
Submitted:
18 July 2023
Posted:
18 July 2023
You are already at the latest version
Abstract
The WRKY gene family in plants regulates the plant's response to drought through regulatory networks and hormone signaling. AfWRKY20 (MT859405) was cloned from A. fruticosa (Amorpha fruticosa,A. fruticosa) seedlings using RT-PCR. The binding properties of the AfWRKY20 protein and the W-box (a DNA cis-acting element) were verified both in vivo and in vitro using EMSA and Dual-Luciferase activity assays. RT-qPCR detected that the total expression level of AfWRKY20 in leaves and roots was 22 times higher in the 30% PEG6000 simulated drought treatment compared to the untreated group. Under the simulated drought stress treatments of sorbitol and ABA (abscisic acid,ABA), the transgenic tobacco with the AfWRKY20 gene showed enhanced drought resistance at the germination stage, with significantly increased germination rate, green leaf rate, fresh weight, and root length compared to the WT (wild type,WT) tobacco. In addition, the SOD (superoxide dismutase,SOD)activity, chlorophyll content, and Fv/Fm ratio of AfWRKY20 transgenic tobacco were significantly higher than those of the WT tobacco under natural drought stress, while the MDA (malondialdehyde,MDA) content and DAB (3,3'-diaminobenzidine,DAB) and NBT (nitroblue tetrazolium,NBT) staining levels were lower. The expression levels of oxidation kinase genes (NbSOD, NbPOD, and NbCAT) in transgenic tobacco under drought stress were significantly higher than those in WT tobacco. This enhancement in gene expression improved the ability of transgenic tobacco to detoxify ROS (reactive oxygen species,ROS). The survival rate of transgenic tobacco after natural drought rehydration was four times higher than that of WT tobacco. In summary, this study revealed the regulatory mechanism of AfWRKY20 in response to drought stress-induced ABA signaling, particularly in relation to the ROS. This finding provides a theoretical basis for understanding the pathways of WRKY20 involved in drought stress, and offers genetic resources for molecular plant breeding aimed at enhancing drought resistance.
Keywords:
Amorpha fruticose
; WRKY transcription factor
; drought stress
; tobacco genetic transformation
; stress resistance
1. Introduction
Drought is a prominent abiotic stress factor that poses a threat to the growth and development of plants, resulting in a significant decrease in crop yield [1]. Such stressful conditions lead to the accumulation of osmotic and oxidative regulators, which in turn induce the expression of stress-related genes [2].The efficiency of photosynthesis determines a plant's ability to withstand natural drought stress. Chlorophyll fluorescence parameters are key to the study of photosynthesis [3]. Under drought stress conditions, the process of light absorption and electron transport in photosynthesis can result in the accumulation of ROS not only in chloroplasts [4], but also in plants. This accumulation can damage the cell membrane system and cause oxidative stress. Fortunately, plants can not only utilize various mechanisms to reduce ROS production but also acquire ROS scavenging systems to protect themselves [5]. Plants transfer stress signals through signal transduction pathways in vivo and regulate the enzymatic antioxidant system, which includes SOD, POD (peroxidase,POD), CAT (catalase,CAT), and other antioxidant enzymes. This system helps remove excessive ROS components in cells [6]. In addition, for ROS accumulated in chloroplasts, non-photochemical quenching, photorespiration, and the CBB (Calvin-Benson-Bassham cycle,CBB) can dissipate the energy of excess electrons in chloroplasts and decrease the production of ROS [7,8]. In addition, drought stress triggers the accumulation of the plant hormone ABA through hyperosmotic signaling. This accumulation leads to the development of multiple ABA signaling pathways in plants. These pathways can promote adaptive responses to drought stress. Even in the absence of water stress, ABA inhibits plant stem and root growth [9].
Thus plants have developed various molecular mechanisms, including signal transduction and gene expression, to adapt to abiotic stresses. Evolved transcription factors (TFs), such as the CBF[10], ERF[11], BHLH[12], bZip[13], ZFP[14], MYB[15], NAC[16], C2H2[17], Dof[18], HSF[19] and WRKY[20] families, can regulate gene network expression to combat environmental stresses. The WRKY gene family is a group of plant-specific transcription factors (TFs) that play important roles in various aspects, including plant defense response [21], plant growth and development [22] and regulation of leaf senescence [23]. WRKY TFs are characterized by a conserved domain, which includes a WRKYGQK motif at the N-terminus and a zinc finger motif at the C-terminus [24]. According to the number of conserved WRKY domains and the structural characteristics of zinc finger motifs, they can be divided into three groups: Group Ⅰ, Group Ⅱ and Group Ⅲ. Group Ⅱ can be further divided into five subgroups: Ⅱa, Ⅱb, Ⅱc, Ⅱd and Ⅱe[25]. When plants are exposed to external stimuli, WRKY TFs are regulated by a cascade of defense signaling networks. They can recognize and bind to the W-box sequence [TTGAC(C/T)] present in the promoter region of the target gene, thereby participating in the regulatory network and enhancing the defense ability of plants [26].
WRKY TFs also play key roles in the transcriptional regulation and signal transduction processes in plants. They extensively regulate the expression changes of target genes in various physiological programs and are involved in various stress pathways [27]. Certain Arabidopsis WRKY TFs function as positive regulators in the pathway of ABA-mediated stomatal closure [28]. In addition, the WRKY20 gene in soybean is sensitive to ABA in terms of regulating stomatal closure. This sensitivity can enhance the plant's tolerance to drought stress [29]. In addition, WRKY TFs can also positively regulate drought resistance by improving ABA biosynthesis, such as the WRKY TF ZmWRKY79 in maize has been demonstrated to have this capability [30]. In the signal transduction pathways mediated by JA (jasmonate,JA) and SA (salicylic acid,SA), the WRKY70 TF is activated by SA, while its expression is suppressed by JA [31]. In terms of regulating plant growth and development, VvWRKY2 is specifically expressed in lignified cells of young grape stems. This expression affects the lignin biosynthesis pathway, which in turn impacts xylem development [32].
There is still a need to further explore the potential of WRKY TFs with different domains in various species. Recent transcriptome studies have shown that herbaceous plants, such as wheat (Triticum aestivum L.) [33], licorice (Glycyrrhiza glabra L.) [34], broad bean (Vicia faba L.) [35], and millet (Panicum miliaceum) [36], which upregulated WRKY TFs in response to drought stress. WRKY TFs in various woody plants, including Xanthoceras sorbifolium [37], Myrothamnus flabellifolia [38], and oil palm (Elaeis guineensis Jacq.) [39], which also responds to drought stress. However, functional studies of WRKY TFS in the woody plant A. fruticosa are limited. A. fruticosa is a perennial leguminous woody plant with strong adaptability. It can survive in adverse conditions, including cold, windy, and saline-alkali environments in Northeast China. It can also be used as a plant for greening, soil improvement, windbreak, and forest stabilization [40,41]. In addition, the roots, stems, leaves, and fruits of A. fruticosa not only possess medicinal properties for reducing dampness and swelling, but also have significant economic and practical value [42,43]. More importantly, A. fruticosa has a high tolerance to drought resistance [44].
Although studies on the drought resistance of A. fruticosa WRKY TFs are limited, transcriptome sequencing analysis of A. fruticosa has shown that the AfWRKY20 gene is upregulated in response to drought-induced expression [45]. In this study, the AfWRKY20 gene was cloned from the transcriptome sequencing of A. fruticosa under drought stress using RT-PCR technology.Subsequently, bioinformatics analysis, phylogenetic tree construction, and subcellular localization verification were conducted. The binding properties of AfWRKY20 and W-box were verified both in vivo and in vitro through EMSA and dual-Luciferase activity assays. The expression pattern of AfWRKY20 in response to abiotic stress was investigated through qRT-PCR analysis. To investigate the resistance of AfWRKY20 transgenic tobacco lines to drought stress at different growth stages, The study first subjected them to sorbitol stress and ABA stress as a means of simulating drought stress treatment. The study then measured phenotypic data, including germination rate, green leaf rate, fresh weight, and root length of the AfWRKY20 transgenic tobacco lines during the germination stage. Secondly, in this experiment, one-month-old and two-month-old AfWRKY20 transgenic tobacco lines were subjected to natural drought stress. The chlorophyll fluorescence parameters and the survival rate of the one-month-old AfWRKY20 transgenic tobacco lines after rewatering were measured. To investigate the potential of AfWRKY20 in enhancing the detoxification of ROS in tobacco, the study measured the activity of SOD, MDA content, levels of DAB and NBT staining, as well as the expression levels of oxidation kinase genes (NbSOD, NbPOD, and NbCAT) in AfWRKY20 transgenic tobacco lines after 2 months of natural drought. This study provides a reference for the role of AfWRKY20 in regulating reactive oxygen species in the ABA signaling response induced by drought stress. It also provides an experimental and theoretical basis for understanding the drought tolerance function of this TF by elucidating the molecular mechanism of AfWRKY20 in enhancing drought stress regulation.
2. Results
2.1. Cloning and bioinformatics analysis of AfWRKY20
Total RNA was extracted from A. fruticosa leaves (Figure S1-A), and a 20 μl cDNA reverse transcription product was prepared. The target band, which was approximately 1758 bp, was then amplified (Figure S1-B). The recombinant plasmid pMD18-T-AfWRKY20 was successfully constructed and sent for sequencing. The NCBI BLAST alignment showed that the nucleotide sequence was correct. AfWRKY20 (c194398, graph_c0) was predicted by the SMART online website to contain two WRKY domains (231-289 aa, 407-466 aa), a C2C2-type zinc finger protein, and a C2H2-type zinc finger protein (Figure 1A). The predictions for the secondary and tertiary structures were consistent (Figure 1B-C). In addition, Plant-mPLoc predicted that AfWRKY20 is localized in the nucleus (Figure S2-B). Using the MEME online website to predict the two WRKY domains of A. fruticosa WRKY20, it was found that the two WRKY domains were highly conserved (Figure 1D).
2.2. Sequence alignment and phylogenetic analysis
Phylogenetic relationships between Arabidopsis WRKYs and WRKYs in the transcriptome sequencing of A. fruticosa under drought stress were analyzed. Under drought stress, 51 WRKY TFs of A. fruticosa were identified from the transcriptome sequencing data. These TFs were categorized into four main groups: Group Ⅰ, Group Ⅱ, Group Ⅲ, and an unclassified group. These groups contained 6, 33, 7, and 5 A. fruticosa WRKY TFs, respectively. Group Ⅱ was further subdivided into five subgroups (Ⅱc, Ⅱa, Ⅱb, Ⅱd, and Ⅱe) containing 13, 3, 7, 5, and 5 A. fruticosa WRKY TFs, respectively. AfWRKY20 (c194398, graph_c0) contains two WRKY domains, and its zinc finger domain is of the C2H2 type. Therefore, it was classified as part of the first group. Seventy Arabidopsis WRKY transcription factors were divided into three groups: Group I contains 14 Arabidopsis WRKY transcription factors, Group Ⅱ contains 43 Arabidopsis WRKY transcription factors, and Group Ⅲ contains 13 Arabidopsis WRKY transcription factors. Group Ⅱ is further subdivided into five subgroups (Ⅱc, Ⅱa, Ⅱb, Ⅱd and Ⅱe). They contained 17, 3, 8, 7, and 8 Arabidopsis WRKY TFs, respectively. The branches of the phylogenetic tree showed that Arabidopsis WRKY20 and A. fruticosa WRKY20 in Group I were closely related (Figure 2).
In addition, a sequence comparison of A. fruticosa WRKY20 with 32 reported drought-resistant WRKY transcription factors from different species showed that, except for GmWRKY, IgWRKY50 had a unique WRKYGKK residue, while the rest had WRKYGQG, C, and H residues (Figure 3A). Phylogenetic tree construction revealed that AfWRKY20 was most closely related to GSWRKY20 (Figure 3B). In addition, to explore the species origin of A. fruticosa WRKY20, a phylogenetic tree was constructed to compare it with different species. The results showed that A. fruticosa WRKY20 was closely related to the WRKY20 of peanut and red bean (Figure S2-A).
2.3. Subcellular localization of AfWRKY20 protein
The control group (35s-GFP) was observed to be localized in the nucleus, cytoplasm, and cell membrane using confocal fluorescence microscopy (Olympus). In order to investigate the subcellular localization of AfWRKY20, the fusion protein of green fluorescent protein (GFP) - AfWRKY20 was expressed transiently in N. benthamiana's leaves, driven by the 35S promoter. AfWRKY20 localizes in the nucleus and overlaps with the signal of the nuclear-specific dye DAPI (Figure 4). The results of the AfWRKY20 protein experiment were consistent with the subcellular localization of the AfWRKY20 protein predicted by Plant-mPLoc, further confirming that the AfWRKY20 protein is located in the nucleus.
2.4. Analysis of binding properties of AfWRKY20 protein and W-box cis-acting elements
In order to explore the binding characteristics of the AfWRKY20 coding protein to the DNA cis-element W-box of the WRKY TF both in vivo and in vitro, we utilized electrophoretic mobility shift assay (EMSA) and Dual-Luc assay to validate the binding characteristics. EMSA kits were used to detect the gel-blocking signals of biotin-labeled probes and GST-AfWRKY20 fusion proteins (Figure 5A). The results showed that no band transfer was observed when biotin-labeled probes and GST proteins were added to the mixture. This finding suggests that the GST protein does not bind to the DNA probe. However, when the fusion protein (GST-AfWRKY20) was mixed with biotin-PR, a DNA-binding band was detected. The addition of competing probes weakened the composite signal of AfWRKY20 binding to the W-box and enhanced the signal of the free probe. This result indicates that the fusion protein is unable to bind to the cis-element W-box after binding to the TTGAC sequence of the DNA probe. Collectively, these experimental results confirm that the protein encoded by AfWRKY20 possesses the functional characteristics of a WRKY TF that binds to the W-box in vitro.
To confirm whether the AfWRKY20 protein can bind to W-box elements in vivo, this study was performed using a Dual-Luc reporter assay in N. benthamiana's tobacco. When mW-box-0800 was co-transformed with the AfWRKY20-62-SK vector, no LUC luminescence signal was observed. When W-box-0800 was co-transformed with the AfWRKY20-62-SK vector, a clear LUC luminescence signal was generated (Figure 5B). Thus, the AfWRKY20 protein is a representative WRKY protein that exhibits a specific binding affinity for the W-box element, as observed through in vitro and in vivo experiments.
2.5. Characterisation of gene expression of AfWRKY20
RT-qPCR was used to detect the expression of AfWRKY20 in various tissues and organs of A. fruticosa. The results showed that the gene was expressed in roots, stems, leaves, and flowers. Its expression level was highest in leaves and lowest in roots (Figure 6A). Under different concentrations (0%, 10%, 20%, and 30%) of PEG6000 to simulate drought stress, the expression of AfWRKY20 increased with the increase in stress concentration. In addition, the overall expression level of AfWRKY20 in leaves and roots significantly increased when treated with 30% PEG6000 compared to the control group, reaching 22 times that of the control group (Figure 6B).
The effects of three stress conditions (30% PEG6000, 150 mmol/L NaCl, and 30 mmol/L NaHCO3) on the expression of AfWRKY20 in the roots and leaves of A. fruticosa were analyzed. The results of the study showed that the expression of the AfWRKY20 gene increased most significantly at 12h of A. fruticosa leaf treatment compared to the control under 30% PEG6000 treatment (Figure 6C). However, the expression of the AfWRKY20 gene was significantly decreased at 48 hours of root treatment (Figure 6D). Under 150 mmol/L NaCl stress, the expression level of AfWRKY20 in the leaves and roots of A. fruticosa exhibited unstable fluctuations. The expression of the AfWRKY20 gene was 9-fold higher in leaves after 24 hours of stress (Figure 6E). The expression of AfWRKY20 was significantly reduced in roots after being stressed for 24 hours (Figure 6F). Under the treatment of 30 mmol/L NaHCO3, the expression level of AfWRKY20 initially increased in the leaves and roots of A. fruticosa, but then decreased. The expression of AfWRKY20 in leaves reached its highest level at 12h, which was 3.7-fold higher than that of the control (Figure 6G). The expression of AfWRKY20 peaked at 6 hours after A. fruticosa root treatment, which was 2.7 times higher than that of the control (Figure 6H).
2.6. Genetic transformation and drought resistance analysis of AfWRKY20 transgenic tobacco
2.6.1. Acquisition of AfWRKY20 transgenic tobacco
PCR molecular identification of T0 generation AfWRKY20 transgenic tobacco lines showed that the target band size of AfWRKY20 transgenic tobacco lines (#4, #5, #6, #7) was identical to that of the positive control plasmid pBI121-AfWRKY20-GFP (CK+). However, the negative control did not exhibit any band. This indicates that AfWRKY20 has been successfully integrated into the genomic DNA of tobacco (Figure S3-A). The expression of the transgenic tobacco lines was analyzed using RT-qPCR. The results showed that the expression level of the AfWRKY20 gene was significantly higher in transgenic tobacco lines 5, 6, and 7 compared to the WT (Figure S3-B).
The seeds of WT and T3 transgenic tobacco lines were sown on 1/2 MS medium and a kanamycin (50 mg/L) resistant medium. The results showed that the seeds of both the WT and AfWRKY20 transgenic lines grew normally on 1/2 MS medium. It was proven that the seeds from both the experimental group and the control group were able to germinate normally (Figure S3-C-a). Screening on Kana (50 mg/L) resistant medium showed that all WT unrooted and AfWRKY20 transgenic tobacco lines (#5, #6, and #7) grew normally, indicating that pure and AfWRKY20 transgenic tobacco lines had been obtained (Figure S3-C-b). Therefore, T3 generation AfWRKY20 transgenic tobacco #5, #6, and #7, which are both pure and highly expressed, were selected as the experimental materials for subsequent drought resistance stress.
2.6.2. Tolerance analysis of AfWRKY20 transgenic tobacco at germination stage under sorbitol and ABA simulated drought stress
When transgenic tobacco seeds containing AfWRKY20 were treated with different concentrations of sorbitol (0, 200, and 300 mM) for 15 days, the germination rate of the transgenic tobacco lines was higher compared to that of the WT tobacco lines. In addition, as the concentration of sorbitol stress gradually increased, the germination advantage of the transgenic lines became more apparent (Figure 7B). When exposed to sorbitol stress for 20 days, the transgenic tobacco lines exhibited a significantly higher rate of green leaf retention compared to the WT. The most significant difference was observed under 200 mM sorbitol stress (Figure 7C). At the same time, transgenic tobacco seeds with AfWRKY20 at different concentrations of ABA stress (0, 2, 2.5, 3, and 5 μM) during the germination stage showed a higher germination rate and green leaf rate compared to WT tobacco(Figure S5). These results suggest that the AfWRKY20 gene may enhance the drought stress tolerance of tobacco seeds during the germination stage under conditions simulated by sorbitol and ABA.
Statistical analysis of root length (Figure 8B) and fresh weight (Figure 8C) of WT and AfWRKY20 transgenic tobacco at the trifoliate stage after 10 days of stress with different concentrations of sorbitol (0, 200, 250, and 300 mM) showed that the fresh weight and root length of transgenic tobacco lines were significantly greater than those of WT tobacco lines. The most significant difference was observed at 200 mM sorbitol. After 15 days of stress with different concentrations of ABA (0, 5, 7.5, and 10 μM) at the trifoliate stage, both WT and AfWRKY20 transgenic tobacco lines showed significant improvements in root length (Figure S6-B) and fresh weight (Figure S6-C) compared to the WT line. These results suggest that the AfWRKY20 gene may regulate the growth and development of tobacco under conditions of sorbitol and ABA, simulating drought stress. This regulation may enhance tobacco's tolerance to drought stress.
2.6.3. Analysis of photosynthetic characteristics of AfWRKY20 transgenic tobacco pot seedlings under natural drought stress
The 1-month-old T3 generation AfWRKY20 transgenic tobacco pot seedlings were subjected to natural drought conditions for 0, 10, and 15 days before being rewatered for 3 days. Images were taken under natural light (Figure 9A), and then chlorophyll fluorescence was imaged using the FluorCam open chlorophyll fluorescence imaging system in Fv/Fm mode (Figure 9B). The photosynthetic characteristics of transgenic tobacco pot seedlings and wild-type pot seedlings were analyzed under natural drought stress. The results showed that the trend of F0 of WT was more pronounced than that of the transgenic lines during the 15-day drought treatment (Figure S4-A). The Fm value of the WT decreased compared to transgenic lines 5 and 6, but was higher compared to transgenic line 7 (Figure S4-B). The decrease in QL was more pronounced in the WT than in the transgenic strain (Figure S4-C). NPQ increased more slowly in the early stages of drought stress (0-10 D) and showed a larger decrease in the later stages (10-15d) compared to the transgenic lines (Figure S4-D). These results indicate that the PSII reaction center is more damaged in the WT tobacco than in AfWRKY20 transgenic tobacco.
The transgenic tobacco pot seedlings treated with drought treatment for 0, 10, and 15 days, followed by rewatering for 3 days. The wild-type tobacco plants were also included in the experiment. The plants were photographed under natural light (Figure 9A), revealing that the growth and development of the transgenic tobacco plants were superior to those of WT tobacco. A chlorophyll fluorescence imaging system was also used to capture chlorophyll fluorescence images at Fv/Fm (range: 0.2-0.8) (Figure 9B). It was observed that the fluorescence color of the drought-treated WT tobacco was not as red as that displayed by the transgenic tobacco. The chlorophyll fluorescence imaging system was used to quantitatively analyze the maximum optical quantum efficiency (Fv/Fm) collected after 0, 10, and 15 days of drought treatment, followed by 3 days of rewatering. The results showed that the Fv/Fm values of both the WT and transgenic tobacco decreased significantly after drought treatment compared to untreated tobacco. However, the Fv/Fm value of the transgenic tobacco was still significantly higher than that of the WT tobacco (Figure 9C). Although the Fv/Fm value of both the WT and transgenic tobacco recovered after three days of rehydration, the Fv/Fm value of the transgenic tobacco remained higher than that of the WT tobacco (Figure 9C). In addition, the survival rate of transgenic tobacco after rehydration was significantly higher than that of the WT tobacco (Figure 9D). These results suggest that AfWRKY20 can reduce the photosynthetic damage caused by drought stress and participate in the molecular mechanism of tobacco photosystem II, thereby enhancing the drought resistance of tobacco.
2.6.4. Determination of physiological indices of drought in AfWRKY20 transgenic tobacco
The T3 generation of AfWRKY20 transgenic lines grown for two months were subjected to 0, 10, and 15 days before being rewatered for 3 days under natural drought conditions. Before treatment, all plants exhibited a consistent growth state (Figure 10A). However, during the drought treatment, the transgenic tobacco exhibited distinct phenotypic changes compared to the WT tobacco. At 10 days of drought treatment, both the transgenic and the WT tobacco leaves exhibited slight wilting and yellowing. However, the changes were more noticeable in the WT tobacco (Figure 10B). After 15 days of drought treatment, the growth condition of transgenic tobacco leaves was significantly superior to that of WT leaves (Figure 10C). At 3 days of rewatering after the drought treatment, the tobacco plants that were overexpressed exhibited improved resilience (Figure 10D) and significantly higher Fv/Fm values (Figure 10E) compared to the WT tobacco. The chlorophyll content (SPAD) of transgenic tobacco was higher than that of WT tobacco throughout the natural drought treatment (Figure 10F).
Histochemical analysis using DAB and NBT as ROS indicators revealed deeper staining in the WT tobacco leaves than in transgenic leaves under drought stress. This finding indicates that the WT tobacco accumulated more hydrogen peroxide and superoxide anions during drought stress than transgenic tobacco, resulting in severe leaf damage (Figure 11A). SOD activity reflects the plant's ability to scavenge oxygen free radicals. After the drought treatment, the SOD content of the plants gradually increased with the extension of drought stress, reaching its peak at 10 days of drought (Figure 11B). In addition, the MDA content of the transgenic plants was significantly lower than that of the WT tobacco (Figure 11C). The gene expression levels of NbSOD (Figure 11D), NbPOD (Figure 11E), and NbCAT (Figure 11F) in transgenic tobacco were significantly higher than those in the WT tobacco, as determined by real-time fluorescence quantitative PCR. This increase in gene expression resulted in an enhancement of antioxidant enzyme levels in the plant, thereby improving the plant's ability to scavenge ROS in transgenic tobacco. These experimental results indicate that the overexpression of the AfWRKY20 gene enhances tobacco's tolerance to drought stress. This study provides a molecular basis for further investigating the potential of AfWRKY20 in enhancing stress tolerance in plants.
3. Discussion and Conclusions
The current drought has a significant impact on plant growth and development [46]. WRKY TFs are critical in regulating plant responses to abiotic stresses [47,48]. Related studies have shown that a new WRKY transcription factor, MuWRKY3 (Macrotyloma uniflorum Lam. Verdc.), can enhance drought resistance in transgenic peanut (Arachis hypogaea L.) plants [49]. In addition to the search for novel WRKY transcription factors, further investigation is needed to understand the resistance function of WRKY transcription factors in various species. Therefore, AfWRKY20 (c194398, graph_c0) was screened during transcriptome sequencing of A. fruticosa under drought stress to investigate the molecular mechanisms regulating drought stress. Bioinformatics analysis revealed that AfWRKY20 contains two WRKY domains (231-289 aa, 407-466 aa) and a C2H2-type zinc finger protein (Figure 1A). According to the classical classification criteria, this protein was classified into Group I, indicating its close relationship with the growth, development, and stress tolerance of nuclear organisms (Figure 1A) [50].
Based on the construction of the phylogenetic tree, A. fruticosa WRKY20 was found to be most closely related to Arabidopsis WRKY20 (Figure 2). Related studies have shown that species with high homology have similar gene functions [51]. AtWRKY20 coregulates the ABA signaling pathway with ABSCISIC ACID-INSENSITIVE 5 (ABI5). Thus, it plays an important role in biological processes such as seed germination, dormancy, anthocyanin synthesis, and response to stress [52]. To further explore the correlation between A. fruticosa WRKY20 and drought-resistant WRKY transcription factors in other species, a phylogenetic tree was constructed. The results revealed a significant similarity between AfWRKY20 and GsWRKY20 (Figure 3). GsWRKY20 plays an important role in enhancing drought tolerance and regulating ABA signaling [53]. Therefore, the present study hypothesized that AfWRKY20 enhances drought stress tolerance in tobacco by regulating the ABA signaling pathway.
In this study, validation experiments were performed to determine the subcellular localization of the pBI121::AfWRKY20::GFP fusion expression vector. The experimental results were consistent with the software's prediction that the protein would be localized in the nucleus. Therefore, AfWRKY20 may play a role in regulating cell signaling molecules (Figure 4)[54]. The binding properties of the AfWRKY20 TF and the DNA cis-element W-box were confirmed through EMSA experiments (Figure 5A) and double LUC experiments (Figure 5B). The W-box mainly exists in the promoter regions of resistance genes associated with disease and insect resistance, drought, low temperature, saline-alkali, and other factors. It can regulate plant resistance by mediating hormone signal transduction pathways. The molecular mechanism of the AfWRKY20 protein regulating drought stress may be similar to that of SbWRKY30. Both of them induce the expression of drought tolerance genes by binding to W-box elements in the promoters of drought tolerance genes in plants, thereby improving the plant's drought tolerance [55].
When A. fruticosa was treated with different concentrations (0%, 10%, 20%, and 30%) of PEG6000 to simulate drought stress, the total expression level of AfWRKY20 in leaves and roots was 22 times higher in the 30% PEG6000 treatment compared to the untreated group (Figure 6B). In addition, the effects of three different stress conditions (30% PEG6000, 150 mmol/L NaCl, and 30 mmol/L NaHCO3) on the expression level of AfWRKY20 in the roots and leaves of A. fruticosa were analyzed. Under 30% PEG6000 treatment, the expression of the AfWRKY20 gene increased most significantly at 12 hours of A. fruticosa leaf treatment compared to the control. Under the stress of 150 mmol/L NaCl, the expression levels of AfWRKY20 in the leaves and roots of A. fruticosa exhibited unstable fluctuations (Figure 6E-F). Under the treatment of 30 mmol/L NaHCO3, the expression levels of AfWRKY20 in the leaves and roots of A. fruticosa showed an overall trend of initially increasing and then decreasing (Figure 6G-H). AfWRKY20 is differentially expressed under these three distinct stress conditions. Therefore, it is speculated that the AfWRKY20 gene may regulate multiple stress pathways. For example, AtWRKY53 can respond to both drought stress and salt stress, and AtWRKY6 can regulate both mechanical damage and oxidative stress [56].
When tobacco plants overexpressing AfWRKY20 were exposed to different concentrations of sorbitol stress (Figure 7-8) and ABA stress (Figure S5-6), the germination rate, green leaf rate, root length, fresh weight, and other phenotypes were significantly higher compared to those of WT tobacco. Our results suggest that AfWRKY20 may regulate the growth and development of tobacco by influencing the ABA signaling pathway, thereby enhancing its drought resistance.
Analysis of the photosynthetic characteristics of transgenic seedlings under drought stress indicated that AfWRKY20 could reduce the damage caused by stress on photosynthesis and enhance drought stress tolerance. This finding was confirmed by measuring the chlorophyll fluorescence parameters F0, Fm, Fv/Fm, QL, and NPQ of transgenic tobacco plants under natural drought stress (Figure S4). After rehydration, the Fv/Fm values and survival rate of the transgenic plants were significantly higher than those of the WT plants (Figure 9). This suggests that AfWRKY20 plays a crucial role in regulating photosystem damage caused by drought stress [57].
AfWRKY20 overexpression in tobacco under natural drought stress had lower MDA content (Figure 11-C) and reduced DAB and NBT staining (Figure 11-A). These findings indicate a decrease in ROS in the transgenic lines. At the same time, it had higher SOD enzyme activity in vivo (Figure 11-B). The expression levels of NbSOD, NbPOD, and NbCAT in transgenic tobacco were significantly higher than those in the WT tobacco, as determined by real-time fluorescence quantitative PCR (Figure 11 D-F). This indicates an enhancement in the content of antioxidant enzymes in the plant. It is speculated that the regulatory mechanism of AfWRKY20 may be similar to that of maize ZmWRKY40. This mechanism involves reducing the levels of ROS in transgenic lines during drought stress. This is achieved by enhancing the activities of POD and CAT , which ultimately enhances the drought resistance of the transgenic lines [58].
The above results suggest that the AfWRKY20 TF can act as a node in the drought stress signaling pathway through ROS balance and ABA signaling pathways, which is similar to the mechanism of XsWRKY20 as a positive regulator [59]. This study revealed the regulatory mechanism of AfWRKY20 in response to drought stress-induced ABA signaling, involving ROS. These findings lay the foundation for further studies on the mechanisms through which AfWRKY20 enhances drought tolerance, providing genetic resources for molecular plant breeding aimed at developing drought-resistant varieties.
4. Materials and Methods
4.1. Plant material
Seeds of A. fruticosa Linn were obtained from Wu Songquan’s research group at Yanbian University, and those of N. benthamiana were acquired from Bu Qingyun's research group at the Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences.
4.2. Strains, vectors, and reagents
Escherichia coli (Top10) competent and Agrobacterium tumefaciens (EHA105) were preserved in the Key Laboratory of Northeast Saline Alkaline Vegetation Restoration and Reconstruction, which is under the Ministry of Education.
PMD18-T vector was acquired from Takara, and the Gateway series entry vector PQBV3, plant expression vector pGWB18, and pBI121-MCS-GFP plant expression vector were obtained from the Key Laboratory of Northeast Salt Vegetation Restoration and Reconstruction Ministry of Education. pGreenⅡ0800-LUC and pGreenⅡ62-SK- LUC carrier were gifted by the Institute of Geography, Chinese Academy of Sciences. T4-DNA ligase, 2×Ex-Taq DNA polymerase, and real-time fluorescent dye 2×SYBR Green qPCR Master Mix were supplied by TaKaRa. Acetosyringone was purchased from Solarbio Biologics, and D-luciferin was obtained from PROMEGA Company. RNA extraction kits, gel recovery kits, and plasmid extraction kits were purchased from Beijing Kangwei Century Company. Domestic analytical reagents were used for all other experiments.
4.3. Cloning and bioinformatics analysis of AfWRKY20 gene
4.3.1. Gene cloning
In the transcriptome data of A. fruticosa under 20% PEG6000 simulated drought stress, after mining of highly differentially expressed genes under drought stress, c194398 and graph_c0 were found to respond to drought stress. Therefore, according to the CDS sequence of c194398, graph_c0, the gene has been designated as AfWRKY20 (accession number: MT859405). Specific primers, AfWRKY20 F1 and AfWRKY20 R1, (AfWRKY20 F1/R1 in table S1)were designed using Primer 5.0 software. The full-length sequence of AfWRKY20 was amplified by RT-PCR and then ligated into the pMD18-T vector. The recombinant vector was transformed into TOP10 through heat shock transformation. The plasmids in the successful bacterial solution were then identified by PCR and extracted using the Beijing Kangwei Century Plasmid Extraction Kit. The recombinant plasmids were then sent to Kumei company for sequencing.
4.3.2. Bioinformatics analysis
The SMART (SMART: Main page (embl-heidelberg.de)) [60] was used to predict the primary structure of AfWRKY20, while The SOPMA (NPSP@ : SOPMA secondary structure prediction (ibcp.fr)) was used to predict its secondary structure. The SWISS-MODEL (The SWISS-MODEL Interactive Workspace (expasy.org)) [61] was used to predict the tertiary structure of AfWRKY20. The Plant-mPLoc (Plant-mPLoc server (sjtu.edu.cn)) [62] was used to predict the subcellular localization of AfWRKY20. The MEME(Introduction-MEME Suite (meme-suite.org)) [63] was used to predict the conservation of the WRKY domain in AfWRKY20.
4.4. Sequence alignment and construction of phylogenetic trees
Multiple sequence alignments were performed on WRKY amino acid sequences using ClustalW in MEGA 7.0 with default parameters. A phylogenetic tree was constructed using the neighbor-joining (NJ) method with MEGA 7.0 software. These parameters were used in the NJ method: bootstrap (1000 replicates), complete deletion, and amino: p distance[65]. This setup was employed for constructing all phylogenetic trees in this paper.
In order to compare the phylogenetic relationships of 70 Arabidopsis protein WRKYs and 51 drought-stressed A. fruticosa WRKYs, all 121 WRKY protein sequences were divided into four groups (Ⅰ, Ⅱ, Ⅲ, and unclassified). Group Ⅱ was further divided into five subgroups (Ⅱa, Ⅱb, Ⅱc, Ⅱd, Ⅱe). In order to investigate the phylogenetic relationship between AfWRKY20 and other drought-resistant WRKY transcription factors, we compared the amino acid sequences of 32 reported drought-resistant WRKY transcription factors with AfWRKY20 and constructed a phylogenetic tree. Additionally, a phylogenetic tree was constructed to explore the homology between AfWRKY20 and different species of WRKY.
4.5. Characterisation of gene expression of AfWRKY20
To investigate the differential expression of AfWRKY20 in various tissues and organs of A. fruticosa, the seeds of A. fruticosa were cultured in 96-well plates using the hydroponic method for four weeks. Different tissues (root, stem, leaf, and flower) were selected from the growth and development of A. fruticosa to extract total RNA. To investigate the gene expression characteristics of AfWRKY20 under different stress conditions, A. fruticosa plants with the same growth rate as A. fruticosa were selected. These plants were then treated with 30 mmol/L NaHCO3, 150 mmol/L NaCl, and 30% PEG6000. The roots and leaves were collected at 0, 6, 12, 24, and 48 hours. Afterwards, A. fruticosa seedlings were treated with various concentrations of PEG6000 (0%, 10%, 20%, and 30%) for 48 hours, and then leaves and roots were collected. Expression of AfWRKY20 gene at 0 h under adversity stress was used as a control.The total RNA was extracted from the samples mentioned above, and the concentration of RNA was examined. One microgram (μg) of total RNA was reverse transcribed into complementary DNA (cDNA) and diluted 100-fold as a template for fluorescence quantitative PCR. After conducting RT-qPCR analysis using primers for the internal reference gene (AfTubu.F/R in Table S1) and specific primers for AfWRKY20 (qAfWRKY20 F/R in Table S1), data were collected using the MxPro-Mx3000P system in this study. Each set of experimental data was subjected to three biological replicates, as well as technical replicates.
4.6. Subcellular localization analysis of AfWRKY20
ORF-specific primers (AfWRKY20-F1/R1 in table S1) were designed using the coding sequence (CDS) of AfWRKY20 and identified by PCR reaction using the pMD18-T-AfWRKY20 plasmid as a template. Then, the recovered product of the target band was ligated with the recovered product of the pBI121::GFP vector, which had been digested with XbaI and SalI. This resulted in the creation of the two-component plant recombinant expression vector pBI121::AfWRKY20::GFP, which was placed under the control of the CaMV 35S promoter. In this study, the recombinant vector (35S::AfWRKY20::GFP) as an experimental group and the empty vector (35S-GFP) as a control group were transformed into EHA105 using electroporation. Using the transient transformation method, tobacco plants were successfully transformed by EHA105. The identified strain of EHA105 was injected into the epidermal cells of N. benthamiana's tobacco. The plants were then incubated in the dark for 12 to 16 hours, followed by incubation under natural light for 3 days. Confocal laser fluorescence microscopy (Olympus) was used to observe the green fluorescence channel, DAPI staining channel, and bright field channel. The superimposed images of the channels were used to confirm the subcellular localization of AfWRKY20.[66] Finally, it was verified whether it was consistent with the subcellular localization results predicted by Plant-mPLoc .
4.7. Analysis of binding properties of AfWRKY20 protein and W-box cis-acting elements
In this study, EMSA was used to investigate the binding characteristics between AfWRKY20 and the W-box element. The prokaryotic expression system was used to express and purify the fusion protein GST-AfWRKY20. Glutathione Sepharose 4B (GE) GST affinity chromatography was used to obtain purified GST-AfWRKY20 fusion protein for the subsequent gel blocking assay. Oligonucleotide probes were prepared using synthetic W-box probe primers (W-box F1/R2 in table S1) and synthesized by Doctor Biology Co. The oligonucleotide probes were labeled with biotin, and a reverse complementary DNA was used as a cold competitive probe (Cold-Pr). The preparation was performed according to the instructions of the EMSA kit (Biyuntian Biological Co., Ltd.) [67,68].
Four-week-old leaves of N. benthamiana's tobacco were used for dual-Luciferase activity assays [69]. The sequences of the W-box (W-box F/R in table S1) and mW-box (mW-box F/R in table S1) elements, which consist of three tandem repeats, were synthesized using oligonucleotide sequencing and then seamlessly cloned into the pGreenII 0800-Luc vector. To construct the recombinant plasmid AfWRKY20-pGreenII62sk-LUC, specific primers (Table S1 AfWRKY20-62-SK F/R) were designed to incorporate the homology arm and SalI and BamHI restriction sites. The recombinant plasmid (Figure S1-D) was successfully identified through double enzyme digestion and then electrotransformed into EHA105. W-box-0800, mW-box-0800, pGreenII 0800-LUC empty vectors, pGreenII 62SK,and AfWRKY20-62-SK were used to transiently express tobacco leaves.The Agrobacterium hybrid system of pGreenII 0800-LUC vectors and pGreenII 62SK vectors was used as a negative control for transforming N. benthamiana’s tobacco.Leaves were incubated in the dark for 12 hours and then exposed to normal light for 3 days. After cutting the leaves, the reaction solution containing 1 mmol/Lof the fluorescein substrate D-fluorescein was injected into the wound. The leaves were then left in the dark for 5-7 minutes. Subsequently, the leaves were examined using a fluorescence imaging system, and the exposure time was adjusted based on the experimental results. Finally, the samples were imaged.
4.8. Genetic transformation and drought resistance analysis of AfWRKY20 overexpressed tobacco
4.8.1. Acquisition of AfWRKY20 transgenic tobacco
In this study, tobacco leaves were infected with EHA105 carrying the pBI121::AfWRKY20::GFP recombinant plasmid. The infected tobacco leaves were then soaked and placed on a tobacco co-medium (1/2MS+As). The leaves were cultured under ambient conditions in the dark for three days. Bud differentiation was then induced on tobacco selective differentiation medium containing 50 mg/L of kanamycin. Finally, rooting was induced in a rooting medium (1/2MS + 50 mg/L Kanamycin + 250 mg/L Carbenicillin), resulting in a T0 generation of AfWRKY20 transgenic tobacco lines.
In this study, CTAB was used to extract genomic DNA from T0 generation AfWRKY20 transgenic tobacco lines (#4, #5, #6, and #7) and the WT. AfWRKY20 overexpressed plants of the T0 generation were identified using PCR molecular biology techniques with specific primers (AfWRKY20 F1/R1 in table S1). DNA from the WT was used as the negative control (CK-), the PBI121-AfWRKY20-GFP plasmid was used as the positive control (CK+), and DNA from the T0 generation AfWRKY20 transgenic tobacco lines (#4, #5, #6, and #7) was used as the experimental group. The expression of the AfWRKY20 gene was identified in different transgenic tobacco lines (#4, #5, #6, and #7) using real-time fluorescence quantitative detection technology. RNA was extracted from the transgenic tobacco lines AfWRKY20 (#4, #5, #6, and #7) as the experimental group, while the WT was used as the control group. The extracted RNA was then reverse transcribed into cDNA. A 100-fold dilution of cDNA was used as the template. The internal reference primer, AfTubu.F/R (AfTubu.F/R in table S1), and the specific primer sequence for AfWRKY20 real-time fluorescence quantification, AfWRKY20 F/R (qAfWRKY20 F/R in table S1), were used. Finally, data were collected using the MxPro-Mx3000P system. Three biological replicates and three technical replicates were performed for each set of experimental data. Finally, the tobacco lines of the experimental group were screened on the 1/2MS medium with kanamycin (50 mg/L) resistance. The aim was to determine if all the seeds of the transgenic tobacco lines germinated and if the T3 generation AfWRKY20 transgenic seeds were obtained in a pure state [70].
4.8.2. Tolerance analysis of AfWRKY20 transgenic tobacco at germination stage under sorbitol and ABA simulated drought stress
The seeds of T3 generation AfWRKY20 transgenic tobacco (#5, #6, and #7) and the WT tobacco were sterilized and placed in 1/2MS sorbitol solutions at different concentrations (0, 200, and 300 mM) and 1/2MS ABA stress solutions at different concentrations (0, 2, 2.5, 3, and 5 μM). T3 generation AfWRKY20 transgenic tobacco seeds (#5, #6, and #7) were used as the experimental groups, while wild type tobacco seeds (WT) were used as the control group. The experimental and control groups were seeded with 30 seeds on a 1/2MS plate containing different concentrations of sorbitol stress. The plates were then placed at -4°C and subjected to vernalization at a low temperature for three days. They were then cultured horizontally at 25°C (8 hours light/16 hours dark). Germination rate and green leaf rate were counted at the beginning of seed germination in 1/2MS medium under various stress conditions. Three sets of biological replicates and three sets of technical replicates were performed for each experimental set.
Seeds from both the experimental and control groups were placed in 1/2MS medium and cultured vertically until reaching the trifoliate stage. The tobacco plants from both the experimental and control groups, which had the same growth rate, were exposed to various concentrations of sorbitol stress (0, 200, 250, and 300 mM) and different concentrations of ABA stress (0, 5, 7.5, and 10 μM) in 1/2MS medium. After 15 days of vertical cultivation at 25°C (8/16 h-light/dark), we measured the fresh weight and root length under various stress concentrations using ImageJ software for analysis. Three sets of biological replicates and three sets of technical replicates were performed for each experimental group.
4.8.3. Analysis of photosynthetic characteristics of AfWRKY20 transgenic tobacco seedlings under natural drought stress
The T3 generation AfWRKY20 transgenic tobacco lines (#5, #6, and #7) mentioned above were vertically cultured until the trifoliate stage. These lines were used as the experimental groups, while the WT tobacco was used as the control group. The tobacco seedlings from both the experimental group and the control group, which had similar growth, were selected for soil cultivation. Sixteen seedlings from each group (4x4) were placed in blue pots and cultured at a temperature of 25℃ (8/16 h-light/dark) for one month. The open chlorophyll fluorescence imaging system was used to investigate the sensitivity of the plant photosystem II response to drought stress in both the experimental and control groups after 0, 10, and 15 days of natural drought treatment. Chlorophyll fluorescence parameters measured by the system include F0 (minimum fluorescence yield in the absence of photosynthetic light), Fm (maximum fluorescence yield in the absence of photosynthetic light), FV/Fm (maximum efficiency of PSII), QL (photochemical quenching coefficient based on the lake model), and NPQ (extent of excess energy dissipation in the form of heat) [71]. At the same time, photographs of tobacco plants were taken under natural light on days 0, 10, and 15 of the natural drought treatment, as well as on the third day after rewatering. In addition, the tobacco's chlorophyll fluorescence imager (with a cursor range of 0.2-0.8) was used to capture images in Fv/Fm mode. The survival rate of tobacco was then calculated after 3 days of rehydration [72]. Each experimental group had three biological replicates and three technical replicates.
4.8.4. Determination of physiological indices in AfWRKY20 transgenic tobacco under natural drought treatment
AfWRKY20 transgenic tobacco lines (#5, #6, and #7) and wild-type tobacco lines of the T3 generation, which exhibited consistent growth for two months, were selected. These lines were exposed to natural drought conditions for 0, 10, and 15 days, followed by a period of rewatering for 3 days. Fv/Fm values of tobacco were collected using the FluorCam open chlorophyll fluorescence imaging system on day 0, day 15 of the natural drought treatment, and day 3 of rewatering. The chlorophyll content, MDA content, and SOD activity of tobacco leaves were measured during the natural drought stage and rewatering stage [73]. The tobacco leaves with the most noticeable phenotypic changes were selected for DAB and NBT chemical staining analysis after 15 days of drought treatment [74]. RNA was extracted from transgenic tobacco lines expressing AfWRKY20 (#5, #6, and #7) after 15 days of drought stress as the experimental group. Wild-type RNA tobacco was used as the control group. The extracted RNA was then reverse transcribed into cDNA.
Using a 100-fold diluted cDNA as a template and NbActin F/R (NbActin F/R in Table S1) as the internal reference primer, specific primer sequences NbSOD F/R, NbPOD F/R, and NbCAT F/R (NbSOD F/R, NbPOD F/R, and NbCAT F/R in Table S1) were used for real-time fluorescence quantification.Finally, data were collected using the MxPro-Mx3000P system. Three sets of biological replicates and three sets of technical replicates were performed for each experimental set.
4.9. Statistical Analysis
All data were processed using the Paired Comparison plot in ORIGIN software. The following parameters were used in the Paired Comparison plot: Error bar - SD, Mean comparison methods - Tukey, and Significance level - 0.05. Lowercase letters (a, b, c, etc.) represent statistical differences with p < 0.05.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, D.L. and Q.G.; methodology, D.L. and Q.G.; Software, D.L.; validation, D.L., B.G., C.H., J.S., X.W.and S.M.; formal analysis, D.L.; investigation, D.L., J.G., and R.Y.; resources, Q.G.; data curation, D.L.; writing—original draft preparation, D.L.; writing—review and editing, D.L.; visualization, D.L.; supervision, Q.G.; project administration, Q.G.; funding acquisition, Q.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32171989) and the College Students’ innovation and Entrepreneurship Project (DC-2023187).
Institutional Review Board Statement
Exclude this statement.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data from this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
We declare no conflict of interest.
References
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.-K. Abiotic stress responses in plants. Nat. Rev. Genet. 2021, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Valliyodan, B.; Nguyen, H.T. Understanding regulatory networks and engineering for enhanced drought tolerance in plants. Curr. Opin. Plant Biol. 2006, 9, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Bueno, M.L.; Pineda, M.; Barón, M. Phenotyping Plant Responses to Biotic Stress by Chlorophyll Fluorescence Imaging. Front. Plant Sci. 2019, 10, 1135. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Chaux, F.; Burlacot, A.; Mekhalfi, M.; Auroy, P.; Blangy, S.; Richaud, P.; Peltier, G. Flavodiiron Proteins Promote Fast and Transient O2 Photoreduction in Chlamydomonas. Plant Physiol. 2017, 174, 1825–1836. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive Oxygen Species in Plant Signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef]
- Kozaki, A.; Takeba, G. Photorespiration protects C3 plants from photooxidation. Nature 1996, 384, 557–560. [Google Scholar] [CrossRef]
- Saroussi, S.; Karns, D.A.J.; Thomas, D.C.; Bloszies, C.; Fiehn, O.; Posewitz, M.C.; Grossman, A.R. Alternative outlets for sustaining photosynthetic electron transport during dark-to-light transitions. Proc. Natl. Acad. Sci. 2019, 116, 11518–11527. [Google Scholar] [CrossRef]
- Aslam, M.M.; Waseem, M.; Jakada, B.H.; Okal, E.J.; Lei, Z.; Saqib, H.S.A.; Yuan, W.; Xu, W.; Zhang, Q. Mechanisms of Abscisic Acid-Mediated Drought Stress Responses in Plants. Int. J. Mol. Sci. 2022, 23, 1084. [Google Scholar] [CrossRef]
- Lee, E.S.; Park, J.H.; Wi, S.D.; Kang, C.H.; Chi, Y.H.; Chae, H.B.; Paeng, S.K.; Ji, M.G.; Kim, W.-Y.; Kim, M.G.; et al. Redox-dependent structural switch and CBF activation confer freezing tolerance in plants. Nat. Plants 2021, 7, 914–922. [Google Scholar] [CrossRef]
- Nowak, K.; Morończyk, J.; Grzyb, M.; Szczygieł-Sommer, A.; Gaj, M.D. miR172 Regulates WUS during Somatic Embryogenesis in Arabidopsis via AP2. Cells 2022, 11, 718. [Google Scholar] [CrossRef] [PubMed]
- An, J.-P.; Zhang, X.-W.; Liu, Y.-J.; Wang, X.-F.; You, C.-X.; Hao, Y.-J. ABI5 regulates ABA-induced anthocyanin biosynthesis by modulating the MYB1-bHLH3 complex in apple. J. Exp. Bot. 2020, 72, 1460–1472. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, Y.; Makita, Y.; Shimohira, H.; Matsui, M. Time-Course Transcriptome Study Reveals Mode of bZIP Transcription Factors on Light Exposure in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 1993. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Farman, K.; Waseem, M.; Rana, R.M.; Nawaz, M.A.; Rehman, H.M.; Abbas, T.; Baloch, F.S.; Akrem, A.; Huang, J.; et al. Genome-wide identification, classification, expression profiling and DNA methylation (5mC) analysis of stress-responsive ZFP transcription factors in rice (Oryza sativa L.). Gene 2019, 718, 144018. [Google Scholar] [CrossRef]
- Tiwari, P.; Indoliya, Y.; Chauhan, A.S.; Pande, V.; Chakrabarty, D. Over-expression of rice R1-type MYB transcription factor confers different abiotic stress tolerance in transgenic Arabidopsis. Ecotoxicol. Environ. Saf. 2020, 206, 111361. [Google Scholar] [CrossRef]
- Trishla, V.S.; Kirti, P.B. Structure-function relationship of Gossypium hirsutum NAC transcription factor, GhNAC4 with regard to ABA and abiotic stress responses. Plant Sci. 2020, 302, 110718. [Google Scholar] [CrossRef]
- Kaur, K.; Megha, S.; Wang, Z.; Kav, N.N.; Rahman, H. Identification and expression analysis of C2H2-zinc finger protein genes reveals their role in stress tolerance in Brassica napus. Genome 2023, 66, 91–107. [Google Scholar] [CrossRef]
- Sugiyama, T.; Ishida, T.; Tabei, N.; Shigyo, M.; Konishi, M.; Yoneyama, T.; Yanagisawa, S. Involvement of PpDof1 transcriptional repressor in the nutrient condition-dependent growth control of protonemal filaments in Physcomitrella patens. J. Exp. Bot. 2012, 63, 3185–3197. [Google Scholar] [CrossRef]
- Scharf, K.-D.; Berberich, T.; Ebersberger, I.; Nover, L. The plant heat stress transcription factor (Hsf) family: Structure, function and evolution. Biochim. et Biophys. Acta (BBA) - Gene Regul. Mech. 2012, 1819, 104–119. [Google Scholar] [CrossRef]
- Babitha, K.C.; Ramu, S.V.; Pruthvi, V.; Mahesh, P.; Nataraja, K.N.; Udayakumar, M. Co-expression of AtbHLH17 and AtWRKY28 confers resistance to abiotic stress in Arabidopsis. Transgenic Res. 2012, 22, 327–341. [Google Scholar] [CrossRef]
- Wani, S.H.; Anand, S.; Singh, B.; Bohra, A.; Joshi, R. WRKY transcription factors and plant defense responses: latest discoveries and future prospects. Plant Cell Rep. 2021, 40, 1071–1085. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, W.; Xu, Z.; Chen, M.; Yu, D. Functions of WRKYs in plant growth and development. Trends Plant Sci. 2023, 28, 630–645. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, Y.; Wang, C.; Zhang, D.; Liu, J.; He, M.; Chen, M.; Guo, Y. Brassica napus Transcription Factor Bna.A07.WRKY70 Negatively Regulates Leaf Senescence in Arabidopsis thaliana. Plants 2023, 12, 347. [Google Scholar] [CrossRef] [PubMed]
- Rinerson, C.I.; Rabara, R.C.; Tripathi, P.; Shen, Q.J.; Rushton, P.J. The evolution of WRKY transcription factors. BMC Plant Biol. 2015, 15, 66. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Ülker, B.; Somssich, I.E. WRKY transcription factors: from DNA binding towards biological function. Curr. Opin. Plant Biol. 2004, 7, 491–498. [Google Scholar] [CrossRef]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Rushton, D.L.; Tripathi, P.; Rabara, R.C.; Lin, J.; Ringler, P.; Boken, A.K.; Langum, T.J.; Smidt, L.; Boomsma, D.D.; Emme, N.J.; et al. WRKY transcription factors: key components in abscisic acid signalling. Plant Biotechnol. J. 2011, 10, 2–11. [Google Scholar] [CrossRef]
- Luo, X.; Bai, X.; Sun, X.; Zhu, D.; Liu, B.; Ji, W.; Cai, H.; Cao, L.; Wu, J.; Hu, M.; et al. Expression of wild soybean WRKY20 in Arabidopsis enhances drought tolerance and regulates ABA signalling. J. Exp. Bot. 2013, 64, 2155–2169. [Google Scholar] [CrossRef]
- Gulzar, F.; Fu, J.; Zhu, C.; Yan, J.; Li, X.; Meraj, T.A.; Shen, Q.; Hassan, B.; Wang, Q. Maize WRKY Transcription Factor ZmWRKY79 Positively Regulates Drought Tolerance through Elevating ABA Biosynthesis. Int. J. Mol. Sci. 2021, 22, 10080. [Google Scholar] [CrossRef]
- Li, J.; Brader, G.; Palva, E.T. The WRKY70 Transcription Factor: A Node of Convergence for Jasmonate-Mediated and Salicylate-Mediated Signals in Plant Defense. Plant Cell 2004, 16, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Guillaumie, S.; Mzid, R.; Méchin, V.; Léon, C.; Hichri, I.; Destrac-Irvine, A.; Trossat-Magnin, C.; Delrot, S.; Lauvergeat, V. The grapevine transcription factor WRKY2 influences the lignin pathway and xylem development in tobacco. Plant Mol. Biol. 2009, 72, 215–234. [Google Scholar] [CrossRef] [PubMed]
- Kuki, Y.; Ohno, R.; Yoshida, K.; Takumi, S. Heterologous expression of wheat WRKY transcription factor genes transcriptionally activated in hybrid necrosis strains alters abiotic and biotic stress tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2020, 150, 71–79. [Google Scholar] [CrossRef]
- Goyal, P.; Manzoor, M.M.; Vishwakarma, R.A.; Sharma, D.; Dhar, M.K.; Gupta, S. A Comprehensive Transcriptome-Wide Identification and Screening of WRKY Gene Family Engaged in Abiotic Stress in Glycyrrhiza glabra. Sci. Rep. 2020, 10, 373. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, S.S.; Khan, M.A.; Ammar, M.H.; Sun, Q.; Huang, L.; Migdadi, H.M.; El-Harty, E.H.; Al-Faifi, S.A. Characterization of drought stress-responsive root transcriptome of faba bean (Vicia faba L.) using RNA sequencing. 3 Biotech 2018, 8, 502. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Hu, Y.; Song, J.; Feng, H.; Wang, J.; Chen, L.; Wang, L.; Diao, X.; Wan, Y.; Liu, S.; et al. Transcriptome Sequencing and Metabolome Analysis Reveals the Molecular Mechanism of Drought Stress in Millet. Int. J. Mol. Sci. 2022, 23, 10792. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Song, L.; Xiao, Y.; Zhong, X.; Wang, J.; Xu, W.; Jiang, C.-Z. Overexpression of Myrothamnus flabellifolia MfWRKY41 confers drought and salinity tolerance by enhancing root system and antioxidation ability in Arabidopsis. Front. Plant Sci. 2022, 13, 967352. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Zhao, S.; Yu, X.; Sun, Y.; Li, H.; Ruan, C.; Li, J. Yellowhorn drought-induced transcription factor XsWRKY20 acts as a positive regulator in drought stress through ROS homeostasis and ABA signaling pathway. Plant Physiol. Biochem. 2020, 155, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.C.; Yeap, W.C.; Appleton, D.R.; Ho, C.-L.; Kulaveerasingam, H. Identification of drought responsive Elaeis guineensis WRKY transcription factors with sensitivity to other abiotic stresses and hormone treatments. BMC Genom. 2022, 23, 164. [Google Scholar] [CrossRef]
- Liang, Y.; Li, X.; Gu, Z.; Qin, P.; Ji, M. Toxicity of Amorphigenin from the Seeds of Amorpha fruticosa against the Larvae of Culex pipiens pallens (Diptera: Culicidae). Molecules 2015, 20, 3238–3254. [Google Scholar] [CrossRef]
- Dehaan, L.R.; Ehlke, N.J.; Sheaffer, C.C.; Wyse, D.L. Evaluation of diversity among north American accessions of false indigo( amorpha fruticosa L) for forage and biomass. Genetic Resources and Crop Evoluton 2006, 53, 1463–1476.. [Google Scholar] [CrossRef]
- Cui, X.; Guo, J.; Lai, C.-S.; Pan, M.-H.; Ma, Z.; Guo, S.; Liu, Q.; Zhang, L.; Ho, C.-T.; Bai, N. Analysis of bioactive constituents from the leaves of Amorpha fruticosa L. J. Food Drug Anal. 2016, 25, 992–999. [Google Scholar] [CrossRef] [PubMed]
- Muharini, R.; Díaz, A.; Ebrahim, W.; Mándi, A.; Kurtán, T.; Rehberg, N.; Kalscheuer, R.; Hartmann, R.; Orfali, R.S.; Lin, W.; et al. Antibacterial and Cytotoxic Phenolic Metabolites from the Fruits ofAmorpha fruticosa. J. Nat. Prod. 2016, 80, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Zheng, S.; Zhong, Y.; Shangguan, Z. Contrasting dynamics of leaf potential and gas exchange during progressive drought cycles and recovery in Amorpha fruticosa and Robinia pseudoacacia. Sci. Rep. 2017, 7, 4470. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Hu, S.; Wang, X.; Liu, H.; Zhou, Y.W.; Guan, Q. De novo assembly of Amorpha fruticosa L. transcriptome in response to drought stress provides insight into the tolerance mechanisms. PeerJ 2021, 9, e11044. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, A. WRKY Proteins: Signaling and Regulation of Expression during Abiotic Stress Responses. Sci. World J. 2015, 2015, 807560. [Google Scholar] [CrossRef]
- Khan, S.-A.; Li, M.-Z.; Wang, S.-M.; Yin, H.-J. Revisiting the Role of Plant Transcription Factors in the Battle against Abiotic Stress. Int. J. Mol. Sci. 2018, 19, 1634. [Google Scholar] [CrossRef] [PubMed]
- Kiranmai, K.; Rao, G.L.; Pandurangaiah, M.; Nareshkumar, A.; Reddy, V.A.; Lokesh, U.; Venkatesh, B.; Johnson, A.M.A.; Sudhakar, C. A Novel WRKY Transcription Factor, MuWRKY3 (Macrotyloma uniflorum Lam. Verdc.) Enhances Drought Stress Tolerance in Transgenic Groundnut (Arachis hypogaea L.) Plants. Front. Plant Sci. 2018, 9, 346. [Google Scholar] [CrossRef]
- Goyal, P.; Devi, R.; Verma, B.; Hussain, S.; Arora, P.; Tabassum, R.; Gupta, S. WRKY transcription factors: evolution, regulation, and functional diversity in plants. Protoplasma 2022, 260, 331–348. [Google Scholar] [CrossRef]
- Proietti, S.; Bertini, L.; Van der Ent, S.; Leon-Reyes, A.; Pieterse, C.M.J.; Tucci, M.; Caporale, C.; Caruso, C. Cross activity of orthologous WRKY transcription factors in wheat and Arabidopsis. J. Exp. Bot. 2010, 62, 1975–1990. [Google Scholar] [CrossRef]
- Li, X. Study on the mechanism of synergistic regulation of ABA signaling pathway by Arabidopsis WRKY20 and ABI5. Shandong Agricultural University [2023-07-03].
- Luo, X.; Bai, X.; Sun, X.; Zhu, D.; Liu, B.; Ji, W.; Cai, H.; Cao, L.; Wu, J.; Hu, M.; et al. Expression of wild soybean WRKY20 in Arabidopsis enhances drought tolerance and regulates ABA signalling. J. Exp. Bot. 2013, 64, 2155–2169. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, F.; Zhang, X.; Wang, W.; Sun, T.; Chen, Y.; Dai, M.; Yu, S.; Xu, L.; Su, Y.; et al. Expression Characteristics and Functional Analysis of the ScWRKY3 Gene from Sugarcane. Int. J. Mol. Sci. 2018, 19, 4059. [Google Scholar] [CrossRef]
- An, X.; Jin, G.; Luo, X.; Chen, C.; Li, W.; Zhu, G. Transcriptome analysis and transcription factors responsive to drought stress in Hibiscus cannabinus. PeerJ 2020, 8, e8470. [Google Scholar] [CrossRef] [PubMed]
- Khoso, M.A.; Hussain, A.; Ritonga, F.N.; Ali, Q.; Channa, M.M.; Alshegaihi, R.M.; Meng, Q.; Ali, M.; Zaman, W.; Brohi, R.D.; et al. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stresses in plants. Front. Plant Sci. 2022, 13, 1039329. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Sun, D.; Cen, H.; Xu, H.; Weng, H.; Yuan, F.; He, Y. Phenotyping of Arabidopsis Drought Stress Response Using Kinetic Chlorophyll Fluorescence and Multicolor Fluorescence Imaging. Front. Plant Sci. 2018, 9, 603. [Google Scholar] [CrossRef]
- Wang, C.-T.; Ru, J.-N.; Liu, Y.-W.; Yang, J.-F.; Li, M.; Xu, Z.-S.; Fu, J.-D. The Maize WRKY Transcription Factor ZmWRKY40 Confers Drought Resistance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2018, 19, 2580. [Google Scholar] [CrossRef]
- Xiong, C.; Zhao, S.; Yu, X.; Sun, Y.; Li, H.; Ruan, C.; Li, J. Yellowhorn drought-induced transcription factor XsWRKY20 acts as a positive regulator in drought stress through ROS homeostasis and ABA signaling pathway. Plant Physiol. Biochem. 2020, 155, 187–195. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2017, 46, D493–D496. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Chou, K.-C.; Shen, H.-B. Large-scale plant protein subcellular location prediction. J. Cell. Biochem. 2006, 100, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Chen, H.; Yang, Z.; Lu, J.; Wu, D.; Luo, Q.; Jia, J.; Tan, J. Identification of WRKY transcription factor family genes in Pinus massoniana Lamb. and their expression patterns and functions in response to drought stress. BMC Plant Biol. 2022, 22, 424. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; He, M.; Mei, E.; Zhang, B.; Tang, J.; Xu, M.; Liu, J.; Li, X.; Wang, Z.; Tang, W.; et al. WRKY53 integrates classic brassinosteroid signaling and the mitogen-activated protein kinase pathway to regulate rice architecture and seed size. Plant Cell 2021, 33, 2753–2775. [Google Scholar] [CrossRef] [PubMed]
- Bo, C.; Cai, R.; Fang, X.; Wu, H.; Ma, Z.; Yuan, H.; Cheng, B.; Fan, J.; Ma, Q. Transcription factor ZmWRKY20 interacts with ZmWRKY115 to repress expression of ZmbZIP111 for salt tolerance in maize. Plant J. 2022, 111, 1660–1675. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ajab, Z.; Liu, C.; Hu, S.; Liu, J.; Guan, Q. Overexpression of transcription factor SlWRKY28 improved the tolerance of Populus davidiana × P. bolleana to alkaline salt stress. BMC Genet. 2020, 21, 103. [Google Scholar] [CrossRef] [PubMed]
- Hellens, R.P.; Allan, A.C.; Friel, E.N.; Bolitho, K.; Grafton, K.; Templeton, M.D.; Karunairetnam, S.; Gleave, A.P.; Laing, W.A. Transient expression vectors for functional genomics, quantification of promoter activity and RNA silencing in plants. Plant Methods 2005, 1, 13. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Chunxia, L.; Hongyan, W. Establishment of a tissue culture and genetic transformation system for native tobacco. Shandong Sci. 2006, 19, 23–27. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Baczewska-Dąbrowska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M.; Wójcik-Jagła, M.; Pawluśkiewicz, B.; Bąba, W.; Brestic, M. Exploration of Chlorophyll a Fluorescence and Plant Gas Exchange Parameters as Indicators of Drought Tolerance in Perennial Ryegrass. Sensors 2019, 19, 2736. [Google Scholar] [CrossRef]
- Tsai, Y.-C.; Chen, K.-C.; Cheng, T.-S.; Lee, C.; Lin, S.-H.; Tung, C.-W. Chlorophyll fluorescence analysis in diverse rice varieties reveals the positive correlation between the seedlings salt tolerance and photosynthetic efficiency. BMC Plant Biol. 2019, 19, 403. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Nahakpam, S. Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars. Plant Physiol. Biochem. 2012, 57, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.; Liao, X.; He, M.; Li, X.; Wang, Z.; Ma, H.; Yu, S.; Liu, S. Tolerance analysis of chloroplast OsCu/Zn-SOD overexpressing rice under NaCl and NaHCO3 stress. PLOS ONE 2017, 12, e0186052. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Results of bioinformatics analysis of AfWRKY20. (A) Primary structure of the AfWRKY20 protein. (B) The secondary structure of the AfWRKY20 protein is depicted, with the α-helix shown in blue, the β-turn in green, the extended strand in red, and the irregular coil in purple. (C) Tertiary structure of the AfWRKY20 protein. (D) The AfWRKY20 motif consists of a conserved domain, with the two red dashed rectangular regions representing the two WRKY domains.
Figure 1.
Results of bioinformatics analysis of AfWRKY20. (A) Primary structure of the AfWRKY20 protein. (B) The secondary structure of the AfWRKY20 protein is depicted, with the α-helix shown in blue, the β-turn in green, the extended strand in red, and the irregular coil in purple. (C) Tertiary structure of the AfWRKY20 protein. (D) The AfWRKY20 motif consists of a conserved domain, with the two red dashed rectangular regions representing the two WRKY domains.
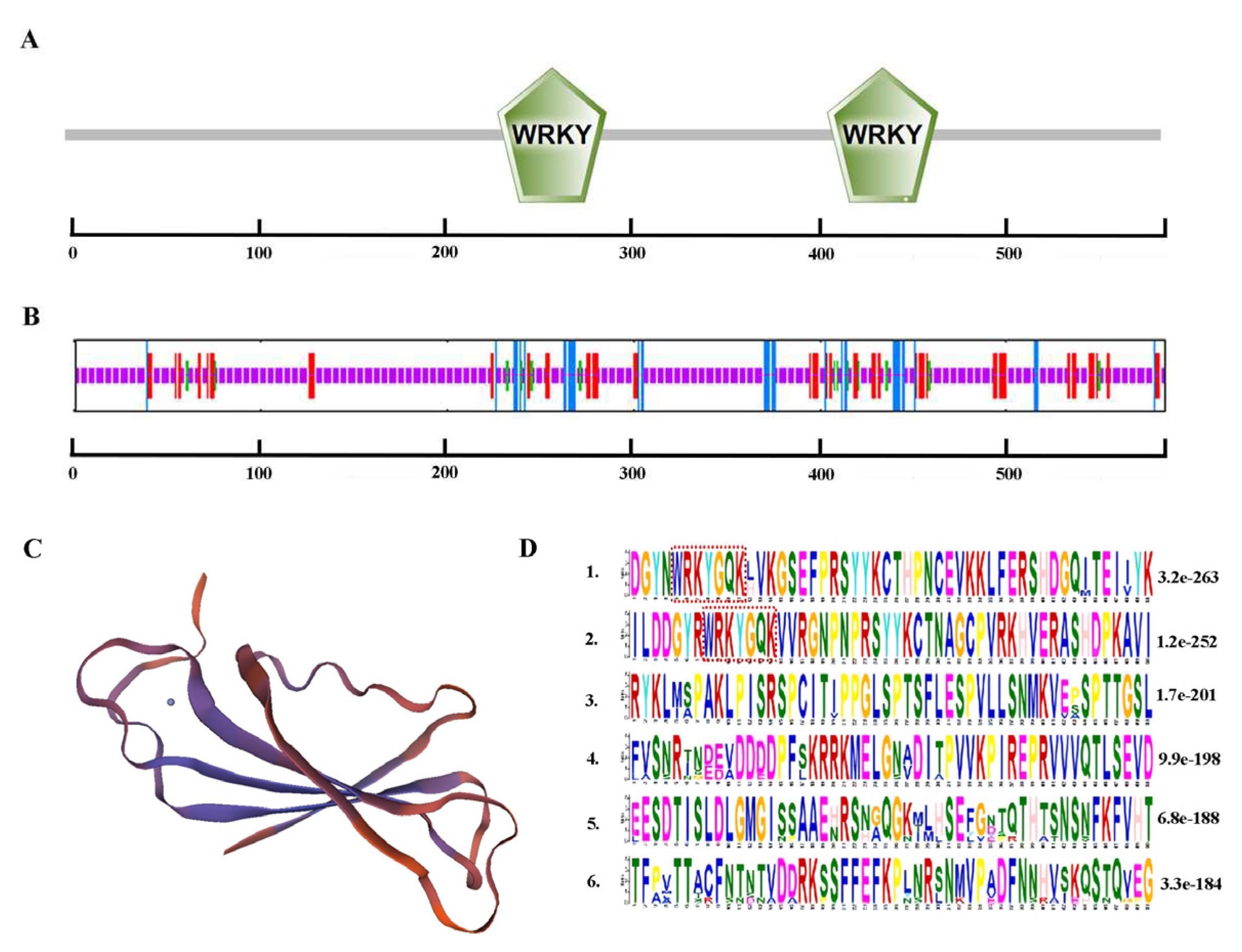
Figure 2.
Construction of phylogenetic trees for drought stress WRKY transcription factors in A. fruticosa and Arabidopsis WRKY transcription factors. The red triangle represents WRKY20 of A. fruticosa. Triangles indicate WRKY transcription factors in the A. fruticosa drought transcriptome. Circles indicate WRKY transcription factors in Arabidopsis. The classification of WRKY transcription factors is distinguished by different colors.
Figure 2.
Construction of phylogenetic trees for drought stress WRKY transcription factors in A. fruticosa and Arabidopsis WRKY transcription factors. The red triangle represents WRKY20 of A. fruticosa. Triangles indicate WRKY transcription factors in the A. fruticosa drought transcriptome. Circles indicate WRKY transcription factors in Arabidopsis. The classification of WRKY transcription factors is distinguished by different colors.
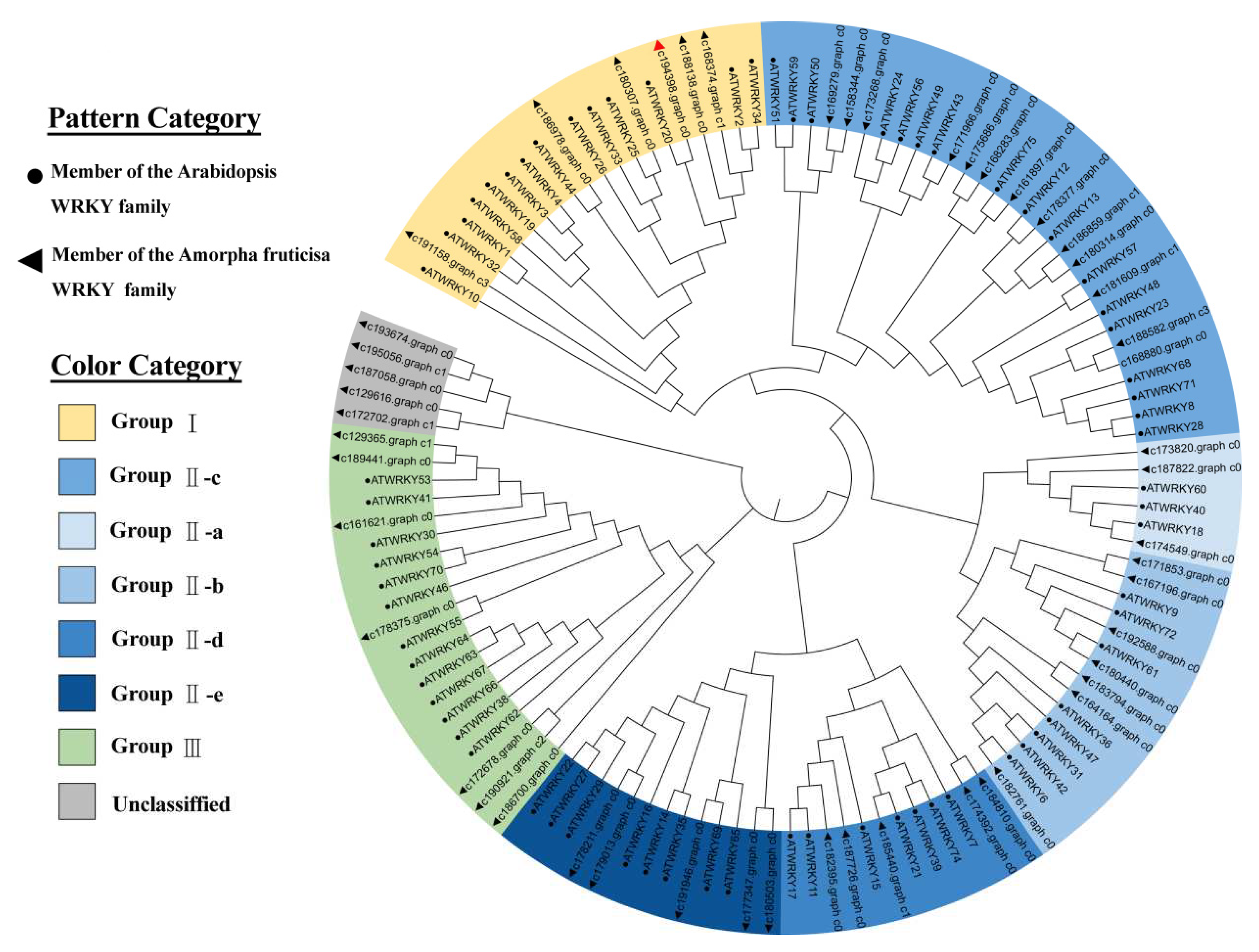
Figure 3.
(A) The amino acid sequences of the drought-resistant WRKY transcription factors of A. fruticosa WRKY20 and various species were compared using MEGA 7.0 software. The WRKY domain, C, and H residues in the zinc finger motif are represented by rectangles, triangles, and circles, respectively. The region of the red line represents WRKYGKK, which is not the regular WRKY domain motif (WRKYGQK). (B) Phylogenetic trees were constructed for A. fruticosa WRKY20 and drought-resistant WRKYs from various species. The red circles indicate AfWRKY20. The blue regions represent drought-resistant WRKY transcription factors with a very high homology to AfWRKY20. The yellow region represents the drought-resistant WRKY transcription factors that show homology to AfWRKY20.
Figure 3.
(A) The amino acid sequences of the drought-resistant WRKY transcription factors of A. fruticosa WRKY20 and various species were compared using MEGA 7.0 software. The WRKY domain, C, and H residues in the zinc finger motif are represented by rectangles, triangles, and circles, respectively. The region of the red line represents WRKYGKK, which is not the regular WRKY domain motif (WRKYGQK). (B) Phylogenetic trees were constructed for A. fruticosa WRKY20 and drought-resistant WRKYs from various species. The red circles indicate AfWRKY20. The blue regions represent drought-resistant WRKY transcription factors with a very high homology to AfWRKY20. The yellow region represents the drought-resistant WRKY transcription factors that show homology to AfWRKY20.
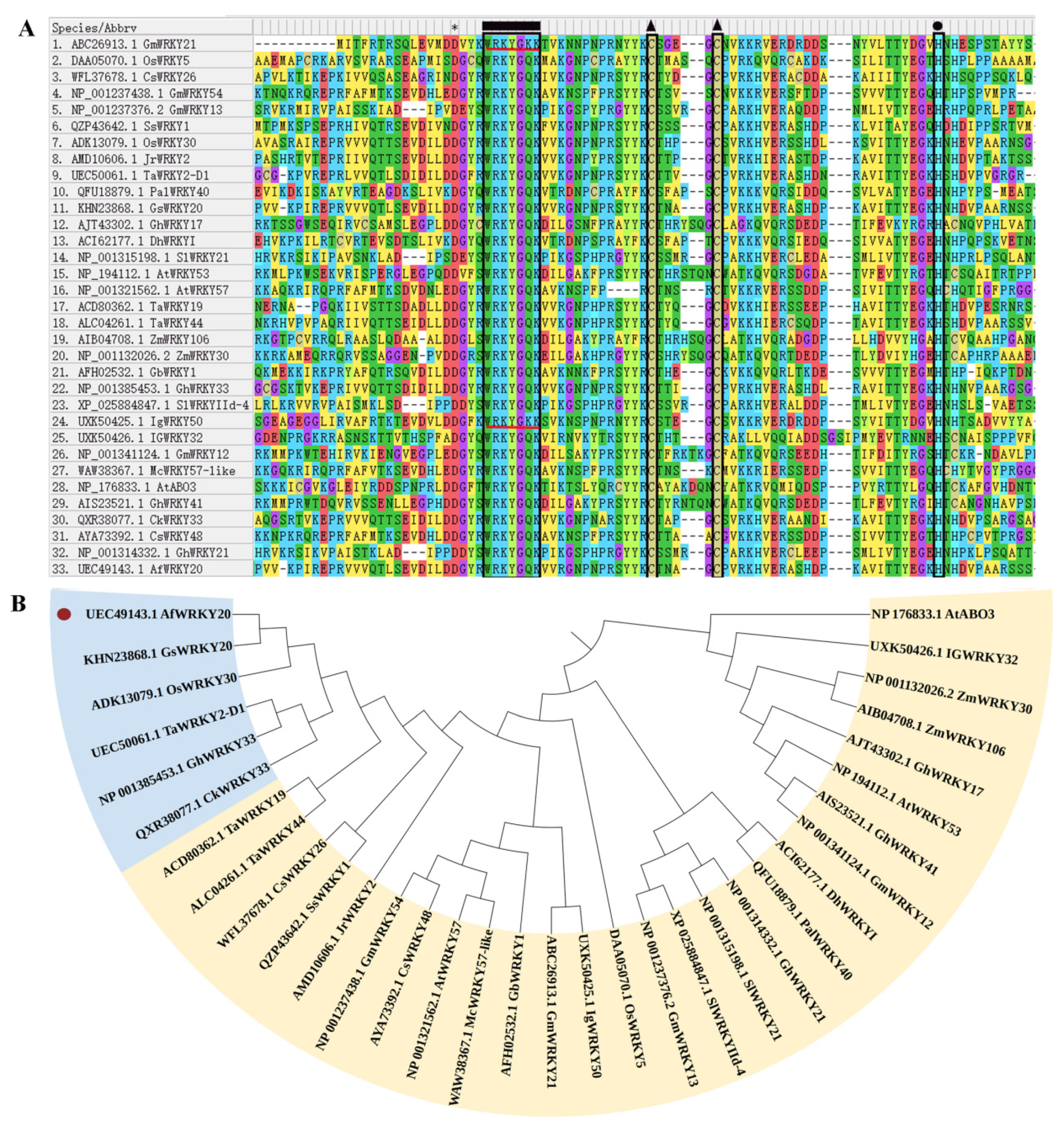
Figure 4.
Subcellular localization of AfWRKY20 in tobacco cells. 35s-GFP was used as the control group. 35s-AfWRKY20-GFP was used as the experimental group. Ruler: 20 μm.
Figure 4.
Subcellular localization of AfWRKY20 in tobacco cells. 35s-GFP was used as the control group. 35s-AfWRKY20-GFP was used as the experimental group. Ruler: 20 μm.
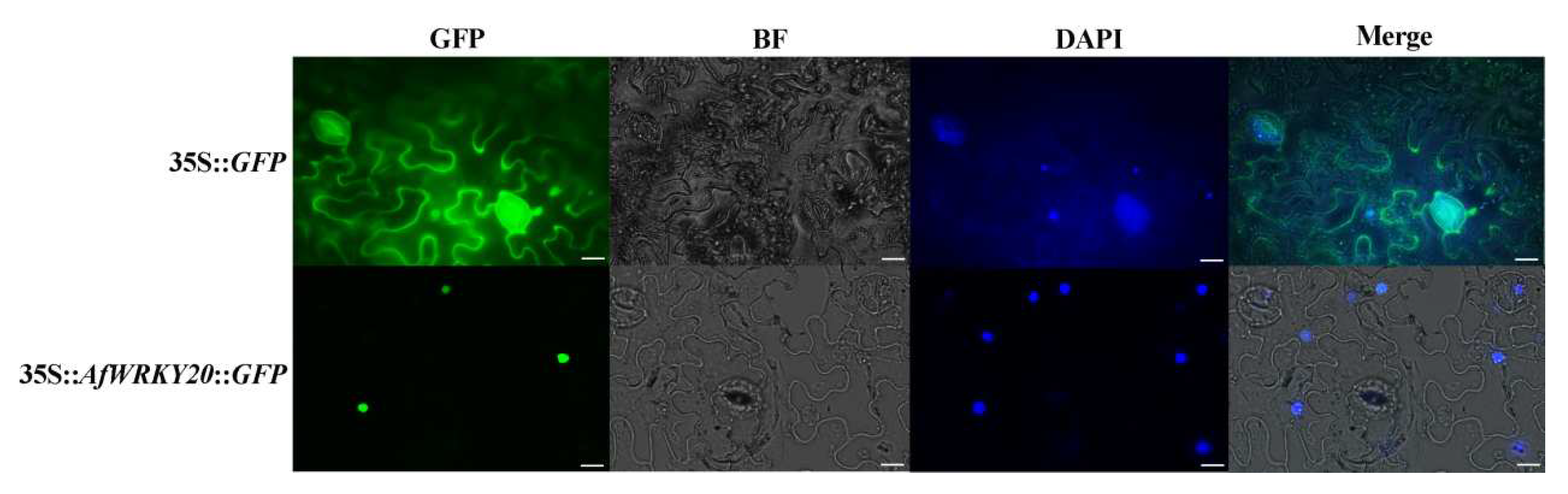
Figure 5.
(A) Binding of the GST-AfWRKY20 fusion protein to the W-box element. (1) GST+ protein and a biotin-labeled oligonucleotide probe (Biotin-Pr) were added; (2) Adding GST-AfWRKY20 fusion protein and biotin-Pr; (3) Adding GST-AfWRKY20 fusion protein, biotin-Pr and unlabeled oligonucleotide probe (Cold-Pr). (B) Dual-LUC analysis of W-box and mW-box elements.
Figure 5.
(A) Binding of the GST-AfWRKY20 fusion protein to the W-box element. (1) GST+ protein and a biotin-labeled oligonucleotide probe (Biotin-Pr) were added; (2) Adding GST-AfWRKY20 fusion protein and biotin-Pr; (3) Adding GST-AfWRKY20 fusion protein, biotin-Pr and unlabeled oligonucleotide probe (Cold-Pr). (B) Dual-LUC analysis of W-box and mW-box elements.
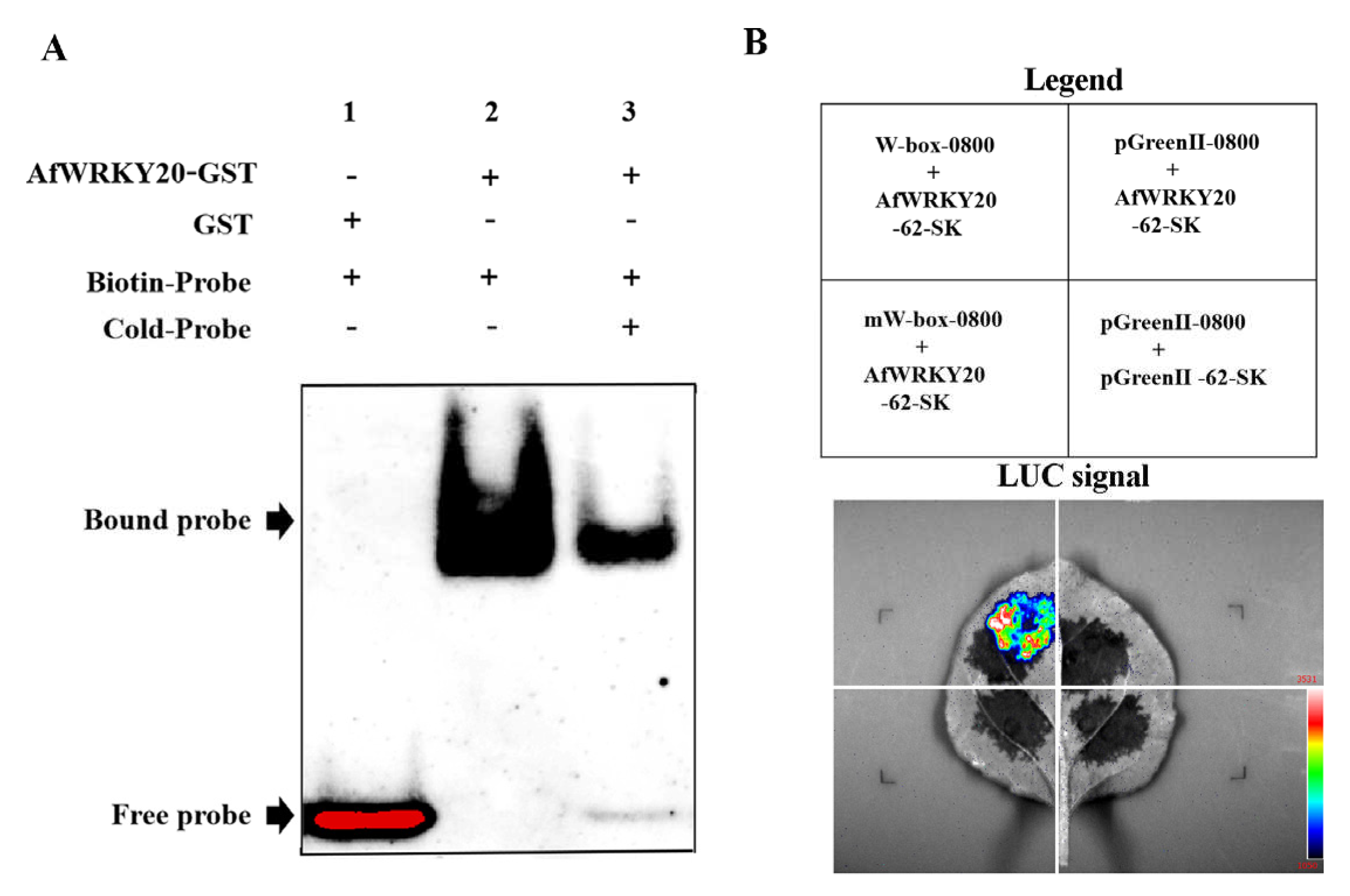
Figure 6.
Expression characteristics of the AfWRKY20 gene in tissues and organs under PEG6000-induced stress. (A) Tissues and organs: roots, stems, leaves, flowers, and peel. (B) Expression of AfWRKY20 in leaves and roots under 0%, 10%, 20%, and 30% PEG6000 stress. (C) Expression of the AfWRKY20 gene in leaves under 30% PEG6000 stress at 0, 6, 12, 24, and 48 hours. (D) Expression of the AfWRKY20 gene in roots under 30% PEG6000 stress at 0, 6, 12, 24, and 48 hours. (E) Expression of the AfWRKY20 gene in leaves exposed to 150 mmol/L NaCl stress at 0, 6, 12, 24, and 48 hours. (F) Expression of the AfWRKY20 gene in roots under 150 mmol/L NaCl stress at 0, 6, 12, 24, and 48 hours. (G) Expression of the AfWRKY20 gene in leaves exposed to 30 mmol/L NaHCO3 stress at 0, 6, 12, 24, and 48 hours. (H) Expression of the AfWRKY20 gene in roots under 30 mmol/L NaHCO3 stress at 0, 6, 12, 24, and 48 hours. Error bars represent standard errors of three biological replicates, with significant differences at the p<0.05 level.
Figure 6.
Expression characteristics of the AfWRKY20 gene in tissues and organs under PEG6000-induced stress. (A) Tissues and organs: roots, stems, leaves, flowers, and peel. (B) Expression of AfWRKY20 in leaves and roots under 0%, 10%, 20%, and 30% PEG6000 stress. (C) Expression of the AfWRKY20 gene in leaves under 30% PEG6000 stress at 0, 6, 12, 24, and 48 hours. (D) Expression of the AfWRKY20 gene in roots under 30% PEG6000 stress at 0, 6, 12, 24, and 48 hours. (E) Expression of the AfWRKY20 gene in leaves exposed to 150 mmol/L NaCl stress at 0, 6, 12, 24, and 48 hours. (F) Expression of the AfWRKY20 gene in roots under 150 mmol/L NaCl stress at 0, 6, 12, 24, and 48 hours. (G) Expression of the AfWRKY20 gene in leaves exposed to 30 mmol/L NaHCO3 stress at 0, 6, 12, 24, and 48 hours. (H) Expression of the AfWRKY20 gene in roots under 30 mmol/L NaHCO3 stress at 0, 6, 12, 24, and 48 hours. Error bars represent standard errors of three biological replicates, with significant differences at the p<0.05 level.
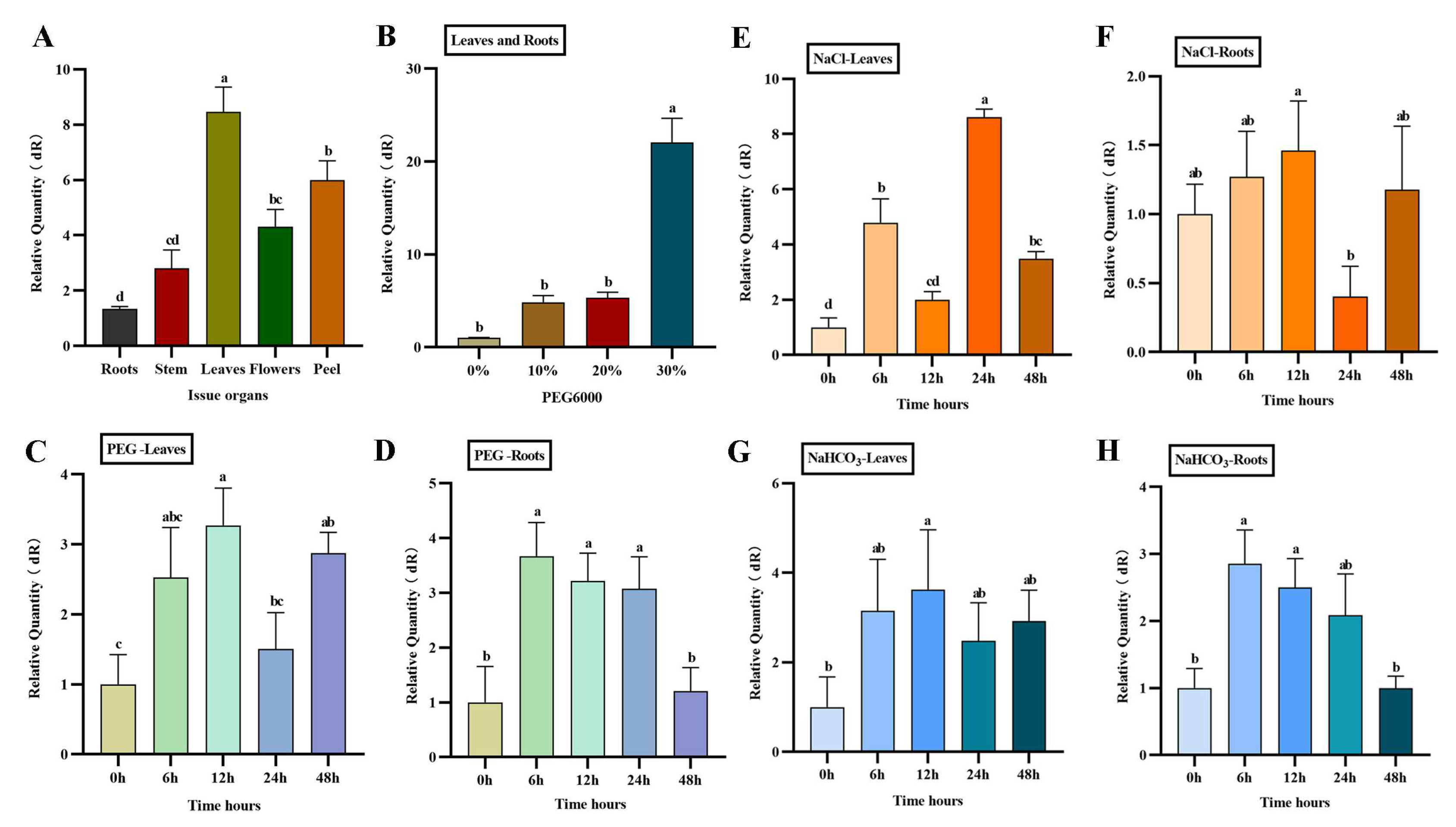
Figure 7.
Analysis of the green leaf rate and germination rate of AfWRKY20 overexpression lines under sorbitol stress at various concentrations. (A) Sorbitol stress phenotypes at different concentrations; (B) Measurement of the percentage of green leaves. (C) Measurement of germination rate. Error bars indicate the standard errors of three biological replicates, which are significantly different at the p < 0.05 level.
Figure 7.
Analysis of the green leaf rate and germination rate of AfWRKY20 overexpression lines under sorbitol stress at various concentrations. (A) Sorbitol stress phenotypes at different concentrations; (B) Measurement of the percentage of green leaves. (C) Measurement of germination rate. Error bars indicate the standard errors of three biological replicates, which are significantly different at the p < 0.05 level.
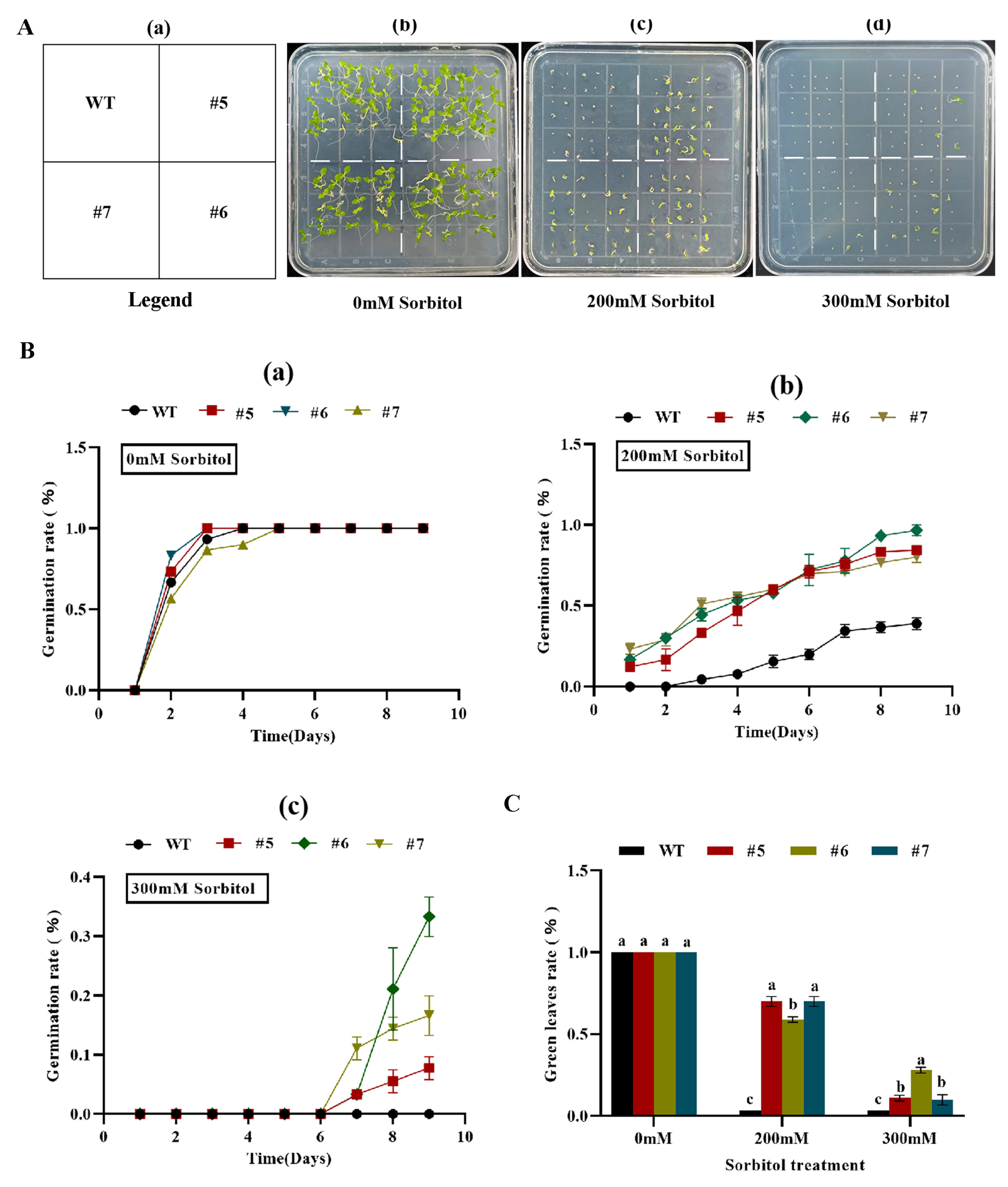
Figure 8.
Analysis of fresh weight and root length of AfWRKY20 overexpression tobacco lines under sorbitol stress at various concentrations.(A) The phenotypes of tobacco under sorbitol stress. (B) Measurement of fresh weight of plants. (C) Measurement of plant root length.
Figure 8.
Analysis of fresh weight and root length of AfWRKY20 overexpression tobacco lines under sorbitol stress at various concentrations.(A) The phenotypes of tobacco under sorbitol stress. (B) Measurement of fresh weight of plants. (C) Measurement of plant root length.
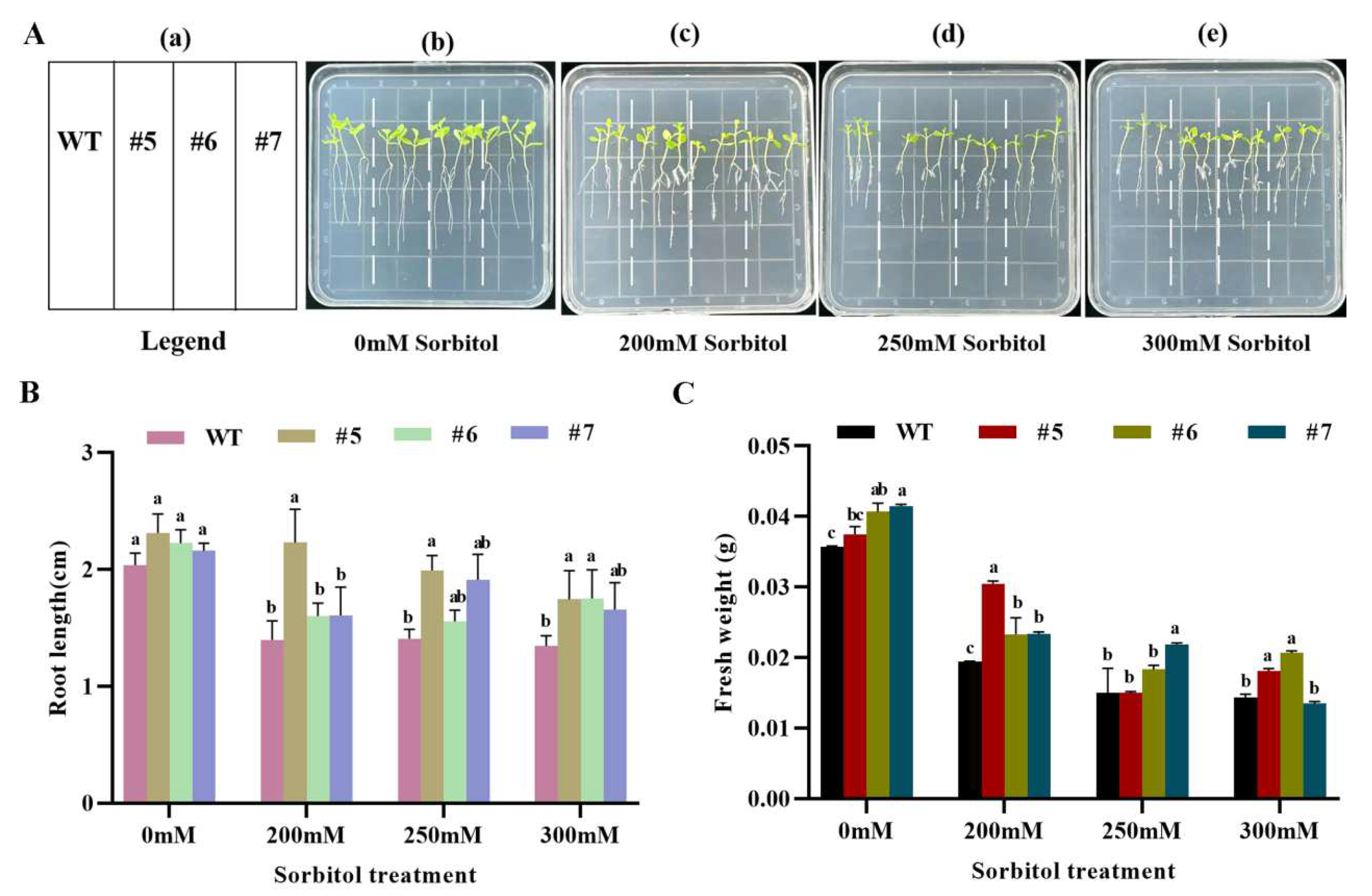
Figure 9.
Analysis of photosynthetic characteristics of AfWRKY20 transgenic tobacco pot seedlings under natural drought stress. (A) Phenotyping of tobacco plants after 0, 10, and 15 days of natural drought treatment, followed by 3 days of rehydration. (B) Images of chlorophyll fluorescence Fv-Fm in transgenic tobacco and the WT tobacco under drought stress treatment, with a cursor range of 0.2-0.8. (C) Fv/Fm values of WT and transgenic tobacco were measured at 0, 10, and 15 days of drought treatment, as well as 3 days after rehydration. (D) Survival rate of tobacco plants after 3 days of rehydration.
Figure 9.
Analysis of photosynthetic characteristics of AfWRKY20 transgenic tobacco pot seedlings under natural drought stress. (A) Phenotyping of tobacco plants after 0, 10, and 15 days of natural drought treatment, followed by 3 days of rehydration. (B) Images of chlorophyll fluorescence Fv-Fm in transgenic tobacco and the WT tobacco under drought stress treatment, with a cursor range of 0.2-0.8. (C) Fv/Fm values of WT and transgenic tobacco were measured at 0, 10, and 15 days of drought treatment, as well as 3 days after rehydration. (D) Survival rate of tobacco plants after 3 days of rehydration.
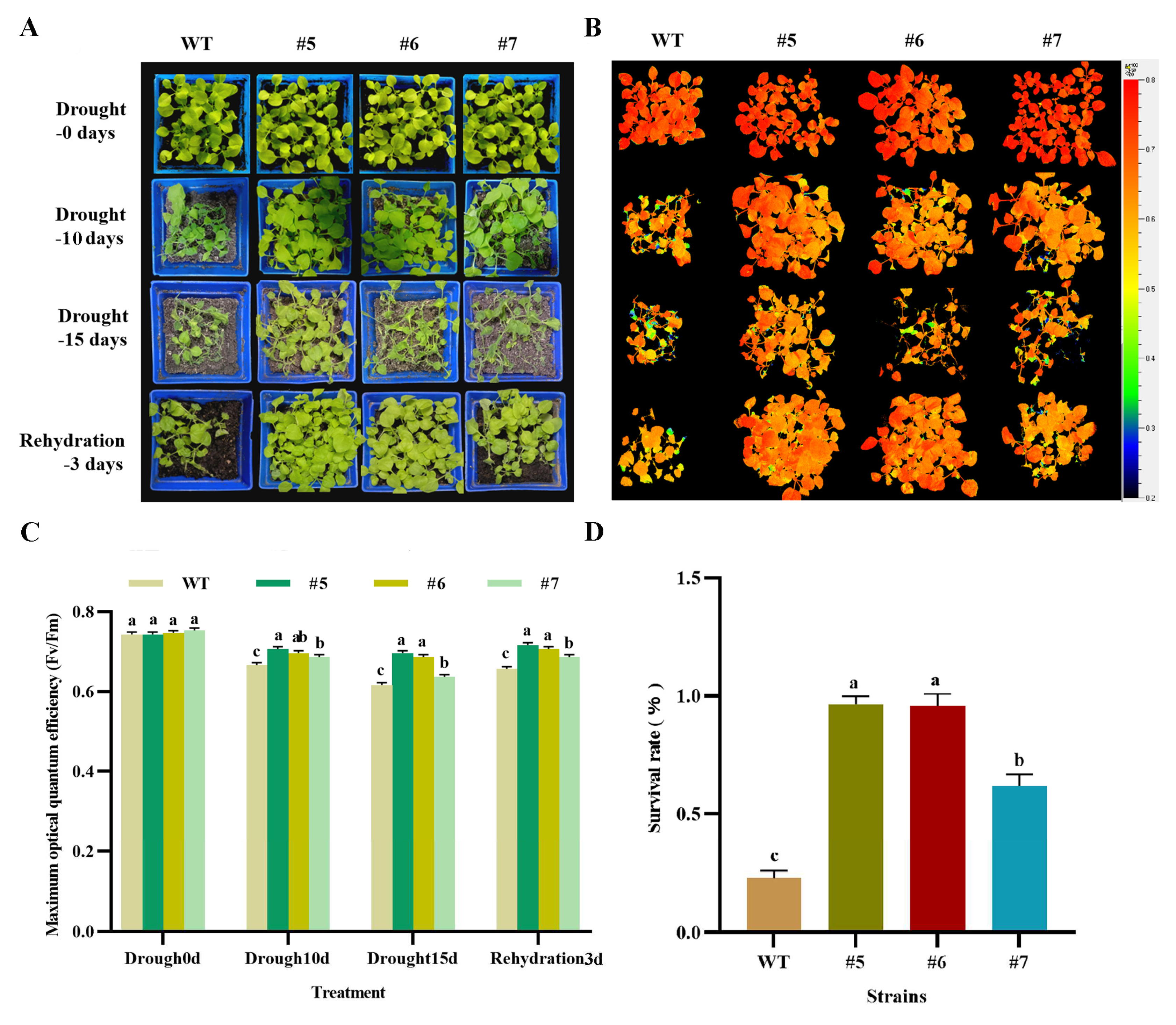
Figure 10.
Phenotypic changes and photosynthetic characteristics of AfWRKY20 transgenic tobacco under natural drought stress. (A) Phenotype of tobacco on 0 day of drought treatment. (B) Phenotypes of tobacco plants after 10 days of drought treatment. (C) Phenotypes of tobacco plants after 15 days of drought treatment. (D) The phenotypes of tobacco after 3 days of rewatering; (E) The Fv/Fm values comparing AfWRKY20 overexpression tobacco with the WT tobacco at 0 day of drought treatment, 15 days of drought treatment, and 3 days after rewatering. (F) Changes in chlorophyll content in tobacco during drought treatment and subsequent rewatering.
Figure 10.
Phenotypic changes and photosynthetic characteristics of AfWRKY20 transgenic tobacco under natural drought stress. (A) Phenotype of tobacco on 0 day of drought treatment. (B) Phenotypes of tobacco plants after 10 days of drought treatment. (C) Phenotypes of tobacco plants after 15 days of drought treatment. (D) The phenotypes of tobacco after 3 days of rewatering; (E) The Fv/Fm values comparing AfWRKY20 overexpression tobacco with the WT tobacco at 0 day of drought treatment, 15 days of drought treatment, and 3 days after rewatering. (F) Changes in chlorophyll content in tobacco during drought treatment and subsequent rewatering.
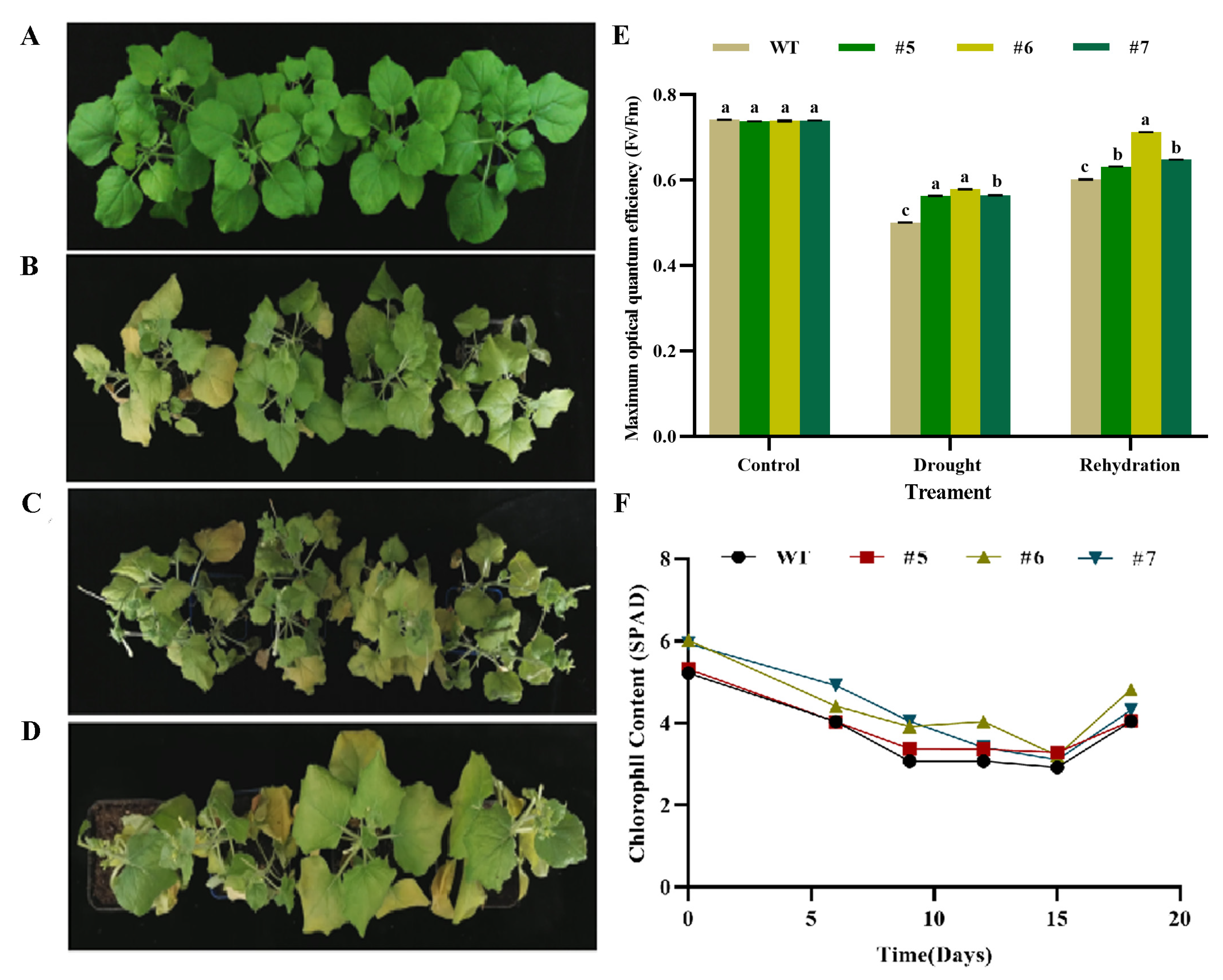
Figure 11.
Analysis of oxygen free radicals in tobacco plants under natural drought conditions. (A) Histochemical analysis of DAB and NBT in tobacco (B) Determination of the SOD content in tobacco under natural drought conditions. (C) Determination of MDA content in tobacco under natural drought conditions. (D) Real-time quantitative expression of NbSOD during drought stress. (E) Real-time quantitative expression of NbPOD during drought stress. (F) Real-time quantitative expression of NbCAT during drought stress.
Figure 11.
Analysis of oxygen free radicals in tobacco plants under natural drought conditions. (A) Histochemical analysis of DAB and NBT in tobacco (B) Determination of the SOD content in tobacco under natural drought conditions. (C) Determination of MDA content in tobacco under natural drought conditions. (D) Real-time quantitative expression of NbSOD during drought stress. (E) Real-time quantitative expression of NbPOD during drought stress. (F) Real-time quantitative expression of NbCAT during drought stress.
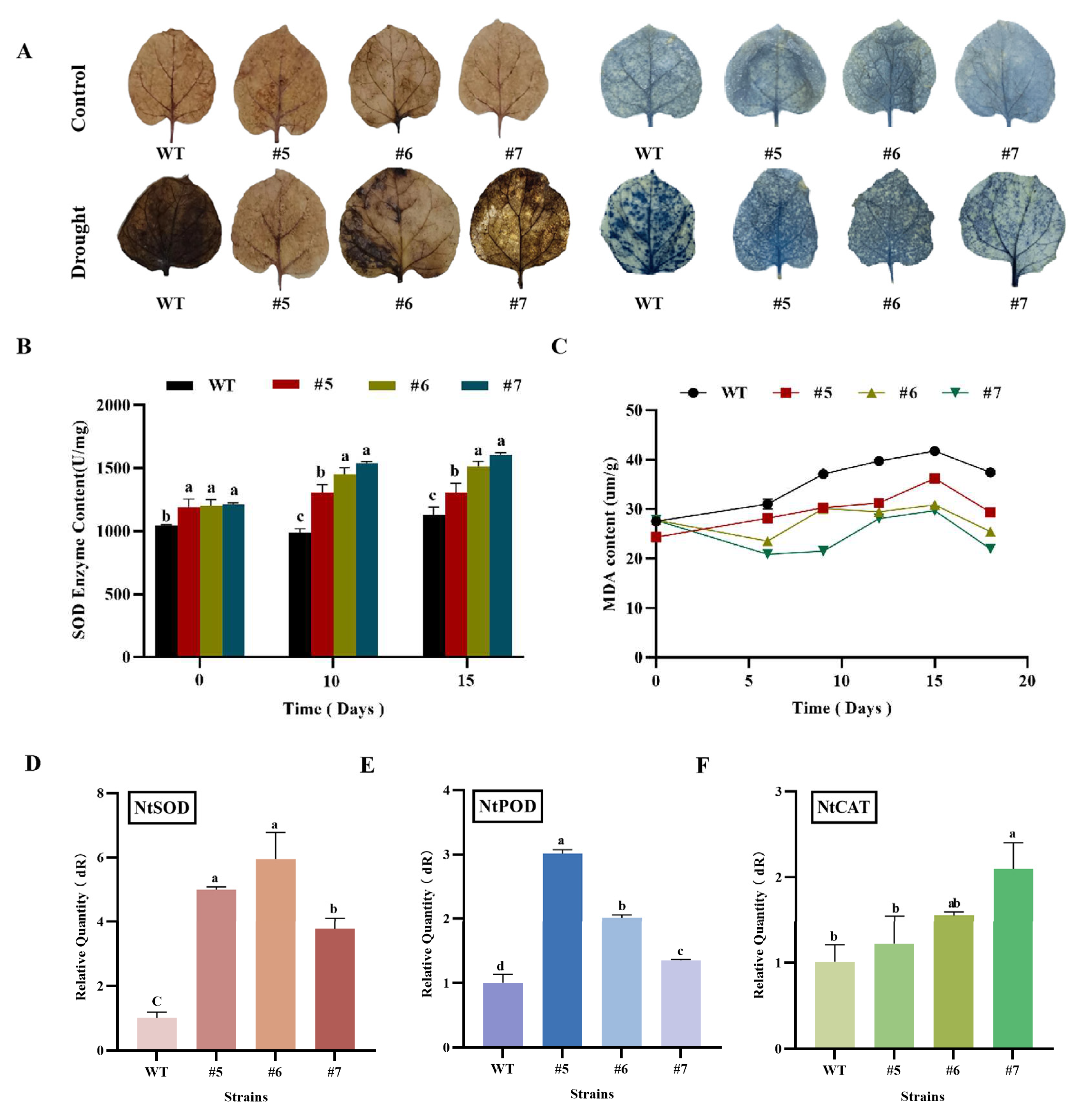
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



