Submitted:
11 July 2023
Posted:
12 July 2023
You are already at the latest version
Abstract
Cumulative exposure to multiple early life stressors are expected to affect behavioral development causing increased susceptibility towards neuropsychiatric disorders. The present study was designed to mimic such conditions in rat model to study behavioral impairments during adolescence and adulthood. Female Wistar rats (n=32; 140-150 gm) were switched to low protein (LP; 8% protein) or Control (20% protein) diet, 15 days prior to conception and then diet regime was maintained throughout the experimental period. Pups born to Control and LP dams were intraperitoneally injected with deltamethrin (DLT-pyrethroid insecticide; 0.7mg/kg body weight; PND 1 to 7) or lipopolysaccharide (LPS-bacterial endotoxin; 0.3mg/kg body weight; PND 3 and 5) or DLT+LPS, on designated days forming 8 experimental groups (Control, LP, Control+LPS, LP+LPS, Control+DLT, LP+DLT, Control+DLT+LPS and LP+DLT+LPS). Neurobehavioral assessments were performed in F1 rats (1, 3, 6 months) by Open field, Elevated plus maze, Light and Dark box, and Rotarod tests. LP rats were found to be more susceptible to either singular or cumulative exposure as compared to their age matched control counterparts, showing significantly severe behavioural abnormalities, as hyperactivity, attention deficits, impulsiveness and low anxiety, the hallmark symptoms of neuropsychiatric disorders like schizophrenia and ADHD, suggesting thereby that early life multi-hit exposure may predispose individuals to developmental disorders.
Keywords:
Multi-hit stress
; Protein malnourishment
; Low anxiety
; Impulsiveness
; Schizophrenia
; ADHD
1. Introduction
Perinatal stressors most notably protein malnutrition, infections and neurotoxicant exposure have been implicated as risk factors for developing brain, leading to cognitive, behavioral and emotional impairments in animals and humans, and raising susceptibility to neuropsychiatric and neurodegenerative illnesses later in life [1,2]. Exposure to a wide variety of environmental factors during perinatal life has been variously suggested to cause an adequate increase in the chances of developing schizophrenia [3]. The early stages of life are marked by rapid brain growth and a dynamic process of synapse sculpting and pruning, making this time particularly prone to harmful disruptions [4]. Moreover, the exposure of stress inducing factors during early life are also known to result in resilient or maladaptive behaviour in human including anxiety or reduced executive functions due to failure to optimise brain circuit refinement [5]. Clinical investigations have revealed how exposure to environmental stressors throughout the developmental years of childhood and adolescence have a significant impact on cognition later in life. Schizophrenia is a key example that is related due to a stressful early life environment in numerous ways, particularly when multiple types of stressors (multi-hit), such as exposure to prenatal and postnatal malnutrition [6,7], viral and bacterial infections, trauma, and social maltreatment and neurotoxins [8,9] act in variable combinations, and profoundly disrupt the brain development, thus may increase many fold the pathophysiology of schizophrenia in affected individuals. Moreover, maternal inflammatory reactions during pregnancy [10], antenatal infection in preterm newborns [11] and neonatal infection [12] have also been linked to a lower intelligence quotient later in life, as well as to an increased chances of developing neuropsychiatric disorders such as schizophrenia and depression [13]. A variety of genetic and environmental variables are likely to interact throughout the course of schizophrenia, resulting in the development, maintenance, and evolution of the disease symptoms, but determining the exact origin of the disease is challenging [14,15].
Epidemiological data suggests that maternal malnutrition, a nutritional stress has long been associated with changes in neurodevelopment, physical growth indices, and brain structure [16,17]. Furthermore, the experimental studies conducted in rodents indicate that protein malnutrition affect genesis, migration, plasticity and differentiation of neurons and glial cells during the critical periods of brain development (18–20] leading to risk assessment behavioral outcomes in later life [21,22,23]. Changes in cell proliferation, differentiation, migration, synaptogenesis and dendritic arborization are key factors in impaired neurocognitive development. Protein malnutrition has also been linked to an increased risk of neurodevelopmental and metabolic abnormalities in offspring, including autism spectrum disorder (ASD), attention deficit hyperactivity disorder (ADHD) and schizophrenia [24,25]. This is thought to happen through disrupted neuroendocrine system, hypothalamic circuitry, and neuroimmune communication in the developing brain through increased foetal proinflammatory cytokine and chemokine levels.
Experimental studies have also revealed behavioral and cognitive deficits, as well as physical development retardation and poor motor coordination in animal models following exposure to stressors such as maternal and early life infections, perinatal protein and protein calorie malnutrition [22,26]. Moreover, perinatal protein malnutrition is also known to cause low anxiety and/or impulsiveness in rodents. Such effects were directly proportional to the length of the period of malnutrition and the longer exposure to deficient diet lead to more and long-lasting anxiolytic-like effects and higher impulsiveness [18,27].
Furthermore, the unhealthy lifestyle in which the children grow, lead them to encounter viral and bacterial infections, which are common problems in already malnourished children especially from low socio-economic background due to the compromised immune system [28]. Perinatal immune activation by viral and bacterial infections can negatively impact the ongoing brain development and may enhance the risk of developing schizophrenia and other developmental disorders [29,30,31]. Lipopolysaccharide (LPS) is a Gram-negative bacterial cell wall component that promotes neuronal death, inhibits neurogenesis and reduces synaptic plasticity and memory by alleviating the levels of brain-derived neurotrophic factor (BDNF) in hippocampus, parietal and temporal cortex [32,33]. LPS- induced peripheral infection signals to the brain and cause microglial activation and extensive production of inflammatory cytokines leading to behavioral pathology and cognitive impairment [34,35]. Prenatal exposure of LPS has been reported to alter developmental trajectories in neuron-microglia communication in the brains of young offspring along with the imbalance in offspring immune system. These prenatally exposed animals exhibit behavioral changes resembling schizophrenia like phenotype at adulthood [36]. LPS- induced inflammation has also been associated with increased chances of anxiety disorders [37] and major depressive disorders [38].
Deltamethrin (DLT), a type II synthetic pyrethroid insecticide (cyano group at carboxyl α- position) is largely used in agricultural, medical and domestic applications all over the world for more than 30 years [39,40]. Deltamethrin has also been reported as a potential neurotoxicant and the epidemiological data suggest a clear link between the early life exposure of pyrethroid insecticides and neurodevelopmental disorders viz., ADHD, ASD and developmental delay suggesting that deltamethrin is developmentally neurotoxic [40,41]. Developing brain is highly susceptible to such insults leading to many potentially and permanently altered events and adverse long term effects on behavioral and cognitive abilities of an individual. Thus the pyrethroids exposure impose the public health concern and the neurobehavioral performance deficiencies include decreased motor activity, including impaired schedule regulated operant response, lower grip strength uncoordinated movements, anxiety, fear, and nervousness along with cognitive impairments including the performance in Y- and Morris Water Maze [42,43,44]. Occupational pesticide exposure has also been related to Parkinson’s diseases (PD) [45], Alzheimer’s disease (AD) [46], amyotrophic lateral sclerosis (ALS) [47], dementia and cognitive impairments in several other diseases [48]. Other functions affected include delayed perceptual thinking and linguistic comprehension levels in children, as well as early growth and development impairments [49].
Thus the adverse early life environment greatly contribute to multiple behavioral and cognitive abnormalities because of compromised brain circuit development and refinement. As maternal PMN is one of the prime stressor used in this study, the F1 pups born to nutritionally challenged mothers remain highly susceptible to infections and other environmental exposures. Thus the concept of the multi-hit in the current context arises because of the fact that the early life environment is susceptible to numerous stressors and potential of several types of stressors acting synergistically is extremely high both during pre- and postnatal development of the brain.
Thus it is crucial to understand how the cumulative interplay of protein malnutrition, bacterial infection and a neurotoxicant pesticide will affect the behavioral development in adolescence and adult rats in terms of locomotor activity and anxiety status as well as to focus, if there was any relationship between the cumulative multiple stressor exposure on the development of neurological disorders.
2. Materials and Methodology
Wistar albino rats used in the study were maintained under standard animal house facility of School of Studies in Neuroscience, Jiwaji University, Gwalior, in groups of 3 rats/cage (polypropylene cages; 52 cm x 28cm x 22cm) with clean and dust free husk bedding. The animals were maintained at a controlled temperature (25±2° C) and humidity (50-65%) with fixed 12:12h light dark cycle and ad libitum food and water. The maintenance of animal house was carried out as per the approved conditions and requirements of the Institutional Animal Ethics Committee of Jiwaji University, Gwalior (M.P), India.
Fresh three month age virgin female rats (n= 32; body weight 140-150 gm) were selected from the breeding colony and shifted to control (20% protein; n=16) and LP (8% protein; n=16) diets for 15 days prior to mating to acquire protein malnourished female rats before conception. Following acclimatization to the diet regimes, the females were put for mating with healthy male rats in a ratio of 2:1. The pregnancy was confirmed through vaginal smear test followed by increased body weights. Males were separated and the pregnant females were maintained on their respective diets throughout gestation and lactation. The day of birth was marked as postnatal day 0 for F1 pups. Both litter weight and size were recorded, pups were properly monitored for any abnormality and reared with their mothers in aseptic conditions. Litter size was adjusted to eight per dam to prevent dissimilarity among the groups due to different litter size. Both the Control and LP diets were designed and procured from National Institute of Nutrition (NIN) Hyderabad, India.
2.1. Experimental Groups
The F1 pups obtained from both Control and LP mothers were further grouped into eight groups (equal number of males and females), depending on the type of exposure (Figure 1).
Control, Control+LPS, Control+DLT and Control+DLT+LPS groups:
Control group: F1 pups (n=12; from four different dams) born to females fed with 20% protein diet were considered as controls and used to assess various behavioral abilities at the age of 1, 3 and 6 months.
Control rats exposed to bacterial mimetic LPS (Control+LPS group): A set of control F1 pups (n=12; from four different dams) were exposed to LPS at the dose of 0.3mg/kg body weight intraperitoneally on PND 3 followed by a booster dose on PND 5 and used to assess various behavioral abilities at the age of 1, 3 and 6 months.
Control rat pups exposed to deltamethrin (Control+DLT group): Another set of Control F1 pups (n=12; from four different dams) were exposed to DLT at the daily dose of 0.7mg/kg body weight, intraperitoneally from PND 1-7 and used to assess various behavioral abilities at the age of 1, 3 and 6 months.
Control F1 rat pups exposed to both LPS and DLT (Control+DLT+LPS group): Another set of Control F1 pups (n=12; from four different dams) were exposed to both DLT and LPS at above specified time points and doses and used to assess various behavioral abilities at the age of 1, 3 and 6 months.
Protein malnourished (PMN) groups:
PMN (LP) group: LP F1 pups (n=12; from four different dams) born to females fed with 8% protein diet and maintained on the same diet throughout the experimental period were considered as LP/PMN group. These LP F1 rats were maintained on LP diet and assessed for various behavioral abilities at the age of 1, 3 and 6 months.
LP F1 rat pups exposed to bacterial mimetic LPS (LP+LPS double-hit group): LP F1 pups (n=12 from four different dams) were exposed to LPS at the dose of 0.3mg/kg body weight, intraperitoneally on PND 3 followed by a booster dose on PND 5. These LP+LPS rats were maintained on LP diet and assessed for various behavioral abilities at the age of 1, 3 and 6 months.
LP F1 rat pups exposed to deltamethrin (LP+DLT double-hit group): Another set of LP F1 pups (n=12; from four different dams) were exposed to DLT at a daily dose of 0.7mg/kg body weight intraperitoneally from PND 1-7. These LP+DLT rats were maintained on LP diet and assessed for various behavioral abilities at the age of 1, 3 and 6 months.
LP F1 rat pups exposed to both LPS and DLT (LP+DLT+LPS multi-hit group): A third set of LP F1 pups (n=12; from four different dams) were exposed to both DLT and LPS at above specified time points and doses. These LP+DLT+LPS rats were maintained on LP diet and assessed for various behavioral abilities at the age of 1, 3 and 6 months.
Both LPS (D9315-10MG) and DLT (E. coli, serotype O11:B4) were procured from Sigma Aldrich. The LPS solution was prepared by dissolving 0.3mg of LPS in 1 ml of sterilized phosphate-buffer saline (PBS), while DLT was dissolved in dimethyl-sulphoxide (DMSO) to obtain 1mg/ml stock solution.
To perform low volume and error free injection of DLT and LPS at a steady flow rate of 30µl/mint and to confirm absolute absorption, Stoelting Nanoinjector and Hamilton micro-syringe were used under hygienic settings. To counter any bias, Control animals were injected with the vehicle alone. No anaesthetic procedure was followed during injection, rather pups were gently handled during the delivery of injection and then quickly transferred to their respective dams to minimize the separation stress from mother. To overcome sex specific differences, equal number of males and females were used in the study as post hoc analysis did not bring any sex specific variations during data analysis.
2.2. Behavioural Studies
Specific behavioural abilities were assessed in F1 generation rats (n=12/group/time point, consisting of 6 males and 6 females from different dams) of all the groups (Control, LP, Control+LPS, LP+LPS, Control+DLT, LP+DLT, Control+LPS+DLT and LP+DLT+LPS) at the age of 1, 3 and 6 months.
2.2.1. Open Field Test
To quantify locomotor activity in F1 rats and track the animal path, Autotrack system (Columbus Instruments, OH, USA, v4.41) was used. The system tracks the motion of the animal with the help of infrared photocells placed around the open field arena made of transparent acrylic box (43x43x22 cm) connected to the computer interface with preinstalled Optovarimax Autotrack software version 4.4. The test animals were subjected to the testing cage of the activity monitor, 5 minutes prior to the final data recording to avoid the novelty induced exploration, followed by recording of the locomotor behaviour for 20 minutes test session. The test arena was cleaned with 70% alcohol after every trail to avoid the interference from animal’s odors i.e., urination and defecation due to presence of pheromones. The parameters recorded include the total distance travelled (DT), resting time (RT), stereotypic time (ST), ambulatory time (AT), horizontal count (HC) and center zone activity (center zone entries and time) as indicative of spontaneous motor activity as described previously [22,30,50]. From each open field track report, time in square analysis was conducted, animals spending more time in the centre zone was considered as indicative of low anxiety. The amount of time an animal spends engaging in stereotypic behaviour such as abnormal repeated scratching, rearing and grooming that breaks a single optical beam is known as stereotypic time. To minimize the circadian cycle effect on open field behaviour, testing of all the animals was performed between 9:00 AM to 5:30 PM.
2.2.2. Elevated Plus Maze
To assess the anxiety and fear behaviour in F1 generation rats, elevated plus maze elevated 50 cm above the base, consisting of two opposite closed arms (with 15cm high walls) and two opposite open arms (without walls) was used. This forms a setup of four arms originating from a common point called center zone. Before starting the test, the rats were acclimatized for 45 minutes to the experimental room. For testing, the animal was placed in the center zone facing one of the open arms and exploratory behaviour was recorded by preinstalled real-time video tracking system camera placed above the apparatus and attached to a computer with ANY- maze software (Columbus instruments, USA, v4.82) for 2 minutes with three trials/animal with a gap of 2h between trails. The parameters of time spent and number of entries to the open and closed arms [22,30,50], center zone time and entries and total open arm entries were automatically recorded and tracked by the ANY-maze software (Columbus Instruments, USA). The data was finally analyzed and expressed as percent time spent, percent entries in the open arms; time and entries in the center zone and total open arm entries. The percent time spent and number of entries (%) in open arms was calculated by the following formula: time spent in open arms/total test time × 100; entries in open arms/total entries × 100 respectively.
2.2.3. Light and Dark Test
To assess the anxiety and depression like behaviour in F1 rats, light and dark test was performed. This test is used to analyse the animal’s reaction to the aversion and anxiety prone context of being in brightly illuminated areas. The apparatus consists of two compartments; the light compartment is 2/3 of the box and is brightly lit and open, while the dark compartment is 1/3 of the box and is covered and dark. Both the compartments are connected by small doorway with opening of 7 cm diameter. Rats were placed in the dark compartment and allowed 5 min to explore freely. To avoid a bias produced by novelty induced exploration, the first 1 min of data was excluded from the analysis. To record and analyse the rats activity automatically i.e., time spent and number of entries in each chamber [50], an overhead real- time video tracking system camera was mounted on the ceiling focused on the middle of the apparatus box and attached to the computer set up with preinstalled ANY- maze software version v4.82 (Stoelting, USA). The apparatus inner arena was cleaned with 70% alcohol after each animal testing, to avoid biasing with the previous tested animal left due to urination and defecation. The data was finally analyzed and expressed as percent time spent and entries in the light chamber. All the procedures were performed between 9:00 AM to 5:30 PM to minimize the influence of possible circadian changes on the behaviour. The percent time spent and number of entries (%) in light zone was calculated by the following formula: time spent in the light zone/total test time × 100; entries in light zone/total entries × 100 respectively.
2.2.4. Rotarod Test
Rotarod was used to assess the neuromuscular coordination in terms of motor coordination and motor learning in F1 generation rats as their ability to balance on an accelerating rotating rod. The apparatus consists of 40 cm long rubber coated horizontal metal rod (3-inch diameter) attached with a motor through which the speed of the rotating rod is maintained. It is divided into 4 sections, separated by 10 cm long plastic discs to enable testing of 4 animals at once. Rotarod was operated with a computer interface installed with Rotamex version 1.2.3 software. The activity of the rat was detected with the infrared sensors (emitter and detector) attached to the instrument as the rat run on the rotating rod with any particular rpm selected from the software. The test procedure consisted of two periods. The first period was acclimatization in which rats were acclimatized for 3 days with three trials per day per animal (Start speed: 2 rpm, Maximum speed: 8 rpm, duration: 100 seconds). Another period was retention period/final day test which was performed after 24 hours of last acclimatization with a starting speed of 2 rpm and maximum speed of 40 rpm for 420 seconds. The latency to fall (off) from the accelerating rod (during the provided experimental time) was automatically detected by the infrared sensors and recorded using the Rotamex software [50]. All the animals were tested on the same day from 9:00 AM to 5:30 PM in order to minimize the influence of possible circadian changes on the behaviour.
2.2.5. Statistical Analysis:
Results were expressed as standard error of mean (± SEM). The standard statistical software Sigma Stat 13.5 was used for all the statistical analyses. The data was analyzed using one-way (ANOVA) for (group wise comparison) and two-way (ANOVA) (for comparison between groups with two independent variables, i.e., diet and infection) followed by post hoc Holm-Sidak test. Significance level was set at a P- values of ≤ 0.001 for highly significant and ≤ 0.05 were considered as significant.
3. Results
3.1. Physical Development
No statistically significant difference was noticed in the appearance of developmental physical landmarks including ear pinna detachment, eye opening, incisor eruption, vaginal opening and testes descent amongst all the eight groups. However, LP and LP exposed group animals (LP+LPS; LP+DLT; LP+DLT+LPS) displayed drastic fur loss with stunted body growth whereas the control group and control treated animals revealed healthy physical appearance in terms of both hair and body growth.
Body weight (gm)
The F1 offspring of LP mothers exhibited a clear compromise in physical growth, with lower body weight with age as compared to the controls. Furthermore, exposure with either LPS or DLT (LP+LPS and LP+DLT) or cumulative exposure of both LPS and DLT to LP rats (LP+DLT+LPS) showed further significant reduced body weight at the age of 1 and 3 months as compared to age matched control and LP alone animals. However, LP+DLT+LPS rats also revealed significant reduced body weight as compared to age matched control and control treated rats at 6 months of age. Furthermore, body weight of the LP treated group rats also remained significantly low as compared to their corresponding age-matched control treated groups (Figure 2A–E). The statistical mean and f-values along with level of significance are given in Table 1.1 and Table 1.2.
3.2. Double Hit and Multi-Hit Exposure of Lipopolysaccharide and Deltamethrin Induced Hyperactivity and Low Anxious Behaviour in Protein Malnourished Rats
Open field analysis demonstrated that maternal protein malnourishment resulted in hyperactivity, motor impulsivity and low anxiety-like behaviour, as revealed by significantly increased DT (Figure 3B); ST (Figure 3C); AT (Figure 3D); HC (Figure 3E); center zone entries (3F); center zone time (3G) and a significantly decreased resting time (Figure 3A) in LP rats at the age of 1, 3 and 6 months as compared to the age matched control groups showing the effects of maternal protein malnutrition. Exposure with either LPS or DLT (LP+LPS and LP+DLT) or cumulative exposure of both LPS and DLT to LP rats (LP+LPS+DLT) resulted in further exaggeration in the hyperactivity, motor impulsivity and anxiety-like behaviour with a highly significant increase in DT; AT; ST and HC at the age of 1, 3 and 6 months as compared to the age matched control and LP alone animals. Such hyperactivity, impulsivity and low anxiety behaviour was also observed in control animals exposed to either LPS or DLT or both LPS+DLT but the mean values for DT; AT; ST and HC remained comparatively low as compared to their respective LP groups. Time in square analysis from open field tracks showed that LP multi-hit rats (LP+DLT+LPS) spent significantly more time in the center zone with increased center zone entries as compared to age matched control and LP alone animals. The locomotive hyperactivity behaviour was highly pronounced at 3 months of age and the animals remained hyperactive and anxious throughout their life, i.e., by 6 months of age studied in this investigation.
Open field track reports also confirmed the above data showing the low anxiety like behavioral profile and locomotive hyperactivity in LP and LP treated animals compared to age matched controls which showed normal exploratory behaviour (Figure 4a,e,i). Combo exposure of LPS and DLT to both control and LP group animals, further reduced the anxiety levels (Figure 4b–d,f–h,j–l) comparatively more in LP treated animals (Figure 4n,o,p,r,s,t,v,w,x), with very frequent haphazard center zone arena exploration. Furthermore, multi-hit animals showed drastic behavioral impairments accompanied with irregular track activity with maximum periphery exploration/wall clinging called thigmotaxis and increased time spent in the center zone and corners showing severe hyperactivity, motor impulsivity or low anxiety and fearless behaviour. Additionally, the dense area in the centre zone depicts stereotyped repetitive rearing and horizontal back and forth movements revealed by multi-hit animals further suggests their low-anxiety profile. The statistical mean and f-values along with level of significance are given in Table 2.1, Table 2.2, Table 2.3, Table 2.4, Table 2.5, Table 2.6, Table 2.7, Table 2.8, Table 2.9, Table 2.10, Table 2.11, Table 2.12, Table 2.13 and Table 2.14.
3.3. Elevated Plus Maze Test (EPM) Revealed Low Anxiety Phenotype in Protein Malnourished Rats Treated with Lipopolysaccharide and Deltamethrin
Elevated plus maze (EPM) test also confirmed the low anxiety behaviour following LPS and DLT exposure to LP rats. Data analysis from the EPM test revealed that LP animals spent significantly more time in open arms (% open arm time; Figure 5B) along with significantly increased number of open arm entries (% open arm entries) (Figure 5A). In addition, both the time spent in center zone and center zone entries (Figure 5C,D) were also significantly more in LP animals. However, upon single or combined treatment of LPS and DLT to LP animals, there was a further significant increase in both % open arm entries and % time spent in open arms, as well as the time spent in center zone and center zone entries with the maximum increase in multi-hit treatment group (LP+LPS+DLT) demonstrating highly hyperactive, motor impulsive and low anxiety like phenotype as compared to age matched control animals. The control animals also responded similarly to LPS or DLT or LPS+DLT treatments with a significant increase in % open arm entries and % open arm time as well as time in center zone and center zone entries at 1, 3 and 6 months, but to a lower degree as compared to their corresponding LP group rats. The number of open arm entries increased gradually with complexity of exposure, being highest in the LP multi-hit group (Figure 5E). Such behaviour persisted consistently till the age of 6 months as studied in this investigation. The data showing the mean and the f- values are tabulated in Table 3.1, Table 3.2, Table 3.3, Table 3.4, Table 3.5, Table 3.6, Table 3.7 and Table 3.8.
Elevated plus maze track reports also confirmed the above data showing the low anxiety like behavioral profile in LP and LP treated animals as compared to the respective age matched control which staying away from the open arms edges (Figure 6 a,i, q). Combo exposure of LPS and DLT to both control (Control+LPS; Control+DLT; Control+DLT+LPS) and LP (LP+LPS; LP+DLT; LP+DLT+LPS) group animals, further reduced the anxiety levels (Figure 6 b, c, d, j, k, l, r, s, t), showing highly anxious profile in LP treated animals (Figure 6 f, g, h, n, o, p, v, w, x), with very frequent open arm and center zone entries. Moreover, the multi-hit group rats frequently explored the open-arms till the extreme distal ends depicting the hyperactivity, motor impulsivity or low-fear behaviour (Figure 6 d, l, t, h, p, x).
3.4. Light and Dark Box Test also Revealed the Low Anxiety Behaviour in Protein Malnourished Rats Treated with LPS and DLT
OFT and EPM results, were further supported by light and dark box test. Data assessment from histograms of LD box revealed that LP animals showed significantly increased % light zone entries (Figure 7A) and % light zone time at 1, 3 and 6 months (Figure 7B) which indicate the low anxiety like behaviour with an increased tendency to explore the light zone in contrast to the biased normal nocturnal behaviour of rats shown by age matched control animals, revealing the impact of maternal protein malnutrition. Further exposure of LPS and DLT or both LPS+DLT to the control and LP F1 rats caused a sharp increase in both the % light zone entries and % light zone time, suggesting the impact of such double or multi-hit exposure in inducing motor impulsivity and low anxiety. However, these changes were significantly higher in the LP multi-hit treated group, which indicate their higher susceptibility to developing such phenotypes.
Light and dark test track reports of animals also supported the above data showing low anxiety profile in LP and LP treated animals as compared to age matched control animals (Figure 7C a, i, q). Combo or multi-hit exposure of LPS and DLT in both Control (Control+LPS; Control+DLT; Control+DLT+LPS) and LP (LP+LPS; LP+DLT; LP+DLT+LPS) group animals further showed reduced anxiety levels (Figure 7C b, c, d, j, k, l, r, s, t), comparatively more in LP treated animals (Figure 7C f, g, n, o, v, w) with irregular, random locomotor activity due to frequent entries into light compartment. Additionally, double or multi-hit animals depicted immobility at the doorway opening towards light compartment thus suggesting freezing behaviour (Figure 7C c, f, h, l, p, s, t, w, x). The statistical mean and f-values along with level of significance are tabulated in Table 4.1, Table 4.2, Table 4.3 and Table 4.4.
3.5. Cumulative Exposure of LPS and DLT to Protein Malnourished (Multi-Hit) Rats Resulted in Hyperlocomotion and Motor Impulsivity Phenotype
Rotarod test was used to evaluate the motor coordination. Interestingly LP animals showed significantly increased latency to fall off from accelerating rod of the rotarod at the age of 3 and 6 months as compared to age matched control animals indicating increased stereotyped behaviour in LP animals which tend them to keep moving doing abnormal activities. Upon single or combined exposure of LPS and DLT to LP animals (LP+LPS, LP+DLT and LP+DLT+LPS) there was further significant increase in latency to fall both at the age of 3 and 6 months. However, the multi-hit (LP+DLT+LPS) group rats were able to retain themselves for the maximum time on the accelerating rod showing highest latency to fall off both at 3 and 6 months of age which reveal increased hyperactivity, motor impulsivity and stereotyped behaviour when compared to age matched control group. Such behavioral deficits were also significantly observed in Control+LPS, Control+DLT and Control+DLT+LPS group at 3 and 6 months of age when compared to age matched normal control group. Such significantly higher retention time on rotating rotarod by the double and multi-hit group rats indicate impulsiveness due to increased hyperactivity and stereotyped behaviour. The statistical mean and f-values along with level of significance are mentioned in Figure 8 and Table 5.1 and Table 5.2.
4. Discussion
Perinatal stress has been widely documented as a cause of neurological illnesses such as schizophrenia, ADHD, AD, PD and many others [51,52,53]. Early childhood is a crucial and vulnerable period when environmental impressions on later life are formed. Several research investigations have concentrated on the dual-hit hypothesis, which states that a first genetic hit during critical developmental periods increases the probability of a second-hit, which is usually an environmental insult such as infections, nutritional inadequacy or exposure to neurotoxicant [30,54,55]. Due to the large spectrum of stressors and their variable modes of action, the dual-hit hypothesis alone is insufficient to establish a relation between stressors and neurological diseases. The present research link early life multi-hit stressors i.e., perinatal PMN and neonatal bacterial infection and pyrethroid (neurotoxicant) exposure to the early onset symptoms of neurological disease like schizophrenia and ADHD.
The severity of neurological problems is directly associated with loss of body weight of an individual as it can increase the risk of systemic infections and may also raise chances of morbidity and mortality in later life [56,57]. In this investigation we have recorded a persistent and significant loss of body weight of LP and LP treated rats by the age of 1,3 and 6 months as compared to control rats, while in double-hit group rats (LP+LPS and LP+DLT) there was further reduction in body weight at all the three time points studied as compared to control and control treated animals. Additionally, the cumulative exposure of all these early life stressors (LP+DLT+LPS) drastically deteriorated the body weight in multi-hit group animals. However, by the age of 6 months single or multi-hit exposed control and LP rats showed increased body weight as compared to their respective controls. This change may be due to persistent overactivation of hypothalamic-pituitary-adrenal (HPA) axis responses towards early life stressors [58], contributing to development of metabolic dysregulation [59] reported in line with studies in animal models [60] and human subjects [61]. Considering the severity of the protein restriction, a drop in body weight in PMN group rats was expected as protein deprivation frequently leads to emaciation due to shrinkage of body size, as also reported earlier from our lab [22,23]. Following singular or multi-hits LP F1 rats showed further decrease in body weight because they responded more aggressively to a subsequent cumulative stress exposure. There is a direct link between immune system and brain development. Both the brain and the immune system are not fully formed at birth, but rather continue to mature in response to the postnatal environment making it susceptible to early life stressors and in turn affecting the brain functions at adulthood [62]. In neonatal age, a drop in body weight after any infection is probably an indication of sickness behaviour [63] as reported both in humans [64] and animal models [65,66]. The population-based birth cohort studies have also found a link between low birth weight and many neuropsychological deficits such as schizophrenia [67,68] and ADHD [69,70]. All these studies, prompted us to suggest that compromised physical growth and immune system following cumulative multi-hits may predispose the individuals more prone to develop neuropsychiatric deficits, simulating schizophrenia and ADHD.
To further investigate how the cumulative interplay impact of all these stressors affect the behaviour of multi-hit rats, a battery of behavioral tests were used to assess the attention deficit and hyperactivity behaviour, anxiety-like and motor impulsivity related phenotype. The open field test, frequently used to assess rodent locomotion, anxiety and stereotyped behaviour including grooming and rearing [71,72], demonstrated that maternal PMN resulted in hyperactivity and stereotyped behaviour with low anxiety-like behavioral phenotype, in terms of significant increase in DT, ST, AT, HC, center zone entries and center zone time. In addition, there was a significant decrease in RT suggesting the restless behaviour. Such changes were further pronounced following exposure to LPS or DLT or cumulative exposure of both LPS+DLT to LP F1 rats, showing severe behavioral deficits with frequent central zone exploration as shown by increase in center zone entries and time depicting hyper-locomotion and impulsive jumping against the wall of the open field arena or wall-hugging as a stereotyped behaviour, which persisted through adolescence and adulthood. In addition to locomotion, the OFT test also revealed the level of the anxiety measured as the fraction of time spent in the perimeter vs the center zone. Increased peripheral zone activity (thigmotaxis) is considered as higher anxiety, while the more center zone exploration as low anxiety as the rodents have a natural tendency to avoid the open center [73]. In addition, the increased horizontal and vertical counts suggests increased exploratory activity [74,75]. Stereotypies or the repetitive behaviour are the repetitions of certain motor patterns with no apparent goals or functions. Such stereotypic behaviour like scratching, licking of head, neck and trunk and other grooming activities were persistent through adolescence and adulthood in LP F1 rats exposed to a combination of both LPS and DLT. This coincides with the theory of being unable to translate cognition into actions. Thus, LP multi-hit rats lose self-control mechanism such as control of attention or emotions because they have lost chain between knowledge and actions and lack the capacity of refraining themselves from acting impulsively [76,77]. In addition the hyperactivity and thigmotaxis also indicate the stereotypic activity traversing the same locomotor trajectory repeatedly [78]. Although the equivalence of rodents and human repetitive behaviour is variously argued, the probability of repetitive behaviour and increased self-grooming in rodents is directly correlated with hyperactivity and impulsivity and may also indicate to impulsive aggression which has long been linked with obsessive compulsive disorder (OCD). Grooming in rodents has become a useful strategy for modelling different mood and psychiatric disorders and to understand neural circuitries underlying complex motor patterns [79]. Thus the increased self-grooming in rodents indicating complex repetitive, self-directed and sequentially patterned behaviours may indirectly correlate to human brain disorders including chain of motor actions and complex patterning of motor activities [79]. Moreover, the anxiety like states also alter rodent self-grooming and its sequencing [80,81]. The increased self-grooming in rodents has been modelled with disease symptoms in OCD [82,83], ASD [84,85], anxiety and panic disorders and schizophrenia [86,87]. Interestingly the LP F1 rats from early adolescence through late adulthood, exhibited increased central time duration in addition to overall increased locomotion, indicating hyperactivity and impulsivity symptoms that are typical of both ADHD [88] and schizophrenic [89,90] patients. This suggests that increased movement and stereotypic behaviour in LP and LP multi-hit group rats is indicative of their inability to adapt to novel conditions or signs of inadequate habituation [91,92]. When tested with rotarod, these single and multi-hit animals maintained themselves on the accelerating rod of the rotarod for longer time showing significantly increased latency to fall-off when compared with the control animals. Such higher motor coordination behaviour of the multi-hit animals might indicate their motor impulsivity phenotype and increased motor stereotyped behaviour which tend them to keep moving abnormally. The motor stereotypies have been described as complex, repetitive, rhythmic, often bilateral movements with a typical onset in early childhood [93]. Such movements in the multi-hit rats may be the resultant of hyperactivity and low anxiety and fearless behaviours as revealed by open field test, elevated plus maze and light and dark test. Over time, the emotional responses (anxiety and impulsive-like response) may become dissipated and replaced by hyperactivity and stereotyped responses. Here motor stereotypy refers to an excessive repetition of one type of motor response [94], i.e., prolonged maintenance on the rotating rod of the rotarod and increased latency to fall-off by multi-hit animals. Such exaggerated hyperactivity, impulsiveness and stereotyped behaviour, also indirectly indicate an inattention behaviour, as rats are reluctant to engage in tasks requiring sustained attention. Thus typical behaviour shown by multi-hit rats is also seen in many neuropsychiatric conditions, such as schizophrenia [95,96], OCD [97], autism spectrum disorder (ASD) and ADHD [98,99]. Recently, exposure to DLT alone has been linked to long-term neurobehavioral deficits in rodents when exposed postnatally [41] or in offspring when exposed during pregnancy [100]. Locomotor hyperactivity, increased impulsivity, attention deficits and elevated dopamine transporter and receptor levels commonly reported following DLT exposure are also typically seen in children with ADHD [100]. Studies examined with exposure to maternal protein deficiency or new-born viral or bacterial infections have similarly demonstrated impairment in behaviour as measured by OFT paradigm [22,23,30,101,102] consistent to what has been observed in present study.
These OFT results were further supported by EPM data, another commonly used test to assess anxiety-like phenotype by using innate fear for height in rodents which stand in conflict with curiosity and a drive for spontaneous exploration [103,104]. Higher degree of anxiety is often indicated by more frequent entries and time spent in the closed arms of EPM. The open arms are symbolised to represent anxiety-provoking environment [72,103,105]. Rodents naturally favour closed arms and in a typical 10-min experiment, normal mice spend majority of their time in the enclosed arms [105,106]. However cumulative exposure of DLT and LPS stressors was found to heighten the hyperactive, low-anxiety and fearless phenotype as multi-hit LP animals spent significantly increased % time with increased % entries and significantly more center zone time and entries in open arms of EPM. Such changes increase gradually with complexity of exposure and age, revealing that perinatal stressors lead to prolonged and consistent behavioral changes through adolescence to adulthood as also seen in most of the schizophrenic patients. Low anxiety phenotype was also reported following single-hit exposure in rats [22,23] and exposure of viral and bacterial infection in rats [30] and humans [107,108]. Additionally, mouse models of tauopathy [109] and bisphenol A exposure [110] have also been implicated to replicate hyperactivity and reduced anxiety like phenotype.
To a greater degree of assessing unconditional anxiety responses in rodents, the light and dark zone exploration test designed by [111] was used that served as a precursor to the EPM. Similar to the EPM, the subject is placed in an open arena that includes protected (dark compartment) and unprotected (bright compartment) areas with free access between the two [112]. Most rodents exhibit a natural affinity for protected, dark spaces [72,107,113] and increased avoidance of the light chamber is interpreted as increased anxiety [114]. However, increased time spent in the light zone or shorter entry delays are interpreted as signs of anxiolytic behaviour or decreased anxiety-like phenotype [111,115]. Light and dark box also revealed hyperactivity, low anxiety and impulsivity like behaviour as multi-hit LP rats showed significant increase in both the % light zone entries and % light zone time and also increased tendency to explore the light zone in contrast to the normal nocturnal behaviour mimicked as dark zone exploration shown by age matched control and control treated animals. This could be because of the robust immune activation that makes these multi-hit rats highly susceptible to develop such phenotypes. Individuals with schizophrenia also have active innate immune system which triggers TLR, inflammasome and apoptotic pathways [116] that enhance the risk of schizophrenia like behaviour in later stages of life. The increased time spent on the light side of the box or shorter entry delays are interpreted as signs of anxiolytic behaviour or low anxiety-like phenotype [111,115]. The present study thus elucidate the persistent effect of perinatal multi-hit exposure of PMN, LPS and deltamethrin exposure pertained to the decrease in body and brain weight, hyperactivity, impulsivity and low anxiety phenotype which are hallmark symptoms of neuropsychiatric disorders like schizophrenia and ADHD. As the heritability of ADHD and schizophrenia is approximately less than 1% [117], thus environmental influences during prenatal and postnatal development have been considered to be the major concern to play substantial role in causing these diseases as well as dysfunction during later life [118]. Thus, it is suggested that cumulative early life exposure to maternal protein malnutrition, bacterial infection and pesticide exposure may act as crucial risk factors for schizophrenia, ADHD and other neuropsychiatric disorders and may pre-dispose individuals to develop such psychopathologies during adolescence and adulthood.
5. Conclusion
The present results suggests that synergistic exposure of multiple perinatal stressors during early life, may lead to severe behavioural abnormalities, in terms of hyperactivity or attention deficit like symptoms, impulsiveness and low-anxiety phenotype at adulthood, which mimic the pathophysiology of neuropsychiatric disorders like schizophrenia, suggesting thereby that cumulative exposure to numerous early-life stressors enhances the risk of developing schizophrenia and other neuropsychiatric disorders at adulthood.
Author Contributions
The study was ratified and planned by Ishan Kumar Patro. Nisha Patro and Syed Mujtaba performed the experiments, analyzed data and wrote the manuscript. Ishan Kumar Patro and Nisha Patro then finalized the manuscript.
Institutional Review Board Statement
The experimental protocols were pre-approved by the Institutional Animal Ethics Committee (1854/GO/Re/16/CPCSEA) via letter No. JU/IAEC/05.
Acknowledgements
The authors would like to acknowledge Indian Council of Medical Research and Basic Scientific Research- University Grants commission, New Delhi, India for providing grant.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
PMN, protein malnutrition; LP, low protein; DLT, deltamethrin; LPS, lipopolysaccharide; PND, postnatal day; OFT, open field test; EPM, elevated plus maze; LD, light and dark box; ADHD, attention deficit hyperactivity disorder; ASD, autism spectrum disorder; BDNF, brain derived neurotrophic factors; PD, Parkinson’s disease; AD, Alzheimer’s disease; ALS, amyotrophic lateral sclerosis; NIN, National institute of nutrition; PBS, phosphate buffer saline; DMSO, dimethyl-sulphoxide; DT, distance travelled; ST, stereotypic time; AT, ambulatory time; RT, resting time; HC, horizontal count; mRNA, messenger ribonucleic acid; MRI, magnetic resonance imaging; TNF, tumour necrosis factor; OCD, obsessive compulsive disorder; TLR, toll-like receptor.
References
- Nestler, E.J., Hyman, S.E., 2010. Animal models of neuropsychiatric disorders. Nat. Neurosci. 13(10), 1161-1169. [CrossRef]
- Nelson III, C.A., Gabard-Durnam, L.J., 2020. Early adversity and critical periods: neurodevelopmental consequences of violating the expectable environment. Trends Neurosci. 43(3), 133-143. [CrossRef]
- Eyles, D.W., 2021. How do established developmental risk-factors for schizophrenia change the way the brain develops? Transl. Psychiatry. 11(1), 1-15. [CrossRef]
- Stolp, H., Neuhaus, A., Sundramoorthi, R., Molnár, Z., 2012. The long and the short of it: gene and environment interactions during early cortical development and consequences for long-term neurological disease. Front. Psychiatry. (3), 50. [CrossRef]
- Milbocker, K.A., Campbell, T.S., Collins, N., Kim, S., Smith, I.F., Roth, T.L., Klintsova, A.Y., 2021. Glia-driven brain circuit refinement is altered by early-life adversity: Behavioral outcomes. Front. Behav. Neurosci. 2(15), 15:786234. [CrossRef]
- Sarkar, T., Patro, N., Patro, I.K., 2020. Neuronal changes and cognitive deficits in a multi-hit rat model following cumulative impact of early life stressors. Biol. Open. 9(9), bio054130. [CrossRef]
- Onaolapo, O.J., Onaolapo, A.Y., 2021. Nutrition, nutritional deficiencies, and schizophrenia: An association worthy of constant reassessment. World J. Clin. Cases. 9(28), 8295. [CrossRef]
- Sarkar, T., Patro, N., Patro, I.K., 2019. Cumulative multiple early life hits-a potent threat leading to neurological disorders. Brain Res. Bull. 147, 58-68. [CrossRef]
- Teissier, A., Le Magueresse, C., Olusakin, J., Andrade da Costa, B.L., De Stasi, A.M., Bacci, A., Imamura, Kawasawa, Y., Vaidya, V.A., Gaspar, P., 2020. Early-life stress impairs postnatal oligodendrogenesis and adult emotional behaviour through activity-dependent mechanisms. Mol. Psychiatry. 25(6), 1159-1174. [CrossRef]
- Smith, S.E., Li, J., Garbett, K., Mirnics, K., Patterson, P.H., 2007. Maternal immune activation alters fetal brain development through interleukin-6. J. Neurosci. 27(40), 10695-10702. [CrossRef]
- Dammann, O., Kuban, K.C., Leviton, A., 2002. Perinatal infection, fetal inflammatory response, white matter damage, and cognitive limitations in children born preterm. Ment. retard. dev. disabil. res. rev. 8(1), 46-50. [CrossRef]
- Koponen, H., Rantakallio, P., Veijola, J., Jones, P., Jokelainen, J., Isohanni, M., 2004. Childhood central nervous system infections and risk for schizophrenia, Eur Arch Psychiatry Clin Neurosci. 254(1) (2004), 9-13, . [CrossRef]
- Miller, B.J., Culpepper, N., Rapaport, M.H., Buckley, P., 2013. Prenatal inflammation and neurodevelopment in schizophrenia: a review of human studies. Prog. Neuropsychopharmacol. Biol. Psychiatry. (42), 92-100. [CrossRef]
- Akdeniz C., Tost H., Streit, F., Haddad, L., Wüst, S., Schäfer, A., Schneider, M., Rietschel, M., Kirsch, P., Meyer-Lindenberg, A., 2014. Neuroimaging evidence for a role of neural social stress processing in ethnic minority–associated environmental risk. JAMA psychiatry. 71(6), 672-680. [CrossRef]
- Uher, R., 2014. Gene–environment interactions in severe mental illness. Front. Psychiatry. (5), 48. [CrossRef]
- Georgieff, M.K., Ramel, S.E., Cusick, S.E., 2018. Nutritional influences on brain development. Acta Paediatr. 107(8), 1310-1321. [CrossRef]
- Cortés-Albornoz, M.C., García-Guáqueta, D.P., Velez-van-Meerbeke, A., Talero- Gutiérrez, C., 2021. Maternal nutrition and neurodevelopment: A scoping review. Nutrients. 13(10), 3530. [CrossRef]
- Alamy, M., Bengelloun, W.A., 2012. Malnutrition and brain development: an analysis of the effects of inadequate diet during different stages of life in rat, Neurosci. Biobehav. Rev. 36(6), 1463-1480. [CrossRef]
- Naik, A.A., Patro, N., Seth, P., Patro, I.K., 2017. Intra-generational protein malnutrition impairs temporal astrogenesis in rat brain. Biol. Open. 6(7), 931-942. [CrossRef]
- Patro, N., Naik, A.A., Patro, I.K., 2019. Developmental changes in oligodendrocyte genesis, myelination, and associated behavioral dysfunction in a rat model of intra-generational protein malnutrition. Mol. Neurobiol. 56(1), 595-610. [CrossRef]
- Reyes-Castro, L.A., Rodriguez, J.S., Charco, R., Bautista, C.J., Larrea, F., Nathanielsz, P.W., Zambrano, E., 2012. Maternal protein restriction in the rat during pregnancy and/or lactation alters cognitive and anxiety behaviors of female offspring. Int. J. Dev. Neurosci. 30(1), 39-45. [CrossRef]
- Naik, A.A., Patro, I.K. Patro, N., 2015. Slow physical growth, delayed reflex ontogeny, and permanent behavioral as well as cognitive impairments in rats following intra-generational protein malnutrition. Front. Neurosci. 9, 446. [CrossRef]
- Sinha, S., Patro, N., Tiwari, P.K., Patro, I.K., 2020. Maternal Spirulina supplementation during pregnancy and lactation partially prevents oxidative stress, glial activation and neuronal damage in protein malnourished F1 progeny. Neurochem. Int. (141), 104877. [CrossRef]
- Smith, B.L., Reyes, T.M., 2017. Offspring neuroimmune consequences of maternal malnutrition: potential mechanism for behavioral impairments that underlie metabolic and neurodevelopmental disorders. Front. Neuroendocrinol. (47), 109-122. [CrossRef]
- Eyles, D.W., Dean, A.J., 2016. Maternal nutritional deficiencies and schizophrenia: Lessons from animal models with a focus on developmental vitamin D deficiency. In Handbook of Behavioral Neuroscience (Vol. 23, pp. 243-264). Elsevier.
- Laus, M.F., Vales, L.D.M.F., Costa, T.M.B., Almeida, S.S., 2011. Early postnatal protein-calorie malnutrition and cognition: a review of human and animal studies. Int. J. Environ. Res. Public Health. 8(2) (2011), 590-612. [CrossRef]
- Françolin-Silva, A.L., da Silva Hernandes, A., Fukuda, M.T.H., Valadares, C.T., Almeida, S.S., 2006. Anxiolytic-like effects of short-term postnatal protein malnutrition in the elevated plus-maze test. Behav. Brain Res. 173(2), 310-314. [CrossRef]
- Walson, J.L., Berkley, J.A., 2018. The impact of malnutrition on childhood infections. Curr. Opin. Infect. Dis. 31(3), 231. [CrossRef]
- Meyer, U. 2014. Prenatal poly (i: C) exposure and other developmental immune activation models in rodent systems. Biol. Psychiatry, 75(4), 307-315. [CrossRef]
- Sarkar, T., Patro, N., Patro, I.K., 2022. Perinatal exposure to synergistic multiple stressors leads to cellular and behavioral deficits mimicking Schizophrenia-like pathology. Biol. Open. 11(3), bio058870. [CrossRef]
- Eyles, D.W., 2021. How do established developmental risk-factors for schizophrenia change the way the brain develops? Transl. Psychiatry. 11(1), 1-15. [CrossRef]
- Guan, Z., Fang, J., 2006. Peripheral immune activation by lipopolysaccharide decreases neurotrophins in the cortex and hippocampus in rats. Brain Behav. Immun. 20(1), 64-71. [CrossRef]
- Zhang, X.Y., Liang, J., Chen, D.C., Xiu, M.H., De Yang, F., Kosten, T.A., Kosten, T.R., 2012. Low BDNF is associated with cognitive impairment in chronic patients with schizophrenia. Psychopharmacology Berl. 222(2), 277-284. [CrossRef]
- Smith, J.A., Das, A., Ray, S.K., Banik, N.L., 2012. Role of pro-inflammatory cytokines released from microglia in neurodegenerative diseases. Brain Res. Bull. 87(1), 10-20. [CrossRef]
- Zhao, J., Bi, W., Xiao, S., Lan, X., Cheng, X., Zhang, J., Lu, D., Wei, W., Wang, Y., Li, H., Fu, Y., 2019. Neuroinflammation induced by lipopolysaccharide causes cognitive impairment in mice. Scientific reports, 9(1), 1-12. [CrossRef]
- Chamera, K., Trojan, E., Szuster-Głuszczak, M., Basta-Kaim, A., 2020. The potential role of dysfunctions in neuron-microglia communication in the pathogenesis of brain disorders. Curr. Neuropharmacol. 18(5), 408-430. [CrossRef]
- Vogelzangs, N., De Jonge, P., Smit, J.H., Bahn, S., Penninx, B.W., 2016. Cytokine production capacity in depression and anxiety. Transl. Psychiatry. 6(5), e825-e825. [CrossRef]
- Gaspersz, R., Lamers, F., Kent, J.M., Beekman, A.T., Smit, J.H., van Hemert, A.M., Schoevers, R.A., Penninx, B.W., 2017. Anxious distress predicts subsequent treatment outcome and side effects in depressed patients starting antidepressant treatment. J. Psychiatr. Res. 84, 41-48. [CrossRef]
- Casida, J.E., Quistad, G.B., 1998. Golden age of insecticide research: past, present, or future? Annu. Rev. Entomol. 43 1-16, . [CrossRef]
- Pitzer, E.M., Sugimoto, C., Gudelsky, G.A., Huff Adams, C.L., Williams, M.T., Vorhees, C.V., 2019. Deltamethrin exposure daily from postnatal day 3–20 in Sprague-Dawley rats causes long-term cognitive and behavioral deficits. Toxicol. Sci. 169(2), 511-523. [CrossRef]
- Pitzer, E.M., Williams, M.T., Vorhees, C.V., 2021. Effects of pyrethroids on brain development and behavior: Deltamethrin. Neurotoxicol Teratol. (87), 106983. [CrossRef]
- Patro, N., Shrivasta va, M., Tripathi, S., Patro, I.K., 2009. S100β upregulation: a possible mechanism of deltamethrin toxicity and motor coordination deficits. Neurotoxicol Teratol. 31(3), 169-176. [CrossRef]
- Chen, N.N., Luo, D.J., Yao, X.Q., Yu, C., Wang, Y., Wang, Q., Wang, J.Z., Liu, G.P., 2012. Pesticides induce spatial memory deficits with synaptic impairments and an imbalanced tau phosphorylation in rats. J. Alzheimer’s Dis. 30(3), 585-594. [CrossRef]
- Hossain, M.M., Belkadi, A., Al-Haddad, S., Richardson, J.R., 2020. Deltamethrin exposure inhibits adult hippocampal neurogenesis and causes deficits in learning and memory in mice. Toxicological Sciences, 178(2), 347-357. [CrossRef]
- Islam, M.S., Azim, F., Saju, H., Zargaran, A., Shirzad, M., Kamal, M., Fatema, K., Rehman, S., Azad, M.M., Ebrahimi-Barough, S., 2021. Pesticides and Parkinson’s disease: Current and future perspective. J. Chem. Neuroanat. 115,101966. [CrossRef]
- Li, Y., Fang, R., Liu, Z., Jiang, L., Zhang, J., Li, H., Liu, C. and Li, F., 2021. The association between toxic pesticide environmental exposure and Alzheimer’s disease: A scientometric and visualization analysis. Chemosphere. 263,128238. [CrossRef]
- Kamel, F., Umbach, D.M., Bedlack, R.S., Richards, M., Watson, M., Alavanja, M.C., Blair, A., Hoppin, J.A., Schmidt, S., Sandler, D.P., 2012. Pesticide exposure and amyotrophic lateral sclerosis. Neurotoxicology. 33(3), 457-462. [CrossRef]
- Parrón, T., Requena, M., Hernández, A.F., Alarcón, R., 2011. Association between environmental exposure to pesticides and neurodegenerative diseases. Toxicol. Appl. Pharmacol. 256(3), 379-385. [CrossRef]
- Xue, Z., Li, X., Su, Q., Xu, L., Zhang, P., Kong, Z., Xu, J., Teng, J., 2013. Effect of synthetic pyrethroid pesticide exposure during pregnancy on the growth and development of infants. Asia Pac J Public Health. 25(4_suppl), 72S-79S. [CrossRef]
- Sinha, S., Patro, N. and Patro, I.K., 2020. Amelioration of neurobehavioral and cognitive abilities of F1 progeny following dietary supplementation with Spirulina to protein malnourished mothers. Brain Behav. Immun. 85, 69-87. [CrossRef]
- Prado, E.L., Dewey, K.G., 2014. Nutrition and brain development in early life. Nutr. Rev. 72(4), 267-284. [CrossRef]
- Hoeijmakers, L., Lucassen, P.J., Korosi, A., 2015. The interplay of early-life stress, nutrition, and immune activation programs adult hippocampal structure and function. Front. Mol. Neurosci. (7), 103. [CrossRef]
- Monk, C., Lugo-Candelas, C., Trumpff, C., 2019. Prenatal developmental origins of future psychopathology: mechanisms and pathways. Annu Rev Clin Psychol. 15, 317. [CrossRef]
- Bayer, T.A., Falkai, P., Maier, W., 1999. Genetic and non-genetic vulnerability factors in schizophrenia: the basis of the two hit hypothesis. J. Psychiatr. 33(6), 543-8. [CrossRef]
- Maynard, T.M., Sikich, L., Lieberman, J.A., LaMantia, A.S., 2001. Neural development, cell-cell signaling, and the “two-hit” hypothesis of schizophrenia. Schizophr. Bull. 27(3), 457-476. [CrossRef]
- Cardoso, F.L., Herz, J., Fernandes, A., Rocha, J., Sepodes, B., Brito, M.A., McGavern, D.B., Brites, D., 2015. Systemic inflammation in early neonatal mice induces transient and lasting neurodegenerative effects. J. Neuroinflammation. 12(1) (2015), 1-18. [CrossRef]
- Dobner, J., Kaser, S., 2018. Body mass index and the risk of infection-from underweight to obesity. Clin. Microbiol. Infect. 24(1) (2018), 24-28. [CrossRef]
- Malik, S., Spencer, S.J., 2019. Early life stress and metabolism. Curr Opin Behav Sci. 28, 25-30. [CrossRef]
- Spencer, S.J., 2013. Perinatal programming of neuroendocrine mechanisms connecting feeding behavior and stress. Front. Neurosci. 7, 109. [CrossRef]
- Peña, C.J., Smith, M., Ramakrishnan, A., Cates, H.M., Bagot, R.C., Kronman, H.G., Patel, B., Chang, A.B., Purushothaman, I., Dudley, J. and Morishita, H., 2019. Early life stress alters transcriptomic patterning across reward circuitry in male and female mice. Nat.Commun.10(1), 5098. [CrossRef]
- Surkan, P.J., Ettinger, A.K., Hock, R.S., Ahmed, S., Strobino, D.M. and Minkovitz, C.S., 2014. Early maternal depressive symptoms and child growth trajectories: a longitudinal analysis of a nationally representative US birth cohort. BMC pediatrics, 14(1), 1-8. http://www.biomedcentral.com/1471-2431/14/185.
- Danese, A., Lewis, S. J., 2017. Psychoneuroimmunology of early-life stress: the hidden wounds of childhood trauma? Neuropsychopharmacology. 42(1), 99-114. [CrossRef]
- Tizard, I., 2008. Sickness behavior, its mechanisms and significance. Anim. Health Res. Rev. 9(1), 87-99. [CrossRef]
- Shattuck, E.C., Muehlenbein, M.P., 2015. Human sickness behavior: ultimate and proximate explanations. Am. J. Phys. Anthropol. 157(1), 1-18. [CrossRef]
- Nemzek, J.A., Hugunin, K., Opp, M.R., 2008. Modeling sepsis in the laboratory: merging sound science with animal well-being, Comp. Med. 58(2), 120-128.
- Bay-Richter, C., Janelidze, S., Hallberg, L., Brundin, L., 2011. Changes in behaviour and cytokine expression upon a peripheral immune challenge. Behav. Brain Res. 222(1), 193-199. [CrossRef]
- Jones, P.B., Rantakallio, P., Hartikainen, A.L., Isohanni, M., Sipila, P., 1998. Schizophrenia as a long-term outcome of pregnancy, delivery, and perinatal complications: a 28-year follow-up of the 1966 north Finland general population birth cohort, Am. J. Psychiatry. 155(3), 355-364. [CrossRef]
- Wahlbeck, K., Forsén, T., Osmond, C., Barker, D.J., Eriksson, J.G., 2001. Association of schizophrenia with low maternal body mass index, small size at birth, and thinness during childhood. Arch. Gen. Psychiatry. 58(1), 48-52. [CrossRef]
- Hatch, B., Healey, D.M., Halperin, J.M., 2014. Associations between birth weight and attention-deficit/hyperactivity disorder symptom severity: indirect effects via primary neuropsychological functions. J Child Psychol Psychiatry. 55(4), 384-392. [CrossRef]
- Kim, J.H., Kim, J.Y., Lee, J., Jeong, G.H., Lee, E., Lee, S., Lee, K.H., Kronbichler, A., Stubbs, B., Solmi, M., Koyanagi, A., 2020. Environmental risk factors, protective factors, and peripheral biomarkers for ADHD: an umbrella review. Lancet Psychiatry. 7(11), 955-970. [CrossRef]
- Seibenhener, M.L., Wooten, M.C., 2015. Use of the open field maze to measure locomotor and anxiety-like behavior in mice. JoVE (Journal of Visualized Experiments), (96), p.e52434. [CrossRef]
- Lezak, K. R., Missig, G., Carlezon Jr, W. A., 2017. Behavioral methods to study anxiety in rodents. Dialogues Clin. Neurosci. 19(2), 181-191. https://doi 10.31887/DCNS.2017.19.2/wcarlezon.
- La-Vu, M., Tobias, B.C., Schuette, P.J., Adhikari, A., 2020. To approach or avoid: an introductory overview of the study of anxiety using rodent assays. Front. Behav. Neurosci. (14), 145. [CrossRef]
- Bilkei-Gorzo, A., Racz, I., Michel, K., Zimmer, A., 2002. Diminished anxiety-and depression-related behaviors in mice with selective deletion of the Tac1 gene. J. Neurosci. 22(22) (2002), 10046-10052. [CrossRef]
- Pawlak, C.R., Ho, Y.J., Schwarting, R.K., 2008. Animal models of human psychopathology based on individual differences in novelty-seeking and anxiety. Neurosci. Biobehav. Rev. 32(8), 1544-1568. [CrossRef]
- Monterosso, J., Ainslie, G., 1999. Beyond discounting: possible experimental models of impulse control. J. Psychopharmacol. 146(4), 339-347. [CrossRef]
- Arce, E., Santisteban, C., 2006. Impulsivity: a review. Psicothema. 18 (2) 213-220.
- Ralph, R.J., Paulus, M.P., Fumagalli, F., Caron, M.G., Geyer, M.A., 2001. Prepulse inhibition deficits and perseverative motor patterns in dopamine transporter knock-out mice: differential effects of D1 and D2 receptor antagonists. J. Neurosci. 21(1), 305-313. [CrossRef]
- Kalueff, A.V., Stewart, A.M., Song, C., Berridge, K.C., Graybiel, A.M., Fentress, J.C., 2016. Neurobiology of rodent self-grooming and its value for translational neuroscience. Nat. Rev. Neurosci. 17(1), 45-59. [CrossRef]
- Kalueff, A.V., Tuohimaa, P., 2005. The grooming analysis algorithm discriminates between different levels of anxiety in rats: potential utility for neurobehavioural stress research. J. Neurosci. Methods. 143(2), 169-177. [CrossRef]
- Denmark, A., Tien, D., Wong, K., Chung, A., Cachat, J., Goodspeed, J., Grimes, C., Elegante, M., Suciu, C., Elkhayat, S., Bartels, B., 2010. The effects of chronic social defeat stress on mouse self-grooming behavior and its patterning. Behav. Brain Res. 208(2), 553-559. [CrossRef]
- Joel, D., 2006. Current animal models of obsessive compulsive disorder: a critical review. Prog. Neuropsychopharmacol, Biol. Psychiatry, 30(3), 374-388. [CrossRef]
- Feusner, J.D., Hembacher, E., Phillips, K.A., 2009. The mouse who couldn’t stop washing: pathologic grooming in animals and humans. CNS Spectr. 14(9), 503-513. [CrossRef]
- Schmeisser, M.J., Ey, E., Wegener, S., Bockmann, J., Stempel, A.V., Kuebler, A., Janssen, A.L., Udvardi, P.T., Shiban, E., Spilker, C., Balschun, D., 2012. Autistic-like behaviours and hyperactivity in mice lacking ProSAP1/Shank2. Nature. 486(7402), 256-260. [CrossRef]
- Sungur, A.Ö., Vörckel, K.J., Schwarting, R.K., Wöhr, M., 2014. Repetitive behaviors in the Shank1 knockout mouse model for autism spectrum disorder: developmental aspects and effects of social context. J. Neurosci. Methods. (234), 92-100. [CrossRef]
- Wolff, S., Chess, S., 1964. A behavioural study of schizophrenic children, Acta Psychiatr. Scand. 40(4), 438-66. https://:10.1111/j.1600-0447.1964.tb07496.x.
- Roehr, B., 2013. American psychiatric association explains DSM-5. 346:f3591346. [CrossRef]
- Lange, K.W., 2020. Micronutrients and diets in the treatment of attention-deficit/hyperactivity disorder: chances and pitfalls. Front. Psychiatry, 11, 102. https//doi:10.3389/fpsyt.2020.00102.
- Marx, W., Moseley, G., Berk, M., Jacka, F., 2017. Nutritional psychiatry: the present state of the evidence. Proc Nutr Soc. 76(4), 427-436. [CrossRef]
- Teasdale, S.B., Müller-Stierlin, A.S., Ruusunen, A., Eaton, M., Marx, W., Firth, J., 2021. Prevalence of food insecurity in people with major depression, bipolar disorder, and schizophrenia and related psychoses: A systematic review and meta-analysis. Crit Rev Food Sci Nutr.1-18. [CrossRef]
- Almeida, S.S., Garcia, R.A., Cibien, M.M., De Araujo, M., Moreira, G., De Oliveira, L.M., 1994. The ontogeny of exploratory behaviors in early-protein-malnourished rats exposed to the elevated plus-maze test. Psychobiol. 22(4), 283-288.
- Bobyn, P.J., Corbett, D., Saucier, D.M., Noyan-Ashraf, M.H., Juurlink, B.H., Paterson, P.G., 2005. Protein-energy malnutrition impairs functional outcome in global ischemia. Exp. Neurol. 196(2), 308-315. [CrossRef]
- Péter, Z., Oliphant, M.E. and Fernandez, T.V., 2017. Motor stereotypies: a pathophysiological review. Front. Neurosci. 11(171), https:// doi: 10.3389/fnins.2017.00171.
- Ridley, R.M., 1994. The psychology of perseverative and stereotyped behaviour. Prog. Neurobiol. 44(2), 221-231. [CrossRef]
- Hoptman, M.J., Ahmed, A.O., 2016. Neural foundations of mood-induced impulsivity and impulsive aggression in schizophrenia. Curr. Behav. Neurosci. Rep. 3(3), 248-255. [CrossRef]
- van Erp, T.G., Baker, R.A., Cox, K., Okame, T., Kojima, Y., Eramo, A., Potkin, S.G., 2020. Effect of brexpiprazole on control of impulsivity in schizophrenia: a randomized functional magnetic resonance imaging study. Psychiatry Res. Neuroimaging. (301), 111085. [CrossRef]
- Grassi, G., Pallanti, S., Righi, L., Figee, M., Mantione, M., Denys, D., Piccagliani, D., Rossi, A., Stratta, P., 2015. Think twice: Impulsivity and decision making in obsessive–compulsive disorder. J. Behav. Addict. 4(4), 263-272. [CrossRef]
- Krakowski, A.D., Cost, K.T., Anagnostou, E., Lai, M.C., Crosbie, J., Schachar, R., Georgiades, S., Duku, E., Szatmari, P., 2020. Inattention and hyperactive/impulsive component scores do not differentiate between autism spectrum disorder and attention-deficit/hyperactivity disorder in a clinical sample. Mol. Autism. 11(1), 1-13. [CrossRef]
- Grimm, O., Kranz, T.M., Reif, A., 2020. Genetics of ADHD: what should the clinician know? Curr. Psychiatry Rep. 22(4), 1-8, . [CrossRef]
- Richardson, J.R., Taylor, M.M., Shalat, S.L., Guillot III, T.S., Caudle, W.M., Hossain, M.M., Mathews, T.A.S.R., Cory-Slechta, D.A., Miller, G.W., 2015. Developmental pesticide exposure reproduces features of attention deficit hyperactivity disorder. FASEB J. 29(5), 1960-1972. [CrossRef]
- Singh, K. Patro, N., Pradeepa, M., Patro, I., 2017. Neonatal lipopolysaccharide infection causes demyelination and behavioral deficits in adult and senile rat brain. Ann. Neurosci. , 24(3), 146-154. [CrossRef]
- Baghel, M.S., Singh, B., Dhuriya, Y.K., Shukla, R.K., Patro, N., Khanna, V.K., Patro, I.K., Thakur, M.K., 2018. Postnatal exposure to poly (I: C) impairs learning and memory through changes in synaptic plasticity gene expression in developing rat brain. Neurobiol. Learn. Mem. 155, 379-389. [CrossRef]
- Handley, S.L., Mithani, S., 1984. Effects of alpha-adrenoceptor agonists and antagonists in a maze-exploration model of ‘fear’-motivated behaviour. Naunyn Schmiedebergs Arch. Pharmacol. 327(1), 1-5. [CrossRef]
- Atrooz, F., Alkadhi, K.A., Salim, S., 2021. Understanding stress: Insights from rodent models, Current Research in Neurobiology, 2, 100013, . [CrossRef]
- Pellow, S., Chopin, P., File, S.E., Briley, M., 1985. Validation of open: closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Methods. 14(3), 149-167. [CrossRef]
- Komada, M., Takao, K., Miyakawa, T., 2008. Elevated plus maze for mice. J. Vis. Exp. (22), p.e1088. [CrossRef]
- Li, J., Olsen, J., Vestergaard, M., Obel, C., 2010. Attention-deficit/hyperactivity disorder in the offspring following prenatal maternal bereavement: a nationwide follow-up study in Denmark. Eur. Child Adolesc. Psychiatry. 19(10), 747-753. [CrossRef]
- Menet, J.S., Rosbash, M., 2011. When brain clocks lose track of time: cause or consequence of neuropsychiatric disorders. Curr. Opin. Neurobiol. 21(6), 849-857. [CrossRef]
- Takeuchi, H., Iba, M., Inoue, H., Higuchi, M., Takao, K., Tsukita, K., Karatsu, Y., Iwamoto, Y., Suhara, T., Miyakawa, T., Trojanowski, J.Q., 2011. P301S mutant human tau transgenic mice manifest early symptoms of human tauopathies with dementia and altered sensorimotor gating. PloS one. 6(6), e21050. [CrossRef]
- Tian, Y.H., Baek, J.H., Lee, S.Y., Jang, C.G., 2010. Prenatal and postnatal exposure to bisphenol a induces anxiolytic behaviors and cognitive deficits in mice. Synapse. 64(6), 432-439. [CrossRef]
- Crawley, J., Goodwin, F.K., 1980. Preliminary report of a simple animal behavior model for the anxiolytic effects of benzodiazepines. Pharmacol Biochem Behav. 13(2), 167-170. [CrossRef]
- Birkett, M.A., Shinday, N.M., Kessler, E.J., Meyer, J.S., Ritchie, S., Rowlett, J.K., 2011. Acute anxiogenic-like effects of selective serotonin reuptake inhibitors are attenuated by the benzodiazepine diazepam in BALB/c mice. Pharmacol. Biochem. Behav. 98(4), 544-551. [CrossRef]
- Bailey, K.R., Crawley, J.N., 2009. Anxiety-Related Behaviors in Mice, In: Buccafusco JJ, editor. Methods of Behavior Analysis in Neuroscience. 2nd ed. Boca Raton (FL): CRC Press/Taylor & Francis; 2009. Chapter 5. PMID: 21204329.
- Kulesskaya, N., Voikar, V., 2014. Assessment of mouse anxiety-like behavior in the light–dark box and open-field arena: role of equipment and procedure. Physiol. Behav. (133), 30-38. [CrossRef]
- Bourin, M., Hascoët, M., 2003. The mouse light/dark box test. Eur. J. Pharmacol. 463(1-3), 55-65. [CrossRef]
- Talukdar, P.M., Abdul, F., Maes, M., Berk, M., Venkatasubramanian, G., Kutty, B.M., Debnath, M., 2021. A proof-of-concept study of maternal immune activation mediated induction of Toll-like receptor (TLR) and inflammasome pathways leading to neuroprogressive changes and schizophrenia-like behaviours in offspring. Eur Neuropsychopharmacol. (5), 48-61. [CrossRef]
- Faraone, S.V., Mick, E., 2010. Molecular genetics of attention deficit hyperactivity disorder. Psychiatr. Clin. 33(1), 159-180. [CrossRef]
- Faraone, S.V., Larsson, H., 2019. Genetics of attention deficit hyperactivity disorder. Mol. Psychiatry. 24(4), 562-575. [CrossRef]
Figure 1.
Overview of the experimental design and procedures followed.

Figure 2.
Effects of singular or cumulative exposure of LPS and DLT to maternally PMN F1 rats on physical development: LP group animals showed significantly reduced body weight as compared to age matched control group. However, LP animals when treated either with LPS (LP+LPS) or DLT (LP+DLT) or both (LP+DLT+LPS) revealed further reduction in body weight as compared to their respective control groups (n=12/group). Figs. 2B-E reveals the difference in the body appearance of multi-hit rats and their age matched controls at the age of PND7, 1M, 3M and 6M. Data was analyzed by one-way (comparison with control) and two-way (comparison between diet and infection) ANOVA and presented as mean ± SEM (n=12); **p≤ 0.01, ***p≤ 0.001, for comparison between Control group and treated animals; #p≤0.05, ###p≤ 0.001, for comparison between LP group and treated group; $$$p≤ 0.001; for comparison between Control+LPS and LP+LPS group; @@@p≤ 0.001, for comparison between Control+DLT and LP+DLT group; &&&p≤ 0.001, for comparison between Control+DLT+LPS and LP+DLT+LPS group.
Figure 2.
Effects of singular or cumulative exposure of LPS and DLT to maternally PMN F1 rats on physical development: LP group animals showed significantly reduced body weight as compared to age matched control group. However, LP animals when treated either with LPS (LP+LPS) or DLT (LP+DLT) or both (LP+DLT+LPS) revealed further reduction in body weight as compared to their respective control groups (n=12/group). Figs. 2B-E reveals the difference in the body appearance of multi-hit rats and their age matched controls at the age of PND7, 1M, 3M and 6M. Data was analyzed by one-way (comparison with control) and two-way (comparison between diet and infection) ANOVA and presented as mean ± SEM (n=12); **p≤ 0.01, ***p≤ 0.001, for comparison between Control group and treated animals; #p≤0.05, ###p≤ 0.001, for comparison between LP group and treated group; $$$p≤ 0.001; for comparison between Control+LPS and LP+LPS group; @@@p≤ 0.001, for comparison between Control+DLT and LP+DLT group; &&&p≤ 0.001, for comparison between Control+DLT+LPS and LP+DLT+LPS group.
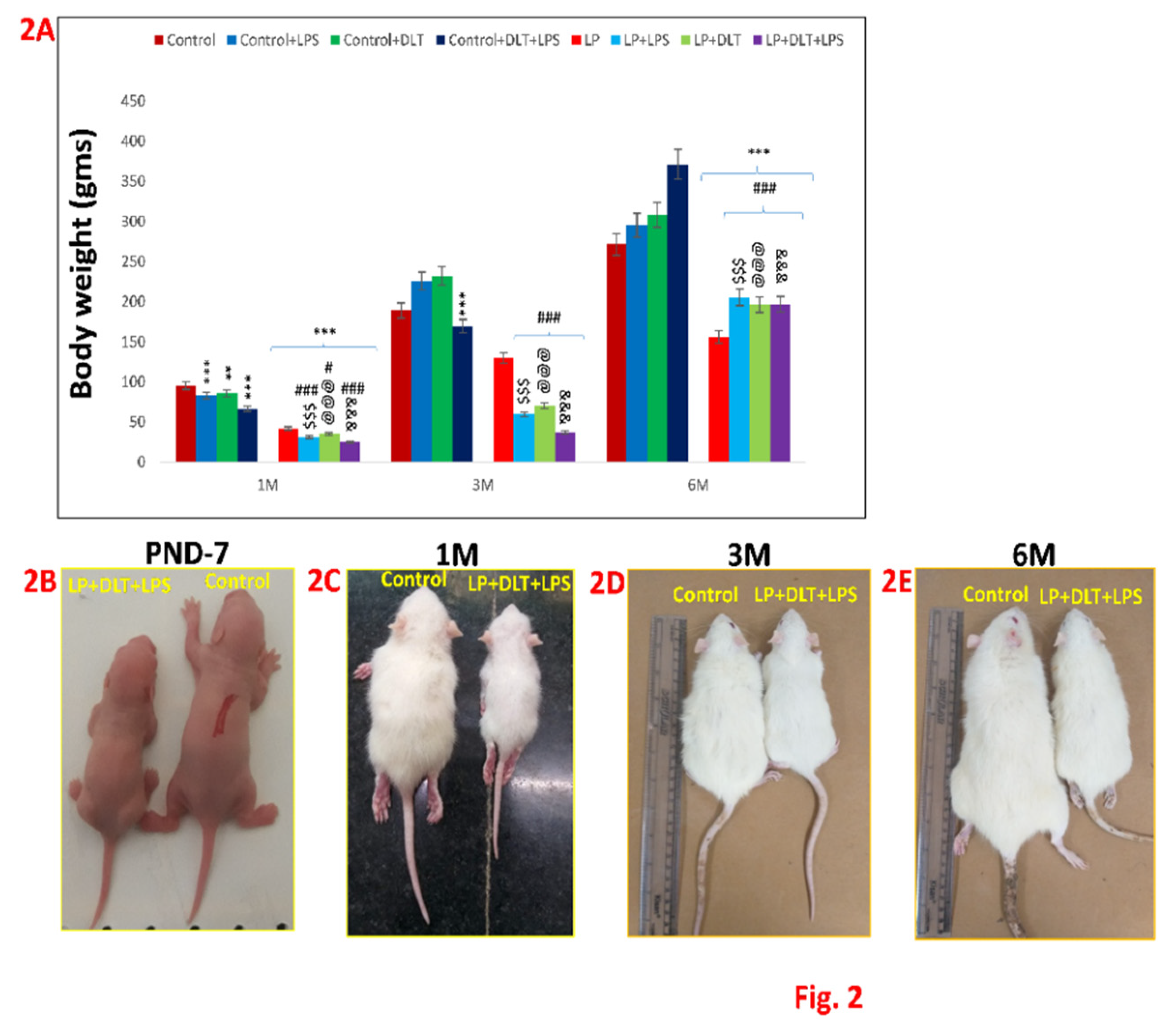
Figure 3 & 4.
Open field test data showing hyperactivity/inattention, motor impulsivity and low anxiety like symptoms in LP+DLT+LPS group rats at adolescence and adulthood: Bar graphs represent, resting time (A), distance travelled (B), stereotypic time (C), ambulatory time (D), horizontal count (E), center zone entries (F) and center zone time (G) at 1, 3 and 6 months of age. All the treated group rats revealed significantly increased distance travelled, stereotypic time, ambulatory time, horizontal count, center zone entries, center zone time with low resting time when compared to age matched control. However, LP+DLT+LPS group rats displayed highest level changes in all parameters as compared to control and LP alone rats. Representative open field activity track reports (Figure 4) revealed that control animals safely explore the open arena (Figure 4a,e,i), while LP F1 rats showed increased exploration (Figure 4m,q,u) and LP+LPS (Figure 4n,r,v), LP+DLT (Figure 4o,s,w) and LP+DLT+LPS (Figure 4p,t,x) group rats exhibit further behavioral impairments. Moreso, LP+DLT+LPS group rats also present increased and haphazard center zone exploration (S6, S7, S10 and S11). Data was analyzed by one-way (comparison with control) and two-way (comparison between diet and infection) ANOVA and presented as mean ± SEM; ***p≤ 0.001, for comparison between Control group and treated animals; ###p≤ 0.001, for comparison between LP group and treated group; $$$p≤ 0.001, for comparison between Control+LPS and LP+LPS group; @@p≤ 0.01, @@@p≤ 0.001, for comparison between Control+DLT and LP+DLT group; &p≤0.05, &&&p≤ 0.001, for comparison between Control+DLT+LPS and LP+DLT+LPS group.
Figure 3 & 4.
Open field test data showing hyperactivity/inattention, motor impulsivity and low anxiety like symptoms in LP+DLT+LPS group rats at adolescence and adulthood: Bar graphs represent, resting time (A), distance travelled (B), stereotypic time (C), ambulatory time (D), horizontal count (E), center zone entries (F) and center zone time (G) at 1, 3 and 6 months of age. All the treated group rats revealed significantly increased distance travelled, stereotypic time, ambulatory time, horizontal count, center zone entries, center zone time with low resting time when compared to age matched control. However, LP+DLT+LPS group rats displayed highest level changes in all parameters as compared to control and LP alone rats. Representative open field activity track reports (Figure 4) revealed that control animals safely explore the open arena (Figure 4a,e,i), while LP F1 rats showed increased exploration (Figure 4m,q,u) and LP+LPS (Figure 4n,r,v), LP+DLT (Figure 4o,s,w) and LP+DLT+LPS (Figure 4p,t,x) group rats exhibit further behavioral impairments. Moreso, LP+DLT+LPS group rats also present increased and haphazard center zone exploration (S6, S7, S10 and S11). Data was analyzed by one-way (comparison with control) and two-way (comparison between diet and infection) ANOVA and presented as mean ± SEM; ***p≤ 0.001, for comparison between Control group and treated animals; ###p≤ 0.001, for comparison between LP group and treated group; $$$p≤ 0.001, for comparison between Control+LPS and LP+LPS group; @@p≤ 0.01, @@@p≤ 0.001, for comparison between Control+DLT and LP+DLT group; &p≤0.05, &&&p≤ 0.001, for comparison between Control+DLT+LPS and LP+DLT+LPS group.
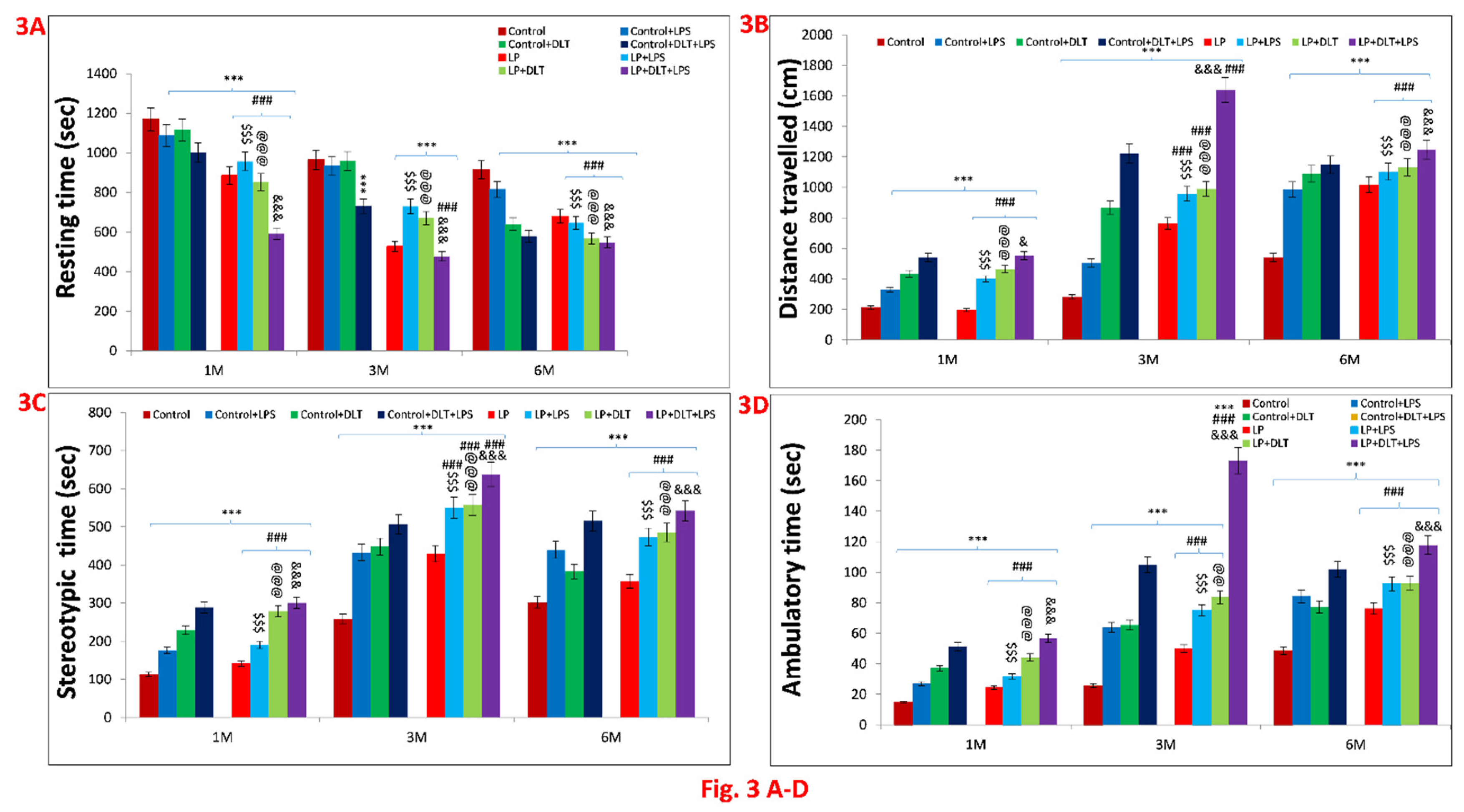
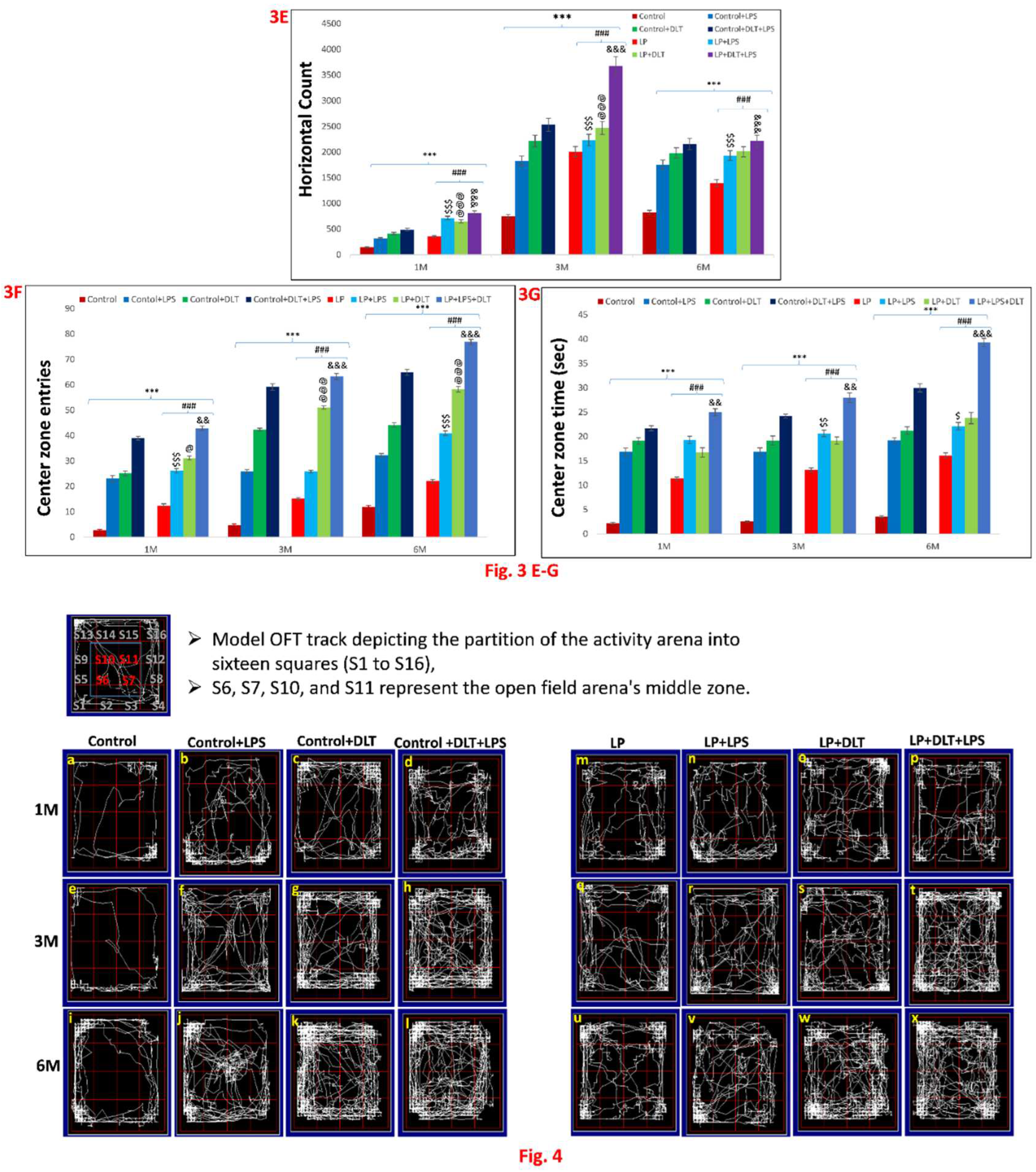
Figure 5 & 6.
Elevated plus maze data also showed low anxiety, hyperactivity/inattention, impulsivity and low-fear like symptoms in LP+DLT+LPS group rats: Bar graphs representing the percent open arm entries (A), percent open arm time (B), center zone entries (C), center zone time (D) and total open arm entries (E) at 1, 3 and 6 months of postnatal age: From bar graphs it was revealed that LP F1 rats prefer open arms and center zone exploration over closed arms thus showing significantly increasing number of open arm entries and time (Figure 5 A, B), center zone entries and time (Figure 5 C, D) and total open arm entries (Figure 5 E) as compared to control group. However, LP+DLT+LPS group rats displayed further increase in all parameters as compared to control and LP alone animals. Track records (Figure 6) indicate that control group rats behave normally while exploring the EPM arms with little percent open arm entries and time (Figure 6 a, i, q). However, exposed control and LP F1 rats favoured exploring open arms over closed ones and center zone. Such behavioral impairment was more severe in the LP+DLT+LPS group animals showing highly anxious profile with very frequent open arm and center zone entries (Figure 6 h, p, x). Moreso, the multi-hit group rats repeatedly explored the open arms to their most distal ends. Data was analyzed by one-way (comparison with control) and two-way (comparison between diet and infection) ANOVA and presented as mean ± SEM; *p≤0.05, ***p≤ 0.001, for comparison between Control group and treated animals; ###p≤ 0.001, for comparison between LP group and treated group; $p≤0.05, $$p≤ 0.01, $$$p≤ 0.001, for comparison between Control+LPS and LP+LPS group; @p≤0.05, @@p≤ 0.01, @@@p≤ 0.001, for comparison between Control+DLT and LP+DLT group; &&p≤ 0.01, &&&p≤ 0.001, for comparison between Control+DLT+LPS and LP+DLT+LPS group.
Figure 5 & 6.
Elevated plus maze data also showed low anxiety, hyperactivity/inattention, impulsivity and low-fear like symptoms in LP+DLT+LPS group rats: Bar graphs representing the percent open arm entries (A), percent open arm time (B), center zone entries (C), center zone time (D) and total open arm entries (E) at 1, 3 and 6 months of postnatal age: From bar graphs it was revealed that LP F1 rats prefer open arms and center zone exploration over closed arms thus showing significantly increasing number of open arm entries and time (Figure 5 A, B), center zone entries and time (Figure 5 C, D) and total open arm entries (Figure 5 E) as compared to control group. However, LP+DLT+LPS group rats displayed further increase in all parameters as compared to control and LP alone animals. Track records (Figure 6) indicate that control group rats behave normally while exploring the EPM arms with little percent open arm entries and time (Figure 6 a, i, q). However, exposed control and LP F1 rats favoured exploring open arms over closed ones and center zone. Such behavioral impairment was more severe in the LP+DLT+LPS group animals showing highly anxious profile with very frequent open arm and center zone entries (Figure 6 h, p, x). Moreso, the multi-hit group rats repeatedly explored the open arms to their most distal ends. Data was analyzed by one-way (comparison with control) and two-way (comparison between diet and infection) ANOVA and presented as mean ± SEM; *p≤0.05, ***p≤ 0.001, for comparison between Control group and treated animals; ###p≤ 0.001, for comparison between LP group and treated group; $p≤0.05, $$p≤ 0.01, $$$p≤ 0.001, for comparison between Control+LPS and LP+LPS group; @p≤0.05, @@p≤ 0.01, @@@p≤ 0.001, for comparison between Control+DLT and LP+DLT group; &&p≤ 0.01, &&&p≤ 0.001, for comparison between Control+DLT+LPS and LP+DLT+LPS group.
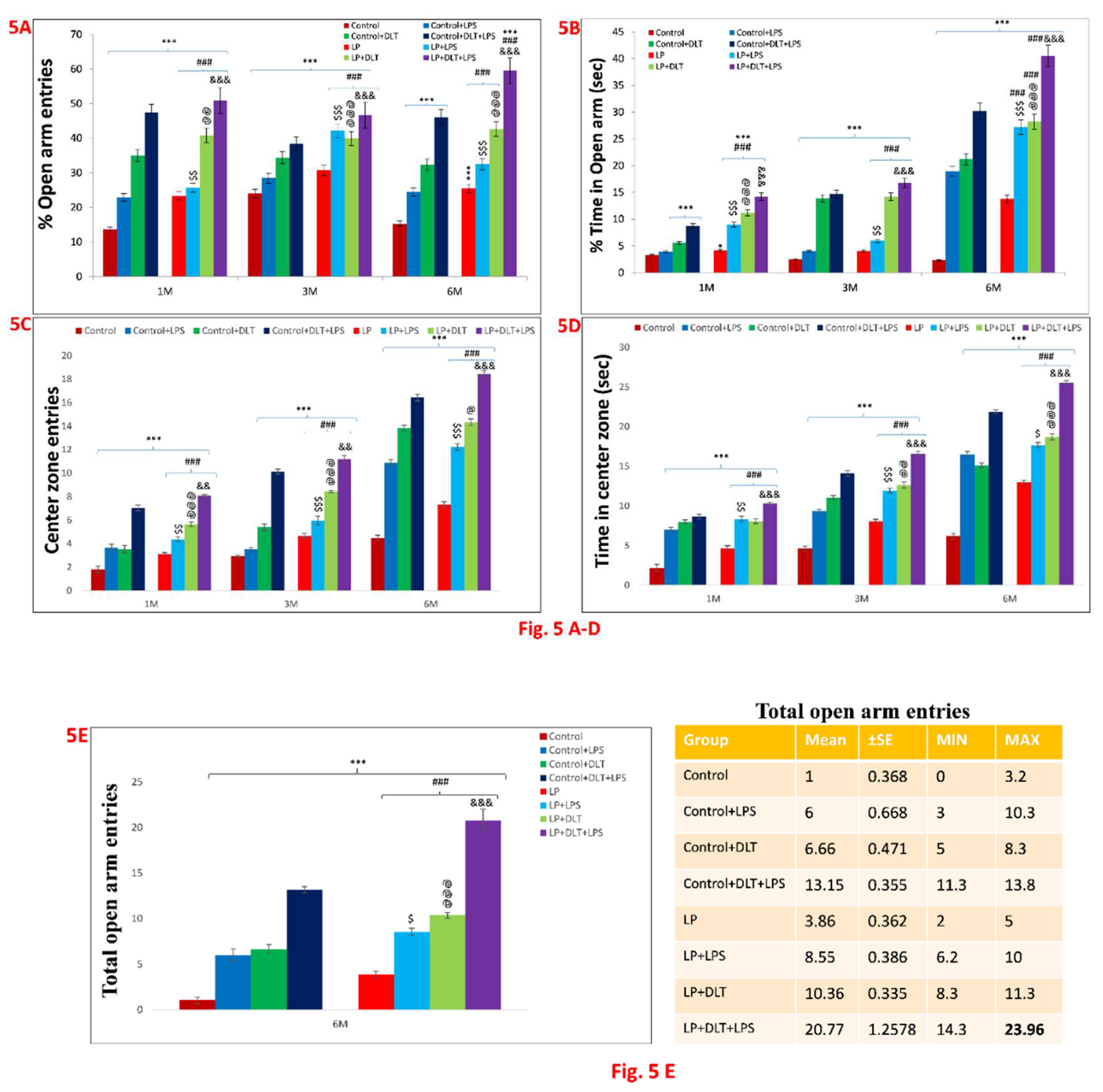
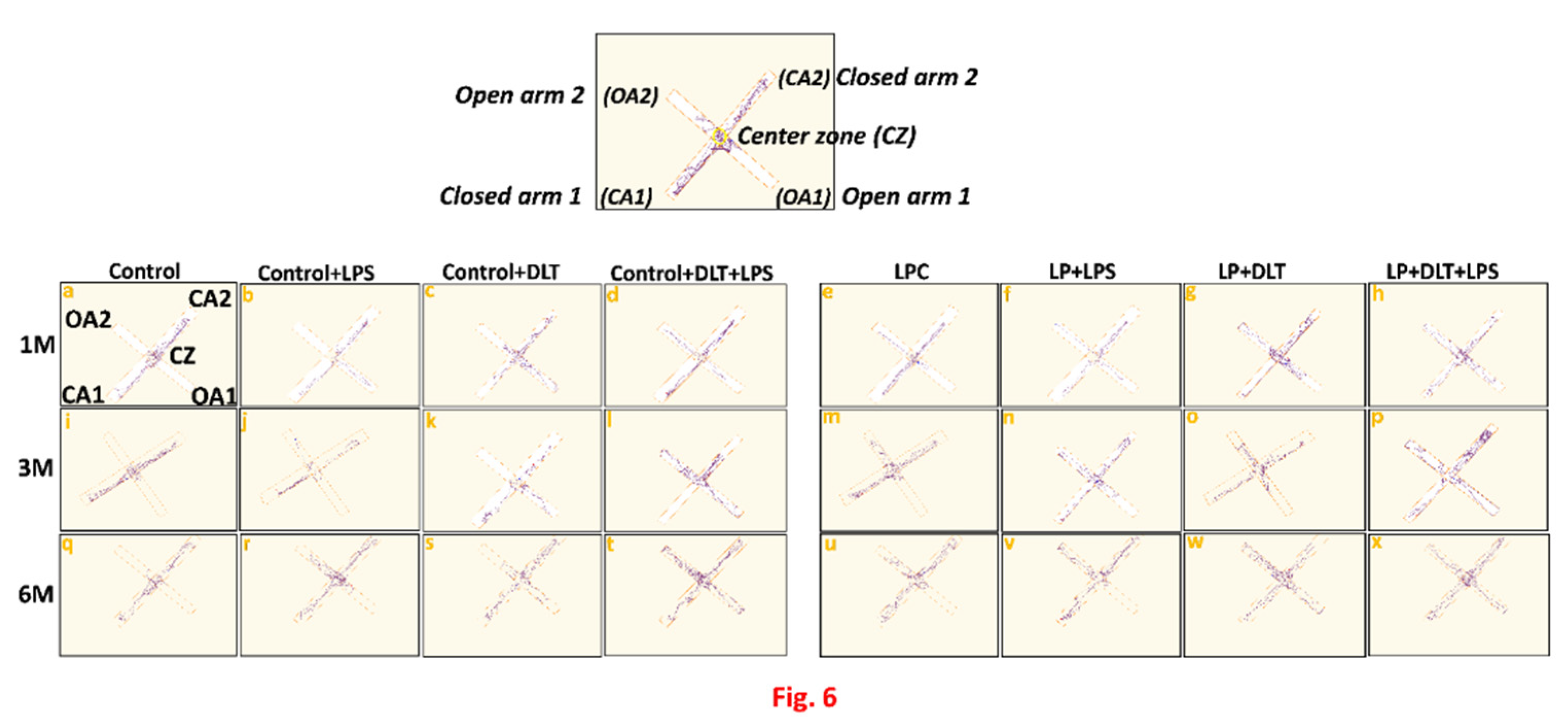
Figure 7.
Light and Dark box data also indicated low anxiety, hyperactivity/inattention, impulsivity and low-fear like symptoms in LP+DLT+LPS group: Bar graphs representing percent light zone entries (A) and percentage light zone time (B) at 1, 3 and 6 months of postnatal age: Data analysis showed that LP animals had significantly higher percent light zone entries (Figure 7A) and percent light zone time (Figure 7B) in contrast to age-matched control animals which preferred dark zone. However, after exposure of either LPS or DLT or both LPS+DLT in Control and LP F1 rats, showed further significant rise in the percentage of light zone entries and light zone time. Furthermore, such changes were significantly higher in the LP F1 stressed animals with maximum in LP+DLT+LPS group rats. Track records also indicate that control group rats showed normal exploratory behaviour upon exploring the light and dark zone with few percent light zone entries and time (Figure 7C a, i, q). However, cumulative exposure of LPS and DLT to Control and LP F1 group rats revealed further increase in locomotor activity evident by frequent entries into the light compartment. Such behavioral impairment was significantly maximum in LP+DLT+LPS group animals as compared to age matched control and other groups (Figure 7C h, p, x). Data was analyzed by one-way (comparison with control) and two-way (comparison between diet and infection) ANOVA and presented as mean ± SEM; ***p≤ 0.001, for comparison between Control group and treated animals; ###p≤ 0.001, for comparison between LP group and treated group; $$$p≤ 0.001, for comparison between Control+LPS and LP+LPS group; @@@p≤ 0.001, for comparison between Control+DLT and LP+DLT group; &&&p≤ 0.001, for comparison between Control+DLT+LPS and LP+DLT+LPS group.
Figure 7.
Light and Dark box data also indicated low anxiety, hyperactivity/inattention, impulsivity and low-fear like symptoms in LP+DLT+LPS group: Bar graphs representing percent light zone entries (A) and percentage light zone time (B) at 1, 3 and 6 months of postnatal age: Data analysis showed that LP animals had significantly higher percent light zone entries (Figure 7A) and percent light zone time (Figure 7B) in contrast to age-matched control animals which preferred dark zone. However, after exposure of either LPS or DLT or both LPS+DLT in Control and LP F1 rats, showed further significant rise in the percentage of light zone entries and light zone time. Furthermore, such changes were significantly higher in the LP F1 stressed animals with maximum in LP+DLT+LPS group rats. Track records also indicate that control group rats showed normal exploratory behaviour upon exploring the light and dark zone with few percent light zone entries and time (Figure 7C a, i, q). However, cumulative exposure of LPS and DLT to Control and LP F1 group rats revealed further increase in locomotor activity evident by frequent entries into the light compartment. Such behavioral impairment was significantly maximum in LP+DLT+LPS group animals as compared to age matched control and other groups (Figure 7C h, p, x). Data was analyzed by one-way (comparison with control) and two-way (comparison between diet and infection) ANOVA and presented as mean ± SEM; ***p≤ 0.001, for comparison between Control group and treated animals; ###p≤ 0.001, for comparison between LP group and treated group; $$$p≤ 0.001, for comparison between Control+LPS and LP+LPS group; @@@p≤ 0.001, for comparison between Control+DLT and LP+DLT group; &&&p≤ 0.001, for comparison between Control+DLT+LPS and LP+DLT+LPS group.
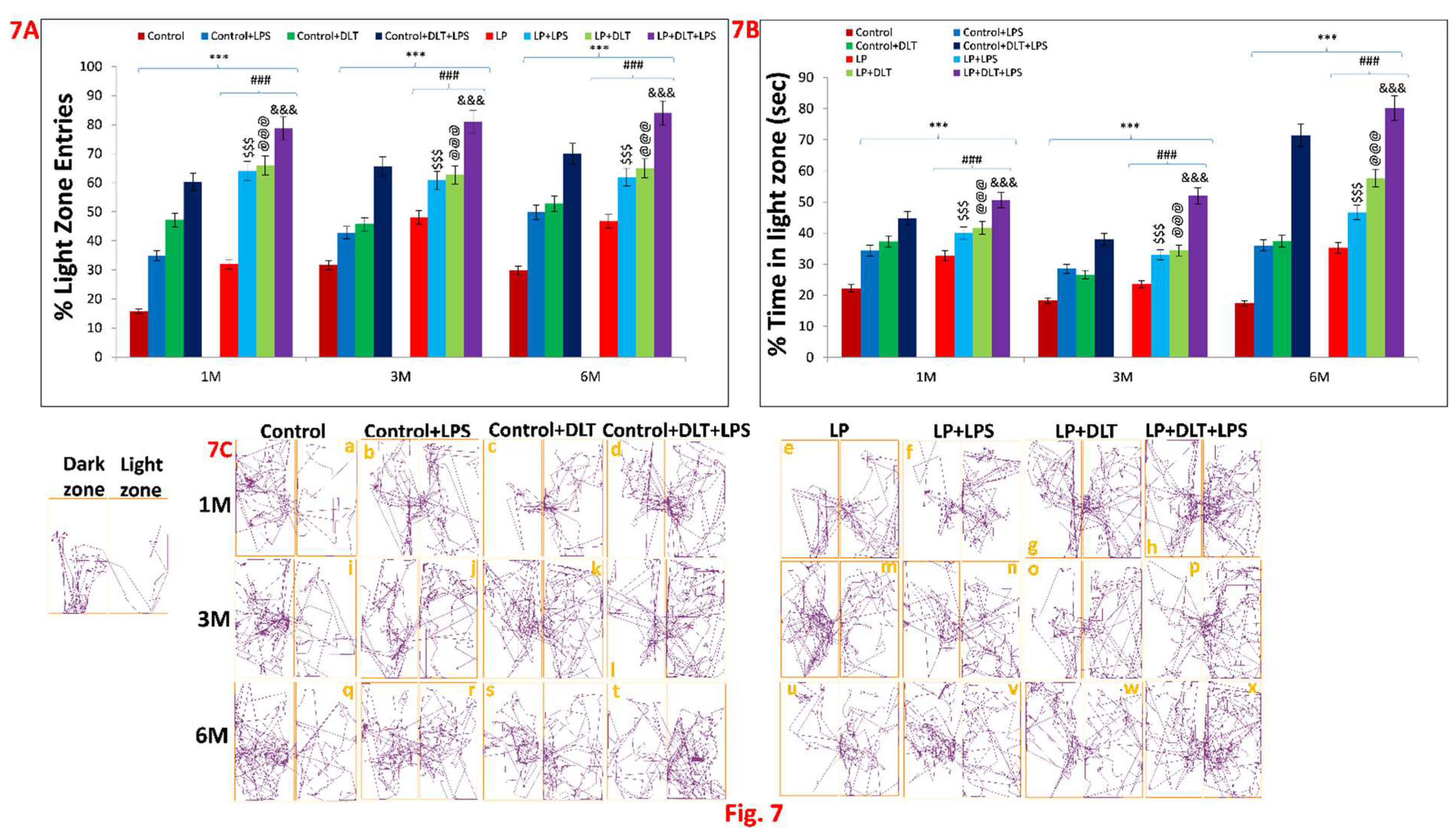
Figure 8.
Rotarod performance displayed abnormal stereotypic behaviour with motor impulsivity and inattention/hyperactive like symptoms in LP+DLT+LPS group: Bar graph indicate the retention time on the accelerating rotating rod at 3 and 6 months of postnatal age: Control and LP F1 group rats exposed to either LPS or DLT or both LPS+DLT showed a significant increase in latency to fall-off from the accelerating rotarod indicating abnormal stereotyped behaviour, which was more prominent in animals belonging to LP F1 treated groups and LP+LPS+DLT group rats indicated significantly maximum increased latency to fall-off among all other groups showing a prominently abnormal stereotypic behaviour than age matched control rats. Data was analyzed by one-way (comparison with control) and two-way (comparison between diet and infection) ANOVA and presented as mean ± SEM; ***p≤ 0.001; for comparison between Control group and treated animals; ###p≤ 0.001; for comparison between LP group and treated group; $$p≤ 0.01, $$$p≤ 0.001, for comparison between Control+LPS and LP+LPS group; @@@p≤ 0.001, for comparison between Control+DLT and LP+DLT group; &&&p≤0.001, for comparison between Control+DLT+LPS and LP+DLT+LPS group.
Figure 8.
Rotarod performance displayed abnormal stereotypic behaviour with motor impulsivity and inattention/hyperactive like symptoms in LP+DLT+LPS group: Bar graph indicate the retention time on the accelerating rotating rod at 3 and 6 months of postnatal age: Control and LP F1 group rats exposed to either LPS or DLT or both LPS+DLT showed a significant increase in latency to fall-off from the accelerating rotarod indicating abnormal stereotyped behaviour, which was more prominent in animals belonging to LP F1 treated groups and LP+LPS+DLT group rats indicated significantly maximum increased latency to fall-off among all other groups showing a prominently abnormal stereotypic behaviour than age matched control rats. Data was analyzed by one-way (comparison with control) and two-way (comparison between diet and infection) ANOVA and presented as mean ± SEM; ***p≤ 0.001; for comparison between Control group and treated animals; ###p≤ 0.001; for comparison between LP group and treated group; $$p≤ 0.01, $$$p≤ 0.001, for comparison between Control+LPS and LP+LPS group; @@@p≤ 0.001, for comparison between Control+DLT and LP+DLT group; &&&p≤0.001, for comparison between Control+DLT+LPS and LP+DLT+LPS group.
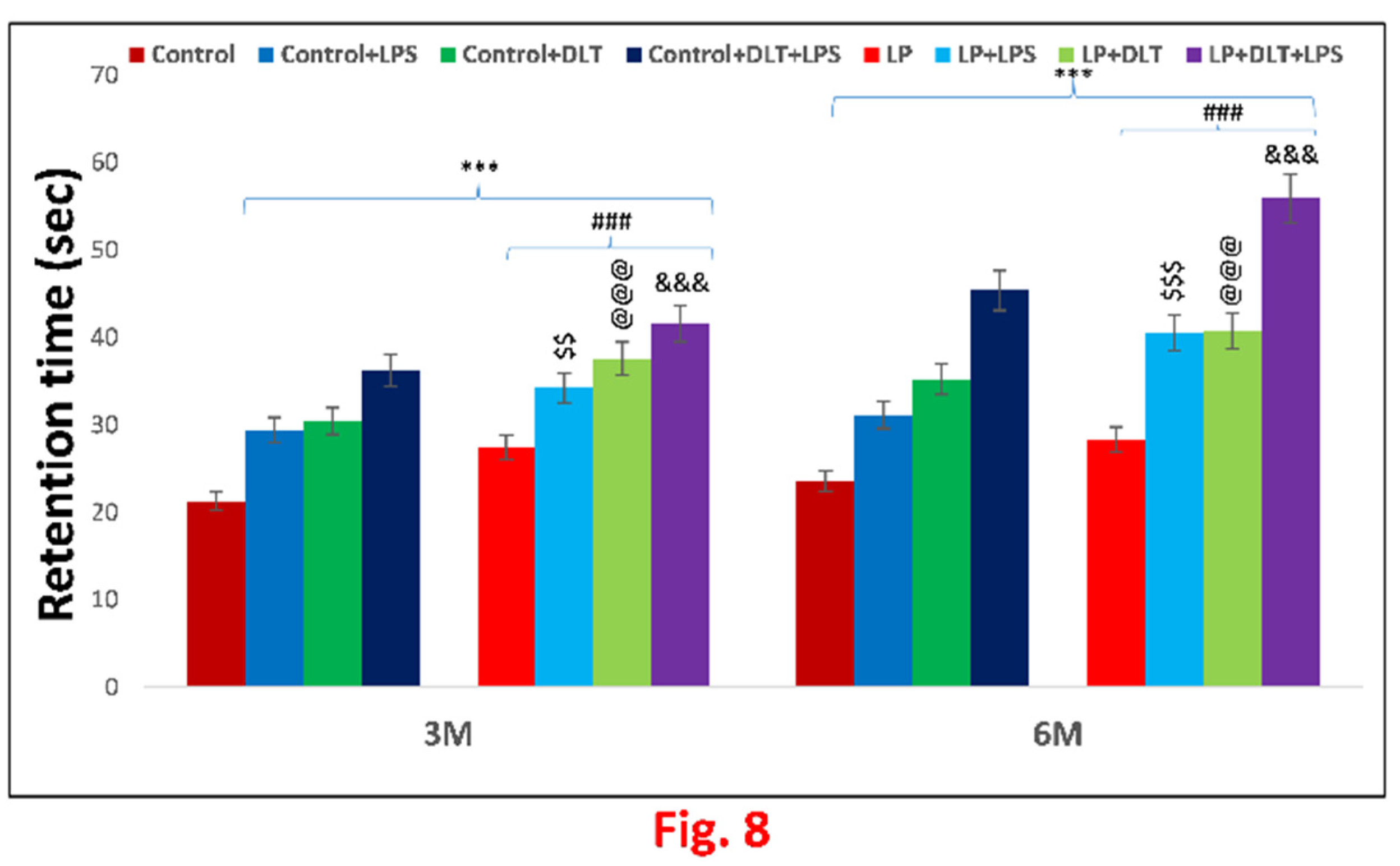

Table 1.1.
Body weight (gm) represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
Table 1.1.
Body weight (gm) represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =25.385, p≤0.001] | [F (7,95) =23.894, p≤0.001] | [F (7,95) =27.811, p≤0.001] |
| 3M | [F (7,95) =35.895, p≤0.001] | [F (7,95) =32.917, p≤0.001] | [F (7,95) =42.173, p≤0.001] |
| 6M | [F (7,95) =14.956, p≤0.001] | [F (7,95) =17.030, p≤0.001] | [F (7,95) =16.900, p≤0.001] |
Table 1.2.
Body weight (gm) represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
Table 1.2.
Body weight (gm) represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =21.125, p≤0.001] | [F (1,95) =4.952, p≤0.001] | [F (1,95) =3.951, p≤0.015] | [F (1,95) =11.425, p≤0.001] | [F (1,95) =4.261, p≤0.001] | [F (1,95) =2.769, p=0.025] | [F (1,95) =6.686, p≤0.001] | [F (1,95) =20.433, p≤0.001] | [F (1,95) =19.942, p≤0.001] | [F (1,95) =16.386, p≤0.001] |
| 3M | [F (1,95) =16.347, p≤0.001] | - | - | [F (1,95) =5.417, p≤0.001] | [F (1,95) =19.548, p≤0.001] | [F (1,95) =16.569, p≤0.001] | [F (1,95) =25.826, p≤0.001] | [F (1,95) =46.184, p≤0.001] | [F (1,95) =44.881, p≤0.001] | [F (1,95) =36.756, p≤0.001] |
| 6M | [F (1,95) =30.977, p≤0.001] | - | - | - | [F (1,95) =11.197, p≤0.001] | [F (1,95) =9.123, p≤0.001] | [F (1,95) =9.252, p≤0.001] | [F (1,95) =24.164, p≤0.001] | [F (1,95) =29.988, p≤0.001] | [F (1,95) =46.891, p≤0.001] |
Table 2.1.
Resting time (sec) represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
Table 2.1.
Resting time (sec) represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M |
[F (7,95) =28.952, p≤0.001] |
[F (7,95) =43.230, p≤0.001] |
[F (7,95) =79.067, p≤0.001] |
| 3M |
[F (7,95) =16.061, p≤0.001] |
[F (7,95) =20.097, p≤0.001] |
[F (7,95) =33.299, p≤0.001] |
| 6M | [F (7,95) =23.689, p≤0.001] | [F (7,95) =30.616, p≤0.001] | [F (7,95) =32.370, p≤0.001] |
Table 2.2.
Resting time (sec) represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
Table 2.2.
Resting time (sec) represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) 38.631, p≤0.001] | [F (1,95) =11.047, p≤0.001] | [F (1,95) =7.105, p≤0.015] |
[F (1,95) =25.412, p≤0.001] |
[F (1,95) =15.910, p≤0.001] | [F (1,95) =30.441, p≤0.001] | [F (1,95) =66.914, p≤0.001] | [F (1,95) =17.878, p≤0.001] | [F (1,95) =36.098, p≤0.001] | [F (1,95) =56.166, p≤0.001] |
| 3M | [F (1,95) =29.858, p≤0.001] | - | - | [F (1,95) =15.993, p≤0.001] | - | - | [F (1,95) =3.441, p≤ 0.001] | [F (1,95) =13.893, p≤0.001] | [F (1,95) =19.687, p≤0.001] | [F (1,95) =17.306, p≤0.001] |
| 6M | [F (1,95) =20.701, p≤0.001] | [F (1,95) =8.822, p≤0.001] | [F (1,95) =24.253, p≤0.001] | [F (1,95) =29.594, p≤0.001] | [F (1,95) =2.988, p= 0.008] | [F (1,95) =9.914, p≤0.001] | [F (1,95) =11.844, p≤ 0.001] | [F (1,95) =14.867, p≤0.001] | [F (1,95) =6.363, p≤0.001] | [F (1,95) =2.952, p= 0.004] |
Table 2.3.
Distance travelled (cm) represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
Table 2.3.
Distance travelled (cm) represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =30.488, p≤0.001] | [F (7,95) =40.905, p≤0.001] | [F (7,95) =55.327 p≤0.001] |
| 3M | [F (7,95) =39.593, p≤0.001] | [F (7,95) =41.488, p≤0.001] | [F (7,95) =79.490, p≤0.001] |
| 6M | [F (7,95) =57.726, p≤0.001] | [F (7,95) =60.553, p≤0.001] | [F (7,95) =72.608, p≤0.001] |
Table 2.4.
Distance travelled (cm) represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
Table 2.4.
Distance travelled (cm) represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
| Age | Control vsLP | Controlvs Control+LPS | Control vs Control+DLT | ControlvsControl+DLT+LPS | LPvsLP+LPS | LPvsLP+DLT | LPvsLP+DLT+LPS | Control+LPSvs LP+LPS | Control+DLTvsLP+DLT | Control+DLT+LPSvsLP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =2.914, p 0.005] | [F (1,95) =19.061, p≤0.001] | [F (1,95) =36.054, p≤0.001] | [F (1,95) =53.401, p≤0.001] | [F (1,95) =33.401, p≤0.001] | [F (1,95) =43.819, p≤0.001] | [F (1,95) =58.240, p≤0.001] | [F (1,95) =11.427, p≤0.001] | [F (1,95) =4.851, p≤0.001] | [F (1,95) =2.084, p=0.041] |
| 3M | [F (1,95) =28.132, p≤0.001] | [F (1,95) =13.004, p≤0.001] | [F (1,95) =34.270, p≤0.001] | [F (1,95) =55.114, p≤0.001] | [F (1,95) =11.461, p≤0.001] | [F (1,95) =13.356, p≤0.001] | [F (1,95) =51.358, p≤0.001] | [F (1,95) =26.589, p≤0.001] | F (1,95) =7.218, p≤0.001] | [F (1,95) =24.376, p≤0.001] |
| 6M | [F (1,95) =48.928,p≤ 0.001] | [F (1,95) =45.814, p≤0.001] | [F (1,95) =56.384, p≤0.001] | [F (1,95) =62.304, p≤0.001] | [F (1,95) =8.798, p≤0.001] | [F (1,95) =11.625, p≤0.001] | [F (1,95) =23.680, p≤0.001] | [F (1,95) =11.912, p≤0.001] | [F (1,95) =4.169, p≤0.001] | [F (1,95) =10.304, p≤0.001] |
Table 2.5.
Stereotypic time (sec) represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
Table 2.5.
Stereotypic time (sec) represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =52.712, p≤0.001] | [F (7,95) =112.616, p≤0.001] | [F (7,95) =127.405 p≤0.001] |
| 3M | [F (7,95) =23.211, p≤0.001] | [F (7,95) =23.769, p≤0.001] | [F (7,95) =30.169, p≤0.001] |
| 6M | [F (7,95) =35.539, p≤0.001] | [F (7,95) =37.990, p≤0.001] | [F (7,95) =49.767, p≤0.001] |
Table 2.6.
Stereotypic time (sec) represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
Table 2.6.
Stereotypic time (sec) represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =18.233, p≤0.001] | [F (1,95) =39.610, p≤0.001] | [F (1,95) =73.058, p≤0.001] | [F (1,95) =110.216, p≤0.001] | [F (1,95) =33.039, p≤0.001] | [F (1,95) =86.136, p≤0.001] | [F (1,95) =99.779, p≤0.001] | [F (1,95) =9.973, p≤0.001] | [F (1,95) =33.785, p≤0.001] | F (1,95) =8.480, p≤0.001] |
| 3M | [F (1,95) =13.590, p≤0.001] | [F (1,95) =13.829, p≤0.001] | [F (1,95) =15.065, p≤0.001] | [F (1,95) =19.783, p≤0.001] | [F (1,95) =9.621, p≤0.001] | [F (1,95) =10.179, p≤0.001] | [F (1,95) =16.529, p≤0.001] | [F (1,95) =9.381, p≤0.001] | [F (1,95) =8.704, p≤0.001] | [F (1,95) =10.386, p≤0.001] |
| 6M | [F (1,95) =11.466, p≤0.001] | [F (1,95) =28.519, p≤0.001] | [F (1,95) =16.824, p≤0.001] | [F (1,95) =44.242, p≤0.001] | [F (1,95) =24.074, p≤0.001] | [F (1,95) =26.524, p≤0.001] | [F (1,95) =38.302, p≤0.001] | [F (1,95) =7.021, p≤0.001] | [F (1,95) =21.166, p≤0.001] | [F (1,95) =5.525, p≤0.001] |
Table 2.7.
Ambulatory time (sec) represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
Table 2.7.
Ambulatory time (sec) represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =11.878, p≤0.001] | [F (7,95) =20.682, p≤0.001] | [F (7,95) =29.346 p≤0.001] |
| 3M | [F (7,95) =9.254, p≤0.001] | [F (7,95) =10.855, p≤0.001] | [F (7,95) =27.597, p≤0.001] |
| 6M | [F (7,95) =17.414, p≤0.001] | [F (7,95) =17.573, p≤0.001] | [F (7,95) =27.530, p≤0.001] |
Table 2.8.
Ambulatory time (sec) represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
Table 2.8.
Ambulatory time (sec) represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =6.847, p≤0.001] | [F (1,95) =8.594, p≤0.001] | [F (1,95) =16.244, p≤0.001] | [F (1,95) =25.573, p≤0.001] | [F (1,95) =4.997, p≤0.001] | [F (1,95) =13.834, p≤0.001] | [F (1,95) =22.498, p≤0.001] | [F (1,95) =3.284, p= 0.003] | [F (1,95) =5.031, p≤0.001] | F (1,95) =3.773, p≤0.001] |
| 3M | [F (1,95) =4.537, p≤0.001] | [F (1,95) =5.849, p≤0.001] | [F (1,95) =7.443, p≤0.001] | [F (1,95) =14.849, p≤0.001] | [F (1,95) =4.717, p≤0.001] | [F (1,95) =6.318, p≤0.001] | [F (1,95) =23.060, p≤0.001] | [F (1,95) =2.121, p= 0.037] | [F (1,95) =3.412, p= 0.001] | [F (1,95) =12.749, p≤0.001] |
| 6M | [F (1,95) =11.028, p≤0.001] | [F (1,95) =14.201, p≤0.001] | [F (1,95) =11.424, p≤0.001] | [F (1,95) =21.262, p≤0.001] | [F (1,95) =6.387, p≤0.001] | [F (1,95) =6.545, p≤0.001] | [F (1,95) =16.502, p≤0.001] | [F (1,95) =3.213, p= 0.002] | [F (1,95) =6.149, p≤0.001] | [F (1,95) =6.268, p≤0.001] |
Table 2.9.
Horizontal count represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
Table 2.9.
Horizontal count represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =63.305, p≤0.001] | [F (7,95) =56.219, p≤0.001] | [F (7,95) =74.260 p≤0.001] |
| 3M | [F (7,95) =379.330, p≤0.001] | [F (7,95) =438.713, p≤0.001] | [F (7,95) =746.570, p≤0.001] |
| 6M | [F (7,95) =72.442, p≤0.001] | [F (7,95) =78.269, p≤0.001] | [F (7,95) =90.936, p≤0.001] |
Table 2.10.
Horizontal count represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
Table 2.10.
Horizontal count represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =23.069, p≤0.001] | [F (1,95) =20.677, p≤0.001] | [F (1,95) =32.187, p≤0.001] |
[F (1,95) =41.210, p≤0.001] |
[F (1,95) =43.336, p≤0.001] | [F (1,95) =35.695, p≤0.001] | [F (7195) =55.135, p≤0.001] | [F (1,95) =47.650, p≤0.001] | [F (1,95) =28.354, p≤0.001] | [F (1,95) =38.771, p≤0.001] |
| 3M | [F (1,95) =24.846, p≤0.001] | [F (1,95) =19.705, p≤0.001] | [F (1,95) =26.677, p≤0.001] |
[F (1,95) =32.427, p≤0.001] |
[F (1,95) =4.125, p≤0.001] | [F (1,95) =8.358, p≤0.001] | [F (1,95) =30.306, p≤0.001] | [F (1,95) =7.338, p≤0.001] | [F (1,95) =4.600, p≤0.001] | [F (1,95) =20.797, p≤0.001] |
| 6M | [F (1,95) =37.229, p≤0.001] | [F (1,95) 60.865, p≤0.001] | [F (1,95) =75.750, p≤0.001] | [F (1,95) =85.633, p≤0.001] | [F (1,95) =35.213, p≤0.001] | [F (1,95) =39.288, p≤0.001] | [F (1,95) =53.707, p≤0.001] | [F (1,95) =11.577, p≤0.001] | - | [F (1,95) =3.896, p≤0.001] |
Table 2.11.
Center zone entries OFT represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
Table 2.11.
Center zone entries OFT represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =19.513, p≤0.001] | [F (7,95) =23.682, p≤0.001] | [F (7,95) =33.439 p≤0.001] |
| 3M | [F (7,95) =19.025, p≤0.001] | [F (7,95) =41.747, p≤0.001] | [F (7,95) =52.746, p≤0.001] |
| 6M | [F (7,95) =22.483, p≤0.001] | [F (7,95) =35.849, p≤0.001] | [F (7,95) =50.219, p≤0.001] |
Table 2.12.
Center zone entries OFT represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
Table 2.12.
Center zone entries OFT represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =8.089, p≤0.001] | [F (1,95) =16.928, p≤0.001] | [F (1,95) =18.679, p≤0.001] | [F (1,95) =30.187, p≤0.001] | [F (1,95) =11.424, p≤0.001] | [F (1,95) =15.594, p≤0.001] | [F (1,95) =25.350, p≤0.001] | [F (1,95) =2.585, p=0.012] | [F (1,95) =5.003, p≤0.001] | [F (1,95) =3.252, p=0.002] |
| 3M | [F (1,95) =9.377, p≤0.001] | [F (1,95) =19.025, p≤0.001] | [F (1,95) =33.903, p≤0.001] | [F (1,95) =49.051, p≤0.001] | [F (1,95) =14.517, p≤0.001] | [F (1,95) =32.370, p≤0.001] | [F (1,95) =43.370, p≤0.001] | [F (1,95) =4.869, p≤0.001] | [F (1,95) =7.845, p≤0.001] | [F (1,95) =3.697, p≤0.001] |
| 6M | [F (1,95) =7.881, p≤0.001] | [F (1,95) =15.761, p≤0.001] | [F (1,95) =24.955, p≤0.001] | [F (1,95) =41.025, p≤0.001] | [F (1,95) =14.602, p≤0.001] | [F (1,95) =27.968, p≤0.001] | [F (1,95) =42.339, p≤0.001] | [F (1,95) =6.722, p≤0.001] | [F (1,95) =10.894, p≤0.001] | [F (1,95) =9.194, p≤0.001] |
Table 2.13.
Center zone time (sec) OFT represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
Table 2.13.
Center zone time (sec) OFT represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =18.531, p≤0.001] | [F (7,95) =15.732, p≤0.001] | [F (7,95) =24.720 p≤0.001] |
| 3M | [F (7,95) =18.608, p≤0.001] | [F (7,95) =17.114, p≤0.001] | [F (7,95) =26.103, p≤0.001] |
| 6M | [F (7,95) =17.300, p≤0.001] | [F (7,95) =18.887, p≤0.001] | [F (7,95) =33.352, p≤0.001] |
Table 2.14.
Center zone time (sec) OFT represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
Table 2.14.
Center zone time (sec) OFT represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =9.893, p≤0.001] | [F (1,95) =15.937, p≤0.001] | [F (1,95) =18.531, p≤0.001] | [F (1,95) =24.720, p≤0.001] | [F (1,95) =8.638, p≤0.001] | [F (1,95) =5.839, p≤0.001] | [F (1,95) =14.824, p≤0.001] | - | [F (1,95) =2.572, p=0.048] | [F (1,95) =3.606, p=0.005] |
| 3M | [F (1,95) =10.855, p≤0.001] | [F (1,95) =14.778, p≤0.001] | [F (1,95) =17.114, p≤0.001] | [F (1,95) =26.1031, p≤0.001] | [F (1,95) =7.753, p≤0.001] | [F (1,95) =6.259, p≤0.001] | [F (1,95) =15.248, p≤0.001] | [F (1,95) =3.830, p=0.002] | - | [F (1,95) =3.858, p=0.002] |
| 6M | [F (1,95) =11.701, p≤0.001] | [F (1,95) =14.194, p≤0.001] | [F (1,95) =16.460, p≤0.001] | [F (1,95) =33.352, p≤0.001] | [F (1,95) =5.599, p≤0.001] | [F (1,95) =7.186, p≤0.001] | [F (1,95) =21.651, p≤0.001] | [F (1,95) =2.706, p=0.042] | - | [F (1,95) =8.679, p≤0.001] |
Table 3.1.
% Open arm entries represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
Table 3.1.
% Open arm entries represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =8.049, p≤0.001] | [F (7,95) =12.819, p≤0.001] | [F (7,95) =19.974 p≤0.001] |
| 3M | [F (7,95) =23.156, p≤0.001] | [F (7,95) =16.259, p≤0.001] | [F (7,95) =23.156, p≤0.001] |
| 6M | [F (7,95) =9.237, p≤0.001] | [F (7,95) =14.676, p≤0.001] | [F (7,95) =23.732, p≤0.001] |
Table 3.2.
% Open arm entries represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
Table 3.2.
% Open arm entries represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =4.562, p≤0.001] | [F (1,95) =4.339, p≤0.001] | [F (1,95) =10.084, p≤0.001] | [F (1,95) =15.952, p≤0.001] | [F (1,95) =3.487, p=0.010] | [F (1,95) =8.257, p≤0.001] | [F (1,95) =15.412, p≤ 0.001] | [F (1,95) =3.710, p=009] | [F (1,95) =2.735, p=0.017] | [F (1,95) =4.022, p=0.001] |
| 3M | [F (1,95) =6.927, p≤0.001] | [F (1,95) =4.553, p≤0.001] | [F (1,95) =10.596, p≤0.001] | [F (1,95) =14.716, p≤0.001] | [F (1,95) =11.578, P≤0.001] | [F (1,95) =9.332, p≤0.001] | [F (1,95) =16.229, p≤0.001] | [F (1,95) =13.952, p≤0.001] | [F (1,95) =5.664, p≤0.001] | [F (1,95) =8.441, p≤0.001] |
| 6M | [F (1,95) =5.431, p=0.001] | [F (1,95) =4.894, p≤0.001] | [F (1,95) =9.152, p≤0.001] | [F (1,95) =23.732, p≤0.001] | [F (1,95) =3.806, p =0.001] | [F 1,95) =9.245, p≤0.001] | [F (1,95) =18.301, p≤0.001] | [F (1,95) =4.344, p≤0.001] | [F (1,95) =5.524, p=≤0.001] | [F (1,95) =7.197, p≤0.001] |
Table 3.3.
% Open arm time represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
Table 3.3.
% Open arm time represented as one-way ANOVA (comparison with Control) mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =20.987, p≤0.001] | [F (7,95) =29.663, p≤0.001] | [F (7,95) =41.150 p≤0.001] |
| 3M | [F (7,95) =30.792, p≤0.001] | [F (7,95) =103.914, p≤0.001] | [F (7,95) =126.484, p≤0.001] |
| 6M | [F (7,95) =12.004, p≤0.001] | [F (7,95) =12.755, p≤0.001] | [F (7,95) =18.094, p≤0.001] |
Table 3.4.
% Open arm time represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
Table 3.4.
% Open arm time represented as two-way ANOVA (comparison between diet and infection) mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =3.573, p=0.004] | - | [F (1,95) =8.545, p≤0.001] | [F (1,95) =20.334, p≤0.001] | [F (1,95) =17.414, p≤0.001] | [F (1,95) =26.091, p≤0.001] | [F (1,95) =37.577, p≤ 0.001] | [F (1,95) =18.738, p≤0.001] | [F (1,95) =21.118, p≤0.001] | [F (1,95) =20.816, p≤0.001] |
| 3M | [F (1,95) =14.714, p≤0.001] | [F (1,95) =14.147, p≤0.001] | [F (1,95) =101.005, p≤0.001] | [F (1,95) =108.576, p≤0.001] | [F (1,95) =16.506, p p≤0.001] | [F (1,95) =89.628, p p≤0.001] | [F (1,95) =112.198, p≤0.001] | [F (1,95) =16.644, p≤0.001] | - | [F (1,95) =17.908, p≤0.001] |
| 6M | [F (1,95) =5.454, p≤0.001] | [F (1,95) =7.897, p≤0.001] | [F (1,95) =8.960, p≤0.001] | [F (1,95) =13.212, p≤0.001] | [F (1,95) =6.550, p = p≤0.001] | [F (1,95) =7.301, p≤0.001] | [F (1,95) =12.650, p≤0.001] | [F (1,95) =4.107, p≤0.001] | [F (1,95) =3.795, p=0.002] | [F (1,95) =4.883, p≤0.001] |
Table 3.5.
Center zone entries represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
Table 3.5.
Center zone entries represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =8.948, p≤0.001] | [F (7,95) =11.968, p≤0.001] | [F (7,95) =19.565 p≤0.001] |
| 3M | [F (7,95) =8.635, p≤0.001] | [F (7,95) =17.375, p≤0.001] | [F (7,95) =26.115, p≤0.001] |
| 6M | [F (7,95) =21.089, p≤0.001] | [F (7,95) =27.725, p≤0.001] | [F (7,95) =13.933, p≤0.001] |
Table 3.6.
Center zone entries represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
Table 3.6.
Center zone entries represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =4.059, p=0.001] | [F (1,95) =5.828, p≤0.001] | [F (1,95) =5.412, p≤0.001] | [F (1,95) =16.339, p≤0.001] | [F (1,95) =4.889, p≤0.001] | [F (1,95) =7.909, p≤0.001] | [F (1,95) =15.507, p≤ 0.001] | [F (1,95) =3.120, p=0.013] | [F (1,95) =6.556, p≤0.001] | [F (1,95) =3.226, p =0.011] |
| 3M | [F (1,95) =5.477, p≤0.001] | - | [F (1,95) =7.793, p≤0.001] | [F (1,95) =22.744, p≤0.001] | [F (1,95) =3.158, p=0.009] | [F (1,95) =11.898, p≤0.001] | [F (1,95) =20.638, p≤ 0.001] | [F (1,95) =6.740, p≤0.001] | [F (1,95) =9.582, p≤0.001] | [F (1,95) =3.371, p =0.006] |
| 6M | [F (1,95) =7.635, p≤0.001] | [F (1,95) =17.362, p p≤0.001] | [F (1,95) =25.452, p≤0.001] | [F (1,95) =32.543, p≤0.001] | [F (1,95) =13.453, p≤0.001] | [F (1,95) =20.089, p≤0.001] | [F (1,95) =30.361, p≤ 0.001] | [F (1,95) =3.727, p≤0.001] | [F (1,95) =2.273, p=0.026] | [F (1,95) =5.454, p≤0.001] |
Table 3.7.
Center zone time (sec) represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
Table 3.7.
Center zone time (sec) represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =13.816, p≤0.001] | [F (7,95) =13.262, p≤0.001] | [F (7,95) =18.227 p≤0.001] |
| 3M | [F (7,95) =16.950, p≤0.001] | [F (7,95) =18.527, p≤0.001] | [F (7,95) =27.690, p≤0.001] |
| 6M | [F (7,95) =25.785, p≤0.001] | [F (7,95) =28.142, p≤0.001] | [F (7,95) =43.407, p≤0.001] |
Table 3.8.
Center zone time (sec) represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
Table 3.8.
Center zone time (sec) represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =5.534, p≤0.001] | [F (1,95) =10.791, p≤0.001] | [F (1,95) =13.037, p≤0.001] | [F (1,95) =14.542, p≤0.001] | [F (1,95) =8.282, p≤0.001] | [F (1,95) =7.728, p≤0.001] | [F (1,95) =12.693, p≤ 0.001] | [F (1,95) =3.025, p=0.031] | - | [F (1,95) =3.684, p =0.004] |
| 3M | [F (1,95) =7.918, p≤0.001] | [F (1,95) =10.902, p≤0.001] | [F (1,95) =14.933, p≤0.001] | [F (1,95) =21.890, p≤0.001] | [F (1,95) =9.032, p≤0.001] | [F (1,95) =10.609, p≤0.001] | [F (1,95) =19.772, p≤ 0.001] | [F (1,95) =6.048, p≤ 0.001] | [F (1,95) =3.594, p=0.003] | [F (1,95) =5.800, p≤ 0.001] |
| 6M | [F (1,95) =15.265, p≤0.001] | [F (1,95) =23.219, p≤0.001] | [F (1,95) =20.098, p≤0.001 | [F (1,95) =35.273, p≤0.001] | [F (1,95) =10.521, p≤0.001] | [F (1,95) =12.878, p≤0.001] | [F (1,95) =28.142, p≤ 0.001] | [F (1,95) =2.566, p=0.025] | [F (1,95) =8.044, p≤ 0.001] | [F (1,95) =8.134, p≤ 0.001] |
Table 4.1.
% Light zone entries represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
Table 4.1.
% Light zone entries represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =11.996, p≤0.001] | [F (7,95) =12.475, p≤0.001] | [F (7,95) =15.620 p≤0.001] |
| 3M | [F (7,95) =9.523, p≤0.001] | [F (7,95) =10.163, p≤0.001] | [F (7,95) =16.124, p≤0.001] |
| 6M | [F (7,95) =11.723, p≤0.001] | [F (7,95) =12.827, p≤0.001] | [F (7,95) =19.761, p≤0.001] |
Table 4.2.
% Light zone entries represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
Table 4.2.
% Light zone entries represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =4.020, p=0.001] | [F (1,95) =4.722, p≤0.001] | [F (1,95) =7.770, p≤0.001] | [F (1,95) =11.052, p≤0.001] | [F (1,95) =7.976, p≤0.001] | [F (1,95) =8.455, p≤0.001] | [F (1,95) =11.599, p≤ 0.001] | [F (1,95) =7.274, p≤0.001] | [F (1,95) =4.705, p≤0.001] | [F (1,95) =4.567, p≤ 0.001] |
| 3M | [F (1,95) =5.345, p≤0.001] | [F (1,95) =3.643, p=0.004] | [F (1,95) =4.558, p≤0.001] | [F (1,95) =11.094, p≤0.001] | [F (1,95) =4.178, p≤0.001] | [F (1,95) =4.817, p≤0.001] | [F (1,95) =10.779, p≤0.001] | [F (1,95) =5.880, p≤0.001] | [F (1,95) =5.605, p≤0.001] | [F (1,95) =5.030, p≤0.001] |
| 6M | [F (1,95) =6.172, p≤0.001] | [F (1,95) =7.316, p≤0.001] | [F (1,95) =8.367, p≤0.001] | [F (1,95) =14.752, p≤0.001] | [F (1,95) =5.551, p≤0.001] | [F (1,95) =6.655, p≤0.001] | [F (1,95) =13.619, p≤0.001] | [F (1,95) =4.408, p≤0.001] | [F (1,95) =4.460, p≤0.001] | [F (1,95) =5.039, p≤0.001] |
Table 4.3.
% Light zone time (sec) represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
Table 4.3.
% Light zone time (sec) represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 1M | [F (7,95) =8.413, p≤0.001] | [F (7,95) =9.659, p≤0.001] | [F (7,95) =9.511 p≤0.001] |
| 3M | [F (7,95) =9.121, p≤0.001] | [F (7,95) =10.434, p≤0.001] | [F (7,95) =13.099, p≤0.001] |
| 6M | [F (7,95) =13.8112, p≤0.001] | [F (7,95) =20.132, p≤0.001] | [F (7,95) =67.694, p≤0.001] |
Table 4.4.
% Light zone time (sec) represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
Table 4.4.
% Light zone time (sec) represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1M | [F (1,95) =4.856, p≤0.001] | [F (1,95) =4.287, p≤0.001] | [F (1,95) =5.543, p≤0.001] | [F (1,95) =9.548, p≤0.001] | [F (1,95) =3.557, p≤0.001] | [F (1,95) =4.265, p≤0.001] | [F (1,95) =8.956, p≤0.001] | [F (1,95) p≤0.001=4.124] | [F (1,95) =3.578, p≤0.001] | [F (1,95) =4.265, p≤0.001] |
| 3M | [F (1,95) =4.480, p≤0.001] | [F (1,95) =5.040.467, p≤0.001] | [F (1,95) =6.210, p≤0.001] | [F (1,95) =12.388, p≤0.001 | [F (1,95) =5.180, p≤0.001] | [F (1,95) =5.954, p≤0.001] | [F (1,95) =15.652, p≤0.001] | [F (1,95) =4.619, p p≤0.001] | [F (1,95) =4.224, p≤0.001] | [F (1,95) =7.744, p≤0.001] |
| 6M | [F (1,95) =5.798, p≤0.001] | [F (1,95) =6.054, p≤0.001] | [F (1,95) =6.490, p≤0.001] | [F (1,95) =17.577, p≤0.001] | [F (1,95) =3.713, p≤0.001] | [F (1,95) =7.301, p≤0.001] | [F (1,95) =16.261, p≤0.001] | [F (1,95) =3.457, p≤0.001] | [F (1,95) =6.607, p≤0.001] | [F (1,95) =4.482, p≤0.001] |
Table 5.1.
Retention time (sec) represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
Table 5.1.
Retention time (sec) represented one-way ANOVA (comparison with Control) as mean values along with level of significance.
| Age | Control vs LP+LPS |
Control vs LP+DLT |
Control vs LP+LPS+DLT |
|---|---|---|---|
| 3M | [F (7,95) =8.476, p≤0.001] | [F (7,95) =10.659, p≤0.001] | [F (7,95) =13.238, p≤0.001] |
| 6M | [F (7,95) =14.982, p≤0.001] | [F (7,95) =14.755, p≤0.001] | [F (7,95) =28.235, p≤0.001] |
Table 5.2.
Retention time (sec) represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
Table 5.2.
Retention time (sec) represented two-way ANOVA (comparison between diet and infection) as mean values along with level of significance.
| Age | Control vs LP |
Control vs Control+LPS |
Control vs Control+DLT |
Control vs Control+DLT+LPS |
LP vs LP+LPS |
LP vs LP+DLT |
LP vs LP+DLT+LPS |
Control+LPS vs LP+LPS |
Control+DLT vs LP+DLT |
Control+DLT+LPS vs LP+DLT+LPS |
|---|---|---|---|---|---|---|---|---|---|---|
| 3M | [F (1,95) =3.542, p≤0.001] | [F (1,95) =4.801, p≤0.001] | [F (1,95) =5.486, p≤0.001] | [F (1,95) =9.791, p≤0.001] | [F (1,95) =4.447, p≤0.001] | [F (1,95) =6.630, p= 0.001] | [F (1,95) =9.209, p≤ 0.001] | [F (1,95) =3.175, p= 0.002] | [F (1,95) =4.666, p≤0.001] | [F (1,95) =3.447, p= 0.001] |
| 6M | [F (1,95) =4.431, p≤0.001] | [F (1,95) =7.044, p≤0.001] | [F (1,95) =10.955, p≤0.001] | [F (1,95) =20.354, p≤0.001] | [F (1,95) =11.616, p≤0.001] | [F (1,95) =11.373, p≤0.001] | [F (1,95) =25.811, p≤0.001] | [F (1,95)) =9.003, p≤0.001] | [F (1,95) =4.849, p≤0.001] | [F (1,95) =9.888, p≤0.001] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



