Submitted:
25 August 2023
Posted:
29 August 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Delineating the epidemic of vaccine injury from the coterminous condition long covid is a challenging prospect, but one with many implications not just for treatment, but also has important legal considerations for settlements of vaccine injury. The shared etiological factor of the spike protein in both vaccine injury and long covid make differentiation difficult, and while treatment is largely similar between vaccine injury and long covid, there are important distinctions. Furthermore, diagnostics are important for monitoring treatment progress and assessing the extent of subclinical vaccine injury in population having received a covid-19 vaccine. The development of rigorous diagnostics is an important step towards the recognition of both long covid and vaccine injury, as those suffering these conditions have faced immense challenges in having their conditions recognized, treated, and compensated by insurance companies or national health services.
Keywords:
Vaccine adverse event
; Covid-19
; spike protein
; diagnosis
Introduction
Adverse events after vaccination have been far more common with Covid-19 vaccines than any other licensed vaccine[1]. Not only are the rates of clinical conditions associated with Covid-19 vaccines high, but there is also extensive subclinical damage. Due to the pathological mechanisms of the vaccine encoded spike protein, the potential for damage can exist at low levels for long periods of time, and those having received a vaccine can be in a ‘sword of Damocles’ situation for years or even decades. Frankly, there is a lot left unknown about the long-term effects of Covid-19 vaccines[2].
The extent of subclinical danger, as well as the increase in sudden and unexplained deaths motivates the diagnosis of vaccine injury through biomarkers. One immediate biomarker that comes to the fore is testing for the presence of the spike protein or its subunits in plasma[3], as it is a major pathological agent driving vaccine injury, long covid, as well as acute covid-19 infection[4].
This conflation has important legal implications for those seeking compensation after injury following Covid-19 vaccination. Pathologists can assign causality to covid-19 vaccination (as opposed to infection) by immunostaining for the nucleocapsid (N) protein in addition to the spike (S) protein[5]. Since Covid-19 infection will express both N and S proteins, whereas vaccination only expresses the S protein, the simultaneous presence of the S protein and absence of the N protein is strong evidence for vaccine induced causation[5].
Furthermore, there are two important differentiations between the spike protein induced by vaccination and the spike protein from infection. First, the viral spike protein will change as the virus mutates, whereas the vaccine spike protein only changes when the sequence is updated (as with the bivalent booster). Secondly, the vaccine spike protein is locked into a prefusion conformation through two proline mutations, and will adopt a more rigid conformation than the viral spike protein[6].
There are three important ways to differentiate spike protein from viral infection from that of the vaccine. The following factors can be used to differentiate vaccine damage from viral damage (Table 1).
Diagnostics
General
In guiding treatment, there are multiple biomarkers that one can test with to gain insight into the progression of the injury sustained from the vaccine. These are non-specific to vaccine injury and are general biomarkers of cardiovascular risk. These include troponin, D-dimer and C-reactive protein[3].
[4]. These biomarkers are specific to cardiac injury, and will not be able to determine disease aetiology.
Troponin is a general biomarker associated with diagnosis of acute coronary syndromes[7,8], as troponins are released into the blood following damage to cardiac muscle[9]. D-dimer is a biomarker associated with the breakdown of fibrin clots by the fibrinolytic system [10]. As the test measures breakdown of clots, a high measure can indicate a high level of clot burden, as well as a high degree of breakdown[11], and this must be taken into consideration by the clinician.
C-reactive protein is an inflammatory biomarker, and higher values are associated with increased cardiovascular risk[12].

Table 2.
Altered biomarkers in vaccine injury. * For patients 50 years or older.
| Biomarker | Upper limit of normal |
| Peak cardiac troponin (T) | 14 ng/L [3] |
| Brain natriuretic Peptide (BNP) |
100pg/mL [3] |
| N-terminal prohormone of brain natriuretic peptide (NT-proBNP) | 450pg/mL [3] |
| C-reactive protein (CRP) | 8mg/L [3] |
| D-dimer | (patient’s age in years x 10mcg/L)* [13] |
Specific
A recent paper by Yonker surveyed the biomarkers of vaccinated individuals, both with and without post-vaccination myocarditis. The main differentiator between the group with myocarditis and those without was the persistence of full length spike protein, unbound by antibodies[3]. Given that this is the sole gene encoded by most of the vaccines and has multiple documented pathological mechanisms[4], it is a likely aetiological factor in post-vaccination syndrome.
Cases of blood thrombosis after vaccination typically occur within one month of receiving the injection[14,15]. A test for spike protein contains two important quantities, the concentration of spike protein, as well as the time since vaccination. While most often after injection spike protein concentration drops off quickly after one week [16], persistence of high levels of spike protein for months after injection has been documented in a subset of vaccinated individuals [17]. It is unclear what the individual factors are affecting long-term spike protein levels; we propose a model for the long term persistence of spike protein.
The first factors are the variations in the initial dose of spike protein encoding mRNA, which can vary due to storage, dilution and administration. Once the mRNA is in the body, the level of spike is in competition between mRNA degradation and protein expression from the mRNA (Figure 1). We also propose a third alternative between degradation and expression, that of conversion to a reservoir. Reverse transcription into the genome is possible [18]. Additionally, a discovery of DNA contamination in a broad swathe of mRNA vaccine vials[19], potentially opening the possibility of but microbiota transfection through the mechanism of horizontal gene transfer [20].
While the half-life of RNA is well known, and endogenous mRNA has a half life of approximately 10 hours [21], it is known that pseudouridinylated RNA is far more persistent[22,23], and less is known about the degradation of the N1-methyl-psuedouridnylated RNA used in the mRNA vaccines[2]. Persistence of spike protein appears to be the factor which differentiates those with post-vaccination myocarditis vs vaccinated people without myocarditis[3].
- Causes of interpersonal variation
In interpreting the causes of interpersonal variation in vaccine response, we will restrict our analysis to disease etiologies tracing to the spike protein in the vaccines. While there may be other contributing factors, they are outside the scope of this review.
- Administration and dosing
Variation in the dosing of the vaccine can greatly influence the response to the vaccine. It is observed that there was significant variation in the adverse event rate of different batches. Furthermore, the rates of AEs differed significantly depending on whether or not aspiration was performed. A strong relationship has also been shown between the dose of mRNA received and the rates of AEs, both when comparing brands (Moderna Covid-19 vaccine delivers a higher dose of mRNA than the Pfizer BNT162b2) and when comparing AE rates by total cumulative dose (i.e. comparing those receiving boosters with those receiving the initial series).
However, assuming the delivery is constant, there are still important sources of individual variation which can impact the vaccination response.
- Individual variation
Viewing the series of events sequentially, the RNA must enter the cell, be read by the ribosome to express spike protein, after which it is degraded (in principle). Lacking an intact degradation pathway, the modified RNA will linger around and keep producing spike protein in principle. Minimal information is known about the biodegradation of modified mRNA in humans [2].
- Variation in cell uptake and biodistribution
The most important factor here is whether the injection was performed into the deltoid muscle, as intended, or some escaped into the bloodstream, which is largely set by the conditions of the injection itself (if aspiration was standard procedure). Still, individuals vary in their cellular affinity to take up extracellular RNA via endocytosis [24,25,26]. While it is unclear of their relevance in this case, there are genetic variations in endocytic pathways [27,28]
It is possible that gross blood flow parameters, such as coagulability and blood pressure influence biodistribution. As hypercoagulation is an etiology behind vaccine injury[29] and long covid[30], genetic factors underlying coagulability may be associated with the prognosis of either condition. A Genome wide association study (GWAS) found that gene polymorphisms of proteins involved in coagulation were associated with acute Covid-19 prognosis[31]. These include MBL2[32], ADAMTS13[33], F8[34] and PDGFRL[34], which are involved in clotting and have also been observed to be associated with acute Covid-19 severity. Our search reveals limited GWAS studies of long covid[35,36] or Covid-19 vaccine adverse events[37], though GWAS studies of SARS-CoV-2 vaccine antibody responses have been performed[38,39] and several mechanistic factors behind the differential vaccine response are identified [40]. Genetic correlates of vaccine adverse reactions have been studied in the context of Hepatitis B, measles, rubella, and other vaccines [41].
Several factors impact the immune response to Covid-19 vaccines, including time of day[42,43], level of recent sleep [44,45,46,47] and prior infection[48]. Several interventions have undergone clinical trials to determine their impact on the vaccine immune response, including iron supplementation (NCT04915820), mushroom mixtures (NCT04951336), immunomodulators (NCT04877496 [49] and NCT05060991), the ketogenic diet (NCT05163743), metformin (NCT03996538), probiotics (NCT05195151) and osteopathy (NCT04928456 and NCT05069636). Of these trials, only the trial on the immune impact of anti-CD20 therapy has results, which were not significantly different from controls [49].
The existing study on genetic determinants of Covid-19 vaccine adverse events is very limited[37]; likewise, we may have to proceed from mechanistic understanding. After endocytosis, the modified mRNA is present in the cytosol to be expressed, which can be highly variable depending on host genetics[50,51]. From this point on, the major factor regulating the amount of pathogenic spike protein produced are largely the clearance and degradation mechanisms acting on the chemically modified mRNA. As mentioned before, while pseudouridinylated RNA is far more stable than RNA[22], the degradation curve of N1-methyl pseudouridinylated RNA is less known, though N1-methyl-pseudouridine in mRNA has been observed to enhance translation[52]. Individual genetic variation influences the degradation of exogenous RNA [53].
Furthermore, other wildcards include the potential for reverse transcription into the host genome[18]. Another point of concern is the observed contamination of mRNA vaccine vials with the DNA plasmid vector[19], which may possibly be a factor in the observed persistence of spike protein in vaccine injured patients [3]. This could be explained by transfection of bacteria in the human gut [54,55,56]. Were this a significant factor, we would expect differential vaccine injury susceptibility based on gut microbiota composition. Long covid susceptibility varies according to gut barrier health[57], and acute SARS-CoV-2 infection severity is negatively associated with gut microbial diversity[58]. Still, it is unknown to what degree this ‘gut reservoir’ hypothesis for spike protein persistence means. Long covid is associated with higher levels of Ruminococcus gnavus, Bacteroides vulgatus and lower levels of Faecalibacterium prausnitzii [59].
Causation
There is an unprecedented wave of vaccine injury, in addition to any disease burden from long Covid. The origins of Covid notwithstanding, establishing causation for those experiencing vaccine injury is an important step both in allowing them to receive just compensation, as well as to establish the true role of vaccination in the mortality and morbidity burdens. The latter is useful both for informing regulatory policy going forward, as it is necessary for regulators and the public to know the true risk profile of this class of intervention. Additionally, establishing causation is useful for legal settings, including compensation for injured recipients as well as prosecution of any wrongdoing. The extent of liability is important, as those seeking treatment often have few options, and little resources, owing to the often debilitating nature of their illness, and its lack of acknowledgement and subsequent compensation by health systems[60]. Vaccine injury compensation schemes are uncommon [61,62].
The experience of the vaccine injured has largely been one of gaslighting and being ignored, and only now are their concerns being heard[60]. Still, treatment is limited, and limited resources exit for injury compensation [63]. Treatment of long Covid is receiving some attention and research funds [64], while treatment of vaccine injury is limited. For example, in the US clinical trials database (clinicaltrials.gov, accessed July 11, 2023) there is currently one study to test treatment of Covid-19 vaccine injury; the study is not yet recruiting and was last updated May 24, 2022. Multiple studies exist to treat long Covid, reviewed in [65]. No large university hospital or academic medical center has published a treatment protocol for vaccine injury, and the current literature is scant [65].
Conclusion
While the situation of the vaccine injured presents a pessimistic view, the situation is improving. Vaccine injury is increasingly being recognized, as a recent acknowledgement by German health minister Karl Lauterbach exemplifies [66]. Still, for those affected, it is a long road to recovery. Diagnostics are a necessary part of the path towards health in those experiencing post-vaccination syndrome and long covid. In some cases, the diagnostics are similar, but the potential also exists to discriminate the two conditions with diagnostics, as well as by patient history.
Developing rigorous diagnostics is an important step towards gauging treatment progress and informing the science of treating vaccine injury, as well as long covid. Diagnostic development ensures that those affected receive the recognition and treatment they deserve, and ensures the integrity of compensation claims, a and can inform legal action against regulators, pharmaceutical manufacturers and public health officials.
References
- Kim, M.S.; Jung, S.Y.; Ahn, J.G.; Park, S.J.; Shoenfeld, Y.; Kronbichler, A.; Koyanagi, A.; Dragioti, E.; Tizaoui, K.; Hong, S.H.; et al. Comparative Safety of MRNA COVID-19 Vaccines to Influenza Vaccines: A Pharmacovigilance Analysis Using WHO International Database. Journal of Medical Virology 2022, 94, 1085–1095. https://doi.org/10.1002/jmv.27424. [CrossRef]
- Halma, M.T.J.; Rose, J.; Lawrie, T. The Novelty of MRNA Viral Vaccines and Potential Harms: A Scoping Review. J 2023, 6, 220–235. https://doi.org/10.3390/j6020017. [CrossRef]
- Yonker, L.M.; Swank, Z.; Bartsch, Y.C.; Burns, M.D.; Kane, A.; Boribong, B.P.; Davis, J.P.; Loiselle, M.; Novak, T.; Senussi, Y.; et al. Circulating Spike Protein Detected in Post–COVID-19 MRNA Vaccine Myocarditis. Circulation 2023, 147, 867–876. https://doi.org/10.1161/CIRCULATIONAHA.122.061025. [CrossRef]
- Theoharides, T.C.; Conti, P. Be Aware of SARS-CoV-2 Spike Protein: There Is More than Meets the Eye. J Biol Regul Homeost Agents 2021, 35, 833–838. https://doi.org/10.23812/THEO_EDIT_3_21. [CrossRef]
- Mörz, M. A Case Report: Multifocal Necrotizing Encephalitis and Myocarditis after BNT162b2 MRNA Vaccination against COVID-19. Vaccines 2022, 10, 1651. https://doi.org/10.3390/vaccines10101651. [CrossRef]
- Riley, T.P.; Chou, H.-T.; Hu, R.; Bzymek, K.P.; Correia, A.R.; Partin, A.C.; Li, D.; Gong, D.; Wang, Z.; Yu, X.; et al. Enhancing the Prefusion Conformational Stability of SARS-CoV-2 Spike Protein Through Structure-Guided Design. Frontiers in Immunology 2021, 12. [CrossRef]
- Mahajan, V.S.; Jarolim, P. How to Interpret Elevated Cardiac Troponin Levels. Circulation 2011, 124, 2350–2354. https://doi.org/10.1161/CIRCULATIONAHA.111.023697. [CrossRef]
- Melanson, S.E.F.; Morrow, D.A.; Jarolim, P. Earlier Detection of Myocardial Injury in a Preliminary Evaluation Using a New Troponin I Assay with Improved Sensitivity. Am J Clin Pathol 2007, 128, 282–286. https://doi.org/10.1309/Q9W5HJTT24GQCXXX. [CrossRef]
- Apple, F.S.; Sandoval, Y.; Jaffe, A.S.; Ordonez-Llanos, J.; IFCC Task Force on Clinical Applications of Cardiac Bio-Markers Cardiac Troponin Assays: Guide to Understanding Analytical Characteristics and Their Impact on Clinical Care. Clin Chem 2017, 63, 73–81. https://doi.org/10.1373/clinchem.2016.255109. [CrossRef]
- Thachil, J.; Lippi, G.; Favaloro, E.J. D-Dimer Testing: Laboratory Aspects and Current Issues. In Hemostasis and Thrombosis: Methods and Protocols; Favaloro, E.J., Lippi, G., Eds.; Methods in Molecular Biology; Springer: New York, NY, 2017; pp. 91–104 ISBN 978-1-4939-7196-1.
- Linkins, L.-A.; Takach Lapner, S. Review of D-Dimer Testing: Good, Bad, and Ugly. International Journal of Laboratory Hematology 2017, 39, 98–103. https://doi.org/10.1111/ijlh.12665. [CrossRef]
- Ridker, P.M. A Test in Context. Journal of the American College of Cardiology 2016, 67, 712–723. https://doi.org/10.1016/j.jacc.2015.11.037. [CrossRef]
- Urban, K.; Kirley, K.; Stevermer, J.J. PURLs: It’s Time to Use an Age-Based Approach to D-Dimer. J Fam Pract 2014, 63, 155–158.
- Bilotta, C.; Perrone, G.; Adelfio, V.; Spatola, G.F.; Uzzo, M.L.; Argo, A.; Zerbo, S. COVID-19 Vaccine-Related Thrombosis: A Systematic Review and Exploratory Analysis. Front Immunol 2021, 12, 729251. https://doi.org/10.3389/fimmu.2021.729251. [CrossRef]
- Mani, A.; Ojha, V. Thromboembolism after COVID-19 Vaccination: A Systematic Review of Such Events in 286 Patients. Annals of Vascular Surgery 2022, 84, 12-20.e1. https://doi.org/10.1016/j.avsg.2022.05.001. [CrossRef]
- Ogata, A.F.; Cheng, C.-A.; Desjardins, M.; Senussi, Y.; Sherman, A.C.; Powell, M.; Novack, L.; Von, S.; Li, X.; Baden, L.R.; et al. Circulating Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Vaccine Antigen Detected in the Plasma of MRNA-1273 Vaccine Recipients. Clinical Infectious Diseases 2022, 74, 715–718. https://doi.org/10.1093/cid/ciab465. [CrossRef]
- Bansal, S.; Perincheri, S.; Fleming, T.; Poulson, C.; Tiffany, B.; Bremner, R.M.; Mohanakumar, T. Cutting Edge: Circulating Exosomes with COVID Spike Protein Are Induced by BNT162b2 (Pfizer–BioNTech) Vaccination Prior to Development of Antibodies: A Novel Mechanism for Immune Activation by MRNA Vaccines. The Journal of Immunology 2021, 207, 2405–2410. https://doi.org/10.4049/jimmunol.2100637. [CrossRef]
- Aldén, M.; Olofsson Falla, F.; Yang, D.; Barghouth, M.; Luan, C.; Rasmussen, M.; De Marinis, Y. Intracellular Reverse Transcription of Pfizer BioNTech COVID-19 MRNA Vaccine BNT162b2 In Vitro in Human Liver Cell Line. Curr Issues Mol Biol 2022, 44, 1115–1126. https://doi.org/10.3390/cimb44030073. [CrossRef]
- McKernan, K.; Helbert, Y.; Kane, L.T.; McLaughlin, S. Sequencing of Bivalent Moderna and Pfizer MRNA Vaccines Reveals Nanogram to Microgram Quantities of Expression Vector DsDNA per Dose. OSF Preprints. April 2023, 10. [CrossRef]
- Brito, I.L. Examining Horizontal Gene Transfer in Microbial Communities. Nat Rev Microbiol 2021, 19, 442–453. https://doi.org/10.1038/s41579-021-00534-7. [CrossRef]
- Lugowski, A.; Nicholson, B.; Rissland, O.S. Determining MRNA Half-Lives on a Transcriptome-Wide Scale. Methods 2018, 137, 90–98. https://doi.org/10.1016/j.ymeth.2017.12.006. [CrossRef]
- Karikó, K.; Muramatsu, H.; Welsh, F.A.; Ludwig, J.; Kato, H.; Akira, S.; Weissman, D. Incorporation of Pseudouridine Into MRNA Yields Superior Nonimmunogenic Vector With Increased Translational Capacity and Biological Stability. Molecular Therapy 2008, 16, 1833–1840. https://doi.org/10.1038/mt.2008.200. [CrossRef]
- Leppek, K.; Byeon, G.W.; Kladwang, W.; Wayment-Steele, H.K.; Kerr, C.H.; Xu, A.F.; Kim, D.S.; Topkar, V.V.; Choe, C.; Rothschild, D.; et al. Combinatorial Optimization of MRNA Structure, Stability, and Translation for RNA-Based Therapeutics. Nat Commun 2022, 13, 1536. https://doi.org/10.1038/s41467-022-28776-w. [CrossRef]
- Fang, E.; Liu, X.; Li, M.; Zhang, Z.; Song, L.; Zhu, B.; Wu, X.; Liu, J.; Zhao, D.; Li, Y. Advances in COVID-19 MRNA Vaccine Development. Sig Transduct Target Ther 2022, 7, 1–31. https://doi.org/10.1038/s41392-022-00950-y. [CrossRef]
- Patel, S.; Kim, J.; Herrera, M.; Mukherjee, A.; Kabanov, A.V.; Sahay, G. Brief Update on Endocytosis of Nanomedicines. Advanced Drug Delivery Reviews 2019, 144, 90–111. https://doi.org/10.1016/j.addr.2019.08.004. [CrossRef]
- Doherty, G.J.; McMahon, H.T. Mechanisms of Endocytosis. Annual Review of Biochemistry 2009, 78, 857–902. https://doi.org/10.1146/annurev.biochem.78.081307.110540. [CrossRef]
- Beyer, A.; Koch, T.; Schröder, H.; Schulz, S.; Höllt, V. Effect of the A118G Polymorphism on Binding Affinity, Potency and Agonist-Mediated Endocytosis, Desensitization, and Resensitization of the Human Mu-Opioid Receptor. Journal of Neurochemistry 2004, 89, 553–560. https://doi.org/10.1111/j.1471-4159.2004.02340.x. [CrossRef]
- Zhan, L.; Li, J.; Jew, B.; Sul, J.H. Rare Variants in the Endocytic Pathway Are Associated with Alzheimer’s Disease, Its Related Phenotypes, and Functional Consequences. PLOS Genetics 2021, 17, e1009772. https://doi.org/10.1371/journal.pgen.1009772. [CrossRef]
- Aladdin, Y.; Algahtani, H.; Shirah, B. Vaccine-Induced Immune Thrombotic Thrombocytopenia with Disseminated Intravascular Coagulation and Death Following the ChAdOx1 NCoV-19 Vaccine. Journal of Stroke and Cerebrovascular Diseases 2021, 30, 105938. https://doi.org/10.1016/j.jstrokecerebrovasdis.2021.105938. [CrossRef]
- Turner, S.; Khan, M.A.; Putrino, D.; Woodcock, A.; Kell, D.B.; Pretorius, E. Long COVID: Pathophysiological Factors and Abnormalities of Coagulation. Trends in Endocrinology & Metabolism 2023, 34, 321–344. https://doi.org/10.1016/j.tem.2023.03.002. [CrossRef]
- Pairo-Castineira, E.; Rawlik, K.; Bretherick, A.D.; Qi, T.; Wu, Y.; Nassiri, I.; McConkey, G.A.; Zechner, M.; Klaric, L.; Griffiths, F.; et al. GWAS and Meta-Analysis Identifies 49 Genetic Variants Underlying Critical COVID-19. Nature 2023, 617, 764–768. https://doi.org/10.1038/s41586-023-06034-3. [CrossRef]
- Hultström, M.; Frithiof, R.; Grip, J.; Lindelöf, L.; Rooijackers, O.; Pigazzini, S.; Niemi, M.; Cordioli, M.; Nkambule, L.; Maricic, T.; et al. Genetic Determinants of Mannose-Binding Lectin Activity Predispose to Thromboembolic Complications in Critical COVID-19. Nat Immunol 2022, 23, 861–864. https://doi.org/10.1038/s41590-022-01227-w. [CrossRef]
- Mancini, I.; Baronciani, L.; Artoni, A.; Colpani, P.; Biganzoli, M.; Cozzi, G.; Novembrino, C.; Boscolo Anzoletti, M.; De Zan, V.; Pagliari, M.T.; et al. The ADAMTS13-von Willebrand Factor Axis in COVID-19 Patients. J Thromb Haemost 2021, 19, 513–521. https://doi.org/10.1111/jth.15191. [CrossRef]
- Bordon, Y. A New Set of Genes Linked to Critical COVID-19. Nat Rev Immunol 2022, 22, 208–208. https://doi.org/10.1038/s41577-022-00709-0. [CrossRef]
- Vilma Lammi; Tomoko Nakanishi; Samuel E. Jones; Shea J. Andrews; Juha Karjalainen; Beatriz Cortés; Heath E. O’Brien; Brian E. Fulton-Howard; Hele H. Haapaniemi; Axel Schmidt; et al. Genome-Wide Association Study of Long COVID. medRxiv 2023, 2023.06.29.23292056. https://doi.org/10.1101/2023.06.29.23292056. [CrossRef]
- Ledford, H. Gene Linked to Long COVID Found in Analysis of Thousands of Patients. Nature 2023, 619, 445–445. https://doi.org/10.1038/d41586-023-02269-2. [CrossRef]
- Shun Nogawa; Hajime Kanamori; Koichi Tokuda; Kaoru Kawafune; Miyuki Chijiiwa; Kenji Saito; Shoko Takahashi Identification of Susceptibility Loci for Adverse Events Following COVID-19 Vaccination in the Japanese Population: A Web-Based Genome-Wide Association Study. medRxiv 2021, 2021.11.30.21267043. https://doi.org/10.1101/2021.11.30.21267043. [CrossRef]
- Li, P.; Shi, D.; Shen, W.; Shi, S.; Guo, X.; Li, J.; Xu, S.; Zhang, Y.; Zhao, Z. Pilot Genome-Wide Association Study of Antibody Response to Inactivated SARS-CoV-2 Vaccines. Frontiers in Immunology 2022, 13. [CrossRef]
- Delshad, M.; Sanaei, M.-J.; Pourbagheri-Sigaroodi, A.; Bashash, D. Host Genetic Diversity and Genetic Variations of SARS-CoV-2 in COVID-19 Pathogenesis and the Effectiveness of Vaccination. International Immunopharmacology 2022, 111, 109128. https://doi.org/10.1016/j.intimp.2022.109128. [CrossRef]
- Falahi, S.; Kenarkoohi, A. Host Factors and Vaccine Efficacy: Implications for COVID-19 Vaccines. Journal of Medical Virology 2022, 94, 1330–1335. https://doi.org/10.1002/jmv.27485. [CrossRef]
- Dudley, M.Z.; Gerber, J.E.; Budigan Ni, H.; Blunt, M.; Holroyd, T.A.; Carleton, B.C.; Poland, G.A.; Salmon, D.A. Vaccinomics: A Scoping Review. Vaccine 2023, 41, 2357–2367. https://doi.org/10.1016/j.vaccine.2023.02.009. [CrossRef]
- Filippatos, F.; Tatsi, E.-B.; Efthymiou, V.; Syriopoulou, V.; Michos, A. Time of Day of BNT162b2 COVID-19 Immunization Affects Total SARS-CoV-2 Antibody Levels but Not Neutralizing Activity. J Biol Rhythms 2022, 37, 562–566. https://doi.org/10.1177/07487304221100951. [CrossRef]
- Zhang, H.; Liu, Y.; Liu, D.; Zeng, Q.; Li, L.; Zhou, Q.; Li, M.; Mei, J.; Yang, N.; Mo, S.; et al. Time of Day Influences Immune Response to an Inactivated Vaccine against SARS-CoV-2. Cell Res 2021, 31, 1215–1217. https://doi.org/10.1038/s41422-021-00541-6. [CrossRef]
- Lammers-van der Holst, H.M.; Lammers, G.J.; van der Horst, G.T.J.; Chaves, I.; de Vries, R.D.; GeurtsvanKessel, C.H.; Koch, B.; van der Kuy, H.M. Understanding the Association between Sleep, Shift Work and COVID-19 Vaccine Immune Response Efficacy: Protocol of the S-CORE Study. Journal of Sleep Research 2022, 31, e13496. https://doi.org/10.1111/jsr.13496. [CrossRef]
- Schmitz, N.C.M.; van der Werf, Y.D.; Lammers-van der Holst, H.M. The Importance of Sleep and Circadian Rhythms for Vaccination Success and Susceptibility to Viral Infections. Clocks & Sleep 2022, 4, 66–79. https://doi.org/10.3390/clockssleep4010008. [CrossRef]
- Mason, A.E.; Kasl, P.; Hartogensis, W.; Natale, J.L.; Dilchert, S.; Dasgupta, S.; Purawat, S.; Chowdhary, A.; Anglo, C.; Veasna, D.; et al. Metrics from Wearable Devices as Candidate Predictors of Antibody Response Following Vaccination against COVID-19: Data from the Second TemPredict Study. Vaccines 2022, 10, 264. https://doi.org/10.3390/vaccines10020264. [CrossRef]
- Wagenhäuser, I.; Reusch, J.; Gabel, A.; Mees, J.; Nyawale, H.; Frey, A.; Lâm, T.-T.; Schubert-Unkmeir, A.; Dölken, L.; Kurzai, O.; et al. The Relationship between Mental Health, Sleep Quality and the Immunogenicity of COVID-19 Vaccinations. Journal of Sleep Research n/a, e13929. https://doi.org/10.1111/jsr.13929. [CrossRef]
- Bongiovanni, M.; Liuzzi, G.; Schiavon, L.; Gianturco, L.; Giuliani, G. Evaluation of the Immune Response to COVID-19 Vaccine MRNA BNT162b2 and Correlation with Previous COVID-19 Infection. Journal of Clinical Virology 2021, 143, 104962. https://doi.org/10.1016/j.jcv.2021.104962. [CrossRef]
- Sidler, D.; Born, A.; Schietzel, S.; Horn, M.P.; Aeberli, D.; Amsler, J.; Möller, B.; Njue, L.M.; Medri, C.; Angelillo-Scherrer, A.; et al. Trajectories of Humoral and Cellular Immunity and Responses to a Third Dose of MRNA Vaccines against SARS-CoV-2 in Patients with a History of Anti-CD20 Therapy. RMD Open 2022, 8, e002166. https://doi.org/10.1136/rmdopen-2021-002166. [CrossRef]
- Rockman, M.V.; Kruglyak, L. Genetics of Global Gene Expression. Nat Rev Genet 2006, 7, 862–872. https://doi.org/10.1038/nrg1964. [CrossRef]
- Zhang, W.; Duan, S.; Kistner, E.O.; Bleibel, W.K.; Huang, R.S.; Clark, T.A.; Chen, T.X.; Schweitzer, A.C.; Blume, J.E.; Cox, N.J.; et al. Evaluation of Genetic Variation Contributing to Differences in Gene Expression between Populations. The American Journal of Human Genetics 2008, 82, 631–640. https://doi.org/10.1016/j.ajhg.2007.12.015. [CrossRef]
- Svitkin, Y.V.; Cheng, Y.M.; Chakraborty, T.; Presnyak, V.; John, M.; Sonenberg, N. N1-Methyl-Pseudouridine in MRNA Enhances Translation through EIF2α-Dependent and Independent Mechanisms by Increasing Ribosome Density. Nucleic Acids Res 2017, 45, 6023–6036. https://doi.org/10.1093/nar/gkx135. [CrossRef]
- Lim, J.K.; Lisco, A.; McDermott, D.H.; Huynh, L.; Ward, J.M.; Johnson, B.; Johnson, H.; Pape, J.; Foster, G.A.; Krysztof, D.; et al. Genetic Variation in OAS1 Is a Risk Factor for Initial Infection with West Nile Virus in Man. PLOS Pathogens 2009, 5, e1000321. https://doi.org/10.1371/journal.ppat.1000321. [CrossRef]
- Liu, L.; Chen, X.; Skogerbø, G.; Zhang, P.; Chen, R.; He, S.; Huang, D.-W. The Human Microbiome: A Hot Spot of Microbial Horizontal Gene Transfer. Genomics 2012, 100, 265–270. https://doi.org/10.1016/j.ygeno.2012.07.012. [CrossRef]
- van Reenen, C.A.; Dicks, L.M.T. Horizontal Gene Transfer amongst Probiotic Lactic Acid Bacteria and Other Intestinal Microbiota: What Are the Possibilities? A Review. Arch Microbiol 2011, 193, 157–168. https://doi.org/10.1007/s00203-010-0668-3. [CrossRef]
- Groussin, M.; Poyet, M.; Sistiaga, A.; Kearney, S.M.; Moniz, K.; Noel, M.; Hooker, J.; Gibbons, S.M.; Segurel, L.; Froment, A.; et al. Elevated Rates of Horizontal Gene Transfer in the Industrialized Human Microbiome. Cell 2021, 184, 2053-2067.e18. https://doi.org/10.1016/j.cell.2021.02.052. [CrossRef]
- Sfera, A.; Osorio, C.; Hazan, S.; Kozlakidis, Z.; Maldonado, J.C.; Zapata-Martín del Campo, C.M.; Anton, J.J.; Rahman, L.; Andronescu, C.V.; Nicolson, G.L. Long COVID and the Neuroendocrinology of Microbial Translocation Outside the GI Tract: Some Treatment Strategies. Endocrines 2022, 3, 703–725. https://doi.org/10.3390/endocrines3040058. [CrossRef]
- Hazan, S.; Stollman, N.; Bozkurt, H.S.; Dave, S.; Papoutsis, A.J.; Daniels, J.; Barrows, B.D.; Quigley, E.M.; Borody, T.J. Lost Microbes of COVID-19: Bifidobacterium, Faecalibacterium Depletion and Decreased Microbiome Diversity Associated with SARS-CoV-2 Infection Severity. BMJ Open Gastroenterol 2022, 9, e000871. https://doi.org/10.1136/bmjgast-2022-000871. [CrossRef]
- Liu, Q.; Mak, J.W.Y.; Su, Q.; Yeoh, Y.K.; Lui, G.C.-Y.; Ng, S.S.S.; Zhang, F.; Li, A.Y.L.; Lu, W.; Hui, D.S.-C.; et al. Gut Microbiota Dynamics in a Prospective Cohort of Patients with Post-Acute COVID-19 Syndrome. Gut 2022, 71, 544–552. https://doi.org/10.1136/gutjnl-2021-325989. [CrossRef]
- Fairgrieve, D.; Rizzi, M.; Kirchhelle, C.; Halabi, S.; Howells, G.; Witzleb, N. No-Fault Compensation Schemes for COVID-19 Vaccines: Best Practice Hallmarks. Public Health Rev 2023, 44, 1605973. https://doi.org/10.3389/phrs.2023.1605973. [CrossRef]
- Fairgrieve, D.; Borghetti, J.-S.; Dahan, S.; Goldberg, R.; Halabi, S.; Holm, S.; Howells, G.; Kirchelle, C.; Pillay, A.; Rajneri, E. Comparing No-Fault Compensation Systems for Vaccine Injury. Tul. J. Int’l & Comp. L. 2023, 31, 75.
- Crum, T.; Mooney, K.; Tiwari, B.R. Current Situation of Vaccine Injury Compensation Program and a Future Perspective in Light of COVID-19 and Emerging Viral Diseases. F1000Res 2021, 10, 652. https://doi.org/10.12688/f1000research.51160.2. [CrossRef]
- Demasi, M. Covid-19: Is the US Compensation Scheme for Vaccine Injuries Fit for Purpose? BMJ 2022, 377, o919. https://doi.org/10.1136/bmj.o919. [CrossRef]
- Long Covid Is a ‘National Crisis.’ So Why Are Grants Taking so Long to Get? Available online: https://www.science.org/content/article/long-covid-national-crisis-so-why-are-grants-taking-so-long-get (accessed on 11 July 2023).
- Halma, M.T.J.; Plothe, C.; Marik, P.; Lawrie, T.A. Strategies for the Management of Spike Protein-Related Pathology. Microorganisms 2023, 11, 1308. https://doi.org/10.3390/microorganisms11051308. [CrossRef]
- Post-Vac Syndrome — the Forgotten COVID Victims – DW – 03/21/2023 Available online: https://www.dw.com/en/post-vac-syndrome-the-forgotten-covid-victims/a-65051748 (accessed on 11 July 2023).
Figure 1.
A model for long-term spike protein persistence in vaccine injured individuals.
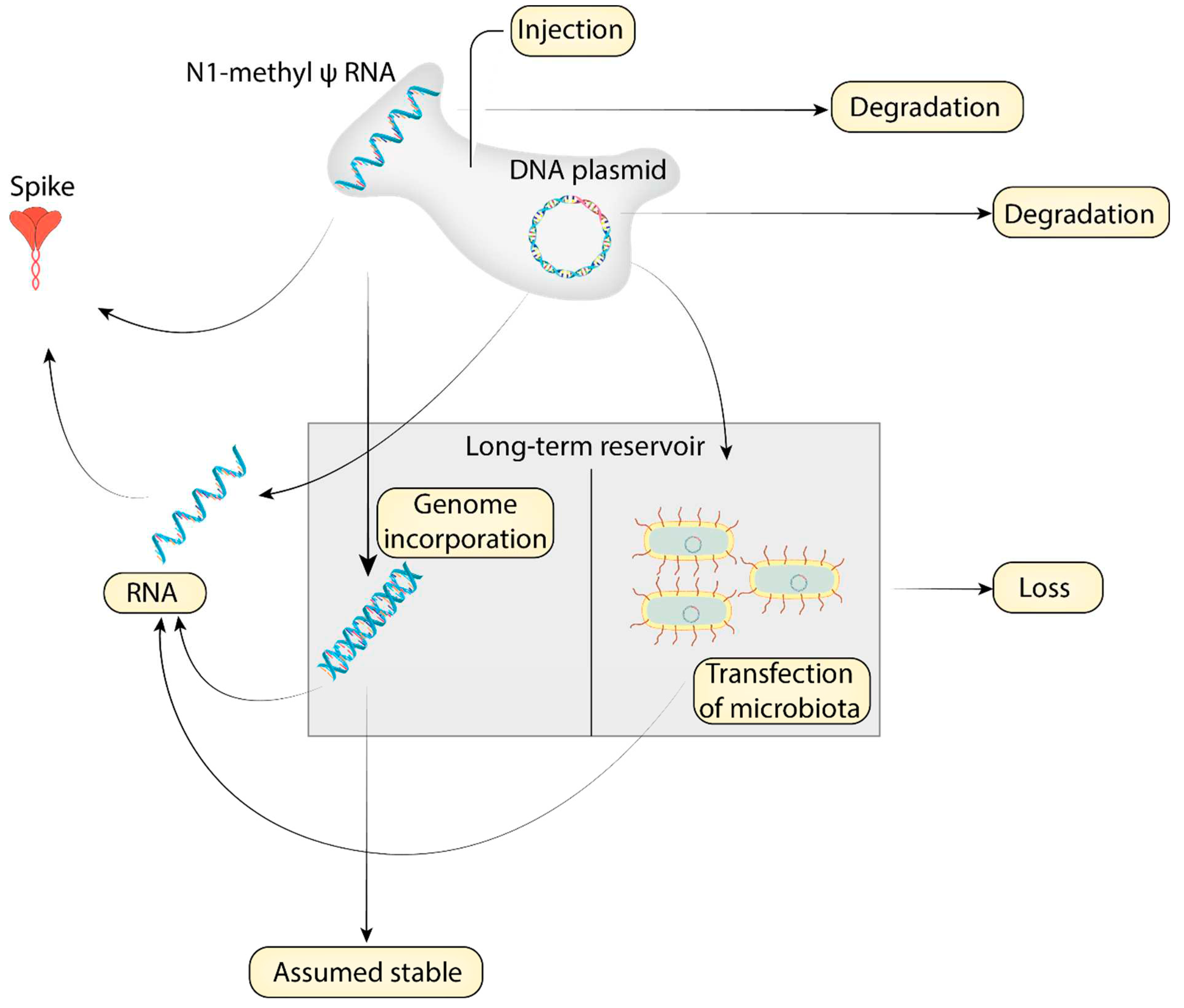
Table 1.
Basis of diagnostic difference between vaccine damage and damage from SARS-CoV-2.
| Vaccine Spike | Viral Spike |
| No N protein present | N protein present |
| Sequence identical to vaccine sequence | Sequence much less constrained, reflects currently circulating variants |
| Locked into prefusion conformation | Conformationally flexible |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



