
Submitted:
14 July 2023
Posted:
17 July 2023
You are already at the latest version
Abstract
Neuropsychiatric disorders are globally public-health concern, in which diagnosis might be based on symptoms that often vary across individuals. Schizophrenia is one of the major neuropsychiatric disorders, which may affect millions worldwide. Detection of predictive biomarkers is the most intensively developing approach for the disease. However, the biochemical alterations have not been comprehensively distinguished up to the present time. In addition, there is less confidence of finding a specific biomarker for neuropsychiatric disorders including schizophrenia, but rather a specific characteristic behavioral pattern. Maternal immune activation has been considered to be one of the important factors for the development of neuropsychiatric disorders. Here, the mouse model of neuropsychiatric disorders has been built, in which poly-I:C, sodium dextran sulfate (DSS) and κ-carrageenan (CGN) were used for the maternal immune activation during the pregnancy of individuals. Subsequently, we challenged to link some of biochemical changes of p62 and GLAST in the offspring mice brain to the alteration of several pathological behaviors. Consequently, it has been shown that autophagy might be involved in the development of neuropsychiatric disorders including schizophrenia. To our knowledge, this is the first report of the significant correlation between pathological behavior and the biochemical alterations in neuropsychiatric disorder model animals. The relative ease of conducting these evaluation-tasks would make them useful for testing novel therapeutics designed to ameliorate the symptoms of several psychiatric disorders.
Keywords:
neuropsychiatric disorder
; schizophrenia
; behavioral test
; poly I:C
; sodium dextran sulfate
; p62
; GLAST
1. Introduction
Neuropsychiatric disorders are globally public-health concern [1]. Schizophrenia is one of the well-known neuropsychiatric disorders with the most severe disabilities that may affect approximately 1% of the human population, which is also complicated by comorbid drug use problems [2]. Schizophrenia is frequently characterized by negative, positive, and cognitive symptoms [3]. Interestingly, Schizophrenia is 1.4 times more frequently diagnosed in males than females [4]. Male patients show an earlier age of onset, decreased social functioning, and worsened negative but reduced depressive symptoms as compared with females [5]. However, sex differences in cellular and molecular mechanisms remain mostly undetermined. A leading theory about the etiology of schizophrenia is the dopaminergic hypothesis, which suggests that this disorder might be caused by dopamine imbalance, which is supported by the beneficial therapeutic effects of anti-dopaminergic drugs [6]. The hypofunction hypothesis of N-methyl-d-aspartate (NMDA) receptor may suggest a complementary elucidation of the etiology of schizophrenia [7]. Patients with schizophrenia may show a reduced expression of NMDA receptor in the prefrontal cortex [8], which seems to be also related to the imbalance of dopamine levels [9].
It has been shown that maternal immune activation is a potential risk factor for schizophrenia [10]. Therefore, maternal immune activation with the treatment of viral RNA imitator, polyriboinosinic-polyribocytidilic acid (poly I:C), during prenatal development could provide as a broadly employed animal model to make behavioral alterations reminiscent of schizophrenia [11].Hence interestingly, behavioral neurological alterations in the Poly I:C model could be prevented if some interventions are performed prior to the symptom manifestation [12,13]. For the disease evaluation of schizophrenia, several behavioral examinations such as the modified forced swim test might be regarded as appropriate tests for the animal model of schizophrenia, which may be thought to imitate psychiatric symptoms [14]. Therefore, the Poly I:C rodent model may be a neurodevelopmental paradigm of neuropsychiatric disorders based on prenatal exposure to a virus-imitating element such as poly I:C, which might lead to a development of schizophrenia-like behaviors in the offspring comparable to those designated in patients [15]. In addition, several behavioral examinations given the impairments in neuropsychiatric disorders have recorded in the offspring treated with polyI:C [16,17], suggesting that patients with a neuropsychiatric disorder might exhibit alterations in multisensory integration [18]. Therefore, prenatal inflammation has been also deliberated as a risk factor for neuropsychiatric disorders [19]. It may be of interest whether intake of some drugs with anti-oxidative and anti-inflammatory properties could neutralize the later development of neuropsychiatric disorder-related consequences. For example, N-acetylcysteine (NAC) might be suggested as a supplement for high-risk pregnancies, as it may considerably improve neonatal consequences [20]. Taken together, the polyI:C model animal might be useful for studying the developmental factors that may alter multisensory integration in neuropsychiatric disorders [21].
At molecular levels in brain, it has been shown that autophagic dysregulation of p62 might potentially lead to cognitive impairment across brain conditions [22]. In addition, the p62 protein expression may be upregulated in cultured neurons isolated from materials with schizophrenia [23], and in brain samples from a mouse model of schizophrenia [24]. Therefore, controlling the p62 protein levels may provide a potential target for therapeutic intervention against symptoms of schizophrenia [22]. Interestingly, Di-(2-ethylhexyl) phthalate (DEHP) could simultaneously increase the number of autophagosomes and the levels of autophagy marker p62, which is known to impair testicular functions and reproduction [25]. DEHP has the ability to cross the blood-placenta and blood-brain barriers, which could increase the proliferation of astrocytes [26]. Abnormalities in regulatory components of the glutamate system could be also important risk factors for schizophrenia. It has been suggested that the dysregulation of glutamate aspartate transporter (GLAST) may also play a significant role in the neuropathogenesis of various neurological disorders including epilepsy, autism and schizophrenia [27]. Increased incidence of a rare genetic variant in the human gene encoding GLAST has been also detected in schizophrenics [28]. In addition, it has been shown that GLAST knockout (KO) mice may display exaggerated locomotor activity in response to the administration of NMDA antagonist [29]. GLAST KO could exhibit phenotypic abnormalities thought to positive symptoms of schizophrenia [30].
Detection of predictive biomarkers is the most intensively developing approach for the disease. Although there is a need for an accurate clinical examination for the diagnosis of neuropsychiatric disorders including schizophrenia at an early stage, the biochemical changes have not been comprehensively characterized up to the present time. In addition, there is less confidence of finding a specific marker for neuropsychiatric disorders including schizophrenia, but rather a specific characteristic behavioral pattern. In the present study, therefore, we challenged to link some of neurobiochemical changes of p62 and GLAST in the poly I:C induced mice model underlying neuropsychiatric disorders to the alteration of several behaviors with the disorder. From results of numerous preliminary experiments with various behavioral examinations and/or diverse scorings, we have selected and utilized the shown original behavioral examinations and Western blotting techniques to investigate the levels of probable relevant behaviors and protein levels observed in those animals for the recognizable relationships.
2. Materials and Methods
2.1. Mice
Male and Female ICR mice (4-week-old) were purchased from Charles River Laboratories Japan, Inc. (Kanagawa, Japan). At the age of 6 weeks after acclimation, female ICR mice were mated with male ICR mice, and the day when the vaginal plug was confirmed was defined as the first day of pregnancy (GD1). The mother mice received DSS + CGN water which was dissolved in 0.5% (w/v) Sodium Dextran Sulfate 5000 (DSS) and 0.2% (w/v) κ-Carrageenan (CGN) in GD 8 to 11. And 5 mg/kg B.W. Poly I: C was administered intraperitoneally on GD10. The day mice were born was defined as PD1, and on day 22 (PD22), mice were separated from their mothers and grouped in net cages separately for males and females. From PD 125, mice were given 2 mg/L DEHP, and three types of behavioral tests: a descent step test, a modified three chambers test, and a light/dark room test were done eight times. After the behavioral tests, some of the mice were dissected, and collected liver, kidney, and brain.
The mice were housed in an environmentally controlled room, at approximately 20°C and 60% humidity with a 12-h light/dark cycle (lights on at 07:00 and off at 19:00). The care and treatment of the experimental animals conformed with the guidelines for the ethical treatment of laboratory animals established by Nara Women’s University (Nara, Japan) (Approval No. 19-02).
2.2. Materials
Poly I: C, DEHP, and Dextran sulfate sodium (DSS, MW5000) were purchased from Fujifilm Wako Pure Chemical Corporation (Osaka, Japan). κ-Carrageenan (CGN) was obtained from Tokyo Chemical Industry Co., LTD. (Tokyo, Japan). Poly I: C was dissolved in saline. DEHP, DSS, and CGN were diluted with sterile water for mice to drink.
2.3. Behavioral Tests
The score of behavioral tests is shown in Table 1.
- (1)
- Descent step test
A box (23 cm x 31 cm x 8.3 cm) was placed in the center of a plastic case (30 cm x 52 cm x 17 cm), and each mouse was placed on the box. After that, we measured whether or not the mouse descended from the box within one minute (Figure 2A).
- (2)
- Modified three chambers test
The field of the plastic case was divided into three compartments with boxes, and the central compartment was dark (27.5 cm x 21 cm). At first, each mouse was placed in a narrow bright place (30 cm × 13 cm), and chambers were set as (I), (II), and (III) in order of proximity. The size of the dark entrance was 4 cm x 2.5 cm. The score was determined by which position you were in 25 seconds after the start (Figure 2B).
- (3)
- Light/dark room test
A dark space (27.5 cm × 21 cm) and a light space (30 cm × 31 cm) were set up in a box in a field of a plastic case, and the boundary between the light space and the dark space was used as the starting point. The size of the dark entrance was 4 cm x 2.5 cm. Mice were allowed to explore freely for 2 minutes, and the time spent in the light was measured (Figure 2C).
2.4. Western Blotting
To extract protein, the whole brain was homogenized with RIPA buffer. The homogenates were centrifuged to obtain supernatants (Tabletop micro-cooled centrifuge Model3500). The supernatants were mixed with sample buffer and adjusted 1mg/mL protein concentration. We used SDS-PAGE to separate proteins and transfer them to membranes (Immobilon-P, Merck KGaA, Darmstadt, Germany). These were blocked with 3% skim milk and then reacted with primary antibodies SQSTM1/p62 polyclonal antibody (Cosmobio) or GLAST polyclonal antibody (Cosmobio) at 1h and peroxidase-conjugated goat anti-rabbit secondary antibodies (Cell Signaling) at 1h. Proteins were detected by ImageQuant LAS500 (GE Healthcare Japan Com., Tokyo, Japan). Each detected band was quantified by ImageJ, and the relative ratio of protein expression was analyzed using GAPDH (Glyceraldehyde 3-phosphate dehydrogenase, FUJIFILM Wako Pure Chemicals Co.) as an internal control protein. The intensities of the detected bands were calculated using ImageJ software.
2.5. Statistical Analyses
Animal data are expressed as the mean ± standard error (SE). All data were analyzed by Pearson’s correlation analysis. P < 0.05 was considered a statistically significant difference. All statistical analyses were performed using GraphPad Prism version 5.0 (GraphPad Software, Inc., San Diego, CA, USA).
3. Results
Overview of the present study design is shown in Figure 1. To facilitate the development of neuropathological condition in offspring, two modifications have been employed in addition to the basic poly I:C method for creating mice with schizophrenia-like behaviors. First, per oral administration of low molecular weight sodium dextran sulfate (DSS) and κ-Carrageenan (CGN) was added to the pregnant mice for augmenting the systemic inflammation levels [31]. Second, the offspring was also forced to drink DEHP in water, which might be related to the augmented oxidative stress and the cause of neurobehavioral disorders [32]. However, with the present method of producing schizophrenia model mice, it was difficult to make all mice be psychiatric. The offspring whose mothers were treated with poly I:C, but without DSS and/or DEHP, were expected to perform similarly to control mice on the tests, which actually resulted in almost no difference (data not shown, personal communication). And no method has been established to determine whether they are affected by neuropathological disorders or not. Actually, behavioral tests revealed that some adult male offspring mice frequently had the alteration of behaviors in this study. (Figure 2) The behavioral tests used in the present study were created and modified from several literatures as references [33]. As shown at Table 1, the psychological behavior index (PBI) score was expressed with a sum scores of the three behavioral examinations. The PBI scores were tracked and documented 8 times at the end period in the trial. (Figure 1) The average scores during the period and the last score were shown in Figure 3. Note that zero or very low value of the PBI score were found in several normal (untreated) offspring (n>10) and their parents (data not shown, personal communication).
Figure 1.
Study design. Schematic representation of the Study design is shown. Female ICR mice were mated with male ICR mice, and the day when the vaginal plug was confirmed was defined as the first day of pregnancy (GD1). The mother mice received DSS + CGN water in GD 8 to 11. And 5 mg/kg B.W. Poly I: C was administered intraperitoneally on GD10. The day of mice were born was defined as PD1, and on day 22 (PD22), the pup mice were separated from their mothers. From PD 125, pup mice were given 2 mg/L DEHP, and from PD 215, mice were conducted for eight times of behavioral test (The day of behavioral tests shows a black and white arrow. White arrow shows the time point of the last behavioral test). DSS; Dextran sodium sulfate, CGN; κ-Carrageenan.
Figure 1.
Study design. Schematic representation of the Study design is shown. Female ICR mice were mated with male ICR mice, and the day when the vaginal plug was confirmed was defined as the first day of pregnancy (GD1). The mother mice received DSS + CGN water in GD 8 to 11. And 5 mg/kg B.W. Poly I: C was administered intraperitoneally on GD10. The day of mice were born was defined as PD1, and on day 22 (PD22), the pup mice were separated from their mothers. From PD 125, pup mice were given 2 mg/L DEHP, and from PD 215, mice were conducted for eight times of behavioral test (The day of behavioral tests shows a black and white arrow. White arrow shows the time point of the last behavioral test). DSS; Dextran sodium sulfate, CGN; κ-Carrageenan.
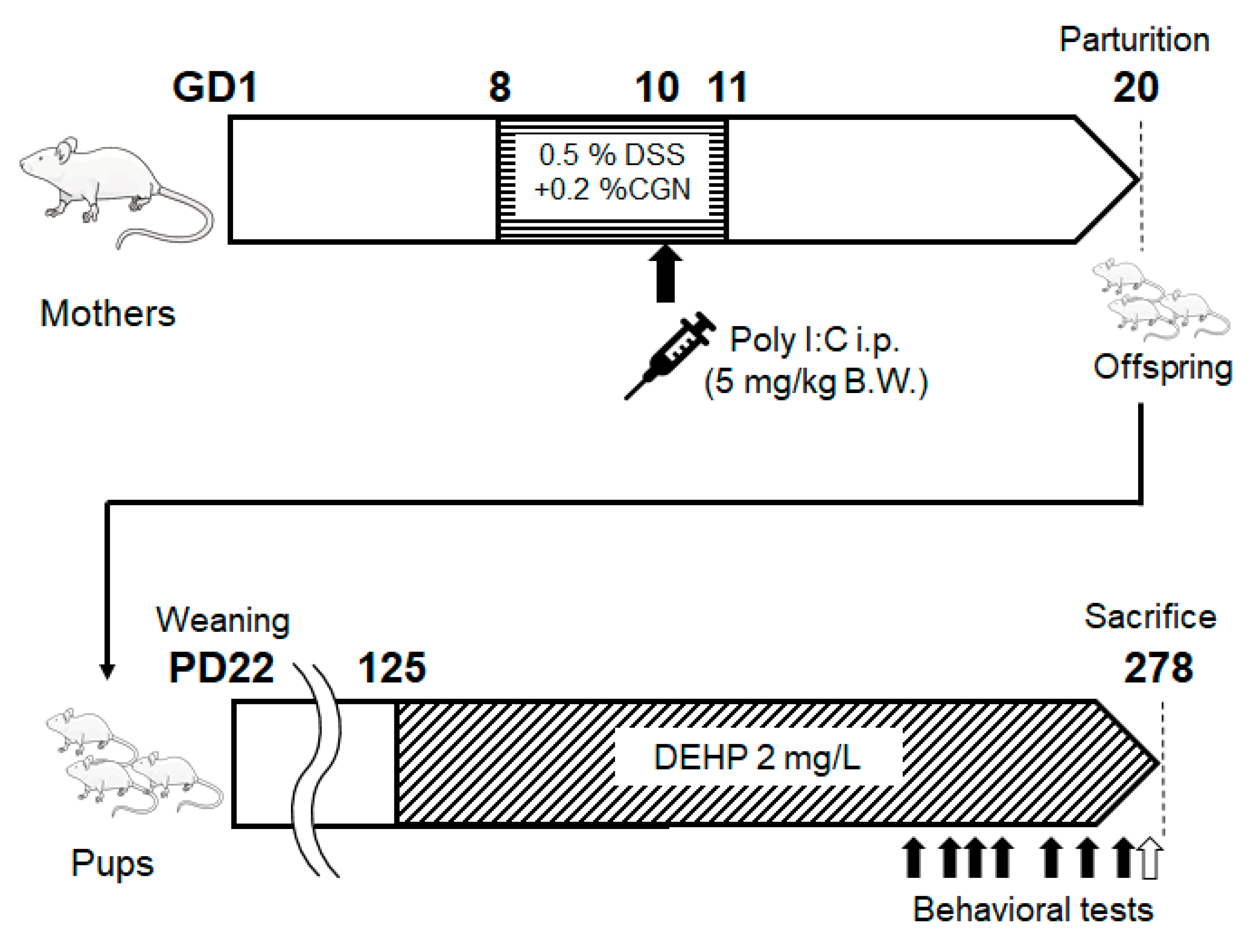
Figure 2.
Behavioral tests. [A] The image of the descent step test. The mouse placed on the box and be measured whether or not the mice descended from the box within one minute. [B] The image of the modified three chambers test. At first, the mouse was placed in (I), and we measured which chamber the mouse was inside after 25 seconds. [C] The image of the light/dark room test. Mice were allowed to explore freely for 2 minutes, and we measured the time spent in the light area.
Figure 2.
Behavioral tests. [A] The image of the descent step test. The mouse placed on the box and be measured whether or not the mice descended from the box within one minute. [B] The image of the modified three chambers test. At first, the mouse was placed in (I), and we measured which chamber the mouse was inside after 25 seconds. [C] The image of the light/dark room test. Mice were allowed to explore freely for 2 minutes, and we measured the time spent in the light area.
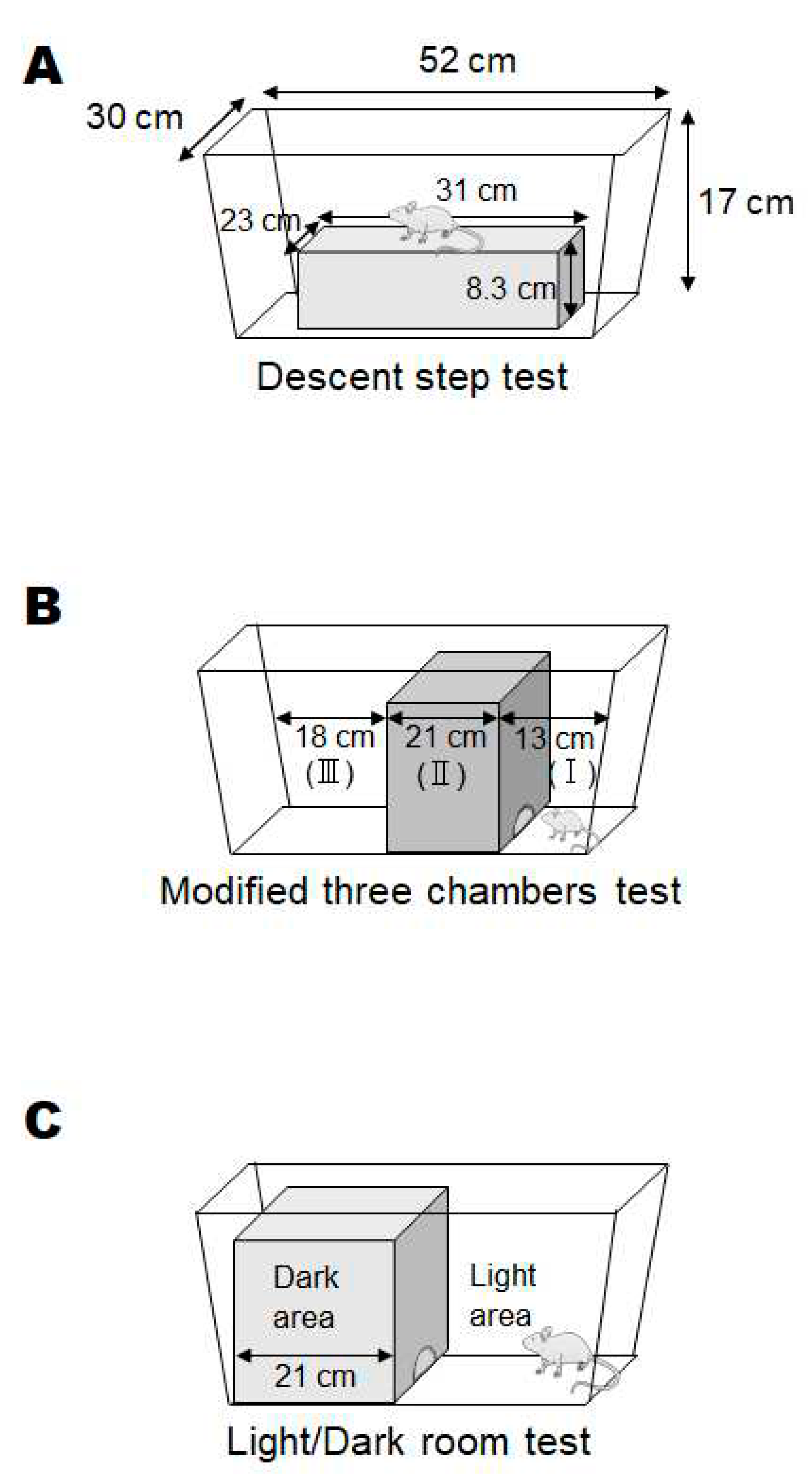
Figure 3.
The results of behavioral tests. The total behavioral tests score (PBI score) of each mouse. The black bar shows the mean value of 8 times behavioral test scores during the trial. White-right upper diagonal bar shows the last behavioral test score before dissection of mice.
Figure 3.
The results of behavioral tests. The total behavioral tests score (PBI score) of each mouse. The black bar shows the mean value of 8 times behavioral test scores during the trial. White-right upper diagonal bar shows the last behavioral test score before dissection of mice.
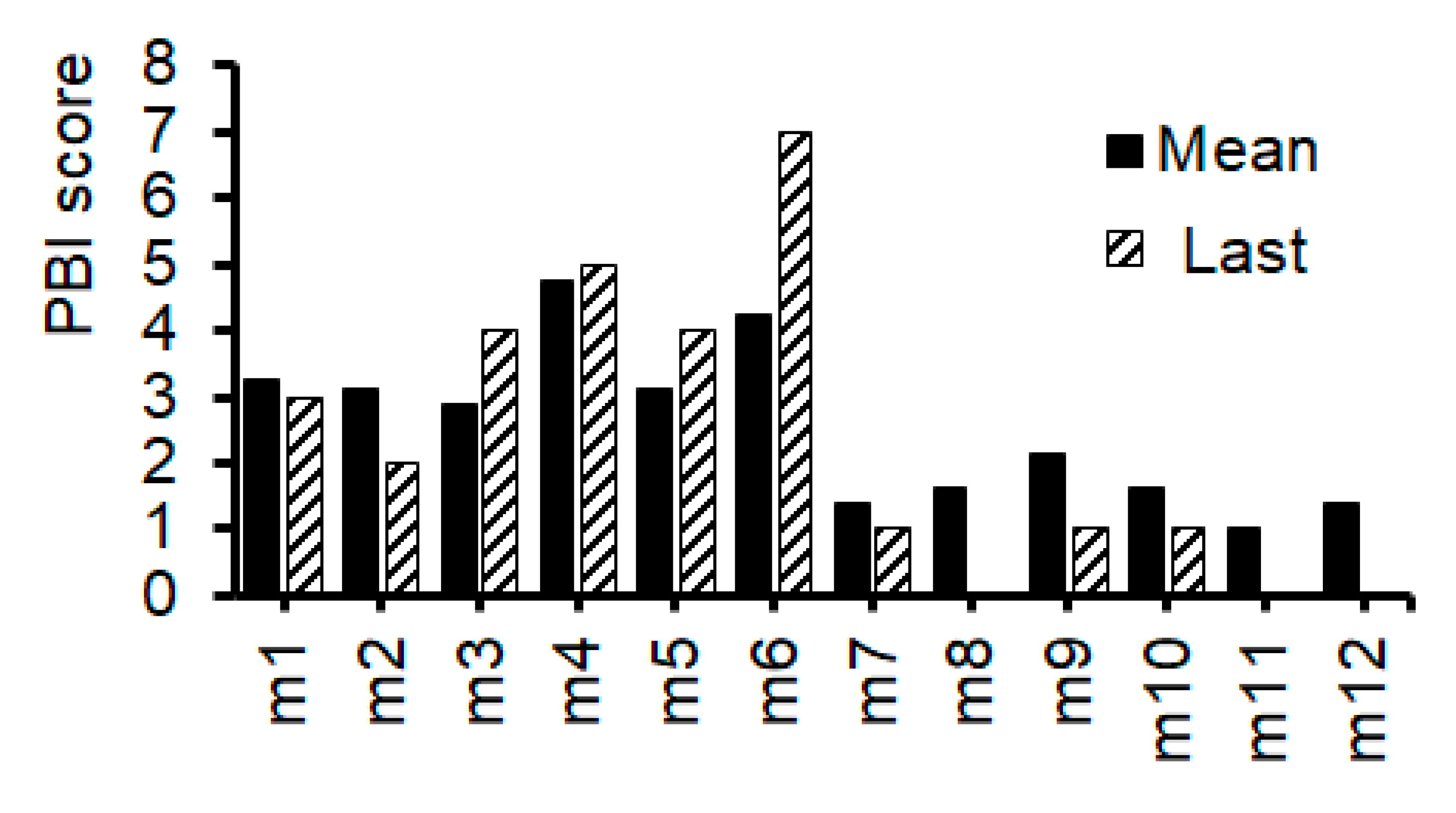
After documented the last PBI scores, all mice were sacrificed and examined for protein expressions in their whole brain. The protein expression levels of p62, GLAST, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) in each mouse were shown in Figure 4A–C). There was a faint, but not significant, relationship between p62 protein expression and GLAST protein expression (R=0.484, P=0.111). (Figure 4D) Then, the correlation between PBI score and p62 protein expression as well as the correlation between PBI score and GLAST protein expression have been also evaluated. (Figure 5) As shown in Figure 6, there were significant correlations both between the last PBI score and p62 protein expression, and between the last PBI score and GLAST protein expression. (Figure 6A,B) The correlation between the last PBI score and p62 protein expression (R=0.82, P=0.001) was more significant rather than the correlation between the last PBI score and GLAST protein expression (R=0.58, P=0.047). (Figure 6A,B) Interestingly, the correlation between the last PBI score and p62 protein expression (R=0.82, P=0.001) was more significant rather than the correlation between mean PBI score and p62 protein expression (R=0.63, P=0.028). (Figure 6A,C) In contrast, the correlation between the mean score and GLAST protein expression (R=0.60, P=0.039) was almost similar, but a little bit more significant, compared to the correlation between the last PBI score and GLAST protein expression (R=0.58, P=0.047). (Figure 6B,D).
4. Discussion
One of the goal in this study was effectively to produce mouse model of neuropsychiatric disorders after exposure to maternal immune activation via the viral nucleic acid imitators including the poly I:C. Almost consistent with earlier reports, we observed that acute poly I:C administered at PD10 plus DSS/CGN supply could elicit significant alteration of behaviors in our original tests for adult offspring animals. The behavioral experiments revealed an array of long-term changes in the offspring following poly I:C, DSS/CGN, and DEHP treatments. Some studies have also proved object recognition deficits in mice prenatally exposed to the similar poly I:C and lipopolysaccharide (LPS) treatment [34,35]. Effects of these poly I:C treatments on the alteration of behaviors observed in the present study had a tendency to increase difference to male mice, and female mice showed reduced discrimination compared to the males in these behavioral tests (unpublished data, personal communication). However, no differences were observed in the other data set. Some discrepancies in details between the present results and those of others may reflect some differences in the tests and/or specific prenatal treatment. Interestingly, sex differences in the effects of prenatal infection for cognition have been also described for fear conditioning in rodents [36,37]. In addition, previous reports suggest that phase of the estrous cycle of female rodent animals may influence object location recognition with upper levels of estrogen and/or progesterone activity [38].
In neurons derived from schizophrenia patients, the sensitivity to PI3K/AKT/GSK3 signaling might be changed [39], which may be involved in the development of schizophrenia [40]. In addition, the AKT activity has been shown to decrease in certain brain regions of patients with major depressive disorder and/or schizophrenia [41]. Interestingly, anti-inflammatory pregnenolone has been indicated to control schizophrenia-like behaviors via the modulation of AKT signaling [42]. It has been shown that the PI3K/AKT signaling pathway might be involved in autophagy [43], therefore, in which autophagy might be also involved in the development of schizophrenia [44]. Autophagy is a membrane trafficking machinery responsible for degrading damaged proteins, lipids, and/or organelles in the lysosome [45]. Remarkably, neuronal autophagy may be related to the cognitive processes via the regulation of synaptic components [23,46]. In addition, the intracellular autophagic activity could control the p62 protein levels [47]. It has been proposed that elevated p62 levels may have functional consequences on the neurotransmission, which might explain the behavioral changes relevant to schizophrenia [23,48]. Fortunately, it has been shown that protein expression levels of p62 seem to be significantly correlated with the score of our behavioral tests. Aberrations in regulatory components of the glutamatergic system could be also important risk factors for schizophrenia, which might be regulated by a family of glutamate transporters including GLAST or excitatory amino-acid transporter 1. Incidence of a genetic variant may be increased in the human gene encoding GLAST within schizophrenia patients [28,49]. Interestingly, some genetic variants may impair metabolic functions of astrocytes and might lead to cognitive dysfunction [49]. In addition, it has been revealed that the GLAST knockout (KO) mice could exhibit exaggerated locomotor activity [28,50], which may be a model for positive symptoms of schizophrenia. Therefore, roles of GLAST protein might be involved in the certain behavior relevant to the symptoms of schizophrenia [30,49,51]. Fortunately again, expression levels of GLAST protein seem to be also correlated with the score of our behavioral tests. These data provide the first demonstration of the correlations between behavioral score and the biochemical alterations of p62 and/or GLAST expression as a result of prenatal immune challenge, which may support the hypothesis that prenatal infection disrupts processing within prefrontal cortex consistent with findings regarding the pathophysiology of neuropsychiatric disorders including schizophrenia and/or autism [52,53].
Over again, the present experiments demonstrate that prenatal immune challenge results in behavioral alterations in mice similar to those reported in patients with neurodevelopmental psychiatric disorders such as schizophrenia. Additionally, the most common pharmacological strategy to elicit schizophrenia-like behaviors might be based on the inhibition of N-methyl-D-aspartate (NMDA) receptor [54]. A non-competitive NMDA receptor antagonist could mimic the hypofunction of the NMDA receptor, which might lead to behavioral effects reminiscent of the schizophrenia [55]. The predictive rationality of that model has been well known by the effective administration of atypical antipsychotic drugs. No other drugs such as antidepressants could inverse the behavioral changes of that model. Whatever the neuropsychiatric disorder is, the approach presented here would make it easy and/or cost effective to determine whether some preventive materials including the use of antipsychotics, hormonal agents, and/or anti-inflammatory drugs should be applied to the adolescent poly I:C animals presented here [56,57]. Continuous and alive evaluation might be possible for the evaluation of therapeutic intervention to the model animal of neuropsychiatric disorders.
The clinical symptom of neuropathological disorders might be intricate. Our results are limited to an individual model animal, in which the phenotype may be partially present in the poly I:C treated mice offspring. Even though the poly I:C model being well-validated, there are presently no animal model which completely mimic a human neuropathological disorder. Therefore, the validity of the behavioral score should be re-evaluated with the model mice with the treatment of NMDA receptor inhibitors. Response to the administration of atypical antipsychotic drugs might be also mandatory. The communication between neurons and glial cells is also important for the appropriate function in brain. Therefore, histopathological study might be helpful for the elucidation of roles with the communication in distinct brain areas. In addition, the larger sample size of experiments might lead to more significant outcomes. As the results of this study have been attained by using a mouse model, the generalization of our findings to humans should be cautiously assessed. Studies on the mechanism of action related to the prevention of schizophrenia-like outcomes in the offspring are also necessary. In this regards, we presume that neuropsychiatric disorders might be categorized in immune related diseases [58], whose pathogenesis may be based on the engram memory system [59,60,61]. Forthcoming studies using a larger number of animals to address the above concerns and/or concepts would be informative. In addition, further research will be required to identify the distinctions and commonalities between the neuropathological mechanisms underlying the behavioral outcomes of prenatal immune activation.
5. Conclusions
Taken together, the mouse model of neuropsychiatric disorders has been built, in which poly-I:C, DSS, and CGN were used for maternal individual during the pregnancy. In addition, the significant correlations between the behavioral test score and the protein expression levels of p62 and GLAST in the whole brain of offspring mice have been detected. For the evaluation of neuropsychiatric disorder-like behaviors, original three behavioral tests have been devised. In conclusion, autophagy regulation might be involved in the occurrence and/or the development of these neuropsychiatric disorders. These findings suggests that certain behavioral tests could be effective for determining some of the brain neuropathological disorder. However, it needs for further investigation to fully comprehend the molecular mechanisms involved.
Author Contributions
Conceptualization, YI, and SM; original draft preparation and editing, YI, AT and SM; visualization, YI, AT and SM; experiment execution, YI, and SM; supervision, SM. Each author (YI, AT and SM) has participated sufficiently in this work of drafting the article and/or revising the article for the important rational content. Then, all authors gave final approval of the version to be submitted. Finally, all authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable
Data Availability Statement
Not applicable.
Competing Interests Statement
The authors declare that they have no competing financial interests.
Abbreviations
| CGN | carrageenan |
| CNS | central nervous system |
| DEHP | 2-ethylhexyl phthalate |
| DSS | sodium dextran sulfate |
| FMT | fecal microbiota transplantation |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| GLAST | glutamate aspartate transporter |
| KO | knockout |
| LPS | lipopolysaccharide |
| mRNA | messenger RNA |
| NAC | N-acetylcysteine |
| NMDA | N-methyl-d-aspartate |
| PBI | psychological behavior index |
| poly I:C | polyriboinosinic-polyribocytidilic acid |
| QOL | quality of life |
References
- Singh, M.; Agarwal, V.; Jindal, D.; Pancham, P.; Agarwal, S.; Mani, S.; Tiwari, R.K.; Das, K.; Alghamdi, B.S.; Abujamel, T.S.; et al. Recent Updates on Corticosteroid-Induced Neuropsychiatric Disorders and Theranostic Advancements through Gene Editing Tools. Diagnostics 2023, 13, 337. [Google Scholar] [CrossRef]
- Khokhar, J.Y.; Dwiel, L.L.; Henricks, A.M.; Doucette, W.T.; Green, A.I. The link between schizophrenia and substance use disorder: A unifying hypothesis. Schizophr. Res. 2017, 194, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Owen, MJ.; Sawa, A.; Mortensen, PB. Schizophrenia. Lancet 2016, 388, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Picchioni, MM.; Murray, RM. Schizophrenia. BMJ 2007, 335, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Abel, K.M.; Drake, R.; Goldstein, J.M. Sex differences in schizophrenia. Int. Rev. Psychiatry 2010, 22, 417–428. [Google Scholar] [CrossRef]
- Ban, T.A. Fifty years chlorpromazine: a historical perspective. Neuropsychiatr. Dis. Treat. 2007, 3, 495–500. [Google Scholar] [CrossRef]
- Adler, C.; Goldberg, T.; Malhotra, A.; Pickar, D.; Breier, A. Effects of Ketamine on Thought Disorder, Working Memory, and Semantic Memory in Healthy Volunteers. Biol. Psychiatry 1998, 43, 811–816. [Google Scholar] [CrossRef]
- Weickert, C.S.; Fung, S.J.; Catts, V.S.; Schofield, P.R.; Allen, K.M.; Moore, L.T.; Newell, K.A.; Pellen, D.; Huang, X.-F.; Catts, S.V.; et al. Molecular evidence of N-methyl-D-aspartate receptor hypofunction in schizophrenia. Mol. Psychiatry 2013, 18, 1185–1192. [Google Scholar] [CrossRef]
- Del, Arco, A.; Segovia, G.; Mora, F. Blockade of NMDA receptors in the prefrontal cortex increases dopamine and acetylcholine release in the nucleus accumbens and motor activity. Psychopharmacology (Berl). 2008, 201, 325–338. [Google Scholar] [CrossRef]
- Hanson, K.L.; Grant, S.E.; Funk, L.H.; Schumann, C.M.; Bauman, M.D. Impact of Maternal Immune Activation on Nonhuman Primate Prefrontal Cortex Development: Insights for Schizophrenia. Biol. Psychiatry 2022, 92, 460–469. [Google Scholar] [CrossRef]
- Connor, C.M.; Dincer, A.; Straubhaar, J.; Galler, J.R.; Houston, I.B.; Akbarian, S. Maternal immune activation alters behavior in adult offspring, with subtle changes in the cortical transcriptome and epigenome. Schizophr. Res. 2012, 140, 175–184. [Google Scholar] [CrossRef]
- Garcia-Partida, J.A.; Torres-Sanchez, S.; MacDowell, K.; Fernández-Ponce, M.T.; Casas, L.; Mantell, C.; Soto-Montenegro, M.L.; Romero-Miguel, D.; Lamanna-Rama, N.; Leza, J.C.; et al. The effects of mango leaf extract during adolescence and adulthood in a rat model of schizophrenia. Front. Pharmacol. 2022, 13, 886514. [Google Scholar] [CrossRef]
- Romero-Miguel, D.; Casquero-Veiga, M.; MacDowell, K.S.; Torres-Sanchez, S.; Garcia-Partida, J.A.; Lamanna-Rama, N.; Romero-Miranda, A.; Berrocoso, E.; Leza, J.C.; Desco, M.; et al. A Characterization of the Effects of Minocycline Treatment During Adolescence on Structural, Metabolic, and Oxidative Stress Parameters in a Maternal Immune Stimulation Model of Neurodevelopmental Brain Disorders. Int. J. Neuropsychopharmacol. 2021, 24, 734–748. [Google Scholar] [CrossRef]
- Tanaka, M.; Bohár, Z.; Martos, D.; Telegdy, G.; Vécsei, L. Antidepressant-like effects of kynurenic acid in a modified forced swim test. Pharmacol. Rep. 2020, 72, 449–455. [Google Scholar] [CrossRef]
- Hadar, R.; Soto-Montenegro, M.L.; Götz, T.; Wieske, F.; Sohr, R.; Desco, M.; Hamani, C.; Weiner, I.; Pascau, J.; Winter, C. Using a maternal immune stimulation model of schizophrenia to study behavioral and neurobiological alterations over the developmental course. Schizophr. Res. 2015, 166, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Howland, J.; Cazakoff, B.; Zhang, Y. Altered object-in-place recognition memory, prepulse inhibition, and locomotor activity in the offspring of rats exposed to a viral mimetic during pregnancy. Neuroscience 2011, 201, 184–198. [Google Scholar] [CrossRef]
- Missig, G.; Mokler, E.L.; O Robbins, J.; Alexander, A.J.; McDougle, C.J.; A Carlezon, W. Perinatal Immune Activation Produces Persistent Sleep Alterations and Epileptiform Activity in Male Mice. Neuropsychopharmacology 2017, 43, 482–491. [Google Scholar] [CrossRef]
- Gröhn, C.; Norgren, E.; Eriksson, L. A systematic review of the neural correlates of multisensory integration in schizophrenia. Schizophr. Res. Cogn. 2022, 27, 100219. [Google Scholar] [CrossRef]
- Otero, A.M.; Antonson, A.M. At the crux of maternal immune activation: Viruses, microglia, microbes, and IL-17A. Immunol. Rev. 2022, 311, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, V.; Afsharian, P.; Shahhoseini, M.; Kalantar, S.M.; Moini, A. A Review on Various Uses of N-Acetyl Cysteine. Cell J. 2017, 19, 11–17. [Google Scholar] [CrossRef]
- Stekelenburg, JJ.; Maes, JP.; Van, Gool, AR.; Sitskoorn, M.; Vroomen, J. Deficient multisensory integration in schizophrenia: an event-related potential study. Schizophr Res. 2013, 147, 253–261. [Google Scholar] [CrossRef]
- Tomoda, T.; Sumitomo, A.; Shukla, R.; Hirota-Tsuyada, Y.; Miyachi, H.; Oh, H.; French, L.; Sibille, E. BDNF controls GABAAR trafficking and related cognitive processes via autophagic regulation of p62. Neuropsychopharmacology 2021, 47, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Sumitomo, A.; Yukitake, H.; Hirai, K.; Horike, K.; Ueta, K.; Chung, Y.; Warabi, E.; Yanagawa, T.; Kitaoka, S.; Furuyashiki, T.; et al. Ulk2 controls cortical excitatory–inhibitory balance via autophagic regulation of p62 and GABAA receptor trafficking in pyramidal neurons. Hum. Mol. Genet. 2018, 27, 3165–3176. [Google Scholar] [CrossRef]
- Sumitomo, A.; Horike, K.; Hirai, K.; Butcher, N.; Boot, E.; Sakurai, T.; Nucifora, FC, Jr.; Bassett, AS.; Sawa, A.; Tomoda, T. A mouse model of 22q11.2 deletions: Molecular and behavioral signatures of Parkinson's disease and schizophrenia. Sci Adv. 2018, 4, eaar6637. [Google Scholar] [CrossRef] [PubMed]
- Yi, W.; Xiang-Liang, T.; Yu, Z.; Bin, L.; Lian-Ju, S.; Chun-Lan, L.; Tao, L.; Da-Wei, H.; Sheng-De, W.; Guang-Hui, W. DEHP exposure destroys blood-testis barrier (BTB) integrity of immature testes through excessive ROS-mediated autophagy. Genes Dis. 2018, 5, 263–274. [Google Scholar] [CrossRef]
- Wójtowicz, A.K.; Sitarz-Głownia, A.M.; Wnuk, A.; Kajta, M.; Szychowski, K.A. Involvement of the peroxisome proliferator-activated receptor gamma (Pparγ) and matrix metalloproteinases-2 and -9 (Mmp-2 and -9) in the mechanism of action of di(2-ethylhexyl)phthalate (DEHP) in cultured mouse brain astrocytes and neurons. Toxicol. Vitr. 2023, 92, 105639. [Google Scholar] [CrossRef] [PubMed]
- Pajarillo, E.; Rizor, A.; Lee, J.; Aschner, M.; Lee, E. The role of astrocytic glutamate transporters GLT-1 and GLAST in neurological disorders: Potential targets for neurotherapeutics. Neuropharmacology 2019, 161, 107559–107559. [Google Scholar] [CrossRef]
- Walsh, T.; McClellan, J.M.; McCarthy, S.E.; Addington, A.M.; Pierce, S.B.; Cooper, G.M.; Nord, A.S.; Kusenda, M.; Malhotra, D.; Bhandari, A.; et al. Rare Structural Variants Disrupt Multiple Genes in Neurodevelopmental Pathways in Schizophrenia. Science 2008, 320, 539–543. [Google Scholar] [CrossRef]
- Karlsson, RM.; Tanaka, K.; Heilig, M.; Holmes, A. Loss of glial glutamate and aspartate transporter (excitatory amino acid transporter 1) causes locomotor hyperactivity and exaggerated responses to psychotomimetics: rescue by haloperidol and metabotropic glutamate 2/3 agonist. Biol Psychiatry. 2008, 64, 810–814. [Google Scholar] [CrossRef]
- Karlsson, R.-M.; Tanaka, K.; Saksida, L.M.; Bussey, T.J.; Heilig, M.; Holmes, A. Assessment of Glutamate Transporter GLAST (EAAT1)-Deficient Mice for Phenotypes Relevant to the Negative and Executive/Cognitive Symptoms of Schizophrenia. Neuropsychopharmacology 2009, 34, 1578–1589. [Google Scholar] [CrossRef]
- Ikeda, Y.; Matsuda, S. Gut Protective Effect from D-Methionine or Butyric Acid against DSS and Carrageenan-Induced Ulcerative Colitis. Molecules 2023, 28, 4392. [Google Scholar] [CrossRef]
- Kang, J.S.; Baek, J.H.; Song, M.Y.; Rehman, N.U.; Chung, H.J.; Lee, D.K.; Yoo, D.Y.; Kim, H.J. Long-term exposure changes the environmentally relevant bis(2-ethylhexyl) phthalate to be a neuro-hazardous substance disrupting neural homeostasis in emotional and cognitive functions. Environ. Pollut. 2023, 324, 121387. [Google Scholar] [CrossRef]
- Chamera, K.; Kotarska, K.; Szuster-Głuszczak, M.; Trojan, E.; Skórkowska, A.; Pomierny, B.; Krzyżanowska, W.; Bryniarska, N.; Basta-Kaim, A. The prenatal challenge with lipopolysaccharide and polyinosinic:polycytidylic acid disrupts CX3CL1-CX3CR1 and CD200-CD200R signalling in the brains of male rat offspring: a link to schizophrenia-like behaviours. J. Neuroinflammation 2020, 17, 247. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, K.; Hashimoto, K.; Kishimoto, T.; Shimizu, E.; Ishikura, H.; Iyo, M. Immune Activation During Pregnancy in Mice Leads to Dopaminergic Hyperfunction and Cognitive Impairment in the Offspring: A Neurodevelopmental Animal Model of Schizophrenia. Biol. Psychiatry 2006, 59, 546–554. [Google Scholar] [CrossRef]
- Coyle, P.; Tran, N.; Fung, J.; Summers, B.; Rofe, A. Maternal dietary zinc supplementation prevents aberrant behaviour in an object recognition task in mice offspring exposed to LPS in early pregnancy. Behav. Brain Res. 2009, 197, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Schwendener, S.; Meyer, U.; Feldon, J. Deficient maternal care resulting from immunological stress during pregnancy is associated with a sex-dependent enhancement of conditioned fear in the offspring. J. Neurodev. Disord. 2009, 1, 15–32. [Google Scholar] [CrossRef]
- Bitanihirwe, B.K.; Peleg-Raibstein, D.; Mouttet, F.; Feldon, J.; Meyer, U. Late Prenatal Immune Activation in Mice Leads to Behavioral and Neurochemical Abnormalities Relevant to the Negative Symptoms of Schizophrenia. Neuropsychopharmacology 2010, 35, 2462–2478. [Google Scholar] [CrossRef]
- Sutcliffe, J.; Marshall, K.; Neill, J. Influence of gender on working and spatial memory in the novel object recognition task in the rat. Behav. Brain Res. 2007, 177, 117–125. [Google Scholar] [CrossRef]
- Stertz, L.; Di, Re, J. ; Pei, G.; Fries, GR.; Mendez, E.; Li, S.; Smith-Callahan, L.; Raventos, H.; Tipo, J.; Cherukuru, R.; et al. Convergent genomic and pharmacological evidence of PI3K/GSK3 signaling alterations in neurons from schizophrenia patients. Neuropsychopharmacology 2021, 46, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Emamian, ES.; Hall, D.; Birnbaum, MJ.; Karayiorgou, M.; Gogos, JA. Convergent evidence for impaired AKT1-GSK3beta signaling in schizophrenia. Nat Genet. 2004, 36, 131–137. [Google Scholar] [CrossRef]
- Emamian, E.S. AKT/GSK3 signaling pathway and schizophrenia. Front. Mol. Neurosci. 2012, 5, 33. [Google Scholar] [CrossRef]
- Karege, F.; Perroud, N.; Burkhardt, S.; Schwald, M.; Ballmann, E.; La, Harpe, R.; Malafosse, A. Alteration in kinase activity but not in protein levels of protein kinase B and glycogen synthase kinase-3beta in ventral prefrontal cortex of depressed suicide victims. Biol Psychiatry. 2007, 61, 240–245. [Google Scholar] [CrossRef]
- Qu, L.; Liu, Y.; Deng, J.; Ma, X.; Fan, D. Ginsenoside Rk3 is a novel PI3K/AKT-targeting therapeutics agent that regulates autophagy and apoptosis in hepatocellular carcinoma. J. Pharm. Anal. 2023, 13, 463–482. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.S.; Lee, S.M.; Hwang, D.; Park, H.J.; Kim, J.W. Association between Unc-51-like autophagy activating kinase 2 gene polymorphisms and schizophrenia in the Korean population. Medicine 2022, 101, e28745. [Google Scholar] [CrossRef]
- Mizushima, N.; Yoshimori, T.; Ohsumi, Y. The Role of Atg Proteins in Autophagosome Formation. Annu. Rev. Cell Dev. Biol. 2011, 27, 107–132. [Google Scholar] [CrossRef]
- Sumitomo, A.; Tomoda, T. Autophagy in neuronal physiology and disease. Curr. Opin. Pharmacol. 2021, 60, 133–140. [Google Scholar] [CrossRef]
- Lippai, M.; Lőw, P. The Role of the Selective Adaptor p62 and Ubiquitin-Like Proteins in Autophagy. BioMed Res. Int. 2014, 2014, 832704. [Google Scholar] [CrossRef]
- Lech, M.A.; Leśkiewicz, M.; Kamińska, K.; Rogóż, Z.; Lorenc-Koci, E. Glutathione Deficiency during Early Postnatal Development Causes Schizophrenia-Like Symptoms and a Reduction in BDNF Levels in the Cortex and Hippocampus of Adult Sprague–Dawley Rats. Int. J. Mol. Sci. 2021, 22, 6171. [Google Scholar] [CrossRef] [PubMed]
- Shevelkin, A.V.; E Terrillion, C.; Hasegawa, Y.; A Mychko, O.; Jouroukhin, Y.; Sawa, A.; Kamiya, A.; Pletnikov, M.V. Astrocyte DISC1 contributes to cognitive function in a brain region-dependent manner. Hum. Mol. Genet. 2020, 29, 2936–2950. [Google Scholar] [CrossRef]
- Karlsson, R.-M.; Adermark, L.; Molander, A.; Perreau-Lenz, S.; Singley, E.; Solomon, M.; Holmes, A.; Tanaka, K.; Lovinger, D.M.; Spanagel, R.; et al. Reduced alcohol intake and reward associated with impaired endocannabinoid signaling in mice with a deletion of the glutamate transporter GLAST. Neuropharmacology 2012, 63, 181–189. [Google Scholar] [CrossRef]
- Uchida, M.; Hida, H.; Mori, K.; Yoshimi, A.; Kitagaki, S.; Yamada, K.; Hiraoka, Y.; Aida, T.; Tanaka, K.; Ozaki, N.; et al. Functional roles of the glial glutamate transporter (GLAST) in emotional and cognitive abnormalities of mice after repeated phencyclidine administration. Eur. Neuropsychopharmacol. 2019, 29, 914–924. [Google Scholar] [CrossRef] [PubMed]
- A Lewis, D.; González-Burgos, G. Neuroplasticity of Neocortical Circuits in Schizophrenia. Neuropsychopharmacology 2008, 33, 141–165. [Google Scholar] [CrossRef]
- Brown, A.S.; Vinogradov, S.; Kremen, W.S.; Poole, J.H.; Deicken, R.F.; Penner, J.D.; McKeague, I.W.; Kochetkova, A.; Kern, D.; Schaefer, C.A. Prenatal Exposure to Maternal Infection and Executive Dysfunction in Adult Schizophrenia. Am. J. Psychiatry 2009, 166, 683–690. [Google Scholar] [CrossRef]
- Bubenikova-Valesova, V.; Stuchlik, A.; Svoboda, J.; Bures, J.; Vales, K. Risperidone and ritanserin but not haloperidol block effect of dizocilpine on the active allothetic place avoidance task. Proc. Natl. Acad. Sci. USA 2008, 105, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Hardingham, G.E.; Do, K.Q. Linking early-life NMDAR hypofunction and oxidative stress in schizophrenia pathogenesis. Nat. Rev. Neurosci. 2016, 17, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Casquero-Veiga, M.; Romero-Miguel, D.; MacDowell, KS.; Torres-Sanchez, S.; Garcia-Partida, JA.; Lamanna-Rama, N.; Gómez-Rangel, V.; Romero-Miranda, A.; Berrocoso, E.; Leza, JC.; et al. Omega-3 fatty acids during adolescence prevent schizophrenia-related behavioural deficits: Neurophysiological evidences from the prenatal viral infection with PolyI:C. Eur Neuropsychopharmacol. 2021, 46, 14–27. [Google Scholar] [CrossRef]
- Felgel-Farnholz, V.; Hlusicka, E.B.; Edemann-Callesen, H.; Garthe, A.; Winter, C.; Hadar, R. Adolescent raloxifene treatment in females prevents cognitive deficits in a neurodevelopmental rodent model of schizophrenia. Behav. Brain Res. 2023, 441, 114276. [Google Scholar] [CrossRef]
- Tsuji, A.; Ikeda, Y.; Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Morikawa, S.; Nakashima, M.; Asai, T.; Matsuda, S. The Tryptophan and Kynurenine Pathway Involved in the Development of Immune-Related Diseases. Int. J. Mol. Sci. 2023, 24, 5742. [Google Scholar] [CrossRef]
- Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Ikeda, Y.; Tsuji, A.; Matsuda, S. A New Concept of Associations between Gut Microbiota, Immunity and Central Nervous System for the Innovative Treatment of Neurodegenerative Disorders. Metabolites 2022, 12, 1052. [Google Scholar] [CrossRef]
- Asai, T.; Yoshikawa, S.; Ikeda, Y.; Taniguchi, K.; Sawamura, H.; Tsuji, A.; Matsuda, S. Encouraging Tactics with Genetically Modified Probiotics to Improve Immunity for the Prevention of Immune-Related Diseases including Cardio-Metabolic Disorders. Biomolecules 2022, 13, 10. [Google Scholar] [CrossRef]
- Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Ikeda, Y.; Tsuji, A.; Matsuda, S. Encouraging probiotics for the prevention and treatment of immune-related adverse events in novel immunotherapies against malignant glioma. Explor. Target. Anti-tumor Ther. 2022, 3, 817–827. [Google Scholar] [CrossRef] [PubMed]
Figure 4.
The protein expression of the brain. [A] The representative image of p62, GLAST, and GAPDH expression in the brain of individual mouse. [B] The protein expression of p62 was measured and normalized to GAPDH by Western blot. [C] The protein expression of GLAST was measured and normalized to GAPDH by Western blot. GLAST; a glutamate transporter protein [D] Positive correlation between the p62 and GLAST protein expression. r=0.484, P=0.111, y=0.8487x + 0.273.
Figure 4.
The protein expression of the brain. [A] The representative image of p62, GLAST, and GAPDH expression in the brain of individual mouse. [B] The protein expression of p62 was measured and normalized to GAPDH by Western blot. [C] The protein expression of GLAST was measured and normalized to GAPDH by Western blot. GLAST; a glutamate transporter protein [D] Positive correlation between the p62 and GLAST protein expression. r=0.484, P=0.111, y=0.8487x + 0.273.
Figure 5.
Overview of the correlation analyses. The image of the relation of behavioral tests, p62, and GLAST. The correlation of behavioral tests and the p62 expression shows in Figure 6B,D. The correlation of behavioral tests and the GLAST expression shows in Figure 6A,C. The correlation between p62 and GLAST expression shows in Figure 4D.
Figure 5.
Overview of the correlation analyses. The image of the relation of behavioral tests, p62, and GLAST. The correlation of behavioral tests and the p62 expression shows in Figure 6B,D. The correlation of behavioral tests and the GLAST expression shows in Figure 6A,C. The correlation between p62 and GLAST expression shows in Figure 4D.

Figure 6.
The correlation between behavioral tests score and the protein expression of p62 and GLAST. [A] Positive correlation between the last behavioral test score and p62 protein expression. r=0.82, P=0.001, y=0.1399x + 0.4459 [B] Positive correlation between the last behavioral test score and GLAST protein expression. r=0.58, P=0.047, y=0.1742x + 0.5219 [C] Positive correlation between the mean behavioral test score and p62 protein expression. r=0.63, P=0.028, y=0.2028x + 0.2569 [D] Positive correlation between the mean behavioral test score and GLAST protein expression. r=0.60, P=0.039, y=0.3381x + 0.0689.
Figure 6.
The correlation between behavioral tests score and the protein expression of p62 and GLAST. [A] Positive correlation between the last behavioral test score and p62 protein expression. r=0.82, P=0.001, y=0.1399x + 0.4459 [B] Positive correlation between the last behavioral test score and GLAST protein expression. r=0.58, P=0.047, y=0.1742x + 0.5219 [C] Positive correlation between the mean behavioral test score and p62 protein expression. r=0.63, P=0.028, y=0.2028x + 0.2569 [D] Positive correlation between the mean behavioral test score and GLAST protein expression. r=0.60, P=0.039, y=0.3381x + 0.0689.
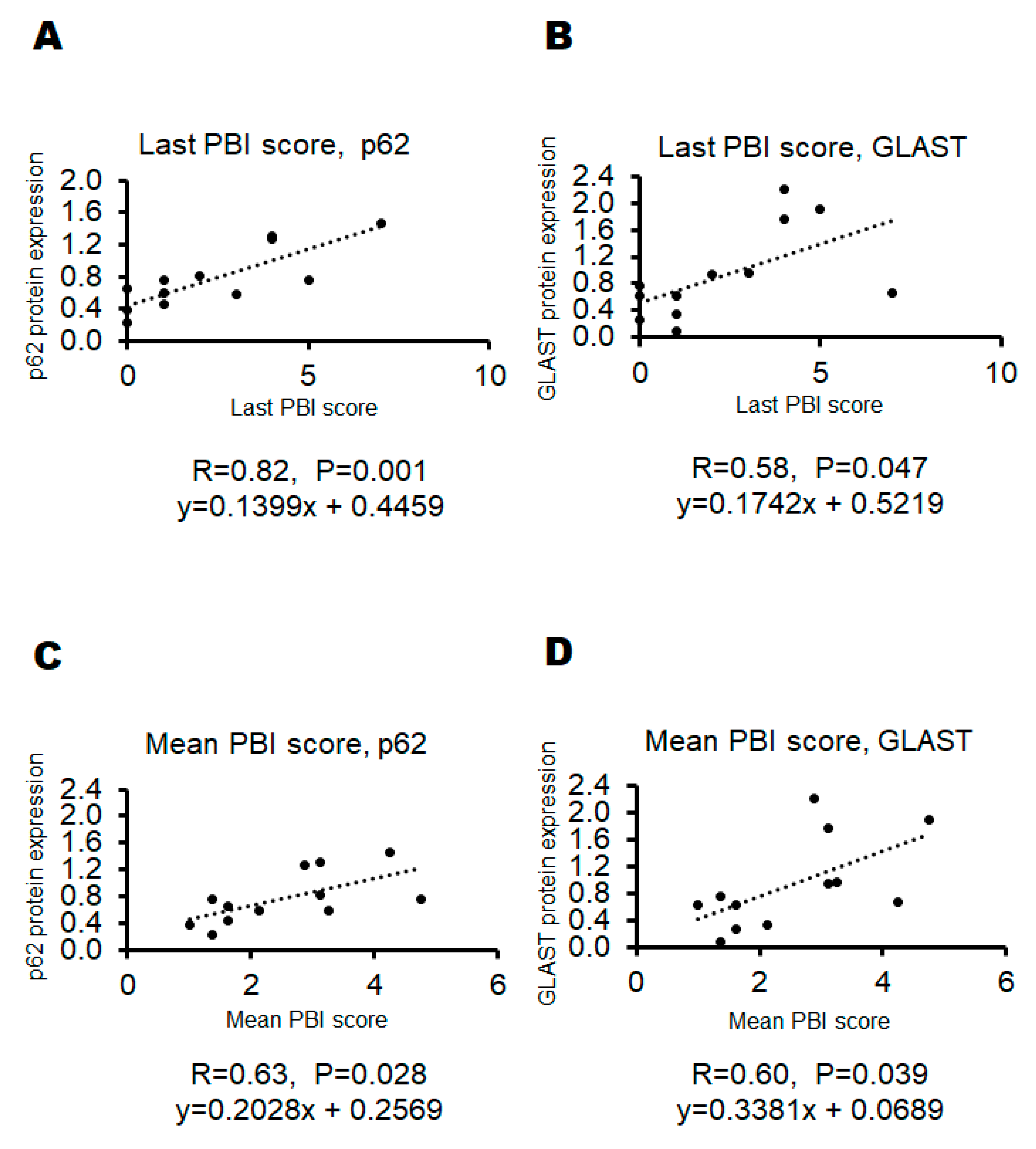

Table 1.
Behavioral tests score.
| Behavioral test | Score | |
|---|---|---|
| Descent step test | Non-descent descent |
0 1 |
| Modified three chambers test | In (III) room In (I) room In (II) room |
1 2 4 |
| Light/Dark room test | Time in light area ≧60 (sec) Time in light area 40-59 (sec) Time in light area 20-39 (sec) Time in light area 0-19 (sec) |
0 1 2 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



