Submitted:
13 July 2023
Posted:
14 July 2023
You are already at the latest version
Abstract
1). BACKGROUND: Cell wall deficient mycobacteria (CWDM) have seldom been isolated in bacterial culture from human subjects. particularly in clinical disease, such as Crohn’s disease. Techniques have focused on the use of polymerase chain reaction (PCR) for the detection of CWDM in association with inflammatory bowel disease. (IBD). This paper is a report of the development of methods to demonstrate the presence of CWDM using the Ziehl-Neelsen (ZN) stain. This is in addition to the detection of the growth of CWDM in fluid media. 2). METHODS: The use of the ZN stain is suitable for the demonstration of CWDM in clinical samples and cultures, when the staining methods that were common prior to the 1970’s, are adopted. Specific culture media are developed and tested in parallel. Alterations in the concentration of basic fuchsin, and changes in the decolourising solutions are necessary. Decolourising solutions such as 20% concentrated sulphuric acid in water are preferable for detection of CWDM in cultures. 3). RESULTS: CWDM can be readily demonstrated using the simple methods outlined in this report. The ability to visualise the presence of CWDM opens the door to the development of specific fluid media for the isolation of CWDM in culture. 4). CONCLUSION: The ZN stain methods optimised for the demonstration of Mycobacterium tuberculosis complex (MTBC) are not suitable for research into infectious causes of inflammatory bowel disease (IBD), as well as for the detection of CWDM in latent tuberculosis. Revisiting the techniques for the detection of CWDM may also refocus attention on inflammatory diseases considered to be idiopathic, but having hallmarks of an infectious process.
Keywords:
cell wall deficient mycobacteria
; mycobacterium paratuberculosis
; crohn’s disease
; ziehl-neelsen stain
; non-tuberculosis mycobacteria
1. Introduction
Mycobacterium species are gram-positive, catalase-positive, aerobic, and occasionally microaerophilic non-spore-forming acid-fast bacilli. They may adopt an infectious life cycle [1] in animals, while the animal response may include an immunological life cycle [2]. Members of the Mycobacterium genus are oligotrophs that evolve to survive in challenging environments. Some members of the genus are able to utilise methanol or carbon monoxide as carbon sources [3]. Culture media for the growth of mycobacteria rely on low nutrient content and are formulated for the growth of bacillary strains of mycobacteria with intact cell walls. Members of the Mycobacterium genus, in common with other bacteria, proceed through four stages of growth; the lag phase, the log (growth) phase, the decline phase, and the death phase. Each growth phase is a response to alterations in atmosphere and nutrients. Mycobacteria, both in culture media and in the host, can remain viable after the decline phase by entering a state variously called latency, dormancy, or non-replicating persistence (NRP). This adaptation enables the life cycle of the Mycobacterium species to reset to the lag phase when a favourable atmosphere and nutrients are encountered.
Bacterial life within a closed culture system is archetypal and most mycobacteria can survive for long periods by adopting reversible quiescent metabolic states that reflect the external environment. There is little known about this aspect of the mycobacterial life cycle, which may involve NRP and the loss of the cell wall. NRP may be a result of the pressures of antimicrobial therapy, adverse environmental conditions or lack of nutrients and oxygen. NRP can be reversed in vivo when favourable conditions for growth eventuate [4].
CWDM represent part of the life cycle of Mycobacterium species. In the latent phase of mycobacterial infection, the microorganism may cease to grow, and enter a cell wall deficient state if nutrients are unavailable and atmospheric conditions are suboptimal. There have been few studies on the fundamental processes that drive mycobacterial latency. In-vitro observations indicate that the CWDM state occurs naturally in old fluid cultures of mycobacteria, and can be artificially induced through chemical stripping of the cell wall, or by long-term incubation of mycobacteria in low nutrient media. [5,6,7].
Development of growth media for previously uncultured organisms [8] is very difficult, and requires patience and innovations [9]. The detection and cultivation of cell wall deficient mycobacteria (CWDM) in blood and tissues is possible using both old and newly developed techniques. [9,10] The ZN stain can be used for monitoring the growth of CWDM in culture [11,12] if alterations to the ZN stain method are made.
Mycobacterial colonisation of blood has been reported in inflammatory bowel disease and in healthy controls using molecular detection methods [13,14,15], including PCR. However, visual confirmation of the existence of CWDM has mostly been lacking, with the exception of electron microscopy of CWDM isolates cultured from tissue collected from patients with Crohn’s disease [16].
Turbidity measurements using mycobacterial culture media such as Middlebrooks 7H9 broth are unsuitable for short term growth monitoring of CWDM, as they contain precipitants. In the absence of methods to detect CWDM, PCR is used as a surrogate test to detect mycobacteria in specimens from patients with IBD, in place of culture. PCR does not differentiate between viable and non-viable mycobacteria, without modifications to the PCR protocols. [17,18] PCR has been used to detect mycobacterial DNA in patients with CD disease as well as in healthy human controls [19]. Some other inflammatory diseases may also involve CWDM in stealth form, or as “non-replicating persister” (NRP) forms [20,21].
The cultivation of CWDM forms from human hosts by the authors has proven to be extremely difficult, but is achievable. In lieu of viable isolates of CWDM from human sources, other researchers have instead simulated CWDM though the chemical deconstruction of acid-fast bacilli [22,23] (AFB). Artificially manufactured CWDM differ from those found in nature, as others have observed [24]. The cell wall deficient state may be a defence against bacteriophage attacks. [25]
MTBC are ontogenetic [26,27], and this trait may be inherited by CWDM. Naturally occurring CWDM produce biofilm, and their inner membranes have been exposed to host antibodies targeting mycobacterial virulence factors [27]. Formation of antigen-antibody complexes [28] will also alter the exposed mycobacterial bacterial inner membrane.
Mycobacterium species are a diverse group of organisms that range from non-pathogenic, to opportunistically pathogenic, to obligately pathogenic. Obligate pathogens such as MTBC are invariably pathogenic in humans. Members of Mycobacterium avium complex (MAC) are opportunistic pathogens, and may cause disease in immunocompromised hosts but are able to transiently colonise healthy subjects without causing diseases.
Non-tuberculous mycobacteria (NTM) are not routinely isolated in clinical laboratories, neither in the bacillary form, nor as CWDM. Partly because they are not actively sought, and partly because they are extremely difficult to isolate using common culture methods. Although they are ubiquitous in the environment, they are seldom seen as laboratory contaminants. There are a wide range of conditions for optimal growth of different members of the Mycobacterium genus, meaning that only a narrow range of NTM are detected in the laboratory. Diagnosis of tuberculosis is a high priority, given its global disease burden, so laboratory methods are optimised to detect MTBC acid fast bacilli(AFB) [29,30]. These methods are slightly different between the two groups.
The clinical significance of a single isolation of NTM is difficult to establish unless it is an obligate pathogen. Isolates of MAC usually require serial isolations from culture to confirm a pathogenic role, because of widespread presence in the environment, including the water supply of domestic homes [31] and hospitals. Hence semi-quantitation is used to differentiate transient acquisition from opportunistic infection. Enumeration of organisms present in samples may suggest a pathogenic role in a subject, as higher numbers of organisms may relate to infection, rather than carriage [32]. The ZN stain of a sample may indicate there are high numbers of acid-fast organisms (AFO) present [32]. Growth on solid media enables numbers of organisms to be estimated.
Quantitation of bacteria is also a useful measure of therapeutic response, both in vitro through antimycobacterial susceptibility testing and in vivo, to monitor a reduction of the number of organisms as well as the viability of the cells over time. In the healthy subject, low numbers of NTM may be present without consequences. This may present difficulties in interpretation, unless transient carriage can be differentiated from infection, through specimen monitoring and clinical evaluation. If culture of CWDM is available then the PCR result will be complementary and confirmatory.
Visualising CWDM in Culture
Since the discovery of the ZN stain in 1883, the stain methodology has undergone modifications to optimise the laboratory detection of MTBC.
Laboratory staining methods are often selected that are convenient, cost effective and technically expedient. The acid-alcohol decolouriser used in most laboratories selects for MTBC and is preferred because it is a “clean” stain and results in a slide without excess carbol-fuchsin (CF) residue. In turn this results in unambiguous reporting of acid-alcohol fast bacilli, strongly indicative of tuberculosis.
The formula of the CF stain used in the ZN stain to visualise CWDM is integral to detection. (Table 1) Concentrations of basic fuchsin vary greatly in recipes for CF. Laboratories making their own CF often use 1% basic fuchsin. This may be sub-optimal for CWDM. The authors recommend the following recipe for the detection of CWDM.
2. Methods
2.1. Ziehl-Neelsen Stain for CWDM
Step 1. Carbol fuchsin stain: (1). Dissolve basic fuchsin in the alcohol and add the water slowly while shaking. (2). Add Phenol. Filter.

Table 1.
Formula for preparing Carbol Fuchsin Stain.
| Basic Fuchsin | 24 gm (CARL ROTH GmbH &Co (C.I 42510) |
| Ethanol 95% | 120 ml |
| Distilled Water | 800 ml |
| Phenol | 48 ml |
Step 2. ZN decolouriser: 20% H2SO4 in distilled water.
Step 3. Counterstain: Loeffler’s methylene blue; 10ml glacial acetic acid; 1 litre Distilled Water; 2.5gm methylene blue; Filter.
2.2. Method for Staining
1. Flood the slide with carbol fuchsin. Gentle heat may be applied to the slide. Leave for 8-10 minutes.
2. Rinse off the carbol fuchsin with water.
3.Decolourise for two minutes using the acid decolouriser with a wash between. i.e., 1 minute of the decolouriser, wash, add more decolouriser, then wash off again. Alternatively, the decolouriser can be applied until clearance of carbol fuchsin can be seen.
4. Cover slide with methylene blue counterstain for 2 minutes; wash off, dry the slide without blotting.
2.3. Microscope Slides
Positively-charged slides enable the retention of blood-containing culture material on the slide during the staining process.
2.4. Sample Collection
Blood samples should be collected in tubes containing sodium citrate as the anticoagulant. EDTA should not be used as the anticoagulant as it is bactericidal [33] and inhibits biofilm formation.
2.5. Detection of CWDM
To confirm methodology and detection of CWDM, the following experiment can be performed using the BD BACTEC™ MGIT™ mycobacterial growth indicator tube (MGIT).
- (1)
- To a MGIT tube supplemented with OADC and PANTA, add enough sterile filtered sucrose solution in water to arrive at a final concentration in the MGIT of 1% sucrose. If MAC is suspected, add mycobactin J. (Allied Monitor Inc., Fayette, MO) according to manufacturer’s instructions.
- (2)
- The buffy coat from a human subject is harvested after centrifugation of the sodium citrated blood sample for 10 minutes at 3000 RPM. Some consideration should be given to separate culture of the erythrocyte component [34].
- (3)
- Inoculate the MGIT tube with buffy coat and incubate for 30 days at 370c, with a primary reading of the culture deposit at 8 days, using the above ZN stain method.
Note: The MGIT tube is optimised for the culture of MTBC in the AFB form, not for CWDM culture.
3. Results
Using additional supplementation and the above ZN stain it is possible to semi-quantitate CWDM to obtain an approximation of numbers of CWDM present in a sample after 30 days incubation. During incubation of the buffy coat in the MGIT tube, the CWDM undergo initial rapid growth, followed by NRP. (Personal data) Nucleases may also be produced extracellularly in the initial growth phase [35,36].
CWDM are readily visible under x 1000 oil immersion as ZN positive spheres. Sizes of CWDM vary from very small to larger than a white blood cell. (WBC) Figure 1, Figure 2 and Figure 3.
Nearly all healthy subjects carry small numbers of intracellular CWDM, possibly to aid immune recognition of mycobacteria (personal data). This has been termed ‘trained immunity. Some IBD patients carry large numbers of CWDM in peripheral blood that can be cultivated using more extensive protocols than are covered in this preliminary report.
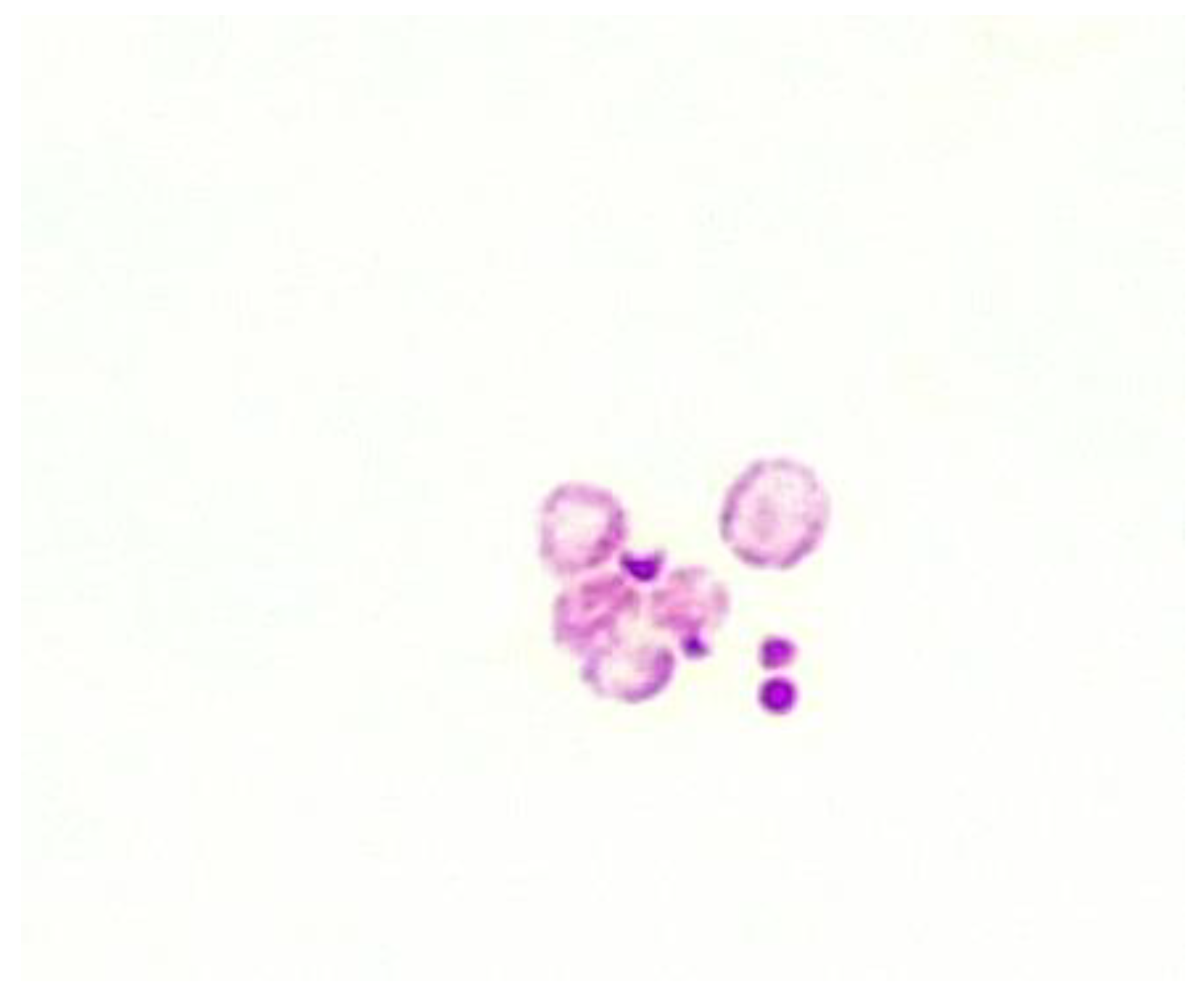
Figure 3.
ZN stain Supplemented MGIT tube 370c incubation for 30 days. Patient diagnosed with refractory Crohn’s disease. Note the variation in size and some evidence of bilateral division (x1000 magnification under oil immersion).
Figure 3.
ZN stain Supplemented MGIT tube 370c incubation for 30 days. Patient diagnosed with refractory Crohn’s disease. Note the variation in size and some evidence of bilateral division (x1000 magnification under oil immersion).
3.1. Fluid Culture Media for CWDM
As we found with the ZN stain, there is no single culture medium that can be considered standard for cultivation of CWDM.
Bacterial culture is necessary for confirmation of the ZN stain result and for the subsequent antimycobacterial susceptibility tests and molecular studies. Other researchers have shown little difference in the performance of supplemented 7H9 medium and MGIT tubes, when used in non-automated manual procedures [37,38]. In-house fluid media has advantages as an experimental vehicle in the development of CWDM culture media, including the ability to supplement the media with specific nutrients, and monitoring for variations in replication times of isolates in response to additives. The authors believe that because of the numerous variables, (nutrients, osmolarity, atmosphere etc.) that in-house bespoke fluid media is cost-effective and sufficiently flexible for further development of media for CWDM, until such time that consensus is reached and commercially manufactured CWDM is available.
CWDM are incapable of growth in 7H9 base medium except when that medium has additional supplements. For the practical purposes of media development, 7H9 medium can be used as a base medium, provided supplementation (i.e. OADC, amino acids, vitamins etc) is possible. Cost can also be a factor in innovation however. Development of media for CWDM requires a large degree of trial and error.
Experiments using 7H9 fluid media as the base are necessary to identify supplementary constituents that may either stimulate or inhibit growth of CWDM. We have observed that CWDM cultivation media must be able to compensate for the effect of nutrient flux on the CWDM. (Titration of media may also be helpful to establish performance limits.)
The rudimentary membrane structure of CWDM may render them susceptible to osmotic and atmospheric changes. The adjustments in constituents of fluid media that must be undertaken to arrive at a “goldilocks zone” to support ongoing growth are further complicated by apparent strain diversity, nutrient requirements, and inhibitory factors. The structure of CWDM is often fragile and isolates may progress through a series of growth changes depending on the strain of CWDM and the duration of incubation. This effort is however, necessary to be able to cultivate the CWDM for susceptibility studies and molecular analysis [9].
Middlebrooks 7H9 medium is a low nutrient, defined medium, and is widely used for isolation of MTBC from a variety of sample types. The use of low nutrient mycobacterial culture medium is based on the restricted availability of nutrients for MTBC in vivo in the host [39]. In vivo the nutrient requirements of MTBC may differ depending on the stage of the disease, and this also applies to CWDM in general.
NTM may require more complex nutrients, amino acids and vitamins (i.e. mycobactin J) to grow in vitro. The effects of nutrient flux may impact more on CWDM, which have only the inner semipermeable membrane to modulate intracellular nutrient availability. Resuscitation promotion factors (RPF) may improve growth considerably. RPF have been associated with the reversion of the CWDM to bacillary forms and may have a role in acceleration or inhibition of growth [40].
3.2. Solid Culture Media for CWDM
Solid culture media such as Lowenstein Jensen (LJ), Middlebrook7H10, and Middlebrook 7H11 are commonly used for isolation of mycobacteria, with a global preference for LJ medium, because of ease of manufacture and cost effectiveness. There is no general agreement on a single solid medium [41]. The selection of media is a mixture of prior experience, personal whims and established laboratory procedures. Supplements can be a major expense for some laboratories, particularly as the revival and growth of CWDM is still somewhat stochastic. Fluid media usually promote mycobacterial growth better than solid media, but solid media is more useful for quantitation. PCR is a poor substitute for growth on solid media. Sheep blood agar (SBA) has been used for cultivation of MTBC on solid agar where it was found to perform better than other standard media, such as Lowenstein Jensen media (LJ) [42,43].
Mycobacterium avium grew better on SBA than on LJ when colonies were counted. SBA may be a higher nutrient medium than LJ. It may also be possible that addition of egg yolk to LJ inhibits growth of CWDM. This suggests that more concentrated, or more specifically formulated nutrient media can be used to advantage. Markova et al reported isolation of CWDM on semi solid agar medium [44]. Other cell wall deficient bacteria can be grown on conventional laboratory semi-solid and solid media [45,46,47].
The authors have not yet had unalloyed success with solid media to grow CWDM, but biphasic media may hold some promise.
4. Discussion
There are over 190 members of the Mycobacterium genus. Members of MAC, which proportionally represent a small number of the total, exhibit significant variances in growth requirements. CWDM similarly show diverse growth requirements in fluid supplemented 7H9 media. There have been few reports of the detection of CWDM in the literature. Coccoid forms of mycobacteria have been reported and are associated with environmental stress [5]. There have been numerous reports supporting or rejecting microbial involvement of CWDM in chronic inflammatory illnesses. Few of these reports have centred on the microbial attributes of CWDM to establish and drive inflammation. This can only be achieved when the pathogen is able to be studied.
Visualisation of CWDM in blood samples from humans is possible using the MGIT tube. Small changes in the Ziehl-Neelsen staining procedure are necessary to allow for differences of NTM. The MGIT tube is useful for initial isolation, but not for replication over a longer period. Extended incubation in CWDM growth media is necessary to collect sufficient DNA from CWDM for whole genome sequencing and prospective employment of MALTI-TOF for identification of isolates. Additionally, susceptibility testing requires that isolates are in the log phase of growth.
Microscopic detection is necessary before growth media for CWDM can be designed and assessed for performance. The ability to cultivate CWDM in broth media means that more specific studies on the molecular genome of isolates may be undertaken, and specific antimycobacterial therapies designed. These steps are important to better understand the involvement (if any) of CWDM in chronic disease states and in latent tuberculosis.
Antimicrobial combination therapy is presently required to be administered for extended periods, from months to years, without supporting laboratory susceptibility data. It is feasible, but untested, that once therapy is removed, the CWDM will reset to a log phase of growth. Future treatments may also require biofilm disrupters, repurposed antimicrobial compounds, and modern vaccine therapy to maintain remission and possibly achieve cures. The possibility that MAP and CD are intimately associated, raises the almost fantastic idea of the defeat of CD through the One Health concept of infection control, targeted at what is possibly a zoonotic disease [48].
Author Contributions
JA Conceptualization, methodology, validation, formal analysis, investigation, resources, data curation, writing—original draft preparation, visualization, supervision, project administration, DA formal analysis, investigation, resources, visualization, All authors have read and agreed to the published version of the manuscript. GA Conceptualization, validation, formal analysis, investigation, data curation, writing—original draft preparation, visualization, supervision, project administration,.
Funding
This research received no external funding.
Institutional Review Board Statement
Not relevant for this study.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Not applicable.
Conflicts of Interest
JA has filed patents in the field of staining and growth of MAP/ mycobacteria. DA and GA have no conflict of interest.
References
- P. T. Elkington, “TUBERCULOSIS: TIME FOR A NEW PERSPECTIVE?,” J Infect, vol. 66, no. 4, pp. 299–302, Apr. 2013. [CrossRef]
- J. D. Ernst, “The immunological life cycle of tuberculosis,” Nat Rev Immunol, vol. 12, no. 8, Art. no. 8, Aug. 2012. [CrossRef]
- S. W. Park et al., “Growth of Mycobacteria on Carbon Monoxide and Methanol,” J Bacteriol, vol. 185, no. 1, pp. 142–147, Jan. 2003. [CrossRef]
- 4. S. Ratnam and S. Chandrasekhar, “The pathogenicity of spheroplasts of Mycobacterium tuberculosis,” American Review of Respiratory Disease, vol. 114, no. 3, pp. 549–554, 1976.
- A. Velayati et al., “Populations of latent Mycobacterium tuberculosis lack a cell wall: Isolation, visualization, and whole-genome characterization,” International Journal of Mycobacteriology, vol. 5, no. 1, pp. 66–73, Mar. 2016. [CrossRef]
- W. R. Burnham, J. E. W. R. Burnham, J. E. Lennard-Jones, J. L. Stanford, and R. G. Bird, “MYCOBACTERIA AS A POSSIBLE CAUSE OF INFLAMMATORY BOWEL DISEASE,” The Lancet, vol. 312, no. 8092, pp. 693–696, Sep. 1978. [CrossRef]
- G. Gitnick et al., “Preliminary report on isolation of mycobacteria from patients with Crohn’s disease,” Digest Dis Sci, vol. 34, no. 6, pp. 925–932, Jun. 1989. [CrossRef]
- D. Raoult et al., “Cultivation of the Bacillus of Whipple’s Disease,” New England Journal of Medicine, vol. 342, no. 9, pp. 620–625, Mar. 2000. [CrossRef]
- J.-C. Lagier, S. Edouard, I. Pagnier, O. Mediannikov, M. Drancourt, and D. Raoult, “Current and past strategies for bacterial culture in clinical microbiology,” Clin Microbiol Rev, vol. 28, no. 1, pp. 208–236, Jan. 2015. 2015. [CrossRef]
- J.-C. Lagier, P. Hugon, S. Khelaifia, P.-E. Fournier, B. La Scola, and D. Raoult, “The rebirth of culture in microbiology through the example of culturomics to study human gut microbiota,” Clin Microbiol Rev, vol. 28, no. 1, pp. 237–264, Jan. 2015. [CrossRef]
- R. B. Gearry, J. M. Aitken, R. L. Roberts, S. Ismail, J. Keenan, and M. L. Barclay, “Images of interest. Gastrointestinal: Mycobacterium avium paratuberculosis and Crohn’s disease,” J Gastroenterol Hepatol, vol. 20, no. 12, p. 1943, Dec. 2005. [CrossRef]
- J. M. Aitken, T. J. Borody, and G. Agrawal, “A revaluation of the use of conventional Ziehl-Neelsen stain for detection of non-tuberculous mycobacteria,” New Zealand Journal of Medical Laboratory Science, vol. 73, no. 3, Art. no. 3, 2019.
- J. T. Kuenstner et al., “Presence of Infection by Mycobacterium avium subsp. paratuberculosis in the Blood of Patients with Crohn’s Disease and Control Subjects Shown by Multiple Laboratory Culture and Antibody Methods,” Microorganisms, vol. 8, no. 12, Art. no. 12, Dec. 2020. [CrossRef]
- H. M. Fidler, W. Thurrell, N. M. Johnson, G. A. Rook, and J. J. McFadden, “Specific detection of Mycobacterium paratuberculosis DNA associated with granulomatous tissue in Crohn’s disease,” Gut, vol. 35, no. 4, pp. 506–510, Apr. 1994. [CrossRef]
- S. A. Naser, S. R. Sagramsingh, A. S. Naser, and S. Thanigachalam, “Mycobacterium avium subspecies paratuberculosis causes Crohn’s disease in some inflammatory bowel disease patients,” World J Gastroenterol, vol. 20, no. 23, pp. 7403–7415, Jun. 2014. [CrossRef]
- R. J. Chiodini, H. J. Van Kruiningen, W. R. Thayer, and J. A. Coutu, “Spheroplastic phase of mycobacteria isolated from patients with Crohn’s disease.,” Journal of Clinical Microbiology, vol. 24, no. 3, pp. 357–363, 1986.
- K. L. Josephson, C. P. Gerba, and I. L. Pepper, “Polymerase chain reaction detection of nonviable bacterial pathogens,” Appl Environ Microbiol, vol. 59, no. 10, pp. 3513–3515, Oct. 1993. [CrossRef]
- H. Kobayashi, M. Oethinger, M. J. Tuohy, G. W. Procop, G. S. Hall, and T. W. Bauer, “Limiting false-positive polymerase chain reaction results: detection of DNA and mRNA to differentiate viable from dead bacteria,” Diagnostic Microbiology and Infectious Disease, vol. 64, no. 4, pp. 445–447, Aug. 2009. [CrossRef]
- R. A. Juste et al., “Association between Mycobacterium avium subsp. paratuberculosis DNA in blood and cellular and humoral immune response in inflammatory bowel disease patients and controls,” International Journal of Infectious Diseases, vol. 13, no. 2, pp. 247–254, Mar. 2009. [CrossRef]
- P. L. Almenoff, A. Johnson, M. Lesser, and L. H. Mattman, “Growth of acid fast L forms from the blood of patients with sarcoidosis.,” Thorax, vol. 51, no. 5, pp. 530–533, 1996.
- S. T. Brown et al., “Recovery of cell wall-deficient organisms from blood does not distinguish between patients with sarcoidosis and control subjects,” Chest, vol. 123, no. 2, pp. 413–417, Feb. 2003. [CrossRef]
- T. Udou, M. Ogawa, and Y. Mizuguchi, “Spheroplast formation of Mycobacterium smegmatis and morphological aspects of their reversion to the bacillary form,” J Bacteriol, vol. 151, no. 2, pp. 1035–1039, Aug. 1982. [CrossRef]
- T. Udou, M. Ogawa, and Y. Mizuguchi, “An improved method for the preparation of mycobacterial spheroplasts and the mechanism involved in the reversion to bacillary form: electron microscopic and physiological study,” Can. J. Microbiol., vol. 29, no. 1, pp. 60–68, Jan. 1983. [CrossRef]
- L. H. Mattman, Cell wall deficient forms: stealth pathogens. Boca Raton: CRC Press, 2001.
- J. C. Wohlfarth et al., “L-form conversion in Gram-positive bacteria enables escape from phage infection,” Nat Microbiol, pp. 1–13, Jan. 2023. [CrossRef]
- S. Gagneux, “Host–pathogen coevolution in human tuberculosis,” Philos Trans R Soc Lond B Biol Sci, vol. 367, no. 1590, pp. 850–859, Mar. 2012. [CrossRef]
- J. W. Saelens, G. Viswanathan, and D. M. Tobin, “Mycobacterial Evolution Intersects With Host Tolerance,” Front Immunol, vol. 10, p. 528, Mar. 2019. [CrossRef]
- L. L. Lu, T. J. Suscovich, S. M. Fortune, and G. Alter, “Beyond binding: antibody effector functions in infectious diseases,” Nat Rev Immunol, vol. 18, no. 1, pp. 46–61, Jan. 2018. [CrossRef]
- G. G. Meynell and E. Meynell, “Theory and Practice in Experimental Bacteriology.,” Theory and Practice in Experimental Bacteriology., 1965. 1965. Available online: https://www.cabdirect.org/cabdirect/abstract/19652704624 (accessed on 19 June 2023).
- G. S. Wilson and M. A. Topley, “Wilson’s Principles of bacteriology, virology and immunity,” London: Edward Arnold, pp. 1855–1863, 1975.
- L. Lande et al., “Mycobacterium avium in Community and Household Water, Suburban Philadelphia, Pennsylvania, USA, 2010–2012,” Emerg Infect Dis, vol. 25, no. 3, pp. 473–481, Mar. 2019. [CrossRef]
- J. Aitken, G. Agrawal, N. Markova, G. Slavchev, W. Chamberlin, and T. J. Borody, “Demonstration of Intracellular Mycobacterium Species in Crohn’s Disease Using Novel Technologies: 1990,” Official journal of the American College of Gastroenterology | ACG, vol. 110, p. S843, Oct. 2015.
- S. Finnegan and S. L. Percival, “EDTA: An Antimicrobial and Antibiofilm Agent for Use in Wound Care,” Adv Wound Care (New Rochelle), vol. 4, no. 7, pp. 415–421, Jul. 2015. 2015. [CrossRef]
- Y. Nishiuchi et al., “Direct Attachment with Erythrocytes Augments Extracellular Growth of Pathogenic Mycobacteria,” Microbiol Spectr, vol. 10, no. 2, p. e0245421, Apr. 2022. [CrossRef]
- E. Mankiewicz and M. G. Tamari, “Lysogeny and Deoxyribonuclease Production in Mycobacteria,” Am Rev Respir Dis, vol. 106, no. 4, pp. 609–610, Oct. 1972. [CrossRef]
- Xinxin Zang et al., “Extracellular DNase MAP3916c attacks the neutrophil extracellular traps and is needed for Mycobacterium avium subsp. paratuberculosis virulence,” Veterinary microbiology, vol. 273, pp. 109529-, 2022. [CrossRef]
- T. Battaglioli, A. Soto, J. Agapito, V. Acurio, and P. Van der Stuyft, “Manual liquid culture on simple Middlebrook 7H9 or MGIT for the diagnosis of smear-negative pulmonary tuberculosis,” Trop Med Int Health, vol. 19, no. 12, pp. 1500–1503, Dec. 2014. [CrossRef]
- J. E. Meneguello et al., “Fast detection of Mycobacterium tuberculosis in culture-positive sputum samples by nitrate reductase activity,” Brazilian Journal of Pharmaceutical Sciences, vol. 54, no. 1, Art. no. 1, Jun. 2018. [CrossRef]
- M. BERNEY and L. BERNEY-MEYER, “Mycobacterium tuberculosis in the Face of Host-Imposed Nutrient Limitation,” Microbiol Spectr, vol. 5, no. 3, p. 10.1128/microbiolspec.TBTB2-0030–2016, Jun. 2017. [CrossRef]
- G. V. Mukamolova, O. Turapov, J. Malkin, G. Woltmann, and M. R. Barer, “Resuscitation-promoting factors reveal an occult population of tubercle Bacilli in Sputum,” Am J Respir Crit Care Med, vol. 181, no. 2, pp. 174–180, Jan. 2010. [CrossRef]
- M. L. Joloba et al., “What is the most reliable solid culture medium for tuberculosis treatment trials?,” Tuberculosis, vol. 94, no. 3, pp. 311–316, May 2014. 20 May. [CrossRef]
- M. Drancourt, P. Carrieri, M.-J. Gévaudan, and D. Raoult, “Blood Agar and Mycobacterium tuberculosis: the End of a Dogma,” J Clin Microbiol, vol. 41, no. 4, pp. 1710–1711, Apr. 2003,. [CrossRef]
- M. Drancourt and D. Raoult, “Cost-Effectiveness of Blood Agar for Isolation of Mycobacteria,” PLOS Neglected Tropical Diseases, vol. 1, no. 2, p. e83, Nov. 2007. [CrossRef]
- N. Markova, G. Slavchev, and L. Michailova, “Filterable forms and L-forms of Mycobacterium bovis BCG,” Human Vaccines & Immunotherapeutics, vol. 8, no. 6, pp. 759–764, Jun. 2012. [CrossRef]
- Petrovic Fabijan et al., “L-Form Switching in Escherichia coli as a Common β-Lactam Resistance Mechanism,” Microbiology Spectrum, vol. 10, no. 5, pp. e02419-22, Sep. 2022. [CrossRef]
- D. Wolf, P. Domínguez-Cuevas, R. A. Daniel, and T. Mascher, “Cell Envelope Stress Response in Cell Wall-Deficient L-Forms of Bacillus subtilis,” Antimicrob Agents Chemother, vol. 56, no. 11, pp. 5907–5915, Nov. 2012. [CrossRef]
- J. Errington, “Cell wall-deficient, L-form bacteria in the 21st century: a personal perspective,” Biochem Soc Trans, vol. 45, no. 2, pp. 287–295, Apr. 2017. [CrossRef]
- “One Health Zoonotic Disease Prioritization (OHZDP) | One Health | CDC,” Dec. 12, 2022. Available online: https://www.cdc.gov/onehealth/what-we-do/zoonotic-disease-prioritization/index.html (accessed on 19 June 2023).
Figure 1.
ZN stain. Blue object is a WBC. The large red object is a CWDM (x1000 magnification under oil immersion).
Figure 1.
ZN stain. Blue object is a WBC. The large red object is a CWDM (x1000 magnification under oil immersion).
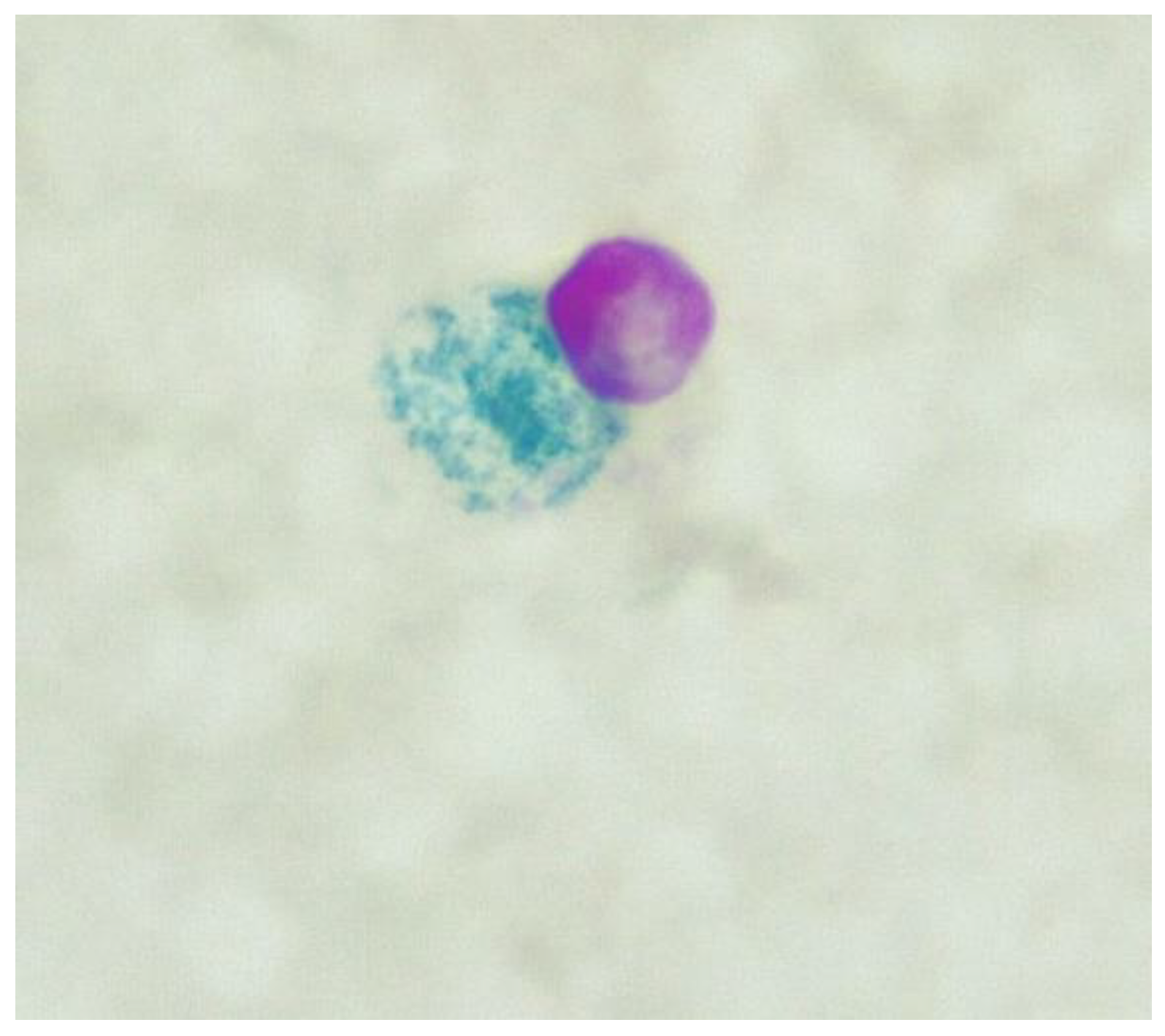
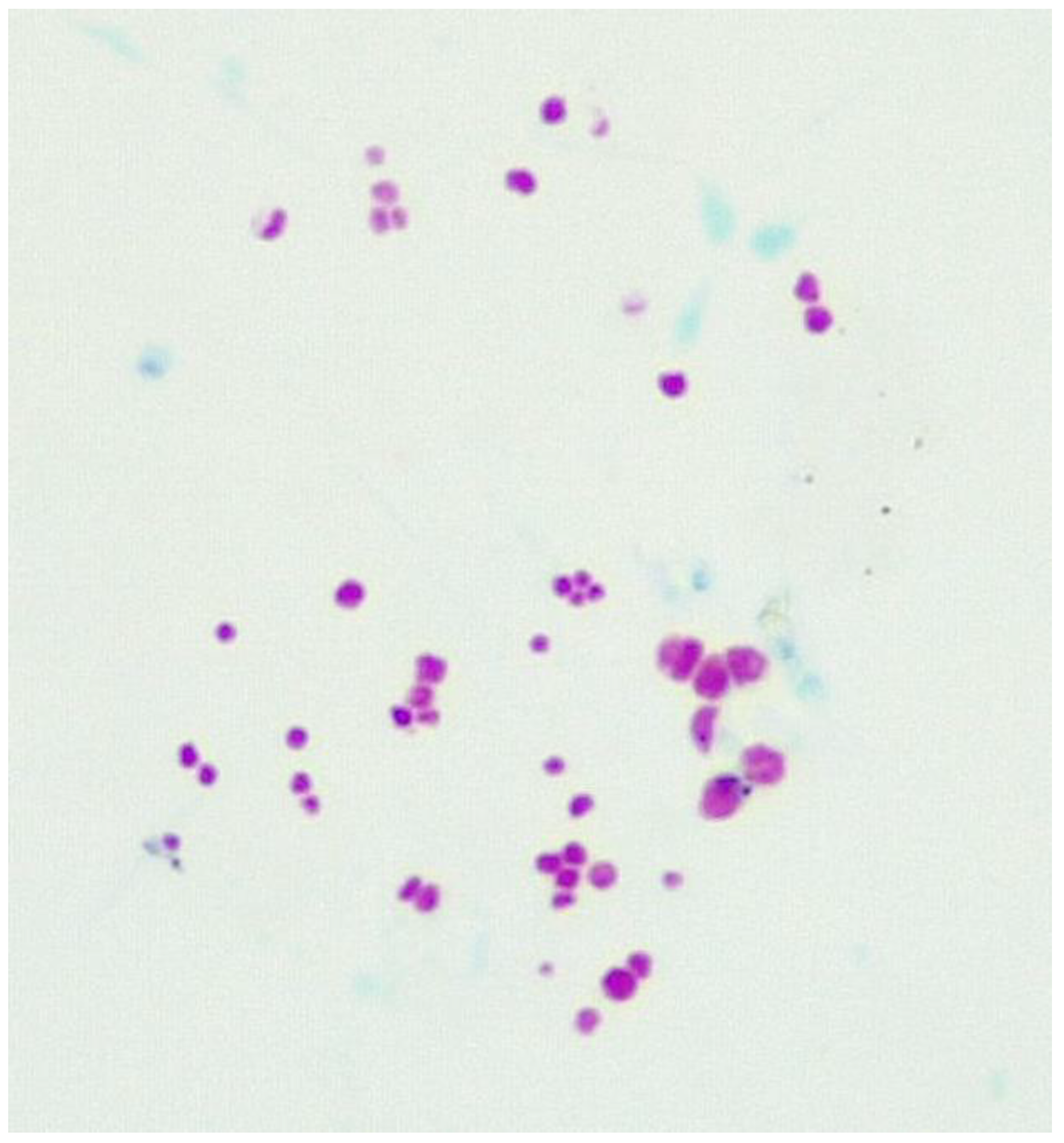
Figure 2.
ZN stain. Blood culture from a subject with CD showing pleomorphic forms (x1000 magnification under oil immersion).
Figure 2.
ZN stain. Blood culture from a subject with CD showing pleomorphic forms (x1000 magnification under oil immersion).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



