the second and fourth iterons was the same as in other plasmids. In four smaller non-conjugative plasmids, the third iteron was also absent, but the distance between the second and fourth iterons was much larger than in the large plasmids, 1788 bp instead of 119 bp (Figure 3b). This resulted from the insertion of an IS3 family element between them (Figure 3b). Thus, in non-conjugative plasmids, only 5 out of 8 iterons can be involved in the the replication initiation. The distances between these iterons and the structure of the downstream region were the same for all plasmids.
In contrast, no iteron-like sequences could be found upstream to the repA2 gene. This further suggests that the gene repA1 of pPPUT-Tik1-1-like plasmids is the main gene involved in the plasmid replication.
2.3. Identification of the replicon and relaxase types of pPPUT-Tik1-1 and related plasmids
According to modern approaches to plasmid classification, the replicon- and relaxase-based typing provide the most reliable results [6,11,12,14,22]. We used both approaches to characterize the plasmid pPPUT-Tik1-1 and related plasmids. When typing replicons, plasmid sequences of different incompatibility groups were used as references, since they directly reflect the structural features of replication proteins. Both replication genes of pPPUT-Tik1-1 were analyzed. The greatest similarity with the repA1 gene of pPPUT-Tik1-1 was observed in plasmids of the recently described IncP-11 group [9] (Table S2). However, the similarity of the repA1 gene with the replication genes from plasmids of this group is only 72% and coverage is 88-96 %. The similarity of the gene repA2 with known replication genes is even more remote (Table S2). Therefore, plasmids related to pPPUT-Tik1-1 could not be attributed to any known group of replicons and should be allocated to a new group.
It is noteworthy that based on the similarity of replication proteins, all plasmids from the pPPUT-Tik1-1 group, including eleven conjugative plasmids and four shorter non-conjugative plasmids should be attributed to the same group (Table 1). However, the structure of the tra1, tra2, trb and mob loci was highly similar in eleven conjugative plasmids, suggesting that they can also be used for classification of pPPUT-Tik1-1 and related plasmids. Earlier, Xu et al. [20], who described the first plasmid from this group, identified its relaxase as a member of the MOBP12 clade. We performed phylogenetic analysis of relaxases for plasmids related to pPPUT-Tik1-1, also taking for analysis proteins encoded by mob genes from the MOB clades P11, P12, P13, P14 [11]. This analysis revealed that the Mob protein encoded by pPPUT-Tik1-1 belongs to the MOBP13 clade (Figure S2). The tra1, tra2 and trb loci belong to the T4SS secretion system [23]. Therefore, for reliable qualification of plasmids it is necessary to take into account the structure of all components of the backbone region, in particular, the RepA1, RepA2 and MobA proteins.
Our analysis revealed that, plasmids having replicons related to pPPUT-Tik1-1 differ greatly in their structure. The loss of conjugation genes could occur only once or several independent times during the evolution of plasmids from the pPPUT-Tik1-1 group. To understand this issue, we performed phylogenetic analysis of three key proteins from the backbone region of plasmids, both replication proteins RepA1, RepA2 and the MobA relaxase. All three phylogenetic trees have a similar topology, which indicates co-evolution of all three proteins (Figure 4). Therefore, the loss of the tra-region occurred only once during the evolution of this group of plasmids. Interestingly, the replication genes of one of the conjugative plasmids, pNY7610-IMP, are closer in structure to the genes of small non-conjugative plasmids. It is noteworthy that this plasmid was found in a strain of P. aeruginosa, the species that is most phylogenetically remoted from the other host plasmid strains from the pPPUT-Tik1-1 group.
2.4. Determination of the host range of Tik1-1 and its stability in different hosts
Previously, we showed that the plasmid pPPUT-Tik1-1 is characterized by a narrow host range, being able to be transferred by conjugation to Pseudomonas strains but not to E. coli and Acinetobacter sp. [18]. In the present research, we analyzed transfer of plasmids related to pPPUT-Tik1-1 in various Pseudomonas species. By the beginning of this study, all plasmids from this group deposited in GenBank were found among strains belonging to the closest groups of P. putida and P. fluorescens [24,25]. Only recently, one plasmid from this group was found in P. aeruginosa (Table 1). Therefore, we initially assumed that the plasmids from this group could have a narrow host range within closely related species in the Pseudomonas genus. To test this assumption, we performed conjugation crosses of the strain Tik1, the original host of pPPUT-Tik1-1, and typical strains of different Pseudomonas species belonging to different groups (Table 2). It was found that the strains of remote group P. aeruginosa, were unable to maintain the replication of pPPUT-Tik1-1 (the conjugation frequency <10-9) (Table 2). At the same time the frequency of transfer of pPPUT-Tik1-1 into strains belonging to more phylogenetically close groups, including P. resinovorans, P. oleovorans, P. putida and P. fluorescence, was 10-3-10-6 (Table 2). The plasmid did not transfer into a strain of P. seleniipraecipitans, likely due to some specific properties of the recipient strain [26]. Thus, the results of the conjugation mating experiments suggested that the plasmid can be transferred only into phylogenetically close species, confirming our assumption about a narrow hosts range of pPPUT-Tik1-1.
However, a plasmid pNY7610-IMP, related to pPPUT-Tik1-1, was isolated from P. aeruginosa (CP096914) (see above), and we also found four unassembled P. aeruginosa genomes containing plasmids from this group (Table S3), suggesting that plasmids from this group can also be maintained in other species of Pseudomonas genus.
An important characteristic that must be considered in during analysis of the host-range of plasmids is their stability in the absence of selective pressure [27]. Therefore, we tested the stability of the plasmid in recipients for more than 10 passages. During this experiment, 100 % cells retained the plasmid after passaging, independently of the phylogenetic position of the recipient, indicating high stability of the plasmid in the bacterial culture.
2.5. The accessory region of pPPUT-Tik1-1 and related plasmids
The accessory regions of plasmids from the pPPUT-Tik1-1 group are characterized by considerable diversity in their size (from 3.5 kb to 80 kb) and the adaptive functions encoded by them (Table 1). And among the adaptive features encoded by accessory genes in plasmids of this group, resistance to heavy metals, toluene biodegradation and antibiotic resistance are found (Tables 1 and S3).
2.5.1. Resistance to heavy metals
Many of the plasmids from the pPPUT-Tik1-1 group (including the 34 plasmids revealed in unassembled genomes) contained mercury resistance determinants. At the same time, determinants of resistance to copper and zinc/cobalt/cadmium (czc operon) were found in only one contig; two contigs contained determinants of chromium resistance (chrAB) (Table S3).
We paid special attention to the study of mercury resistance operons and transposons present in the prototype plasmid pPPUT-Tik1-1 and other plasmids of this group. The plasmid pPPUT-Tik1-1 and its variants (p340252-strA and unnamed plasmid from P. fulva) contained the defective transposon Tn501. It contained a full mer-operon, almost identical to the mer-operon of Tn501, and remnants of the Tn501 transposition module. In particular, only the sequences of the left repeat, the spacer between it and the start of the merR gene, and 43 bp from the res site were preserved. This structure likely arose as a result of site-specific recombination in the res site.
Unlike pPPUT-Tik1-1, the plasmid pCN1276 contained two complete mercury transposons, Tn512-like (Tn5053-family) and Tn5046-like (Tn5041xTn5044 group); ΔTn5041D was revealed in the plasmid p12969-2 (Table 3). The same transposons were revealed in several contigs (Table S3). Given the limited number of plasmids in this study, this suggests the high frequency of mercury resistance transposons and their high diversity in plasmids in various Pseudomonas strains, including clinical strains.
Unexpectedly, we found another transposon (Tn5563a) in pPPUT-Tik1-1, containing an incomplete mer operon (merRTP), described by Yeo et al. [28]. It turned out that this transposon also occurs with high frequency in plasmids from the pPPUT-Tik1-1 group (in 16 from 45). In a number of cases, a complete mer operon was present in the same plasmid simultaneously with Tn5563a (Tables 2 and S3).
2.5.2. Biodegradation
Plasmid pGRT1 and contig node 6 contain genes (ttgGHI) encoding tolerance to toluene; and the pGRT1 was shown to provide the ability to use organic solvents for cell growth [29]. In both cases, the hosts of the plasmids were environmental strains of Pseudomonas putida (Tables 2 and S3).
2.5.3. Antibiotic resistance
Despite the fact that the vast majority of Pseudomonas plasmids (contigs) from the pPPUT-Tik1-1 group were isolated from clinical strains, only a few of them contained genes for resistance to antibiotics (4 out of 45). Thus, plasmid pPPUT-Tik1-1 and p12969-2 contained genes for resistance to streptomycin, pNY7610-IMP contained genes for resistance to streptomycin, gentamicin, kanamycin, carbapenems, cefalosporins and contig from strain Pseudomonas sp. NM1542 contained two copies of blavim-2 gene encoding resistance to beta-lactams. The absence of antibiotic resistance genes in the plasmids of clinical strains likely suggests that such genes can be located in other plasmids or in the chromosome. For instance, the strain P. juntendii 18091276 isolated from urine contained in chromosome gene blaimp encoding resistance to beta-lactams while its plasmid pCN1276 lacked antibiotic resistance genes.
3. Discussion
In this study, we analyzed a novel group of pPPUT-Tik1-1-related plasmids, using previously developed criteria for plasmid classification based on the genetic organization, homology and size of the backbone regions [17]. As a result, a novel group of conjugative Pseudomonas plasmids was discovered. Along with pPPUT-Tik1-1 and other eight completely sequences plasmids not previously described, it included two known plasmids: (i) p12969-2 from a clinical strain of P. putida [20] and (ii) pGRT1 from an environmental strain of P. putida [29]. Comparative analysis of the structure of all plasmids confirmed that they all belong to the same group, characterized by the presence of two replicative proteins RepA1 and RepA2. Despite significant differences in the size and structure of the accessory region, their genes encoding the main replicative and transfer functions (replication, partioning, mobilization, conjugation) are characterized by high levels of homology (Table 1). Furthermore, analysis of the phylogenetic trees were built for both replication initiation proteins and the relaxase MobA indicated co-evolution of the genes from the backbone region encoding different functions. This may indicate coordinated functioning of those genes during plasmid replication.
Based on the example of Pseudomonas plasmid, we have demonstrated the usefulness of a complex approach for plasmid classification, which takes into account the types of both replication and mobilization genes. The use of the classical approach to classify Pseudomonas plasmids by the type of their replicon proteins fails to separate two subgroups of the plasmids, containing or lacking the conjugation genes. Earlier, we found the same problem during the study of Acinetobacter plasmids [17]. The use of a complex approach in the classification of plasmids related to pPPUT-Tik1-1 made it possible to reliably determine the boundaries of pPPUT-Tik1-1 group and separate the two subgroups of these plasmids. However, when considering various groups of plasmids, the situation is complicated by the fact that, on the one hand, many plasmids either do not contain a replication initiation gene, or contain two such genes, and on the other hand, not all plasmids contain mobilization genes. According to Orlek et al. [14] only 85% and 65% of the collected plasmids contain the replicon and MOB genes, respectively. Obviously, in the absence of these genes, it is necessary to develop specific approaches for different cases.
The prototype of the novel plasmid group, the plasmid pPPUT-Tik1-1, was isolated from a permafrost sample aged 220-390 thousand years. This conjugative plasmid possesses a complete set of determinants of the type IVSS conjugation system and contains two genes encoding replication initiation proteins, one of which is adjacent to eight 15-bp iterons. Among the plasmids belonging to this group there are plasmids found in environmental Pseudomonas strains (permafrost, soil, water) as well as in clinical strains. It has been also revealed that the plasmids of the novel group are characterized by a narrow host range; they distributed among strains belonging to the genus Pseudomonas only.
It should be noted that the genus Pseudomonas is very wide and contains phylogenetically distant groups. Our data indicate the possibility of a limited distribution of plasmids related to pPPUT-Tik1-1 among strains belonging to the P. aeruginosa group. Indeed, we were unable to transfer pPPUT-Tik1-1 into any of the six used P. aeruginosa strains by conjugation. However, in the GenBank there is one complete genome of the plasmid pNY7610-IMP revealed into the strain of this species and four plasmids found among unassembled genomes of P. aeruginosa. It is noteworthy that this particular plasmid has a greater similarity of replication genes with a group of short non-conjugative plasmids (Figure 4), which may also explain its ability to replicate in the P. aeruginosa strains. Alternatively, some strains of Pseudomonas may be misannotated in the database [24,25,30]. Additional research is needed to determine the exact host range of the pPPUT-Tik1-1-related plasmids.
While the molecular structure of the backbone region of pPPUT-Tik1-1-related plasmids is characterized by a high level of homology, the accessory regions of these plasmids are highly diverse. Some contain operons (transposons) of resistance to mercury, and/or modules of resistance to other heavy metals (copper, cobalt/zinc/cadmium, as well as chromium) (Tables 1 and S3). Others contain genes of resistance to antibiotics (streptomycin, aminoglycosides, beta-lactams). Some plasmids possess genes encoding resistance to both, heavy metals and antibiotics. Finally, there are plasmids with organic solvent degradation genes. Thus, plasmids with different phenotypes belong to the same incompability group. Previously, the same pattern was described for Pseudomonas plasmids from the IncP-7 group [31] and the IncpRBL16 plasmids [10].
Numerous studies have shown a wide distribution of operons and transposons of mercury resistance in bacteria both in natural conditions and in the clinic [32,33,34,35,36]. The cause is believed to be the ubiquitous presence of mercury compounds on earth, as a result of natural phenomena [37,38,39] as well as human activities [38]. According to recently published data, up to 7.75% of bacterial genomes contain the merA gene (s), encoding mercury reductase, responsible for resistance to mercury [40]. In plasmids (contigs) of the pPPUT-Tik1-1 group, this number is 60% (27 from 45). The reason for this high frequency of occurrence of mercury resistance transposons among plasmids of the pPPUT-Tik1-1 group remains unclear. Also surprising is the wide distribution of the Tn5563a transposon, which contains a defective mer operon carrying intracellular mercury transport genes merP and merT but lacking the mercury reductase gene. It is known that such combination of mer genes may cause hypersensitivity to mercury compounds [41,42]. However, in many cases, the genomes of these strains simultaneously contain complete mer operons, which could neutralize the negative effect of Tn5563a (Tables 2 and S3).
The possible role of plasmids of the novel group in the spread of antibiotic resistance in the clinic remains to be studied. Since most Pseudomonas strains are rare in the clinic, and do not have high virulence, some researchers suggested that they may play a role in the exchange of antibiotic resistance genes to virulent strains of P. aeruginosa [43,44]. Most of the plasmids from the new group do not contain determinants of antibiotic resistance and likely do not play much role in the distribution of these genes or their accumulation. However, this year a complete sequence of a plasmid from a clinical strain of P. aeruginosa NY7610 carrying class 1 integron and six resistance genes to antibiotics of various classes was submitted to the GenBank. As in the case of environmental Acinetobacter mega-plasmids [45], the active use of antibiotics may result in the recruitment of new environmental plasmids to the clinic.
At the same time, the wide presence of pPPUT-Tik1-1-like plasmids in strains inhabiting various ecological niches suggests that they may play a role in adaptability of their host bacteria. This assumption is confirmed by the fact that many pPPUT-Tik1-1-like plasmids encode adaptive genes that are different for different plasmids.
In conclusion, it should be emphasized that the data obtained from comparative analysis of plasmids of the pPPUT-Tik1-1 group convincingly indicate the need and promise of using complex approaches in the classification of plasmids.
4. Materials and Methods
4.1. Media and growth conditions
Bacteria were grown in lysogeny broth (LB) medium or solidified agar LB medium (LA) [46] at 30C. When required, LA medium was supplemented with antimicrobial agents at the following final concentrations (µg/ml): HgCl2 (Hg) 5-8; streptomycin (Sm) 100-200; rifampicin (Rif) 25.
4.2. Bacterial strains
The strain of P. putida Tik1 was isolated from a permafrost sample aged 220-390 thousand years collected in the area of the Laptev Sea Coast. A variety of Pseudomonas species strains were obtained from the All-Russian National Collection of Industrial Microorganisms (VKPM), All-Russian Collection of Microorganisms (VKM) and from the collection of the Institute of Molecular Genetics (IMG) (Table 2). All the Pseudomonas strains used as recipients in conjugative crosses were first marked with spontaneous mutations to rifampicin resistance (rifR). To this goal, bacterial cells grown in LB to OD600=0.8 were plated onto LA supplemented with rifampicin. The colonies were replated on the same medium. Mutants with the best growth in the presence of rifampicin were selected and used as recipients in mating experiments.
4.3. Matings
Plasmid conjugal transfer experiments were carried out as previously described [45]. Briefly, overnight cultures were diluted 50-fold in fresh broth and were grown 3-4 hours (to logarithmic growth stage), then a total of 25 µl of culture of donor strain Tik1 and each of recipient bacteria (Table 2) were mixed together, plated onto LA and incubated at 30°C for 16–18 h. The grown cells was scrapped off the plate, resuspended and plated onto LA supplemented with antimicrobial agents after appropriate dilutions.
4.4. Plasmid stability assays
Plasmid stability assays were performed essentially as described in Deane & Rawlings [47]. Briefly, transconjugants containing pPPUT-Tik1-1 were grown to the stationary phase in the absence of selection and then diluted 100-fold in fresh broth. This procedure was repeated twice a day for 4 days. The final cultures were plated onto LA after appropriate dilutions, and tested for streptomycin and mercury resistance.
4.5. Identification of the backbone region of pPPUT-Tik1-1
The genes involved in conjugation (mob genes and mating pair formation (MPF) genes) were identified by amino acid similarity with genes of previously described plasmids. The plasmid R64 (AB027308.1, NC_005014) from Salmonela enterica and plasmid pA297-3 (KU744946.1) isolated from A. baumannii A297 [48] were used as references for the MPF I group of the T4SS system and CPT4 from the MOBF family.
4.6. Search for plasmids related to pPPUT-Tik1-1 in GenBank
Plasmids related to pPPUT-Tik1-1 from modern Pseudomonas strains were identified using the BLASTn program. The sequence of pPPUT-Tik1-1 was used as query to search for related plasmids in the NCBI database containing complete plasmid genomes on April 1, 2023. All plasmids that had the query cover greater than 50% and the identity of the common region greater than 98.5% were considered as related to pPPUT-Tik1-1. Since all the detected plasmids contained genes encoding the coupling protein TrbC and the putative replication initiator protein Rep, the sequences of these two genes were used as queries to search for related sequences in the NCBI database containing whole-genome shotgun contigs on December 31, 2022.
4.7. Whole-genome sequencing and plasmid assembly
The Illumina (Illumina, United States) and single molecular nanopore sequencing (Oxford Nanopore, United Kingdom) technologies were used for the sequencing of the genome of P. putida Tik1. As a result of sequencing of the genomic DNA library on Illumina MiSeq, 3.3 million read pairs (2 × 300 bp) were obtained. Additionally, genomic DNA was sequenced using the MinION system (Oxford Nanopore, UK), resulting in which 65548 reads with an average length of 19027 bp (a total of 1.2 Gb). Nanopore reads were assembled with the Flye v. 2.7b software. Illumina reads was used to polish assembly with two iterations using pilon v1.22. As a result, a complete circular sequence of the chromosome with a length of 6433697 bp and one circular contig of 153663 bp, representing the plasmid designated pPPUT-Tik1-1, were obtained.
4.8. Bioinformatic analysis
Multiple alignment of relaxase genes was constructed with MUSCLE v3.8.31, the alignment was used as an input for the Maximum likelihood tree construction in PhyML 3.0 with default parameters. All genes encoded by 15 plasmids related to pPPUT-Tik1-1 were clustered using CD-HIT v4.6 program with a minimum global sequence identity of 50%. Plasmids were clustered according to the presence/absence of genes using hclust function in R v4.2.1 programming language, the results of the analysis were visualized with the heatmap.2 function in R. Phylogenetic analysis of RepA1, RepA2 and relaxase MobA was performed using FastTree v.2.1.11 tool with default parameters, MUSCLE was used to build the input alignments.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Whole genome analysis of the plasmids, related to pPPUT-Tik1-1; Figure S2: Phylogenetic tree of the mob genes from clades P11, P12, P13, P14; Table S1: Characteristics of the Pseudomonas plasmids related to pPPU-Tik1-1; Table S2: Identification of the replicon type of pPPUT-Tik1-1; Table S3: List of the pPPUT-Tik1-1 homologs found in whole shortgun contigs sequences.
Author Contributions
Conceptualization, M.P. and S.M; investigation, O.M. and N.P.; validation, O.M., A.B., N.P. and S.M.; formal analysis, A.B., and S.M.; data curation, M.P., A.M. and S.M.; writing—original draft preparation, M.P. and S.M., O.M., A.B.; writing—review and editing, M.P., and A.M.; visualization, A.B. and M.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Genome Research Centre development program «Kurchatov Genome Centre» (agreement No.075-15-2019-1659).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequence of pPPUT-Tik1-1 was submitted to GenBank and can be found under the accession number OQ920555.
Acknowledgments
The authors are grateful to the staff of the collections VKPM and VKM for the Pseudomonas strains provided and to A. Kulbachinskiy for helpful comments and suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wackett, LP. Pseudomonas putida – a versatile biocatalyst. Nat Biotechnol. 2003, 21(2), 136–8. [Google Scholar] [CrossRef]
- Gutiérrez-Barranquero, JA, Cazorla, FM, de Vicente, A, Sund, GW. Complete sequence and comparative genomic analysis of eight native Pseudomonas syringae plasmids belonging to the pPT23A family. BMC Genomics 2017, 18, 365. [CrossRef]
- Li, Z, Cai, Z, Cai, Z, Zhang, Y, Fu, T, Jin, Y, Cheng, Z, Jin, S, Wu, W, Yang, L, Ba,i F.J Molecular genetic analysis of an XDR Pseudomonas aeruginosa ST664 clone carrying multiple conjugal plasmids. J Molecular genetic analysis of an XDR Pseudomonas aeruginosa ST664 clone carrying multiple conjugal plasmids. Antimicrob Chemother. 2020, 75(6), 1443–52. [CrossRef]
- Paul, D, Chanda, DD, Chakravarty, A, Bhattacharjee, A. An insight into analysis and elimination of plasmids encoding metallo-β-lactamases in Pseudomonas aeruginosa. J Glob Antimicrob Resist. 2020, 21, 3–7. [CrossRef]
- Novick, R. P. Plasmid incompatibility. Microbiol. Rev. 1987, 51, 381–95. [Google Scholar] [CrossRef] [PubMed]
- Shintani, M, Sanchez, ZK, Kimbara, K. Genomics of microbial plasmids: classification and identification based on replication and transfer systems and host taxonomy. Front Microbiol. 2015, 6, 242. [CrossRef]
- Bardaji, L, Añorga, M, Ruiz-Masó, JA, Del Solar, G, Murillo, J. Plasmid replicons from Pseudomonas are natural chimeras of functional, exchangeable modules. Front Microbiol. 2017, 8, 190. [CrossRef]
- Sentchilo, VS, Perebituk, AN, Zehnder, AJ, van der Meer, JR. Molecular diversity of plasmids bearing genes that encode toluene and xylene metabolism in Pseudomonas strains isolated from different contaminated sites in Belarus. Appl Environ Microbiol. 2000, 66(7), 2842–52. [CrossRef]
- Bonnin, RA, Bogaerts, P, Girlich, D, Huang, TD, Dortet, L, Glupczynski, Y, Naas, T. Molecular characterization of OXA-198 carbapenemase-producing Pseudomonas aeruginosa clinical isolates. Antimicrob Agents Chemother 2018, 62, 02496-17. [CrossRef]
- Jiang, X, Yin, Z, Yuan, M, Cheng, Q, Hu, L et al. Plasmids of novel incompatibility group IncpRBL16 from Pseudomonas species. J Antimicrob Chemother. 2020, 75(8), 2093–2100. [CrossRef]
- Garcillán-Barcia, MP, Francia, MV, de la Cruz, F. The diversity of conjugative relaxases and its application in plasmid classification. FEMS Microbiol. Rev. 2009, 33, 657–87. [CrossRef] [PubMed]
- Garcillán-Barcia, MP, de la Cruz, F. Ordering the bestiary of genetic elements transmissible by conjugation. Mob Genet Elements. 2013, 3(1), 24263. [CrossRef]
- Smillie, C, Garcillán-Barcia, MP, Francia, MV, Rocha, EP, de la Cruz, F. Mobility of plasmids. Microbiol Mol Biol Rev. 2010, 74(3), 434–52. [CrossRef]
- Orlek, A, Phan, H, Sheppard, AE, Doumith, M, Ellington, M, et al. Ordering the mob: Insights into replicon and MOB typing schemes from analysis of a curated dataset of publicly available plasmids. Plasmid 2017, 91, 42–52. [CrossRef] [PubMed]
- Chen, F, Wang, P, Yin, Z, Yang, H, Hu, L et al. VIM-encoding IncpSTY plasmids and chromosome-borne integrative and mobilizable elements (IMEs) and integrative and conjugative elements (ICEs) in Pseudomonas. Ann Clin Microbiol Antimicrob 2022, 21, 10. [CrossRef]
- Moura, A, Oliveira, C, Henriques, I, Smalla, K, Correia, A. Broad diversity of conjugative plasmids in integron-carrying bacteria from wastewater environments. FEMS Microbiol Lett. 2012, 330, 157–164. [CrossRef]
- Mindlin, S, Beletsky, A, Rakitin, A, Mardanov, A, Petrova, M. Acinetobacter plasmids: diversity and Development of classification strategies. Front Microbiol. 2020, 11, 588410. [CrossRef] [PubMed]
- Mindlin, SZ, Petrova, M., Gorlenko, ZM, Soina, VS, Khachikian, NA, Karaevskaya, EA. Multidrug-resistant bacteria in permafrost: Isolation, biodiversity, phenotypic and genotypic analysis. In New Permafrost and Glacier Research, 1st ed.; Krugger, M.I., Stern, H.P., Eds.; Nova Science: Hauppauge, NY, USA, 2009; pp. 89–105. [Google Scholar]
- Gerdes, K, Moller-Jensen, J, Bugge Jensen, R. Plasmid and chromosome partitioning: surprises from phylogeny. Mol. Microbiol. 2000, 37, 455–466. [CrossRef]
- Xu,Y, Niu,Y, Sun, F, Yang,Y, Luo, W, Wang, Z. The novel Pseudomonas putida plasmid p12969-2 harbors an In127-carrying multidrug-resistant region. Future Microbiol. 2017, 12, 573–84. [CrossRef]
- Mutschler, H, Meinhart, AJ, ε/ζ systems: their role in resistance, virulence, and their potential for antibiotic development. Mol Med (Berl). 2011, 89(12), 1183–94. [CrossRef]
- Carattoli, A, Zankari, E, García-Fernández, A, Voldby Larsen, M, Lund O, Villa, L, Møller Aarestrup, F, Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother. 2014, 58(7), 3895–903. [CrossRef]
- Cabezón, E, Ripoll-Rozada, J, Peña, A, de la Cruz, F, Arechaga, I. Towards an integrated model of bacterial conjugation. FEMS Microbiol Rev. 2015, 39(1), 81–95. [CrossRef]
- Hesse, C, Schulz, F, Bull, CT, Shaffer, BT, Yan, Q, Shapiro, N et al. Genome-based evolutionary history of Pseudomonas spp. Environ Microbiol 2018, 20, 2142–2159. [CrossRef] [PubMed]
- Passarelli-Araujo, H, Franco, GR, Venancio, TM. Network analysis of ten thousand genomes shed light on Pseudomonas diversity and classification. Microbiol Res. 2022, 254, 126919. [CrossRef] [PubMed]
- Virolle, C., Goldlust, K., Djermoun, S, Bigot, S, Lesterlin, C. Plasmid transfer by conjugation in gram-negative bacteria: from the cellular to the community level. Genes 2020, 11, 1239. [CrossRef]
- Gelder, LD, Ponciano, JM, Joyce, P, Top, EM. Stability of a promiscuous plasmid in different hosts: no guarantee for a long-term relationship. Microbiology 2007, 153, 452–463. [CrossRef] [PubMed]
- Yeo, CC, Tham, JM, Kwong, SM, Yiin, S, Poh, CL. Tn5563, a transposon encoding putative mercuric ion transport proteins located on plasmid pRA2 of Pseudomonas alcaligenes. Tn 5563, 165(2), 253–60. [CrossRef]
- Molina, L, Duque, E, Gomez, MJ, Krell, T, Lacal, J, García-Puente, A et al. The pGRT1 plasmid of Pseudomonas putida DOT-T1E encodes functions relevant for survival under harsh conditions in the environment. Environ. Microbiol. 2011, 13(8), 2315-27. [CrossRef]
- Lalucat, J, Mulet, M, Gomila, M, García-Valdés, E. Genomics in Bacterial Taxonomy: Impact on the Genus Pseudomonas. Genes (Basel) 2020, 11(2), 139. [CrossRef]
- Volkova, OV, Panov, AV, Kosheleva, IA, Boronin, AM. IncP-7 plasmids classification based on structural diversity of their basic replicon. Mol Biol (Mosk). 2013, 47(2), 232-42. [CrossRef]
- Osborn, AM, Bruce, KD, Strike, P, Ritchie, DA. Distribution, diversity and evolution of the bacterial mercury resistance (mer) operon. FEMS Microbiol Rev. 1997, 19(4), 239-62. [CrossRef]
- Liebert, CA, Hal,l RM, Summers, AO. Transposon Tn21, flagship of the floating genome. Microbiol Mol Biol Rev. 1999, 63(3), 507-22. [CrossRef]
- Schneiker, S, Keller, M, Dröge, M, Lanka, E, Pühler, A, Selbitschka, W. The genetic organization and evolution of the broad host range mercury resistance plasmid pSB102 isolated from a microbial population residing in the rhizosphere of alfalfa. Nucleic Acids Res. 2001, 29(24), 5169-81. 24. [CrossRef]
- Mindlin, S, Minakhin, L, Petrova, M, Kholodi,i G, Minakhina, S, Gorlenko, Zh, Nikiforov, V. Present-day mercury resistance transposons are common in bacteria preserved in permafrost grounds since the Upper Pleistocene. Res Microbiol. 2005, 156(10), 994-1004. [CrossRef]
- Barkay, T, Miller, SM, Summers, AO. Bacterial mercury resistance from atoms to ecosystems. FEMS Microbiol Rev. 2003, 27(2-3), 355-84. [CrossRef]
- Driscoll, CT, Mason, RP, Chan, HM, Jacob, DJ, Pirrone, N. Mercury as a global pollutant: sources, pathways, and effects. Environ Sci Technol. 2013, 47(10), 4967-83. [CrossRef]
- Obrist, D, Kirk, JL, Zhang, L, Sunderland, EM, Jiskra, M, Selin, NE. A review of global environmental mercury processes in response to human and natural perturbations: Changes of emissions, climate, and land use. Ambio. 2018, 47(2), 116-140. [CrossRef]
- Raj, D, Maiti, SK. Sources, toxicity, and remediation of mercury: an essence review. Environ Monit Assess. 2019, 191(9), 566. [CrossRef]
- Christakis, CA, Barkay, T, Boyd, ES. Expanded diversity and phylogeny of mer genes broadens mercury resistance paradigms and reveals an origin for MerA among thermophilic archaea. Front Microbiol. 2021, 12, 682605. [CrossRef]
- Babich, K, Engle, M, JS, Laddaga, RA. Deletion mutant analysis of the Staphylococcus aureus plasmid pI258 mercury-resistance determinant. Can J Microbiol. 1991, 7(8), 624-31. [CrossRef]
- Hobman, JL, Brown, NL. Overexpression of MerT, the mercuric ion transport protein of transposon Tn501, and genetic selection of mercury hypersensitivity mutations. Mol Gen Genet. 1996, 250(1), 129-34. [CrossRef]
- Juan, C, Zamorano, L, Mena, A, Albertí, S, Pérez, JL, Oliver, A. Metallo-beta-lactamase-producing Pseudomonas putida as a reservoir of multidrug resistance elements that can be transferred to successful Pseudomonas aeruginosa clones. J Antimicrob Chemother. 2010, 65(3), 474-8. [CrossRef]
- Peter, S, Oberhettinger, P, Schuele, L, Dinkelacker, A, Vogel, W. Genomic characterisation of clinical and environmental Pseudomonas putida group strains and determination of their role in the transfer of antimicrobial resistance genes to Pseudomonas aeruginosa. BMC Genomics 2017, 18(1), 859. [CrossRef]
- Mindlin, S, Maslova, O, Beletsky, A, Nurmukanova, V, Zong, Z, Mardanov, A, Petrova M. Ubiquitous conjugative mega-plasmids of Acinetobacter species and their role in horizontal transfer of multi-drug resistance. Front Microbiol. 2021,12, 728644. [CrossRef]
- Sambrook, J, Russell, D. W. (2001). Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, USA, 3d edition.
- Deane, S., Rawlings, D. Plasmid evolution and interaction between the plasmid addiction stability systems of two related broad-host-range IncQ-like plasmids. J Bacteriol. 2004, 186(7), 2123-33. [CrossRef]
- Hamidian, M., Ambrose, SJ, Hall, RM. A large conjugative Acinetobacter baumannii plasmid carrying the sul2 sulphonamide and strAB streptomycin resistance genes. Plasmid 2016, 87-88, 43-50. [CrossRef]
Figure 1.
Schematic map of pPPUT-Tik1-1. Genes are denoted by arrows and colored based on the gene function classification shown in the upper left corner. The innermost circle represents GC-content. The second circle shows positions of the backbone (blue) and accessory (pink) regions. The third circle shows positions of mobile elements (brown). The fourth and fifth circles show genes encoded in the minus and plus plasmid strands, respectively.
Figure 1.
Schematic map of pPPUT-Tik1-1. Genes are denoted by arrows and colored based on the gene function classification shown in the upper left corner. The innermost circle represents GC-content. The second circle shows positions of the backbone (blue) and accessory (pink) regions. The third circle shows positions of mobile elements (brown). The fourth and fifth circles show genes encoded in the minus and plus plasmid strands, respectively.
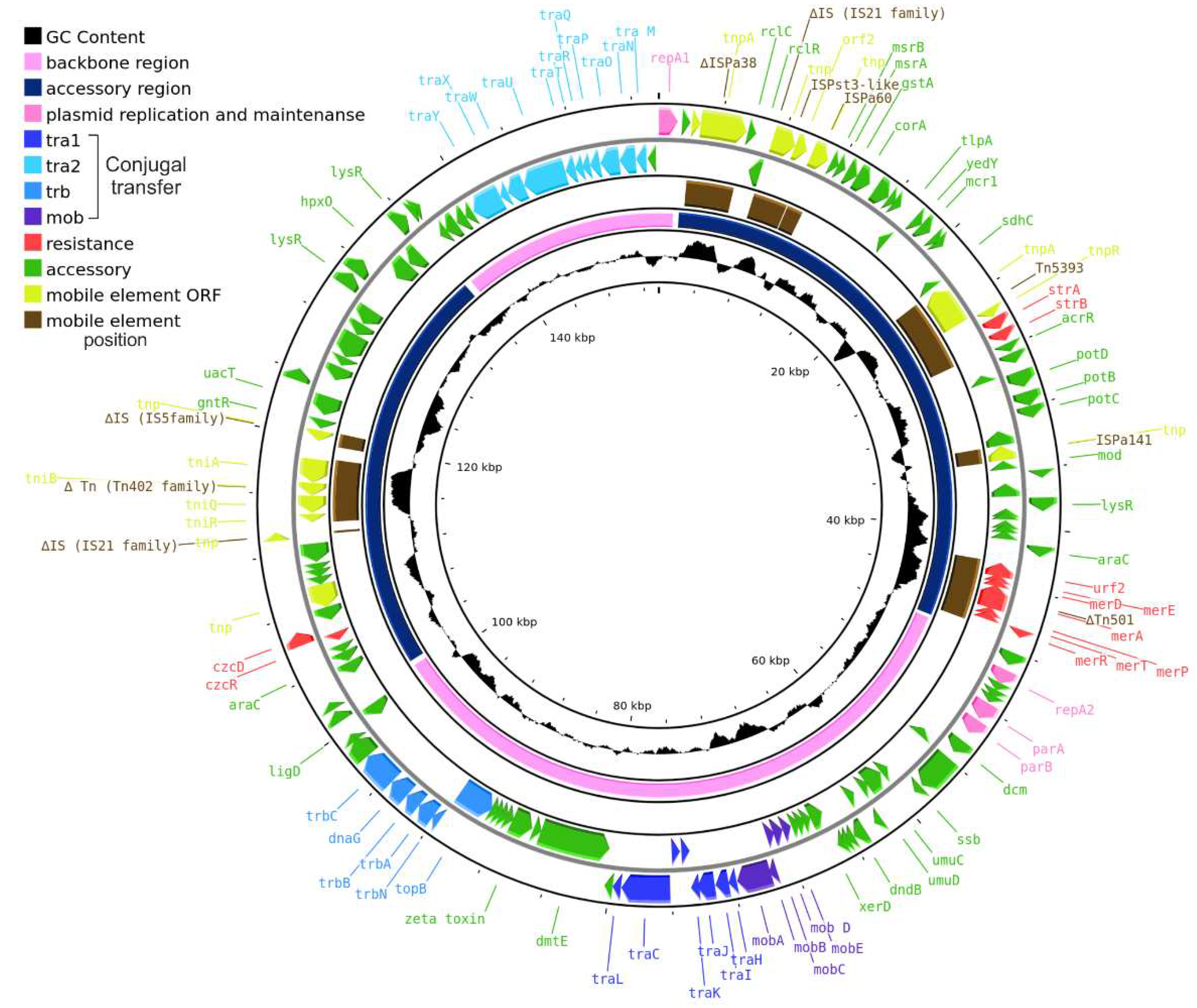
Figure 2.
Comparative molecular structure of the plasmids, related to pPPUT-Tik1-1.
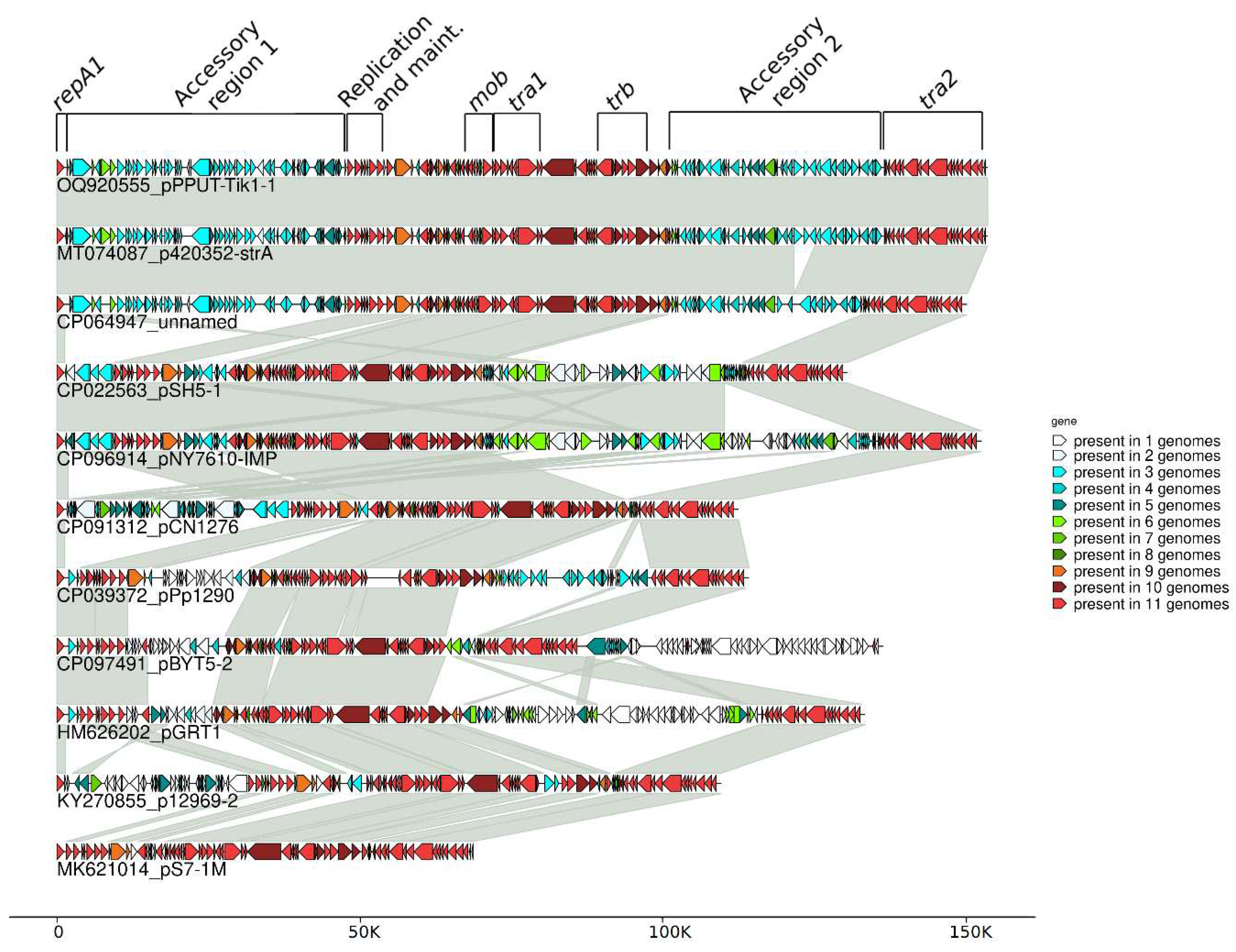
Figure 3.
Schematic representation of the iterons and downstream regions of plasmids related to pPPUT-Tik1-1. The number of nucleotides between iterons and conservative elements of the repA1 gene promoter is indicated.
Figure 3.
Schematic representation of the iterons and downstream regions of plasmids related to pPPUT-Tik1-1. The number of nucleotides between iterons and conservative elements of the repA1 gene promoter is indicated.
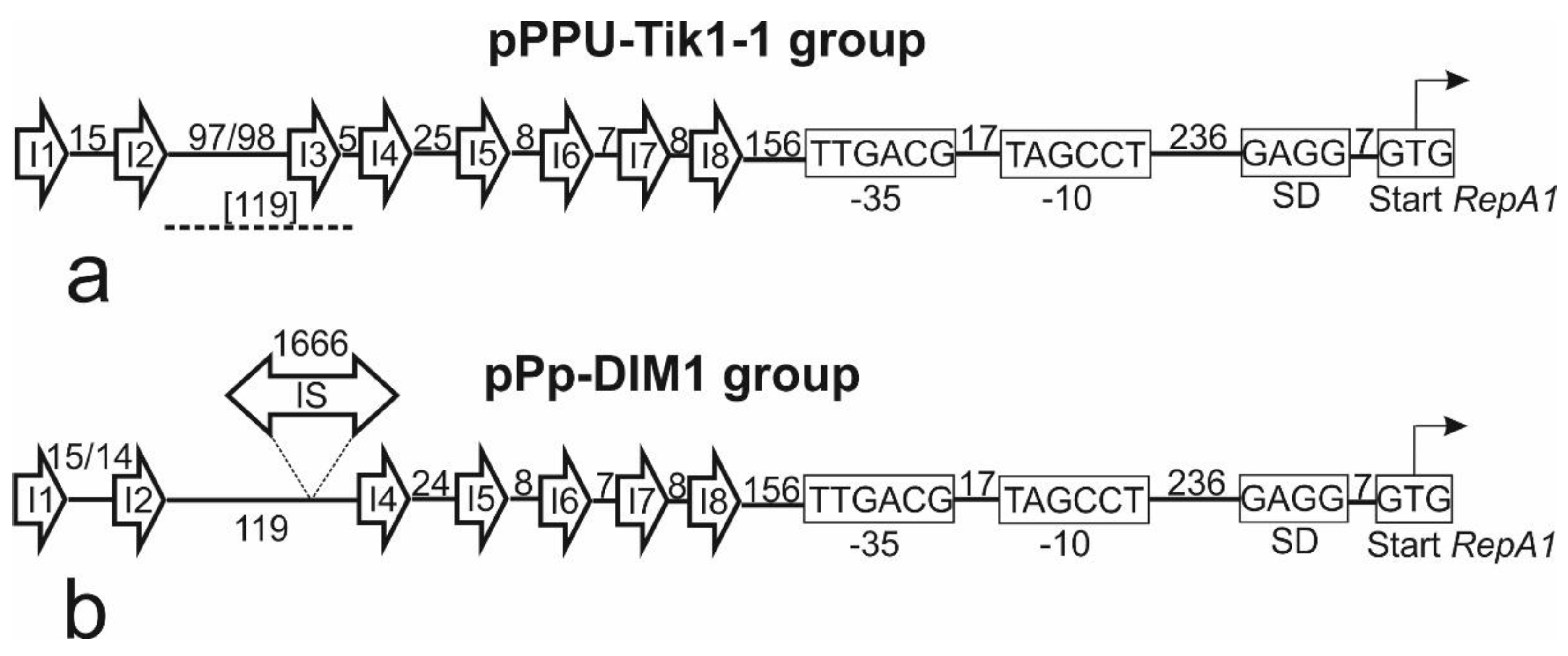
Figure 4.
Phylogenetic trees of the RepA1, RepA2 and MobA proteins.
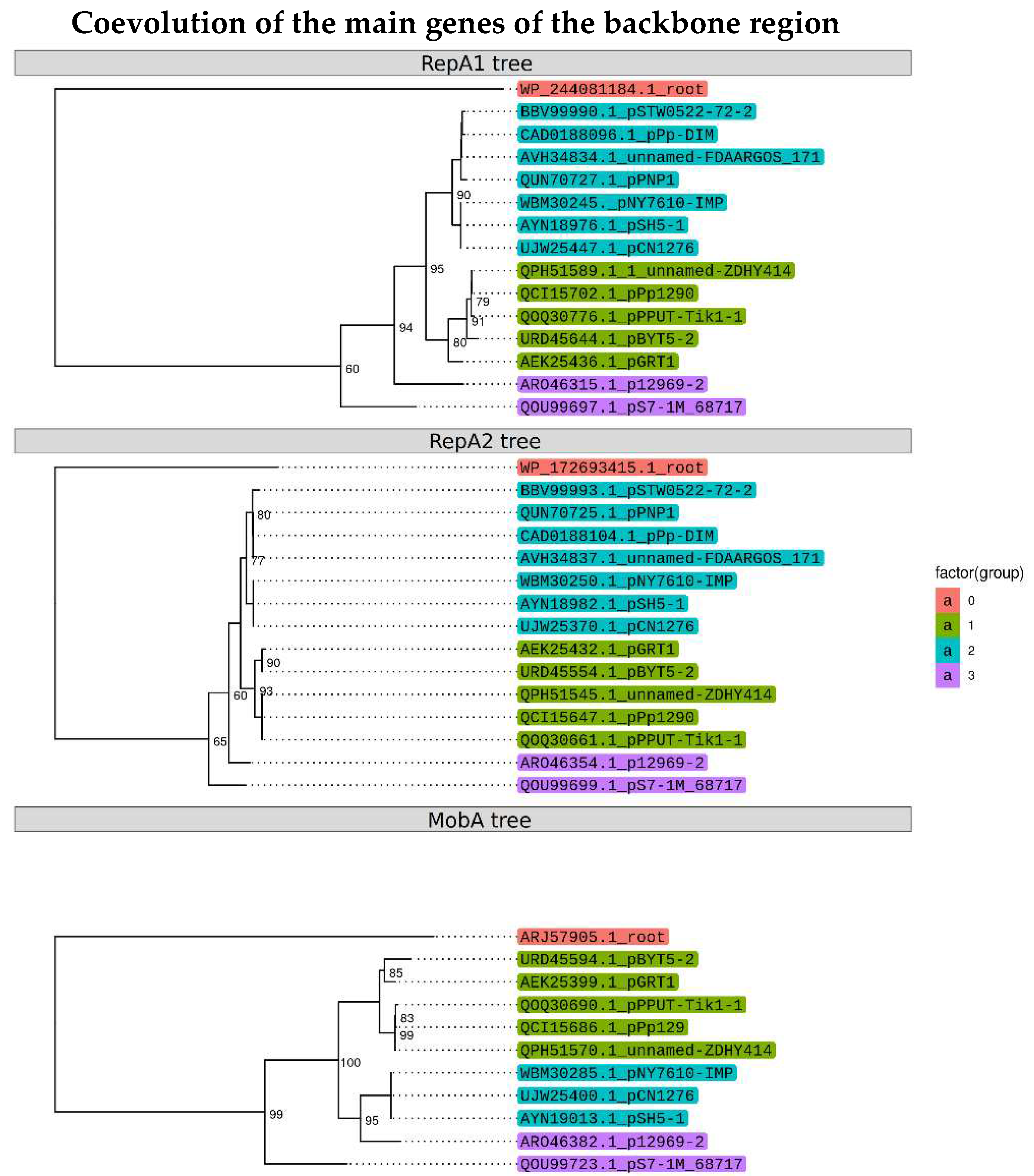

Table 1.
Characteristics of the Pseudomonas plasmids related to pPPUT-Tik1-1.
| Strain | Plasmid | Size (bp) | Source | Accessory region1 | Accession Number |
|---|---|---|---|---|---|
| Conjugative plasmids (Tra+) | |||||
| P. putida Tik1 | pPPUT-Tik1-1 | 153663 | permafrost, Russia | Hg, Str | OQ920555 |
| P. putida 15420352 | p420352-strA | 153678 | urine, China | Hg, Str | MT074087 |
| P.fulva ZDHY414 | Unnamed | 150273 | clinic, China | Hg, Str | CP064947 |
| P. putida 12969 | p12969-2 | 109708 | clinic, China | Hg, Str, Sul | KY270855 |
| P. putida 1290 | pPp1290 | 114265 | pear pyllosphere | Aromatic compounds degradation | CP039372 |
| P. putida DOT-T1E | pGRT1 | 133451 | unknown | Tolerance to toluene, UV-resistance | HM626202 |
| P. juntendi 18091276 | pCN1276 | 112529 | urine, China | Hg | CP091312 |
| P. monteilii B5 | pSH5-1 | 130536 | soil | Str | CP022562 |
| Pseudomonas sp. BYT-5 | pBYT5-2 | 136400 | soil | Molybdate absorption | CP097489 |
| P. aeruginosa NY7610 | pNY7610-IMP | 152682 | clinic, China | Str, Gm, Km, Cb, Cf | CP096914 |
| P. syringae pv. actinidiae S7-1M | pS7-1M | 68717 | kiwifruit orchard, New Zealand | - | MK621014 |
| Non-conjugative plasmids (Tra-) | |||||
| P. putida | pPp-DIM | 69823 | water, Spain | Sul, Km/Gm, Cb, Tc | LR822050 |
| P.monteilii | unnamed | 60588 | sputum, USA | - | CP014061 |
| P.monteilii STW0522-72 | pSTW0522-72-2 | 59057 | hospital sewage, Japan | - | AP022475 |
| Pseudomonas sp | pPNP1 | 125508 | activated sludge, USA | Sul, 4-Nitrophenol degradation | CP073662 |
1 Resistance to: Hg compounds (Hg), streptomycin (Str), sulphonylamides (Sul), gentamicin (Gm), kanamycin (Km), carbapenems (Cb), cefalosporins (Cf), tetracycline (Tc).
Table 2.
Frequency of conjugative transfer of pPPUT-Tik1-1 into Pseudomonas strains from different phylogenetic groups.
Table 2.
Frequency of conjugative transfer of pPPUT-Tik1-1 into Pseudomonas strains from different phylogenetic groups.
| Group1 | Strain | Species (Collection2) | Transconjugants frequency (per recipient)3 |
|---|---|---|---|
| P. aeruginosa | PAO Type | P. aeruginosa (IMG) | <10-9 |
| B-6643 | P. aeruginosa (VKPM) | <10-9 | |
| B-5807 Type | P. aeruginosa (VKPM) | <10-9 | |
| TA37-4 | P. aeruginosa (IMG) | <10-9 | |
| FA8-1 | P. aeruginosa (IMG) | <10-9 | |
| TA40-1 | P. aeruginosa (IMG) | <10-9 | |
| P. resinovorans | B-14151 Type | P. resinovorans (VKPM) | 1.9*10-3 |
| P. stutzeri | B-975 Type | P. stutzeri (VKM) | 2.4*10-6 |
| B-903 | P. stutzeri (VKM) | 3.4*10-4 | |
| P. oleovorans | B-8623 | P. oleovorans (VKPM) | 9.8*10-5 |
| P. straminea | B-14152 Type | P. seleniipraecipitans (VKPM) | <10-9 |
| P.putida | B-4589 Type | P.putida (VKPM) | 3.7*10-3 |
| P. syringae | B-1546 | P. savastanoi (VKM) | 1.2*10-3 |
| P. fluorescens | B-6735 | P. fluorescens (VKPM) | 5.7*10-4 |
| P22-1-2 | P. fluorescens (IMG) | 2.6*10-4 | |
| B14148 Type | P. fluorescens (VKPM) | 4.7*10-3 |
1 According to 24, 25. 2 All-Russian National Collection of Industrial Microorganisms (VKPM), All-Russian Collection of Microorganisms (VKM), the collection of the Institute of Molecular Genetics (IMG). 3 Average of three experiments.
Table 3.
Mercury transposons found in plasmids from pPPUT-Tik1-1 group.
| Strain | Plasmid | mer-operon | Transposon |
|---|---|---|---|
| P. putida Tik1-1 | pPPUT-Tik1-1 | merRTPADE | ΔTn501 |
| P. putida 12969 | p12969-2 | merRTPABGD | ΔTn5041D |
| merRTPCADE | Tn5046 | ||
| P. juntendii 18091276 | pCN1276 | merRTPFADE | Tn512 |
| merRTPCADE | Tn5046 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



