
Submitted:
12 July 2023
Posted:
13 July 2023
You are already at the latest version
Abstract
Quorum sensing (QS), a well-established phenomenon in microorganisms, involves the communication between cells through chemical signals, which is dependent on cell density. Extensive research has been conducted on this microbial ability, encompassing the early stages of understanding QS to the latest advancements in the identification and characterization of its mechanisms. This minireview comprehensively examines the role of QS in various aspects, including biofilm formation, virulence in pathogenic bacteria such as Helicobacter pylori and Pseudomonas sp., as well as its influence on biogeochemical cycling in deep-sea environments. Furthermore, future progress in the field will be achieved by combining state-of-the-art methods for observing QS in the deep sea with a deeper understanding of the underlying processes, which will facilitate the engineering of microorganisms for improved degradation of persistent environmental pollutants and other biotechnological applications.
Keywords:
quorum sensing
; microbial communication
; biofilm formation
; biogeochemical cycling
; biotechnological applications
; marine environment
; deep‐sea environment
1. Introduction
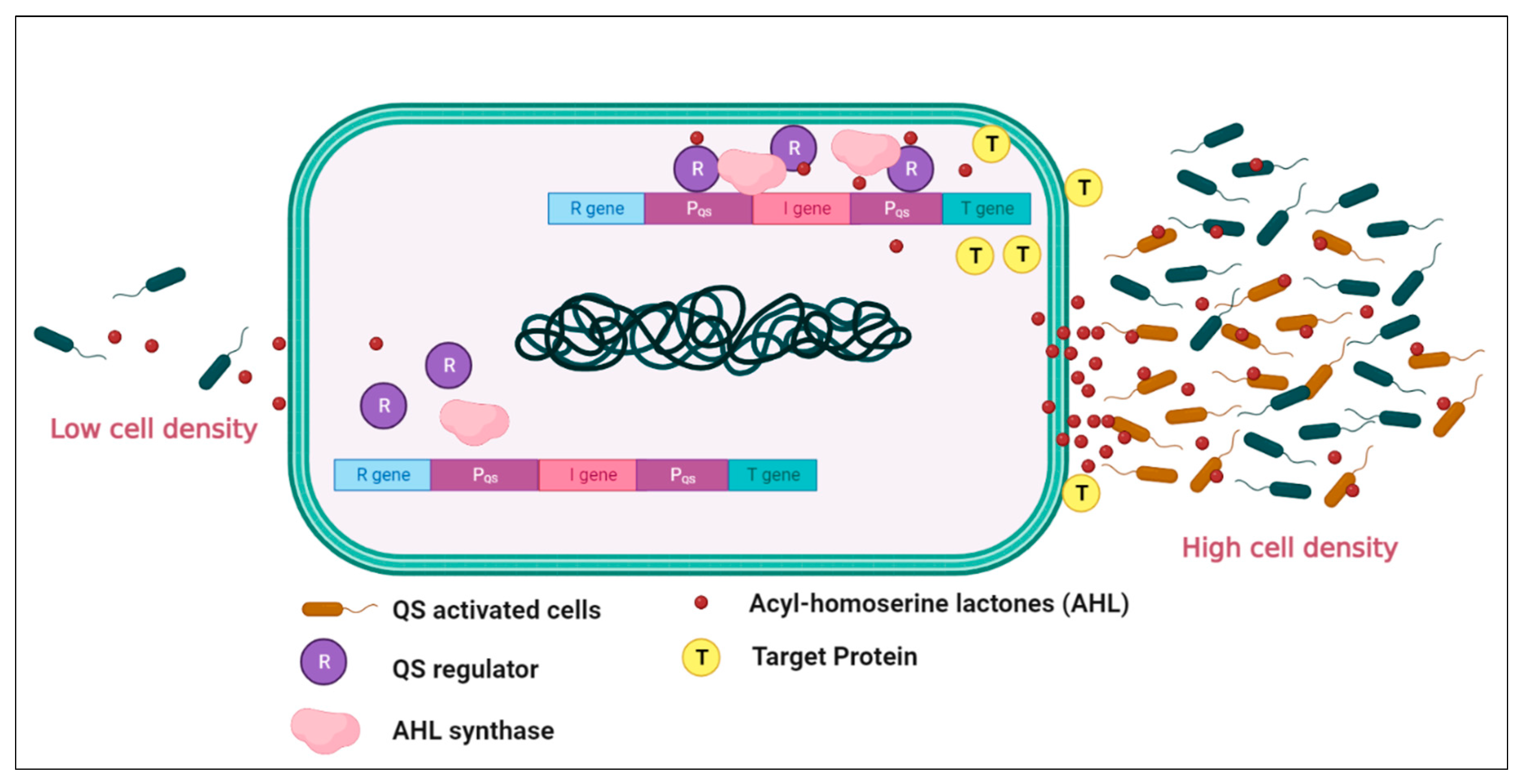
Extensive research has significantly advanced the understanding of bacteria, particularly concerning their lifecycle. From the earliest investigations into basic phenomena like food fermentation, research has progressed to uncovering the presence of disease-causing pathogens in diverse ecological niches, including plant rhizosphere soil, deep ocean zones, alpine peaks, and other extreme environments [1]. Among the notable discoveries is the phenomenon of cell-to-cell communication, known as "quorum sensing," which occurs within bacterial communities, both within and between species. The concept of quorum sensing was first observed over 25 years ago in two marine bacterial species: Vibrio fischeri and Vibrio harveyi; as shown in Figure 1 these bacteria were found to accumulate autoinducer signal molecules at high cell population densities, resulting in the activation of light emission by altering gene expression [2,3,4]. Since this initial observation, researchers have identified homologs of these genes in various bacterial communities, which play a regulatory role in quorum sensing. This communication process has been found to influence a wide range of biological processes, including bioluminescence [5], biofilm formation [6], cell competency [7], horizontal gene transfer [8], virulence factor expression [9], symbiotic relationships [10], sporulation in fungi [11,12];, pigment production [13], motility [14], toxin production [15], and even antibiotic production [16,17,18]. These findings underscore the intricate and diverse ways bacteria communicate and coordinate their behaviours within complex microbial communities. Exploring quorum sensing (QS) has contributed to understanding the mechanisms underlying bacterial interactions. It has opened avenues for further research into manipulating and controlling bacterial behaviour for various applications, including medical, environmental, and industrial contexts.
For bacteria to effectively utilize quorum sensing, they must possess certain capabilities, including the ability to secrete signaling molecules, synthesize autoinducers for detecting changes in signaling molecule concentration, and respond by regulating gene transcription [19,20]. The process heavily relies on the diffusion mechanism of these signaling molecules, which are typically secreted at low concentrations by individual bacteria. These molecules may simply diffuse away from the cell in environments with low cell density. However, as cell density increases, the local concentration of signaling molecules can surpass a threshold level, triggering a shift in gene expression [21].
Numerous quorum sensing systems have been extensively studied and documented in various taxa and habitats (a PubMed keyword search for “quorum sensing” generates over 12,000 results) [22]. Yet, many taxa and habitats remain, including deep-sea microbes, where quorum sensing remains unidentified or poorly characterized. Further research is needed to explore and understand the prevalence and specific mechanisms of quorum sensing in these diverse environments. This ongoing investigation will enhance our knowledge of the ecological significance and evolutionary implications of quorum sensing across a broader spectrum of microbial life.
Traditionally, QS was believed to be closely associated with the regulation of bacterial bioluminescence. However, a recent study by Tanet et al. [23] challenged this notion. The researchers investigated Photobacterium phosphoreum ANT-2200, a bioluminescent strain isolated from the deep-sea waters of the Mediterranean at a depth of 2200 meters. Surprisingly, this strain exhibited higher light emission rates at low cell densities than higher ones. Subsequently, the researchers cultivated the same bacteria under high hydrostatic pressure and found no change in QS gene transcription. These findings led them to conclude that the deep-sea strain of Photobacterium phosphoreum ANT-2200 is not density-dependent and, therefore, not susceptible to quorum sensing control. Another study by Muras et al. [24] investigated the prevalence of quorum sensing in deep-sea environments. The researchers found that the abundance of bacterial members involved in quorum sensing was relatively low in deep-sea samples collected at a depth of 2000 meters compared to surface samples taken at 15 meters. Interestingly, the study also revealed a high abundance of potential enzymes engaged in QS in the deep-sea samples. This observation suggests that QS activity may be an adaptive feature of strains that possess QS capacity in an environment where quorum sensing activities enhance the fitness of bacterial species, or it may indicate a condition in which the concentration of , N-acyl homoserine lactones (AHL) molecules (quorum sensing signal molecules) is high enough to serve as a substantial source of carbon. These studies emphasize the importance of both quorum sensing and quorum quenching activities in deep-sea environments. They highlight the significant role of these activities in deep marine systems. Consequently, this mini-review aims to discuss the phenomenon of quorum sensing and quenching in deep-sea environments, shedding light on its implications and importance in these unique ecosystems.
2. Bacterial Quorum Sensing in the Deep Sea
Quorum sensing (QS) has been discovered in various marine organisms, including the deep-water inhabitant, the bobtail squid (Euprymna scolopes), which utilizes Vibrio fischeri for its bioluminescence trait [25] . QS has also been identified in Epsilonproteobacteria, some of which are pathogens in humans. The thermal origin of this ability in mesophilic and pathogenic Epsilonproteobacteria has been traced back to ocean hydrothermal vents [26]. Among the different classes of signaling molecules, N-acyl homoserine lactones (AHLs), oligopeptides, and LuxS/autoinducer 2 (AI-2) have been extensively studied in marine environments [27] . AI-2 has been proposed as a universal signal for interspecies communication and is synthesized by the LuxS enzyme. It was initially identified in Vibrio harveyi, where it regulates bioluminescence in response to population density. Other microbes found in deep-sea vents, such as Epsilonbacteria (e.g., Sulfurovum lithotrophicum and Caminibacter mediatlanticus), also express the LuxS gene and produce QS signals [26,28].
AI-2 has a precursor molecule called 4,5-dihydroxy-2,3-pentanedione (DPD), which can exist in three forms: 4-hydroxy-5-methyl-3(2H) furanone (MHF), (2R, 4S)-2-methyl-2,3,3,4-tetrahydroxy-tetrahydrofuran (R-THMF), and furanosyl borate diester (S-THMF borate). Epsilonproteobacteria exhibit the highest LuxS gene expression during the mid-exponential growth phase and the lowest during the stationary phase. Conversely, bioluminescence, as detected using Vibrio harveyi strain BB170, is highest during the stationary phase when cell density is high, correlating with AI-2 activity [26,28].
To date, no AI-2 receptors have been identified in Epsilonproteobacteria. However, two main receptor categories, LuxP in Vibrio spp. and LrsB in enteric bacteria, are known. In deep-sea vents, Epsilonproteobacteria act as early-stage colonizers, preparing the stage for colonizing other microbes through QS signaling involving adhesive polymeric secretion. This secretion also serves as a nutrient source, promoting the recruitment of additional microbial species. The extracellular matrix of P. aeruginosa is composed of extracellular DNA (eDNA), which aids biofilm assemblage in a reaction with extracellular calcium by promoting bacterial association when released via QS-dependent and independent pathways. eDNA affects the RNA composition of members of the biofilm, acting as a nutrient source through the contents of the membrane vesicle in which it is enclosed, as well as modulating the expression of genes influencing acidity and antimicrobial resistance [29]. Horizontal gene transfer has been observed between gut-dwelling Helicobacter pylori and Enterococcus faecium, explaining the successful QS responses, such as virulence, in communities of species from different taxa [26,30]. Marine environments, particularly microniches characterized by high organic matter content (e.g., biofilm covers and biotic/abiotic surfaces), create favourable conditions for QS due to the resulting higher cell density in these habitats, leading to the accumulation of AI molecules.

Table 1.
Quorum Sensing Activities in Various Bacterial Species and Environments, includin oceanic settings.
Table 1.
Quorum Sensing Activities in Various Bacterial Species and Environments, includin oceanic settings.
| Bacteria | Activity mediated by QS | Location of occurrence | Signaling Molecules | QS System | Reference |
|---|---|---|---|---|---|
| Staphylococcus aureus | Virulence factor production control Biofilm formation control through the agr system |
Human body | Autoinducing peptides (AIPs) | Agr system | [31,32] |
| Bacillus cereus | Virulence factor control through the PlcR system | Various habitats including soil and food | Cyclic peptide | PlcR system | |
| Pseudomonas aeruginosa | Virulence factor control through LuxI/LuxR-type system | Various habitats including soil, water, and plants | N-acyl homoserine lactones (AHLs) | LasR/RhlR system | [33] |
| Vibrio fischerii | Light emission gene control through the LuxI system | Bobtail squid (Euprymna scolopes) in marine waters | N-acyl homoserine lactone (AHL) | LuxI/LuxR system | [25] |
| Vibrio diabolicus | Biofilm formation control | Polychaete annelid Alvinella pompejana deep-sea hydrothermal vent | Autoinducing peptides (AIPs) | Unknown QS system | [34,35] |
| Escherichia coli | Regulation of biofilm formation, motility, and virulence | Various habitats including the human gastrointestinal tract | Autoinducer-2 (AI-2) | LuxS/AI-2 system | [36,37] |
| Vibrio cholerae | Toxin production regulation and colonization of the human intestine | Hadel zones and human gastrointestinal tract | N-acyl homoserine lactones (AHLs) | LuxO/LuxR system | [38,39] |
| Streptococcus pneumoniae | Competence development and genetic transformation | Human respiratory tract | Peptides | ComDE system | [40] |
| Acinetobacter baumannii | Biofilm formation and antibiotic resistance | Various environments including hospitals and soil | Unknown | Unknown QS system | [41,42] |
| Aliivibrio fischeri | Symbiotic colonization of the Hawaiian bobtail squid | Bobtail squid (Euprymna scolopes) in marine waters | N-acyl homoserine lactone (AHL) | LuxI/LuxR system | [43] |
| Photobacterium phosphoreum | Bioluminescence control | Deep-sea waters | Autoinducer-2 | LuxS/AI-2 system | [23] |
| Sulfitobacter sp. | Production of extracellular enzymes and biofilm formation | Marine environments including surface water and sediments | Unknown | Unknown QS system | [44] |
| Ruegeria sp. | Biofilm formation and production of extracellular enzymes | Marine environments including coastal seawater and sediment | N-acyl homoserine lactone (AHL) | LuxI/LuxR system | [45] |
| Shewanella oneidensis | Regulation of biofilm formation and metal oxide reduction | Marine and freshwater environments | Unknown | Unknown QS system | [46,47] |
| Colwellia psychrerythraea | Cold adaptation and biofilm formation | Cold marine environments | Unknown | Unknown QS system | [48,49] |
| Psychrobacter sp. | Production of extracellular enzymes and biofilm formation | marine sediments | N-acylhomoserine lactones (AHL) | LuxI/LuxR system QS system | [50] |
| Marinobacter sp. | Biofilm formation and quorum quenching | Marine environments including deep-sea sediments | Unknown | Unknown QS system | [51] |
| Sulfurovum lithotrophicum | Quorum sensing in deep-sea vent bacteria | Deep-sea hydrothermal vents | Autoinducer-2 (AI-2) | LuxS/AI-2 system | [26,52] |
| Caminibacter mediatlanticus | Quorum sensing in deep-sea vent bacteria | Deep-sea hydrothermal vents | Autoinducer-2 (AI-2) | LuxS/AI-2 system | [26,52] |
| Thiomicrospira sp. | Symbiotic interactions and biofilm formation | Deep-sea hydrothermal vents | Unknown | Unknown QS system | [53] |
3. Biofilm Formation
Biofilms play a crucial role in the physiological adaptation of bacteria in deep-sea environments, enabling their survival within these complex matrices. The unique characteristics of biofilms provide member cells with distinct advantages, such as heightened resistance against host immune responses, enhanced nutrient availability and utilization, and increased tolerance to antimicrobial agents [54]. Several studies focusing on Vibrio fischeri, Vibrio anguillarum, and Vibrio harveyi have demonstrated the involvement of QS in regulating bioluminescence, virulence, and biofilm formation in these species. Notably, biofilms from extreme environments exhibit the remarkable ability to rapidly transition from early colonization stages to robust production of extracellular polymeric substances (EPS) that contribute to protective mechanisms [55].
In marine ecosystems, EPS production is a common trait observed among various microorganisms, including bacteria, cyanobacteria, and actinobacteria. These EPS molecules contribute to biofilms' development and structural integrity in marine environments, facilitating microbial survival and adaptation [55]). In addition to the physiological advantages biofilms confer in deep-sea environments, QS within these communities plays a vital role in coordinating microbial activities. It enables intercellular communication and facilitates the formation of complex multispecies biofilms commonly observed in marine ecosystems. Studies have revealed that QS regulates bioluminescence, virulence, and biofilm formation and other important processes in marine bacteria. For example, in the bacterium Pseudomonas aeruginosa, QS controls the production of various virulence factors, including exopolysaccharides, proteases, and toxins, contributing to its pathogenicity and colonization ability [56].
Furthermore, QS has been implicated in the regulation of motility and colonization in marine bacteria. In Vibrio cholerae, a pathogenic bacterium responsible for cholera, quorum sensing coordinates the expression of genes involved in flagellar motility and biofilm formation, enabling efficient colonization of aquatic environments and host surfaces [57]. Quorum sensing in marine environments is not limited to bacteria alone. Marine algae, like diatoms, also employ QS mechanisms to coordinate their growth and reproductive processes [58]. This interkingdom communication between bacteria and algae further highlights the ecological significance of QS in marine ecosystems. The deep-sea environment presents unique challenges for microbial communities, including high pressure, low temperature, and limited nutrient availability. Quorum sensing allows bacteria to adapt to these extreme conditions by facilitating cooperative behaviours and resource sharing within biofilms. Moreover, studies have shown that deep-sea bacteria exhibit specific adaptations in their QS systems, likely influenced by the selective pressures of these environments [59,60]. Quorum sensing plays a multifaceted role in marine environments, regulating various physiological processes in bacteria and other microorganisms. It facilitates communication and coordination within biofilms and contributes to the adaptation and survival of microbial communities in diverse marine ecosystems, including the challenging deep-sea environment.
Table 2.
Quorum Sensing and Biofilm Composition in Various Marine Environments.
| Location | Bacteria | Biofilm composition | Functions | Reference | Quorum Sensing Involvement |
|---|---|---|---|---|---|
| Antarctic Waters | Marine bacteria | Charged uronic acid moieties and sulfate groups | Plays a role in cold adaptation | [55] | QS system(s) involved |
| Korean yellow sea | Bacillus sp. I450 | Neutral sugars and uronic acids | Exhibits antimicrobial activity | [55,61] | QS system(s) involved |
| Solar Saltern and Spanish Mediterranean Seaboard | Halomonas maura & Salipiger mucosus | Sulfated polysaccharide with high uronic acid content and fucose-rich polysaccharides | Adapted to high salinity environments and high capacity for binding cations | [62,63] | QS system(s) involved |
| Deep-sea hydrothermal vent | Caminibacter mediatlanticus | Sulfated polysaccharide high in glucosamine | Thrives in high-temperature hydrothermal vents | [26,64] | QS system(s) involved |
| Deep-sea hydrothermal vent | Alteromonas infernus | Lacking lipopolysaccharides (LPS) | Exhibits unique adaptations to extreme conditions, including high-pressure and high-temperature environments | [65] | QS system(s) involved |
| Host in deep-sea hydrothermal vent | Vibrio diabolcus | Large amounts of uronic acid and no sulfate groups | Forms symbiotic relationship with host organism, the polychaete annelid Alvinella pompejana, in deep-sea hydrothermal vent ecosystems | [34] | QS system(s) involved |
| Antarctic Ocean | Psychrobacter sp. | Extracellular polysaccharides | Exhibits cold-adapted enzymatic activity and plays a role in biofilm formation under low temperatures | [66] | Not specified |
| Deep-sea sediments | Marinobacter sp. | Exopolysaccharides | Involved in sediment stabilization and biogeochemical cycling in deep-sea sediments | [67,68] | Not specified |
| Gulf of Mexico | Vibrio vulnificus | Alginate and extracellular DNA | Forms biofilms on oyster shells and contributes to oyster pathogenesis | [69] | QS system(s) involved |
| Deep-sea hydrothermal vent | Thiomicrospira sp. | Extracellular sulfur and polysaccharides | Capable of sulfur oxidation and plays a role in ecosystem functioning in deep-sea hydrothermal vents | [70] | Not specified |
| Mariana Trench | Pseudomonas sp. | Exopolysaccharides | Exhibits unique adaptations to extreme pressures and low nutrient availability in the Mariana Trench | [71,72] | QS system(s) involved |
4. Biogeochemical Cycling and Quorum Sensing
Quorum sensing mechanisms have been found to play a significant role in regulating microbial activities involved in biogeochemical cycling in the ocean. QS allows bacteria to communicate and coordinate their behaviours in response to population density and environmental cues. In marine environments, QS-mediated gene regulation influences the cycling of essential elements such as carbon, nitrogen, and sulfur. For example, QS controls the production of extracellular enzymes involved in the breakdown of complex organic matter, allowing bacteria to efficiently utilize these substrates for energy and nutrient acquisition [56,57]. The coordinated expression of genes involved in enzymatic activities enhances the degradation and cycling of organic compounds in the ocean. Biofilms found in deep-sea vents, characterized by a white sheen on rocks, have long been observed, but their functions were poorly understood. Chemoautotrophic microorganisms are the first to colonize solid substrates at the seafloor and form extensive biofilms that condition the environment for the subsequent colonization of other single-celled organisms. These microorganisms exhibit metabolic versatility and can utilize heavy metals, contributing to their detoxification. The formation of these biofilms is regulated by QS, as evidenced by the expression of QS genes in Caminibacter mediatlanticus and Sulfurovum sp. during biofilm growth and in situ [52]. Quorum sensing plays a crucial role in directing the formation of biofilms, facilitating the breakdown of chemicals in deep-sea vents and contributing to the continuous flow of products in the global biogeochemical cycle.
Quorum sensing also influences the nitrogen cycle by regulating nitrogen fixation, nitrification, and denitrification processes. Some marine bacteria, such as the genus Vibrio, utilize QS to control the expression of nitrogen fixation genes, enabling them to convert atmospheric nitrogen into biologically available forms [43]. Quorum sensing also regulates the expression of genes involved in nitrification and denitrification, affecting the balance of nitrogen species in marine ecosystems. Sulfur cycling in the ocean is another biogeochemical process influenced by QS. QS-controlled gene expression in sulfur-oxidizing bacteria regulates the production of enzymes involved in oxidising reduced sulfur compounds, such as hydrogen sulfide, into sulfate [52,73]. This process is crucial for sulfur recycling in marine environments and contributes to the global sulfur cycle. Furthermore, QS-mediated biofilm formation has implications for biogeochemical cycling. Biofilms provide a structured and protected environment for microbial communities, allowing them to efficiently carry out processes such as nutrient uptake, metabolic cooperation, and extracellular matrix production ( [74]. QS coordinates the formation and maturation of biofilms, enabling microbial consortia to establish complex interactions and perform specialized functions that contribute to biogeochemical cycling.
5. Methods of Monitoring Quorum Sensing
Monitoring QS in the marine environment requires using specific techniques that can capture the dynamic nature of QS interactions. Time-series methods are commonly employed to observe QS dynamics in deep-sea environments. These methods involve collecting samples at multiple time points and analyzing them using various techniques.
Omics approaches, such as metagenomics, metatranscriptomics, and metaproteomics, provide a comprehensive view of the microbial community and its gene expression patterns. These techniques enable the identification of QS-related genes, signaling molecules, and regulatory networks in the marine environment [75]. By analyzing the genetic information encoded in the collected samples, researchers can gain insights into the presence and activity of QS systems. Fluorescence in situ hybridization (FISH) is a microscopy-based technique that allows the visualization and identification of specific microbial populations in their natural habitat. Using fluorescently labeled probes that target QS-related genes or specific microbial taxa, researchers can observe the spatial distribution and abundance of QS-active cells in marine samples [75]. This technique provides valuable information on the localization and dynamics of QS-mediated interactions.
Raman microscopy is another powerful tool used in QS studies. It enables the non-destructive analysis of individual cells and their chemical composition. By detecting molecular vibrations, Raman microscopy can provide insights into cellular metabolism and QS-related molecule production in the marine environment [75,76]. This technique can be combined with other methods to obtain a more comprehensive understanding of QS processes. To study the interactions between microorganisms in the deep sea, researchers often analyze taxonomic data from different habitats or examine time-series samples from the same habitat. These approaches allow for identifying shifts in microbial community composition and QS-related gene expression patterns over time, providing insights into the dynamics of QS-mediated interactions [77]. Single-cell genomics is a powerful approach that involves analyzing the genetic material of individual cells in their natural environment. By targeting genes associated with QS systems, researchers can gain specific insights into the QS activity of individual cells within a microbial community [52,78]. This technique can provide valuable information on the diversity and functionality of QS systems in marine environments. While traditional molecular sequence-based methods, such as 16S and 18S rRNA gene sequencing, have been widely used for microbial ecology studies, they have limitations in capturing microbial communities' metabolic pathways and functional capabilities. PCR biases, mismatches, and the inability to decode metabolic pathways have limited the applicability of these methods in QS research [75,79] ). By combining molecular sequence-based -omics methods with isotope tracing techniques like FISH, researchers can better understand the metabolite flow within microbial communities. Isotope tracing allows for tracking specific metabolites and their utilization by QS-active cells, providing insights into the metabolic activities and interactions within marine microbial communities [75,80].
6. Biotechnological Applications
The biotechnological advancements in QS have led to the discovery and utilization of extracellular polymeric substances (EPS) produced by marine bacteria for various applications in different industries. One notable example is the EPS produced by Vibrio diabolicus and Alteromonas infernus, isolated from deep-sea hydrothermal vents, showing medicinal regenerative properties. These EPS contain glycosaminoglycans are beneficial for tissue repair and remodeling due to their low immunogenicity. One of such product derived from these EPS is Hyalurift®, which utilizes hyaluronic acid to promote tissue remodelling and aid in joint injuries and embryogenesis. The EPS consists of high molecular weight linear tetrasaccharidic repeating units of glucuronic acid and hexosamine (N-acetyl-glucosamine and N-acetyl-galactosamine), resembling a fusion of hyaluronan and non-sulfated chondroitin units [81]. Alteromonas macleodii subsp. fijiensis, strain HYD657, is another species that produces EPS with protective properties for sensitive skin against chemicals and UV radiation. This EPS, marketed as Abyssine, offers potential applications in skin care products for its protective and rejuvenating effects [55].
In the food industry, EPS produced by Labrenzia sp. have gained attention due to their low viscosity and antioxidant properties. These EPS can be utilized in food and pharmaceutical products, providing viscosity control and antioxidant benefits (Di Donato et al., 2016). Deep-sea bacteria have also shown the production of extremozymes, which are enzymes capable of functioning under extreme conditions. These extremozymes have significant applications in bioremediation, particularly in degrading hazardous compounds like crude oil, polycyclic aromatic hydrocarbons (PAHs), and heavy metals in the environment. They offer potential solutions for environmental clean-up and mitigation of pollution [82]. Furthermore, EPS produced by Pseudomonas aeruginosa JP-11 have been investigated for their potential in removing toxic cadmium ions from polluted waters. This suggests the possibility of using EPS-based strategies for the remediation of heavy metal contamination [55].
Some studies applied QS in antibiotic development, where quorum sensing inhibitors (QSIs) have been explored as potential antibiotics to disrupt bacterial communication and combat biofilm formation [83]. Bioremediation processes have also benefited from quorum sensing, as it has been utilized to enhance the degradation of pollutants like crude oil and heavy metals ( [84,85]. In biofilm control, strategies targeting QS have been developed to prevent and disrupt biofilm formation in healthcare, water systems, and food processing industries [86,87,88]). Moreover, manipulating QS has improved the production of biofuels, pharmaceuticals, and enzymes in industrial fermentation processes. In agriculture, quorum sensing has shown promise for developing biocontrol agents that can target plant pathogens and reduce the reliance on chemical pesticides [14,89]. Finally, synthetic quorum sensing systems have been engineered, enabling the programming of microbial cells for desired behaviors, opening up possibilities in biomedicine, biofuel production, and bioremediation. These advancements highlight the broad potential of quorum sensing in various biotechnological applications [14,83,90,91,92].
7. Conclusion
Studying of QS in extreme environments, including deep-sea, poses unique challenges in fully comprehending its intricate systems. However, unravelling the functions, molecular components, and underlying mechanisms of QS is imperative. Deep-sea environments have served as habitats for ancestral microorganisms, making it crucial to investigate quorum sensing to gain a comprehensive understanding of its roles in microbial communities, as many of these microbes have contributed to the cultivation of various organisms observed today. In-depth studies have shed light on strategies to mitigate virulence by disrupting quorum sensing through QS techniques and enhancing therapeutic applications by inducing autoinducer molecules. It is essential to incorporate ecological considerations in microbial QS research, recognizing the influence of evolution within diverse bacterial communities across different environments. Moreover, the future exploration of synthetic communities holds promise for more effective biotechnological applications than current genetic engineering approaches. By delving deeper into these scientific realms, we can unlock the full potential of quorum sensing and its practical implications.
Author Contributions
Conceptualization, E.I. and S.R.; methodology, E.I.; software, S.R.; validation, A.L.K.A. and C.B.C..; formal analysis, A.L.K.A. and C.B.C; data curation, X.X.; writing—original draft preparation, E.I. and S.R.; writing—review and editing, S.R., A.L.K.A. and C.B.C.; visualization, S.R., A.L.K.A. and C.B.C..; supervision, C.B.C. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not Applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sood, U.; Dhingra, G.G.; Anand, S.; Hira, P.; Kumar, R.; Kaur, J.; Verma, M.; Singhvi, N.; Lal, S.; Rawat, C.D.; et al. Microbial Journey: Mount Everest to Mars. Indian J. Microbiol. 2022, 62, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Nealson, K.H.; Platt, T.; Hastings, J.W. Cellular control of the synthesis and activity of the bacterial luminescent system. J. Bacteriol. 1970, 104, 313–322. [Google Scholar] [PubMed]
- Nealson, K.H. Autoinduction of bacterial luciferase: occurrence, mechanism and significance. Arch. Microbiol. 1977, 112, 73–79. [Google Scholar] [CrossRef]
- Engebrecht, J.; Nealson, K.; Silverman, M. Bacterial bioluminescence: isolation and genetic analysis of functions from Vibrio fischeri. Cell 1983, 32, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Bai, L.; Li, S.; Yan, W. Similarities and Differences in Quorum Sensing-Controlled Bioluminescence between Photobacterium phosphoreum T3 and Vibrio qinghaiensis sp.-Q67. Appl. Sci. 2022, 12. [Google Scholar] [CrossRef]
- Preda, V.G.; Săndulescu, O. Communication is the key: biofilms, quorum sensing, formation and prevention. Discoveries 2019, 7, e10. [Google Scholar] [CrossRef]
- Shanker, E.; Federle, M.J. Quorum sensing regulation of competence and bacteriocins in Streptococcus pneumoniae and mutans. Genes (Basel). 2017, 8. [Google Scholar]
- van Gestel, J.; Bareia, T.; Tenennbaum, B.; Dal Co, A.; Guler, P.; Aframian, N.; Puyesky, S.; Grinberg, I.; D’Souza, G.G.; Erez, Z.; et al. Short-range quorum sensing controls horizontal gene transfer at micron scale in bacterial communities. Nat. Commun. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- İnat, G.; Sırıken, B.; Başkan, C.; Erol, İ.; Yıldırım, T.; Çiftci, A. Quorum sensing systems and related virulence factors in Pseudomonas aeruginosa isolated from chicken meat and ground beef. Sci. Rep. 2021, 11, 1–9. [Google Scholar]
- Kai, K. Bacterial quorum sensing in symbiotic and pathogenic relationships with hosts. Biosci. Biotechnol. Biochem. 2018, 82, 363–371. [Google Scholar] [CrossRef]
- Albuquerque, P.; Casadevall, A. Quorum sensing in fungi – a review. Med. Mycol. 2012, 50, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Hull, C.M.; Genet Author manuscript, C. Sporulation: How to survive on planet Earth (and beyond) HHS Public Access Author manuscript. Curr. Genet. 2017, 63, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Joshi, C.; Kothari, V.; Patel, P. Importance of selecting appropriate wavelength, while quantifying growth and production of quorum sensing regulated pigments in bacteria. Recent Pat. Biotechnol. 2016, 10, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Bramhachari, P.V.; Yugandhar, N.M.; Prathyusha, A.; Mohana Sheela, G.; Naravula, J.; Venkateswarlu, N. Quorum sensing regulated swarming motility and migratory behavior in bacteria. In Implication of quorum sensing system in biofilm formation and virulence; Springer, 2018; pp. 49–66. [Google Scholar]
- Butrico, C.E.; Cassat, J.E. Quorum sensing and toxin production in Staphylococcus aureus osteomyelitis: Pathogenesis and paradox. Toxins (Basel). 2020, 12, 516. [Google Scholar] [CrossRef]
- Armes, A.C.; Walton, J.L.; Buchan, A. Quorum Sensing and Antimicrobial Production Orchestrate Biofilm Dynamics in Multispecies Bacterial Communities. Microbiol. Spectr. 2022, 10, e02615-22. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Bimerew, M.; Ma, Y.; Müller, H.; Ovadis, M.; Eberl, L.; Berg, G.; Chernin, L. Quorum-sensing signaling is required for production of the antibiotic pyrrolnitrin in a rhizospheric biocontrol strain of Serratia plymuthica. FEMS Microbiol. Lett. 2007, 270, 299–305. [Google Scholar] [CrossRef]
- Sanchez, L.M.; Mannathan, S.; Lee, C.-K.; Merlo, M.E.; Meng, X.-F.; Nihira, T.; Takano, E.; Minnaard, A.J.; Dijkhuizen, L.; Petrusma, M. Identification and characterisation of enzymes involved in γ-butyrolactone biosynthesis in Streptomyces coelicolor. In Quorum sensing in Streptomyces coelicolor; 2016; pp. 92–137. [Google Scholar]
- Pan, J.; Ren, D. Quorum sensing inhibitors: a patent overview. Expert Opin. Ther. Pat. 2009, 19, 1581–1601. [Google Scholar] [CrossRef]
- Carradori, S.; Di Giacomo, N.; Lobefalo, M.; Luisi, G.; Campestre, C.; Sisto, F. Biofilm and quorum sensing inhibitors: The road so far. Expert Opin. Ther. Pat. 2020, 30, 917–930. [Google Scholar] [CrossRef]
- Bassler, B.L. How bacteria talk to each other: regulation of gene expression by quorum sensing. Curr. Opin. Microbiol. 1999, 2, 582–587. [Google Scholar] [CrossRef]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and promise of bacterial quorum sensing research. Nature 2017, 551, 313–320. [Google Scholar] [PubMed]
- Tanet, L.; Tamburini, C.; Baumas, C.; Garel, M.; Simon, G.; Casalot, L. Bacterial bioluminescence: Light Emission in Photobacterium phosphoreum Is Not under Quorum-Sensing Control. Front. Microbiol. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Muras, A.; López-Pérez, M.; Mayer, C.; Parga, A.; Amaro-Blanco, J.; Otero, A. High prevalence of quorum-sensing and quorum-quenching activity among cultivable bacteria and metagenomic sequences in the mediterranean sea. Genes (Basel). 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Turovskiy, Y.; Kashtanov, D.; Paskhover, B.; Chikindas, M.L. Quorum sensing: fact, fiction, and everything in between. Adv. Appl. Microbiol. 2007, 62, 191–234. [Google Scholar] [PubMed]
- Pérez-Rodríguez, I.; Bolognini, M.; Ricci, J.; Bini, E.; Vetriani, C. From deep-sea volcanoes to human pathogens: A conserved quorum-sensing signal in Epsilonproteobacteria. ISME J. 2015, 9, 1222–1234. [Google Scholar] [CrossRef]
- Keller, L.; Surette, M.G. Communication in bacteria: an ecological and evolutionary perspective. Nat. Rev. Microbiol. 2006, 4, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Schauder, S.; Potier, N.; Van Dorsselaer, A.; Pelczer, I.; Bassler, B.L.; Hughson, F.M. Structural identification of a bacterial quorum-sensing signal containing boron. Nature 2002, 415, 545–549. [Google Scholar] [CrossRef]
- del Mar Cendra, M.; Torrents, E. Pseudomonas aeruginosa biofilms and their partners in crime. Biotechnol. Adv. 2021, 49, 107734. [Google Scholar] [CrossRef]
- Alain, K.; Zbinden, M.; Le Bris, N.; Lesongeur, F.; Quérellou, J.; Gaill, F.; Cambon-Bonavita, M.A. Early steps in microbial colonization processes at deep-sea hydrothermal vents. Environ. Microbiol. 2004, 6, 227–241. [Google Scholar] [CrossRef]
- Derakhshan, S.; Navidinia, M.; Haghi, F. Antibiotic susceptibility of human-associated Staphylococcus aureus and its relation to agr typing, virulence genes, and biofilm formation. BMC Infect. Dis. 2021, 21, 1–10. [Google Scholar] [CrossRef]
- Kong, K.-F.; Vuong, C.; Otto, M. Staphylococcus quorum sensing in biofilm formation and infection. Int. J. Med. Microbiol. 2006, 296, 133–139. [Google Scholar] [CrossRef]
- Wu, W.; Jin, Y.; Bai, F.; Jin, S. Pseudomonas aeruginosa. In Molecular medical microbiology; Elsevier, 2015; pp. 753–767. [Google Scholar]
- Raguénès, G.; Christen, R.; Guezennec, J.; Pignet, P.; Barbier, G. Vibrio diabolicus sp. nov., a new polysaccharide-secreting organism isolated from a deep-sea hydrothermal vent polychaete annelid, Alvinella pompejana. Int. J. Syst. Evol. Microbiol. 1997, 47, 989–995. [Google Scholar]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef]
- Reading, N.C.; Rasko, D.A.; Torres, A.G.; Sperandio, V. The two-component system QseEF and the membrane protein QseG link adrenergic and stress sensing to bacterial pathogenesis. Proc. Natl. Acad. Sci. 2009, 106, 5889–5894. [Google Scholar] [CrossRef]
- Bassler, B.L.; Losick, R. Bacterially Speaking. Cell 2006, 125, 237–246. [Google Scholar] [CrossRef]
- Zhu, J.; Miller, M.B.; Vance, R.E.; Dziejman, M.; Bassler, B.L.; Mekalanos, J.J. Quorum-sensing regulators control virulence gene expression in Vibrio cholerae. Proc. Natl. Acad. Sci. U. S. A. 2002, 99, 3129–3134. [Google Scholar] [CrossRef]
- Liang, J.; Liu, J.; Wang, X.; Sun, H.; Zhang, Y.; Ju, F.; Thompson, F.; Zhang, X.H. Genomic Analysis Reveals Adaptation of Vibrio campbellii to the Hadal Ocean. Appl. Environ. Microbiol. 2022, 88. [Google Scholar] [CrossRef]
- Peterson, S.N.; Sung, C.K.; Cline, R.; Desai, B. V; Snesrud, E.C.; Luo, P.; Walling, J.; Li, H.; Mintz, M.; Tsegaye, G. Identification of competence pheromone responsive genes in Streptococcus pneumoniae by use of DNA microarrays. Mol. Microbiol. 2004, 51, 1051–1070. [Google Scholar] [CrossRef]
- Niu, C.; Clemmer, K.M.; Bonomo, R.A.; Rather, P.N. Isolation and characterization of an autoinducer synthase from Acinetobacter baumannii. J. Bacteriol. 2008, 190, 3386–3392. [Google Scholar] [CrossRef]
- Antunes, L.C.M.; Ferreira, R.B.R.; Buckner, M.M.C.; Finlay, B.B. Quorum sensing in bacterial virulence. Microbiology 2010, 156, 2271–2282. [Google Scholar] [CrossRef]
- Visick, K.L.; Ruby, E.G. Vibrio fischeri and its host: it takes two to tango. Curr. Opin. Microbiol. 2006, 9, 632–638. [Google Scholar] [CrossRef]
- Caruso, G. Microbial colonization in marine environments: Overview of current knowledge and emerging research topics. J. Mar. Sci. Eng. 2020, 8, 1–22. [Google Scholar] [CrossRef]
- Su, Y.; Tang, K.; Liu, J.; Wang, Y.; Zheng, Y.; Zhang, X.-H. Quorum sensing system of Ruegeria mobilis Rm01 controls lipase and biofilm formation. Front. Microbiol. 2019, 9, 3304. [Google Scholar] [CrossRef]
- El-Naggar, M.Y.; Wanger, G.; Leung, K.M.; Yuzvinsky, T.D.; Southam, G.; Yang, J.; Lau, W.M.; Nealson, K.H.; Gorby, Y.A. Electrical transport along bacterial nanowires from Shewanella oneidensis MR-1. Proc. Natl. Acad. Sci. 2010, 107, 18127–18131. [Google Scholar] [CrossRef]
- Gorby, Y.A.; Yanina, S.; McLean, J.S.; Rosso, K.M.; Moyles, D.; Dohnalkova, A.; Beveridge, T.J.; Chang, I.S.; Kim, B.H.; Kim, K.S. Electrically conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and other microorganisms. Proc. Natl. Acad. Sci. 2006, 103, 11358–11363. [Google Scholar] [CrossRef]
- Marx, J.G.; Carpenter, S.D.; Deming, J.W. Production of cryoprotectant extracellular polysaccharide substances (EPS) by the marine psychrophilic bacterium Colwellia psychrerythraea strain 34H under extreme conditions. Can. J. Microbiol. 2009, 55, 63–72. [Google Scholar] [CrossRef]
- Casillo, A.; D’Angelo, C.; Parrilli, E.; Tutino, M.L.; Corsaro, M.M. Membrane and Extracellular Matrix Glycopolymers of Colwellia psychrerythraea 34H: Structural Changes at Different Growth Temperatures. Front. Microbiol. 2022, 13, 820714. [Google Scholar] [CrossRef] [PubMed]
- Packiavathy, I.A.S.V.; Kannappan, A.; Thiyagarajan, S.; Srinivasan, R.; Jeyapragash, D.; Paul, J.B.J.; Velmurugan, P.; Ravi, A.V. AHL-Lactonase producing Psychrobacter sp. from Palk Bay sediment mitigates quorum sensing-mediated virulence production in Gram negative bacterial pathogens. Front. Microbiol. 2021, 12, 634593. [Google Scholar] [CrossRef] [PubMed]
- Rughöft, S.; Jehmlich, N.; Gutierrez, T.; Kleindienst, S. Comparative proteomics of Marinobacter sp. Tt1 reveals corexit impacts on hydrocarbon metabolism, chemotactic motility, and biofilm formation. Microorganisms 2020, 9, 3. [Google Scholar] [CrossRef]
- Sievert, S.M.; Vetriani, C. Chemoautotrophy at deep-sea vents, past, present, and future. Oceanography 2012, 25, 218–233. [Google Scholar] [CrossRef]
- Nunoura, T.; Takaki, Y.; Kazama, H.; Kakuta, J.; Shimamura, S.; Makita, H.; Hirai, M.; Miyazaki, M.; Takai, K. Physiological and genomic features of a novel sulfur-oxidizing gammaproteobacterium belonging to a previously uncultivated symbiotic lineage isolated from a hydrothermal vent. PLoS One 2014, 9, e104959. [Google Scholar] [CrossRef] [PubMed]
- Rumbaugh, K.P.; Sauer, K. Biofilm dispersion. Nat. Rev. Microbiol. 2020, 18, 571–586. [Google Scholar] [CrossRef]
- Di Donato, P.; Poli, A.; Taurisano, V.; Abbamondi, G.R.; Nicolaus, B.; Tommonaro, G. Recent advances in the study of marine microbial biofilm: From the involvement of quorum sensing in its production up to biotechnological application of the polysaccharide fractions. J. Mar. Sci. Eng. 2016, 4. [Google Scholar] [CrossRef]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Mekalanos, J.J. Quorum Sensing-Dependent Biofilms Enhance Colonization in Vibrio cholerae), the phenomenon by which bacteria monitor their cell population density through the extracellular accumulation of signaling molecules called autoinduc. Dev. Cell 2003, 5, 647–656. [Google Scholar] [CrossRef]
- Seyedsayamdost, M.R.; Carr, G.; Kolter, R.; Clardy, J. Roseobacticides: small molecule modulators of an algal-bacterial symbiosis. J. Am. Chem. Soc. 2011, 133, 18343–18349. [Google Scholar] [CrossRef]
- Cude, W.N.; Mooney, J.; Tavanaei, A.A.; Hadden, M.K.; Frank, A.M.; Gulvik, C.A.; May, A.L.; Buchan, A. Production of the antimicrobial secondary metabolite indigoidine contributes to competitive surface colonization by the marine roseobacter Phaeobacter sp. strain Y4I. Appl. Environ. Microbiol. 2012, 78, 4771–4780. [Google Scholar] [CrossRef]
- Krupke, A.; Hmelo, L.R.; Ossolinski, J.E.; Mincer, T.J.; Van Mooy, B.A.S. Quorum sensing plays a complex role in regulating the enzyme hydrolysis activity of microbes associated with sinking particles in the ocean. Front. Mar. Sci. 2016, 3, 55. [Google Scholar] [CrossRef]
- Kumar, C.G.; Joo, H.-S.; Choi, J.-W.; Koo, Y.-M.; Chang, C.-S. Purification and characterization of an extracellular polysaccharide from haloalkalophilic Bacillus sp. I-450. Enzyme Microb. Technol. 2004, 34, 673–681. [Google Scholar] [CrossRef]
- Llamas, I.; Quesada, E.; Martínez-Cánovas, M.J.; Gronquist, M.; Eberhard, A.; Gonzalez, J.E. Quorum sensing in halophilic bacteria: detection of N-acyl-homoserine lactones in the exopolysaccharide-producing species of Halomonas. Extremophiles 2005, 9, 333–341. [Google Scholar] [CrossRef]
- Llamas, I.; Mata, J.A.; Tallon, R.; Bressollier, P.; Urdaci, M.C.; Quesada, E.; Béjar, V. Characterization of the exopolysaccharide produced by Salipiger mucosus A3T, a halophilic species belonging to the Alphaproteobacteria, isolated on the Spanish mediterranean seaboard. Mar. Drugs 2010, 8, 2240–2251. [Google Scholar] [CrossRef]
- Voordeckers, J.W.; Starovoytov, V.; Vetriani, C. Caminibacter mediatlanticus sp. nov., a thermophilic, chemolithoautotrophic, nitrate-ammonifying bacterium isolated from a deep-sea hydrothermal vent on the Mid-Atlantic Ridge. Int. J. Syst. Evol. Microbiol. 2005, 55, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Raguénès, G.H.C.; Peres, A.; Ruimy, R.; Pignet, P.; Christen, R.; Loaec, M.; Rougeaux, H.; Barbier, G.; Guezennec, J.G. Alteromonas infernus sp. nov., a new polysaccharide-producing bacterium isolated from a deep-sea hydrothermal vent. J. Appl. Microbiol. 1997, 82, 422–430. [Google Scholar] [CrossRef]
- Floris, R.; Rizzo, C.; Lo Giudice, A. Biosurfactants from Marine Microorganisms. Metabolomics - New Insights into Biol. Med. 2020. [Google Scholar]
- Gutiérrez-Arnillas, E.; Rodríguez, A.; Sanromán, M.A.; Deive, F.J. New sources of halophilic lipases: Isolation of bacteria from Spanish and Turkish saltworks. Biochem. Eng. J. 2016, 109, 170–177. [Google Scholar] [CrossRef]
- Gutierrez, T.; Singleton, D.R.; Berry, D.; Yang, T.; Aitken, M.D.; Teske, A. Hydrocarbon-degrading bacteria enriched by the Deepwater Horizon oil spill identified by cultivation and DNA-SIP. ISME J. 2013, 7, 2091–2104. [Google Scholar] [CrossRef]
- Heng, S.-P.; Letchumanan, V.; Deng, C.-Y.; Ab Mutalib, N.-S.; Khan, T.M.; Chuah, L.-H.; Chan, K.-G.; Goh, B.-H.; Pusparajah, P.; Lee, L.-H. Vibrio vulnificus: an environmental and clinical burden. Front. Microbiol. 2017, 8, 997. [Google Scholar] [CrossRef]
- Zhou, Z.; St John, E.; Anantharaman, K.; Reysenbach, A.-L. Global patterns of diversity and metabolism of microbial communities in deep-sea hydrothermal vent deposits. Microbiome 2022, 10, 1–22. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, Y.; Liu, Y.; Liu, B.; Wang, D.; Xu, Y.; Wei, Y. Pseudomonas marianensis sp. nov., a marine bacterium isolated from deep-sea sediments of the Mariana Trench. Arch. Microbiol. 2022, 204, 1–7. [Google Scholar] [CrossRef]
- Casillo, A.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Exopolysaccharides from marine and marine extremophilic bacteria: Structures, properties, ecological roles and applications. Mar. Drugs 2018, 16. [Google Scholar] [CrossRef]
- Weiland-Bräuer, N. Friends or foes—microbial interactions in nature. Biology (Basel). 2021, 10, 496. [Google Scholar] [CrossRef]
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.Z.; Sasidharan, R.S.; Alghamdi, H.A.; Dang, H. Understanding Interaction Patterns within Deep-Sea Microbial Communities and Their Potential Applications. Mar. Drugs 2022, 20, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Keleştemur, S.; Avci, E.; Çulha, M. Raman and surface-enhanced Raman scattering for biofilm characterization. Chemosensors 2018, 6, 5. [Google Scholar] [CrossRef]
- Bodelón, G.; Montes-García, V.; López-Puente, V.; Hill, E.H.; Hamon, C.; Sanz-Ortiz, M.N.; Rodal-Cedeira, S.; Costas, C.; Celiksoy, S.; Pérez-Juste, I. Detection and imaging of quorum sensing in Pseudomonas aeruginosa biofilm communities by surface-enhanced resonance Raman scattering. Nat. Mater. 2016, 15, 1203–1211. [Google Scholar] [CrossRef]
- Prescott, R.D.; Decho, A.W. Flexibility and adaptability of quorum sensing in nature. Trends Microbiol. 2020, 28, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Qu, Y.; Wu, W.; Li, S.; Chen, Z.; Lian, S.; Jing, J. QSP: An open sequence database for quorum sensing related gene analysis with an automatic annotation pipeline. Water Res. 2023, 235, 119814. [Google Scholar] [CrossRef]
- May, A.L.; Eisenhauer, M.E.; Coulston, K.S.; Campagna, S.R. Detection and quantitation of bacterial acylhomoserine lactone quorum sensing molecules via liquid chromatography–isotope dilution tandem mass spectrometry. Anal. Chem. 2012, 84, 1243–1252. [Google Scholar] [CrossRef]
- Senni, K.; Gueniche, F.; Changotade, S.; Septier, D.; Sinquin, C.; Ratiskol, J.; Lutomski, D.; Godeau, G.; Guezennec, J.; Colliec-Jouault, S. Unusual glycosaminoglycans from a deep sea hydrothermal bacterium improve fibrillar collagen structuring and fibroblast activities in engineered connective tissues. Mar. Drugs 2013, 11, 1351–1369. [Google Scholar] [CrossRef]
- Montgomery, K.; Charlesworth, J.C.; LeBard, R.; Visscher, P.T.; Burns, B.P. Quorum sensing in extreme environments. Life 2013, 3, 131–148. [Google Scholar] [CrossRef]
- Bzdrenga, J.; Daude, D.; Remy, B.; Jacquet, P.; Plener, L.; Elias, M.; Chabriere, E. Biotechnological applications of quorum quenching enzymes. Chem. Biol. Interact. 2017, 267, 104–115. [Google Scholar] [CrossRef]
- Liao, N.Q.; Li, H.M. Conceivable bioremediation techniques based on quorum sensing. Appl. Mech. Mater. 2013, 295, 39–44. [Google Scholar] [CrossRef]
- Sivasankar, P.; Poongodi, S.; Seedevi, P.; Sivakumar, M.; Murugan, T.; Loganathan, S. Bioremediation of wastewater through a quorum sensing triggered MFC: A sustainable measure for waste to energy concept. J. Environ. Manage. 2019, 237, 84–93. [Google Scholar] [CrossRef]
- Smith, J.L.; Fratamico, P.M.; Novak, J.S. Quorum sensing: a primer for food microbiologists. J. Food Prot. 2004, 67, 1053–1070. [Google Scholar] [CrossRef] [PubMed]
- Tonkin, M.; Khan, S.; Wani, M.Y.; Ahmad, A. Quorum sensing-a stratagem for conquering multi-drug resistant pathogens. Curr. Pharm. Des. 2021, 27, 2835–2847. [Google Scholar] [CrossRef]
- Kameswaran, S.; Ramesh, B. Quenching and Quorum Sensing in Bacterial Bio-films. Res. Microbiol. 2023, 104085. [Google Scholar] [CrossRef] [PubMed]
- Abbamondi, G.R.; Tommonaro, G. Research progress and hopeful strategies of application of quorum sensing in food, agriculture and nanomedicine. Microorganisms 2022, 10, 1192. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Gerchman, Y.; Collins, C.H.; Arnold, F.H.; Weiss, R. A synthetic multicellular system for programmed pattern formation. Nature 2005, 434, 1130–1134. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.C.; Patel, S.K.S.; Kang, Y.C.; Lee, J.-K. Quorum sensing inhibitors as antipathogens: biotechnological applications. Biotechnol. Adv. 2019, 37, 68–90. [Google Scholar] [CrossRef]
- Shaaban, M.; Elgaml, A.; Habib, E.-S.E. Biotechnological applications of quorum sensing inhibition as novel therapeutic strategies for multidrug resistant pathogens. Microb. Pathog. 2019, 127, 138–143. [Google Scholar] [CrossRef]
Figure 1.
An illustration of the working system of the autoinducer mechanism in bacteria. (Image created in Bio-render online platform).
Figure 1.
An illustration of the working system of the autoinducer mechanism in bacteria. (Image created in Bio-render online platform).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



