
Submitted:
13 July 2023
Posted:
14 July 2023
You are already at the latest version
Abstract
Angiotensin II (Ang II) plays a role in several inflammatory diseases and is capable of inducing the release of inflammatory mediators in several cell types. The objective was to investigate the potential of Ang II to induce mRNA expression of inflammatory mediators by cultured primary human periodontal cells. A synergistic effect of a co-treatment with Ang II and IL1β on mRNA expression of inflammatory mediators was also explored. Primary cultures of human fibroblast-like cells isolated from gingival and periodontal ligament tissues from 3 subjects were established using explant techniques. Immunophenotyping of STRO-1, Ang II type 1 receptor (AT1R) and Ang II type 2 receptor (AT2R) was performed by flow cytometry analysis. Cell cultures were challenged with Ang II (1M) for 3, 6 and 24h with or without a co-treatment with IL1β (0.1ng/mL) for 24h. mRNA expression of inflammatory mediators was performed by real time quantitative polymerase chain reaction (RT-qPCR). We present, for the first time, a precise quantification for AT1R and AT2R on human gingival and periodontal fibroblast-like cell types; the percentage of positive immunostaining compared to the total population of cells varied from 3.35% through 5.29% for AT1R and 2.97% through 4.57% for AT2R. Ang II slightly upregulated mRNA expression of IL6 and CCL2/MCP1 in gingival cells and IL8 and PTGS2/COX2 in periodontal ligament cells. IL1β induced upregulation of IL8, IL6 CCL2/MCP1, PTGS2/COX2 and IL1β mRNA in both cell types. Co-treatment with Ang II and IL1β did not show a synergistic effect under these experimental conditions. Ang II shows a low potential to induce mRNA upregulation of inflammatory mediators in cultured primary human gingival and periodontal ligament cells, most likely due to the low percentage of Ang II receptors in such cells, and no synergistic effect with the co-treatment with IL1β.
Keywords:
Periodontium
; Renin-Angiotensin System
; Receptors
; Angiotensin
; Primary Cell Culture
; Molecular Biology
; Inflammation Mediators
1. Introduction
In addition to the ability to regulate several essential functions of the body, such as those related to blood pressure and cardiovascular homeostasis in a systemic manner, the Renin-Angiotensin System (RAS) has been identified locally in numerous tissues, collectively referred to as a “local Renin-Angiotensin System” (local RAS) or a tissue Renin-Angiotensin System (tRAS) [1,2]. Locally, the main functions of these systems are generally related to inflammation, aging, cell proliferation and fibrosis, mainly through modulation of the primary active mediator of the system, Angiotensin II (Ang II), which exerts its effects through the interaction with specific AT1 and AT2 G protein-coupled receptors [1,3]. Several studies have associated the involvement of tRAS and local RAS with the periodontal pathology, suggesting an impact on the progression of periodontitis and bone loss, caused by the unbalance on periodontal tissue [3,4,5,6]. In rats, AT1R blockade by losartan was able to modulate the progression of experimental periodontitis due to the capacity to reduce mRNA expression of proinflammatory mediators and osteoclastogenesis, reducing bone resorption and preventing bone loss [6]. Bone formation was also affected by Ang II acting via the AT1R in an ex vivo model using the embryonic chick femur organotypic culture, where negative effects of Ang II on bone formation was observed [7]. Regarding the role of Ang II/AT1R axis on inflammation, several cell types (both animal and human), such as smooth muscle cells, THP-1 monocytes, bone marrow mesenchymal stem cells, mesangial cells, adipocytes, pancreatic islet, cardiac and lung fibroblasts, and vascular smooth muscle cells, were able to respond to an Ang II challenge, expressing and/or producing proinflammatory mediators like: interleukin (IL)18, IL6, IL1β, C-X-C motif chemokine ligand 8 (CXCL8 or IL8), tumor necrosis factor α (TNFα, monocyte chemoattractant protein-1 (MCP1)/ C-C motif chemokine ligand 2 (CCL2), cyclooxygenase-2 (COX2)/prostaglandin-endoperoxide synthase 2 (PTGS2) and prostaglandin E2 (PGE2) [8,9,10,11,12,13,14,15]. Oral tissue cells were able to produce several inflammatory mediators when challenged by different stimuli. For example, IL1β when added to primary cultured oral fibroblasts, caused an upregulation of IL6, IL1β, IL8 and TNFα [5,16]. IL1β induces AT1R expression in different cell types [32,33] and is an important cytokine in periodontal disease. Studies have been shown synergistic induction of COX-2 in pulmonary fibroblasts, and MCP1 and IL6 in mesangial cells between Ang II and proinflammatory cytokines such as IL1β [28,34].When human oral fibroblasts were challenged by bacterial stimuli such as Porphyromonas gingivalis (P. gingivalis) lipopolysaccharide (LPS), Escherichia coli lipopolysaccharide (EcLPS) or Enterococcus faecalis lipoteichoic acid (EfLTA), inflammatory mediators such as macrophage inflammatory protein 1 alpha (MIP1α)/ C-C motif chemokine ligand 3 (CCL3), stromal derived factor 1 (SDF1), CXCL12, IL6, IL1α, IL1β, IL8, CCL2, CCL5, TNFα and colony stimulating factor 1 (CSF1) were produced [17,18,19]. Due to the ability of primary cultured oral cells to produce several inflammatory mediators, and the capacity of Ang II to induce an inflammatory response in diverse cell types, the hypothesis of this study was that Ang II has a potential to induce mRNA expression of inflammatory mediators in cultured primary human gingival and periodontal ligament cells.
2. Materials and Methods
2.1. Primary Cell Culture
The primary cultures of human fibroblast-like cells from gingival and periodontal ligament tissues from 3 systematically healthy adults subjects (2 females and 1 male; age range from 22-25 years) were established by explant technique [17,18,19] following third molar extraction surgery. This study was approved by the Ethics Committee for Human Research of the Bauru School of Dentistry, University of São Paulo (CAAE: 77365617.3.0000.5417). Tissues were removed and cultured in aseptic conditions. After fragmentation, tissues were incubated for cell growth in Dulbecco’s modified Eagle medium (DMEM) (Gibco™), supplemented with 10% fetal bovine serum (FBS) (Gibco™) and antibiotics (100 U/mL penicillin, 100 μg/mL streptomycin, 0.25 μg/mL amphotericin B and 0.5 μg/mL gentamicin) (Gibco™). Cultures were maintained at 37ºC in a humidified atmosphere of 5% CO2 and 95% air; culture medium was changed every 2-3 days. Cells were used between the fourth and eighth passages.
2.2. Characterization of Primary Cell Cultures
Primary gingival and periodontal ligament cells were characterized as fibroblasts by their morphology and positive staining for fibroblast surface protein (FSP), using immunofluorescence technique, as previously described [5,18,19,20,21]. Immunophenotyping of STRO-1, AT1R and AT2R (Santa Cruz Biotechnology, Dallas, Texas, EUA, Cat. No. sc-47733, sc-515884 and sc-518054) was investigated by flow cytometry in a BD FACSAria™ Fusion Cell Sorter (BD Bioscience, San Jose, CA, United States) equipment and data was analyzed using FlowJo™ analysis software. Briefly, cells were maintained under basal conditions and detached using TrypLE™ Express Enzyme (1X) (Gibco™). After counting, 106 of each cells were separated and processed. Human BD Fc Block™ was used for blocking nonspecific binding sites. Cells were stained with the specific antibodies mentioned above (1:100), for 30 minutes at 4°C, protected from light. Cells not stained were used as negative control.
2.3. Gene Expression Analysis
Cellular viability and cytotoxicity against the challenges were analyzed using AlamarBlue® Cell Viability reagent (Invitrogen™, Ambion, Thermo Fisher Scientific, Waltham, MA), according to the manufacturer’s instructions.
For gene expression analysis, cells were seeded at 2 x 105 cells/well in 6-well plate, in triplicate, in basal culture medium. After an overnight period of cell attachment, medium culture was changed to a reduced FBS (1%). After 24h, Ang II (1μM, Sigma-Aldrich®) was added to the cells for 3, 6 and 24h or IL1β (0.1ng/mL, PeproTech®) for 24h alone or with additional challenge with Ang II for 3, 6 and 24h. After the experimental periods, the supernatants were removed and the cells were lysed for RNA extraction.
Total RNA was obtained directly from cells using PureLink™ RNA Mini Kit (Invitrogen™), according to the manufacturer's instructions. RNA concentration measurement and quality assessment were performed in a spectrophotometer NanoDrop™ 1000 (Thermo Fisher Scientific). cDNA synthesis was performed using the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems™). Real-time quantitative polymerase chain reaction (RT-qPCR) was performed using a gene expression assay and proprietary primers Taqman™ Gene Expression PCR Master Mix (Applied Biosystems™) targeting mRNA for CXCL8/IL8 (Hs00174103_m1), CCL2/MCP1 (Hs00234140_m1), PTGS2/COX2 (Hs00153133_m1), IL6 (Hs00174131_m1), TNFα (Hs00174128_m1), IL1β (Hs01555410_m1) and AGTR1 (Hs99999095_m1). RPL13A (Hs03043885_g1) was used as the reference gene. All experiments were performed in the ViiA™ 7 Real-Time PCR System (Applied Biosystems™) using the comparative cycle threshold (Ct) method (∆∆Ct) as previously described [4].
2.4. Statistical Analysis
Statistical analysis was performed with GraphPad Prism 9 software (GraphPad Software, LLC. San Diego, CA, USA). Data were tested for normal distribution using the Shapiro-Wilk test. When data were parametric, an unpaired t-test was used and when data were non-parametric, the Mann-Whitney test was used observing the differences between the challenged group in relation to the respective control in the same period of evaluation. Data were also presented as means or medians and analyzed by means of one-way ANOVA followed by Tukey's post-test for multiple comparisons for parametric data, and by means of Kruskal-Wallis test followed by Dunn's post-test for non-parametric data. Differences were identified using significance level set at 95% (p<0.05).
3. Results
3.1. Characterization of Primary Cells
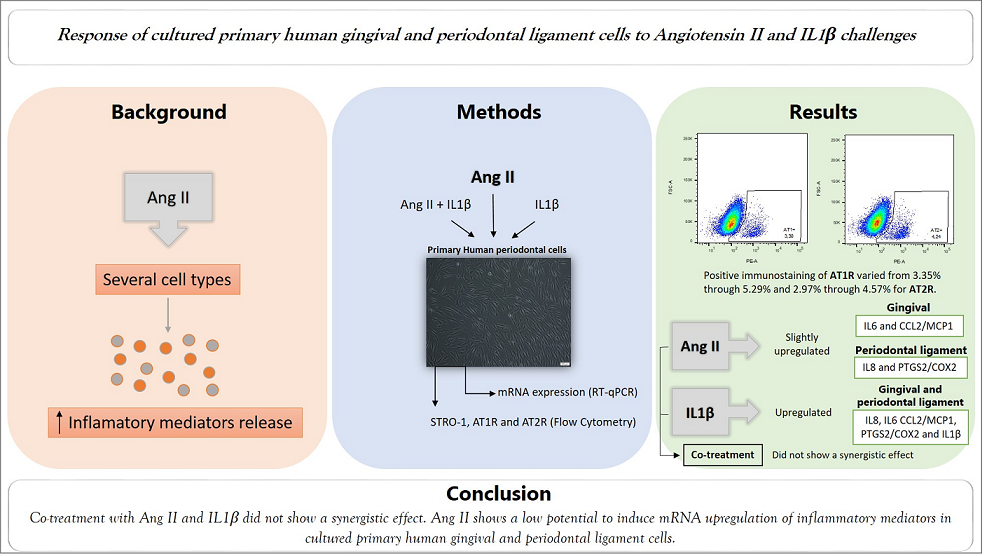
Positivity for FSP by immunofluorescence analysis indicated the fibroblastic phenotype (Figure 1A,B,C). The expression of STRO-1 was also observed, indicating the possible presence of mesenchymal stromal cells/stem cells (MSCs), as shown in Figure 1D,E. In the primary cell culture used in this work, it was possible to observe in the general population a low expression of the STRO-1 marker, which associated with FSP immunostaining, cell morphological analysis and specific culture conditions characterize such cells as fibroblasts. However, the presence of smaller and larger cells was also noted, which allowed us to consider the presence of 2 populations of cells, which we named POP1 and POP2, which are present in very different amounts, ranging from 72.6 to 83.7% for POP1 and 5.68 to 12.8% for POP2. In addition, a slightly positive STRO-1 immunostaining in these POP2 cells was observed. Ang II AT1R and AT2R expression is show in Figure 1F, 1G and 1H. A small percentage of cells express these receptors, varying from 3.35% through 5.29% for AT1R and 2.97% through 4.57% for AT2R, considering the general population.
3.2. mRNA Expression of Inflammatory Mediators in Primary Human Cells from Periodontal Tissues
3.2.1. Ang II Slightly Increased IL6 and CCL2/MCP1 Expression in Gingival Primary Cells and IL8 and PTGS2/COX2 in Periodontal Ligament Cells
mRNA expression of inflammatory mediators by the periodontal cells after Ang II challenge is described in Figure 2 and Figure 3. At time points of 24h and at 3h, Ang II challenge slightly upregulated mRNA expression of IL6 and CCL2, respectively (Figure 2B,C). With addition of Ang II, a slightly upregulated mRNA expression of IL8 and PTGS2 was observed at time points of 3h and 24h, respectively (Figure 3A,D).
3.2.2. IL1β for 24h Induced mRNA Upregulation of Several Inflammatory Mediators in Gingival and Periodontal Ligament Cells and Ang II Did Not Cause a Synergistic Effect
4. Discussion
Several scientific reports have demonstrated inflammatory functions exerted by Ang II are linked primarily to AT1R in several cells and tissues [8,9,22]. Oral cells, mainly fibroblasts, play an important role in inflammatory responses to a wide variety of stimuli [5,16,17,19]. Notably, such cells show positive fluorescent immunostaining for AT1R [4,23]. Therefore, we hypothesized that Ang II, mediated through action on AT1R of fibroblasts from oral tissues, would modulate immunoinflammatory processes in oral pathologies, such as gingivitis and periodontitis.
It is well recognized that gingival and periodontal ligament fibroblasts express AT1R, [4,16] however, the quantification of these receptors had never been directly analyzed. To the best of our knowledge, we present for the first time a precise quantification through immunostaining/flow cytometry for AT1R and AT2R on such cell types. The percentage varied from 3.35% through 5.29% for AT1R and 2.97% through 4.57% for AT2R. Although a quantification was not performed for other types of oral cells, like stem cells of apical papilla, it is known they present a slightly positive immunostaining for AT1R, thus indicating a low presence of AT1R [24]. Besides that, we have to consider the existence of nuclear AT1R and the effects that Ang II can generate intracellularly [16,25].
It is important to consider that, in this work, we restricted the study to fibroblast-like cells, but in the living organism other cells exist in the periodontal tissue, which also contain AT1R and AT2R [4] and those may also play a role in the response to Ang II. Along those lines, it is important to emphasize the anti-inflammatory effects of AT1R antagonism by losartan, which was able to decrease inflammation and bone loss in periodontal tissues of rats with experimentally-induced periodontal disease [6]. Although slight immunostaining for AT1R and AT2R was observed in the cell cultures of the present work, the importance of such receptors cannot be disregarded in the whole animal since other cells of the periodontal tissue may interact. Moreover, for future studies, we consider to perform cell isolation and conduct the experiments just with the positive cells for the AT1R and AT2R. Also, the systemic effects of Ang II binding to its receptors should always be considered in the whole organism.
Primary gingival and periodontal ligaments cells were characterized as fibroblasts by their morphology and positive staining for FSP, using immunofluorescence technique, as previously described [5,18,19,20,21]. However, MSCs also can be isolated from human dental tissues, show a fibroblast-like morphology and express the same surface markers like FSP, being considered phenotypically indistinguishable from fibroblasts [26]. In this study, we used the distinct marker, STRO-1, which is highly expressed in MSCs but not in fibroblasts to allow distinction in these groups of cells [27]. Therefore, we considered that the primary cultures used in this work to be fibroblast-like oral cells with a low, variable proportion of MSCs.
The inflammatory mediators analyzed in this study were chosen based on previous work that detected the modulatory role of Ang II in fibroblasts from other tissues, such as kidney, heart and lung [14,28,29]. Besides that, such mediators are important to the periodontal pathology and inflammatory diseases. CXCL8/IL8 and CCL2/MCP1 are chemokines that have great importance in the inflammatory process due to the ability to attract neutrophils and monocytes, respectively, to the site of inflammation [30,31]. IL6 is also an important proinflammatory cytokine with effects on osteoclastogenesis and bone resorption [5]. PTGS2 can be induced by cytokines and during inflammation; it is responsible for elevated production of prostaglandins, which are also potent inflammatory mediators [14,28]. Rat cardiac fibroblasts challenged with Ang II showed intense expression of COX2 protein in a time-dependent manner, which was correlated with a significant increase in prostaglandin E2 release [14]. Ang II also induced prostaglandin E2 release in human gingival fibroblast [22] and COX2 expression in mouse lung fibroblasts, but not in human lung adenocarcinoma (A549) and normal human bronchial epithelial cells (BEAS-2B) [28]. In the present work, Ang II was not capable of inducing PTGS2 expression in gingival fibroblasts however a slight upregulation was observed in cells derived from the periodontal ligament at the 24h time point.
The mRNA expression of AT1R was downregulated with addition of Ang II at the 24h time point in both cell types. IL1β alone (0.1ng/mL) was not capable of inducing slightly increased of AT1R, differently from what was observed in other studies [32,33]. Taken together, these results suggest that fibroblasts originating from different compartments of the periodontal tissue (gingiva and periodontal ligament) show different abilities to regulate AT1R mRNA. Importantly, IL1β was able to significantly increase mRNA expression of several inflammatory mediators with great importance for periodontal pathology, such as IL8, PTGS2, CCL2, IL6, IL1β, when compared to the non-challenged group of cells. It would be important to measure protein for any cytokines that were significantly altered at the RNA level. Unfortunately, we were not able to do that and, therefore, this is a limitation of the present work.
Although there are reports documenting the synergism between IL1β and Ang II, increasing COX2 production in lung fibroblasts [28] and MCP1 and IL6 in mesangial cells [34], no such interaction was observed in the present study suggesting the intracellular mechanisms facilitating such a molecular interaction are not present in these cells.
5. Conclusions
Ang II challenge to cultured primary human gingival and periodontal ligament cells was capable of inducing only a slight upregulation of inflammatory mediators the profile of which differed between the cell types studied. This attenuated effect, compared to other cell types was most likely due to the low percentage of Ang II receptors in the fibroblast-like cells of the gingiva and periodontal ligament. Compared to Ang II, IL1β challenge produced a robust upregulation of mRNA expression of inflammatory mediators without a synergistic effect with Ang II under the experimental conditions of our experiments.
Author Contributions
All authors have made substantial contributions to conception and design of the study. TFG, TJD, BLCI and RAS have been involved in data collection and data analysis. TFG, TJD, BLCI, RAS, VSL, SHPO, MHF, ASG and CFS have been involved in data interpretation, drafting the manuscript and revising it critically and have given final approval of the version to be published.
Funding
This research was funded by São Paulo Research Foundation (FAPESP), grants: #2015/03965-2 (CFS), #2017/19227-6 and #2018/2393-2 (TFG) and Coordination for the Improvement of Higher Education Personnel (CAPES - Finance Code 001) (TFG).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Ethics Committee of Bauru School of Dentistry, University of São Paulo (CAAE: 77365617.3.0000.5417; date of approval: 05/17/2021).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
In The authors express their gratitude to Adriano de Souza Pessôa (Bauru School of Dentistry/University of São Paulo/Department of Biological Sciences) for his assistance with some flow cytometry data analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Benigni, A.; Cassis, P.; Remuzzi, G. Angiotensin II Revisited: New Roles in Inflammation, Immunology and Aging. EMBO Mol. Med. 2010, 2, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Saravi, B.; Lang, G.; Ülkümen, S.; Burchard, T.; Weihrauch, V.; Patzelt, S.; Boeker, M.; Li, Z.; Woelber, J.P. The Tissue Renin-Angiotensin System (Tras) and the Impact of Its Inhibition on Inflammation and Bone Loss in the Periodontal Tissue. Eur. Cells Mater. 2020, 40, 203–226. [Google Scholar] [CrossRef]
- Santos, C.F.; Akashi, A.E.; Dionísio, T.J.; Sipert, C.R.; Didier, D.N.; Greene, A.S.; Oliveira, S.H.P.; Pereira, H.J.V.; Becari, C.; Oliveira, E.B.; et al. Characterization of a Local Renin-Angiotensin System in Rat Gingival Tissue. J. Periodontol. 2009, 80, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.F.; Morandini, A.C.; Dionísio, T.J.; Faria, F.A.; Lima, M.C.; Figueiredo, C.M.; Colombini-Ishikiriama, B.L.; Sipert, C.R.; Maciel, R.P.; Akashi, A.P.; et al. Functional Local Renin-Angiotensin System in Human and Rat Periodontal Tissue. PLoS ONE 2015, 10, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Gabriele, L.G.; Morandini, A.C.; Dionísio, T.J.; Santos, C.F. Angiotensin II Type 1 Receptor Knockdown Impairs Interleukin-1β-Induced Cytokines in Human Periodontal Fibroblasts. J. Periodontol. 2017, 88, e1–e11. [Google Scholar] [CrossRef] [PubMed]
- Dionísio, T.J.; Souza, G.P.; Colombini-Ishikiriama, B.L.; Garbieri, T.F.; Parisi, V.A.; Oliveira, G.M.; Cano, I.P.; Rodini, C.O.; Oliveira, S.H.P.; Greene, A.S.; et al. AT1 Receptor Antagonism Promotes Bone Loss Attenuation in Experimental Periodontitis, Blocks Inflammatory Mediators, and Upregulates Antioxidant Enzymes and Bone Formation Markers. J. Periodontol. 2020, 91, 533–544. [Google Scholar] [CrossRef]
- Garbieri, T.F.; Martin, V.; dos Santos, C.F.; Gomes, P.d.S.; Fernandes, M.H. The Embryonic Chick Femur Organotypic Model as a Tool to Analyze the Angiotensin II Axis on Bone Tissue. Pharmaceuticals 2021, 14. [Google Scholar] [CrossRef]
- Valente, A.J.; Yoshida, T.; Murthy, S.N.; Sakamuri, S.S.V.P.; Katsuyama, M.; Clark, R.A.; Delafontaine, P.; Chandrasekar, B. Angiotensin II Enhances AT 1-Nox1 Binding and Stimulates Arterial Smooth Muscle Cell Migration and Proliferation through AT 1, Nox1, and Interleukin-18. Am. J. Physiol. - Hear. Circ. Physiol. 2012, 303, 282–296. [Google Scholar] [CrossRef]
- Vogiatzi, K.; Apostolakis, S.; Vlata, Z.; Krabovitis, E.; Spandidos, D.A. Opposite Effect of Angiotensin Receptor Blockade on CXCL8 Production and CXCR1/2 Expression of Angiotensin II-Treated THP-1 Monocytes. Exp. Ther. Med. 2013, 5, 987–991. [Google Scholar] [CrossRef]
- ZHANG, F.; LU, M.; WANG, H.; REN, T. Aspirin Attenuates Angiotensin II-Induced Inflammation in Bone Marrow Mesenchymal Stem Cells via the Inhibition of ERK1/2 and NF-ΚB Activation. Biomed. Reports 2013, 1, 930–934. [Google Scholar] [CrossRef]
- Zhang, F.; Sun, D.; Chen, J.; Guan, N.; Huo, X.; Xi, H. Simvastatin Attenuates Angiotensin IIInduced Inflammation and Oxidative Stress in Human Mesangial Cells. Mol. Med. Rep. 2015, 11, 1246–1251. [Google Scholar] [CrossRef] [PubMed]
- Sauter, N.S.; Thienel, C.; Plutino, Y.; Kampe, K.; Dror, E.; Traub, S.; Timper, K.; Bédat, B.; Pattou, F.; Kerr-Conte, J.; et al. Angiotensin II Induces Interleukin-1b-Mediated Islet Inflammation and β-Cell Dysfunction Independently of Vasoconstrictive Effects. Diabetes 2015, 64, 1273–1283. [Google Scholar] [CrossRef]
- Skurk, T.; Van Harmelen, V.; Hauner, H. Angiotensin II Stimulates the Release of Interleukin-6 and Interleukin-8 from Cultured Human Adipocytes by Activation of NF-ΚB. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1199–1203. [Google Scholar] [CrossRef] [PubMed]
- Scheuren, N.; Jacobs, M.; Ertl, G.; Schorb, W. Cyclooxygenase-2 in Myocardium Stimulation by Angiotensin-II in Cultured Cardiac Fibroblasts and Role at Acute Myocardial Infarction. J. Mol. Cell. Cardiol. 2002, 34, 29–37. [Google Scholar] [CrossRef]
- Han, Y.; Runge, M.S.; Brasier, A.R. Angiotensin II Induces Interleukin-6 Transcription in Vascular Smooth Muscle Cells through Pleiotropic Activation of Nuclear Factor-Κb Transcription Factors. Circ. Res. 1999, 84, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Hasegawa-Nakamura, K.; Sakoda, K.; Matsuyama, T.; Noguchi, K. Involvement of Angiotensin II Type 1 Receptors in Interleukin-1β-Induced Interleukin-6 Production in Human Gingival Fibroblasts. Eur. J. Oral Sci. 2011, 119, 345–351. [Google Scholar] [CrossRef]
- Morandini, A.C.F.; Sipert, C.R.; Gasparoto, T.H.; Greghi, S.L.A.; Passanezi, E.; Rezende, M.L.R.; Sant’ana, A.P.; Campanelli, A.P.; Garlet, G.P.; Santos, C.F. Differential Production of Macrophage Inflammatory Protein-1α, Stromal-Derived Factor-1, and IL-6 by Human Cultured Periodontal Ligament and Gingival Fibroblasts Challenged With Lipopolysaccharide From P. Gingivalis. J. Periodontol. 2010, 81, 310–317. [Google Scholar] [CrossRef]
- Sipert, C.R.; Morandini, A.C.; Dionísio, T.J.; Machado, M.A.A.M.; Oliveira, S.H.P.; Campanelli, A.P.; Kuo, W.P.; Santos, C.F. In Vitro Regulation of CCL3 and CXCL12 by Bacterial By-Products Is Dependent on Site of Origin of Human Oral Fibroblasts. J. Endod. 2014, 40, 95–100. [Google Scholar] [CrossRef]
- Colombini-Ishikiriama, B.L.; Dionisio, T.J.; Garbieri, T.F.; Da Silva, R.A.; Machado, M.A.A.M.; De Oliveira, S.H.P.; Lara, V.S.; Greene, A.S.; Santos, C.F. What Is the Response Profile of Deciduous Pulp Fibroblasts Stimulated with E. Coli LPS and E. Faecalis LTA? BMC Immunol. 2020, 21, 1–9. [Google Scholar] [CrossRef]
- Morandini, A.C.F.; Chaves Souza, P.P.; Ramos-Junior, E.S.; Brozoski, D.T.; Sipert, C.R.; Souza Costa, C.A.; Santos, C.F. Toll-Like Receptor 2 Knockdown Modulates Interleukin (IL)-6 and IL-8 but Not Stromal Derived Factor-1 (SDF-1/CXCL12) in Human Periodontal Ligament and Gingival Fibroblasts. J. Periodontol. 2013, 84, 535–544. [Google Scholar] [CrossRef]
- Sipert, C.R.; Morandini, A.C.d.F.; Modena, K.C.d.S.; Dionisio, T.J.; Machado, M.A.A.M.; de Oliveira, S.H.P.; Campanelli, A.P.; Santos, C.F. CCL3 and CXCL12 Production in Vitro by Dental Pulp Fibroblasts from Permanent and Deciduous Teeth Stimulated by Porphyromonas Gingivalis LPS. J. Appl. Oral Sci. 2013, 21, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Segawa, M.; Nakao, S.; Ogata, Y.; Sugiya, H.; Furuyama, S. Angiotensin II Induces Prostaglandin E2 Release in Human Gingival Fibroblasts. Life Sci. 2003, 72, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Monnouchi, S.; Maeda, H.; Fujii, S.; Tomokiyo, A.; Kono, K.; Akamine, A. The Roles of Angiotensin II in Stretched Periodontal Ligament Cells. J. Dent. Res. 2011, 90, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Pizzatto, L.N.; Meneses, C.C.B.; Diniz, E.A.; Dionísio, T.J.; Santos, C.F.; Sipert, C.R. Angiotensin II Regulates Proliferation and Function of Stem Cells of Apical Papilla. J. Endod. 2020, 46, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Morinelli, T.A.; Raymond, J.R.; Baldys, A.; Yang, Q.; Lee, M.-H.; Luttrell, L.; Ullian, M.E. Identification of a Putative Nuclear Localization Sequence within ANG II AT(1A) Receptor Associated with Nuclear Activation. Am. J. Physiol. Cell Physiol. 2007, 292, C1398–C1408. [Google Scholar] [CrossRef]
- Denu, R.A.; Nemcek, S.; Bloom, D.D.; Goodrich, A.D.; Kim, J.; Mosher, D.F.; Hematti, P. Fibroblasts and Mesenchymal Stromal/Stem Cells Are Phenotypically Indistinguishable. Acta Haematol. 2016, 136, 85–97. [Google Scholar] [CrossRef]
- Du, L.; Yang, P.; Ge, S. Isolation and Characterization of Human Gingiva-Derived Mesenchymal Stem Cells Using Limiting Dilution Method. J. Dent. Sci. 2016, 11, 304–314. [Google Scholar] [CrossRef]
- Matsuzuka, T.; Miller, K.; Pickel, L.; Doi, C.; Ayuzawa, R.; Tamura, M. The Synergistic Induction of Cyclooxygenase-2 in Lung Fibroblasts by Angiotensin II and pro-Inflammatory Cytokines. Mol. Cell. Biochem. 2009, 320, 163–171. [Google Scholar] [CrossRef]
- Yokoyama, T.; Sekiguchi, K.; Tanaka, T.; Tomaru, K.; Arai, M.; Suzuki, T.; Nagai, R. Angiotensin II and Mechanical Stretch Induce Production of Tumor Necrosis Factor in Cardiac Fibroblasts. Am. J. Physiol. Circ. Physiol. 1999, 276, H1968–H1976. [Google Scholar] [CrossRef]
- Sipert, C.R.; Moraes, I.G.; Bernardinelli, N.; Garcia, R.B.; Bramante, C.M.; Gasparoto, T.H.; Figueira, E.A.; Dionísio, T.J.; Campanelli, A.P.; Oliveira, S.H.P.; et al. Heat-Killed Enterococcus Faecalis Alters Nitric Oxide and CXCL12 Production but Not CXCL8 and CCL3 Production by Cultured Human Dental Pulp Fibroblasts. J. Endod. 2010, 36, 91–94. [Google Scholar] [CrossRef]
- Morandini, A.C.; Chaves Souza, P.P.; Ramos-Junior, E.S.; Souza Costa, C.A.; Santos, C.F. MyD88 or TRAM Knockdown Regulates Interleukin (IL)-6, IL-8, and CXCL12 MRNA Expression in Human Gingival and Periodontal Ligament Fibroblasts. J. Periodontol. 2013, 84, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Gurantz, D.; Cowling, R.T.; Varki, N.; Frikovsky, E.; Moore, C.D.; Greenberg, B.H. IL-1β and TNF-α Upregulate Angiotensin II Type 1 (AT 1) Receptors on Cardiac Fibroblasts and Are Associated with Increased AT1 Density in the Post-MI Heart. J. Mol. Cell. Cardiol. 2005, 38, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Imaizumi, T.; Tanji, K.; Sakaki, H.; Metoki, N.; Sato, Y.; Wakabayashi, K.; Kimura, H.; Satoh, K. Interleukin-1β Enhances the Angiotensin-Induced Expression of Plasminogen Activator Inhibitor-1 through Angiotensin Receptor Upregulation in Human Astrocytes. Brain Res. 2006, 1073–1074, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Alique, M.; Sánchez-López, E.; Rayego-Mateos, S.; Egido, J.; Ortiz, A.; Ruiz-Ortega, M. Angiotensin II, via Angiotensin Receptor Type 1/Nuclear Factor-ΚB Activation, Causes a Synergistic Effect on Interleukin-1-β-Induced Inflammatory Responses in Cultured Mesangial Cells. JRAAS - J. Renin-Angiotensin-Aldosterone Syst. 2015, 16, 23–32. [Google Scholar] [CrossRef]
Figure 1.
Phenotypic characterization of cultured primary periodontal cells. Cells were plated (104 cells/well) and showed positive staining for FSP-1 protein in green (A - cells from gingival tissue and B - cells from periodontal ligament tissue) through immunofluorescence analysis. Cell nuclei were stained with DAPI- blue (4′,6- diamidino-2- phenylindole dihydrochloride) C - negative control. Images were captured by a confocal microscope (TCS model, SPE, Leica®, Mannheim, Germany). Scale bars - 20 μm. Representative image of STRO-1 expression by flow cytometry analysis considering the cells divided into two populations (POP1 and POP2) without (D) and with (E) staining. AT1R and AT2R expression by cells considering the whole population. F - Strategy gate to define the stained cells compared to non-stained cells. Percentage of stained cells for AT1R (G) and AT2R (H).
Figure 1.
Phenotypic characterization of cultured primary periodontal cells. Cells were plated (104 cells/well) and showed positive staining for FSP-1 protein in green (A - cells from gingival tissue and B - cells from periodontal ligament tissue) through immunofluorescence analysis. Cell nuclei were stained with DAPI- blue (4′,6- diamidino-2- phenylindole dihydrochloride) C - negative control. Images were captured by a confocal microscope (TCS model, SPE, Leica®, Mannheim, Germany). Scale bars - 20 μm. Representative image of STRO-1 expression by flow cytometry analysis considering the cells divided into two populations (POP1 and POP2) without (D) and with (E) staining. AT1R and AT2R expression by cells considering the whole population. F - Strategy gate to define the stained cells compared to non-stained cells. Percentage of stained cells for AT1R (G) and AT2R (H).
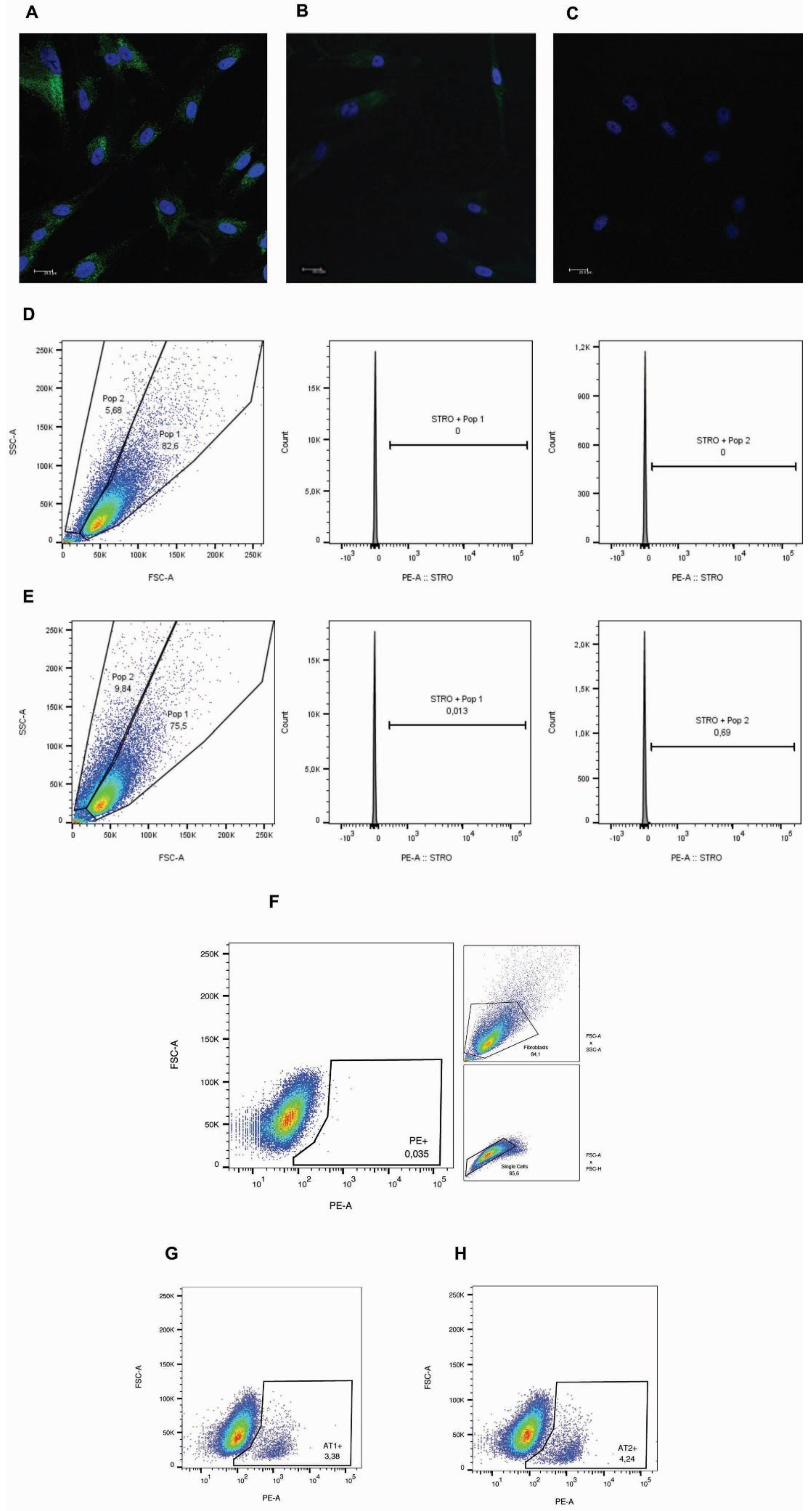
Figure 2.
Gene expression analysis by RT-qPCR of inflammatory mediators and AT1R with Ang II challenge. Relative expression levels of the target mRNA to RPL13A mRNA from 3 donors in triplicate are displayed in graphs. Primary gingival cells were challenged by Ang II (1μM) for 3, 6 and 24h and RT-qPCR analysis were performed for the inflammatory mediators: IL8 (A), IL6 (B), CCL2 (C), PTGS2 (D), TNF𝛼 (E) and AT1R (F). * Indicates significant difference in relation to respective control in the same experimental period (p <0.05).
Figure 2.
Gene expression analysis by RT-qPCR of inflammatory mediators and AT1R with Ang II challenge. Relative expression levels of the target mRNA to RPL13A mRNA from 3 donors in triplicate are displayed in graphs. Primary gingival cells were challenged by Ang II (1μM) for 3, 6 and 24h and RT-qPCR analysis were performed for the inflammatory mediators: IL8 (A), IL6 (B), CCL2 (C), PTGS2 (D), TNF𝛼 (E) and AT1R (F). * Indicates significant difference in relation to respective control in the same experimental period (p <0.05).
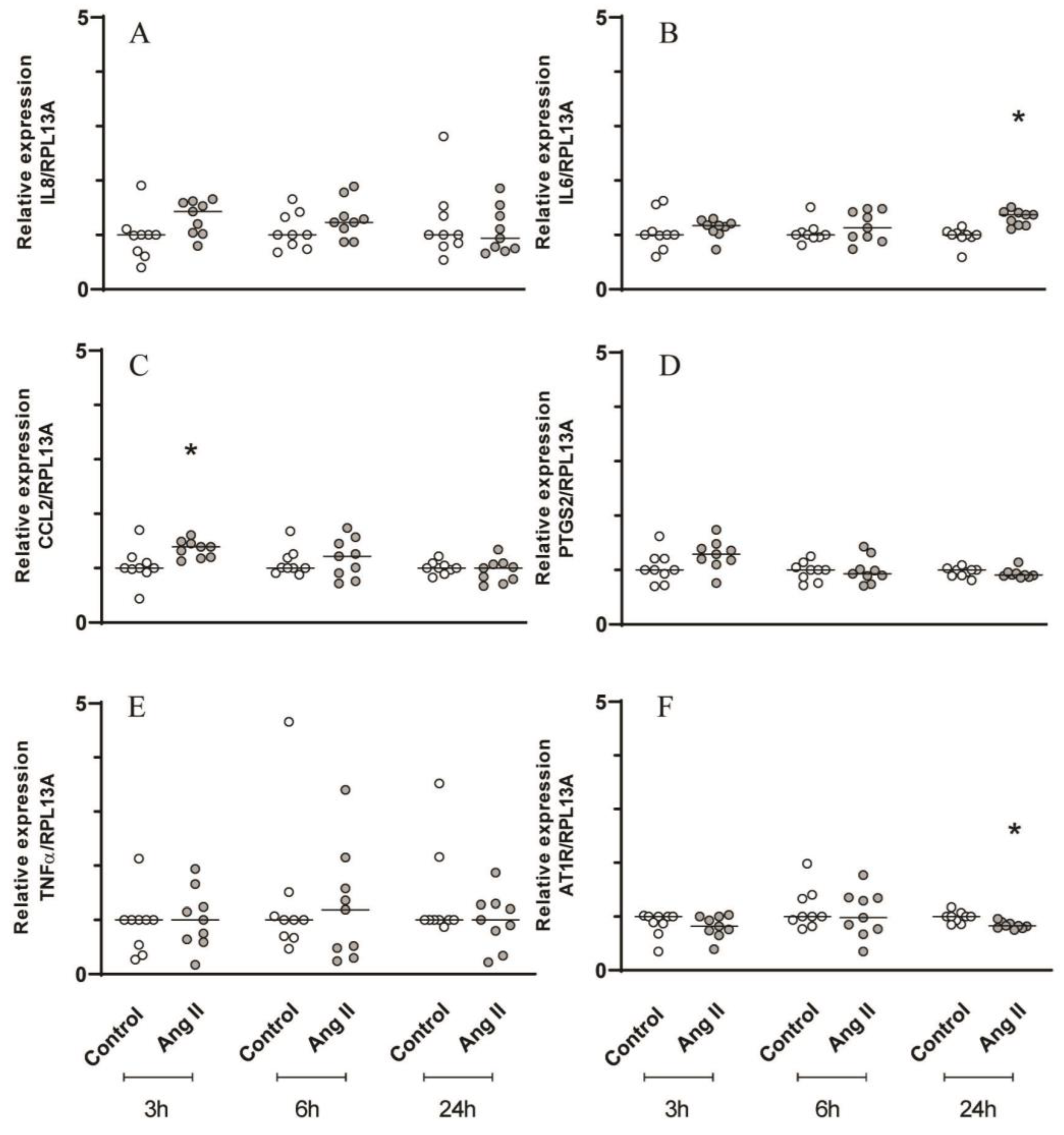
Figure 3.
Gene expression analysis by RT-qPCR of inflammatory mediators and AT1R with Ang II challenge. Relative expression levels of the target mRNA to RPL13A mRNA from 3 donors in triplicate are displayed in graphs. Primary periodontal ligament cells were challenged by Ang II (1μM) for 3, 6 and 24h and RT-qPCR analysis were performed for the inflammatory mediators: IL8 (A), IL6 (B), CCL2 (C), PTGS2 (D), TNF𝛼 (E) and AT1R (F). * Indicates significant difference in relation to respective control in the same experimental period (p <0.05).
Figure 3.
Gene expression analysis by RT-qPCR of inflammatory mediators and AT1R with Ang II challenge. Relative expression levels of the target mRNA to RPL13A mRNA from 3 donors in triplicate are displayed in graphs. Primary periodontal ligament cells were challenged by Ang II (1μM) for 3, 6 and 24h and RT-qPCR analysis were performed for the inflammatory mediators: IL8 (A), IL6 (B), CCL2 (C), PTGS2 (D), TNF𝛼 (E) and AT1R (F). * Indicates significant difference in relation to respective control in the same experimental period (p <0.05).
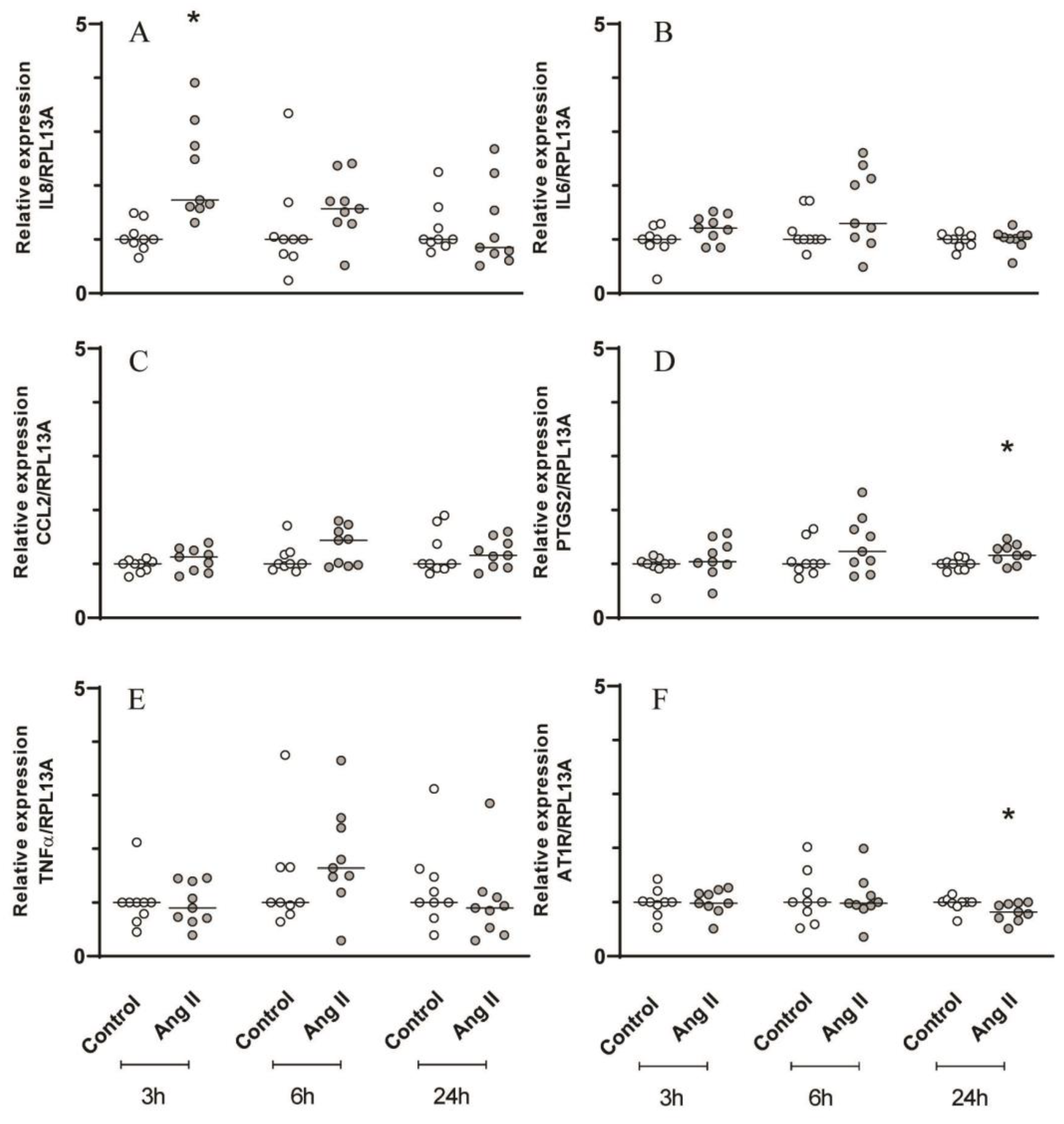
Figure 4.
Gene expression analysis by RT-qPCR of inflammatory mediators and AT1R with IL1β and Ang II challenges. Relative expression levels of the target mRNA to RPL13A mRNA from 3 donors in triplicate are displayed in graphs. Cultured primary gingival cells were challenged by IL1β (0.1 ng/mL) for 24h, 27h, 30h and 48h or IL1β for 24h followed by Ang II (1μM) for 3, 6 and 24h. RT-qPCR analysis was performed for the following inflammatory mediators: IL8 (A), IL6 (B), CCL2 (C), PTGS2 (D), IL1β (E) and AT1R (F). IL1β challenge alone for 27, 30 and 48h was used to compare with those groups that were challenged with IL1β followed by Ang II for 3h, 6h and 24h respectively. * Indicates significant difference in comparison to control (basal medium) (p <0.05).
Figure 4.
Gene expression analysis by RT-qPCR of inflammatory mediators and AT1R with IL1β and Ang II challenges. Relative expression levels of the target mRNA to RPL13A mRNA from 3 donors in triplicate are displayed in graphs. Cultured primary gingival cells were challenged by IL1β (0.1 ng/mL) for 24h, 27h, 30h and 48h or IL1β for 24h followed by Ang II (1μM) for 3, 6 and 24h. RT-qPCR analysis was performed for the following inflammatory mediators: IL8 (A), IL6 (B), CCL2 (C), PTGS2 (D), IL1β (E) and AT1R (F). IL1β challenge alone for 27, 30 and 48h was used to compare with those groups that were challenged with IL1β followed by Ang II for 3h, 6h and 24h respectively. * Indicates significant difference in comparison to control (basal medium) (p <0.05).
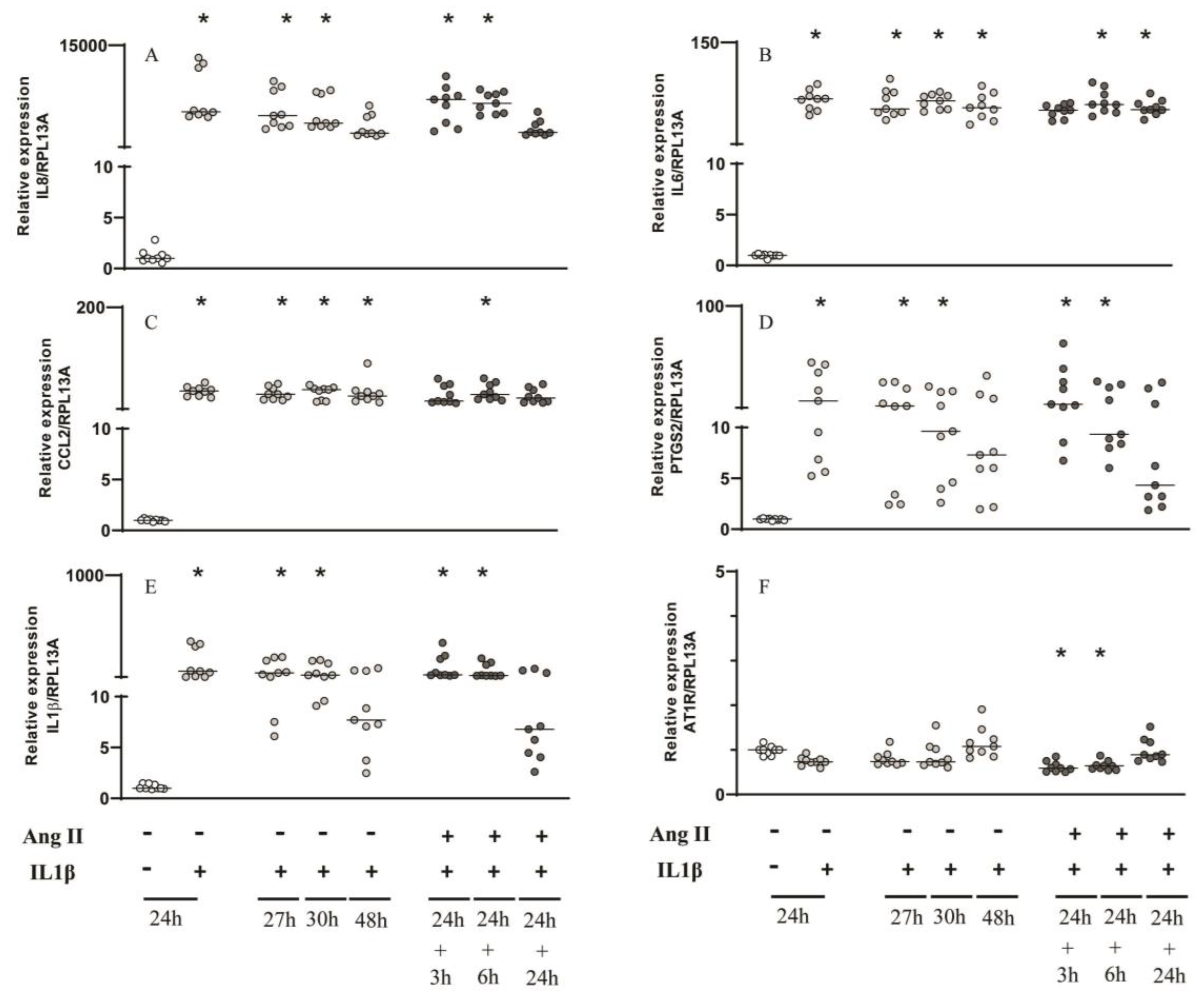
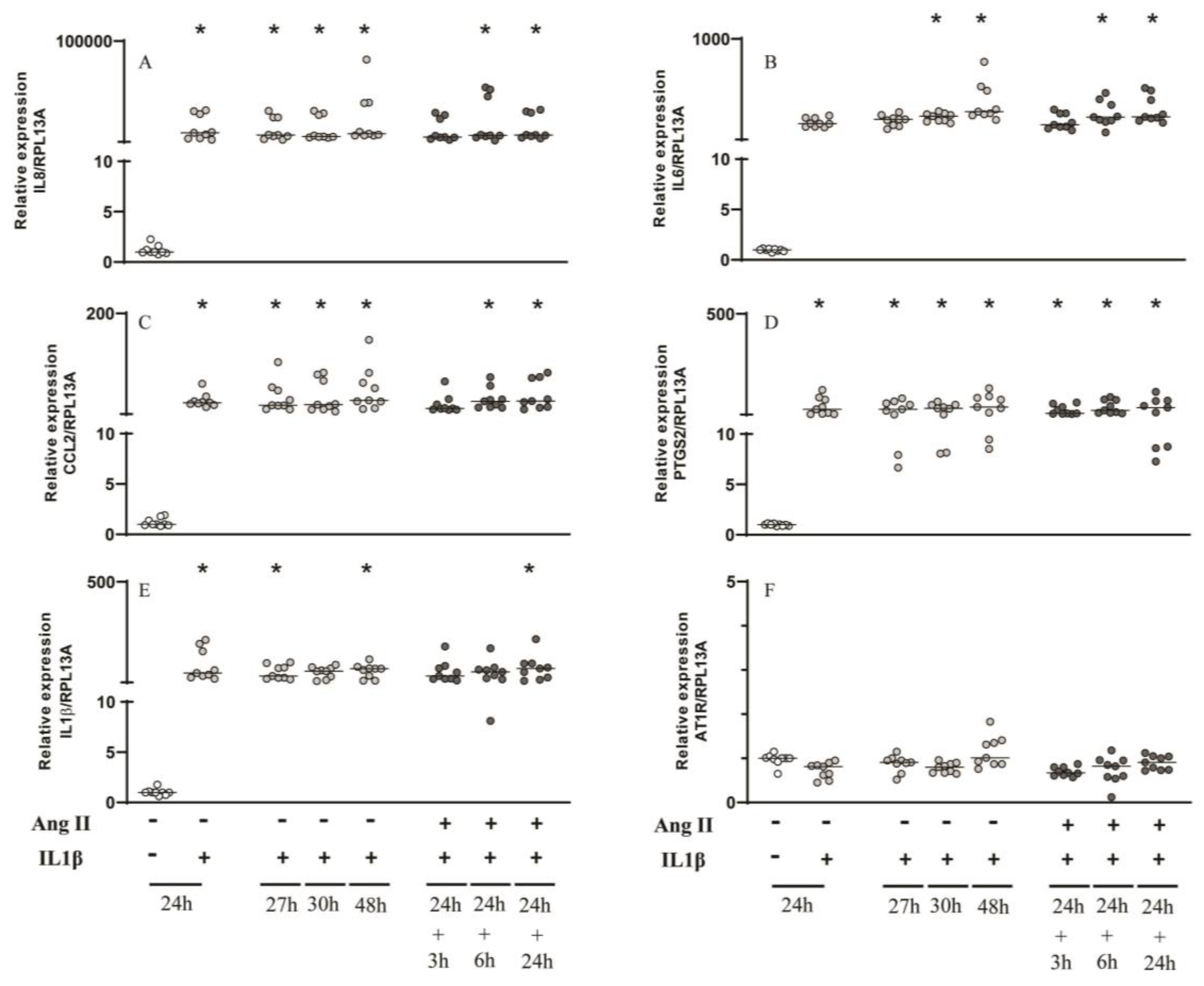
Figure 5.
Gene expression analysis by RT-qPCR of inflammatory mediators and AT1R with IL1β and Ang II challenges. Relative expression levels of the target mRNA to RPL13A mRNA from 3 donors in triplicate are displayed in graphs. Cultured primary periodontal ligament cells were challenged by IL1β (0.1 ng/mL) for 24h, 27h, 30h and 48h or IL1β for 24h followed by Ang II (1μM) for 3, 6 and 24h. RT-qPCR analysis was performed for the following inflammatory mediators: IL8 (A), IL6 (B), CCL2 (C), PTGS2 (D), IL1β (E) and AT1R (F). IL1β challenge alone for 27, 30 and 48h was used to compare with those groups that were challenged with IL1β followed by Ang II for 3h, 6h and 24h respectively. * Indicates significant difference in comparison to control (basal medium) (p <0.05).
Figure 5.
Gene expression analysis by RT-qPCR of inflammatory mediators and AT1R with IL1β and Ang II challenges. Relative expression levels of the target mRNA to RPL13A mRNA from 3 donors in triplicate are displayed in graphs. Cultured primary periodontal ligament cells were challenged by IL1β (0.1 ng/mL) for 24h, 27h, 30h and 48h or IL1β for 24h followed by Ang II (1μM) for 3, 6 and 24h. RT-qPCR analysis was performed for the following inflammatory mediators: IL8 (A), IL6 (B), CCL2 (C), PTGS2 (D), IL1β (E) and AT1R (F). IL1β challenge alone for 27, 30 and 48h was used to compare with those groups that were challenged with IL1β followed by Ang II for 3h, 6h and 24h respectively. * Indicates significant difference in comparison to control (basal medium) (p <0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



