
Submitted:
14 July 2023
Posted:
17 July 2023
You are already at the latest version
Abstract
Drought stress has been considered a major problem in coffee crops, and the higher occurrence of unpredictable environmental events has highlighted the vulnerability of agricultural systems. Several reports have indicated selenium (Se) as a plant biostimulant against environmental stress, including drought. The effects of Se in plants are related to the modulation of enzymatic activity, energetics and osmoprotectants plant content, and nutrient balance in plants. However, investi-gating the effective time of Se application is crucial for better understanding its effects on coffee plants, a drought-sensitive crop. This study aimed to investigate the role of Se supply in improving drought tolerance in coffee seedlings while also evaluating the best timing for Se application. Five times of Se foliar application were assessed during induced drought stress by PEG-6000 using the day of imposing stress as a default, plus two control treatments: i) with osmotic stress and without Se; ii) without osmotic stress and Se. The treatments were compared with both control treatments. Results demonstrated that the osmotic stress (OS) promoted mild stress in the coffee plants, as ex-pected. Plants under stress showed reduced GR and SOD activity compared with the non-stressed ones, and the OS induces starch degradation, which is potentialized by Se foliar supply, mainly when Se is applied before inducing the stress. The decline in the starch content might be considered a strategy to provide energy to coffee seedlings. The seedlings that received foliar Se application 8 days before the stress exhibited higher CAT, APX, and SOD than those that did not receive OS nor Se application. The results obtained in this study suggest that the previous Se foliar spray is more effective than the Se supply after OS to overcome the adverse condition. On the other hand, the post-stress application seems to impose extra stress on the plants, leading them to reduce the water potential.
Keywords:
Beneficial Elements
; Tropical Agriculture
; Coffee Belt.
1. Introduction
Atmospheric carbon dioxide (CO2) has increased over seven decades. It is correlated with gradual and systematic modifications in average climate conditions, such as temperature and precipitation variance [1]. Indeed, such extreme events (e.g., heat waves, floods, and severe drought seasons) expose the remarkable vulnerability of agricultural systems [2,3].
These environmental changes have modified temperature and rain patterns worldwide, making coffee cultivation uncertain in commonly cultivated areas [4,5]. Coffee is a crop sensitive to precipitation variability, and rainfall instability can lead to high losses in coffee production. Arabica coffee requires between 1,000 and 2,700 mm of annual precipitation and from one to three months of dry season annually [6]. Due to its temperature and humidity demands, coffee cultivation is limited to the intertropical region, commonly called The Coffee Belt [7].
The plant side effects of the lack of water in the crop system are drought stress [8]. Drought stress imposes an osmotic stress (OS) due to the lack of water in the plant tissue. The OS promoted changes in plants’ physiological, morphological, ecological, biochemical, and molecular traits [9,10]. Water deficit directly affects crops’ growth, development, and yield [11]. As an immediate response to drought, the stomatal closes, which constrains the transpiration flow and the CO2 fixation. These responses vigorously reduce the photosynthetic rates and hence the production of photoassimilates [12]. The impact of drought on coffee plants reflects negatively in the harvest in progress and future ones [13].
Plant mineral nutrition is considered a strategy to reduce the adverse effects of drought stress. Selenium (Se) is one of the promising approaches to fight the metabolic responses in plants under this type of adverse condition [14,15,16]. Selenium is not a plant nutrient, but several studies have reported its beneficial effects, mainly in stress conditions (e.g., salinity, chilling stress, metals accumulation, and drought stress) [11,17,18,19]. Sousa et al., [19] found that Se can modulate nutrient uptake, carbohydrate breakdown, and enzymatic activity in coffee plants after low-temperature stress, helping the plants to overcome adverse conditions. Meanwhile, Luo et al., [20] showed Se increased photosynthetic parameters during drought stress in rice. Also, the same authors found that Se can promote a higher transcript level of antioxidant-related genes.
Studies determining the most effective time to apply this beneficial element for achieving drought stress mitigation are still lacking in the literature. Yet, plant supplementation using Se before the stress has been responsible for triggering metabolic responses in plants – priming effect [21]. Priming effects were first used to describe the application technique of nutrient and/or plant biostimulants in seeds, to increase the vigor during germination [22]. However, applying biostimulants, such as Se, has been considered a resistance inducer strategy in plants and can be thought as a promising strategy for crop production in response to future climate changes [23,24,25].
In this paper, the foliar application of such biostimulant element is also called “priming” due to the preparation effect that it can promote in the plants and its implication on metabolic responses before the stress [26]. However, the exogenous Se applied post-stress can also be used as a last resource to alleviate the side effects of drought in plants, but the effects of Se on these conditions need to be clarified. Thus, this study aimed to investigate the role of Se supply in improving drought tolerance in coffee seedlings while also assessing the best time for Se application.
2. Results
2.1. Analysis of Se content
Selenium content in leaves was significantly increased by foliar application. There was a statistical difference between all the treatments with Se application and the controls - without Se supply (Figure 1). The Se content in the control treatments was 0.37 and 0.38 mg kg DW-1 for the treatments with stressed (+OS-Se) and non-stressed plants (-OS-Se), respectively. Meanwhile, the average Se content in the remaining treatments’ leaves was 1.95 mg kg DW-1.
2.2. H2O2, MDA, and antioxidant enzymes (SOD, CAT, APX, GR)
There was no marked trend of OS on H2O2 and MDA, even when the control with stressed treatment was compared with the non-stressed one (Figure 2). In the H2O2 essays, even if the OS is considered one of the main triggering agents of reactive oxygen species (ROS), there was no statistical difference between the treatments. On the other hand, the treatments with Se application -4BOS and +4AOS promoted higher values of its content, indicating that these treatments induced lipid peroxidation in the leaves.
When the control treatments were compared, the OS significantly reduced the activity of GR and SOD, but the OS did not affect the activities of APX and CAT (Figure 2), i.e., there was no difference between +OS-Se and -OS-Se.
The Se application 8 days before the plants were submitted to OS (-8BOS) promoted higher APX, CAT, and SOD activity than the treatment +OS-Se. The Se application at -8BOS increased the activity of these enzymes in the order of 356.3, 228.5, 771.1, and 266.5% compared with +OS-Se, respectively, for APX, CAT, and SOD. A reduction in the GR was noticed in the treatments 0OS; +4AOS; +8AOS; and +OS-Se compared with the treatment -OS-Se. Plants previously treated with Se (-8 and -4BOS) had higher GR content than those of the +OS-Se treatment and a similar Se content when compared with non-stressed plants (-OS-Se) (Figure 2).
2.3. Carbohydrates, Protein, Amino Acids, Proline
Regardless of the Se supply and OS, the total free amino acids, reducing sugars, and sucrose content were unaffected (Figure 3). On the other hand, Se foliar supply increased, to some extent, the proline and protein content. The proline content obtained with the application of Se at -4BOS and +8AOS was significantly higher than that observed for stressed plants without Se (+OS-Se). Hence, the Se supplementation might be seen as a strategy to increase these compounds in coffee leaves under OS.
On the other hand, the imposed OS affected the starch content, with all the treatments submitted to the stress showing lower starch content compared with the absolute control treatment (-OS-Se). However, all the treatments with Se application promoted lower starch content than the positive control treatment (+OS-Se), except the treatment with the application on the day on which the stress was imposed (0OS) (Figure 3). Such results indicate that the OS can reduce the starch content, but the Se supply can impose lower starch content than that detected in plants submitted to OS without Se.
2.4. Chlorophyll Fluorescence parameters (MultispeQ®)
The graph of chlorophyll was inserted as Supplementary Data (Supplementary data, Figure 2). There was no statistical difference in the ECSt, Phi2, PhiNO, PhiNPQ, and qL. Selenium supply on the same day the plants were submitted to the OS (0OS) and 8 days after the plants were submitted to the OS increased the LEF – linear electron flux – compared with the treatment without Se supply and OS (Supplementary data, Figure 2). The Se application at +4AOS promoted the highest NPQt, showing that, in a certain way, Se can act to quench the excess of light energy.
2.5. Principal component analysis (PCA)
The variables of reducing sugars, total free amino acids, sucrose, ECSt, Phi2, PhiNO, PhiNPQ, and qL, were excluded in the PCA analysis because they showed little influence of the analyzed treatments as shown in the univariate analysis. Besides, when these variables were added to the PCA, the variation of the explained data was low (43.1%). The contributions of variables when all of them were used in the PCA as well as the contribution of selected variables are shown in the Supplementary data, Table 1.
Results of the PCA evaluating the effect of the treatments on leaf content of starch, proline, antioxidant enzymes, and protein are shown in Figure 4. The PCA explained 64.8% of the data variance; the first axis (PC1) explained 45.0%, and the second axis (PC2) explained 19.8%. The PC1 was affected mainly by APX, SOD, and starch, while the values of GR, CAT, MDA, proline, and protein were explained by the PC2. The Se content in leaves showed a better correlation with APX and SOD, but a low correlation with Starch. A strong correlation between leaf Se content and the antioxidant enzymes CAT and SOD was noticed in the PCA.
Meanwhile, the biplot correlation clusters clearly distinguished the treatments and their respective correlations (Figure 4). The treatment -8BOS showed a clear correlation with GR and CAT, corroborating the previously shown results (Figure 2). The principal component analysis also revealed a strong correlation of Se, APX, and SOD with the treatment respective to the previous Se application 4 days before the stress (-4BOS).
3. Discussion
Osmotic stress encompasses stress-induced decreasing water potential (ψw) in plant cells [27]. Considering that the water flow moves towards the lowest ψw, if the stress continues, the leaves start to lose water, reflecting in the ψw in the leaf [28]. As a typical response to low water potential, the leaves of coffee plants in this trial started to become wilted and flabby 5 days after the imposed stress, leading to leaf prostration because of the turgor loss during abiotic stress. Osmotic stress tolerance involves the maintenance of the plant water status and hence cells turgor. This condition may be reached by stomatal regulation decreasing transpiration loss or osmotic adjustment by the accumulation of osmoprotective substances, such as proline, glycine betaine, soluble proteins, and sugars that help plants conserve their water status [29].
The results of ψw (Supplementary data, Figure 1) showed that all plants treated with PEG-6000 suffered from the OS. The effect of OS can also be illustrated in Figure 5. The OS imposed mild stress in the treatments -8BOS, -4BOS, 0OS, and +OS-Se (ψw from -1.5 to -2.5MPa). Meanwhile, the treatments +4AOS and +8AOS were subjected to severe stress (ψw > -2.5) (Supplementary data, Figure 1; Figure 5). According to Suma [30], non-susceptible plants can keep a minor reduction in ψw (6.9%) compared with a higher reduction (14.4%) in susceptible genotypes of finger millet. Then, after the plants were submitted to the stress, the Se application might have acted as a stressor in coffee plants, leading those plants to higher water potential loss and potentializing the OS response.
Plants treated with Se at all times had a higher relative water content (RWC) than the controls, including at the turgor loss point (RWCtlp), whereas the turgor loss point (tlp) was less negative in all the same plants. The tlp indicates the cell water potential inducing turgor pressure loss, which is crucial to maintain gas exchange and plant growth, and plants with low tlp tend to maintain stomatal and hydraulic conductance, photosynthetic efficiency, and growth at lower external water potential [31]. This parameter is thus correlated with the ability to tolerate stress rather than avoid it.
Although it is considered that a more negative tlp improves drought tolerance, as described above, it is also suggested that a less negative tlp may be helpful because it enables leaves to lose turgor quickly and close their stomata, and thereby maintain a high RWCtlp [32]. This response pattern was observed in our study. Plants treated with Se showed 20% higher RWCtlp than untreated plants. According to DaMatta and Ramalho [7], coffee leaves usually have high RWCtlp, regardless of water availability to avoid stress rather than tolerate it. For the authors, this seems to be more related to stomatal regulation and gas exchange maintenance than turgor. We suggest that in our experiment, Se helped the stomatal regulation in coffee plants under osmotic stress as a strategy to decrease transpiration rates. Similar results were related to yellow sweet clover under drought stress and Se addition [33].
A high RWCtlp despite very low water potential is generally correlated with osmotic adjustment. However, our study did not observe an increase in the concentration of proline or soluble sugars as a standard response to stress or Se application (Figure 3). Furthermore, it has been reported for coffee leaves that accumulation of proline and other solutes does not always correlate well with drought tolerance [7]. In our study, the application of Se 4 days before and 8 days after stress (-4BOS and +8AOS) seems to have stimulated an osmotic adjustment due to the combination of very low water potential, high relative water content, and proline accumulation concerning the controls (with and without stress). But we believe that the high RWCtlp in all treatments with Se application can be better explained by stomatal regulation, as mentioned before.
Stomatal closure in response to stress might limit CO2 absorption by the leaves. In our study, photosystem II efficiency showed no change in response to stress or Se (Supplementary data, Figure 2). Associated with the fact that plants did not show a reduction in growth, we can conclude that there was no photochemical limitation in photosynthesis. In line with this, we also observed no alteration in soluble sugars or sucrose in response to stress, suggesting no significant chemical limitations (Figure 3). Only starch was reduced in response to stress and Se application.
In photosynthetic cells, starch is synthesized mostly using a fraction of the CO2-fixed carbon and temporarily stored in the chloroplast called “transitory”. The transitory starch is usually synthesized during the day and consumed at night to provide a constant flow of carbon and energy without photosynthesis [34]. Starch is considered the major carbohydrate storage in plants [35]. In stressful conditions, starch represents a pool of energy that can induce metabolic responses and help plants overcome harmful circumstances. It can be broken down into low molecular weight compounds. Starch degradation can be stimulated in response to osmotic stress to promote osmotic adjustment, which might explain the response to treatments in which the OS was imposed. Besides this, a noteworthy factor is that ABA biosynthesis is the primary signal for starch degradation in response to osmotic stress [36].
In the extensive literature survey by Thalmann et al., [35], the authors discovered that in 23 of 36 of the considered studies, leaf starch content was said to decrease in response to abiotic stress, regardless of the species assessed. This result highlights the importance of starch in providing energy to deal with abiotic stresses. Then, the starch catabolism displaces carbons to produce osmoprotectants that induce osmotic adjustments and stabilize proteins [37,38], and also promotes signals that induce stress responses [35]. The fact that Se application caused a more substantial reduction of starch content in coffee leaves under drought conditions led us to the hypothesis that reduced starch accumulation during the OS might be a plant strategy to maintain the flow of carbon and energy availability for growth during the harmful condition (Figure 3 and Figure 4) [39].
The higher Se content in the plants supplied with Se was expected since Se supplementation in coffee plants by foliar application (and other plant species) has been studied in the literature [19,40]. Selenium can be supplied by seed, soil, and foliar application [41,42]. However, when applied at the same rate, foliar applications have been considered the most efficient way to increase Se content in plant tissue [41,43,44]. Since an active chemical chain builds the Se assimilation pathway, the addition of Se to stressed plants (+4AOS and +8AOS) possibly consumed the energy used to trigger metabolic responses that was supposed to be used to overcome the stress, making the plants unable to keep the ψw at higher levels in the leaves.
Despite the beneficial effects of Se related in the literature, it can be toxic depending on the tissue levels and plant health condition [45,46]. Due to the chemical similarity of Se and S, selenate (SeO4-) is transported into the plants through sulfate transporters [47,48]. Since it is inside the plant cell, it is metabolized in the plastids via sulfur assimilation pathway to selenocysteine - SeCys or selenomethionine - SeMet [49,50]. Se-SeO4- is first assimilated by an active form via the enzyme adenosine triphosphate sulfurylase (APS) and APS-reductase (APR). Adenosine triphosphate sulfurylase binds selenate with triphosphate sulfurylase (ATP) to form adenosine 5’-phosphoselenate (APSe). After that, APSe is reduced to selenite by APR [47,51,52]. Selenite is then converted into SeCys and available to be converted into other organic compounds – like SeMet and proteins – or stocked in the vacuoles [53]. Notably, these Se-amino acids serve as precursors of ethylene, and the production of this phytohormone is enhanced under stress conditions collaborating with stomatal closure [33,54].
Organic Se in excess, such as SeMet and SeCys, might cause toxicity to plant cells by forming malformed selenoproteins due to the replacement of Cys/Met with SeCys and SeMet in the peptide chain. Changing between Cys and SeCys changes the cellular protein’s structure by changing the disulfide bond to a diselenide bond, which affects the peptide chains redox potential [55]. Protein function might be compromised if the organic selenocompounds (like SeCys and SeMet) are non-specifically integrated into proteins in place of their sulfur (S) equivalents. This condition might trigger the plants’ negative responses and osmotic imbalances [56,57,58]. This result is also supported by the protein content in the leaves of the treatments +4AOS and +8AOS, in which the Protein content found in the treatments was higher than in the stressed plants without Se supply (+OS-Se) (Figure 3).
Several studies have shown the positive effect of Se on increasing antioxidant enzyme activities [40,59]. The wide antioxidant capacity is due to the promotion of the selenoproteins and enzyme cofactor role that enhance the antioxidant enzymes, such as glutathione peroxidase (GPX) and glutathione reductase (GR), which combat ROS during plant osmotic imbalance under stressful situations. The positive correlation between Se and GPX has been described and implicated in the presence of Se-dependent GPX [60,61]. It may be an osmoprotective strategy to mitigate the harmful effects of abiotic stresses, such as drought [18,62], salinity [63], heavy metals [64], and low temperature [19].
Indeed, in this trial, GR was the enzyme that better responded to the application of Se, and only the treatment with Se application 8 days before the stress was induced (-8BOS) was able to increase the content of APX, CAT, SOD, and GR at the same time, showing that prior Se supply is the best way to induce antioxidant activity to trigger metabolic responses to ROS, priming the plants to trigger metabolic responses against the oxidative stress which is coming. These results corroborated with those of Silva et al., [65], who also found that Se foliar application can provide an enhanced antioxidant metabolism by increasing superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX) and glutathione reductase (GR) activity.
The effects of Se on the antioxidant system of plants under abiotic stresses are extensively explored as the primary regulator of plant growth and yield under these conditions [46]. This condition was evident in our study, in which most of the observed results can be explained by factors related to the antioxidant metabolism of the plant (Figure 4). What has also been well discussed is how and to what extent different doses of Se in other species, plant organs, and developmental stages affect plant metabolism, which explains the varied responses to Se described in the literature.
4. Materials and Methods
4.1. Plant Materials and Study Site
The experiment was carried out in a greenhouse using arabica coffee seedlings (Coffea arabica cv. Catuaí), one of Brazil’s most traditional species. The cv. Catuaí is well known for its high beverage quality, good plant health, and high yield [66,67]. The plants used in the trial were at the age of 5-6 fully expanded leaves and were previously selected to keep uniformity and high health. The plants were provided by the National Institute of Science and Technology of Coffee (INCT Café).
The seedlings were produced in 1 L plant grow bags filled with subsoil + cattle manure at a ratio of 3:1, with 5 g of single superphosphate being added to each kilogram of the mixture. After the seedlings reached 5-6 fully expanded leaves, they were installed in a greenhouse at the Soil Science Department at the Federal University of Lavras – UFLA, Minas Gerais State, for 20 days to acclimate. The greenhouse temperature was 25/15 °C day/night, and the relative humidity was 50/85% day/night.
After the acclimation, the substrate was removed from each root system, and plants were transferred to 1-L black plastic pots with nutritive solution [68]. All plants underwent an acclimatization process for two weeks by applying a 20% and 40% ionic strength, respectively, for each week. After that, the plants were randomly selected to compose the treatments. According to Salgado et al., [69], the plants were kept at 40% of the ionic strength until the end of the trial. The black plastic pots were used with a polystyrene layer on top to avoid algae growth into the nutrient solution. In addition, we used a system composed of an air compressor pump and clear PVC flexible tubing to keep the nutrient solution oxygenating during the experiment.
4.2. Experimental design and treatments
The experimental design was in a randomized block with seven treatments and four replicates. The treatments consisted of Se application through foliar supply on five different days compared with the day of induced osmotic stress to establish the best day to apply Se in coffee plants under induced drought stress. The treatments were: i) eight days before induced osmotic stress (-8 BOS); ii) four days before induced osmotic stress (-4 BOS); iii) the same day of the induced osmotic stress (0 OS); iv) four days after induced osmotic stress (+4 AOS); v) eight days after induced osmotic stress (+8 AOS). Two control treatments – vi) induced osmotic stress without Se (+OS -Se); and vii) without stress and Se. The osmotic stress was induced by Polyethylene Glycol (PEG-6000). To induce the priming effect of Se foliar application and the alleviating effect, the treatments consisting of Se foliar application before the osmotic stress were called “priming treatments”, whereas plants treated after the osmotic stress were named “alleviated treatments”.
4.3. Application of foliar treatments
The foliar Se application was performed according to Sousa et al., [19]. In brief, the plants with Se application were sprayed with 5 mL of Se solution at 80 mg L−1 Se + 0.5% v/v of mineral oil, and the remaining plants were sprayed with a mineral-oil solution at the rate of 0.5% v/v. On the day of application, the plants were moved to the outer of the greenhouse to avoid contaminating the remaining plants. The Se source was Na2SeO4 - Sigma Aldrich 98.9%.
4.4. Osmotic stress imposition and leaf water status
Polyethylene Glycol with a molecular weight of 6000 (PEG-6000) was added to the nutritive solution according to Villela et al., [70] to induce the osmotic stress of −0.8 MPa. For this, 261.95 g L-1 of PEG-6000 were added to each plastic pot containing nutritive solution in the respective treatment with stress. The osmotic potential was used based on previous tests with coffee considering the osmotic potential of -0.1; -0.2; -0.4; -0.6; -0.8; e -1.0 MPa. The osmotic potential of -0.8 MPa promoted a leaf water potential (Ѱw) between -1.5 – -2.5, considered moderate stress to the coffee plants [71]. The determination of the Ѱw in each leaf was carried out with a Scholander pressure chamber (model 1000, PMS Instruments, Albany, NY, USA) [72] to confirm the leaf turgor on the day of sample collection and chlorophyll fluorescence parameters evaluation (Supplementary Figure S1).
4.5. Leaf sample collection and preparation
All treatments’ leaf sample collection and photosynthetic parameters were performed seven days after the last Se foliar application (+8 AOS). The second fully expanded pair of leaves from the top to bottom was used to perform the non-invasive analysis of photosynthetic parameters (MultispeQ®) [73]. After the measurement, the leaves were collected and immediately snap-frozen in liquid nitrogen, individually macerated in liquid nitrogen, homogenized in a cooled mortar using 0.1 g of the antioxidant polyvinylpyrrolidone (PVPP), and stored at -80°C. The frozen samples were used to determine the analyses of lipid peroxidation (MDA content), hydrogen peroxide (H2O2), catalase (CAT), superoxide dismutase (SOD), glutathione reductase (GR), and ascorbate peroxidase (APX).
The third and fourth fully expanded pairs of leaves from top to bottom were collected and washed three times with distilled water. All samples were dried at 65°C for 72 h and were subjected to grinding in Willey Grinder. Ground samples were labeled and kept in air-tight plastic containers until they were used to quantify Se content, carbohydrates, protein, total free amino acids, and proline.
4.6. Assessments
4.6.1. Selenium content in leaves and detection limit (LOD and LOQ)
The Se content in the leaves was performed according to the USEPA 3051A protocol (U.S. Environmental Protection Agency - USEPA) with modifications [74]. Briefly, 0.5 g of dried leaves samples was digested with 5 mL of HNO3 in a microwave (Mars 5, CEM Corporation, Matthews, NC, USA). To avoid foaming and splashing, the vessels were allowed in a cool room with a controlled temperature for 30 minutes after the end of the digestion program, opened carefully, and the volume was made up to 50 mL with water. A blank and a certified reference material for Se (White clover, BCR402-IRMM) were included in each batch of samples. The Se content in the leaves was measured using GFAAS (Graphite Furnace Atomic Absorption Spectrometry, Atomic Absorption Spectrometry with Zeeman background correction, and an EDL lamp for Se; Analyst™ 800 AAS, Perkin Elmer). The detection and quantification limits (LOD and LOQ) were determined according to Silva Junior et al., [75]. The LOD and LOQ for Se were respectively 2.49 and 8.32 μg kg−1. The Se recovery rate in the reference material was 96.7% ± 1.28.
4.6.2. Carbohydrates, total protein, total free amino acids
The extraction of carbohydrates and proteins was based on Zanandrea et al., [76]. Dried samples were weighed (0.2 g) and mixed with 5 mL of 100 mM potassium phosphate buffer (pH 7.0) and then warmed in a water bath at 40°C for 30 minutes. The solution was centrifuged at 10,000 g for 20 min and the supernatant was collected. This procedure was made twice, and the supernatant was combined, totalizing 10 mL. The pellet was resuspended and used for starch extraction, mixing 8 mL of potassium acetate buffer (200 mM pH 4.8) and 2 mL of amyloglucosidase (1 mg mL-1; 16 units of enzyme).
4.6.3. Proline
Proline content was estimated by the method described by Bates et al., [81]. Dried leaf samples (0.2 g) were weighed and macerated with sulfosalicylic acid 3% and heated in a water bath for 60 minutes at room temperature. After that, the samples were centrifuged at 10,000 × g for 30 min. The supernatant (0.1 mL) was then mixed with acid ninhydrin and determined by a colorimetric method (520 nm).
4.6.4. Antioxidant enzymes (SOD, CAT, APX, GR)
The extraction of antioxidant enzymes was based on Biemelt et al., [82]. Frozen leaf samples were weighted (0.2 g) and mixed with 1.5 mL of potassium phosphate buffer solution (0.1 mol L−1, pH 7.8 + 0.1 mol L−1 EDTA, pH 7.0, 0.01 mol L−1 ascorbic acid, and 22 mg of PVPP). The solution was centrifuged at 13,000 g for 10 min at 4°C. The enzymatic analyses’ quality assurance and quality control were warranted by using two blanks in each reading plate and operating the samples at 0–4°C. In addition, the enzyme extraction was performed on the day of the analysis to avoid the oxidation of the enzyme extract. The analyses were carried out in triplicates and were measured using an Epoch® Microplate Spectrophotometer (BioTek, United States). The supernatant was used to quantify the activity of superoxide dismutase (SOD, EC: 1.15.1.1), catalase (CAT, EC: 1.11.1.6), ascorbate peroxidase (APX, EC: 1.11.1.11) and glutathione reductase (GR, EC: 1.8.1.7).
The essay on SOD activity was performed by measuring its ability to inhibit the photochemical reduction of nitro blue tetrazolium at 560 nm [83]. Catalase (CAT) activity was assayed by measuring the rate of decomposition of H2O2 at 240 nm [84]. Ascorbate peroxidase (APX) was determined by reducing ascorbate at 290 nm [85]. Glutathione reductase (GR) was assayed according to the methodology proposed by Schaedle and Bassham [86] and adapted by García-Limones et al., [87].
4.6.5. Hydrogen peroxide and lipid peroxidation (malondialdehyde)
The frozen leaf tissue (0.2 g) was homogenized in 5 mL of trichloroacetic acid (TCA), and centrifuged at 12,000 g for 15 min at 4°C. The supernatant was collected to quantify the hydrogen peroxide according to Velikova [88] with modifications [89]. Lipid peroxidation analysis was assayed from the content of malondialdehyde (MDA) using thiobarbituric acid (TBAR) according to Buege and Aust [90] and Silva et al., [91].
4.6.6. Chlorophyll fluorescence parameters (MultispeQ®)
The electron transport and electrochromic shift parameters were measured with the handheld unit MultispeQ® using the PhotosynQ web application (https://photosynq.org) according to Kuhlgert et al., [73]. The following parameters were measured: total electrochromic shift (ECSt); linear electron flux (LEF); total non-photochemical quenching (NPQt); quantum yield of PSII (Phi2); quantum yield of non-regulated energy loss in PSII (PhiNO), the quantum yield of regulated non-photochemical energy loss in PSII (PhiNPQ), and a fraction of PSII centers which are in the open state (qL).
4.7. Statistical analysis and PCA
Due to the lack of homogeneity of the residues of some variables, generalized linear models (GLMs) were constructed to compare the treatments tested to each variable studied. After building the models, a Chi-square test was performed to determine the differences that existed between treatments studied with the ANOVA function [92], complemented by multiple comparisons with the “ghlt function” [93]. The comparisons were carried out as follows: I) Each treatment with water deficit was compared with the treatment without water deficit, and II) comparison involved the different strategies of Se application compared with the cultivation with a water deficit. In addition, principal component analysis (PCA) was performed to determine the relationships among some variables. The variables were selected according to the main effects observed in the univariate analysis, and to increase the explained variance in PCA. All statistical analyses were performed with the R software [94] using the base, stats, nlme, multcomp, FactoMineR, and factoextra packages [94,95,96,97].
5. Conclusions
The OS induced by PEG-6000 imposed significant stress on the Coffea arabica cv. Catuaí, promoting an imbalance in the water relationship. At the same time, the OS reduced the GR and SOD activity compared with the control treatment. Selenium foliar supply revealed great potential for reducing the adverse effects of OS as a priming strategy 8 days before stress, improving the water relations, increasing the enzymatic activity (GR, SOD, and CAT), and potentiating the starch degradation under stress conditions. These findings also assist decision-makers in how to deal with a foreseen drought in coffee plantations, where the earlier administration of foliar Se aids in the setting up of metabolic reactions that help the plants to combat the stress caused by a shortage of water. On the other hand, the post-stress application seems to impose extra stress on the plants, leading them to reduce the water potential.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
G.F.S, M.A.S., M.R.C., and L.G.G.G designed the research; G.F.S., M.A.S., M.R.C., P.A.N.B., G.A.Z van O, and G.G.A.Z van O. conducted the experiments and analyses; G.F.S, M.A.S, and E.G.M. analyzed the data; G.F.S.; M.A.S.; E.G.M.; P.A.N.B., and M. R. C. wrote the original draft; G.F.S, M.A.S., and L.R.G.G. wrote the final text and approved the final version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Research Support Foundation of the State of Minas Gerais (FAPEMIG), the National Institute of Science and Technology of Coffee (INCT/Café), under FAPEMIG grant (CAG APQ 03605/17), and the National Council for Scientific and Technological Development (CNPq). This study was also partially supported by grant #2021/06968-3, São Paulo Research Foundation (FAPESP), and funding from the Coordination for the Improvement of Higher Education Personnel (CAPES).
Data Availability Statement
The data presented in this study are available from the corresponding author upon reasonable request.
Acknowledgments
We are grateful to CNPq, CAPES, and FAPEMIG from Brazil for their financial support for the research project and scholarships.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- IPCC Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change 2014, 151.
- Ortiz-Bobea, A.; Ault, T.R.; Carrillo, C.M.; Chambers, R.G.; Lobell, D.B. Anthropogenic Climate Change Has Slowed Global Agricultural Productivity Growth. Nat. Clim. Chang. 2021, 11, 306–312. [Google Scholar] [CrossRef]
- Kotz, M.; Levermann, A.; Wenz, L. The Effect of Rainfall Changes on Economic Production. Nature 2022, 601, 223–227. [Google Scholar] [CrossRef]
- Cerdán, C.R.; Rebolledo, M.C.; Soto, G.; Rapidel, B.; Sinclair, F.L. Local Knowledge of Impacts of Tree Cover on Ecosystem Services in Smallholder Coffee Production Systems. Agric. Syst. 2012, 110, 119–130. [Google Scholar] [CrossRef]
- Chain-Guadarrama, A.; Martínez-Salinas, A.; Aristizábal, N.; Ricketts, T.H. Ecosystem Services by Birds and Bees to Coffee in a Changing Climate: A Review of Coffee Berry Borer Control and Pollination. Agric. Ecosyst. Environ. 2019, 280, 53–67. [Google Scholar] [CrossRef]
- Davis, A.P.; Govaerts, R.; Bridson, D.M.; Stoffelen, P. An Annotated Taxonomic Conspectus of the Genus Coffea (Rubiaceae). Bot. J. Linn. Soc. 2006, 152, 465–512. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Ramalho, J.D.C. Impacts of Drought and Temperature Stress on Coffee Physiology and Production: A Review. Brazilian J. Plant Physiol. 2006, 18, 55–81. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought Stress in Plants: A Review on Morphological Characteristics and Pigments Composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, Z.; Waraich, E.A.; Akhtar, S.; Anjum, S.; Ahmad, T.; Mahboob, W.; Hafeez, O.B.A.; Tapera, T.; Labuschagne, M.; Rizwan, M. Physiological Responses of Wheat to Drought Stress and Its Mitigation Approaches. Acta Physiol. Plant. 2018, 40, 1–13. [Google Scholar] [CrossRef]
- Nawaz, F.; Ahmad, R.; Ashraf, M.Y.Y.; Waraich, E.A.A.; Khan, S.Z.Z. Effect of Selenium Foliar Spray on Physiological and Biochemical Processes and Chemical Constituents of Wheat under Drought Stress. Ecotoxicol. Environ. Saf. 2015, 113, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of Drought Stress on Photosynthesis and Photosynthetic Electron Transport Chain in Young Apple Tree Leaves. Biol. Open 2018, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Semedo, J.N.; Rodrigues, W.P.; Dubberstein, D.; Martins, M.Q.; Martins, L.D.; Pais, I.P.; Rodrigues, A.P.; Leitão, A.E.; Partelli, F.L.; Campostrini, E.; et al. Coffee Responses to Drought, Warming and High [CO2] in a Context of Future Climate Change Scenarios. In Climate Change Management; Alves, F., Leal Filho, W., Azeiteiro, U., Eds.; Climate Change Management; Springer International Publishing: Cham, 2018; pp. 465–477. ISBN 978-3-319-72873-5. [Google Scholar]
- Feng, R.; Wei, C.; Tu, S. The Roles of Selenium in Protecting Plants against Abiotic Stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Bhadwal, S.; Sharma, S. Selenium Alleviates Carbohydrate Metabolism and Nutrient Composition in Arsenic Stressed Rice Plants. Rice Sci. 2022, 29, 385–396. [Google Scholar] [CrossRef]
- Prasad, R.; Shivay, Y.S. Agronomic Biofortification of Plant Foods with Minerals, Vitamins and Metabolites with Chemical Fertilizers and Liming. J. Plant Nutr. 2020, 43, 1534–1554. [Google Scholar] [CrossRef]
- Abul-Soud, M.; Abd-Elrahman, S. Foliar Selenium Application to Improve the Tolerance of Eggplant Grown under Salt Stress Conditions. Int. J. Plant Soil Sci. 2016, 9, 1–10. [Google Scholar] [CrossRef]
- Ravello, R.A.V.; de Oliveira, C.; Lessa, J.; Boas, L.V.V.; de Castro, E.M.; Guilherme, L.R.G.; Lopes, G. Selenium Application Influenced Selenium Biofortification and Physiological Traits in Water-Deficit Common Bean Plants. Crop Pasture Sci. 2021, 73, 44–55. [Google Scholar] [CrossRef]
- Sousa, G.F. de; Silva, M.A.; de Morais, E.G.; Van Opbergen, G.A.Z.; Van Opbergen, G.G.A.Z.; de Oliveira, R.R.; Amaral, D.; Brown, P.; Chalfun-Junior, A.; Guilherme, L.R.G. Selenium Enhances Chilling Stress Tolerance in Coffee Species by Modulating Nutrient, Carbohydrates, and Amino Acids Content. Front. Plant Sci. 2022, 13, 1–15. [Google Scholar] [CrossRef]
- Luo, H.; Xing, P.; Liu, J.; Pan, S.; Tang, X.; Duan, M. Selenium Improved Antioxidant Response and Photosynthesis in Fragrant Rice (Oryza Sativa L.) Seedlings during Drought Stress. Physiol. Mol. Biol. Plants 2021, 27, 2849–2858. [Google Scholar] [CrossRef]
- Thuc, L.V.; Sakagami, J.I.; Hung, L.T.; Huu, T.N.; Khuong, N.Q.; Vi, L.L.V. Foliar Selenium Application for Improving Drought Tolerance of Sesame (Sesamum Indicum L.). Open Agric. 2021, 6, 93–101. [Google Scholar] [CrossRef]
- Becerra-Vázquez, Á.G.; Coates, R.; Sánchez-Nieto, S.; Reyes-Chilpa, R.; Orozco-Segovia, A. Effects of Seed Priming on Germination and Seedling Growth of Desiccation-Sensitive Seeds from Mexican Tropical Rainforest. J. Plant Res. 2020, 133, 855–872. [Google Scholar] [CrossRef]
- WANG, X.; LIU, F.; JIANG, D. Priming: A Promising Strategy for Crop Production in Response to Future Climate. J. Integr. Agric. 2017, 16, 2709–2716. [Google Scholar] [CrossRef]
- Franzoni, G.; Cocetta, G.; Prinsi, B.; Ferrante, A.; Espen, L. Biostimulants on Crops: Their Impact under Abiotic Stress Conditions. Horticulturae 2022, 8. [Google Scholar] [CrossRef]
- Nephali, L.; Piater, L.A.; Dubery, I.A.; Patterson, V.; Huyser, J.; Burgess, K.; Tugizimana, F. Biostimulants for Plant Growth and Mitigation of Abiotic Stresses: A Metabolomics Perspective. Metabolites 2020, 10, 1–26. [Google Scholar] [CrossRef]
- Jolayemi, O.L.; Malik, A.H.; Ekblad, T.; Fredlund, K.; Olsson, M.E.; Johansson, E. Protein-Based Biostimulants to Enhance Plant Growth—State-of-the-Art and Future Direction with Sugar Beet as an Example. Agronomy 2022, 12. [Google Scholar] [CrossRef]
- Upadhyaya, H.; Sahoo, L.; Panda, S.K. Molecular Physiology of Osmotic Stress in Plants. In Molecular Stress Physiology of Plants; Springer India: India, 2013; ISBN 9788132208075. [Google Scholar]
- Reddy, Y.A.N.; Reddy, Y.N.P.; Ramya, V.; Suma, L.S.; Reddy, A.B.N.; Krishna, S.S. Drought Adaptation: Approaches for Crop Improvement; Elsevier Inc., 2020; ISBN 9780128200896.
- Moustakas, M.; Sperdouli, I.; Moustaka, J. Early Drought Stress Warning in Plants: Color Pictures of Photosystem II Photochemistry. Climate 2022, 10, 179. [Google Scholar] [CrossRef]
- Suma, L.S. Characterization of Selected Germplasm Accessions for Drought Tolerance in Finger Millet (Eleusine Coracana (L.) Gaertn, University of Agricultural Sciences, GKVK., 2014.
- Bartlett, M.K.; Scoffoni, C.; Sack, L. The Determinants of Leaf Turgor Loss Point and Prediction of Drought Tolerance of Species and Biomes: A Global Meta-Analysis. Ecol. Lett. 2012, 15, 393–405. [Google Scholar] [CrossRef]
- Read, J.; Sanson, G.D.; Garine-Wichatitsky, M. d.; Jaffre, T. Sclerophylly in Two Contrasting Tropical Environments: Low Nutrients vs. Low Rainfall. Am. J. Bot. 2006, 93, 1601–1614. [Google Scholar] [CrossRef]
- Kostopoulou, P.; Barbayiannis, N.; Noitsakis, B. Water Relations of Yellow Sweetclover under the Synergy of Drought and Selenium Addition. Plant Soil 2010, 330, 65–71. [Google Scholar] [CrossRef]
- Ribeiro, C.; Stitt, M.; Hotta, C.T. How Stress Affects Your Budget—Stress Impacts on Starch Metabolism. Front. Plant Sci. 2022, 13, 1–9. [Google Scholar] [CrossRef]
- Thalmann, M.; Santelia, D. Starch as a Determinant of Plant Fitness under Abiotic Stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef]
- Thalmann, M.; Pazmino, D.; Seung, D.; Horrer, D.; Nigro, A.; Meier, T.; Kölling, K.; Pfeifhofer, H.W.; Zeeman, S.C.; Santelia, D. Regulation of Leaf Starch Degradation by Abscisic Acid Is Important for Osmotic Stress Tolerance in Plants. Plant Cell 2016, 28, 1860–1878. [Google Scholar] [CrossRef] [PubMed]
- Kempa, S.; Krasensky, J.; Dal Santo, S.; Kopka, J.; Jonak, C. A Central Role of Abscisic Acid in Stress-Regulated Carbohydrate Metabolism. PLoS One 2008, 3. [Google Scholar] [CrossRef]
- Danso, O.P.; Asante-badu, B.; Zhang, Z.; Song, J.; Wang, Z.; Yin, X. Selenium Biofortification : Strategies, Progress and Challenges. 2023, 1–29.
- Sitnicka, D.; Orzechowski, S. Cold-Induced Starch Degradation in Potato Leaves — Intercultivar Differences in the Gene Expression and Activity of Key Enzymes. Biol. Plant. 2014, 58, 659–666. [Google Scholar] [CrossRef]
- Mateus, M.P. de B.; Tavanti, R.F.R.; Tavanti, T.R.; Santos, E.F.; Jalal, A.; Reis, A.R. dos Selenium Biofortification Enhances ROS Scavenge System Increasing Yield of Coffee Plants. Ecotoxicol. Environ. Saf. 2021, 209. [Google Scholar] [CrossRef]
- Silva, M.A.; Sousa, G.F. De; Bañuelos, G.; Amaral, D.; Brown, P.H. Selenium Speciation in Se-Enriched Soybean Grains from Biofortified Plants Grown under Different Methods of Selenium Application. 2023, 1–13.
- Silva, M.A.; de Sousa, G.F.; Corguinha, A.P.B.; de Lima Lessa, J.H.; Dinali, G.S.; Oliveira, C.; Lopes, G.; Amaral, D.; Brown, P.; Guilherme, L.R.G. Selenium Biofortification of Soybean Genotypes in a Tropical Soil via Se-Enriched Phosphate Fertilizers. Front. Plant Sci. 2022, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Liu, K.; Li, M.; Zhang, W.; Zhao, X.; Zhao, Z.; Liu, X. Difference of Selenium Uptake and Distribution in the Plant and Selenium Form in the Grains of Rice with Foliar Spray of Selenite or Selenate at Different Stages. F. Crop. Res. 2017, 211, 165–171. [Google Scholar] [CrossRef]
- Manojlović, M.S.; Lončarić, Z.; Cabilovski, R.R.; Popović, B.; Karalić, K.; Ivezić, V.; Ademi, A.; Singh, B.R. Biofortification of Wheat Cultivars with Selenium. Acta Agric. Scand. Sect. B Soil Plant Sci. 2019, 69, 715–724. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.H.; Winkel, L.H.E.; Lin, Z.-Q. Selenium in Plants: Molecular, Physiological, Ecological and Evolutionary Aspects; Pilon-Smits, E.A.H., Winkel, L.H.E., Lin, Z.-Q., Eds.; Plant Ecophysiology; 1st ed.; Springer International Publishing: Cham, 2017; ISBN 978-3-319-56248-3. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 1–52. [Google Scholar] [CrossRef]
- Gupta, M.; Gupta, S. An Overview of Selenium Uptake, Metabolism, and Toxicity in Plants. Front. Plant Sci. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Zhang, L.; Chu, C. Selenium Uptake, Transport, Metabolism, Reutilization, and Biofortification in Rice. Rice 2022, 15. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Z.; Luo, L.Y.; Fu, P.N.; Wang, Q.; Li, H.F. Selenium Uptake and Biotransformation in Brassica Rapa Supplied with Selenite and Selenate: A Hydroponic Work with HPLC Speciation and RNA-Sequencing. J. Agric. Food Chem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Lin, W.; Jiao, H.; Liu, J.; Chan, L.; Liu, X.; Wang, R.; Chen, T. Uptake, Transport, and Metabolism of Selenium and Its Protective Effects against Toxic Metals in Plants: A Review. Metallomics 2021, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sors, T.G.; Ellis, D.R.; Salt, D.E. Selenium Uptake, Translocation, Assimilation and Metabolic Fate in Plants. Photosynth. Res. 2005, 86, 373–389. [Google Scholar] [CrossRef]
- Lanza, M.G.D.B.; Reis, A.R. dos Roles of Selenium in Mineral Plant Nutrition: ROS Scavenging Responses against Abiotic Stresses. Plant Physiol. Biochem. 2021, 164, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.W.; Pilon-Smits, E.A.H.; Schiavon, M. Mechanisms of Selenium Hyperaccumulation in Plants: A Survey of Molecular, Biochemical and Ecological Cues. Biochim. Biophys. Acta - Gen. Subj. 2018, 1862, 2343–2353. [Google Scholar] [CrossRef]
- Konze, J.R.; Schilling, N.; Kende, H. Enhancement of Ethylene Formation by Selenoamino Acids. Plant Physiol. 1978, 62, 397–401. [Google Scholar] [CrossRef]
- Tangjaidee, P.; Swedlund, P.; Xiang, J.; Yin, H.; Quek, S.Y. Selenium-Enriched Plant Foods: Selenium Accumulation, Speciation, and Health Functionality. Front. Nutr. 2023, 9, 1–19. [Google Scholar] [CrossRef]
- Akladious, S.A. Influence of Different Soaking Times with Selenium on Growth, Metabolic Activities of Wheat Seedlings under Low Temperature Stress. African J. Biotechnol. 2012, 11, 14792–14804. [Google Scholar] [CrossRef]
- Schiavon, M.; Lima, L.W.; Jiang, Y.; Hawkesford, M.J. Effects of Selenium on Plant Metabolism and Implications for Crops and Consumers BT - Selenium in Plants: Molecular, Physiological, Ecological and Evolutionary Aspects. In; Pilon-Smits, E.A.H., Winkel, L.H.E., Lin, Z.-Q., Eds.; Springer International Publishing: Cham, 2017; pp. 257–275. ISBN 978-3-319-56249-0. [Google Scholar]
- Trippe, R.C.; Pilon-Smits, E.A.H. Selenium Transport and Metabolism in Plants: Phytoremediation and Biofortification Implications. J. Hazard. Mater. 2021, 404, 124178. [Google Scholar] [CrossRef]
- Elkelish, A.A.; Soliman, M.H.; Alhaithloul, H.A.; El-Esawi, M.A. Selenium Protects Wheat Seedlings against Salt Stress-Mediated Oxidative Damage by up-Regulating Antioxidants and Osmolytes Metabolism. Plant Physiol. Biochem. 2019, 137, 144–153. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, A. Role of Glutathione in Plant Abiotic Stress Tolerance. React. Oxyg. Nitrogen Sulfur Species Plants Prod. Metab. Signal. Def. Mech. 2019, 1, 159–172. [Google Scholar] [CrossRef]
- Rahman, M.; Rahman, K.; Sathi, K.S.; Alam, M.M.; Nahar, K.; Fujita, M.; Hasanuzzaman, M. Supplemental Selenium and Boron Mitigate Salt-induced Oxidative Damages in Glycine Max L. Plants 2021, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M. Application of Selenium A Useful Way to Mitigate Drought Stress: A Review. Open Access J. Biog. Sci. Res. 2020, 3. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Akbar, A.; Parveen, A.; Rasheed, R.; Hussain, I.; Iqbal, M. Phenological Application of Selenium Differentially Improves Growth, Oxidative Defense and Ion Homeostasis in Maize under Salinity Stress. Plant Physiol. Biochem. 2018, 123, 268–280. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; García-Caparrós, P.; Parvin, K.; Zulfiqar, F.; Ahmed, N.; Fujita, M. Selenium Supplementation and Crop Plant Tolerance to Metal/Metalloid Toxicity. Front. Plant Sci. 2022, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.M.; Rimoldi Tavanti, R.F.; Gratão, P.L.; Alcock, T.D.; Reis, A.R. dos Selenate and Selenite Affect Photosynthetic Pigments and ROS Scavenging through Distinct Mechanisms in Cowpea (Vigna Unguiculata (L.) Walp) Plants. Ecotoxicol. Environ. Saf. 2020, 201, 110777. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, F.M.; Ronchi, C.P.; Maestri, M.; Barros, R.S. Ecophysiology of Coffee Growth and Production. Brazilian J. Plant Physiol. 2007, 19, 485–510. [Google Scholar] [CrossRef]
- van der Vossen, H.; Bertrand, B.; Charrier, A. Next Generation Variety Development for Sustainable Production of Arabica Coffee (Coffea Arabica L.): A Review. Euphytica 2015, 204, 243–256. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. Calif. Agr. Expt. Sta. Circ. 1950, 347, 1–32. [Google Scholar]
- Salgado, O.G.G.; Teodoro, J.C.; Alvarenga, J.P.; de Oliveira, C.; de Carvalho, T.S.; Domiciano, D.; Marchiori, P.E.R.; Guilherme, L.R.G. Cerium Alleviates Drought-Induced Stress in Phaseolus Vulgaris. J. Rare Earths 2020, 38, 324–331. [Google Scholar] [CrossRef]
- Villela, F.A.; Doni Filho, L.; Sequeira, E.L. Tabela de Potencial Osmótico Em Função Da Concentração de Polietileno Glicol 6.000 e Da Temperatura. Pesqui. Agropecuária Bras. 1991, 26, 1957–1968. [Google Scholar]
- Fernandes, I.; Marques, I.; Paulo, O.S.; Batista, D.; Partelli, F.L.; Lidon, F.C.; Damatta, F.M.; Ramalho, J.C.; Ribeiro-Barros, A.I. Understanding the Impact of Drought in Coffea Genotypes: Transcriptomic Analysis Supports a Common High Resilience to Moderate Water Deficit but a Genotype Dependent Sensitivity to Severe Water Deficit. Agronomy 2021, 11, 1–27. [Google Scholar] [CrossRef]
- Scholander, P.F.; Bradstreet, E.D.; Hemmingsen, E.A.; Hammel, H.T. Sap Pressure in Vascular Plants. Science (80-. ). 1965, 148, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Kuhlgert, S.; Austic, G.; Zegarac, R.; Osei-Bonsu, I.; Hoh, D.; Chilvers, M.I.; Roth, M.G.; Bi, K.; TerAvest, D.; Weebadde, P.; et al. MultispeQ Beta: A Tool for Large-Scale Plant Phenotyping Connected to the Open PhotosynQ Network. R. Soc. Open Sci. 2016, 3. [Google Scholar] [CrossRef]
- 74. U.S. EPA Method 3051A (SW-846): Microwave Assisted Acid Digestion of Sediments, Sludges, and Oils. Ятыатат, 2007; 30, 75.
- Silva Junior, E.C.; Wadt, L.H.O.; Silva, K.E.; Lima, R.M.B.; Batista, K.D.; Guedes, M.C.; Carvalho, G.S.; Carvalho, T.S.; Reis, A.R.; Lopes, G.; et al. Natural Variation of Selenium in Brazil Nuts and Soils from the Amazon Region. Chemosphere 2017, 188, 650–658. [Google Scholar] [CrossRef]
- Zanandrea, I.; Alves, J.D.; Deuner, S.; Goulart, P. de F.P.; Henrique, P.D.C.; Silveira, N.M. Tolerance of Sesbania Virgata Plants to Flooding. Aust. J. Bot. 2010, 57, 661. [Google Scholar] [CrossRef]
- DISCHE, Z. General Color Reactions. In Carbohydrate chemistry; WHISTLER, R.L.., WOLFRAM, M.L., Eds.; New York: Academic: New York:, 1962; pp. 477–512. [Google Scholar]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The Estimation of Carbohydrates in Plant Extracts by Anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Biemelt, S.; Keetman, U.; Albrecht, G. Re-Aeration Following Hypoxia or Anoxia Leads to Activation of the Antioxidative Defense System in Roots of Wheat Seedlings. Plant Physiol. 1998, 116, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases. 1977, 309–314.
- Havir, E.A.; McHale, N.A. Biochemical and Developmental Characterization of Multiple Forms of Catalase in Tobacco Leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Schaedle, M.; Bassham, J.A. Chloroplast Glutathione Reductase. Plant Physiol. 1977, 59, 1011–1012. [Google Scholar] [CrossRef] [PubMed]
- García-Limones, C.; Hervás, A.; Navas-Cortés, J.A.; Jiménez-Díaz, R.M.; Tena, M. Induction of an Antioxidant Enzyme System and Other Oxidative Stress Markers Associated with Compatible and Incompatible Interactions between Chickpea (Cicer Arietinum L.) and Fusarium Oxysporum f. Sp. Ciceris. Physiol. Mol. Plant Pathol. 2002, 61, 325–337. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative Stress and Some Antioxidant Systems in Acid Rain-Treated Bean Plants. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Loreto, F.; Velikova, V. Isoprene Produced by Leaves Protects the Photosynthetic Apparatus against Ozone Damage, Quenches Ozone Products, and Reduces Lipid Peroxidation of Cellular Membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal Lipid Peroxidation. In Methods in Enzymology; 1978; Vol. 52, pp. 302–310 ISBN 9780121819521.
- Silva, V.M.; Rimoldi Tavanti, R.F.; Gratão, P.L.; Alcock, T.D.; Reis, A.R. dos Selenate and Selenite Affect Photosynthetic Pigments and ROS Scavenging through Distinct Mechanisms in Cowpea (Vigna Unguiculata (L.) Walp) Plants. Ecotoxicol. Environ. Saf. 2020, 201, 110777. [Google Scholar] [CrossRef]
- Chambers, J.M.; Hastie, T.J. Statistical Models in S; Wadsworth & Brooks: Pacific Grove, CA. , 1992; ISBN 9780534167646.
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biometrical J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing 2022.
- Pinheiro, J.; Bates, D. Linear and Nonlinear Mixed Effects Models Available online: https://cran.r-project.org/package=nlme.
- Kassambara, A.; Mundt, F. Extract and Visualize the Results of Multivariate Data Analyses Available online: https://cran.r-project.org/package=factoextra.
- Lê, S.; Josse, J.; Husson, F. FactoMineR : An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25. [Google Scholar] [CrossRef]
Figure 1.
Leaf Se content as a result of Se application in Coffea arabica cv. Catuaí seedlings under osmotic stress induced by PEG-6000. The values displayed are the distribution of four replicates. Asterisks refer to the significant difference when comparing all treatments with non-stressed plants without Se supply (-OS-Se) (p < 0.05). Dagger refers to the significant difference when comparing all treatments with stressed plants without Se supply (+OS-Se) (p < 0.05). Treatments: -8BOS- Application of Se 8 days before stress (Stressed plants); -4BOS - Application of Se 4 days before stress (Stressed plants); 0OS - Application of Se in stress (Stressed plants); +4AOS - Application of Se 4 days after stress (Stressed plants); +8AOS - Application of Se 8 days after stress (Stressed plants); +OS-Se - Without Se (Stressed plants); -OS-Se - Without Se (Non-stressed plants).
Figure 1.
Leaf Se content as a result of Se application in Coffea arabica cv. Catuaí seedlings under osmotic stress induced by PEG-6000. The values displayed are the distribution of four replicates. Asterisks refer to the significant difference when comparing all treatments with non-stressed plants without Se supply (-OS-Se) (p < 0.05). Dagger refers to the significant difference when comparing all treatments with stressed plants without Se supply (+OS-Se) (p < 0.05). Treatments: -8BOS- Application of Se 8 days before stress (Stressed plants); -4BOS - Application of Se 4 days before stress (Stressed plants); 0OS - Application of Se in stress (Stressed plants); +4AOS - Application of Se 4 days after stress (Stressed plants); +8AOS - Application of Se 8 days after stress (Stressed plants); +OS-Se - Without Se (Stressed plants); -OS-Se - Without Se (Non-stressed plants).
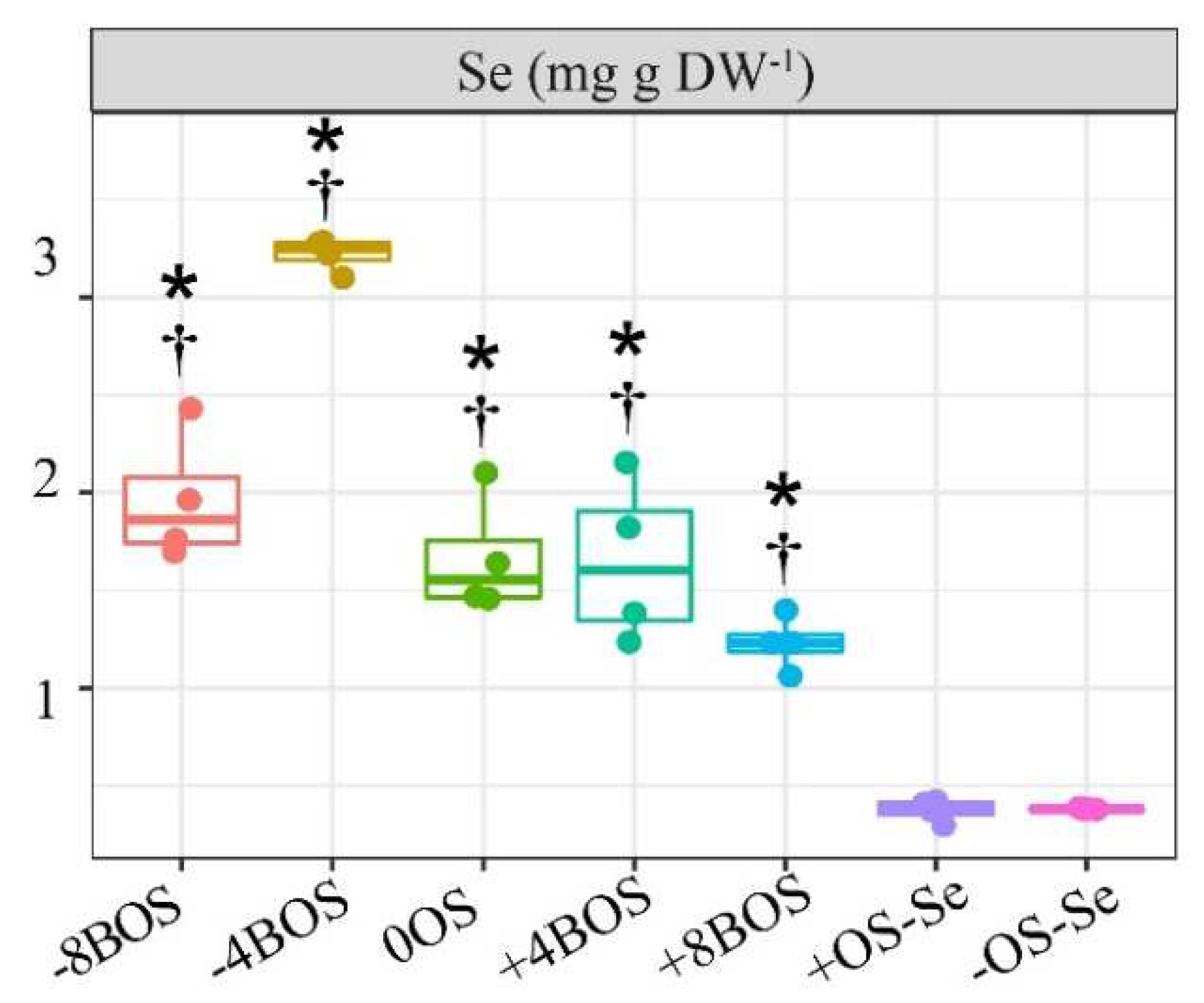
Figure 2.
Hydrogen peroxide (H2O2), MDA, and activity of leaf antioxidant enzymes as a result of Se application in Coffea arabica cv. Catuaí seedlings under osmotic stress induced by PEG-6000. The values displayed are the distribution of four replicates. Asterisks refer to the significant difference when comparing all treatments with non-stressed plants without Se supply (-OS-Se) (p < 0.05). Dagger refers to the significant difference when comparing all treatments with stressed plants without Se supply (+OS-Se) (p < 0.05). Treatments: -8BOS - Application of Se 8 days before stress (Stressed plants); -4BOS - Application of Se 4 days before stress (Stressed plants); 0OS - Application of Se in stress (Stressed plants); +4AOS - Application of Se 4 days after stress (Stressed plants); +8AOS - Application of Se 8 days after stress (Stressed plants); +OS-Se - Without Se (Stressed plants); -OS-Se - Without Se (Non-stressed plants).
Figure 2.
Hydrogen peroxide (H2O2), MDA, and activity of leaf antioxidant enzymes as a result of Se application in Coffea arabica cv. Catuaí seedlings under osmotic stress induced by PEG-6000. The values displayed are the distribution of four replicates. Asterisks refer to the significant difference when comparing all treatments with non-stressed plants without Se supply (-OS-Se) (p < 0.05). Dagger refers to the significant difference when comparing all treatments with stressed plants without Se supply (+OS-Se) (p < 0.05). Treatments: -8BOS - Application of Se 8 days before stress (Stressed plants); -4BOS - Application of Se 4 days before stress (Stressed plants); 0OS - Application of Se in stress (Stressed plants); +4AOS - Application of Se 4 days after stress (Stressed plants); +8AOS - Application of Se 8 days after stress (Stressed plants); +OS-Se - Without Se (Stressed plants); -OS-Se - Without Se (Non-stressed plants).
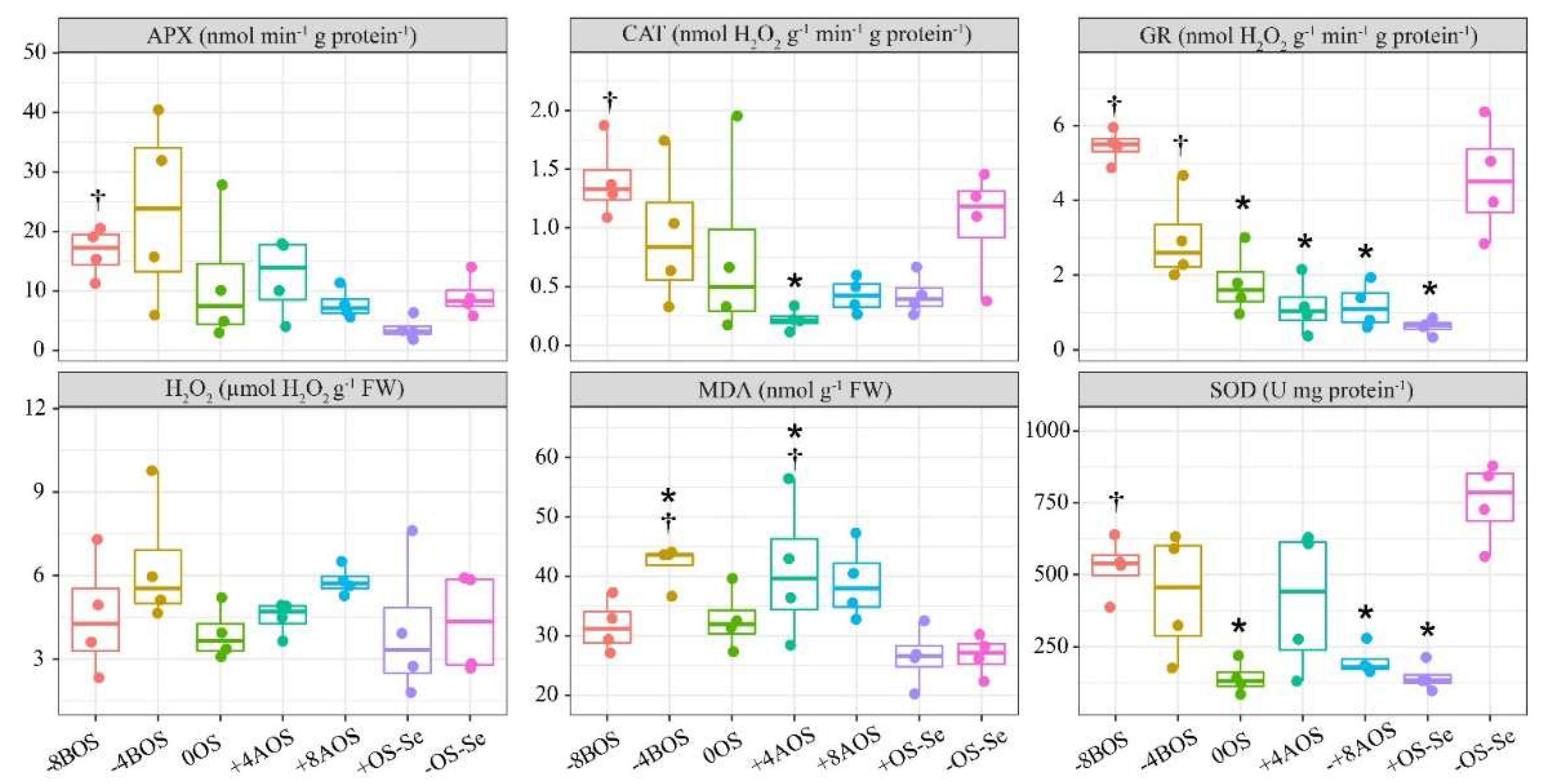
Figure 3.
Total free amino acids (AA), proline (Pro), carbohydrates, and protein (Prt) as a result of Se application in Coffea arabica cv. Catuaí seedlings under osmotic stress induced by PEG-6000. The values displayed are the distribution of four replicates. Asterisks refer to the significant difference when comparing all treatments with non-stressed plants without Se supply (-OS-Se) (p < 0.05). Dagger refers to the significant difference when comparing all treatments with stressed plants without Se supply (+OS-Se) (p < 0.05). Treatments: -8BOS - Application of Se 8 days before stress (Stressed plants); -4BOS - Application of Se 4 days before stress (Stressed plants); 0OS - Application of Se in stress (Stressed plants); +4AOS - Application of Se 4 days after stress (Stressed plants); +8AOS - Application of Se 8 days after stress (Stressed plants); +OS-Se - Without Se (Stressed plants); -OS-Se - Without Se (Non-stressed plants).
Figure 3.
Total free amino acids (AA), proline (Pro), carbohydrates, and protein (Prt) as a result of Se application in Coffea arabica cv. Catuaí seedlings under osmotic stress induced by PEG-6000. The values displayed are the distribution of four replicates. Asterisks refer to the significant difference when comparing all treatments with non-stressed plants without Se supply (-OS-Se) (p < 0.05). Dagger refers to the significant difference when comparing all treatments with stressed plants without Se supply (+OS-Se) (p < 0.05). Treatments: -8BOS - Application of Se 8 days before stress (Stressed plants); -4BOS - Application of Se 4 days before stress (Stressed plants); 0OS - Application of Se in stress (Stressed plants); +4AOS - Application of Se 4 days after stress (Stressed plants); +8AOS - Application of Se 8 days after stress (Stressed plants); +OS-Se - Without Se (Stressed plants); -OS-Se - Without Se (Non-stressed plants).
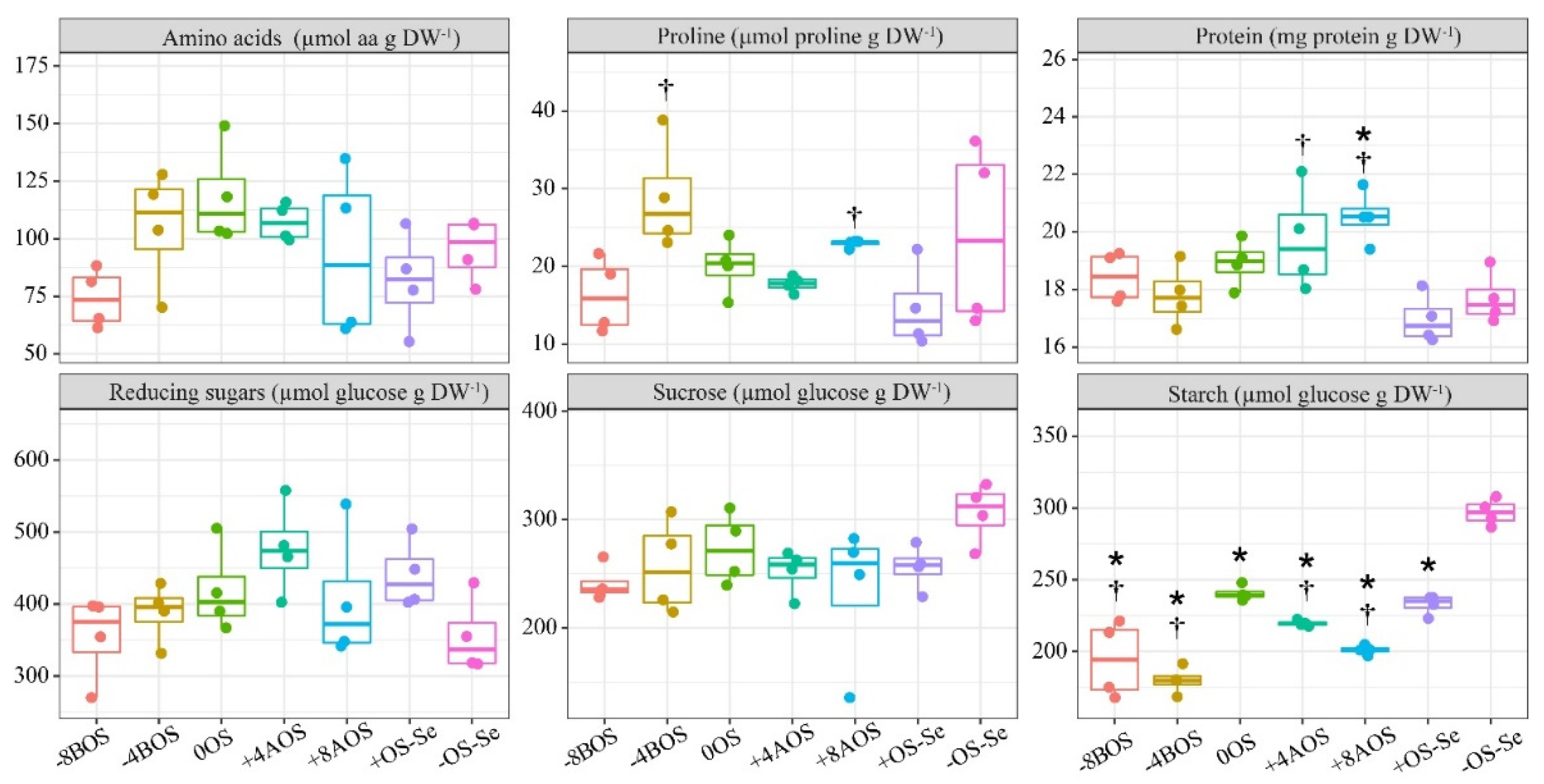
Figure 4.
Principal component analysis (PCA) of leaf compounds and Se content in leaves. The leaf attributes included were Leaf Se content (Se); Ascorbate peroxidase (APX); Superoxide Dismutase (SOD), Catalase (CAT), Glutathione Reductase (GR); Proline; Protein; Lipid Peroxidation (MDA), and Starch. Arrows represent the contribution of leaf compounds on principal component axes. Treatments: -8BOS - Application of Se 8 days before stress (Stressed plants); -4BOS - Application of Se 4 days before stress (Stressed plants); 0OS - Application of Se in stress (Stressed plants); +4AOS - Application of Se 4 days after stress (Stressed plants); +8AOS - Application of Se 8 days after stress (Stressed plants); +OS-Se - Without Se (Stressed plants); -OS-Se - Without Se (Non-stressed plants).
Figure 4.
Principal component analysis (PCA) of leaf compounds and Se content in leaves. The leaf attributes included were Leaf Se content (Se); Ascorbate peroxidase (APX); Superoxide Dismutase (SOD), Catalase (CAT), Glutathione Reductase (GR); Proline; Protein; Lipid Peroxidation (MDA), and Starch. Arrows represent the contribution of leaf compounds on principal component axes. Treatments: -8BOS - Application of Se 8 days before stress (Stressed plants); -4BOS - Application of Se 4 days before stress (Stressed plants); 0OS - Application of Se in stress (Stressed plants); +4AOS - Application of Se 4 days after stress (Stressed plants); +8AOS - Application of Se 8 days after stress (Stressed plants); +OS-Se - Without Se (Stressed plants); -OS-Se - Without Se (Non-stressed plants).
Figure 5.
Coffea arabica cv. Catuaí seedlings under osmotic stress induced by PEG-6000 and Se foliar application. Treatments: -8BOS - Application of Se 8 days before stress (Stressed plants); -4BOS - Application of Se 4 days before stress (Stressed plants); 0OS - Application of Se in stress (Stressed plants); +4AOS - Application of Se 4 days after stress (Stressed plants); +8AOS - Application of Se 8 days after stress (Stressed plants); +OS-Se - Without Se (Stressed plants); -OS-Se - Without Se (Non-stressed plants).
Figure 5.
Coffea arabica cv. Catuaí seedlings under osmotic stress induced by PEG-6000 and Se foliar application. Treatments: -8BOS - Application of Se 8 days before stress (Stressed plants); -4BOS - Application of Se 4 days before stress (Stressed plants); 0OS - Application of Se in stress (Stressed plants); +4AOS - Application of Se 4 days after stress (Stressed plants); +8AOS - Application of Se 8 days after stress (Stressed plants); +OS-Se - Without Se (Stressed plants); -OS-Se - Without Se (Non-stressed plants).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



