
Submitted:
17 July 2023
Posted:
18 July 2023
You are already at the latest version
Abstract
Human rhabdomyosarcomas are rarely cured by surgical resection alone. This is also true for high grade soft tissue sarcomas in dogs. Dogs with spontaneous sarcoma are good models for clinical responses to new cancer therapies. Strategic combinations of immunotherapy and oncolytic virotherapy (OV) could improve treatment responses in canine and human cancer patients. To develop an appropriate combination of immunotherapy and OV for dogs with soft tissue sarcoma (STS), canine cancer cells were inoculated with myxoma viruses (MYXVs) and gene transcripts were quantified. Next, cytokine concentrations in canine cancer cells were altered to evaluate their effect on MYXV replication. These studies indicated that, as in murine and human cells, type I interferons (IFN) play an important role in limiting MYXV replication in canine cancer cells. To reduce type I IFN production during OV, oclacitinib (a JAK1 inhibitor) was administered twice daily to dogs for 14 days starting ~7 days prior to surgery. STS tumors were excised and MYXV deleted for serp2 (MYXV∆SERP2) was administered at the surgical site at two time points post-operatively to treat any remaining microscopic tumor cells. Tumor regrowth in dogs treated with OV was decreased relative to historical controls. However, regrowth was not further inhibited in patients given combination therapy.
Keywords:
cytokines
; oclacitinib
; oncolytic virus
; dogs
; cancer
1. Introduction
Dogs that have spontaneous cancers are excellent models of human response to therapy and directly benefit from new treatment options, as effective treatments are often limited for canine patients [1]. For this study, dogs with spontaneous soft tissue sarcomas were recruited to determine if oclacitinib (Apoquel®, Zoetis, Parsippany, NJ, USA) effected treatment outcomes following surgical excision and post-operative treatment with an oncolytic virus (OV).
Soft tissue sarcomas arise from neoplastic mesenchymal cells within connective tissue components of the body. If complete excision of the STS cannot be achieved, tumor regrowth is expected within approximately one year [2,3]. Frequently, radiation and/or chemotherapy are recommended after surgery to slow the rate of tumor regrowth, but these treatments often do not prevent recurrence [4]. Discovery of alternative therapies, including OV, is important to improve outcomes in patients with STS.
Currently, two OVs are approved for use in people [5,6] but no OVs are available for dogs. Oncolytic poxviruses and herpes simplex virus cause cell death in canine sarcoma cells lines and reduce tumor growth rates in mice bearing canine sarcoma xenografts [7,8,9]. Additionally, there is evidence that OVs cause very few adverse clinical events in dogs [10,11,12,13,14,15,16,17,18,19]. These data indicate that OVs could be beneficial to dogs with STS. In most clinical studies, response to OVs is variable, so OVs are used in combination with other cancer treatments.
It is well established that healthy murine and human cells are stimulated to produce type I IFNs when they are infected with a virus [20,21]. Upregulation of type I IFNs in healthy cells then inhibits viral replication within the infected cells [20]. This is generally believed to happen in many different species and a significant amount of data has shown that this occurs in dogs infected with viruses that are canine pathogens [22]. However, neoplastic murine and human cells often lack an appropriate type I IFN response to virus infection, which allows MYXV replication to occur [23,24]. To our knowledge, the IFN response to virus infection has not been determined in canine cancer cells. Several canine cell cultures were used to evaluate the effects of innate cytokines on MYXV infection, including two soft tissue sarcoma cell cultures.
Type I IFNs have both autocrine and paracrine effects. Secreted type I IFNs bind to the IFNα receptor complex that activates Janus kinase 1 (JAK1) and tyrosine kinase 2 (Tyk2). This activates signal transducer and activator of transcription (STAT) proteins, which translocate to the nucleus. The canonical pathway involves an activated STAT1, STAT2, and IFN-regulatory factor 9 complex. The complex binds to IFN-stimulated response elements in the nucleus and drives anti-viral cellular response [25]. Alternative type I IFN pathways signal cells through gamma activated sequences [26] to produce proteins that drive proliferation and could promote tumor invasion, metastasis, and angiogenesis [27]. Theoretically, type I IFN pathway inhibition could reduce both the anti-viral responses and cell proliferation in tumors.
Oclacitinib is a JAK1 and JAK2 inhibitor that is used to treat allergic disease in dogs by reducing serum IL-31 concentration and decreasing pruritis [28]. Interleukin-6 (IL-6) and IL-13 (which signal through JAK1/2 and Tyk2) are inhibited by oclacitinib therapy in dogs [29]. Since type I IFN signaling occurs through JAK1 and Tyk2 pathways, oclacitinib also is predicted to inhibit IFNα and IFNβ production. Importantly for OV, oclacitinib does not inhibit patient response to vaccination because its inhibitory activity is relatively poor against JAK3 and Tyk2, which are involved in signaling pathways that drive a T helper 1 (cell-mediated) immune response [28,29,30]. Therefore, this drug could limit type I IFN production and allow viral replication to occur in the tumor for a longer period of time while the virus continues to stimulate an anti-tumoral immune response.
We chose to combine oclacitinib treatment with MYXV∆SERP2 therapy in dogs with STS. Myxoma virus is an oncolytic poxvirus that only causes disease in Lagomorphs [31,32,33,34,35,36,37,38] but replicates in murine and human cancer cells that lack appropriate anti-viral IFN responses [23,24]. The data presented in this study indicate that this mechanism also occurs in canine cancer cells. MYXVΔSERP2 was used to treat dogs with STS instead of MYXV because MYXVΔSERP2 is attenuated in rabbits [39] and has improved oncolytic effects in many feline [40] and canine [41] cancer cells. Previous studies indicated that MYXVΔSERP2 treatment is safe in dogs with STS but viral replication lasts < 4 days after injection into tumors [18,42]. We hypothesized that MYXV∆SERP2 oncolytic effects could be improved by pre-treating canine STS patients with oclacitinib to suppress the antiviral IFN immune response before MYXVΔSERP2 was administered.
The goal of this study was to determine the effect of cytokines on replication of myxoma virus (MYXV) in canine cancer cells. The knowledge gained in cell cultures supported testing the use of oclacitinib (a JAK1 inhibitor) to improve outcomes in dogs with high-grade soft tissue sarcomas (STS) that were treated post-operatively with MYXV deleted for serp2 (MYXV∆SERP2). We hypothesized that oclacitinib would inhibit type I interferon (IFN) responses and prolong MXYV∆SERP2 replication within STS cells that were not surgically removed thereby limiting the rate of tumor regrowth better than MYXV∆SERP2 treatment alone.
2. Materials and Methods
2.1. Recombinant Viruses
The Lausanne strain of wild-type myxoma virus (MYXV) and MYXV expressing tandem dimer tomato red (MYXV-red; vMyx-tdTr) [43] were generous gifts from Dr. Grant McFadden. Myxoma virus deleted for serp2 and expressing lacZ (MYXV∆SERP2, MYXV∆serp2::lacZ) was constructed in the Moyer laboratory [39]. Prior to injection into dogs, MYXVΔSERP2 was sucrose pad purified, titered, and diluted to 106 foci-forming units (ffu) of MYXV∆SERP2 per mL phosphate-buffered saline (PBS) as previously described [9]. Virus titers were calculated as ffu/mL using standard plaque assays. Plaque assays were performed in 12-well plates (Thermo Fisher Scientific, Nunc, Roskilde, Denmark) containing confluent rabbit kidney epithelial (RK-13, ATTC CCL-37) cells. Cells were inoculated with 10-fold dilutions of samples containing virus then placed in a water-jacketed incubator at 5% CO2 and 37˚C for one-hour. After incubation, cells were overlaid with 0.5% agarose (Lonza Group Ltd., Basel, Switzerland) and growth media consisting of minimal essential media supplemented with 2 mM glutamine, 50 U/mL penicillin, and 50 µg/mL streptomycin (GE Healthcare, Marlborough, MA, USA), 1 mM sodium pyruvate and 0.1 mM non-essential amino acids (Corning, Corning, NY, USA), and 10% fetal bovine serum (FBS; VWR Life Science Seradigm, Radnor, PA, USA). Cells were incubated for ~5 days then viral foci that formed in the cell monolayers were visualized and counted. Foci counts were multiplied by the dilution factor and divided by the volume of inoculum to calculate the virus titer (ffu/mL) in the sample.
2.2. Canine Cell Cultures
2.2.1. Cell isolation and Validation
Primary soft tissue sarcoma (STS) cancer cells were derived from spontaneous tumors surgically removed from two dogs that were cancer patients at different Veterinary Teaching Hospitals; STS-1 cells were isolated at the University of Illinois and STS-2 cells were isolated at Colorado State University (CSU). Fibroblasts (FB) were cultured posthumously from the linea alba of an 11-year-old female dog prior to necropsy at CSU. Data using STS-1 (a.k.a. STSA-1) cells has been published previously and include the effects of MYXV and MYXVΔSERP2 inoculation in cell culture [41], growth of the cells in a murine xenograft model [8], and identification of genetic modifications associated with the cancerous cells [44].
To isolate and culture the cells, a 1 cm3 section of tumor or linea was placed in growth media (described in subsection 2.1.) in a 35 × 10 mm cell culture dish (Corning, Corning, NY, USA). The tissue was homogenized manually using two 22-gauge needles and placed in a water-jacketed incubator at 5% CO2 and 37˚C. Media was changed daily.
Short Tandem Repeat (STR) analysis to ensure each cell culture was unique and of canine origin and Mycoplasma spp. testing were performed by the Cell Line Validation Core at CSU.
Cytochemical staining for vimentin (Clone AP20, BioCare Medical, Pacheco, CA, USA) and cytokeratin (AE1/AE3+8/18, BioCare Medical) was performed to validate that cells in culture produced proteins that corresponded to the tumor type they were isolated from using previously described methods [41].
2.2.2. Cell Permissivity to MYXV
MYXV-red replication in cells was determined by calculating virus ffu/mL (described in subsection 2.1.) at several hours post-inoculation (hpi) and constructing virus growth curves [45].
2.2.3. Cytotoxicity of MYXV
Cytotoxic effects of MYXV-red inoculation (0.1 moi) were observed in 48-well plates (Corning, Corning, NY, USA) of confluent cells using a Leica DMI4000B inverted microscope. Expression of the reporter protein tdTr (encoded by MYXV-red) was detected in MYXV-red-inoculated cell cultures using a 560/40 nm bandpass excitation filter associated with the microscope.
Cell viability was measured in canine cells cultured to confluence in 96-well plates (Corning, Corning, NY, USA) and inoculated with MYXV (0.1 moi) using a CellTiter-Glo 2.0 Cell Viability Assay (Promega, Madison, WI, USA) according to the manufacturer’s protocols. At 24, 48, and 72 hpi, cell viability data from mock-infected and MYXV-inoculated cells were compared. Data were analyzed using unpaired, nonparametric t-tests (Mann-Whitney tests) in GraphPad Prism version 9.1.0 software (San Diego, CA, USA).
2.3. Evaluation of Cytokine Effects on Viral Infection of Canine Cells
2.3.1. Measurement of mRNAs
2.3.1.1. Canine Immuno-Oncology Panel
A nCounter® Canine IO Panel (NanoString Technologies, Seattle, WA, USA) was used to identify and count transcripts for 800 canine genes (including 20 genes for data normalization). This technology does not require PCR amplification of transcripts.
Primary canine cells STS-1, STS-2, and FB were grown to confluency in 6-well plates (Corning, Corning, NY, USA) and inoculated with wild-type MYXV or MYXVΔSERP2 by removing the growth media (described in subsection 2.1.) and incubating cells with virus (0.1 moi) in media lacking FBS. After incubation in a water-jacketed incubator at 5% CO2 and 37˚C for one-hour, growth media (described in subsection 2.1.) was placed on the cells. Cells were harvested for mRNA 4 and 8 hpi using the Monarch Total RNA Miniprep kit (New England Biolabs, Ipswich, MA, USA) according to the manufacturer’s instructions, including removal of genomic DNA using gDNA Removal Columns provided with the kit.
RNA concentration was determined by measuring absorbance at 260 nm using a 0.5 mm pathlength [ssRNA ng/µL = optical density units (10 mm/0.5 mm) × 40 µg/mL]. RNA purity was estimated by calculating A260/280 and A260/230 ratios. Samples were sent to the University of Arizona Genetics Core (Tucson, AZ, USA) for further evaluation of concentration and purity and for NanoString analysis. They performed an RNA High Sensitivity Qubit quantitative read and an RNA High Sensitivity Fragment Analyzer quality assay, and provided an RNA quality control (QC) summary. They reviewed the NanoString data and reported that “…there are no QC flags and the counts show robustness and some nice diversity. The samples were run targeting between 125 ng and 275 ng in the assay. They were all run with 19-hour 65c hybridizations.”
NanoString data were analyzed using nSolver 4.0 (NanoString Technologies, Seattle, WA, USA) and Rosalind version 3.37.1.0 (Rosalind, San Diego, CA, USA) bioinformatics software.
2.3.1.2. Quantitative PCR
Primary canine cells STS-1 and FB were grown to confluency in 48-well plates (Corning, Corning, NY, USA) and inoculated with wild-type MYXV or MYXVΔSERP2 by removing the growth media (described in subsection 2.1.) and incubating cells with virus (0.1 moi) in media lacking FBS. After incubation in a water-jacketed incubator at 5% CO2 and 37˚C for one-hour, growth media (described in subsection 2.1.) was placed on the cells. Cells were harvested for mRNA 4 and 24 hpi using the RNeasy® QIAGEN kit according to the manufacturer’s instructions. The optional steps to digest DNA were performed. Moloney murine leukemia virus reverse transcriptase with oligonucleotide (DT12-18) primers (Invitrogen, Carlsbad, CA, USA), and 10 mM deoxynucleoside triphosphate (New England Biolabs, Ipswitch , MA, USA) were used to generate cDNA according to the manufacturer’s instructions. Primers (2 µM) against β-actin (housekeeping gene), interferon α (IFNα), IFNβ, tumor necrosis factor α (TNFα), interleukin-1β (IL-1β), IFNγ, IL-4, IL-10, and transforming growth factor β (TGFβ) were combined with 2× SsoFast™ Evagreen® Supermix (BioRad, Hercules, CA, USA) and 5 µL of sample. Primer sequences are listed in Table 1. Cytokine concentrations were determined using a quantitative PCR (qPCR) thermocycler (Applied Biosystems, QuantStudio, Thermo Fisher Scientific, Waltham, MA, USA) under the following amplification conditions: 94˚C for 1 min, followed by 40 cycles of 95˚C for 15 sec, 60˚C for 30 sec, a final melting curve at 95˚C for 15 min, and then 60˚C for 15 sec. Relative quantification (RQ) values were determined using the ΔΔCT method [46]. Eight data points were recorded per group.
2.3.2. Addition of Cytokines to Canine Cell Cultures
Recombinant canine cytokines were purchased from KingFisher Biotec (St. Paul, MN, USA). These cytokines were used in two separate experiments. First, canine cell cultures were exposed to a range of doses of recombinant cytokines and the cytotoxicity of the cytokines was analyzed. The highest dose of the cytokine that did not cause cytotoxicity in canine cell cultures was determined. Next, cells were treated with the highest dose of cytokine(s) that did not cause cytotoxicity and their effect on tdTr expression in MYXV-red-inoculated canine cell cultures was measured. Details of these two experiments are described below.
Toxicity of the cytokines was evaluated by adding individual and combined recombinant proteins onto 48-well plates (Corning, Corning, NY, USA) containing confluent cells in the absence of virus. The doses of cytokines tested [IFNα (50, 500, and 1000 U/mL), IFNβ (50, 500, and 1000 U/mL), TNFα (10, 20, and 40 ng/mL), and IFNγ (20, 100, and 200 U/mL)] were guided by previous studies [24,49,50,51,52]. A Cell Titer Blue® Cell Viability Assay (Promega, Madison, WI, USA) was used according to the manufacturer’s instructions. Cytotoxicity was calculated as the percentage of viable cells after cytokine treatment relative to untreated cells (fluorescence value of treated cells ÷ fluorescence value of the untreated cells). Data is shown for samples that were run in duplicate. Three similar experiments were performed, although different incubation endpoints were collected. Similar conclusions were made from all experiments.
The highest dose of recombinant cytokine that did not cause toxicity was used to determine if cytokines would alter MYXV-red tdTr expression in canine cells. Confluent cells in 48-well plates (Corning, Corning, NY, USA) were inoculated by removing growth media (described in subsection 2.1.) and incubating cells with MYXV-red at 0.1 moi in media lacking FBS. After incubation in a water-jacketed incubator at 5% CO2 and 37˚C for one-hour, recombinant canine IFNα (500 U/mL), IFNβ (500 U/mL), TNFα (20 ng/mL), and IFNγ (100 U/mL) were diluted in growth media (described in subsection 2.1.) and placed on the cells alone and in combination. MYXV-red infection in cells was monitored visually by screening for fluorescent red protein expression using a 560/40 nm bandpass excitation filter and a Leica DMI4000B inverted microscope. Reporter protein expression by MYXV-red was measured using a microplate reader by detecting fluorescence (excitation 554 nm, emission 581 nm) at 24, 48, and 72 hpi. Four to ten data points at 72 hpi were collected for each canine cell tested. Data were analyzed using multiple unpaired t-tests in GraphPad Prism version 9.1.0 software (San Diego, CA, USA).
2.3.3. Inhibition of Cytokines during MYXV-Red Infection
To block the effects of cytokine response to viral infection in canine cells, 100 ng of antibody against canine TNFα, IFNα, and IFNγ (KingFisher Biotec, St. Paul, MN, USA) was added alone and in combination to confluent cells infected with MYXV-red for one-hour at 0.1 moi in 48-well plates (Corning, Corning, NY, USA). Antibody against canine IFNβ was not available for purchase. MYXV-red infection in cells was monitored visually by screening for fluorescent red protein expression using a 560/40 nm bandpass excitation filter and a Leica DMI4000B inverted microscope. Virus titers were determined using a plaque assay 72 hpi. The effect of cytokines on MYXV-red replication was calculated as a percentage of ffu/mL in treated cells relative to untreated cells (ffu/mL of treated cells ÷ ffu/mL of untreated cells). Four to eight data points were collected for each cell line tested. Data were analyzed using unpaired t-tests with Welch’s correction in GraphPad Prism version 9.1.0 software (San Diego, CA, USA).
2.4. MYXVΔSERP2 Oncolytic Therapy in Dogs with Sarcomas
2.4.1. Treatment Protocols for Dogs with Sarcoma
Dogs that were otherwise healthy except for spontaneous growth of a grade II or III soft tissue sarcoma were enrolled in the study as previously described [18]. Post-operative treatment of dogs with MYXVΔSERP2 and combination therapy of dogs with oclacitinib (Apoquel®, Zoetis, Parsippany, NJ, USA) and post-operative MYXVΔSERP2 (O+MYXV ΔSERP2) were approved by the CSU Veterinary Clinical Studies review board (protocols 2016-016 and 2017-134, respectively). The safety of MYXVΔSERP2 treatment was previously described [18], but clinical outcomes data were not available at the time of publication. Outcome data from these dogs are reported in this manuscript (Results subsection 3.4.3.).
Administration of post-operative virus in O+MYXVΔSERP2-treated dogs was performed as previously described [18]. Briefly, sucrose pad purified MYXVΔSERP2 (5-10 mL) was injected into excised tumor margins at 106 ffu/mL immediately following the surgery (Day 0) and boosted with a second treatment approximately two weeks later. The second treatment was delayed an additional two weeks if the surgical site was not healed or if a seroma was present. Animals enrolled in combination therapy began a 14-day treatment with oclacitinib approximately 7 days prior to surgery. Figure 1 indicates the timeline of sample collections and treatments in dogs with STS. Note that both post-operative MYXVΔSERP2 and O+MYXVΔSERP2 treatments were pilot clinical trials in client-owned dogs, therefore the exact timing of treatment varied slightly between patients as appointment times were scheduled around the owners’ availability.
2.4.2. Sample Analysis for Study Dogs
A physical exam, complete blood count (CBC), serum biochemistry profile, and urinalysis were completed for each patient pre-enrollment, and on Days 0, ~14, and ~28 to assess the dogs’ overall health. Three patients also had samples collected ≥ 42 days post-operatively. A board certified veterinary anatomic pathologist assessed surgically excised tumors for tumor grade, presence of inflammation, and residual disease. Patients were examined for tumor regrowth monthly for up to ~1 year post-operatively.
DNA was extracted from blood, urine, and feces collected from the patients at the time of enrollment (pre-treatment) and on the days specified above. A QIAGEN DNeasy Blood and Tissue kit (Hilden, Germany) was used to isolate DNA according to the manufacturer’s instructions. Detection of virus was done using conventional PCR with primers that detect an 818 bp product in the multigene region [M032R, M033R, M034L (DNA polymerase), and M035R] of the MYXV genome (forward 5’CAC CCT CTT TAG TAA AGT ATA CAC C 3’, reverse 5’GAA ATG TTG TCG GAC GGG 3’). A second set of primers covering the M135-M136 genes resulting in an 1182 bp product were also used (forward 5’ CGA CAC CTG TGT ATG TTT G 3’, reverse 5’CCA ATA ACA CAC AGT TCG G 3’). Amplification was performed in a thermocycler under the following conditions: 94˚C for 1 min, followed by 30 cycles of 94˚C for 30 sec, 55˚C for 1 min, 72˚C for 2 min, and a final 72˚C 10 min elongation step.
The immune response of patients to treatment was assessed in several ways. To detect neutralizing antibodies that developed in dogs treated with MXYVΔSERP2, virus neutralization assays were performed using serum collected at the above time points. A plaque reduction neutralization test (PRNT) was used to quantitate anti-viral antibodies as previously described [42,53]. To evaluate subsets of peripheral blood leukocytes, flow cytometric analysis was performed by the Clinical Immunology (Hematopathology) Laboratory at CSU. Numbers of circulating B-lymphocytes (CD21+/MHCII+), T-lymphocytes (CD3+/CD5+/CD4±/CD8±), neutrophils (CD4+/CD5-), monocytes (CD14+/MHCII+), and hematopoietic precursors (CD34+) were quantified. Data were analyzed using Kaluza software (Beckman Coulter Life Sciences, Indianapolis, IN, USA). Additionally, pre-treatment concentrations of serum IL-31 and type I IFNs were compared to post-treatment concentrations using Nori® Canine ELISA kits (Genorise Scientific, Inc., PA, USA).
The time to tumor regrowth in dogs treated with MYXVΔSERP2 was compared to dogs treated with combination therapy (O+MYXVΔSERP2) using an unpaired, nonparametric t-test (Mann-Whitney test) in GraphPad Prism version 9.1.0 software (San Diego, CA, USA).
3. Results
3.1. Characteristics of Canine Cell Lines
Cancer cells from tumors of two canine patients with spontaneously occurring STS were isolated post-operatively. FB were collected from one dog post-humorously. All cell cultures were confirmed to be of unique canine lineage using STR analysis. Immunocytochemistry assays were performed to determine if cells expressed proteins associated with the tumor they were derived from. As expected, all cells expressed vimentin but did not express cytokeratin.
Next, the permissively of the cells to MYXV-red infection was assessed. MYXV-red titers increased logarithmically over time in canine cancer cells inoculated with virus at 0.1 moi, but viral replication in FB was suppressed (Figure 2). Visible cytotoxic effects were minimal up to 72 hpi with MYXV-red at 0.1 moi in canine STS and FB cells (Figure 3). Cell viability assays collected 24, 48, and 72 hpi with MYXV-red (0.1 moi) indicated that by 24 hpi, there was a significant decrease in viability of canine STS cells (Figure 4). Although MYXV-red replication (Figure 2) and reporter protein expression were minimal in healthy canine FB (Figure 3), a significant decrease in FB cell viability was detected 72 hpi (Figure 4). Data indicate that the canine STS cells are fully permissive to MYXV replication and that healthy canine FB are poorly permissive to MYXV.
3.2. Cytokine mRNA Expression Following MYXV Inoculation
Given that canine STS cells were fully permissive for MYXV infection but non-cancerous FB were not, we predicted that IFN and other cytokine transcription levels would be different in infected STS and FB cells. A nCounter® Canine IO Panel (NanoString Technologies, Seattle, WA, USA) was used to determine how mRNA expression changed in STS-1, STS-2, and FB following inoculation with MYXV or MYXVΔSERP2 (0.1 moi) as compared to mock-infected cells at 4 and 8 hpi. MYXVΔSERP2 was evaluated because its safety profile in dogs is known [18] and it was used to treat the dogs with spontaneous sarcoma described in this study.
Using nCounter 4.0 Advanced Analysis software, heatmaps of key cellular pathways associated with neoplastic transformation and immunologic responses were created. They highlighted several differences between the three mock-infected cell cultures (Figure 5A) and between mock-infected and MYXV- or MYXVΔSERP2-inoculated FB cells (Figure 5B), STS-1 cells (Figure 5C), and STS-2 cells (Figure 5D). Of particular interest for this study, pathway scoring analyses indicated that JAK-STAT and cytokine/chemokine signaling scores were increased in FB and decreased in STS cells 4 and 8 hpi with viruses (Figure 6). This was expected, given the role of JAK-STAT and cytokine/chemokine signaling in limiting viral infection of healthy cells, but not in some cancer cells.
Figure 7 shows heatmaps indicating fold-changes in selected cytokine transcripts detected in cells using NanoString technology. Cells inoculated with virus were compared to mock-infected cells collected at the same hpi. IFNα7, TNFSF4, TNFSF13, IL-1β, and IL-10 transcripts were more numerous in all MYXV-inoculated cells at 4 and/or 8 hpi (Figure 7A). This also was observed for IFNβ1 (except in STS-1 cells at 4 hpi and STS-2 cells at 8 hpi) and IL-4 (except in STS-2 cells at 8 hpi) (Figure 7A). IFNγ transcripts remained stable or were down-regulated following MYXV inoculation with the notable exception of FB at 8 hpi (Figure 7A). TGFβ1 and TGFβ2 were stable or slightly increased at 4 hpi in all cells, but were down-regulated in STS-1 and STS-2 cells at 8 hpi (Figure 7A). Interestingly, fold changes in transcript numbers were much more variable following inoculation with MYXVΔSERP2 (Figure 7B). It is possible that expression of lacZ by this recombinant virus (which is driven by an early/late poxvirus promoter) is the cause for the differences observed in cellular transcript changes when MYXV and MYXVΔSERP2 are compared. Table 2 lists genes with > 10-fold differences in expression in MYXV-inoculated as compared to mock-infected FB, STS-1, and STS-2 cells, respectively, at 8 hpi calculated using Rosalind software. Transcripts that were similarly different in MYXVΔSERP2-inoculated cells as compared to mock-infected cells of the same cell type are indicated by an asterisk. Several similarities in up-regulated and down-regulated genes were detected in MYXV- and MYXVΔSERP2-inoculated cells as compared to mock-infected cells of the same type, although fold changes in MYXVΔSERP2-inoculated cells were often less pronounced that in MYXV-inoculated cells. There was no differential expression detected between mock-infected cells of the same type at 4 and 8 hpi.
Figure 8 shows heatmaps of RQ values generated using qPCR for IFNα, IFNβ, TNFα, IL-1β, IFNγ, IL-4, IL-10, and TGFβ mRNA expression in STS-1 and FB following inoculation (0.1 moi) with MYXV (Figure 8A) or MYXVΔSERP2 (Figure 8B) as compared to mock-infected cells. MYXV and MYXVΔSERP2 inoculation caused more similar alterations in transcript levels using this method of detection. At 4 hpi, STS-1 cell mRNA transcripts were relatively stable for all cytokines except IL-4 transcripts, which were increased (Figure 8A,B). The increase in IL-4 transcripts also was observed in FB inoculated with MYXV (0.1 moi) 4hpi (Figure 8A). Virus-inoculated FB also had a mild increase in IFNα RQ values at 4 hpi (Figure 8A,B).
3.3. Effect of Cytokines on MYXV Infection
It was known that treatment of primary human FB with a combination of recombinant human IFNβ and TNF blocked MYXV replication [54]. To determine the effect of adding exogenous cytokines to canine cells following MYXV-red inoculation, canine cells were treated with recombinant canine IFNα, IFNβ, TNFα, and/or IFNγ both alone and in combination. No cytotoxic effects (p < .05) were observed when cancer cells were treated with the cytokines at the following concentrations: 500 U/mL IFNα, 500 U/mL IFNβ, 20 ng/mL TNFα, and 100 U/mL IFNγ (Figure 9). When IFNα was added to media following MYXV-red inoculation, MYXV-red reporter protein expression was significantly (p < .05) decreased in all cells tested (Table 3). Significant reductions in MYXV-red reporter protein expression also were observed when IFNα was combined with IFNβ, TNFα and/or IFNγ. Similarly, virus titers were reduced in STS-1 and STS-2 cells following cytokine treatment with IFNs (Figure 10).
Conversely, when IFNα, TNFα, and IFNγ were blocked using anti-cytokine antibodies, increases in MYXV-red replication were observed in non-permissive canine cells (Figure 11). Specifically, in canine FB, which inhibit MXYV-red replication (Figure 2), significant increases (p < .05) in MYXV-red titers were observed when anti-IFNα antibody was used alone or in combination with anti-TNFα and/or anti-IFNγ antibodies. This also was seen following treatment with a combination of anti-TNFα and anti-IFNγ antibodies. Interestingly, when treated concurrently with anti-IFNα, anti-TNFα, and anti-IFNγ, MYXV-red replication was decreased in STS-1 cells, which are naturally permissive for viral replication (Figure 2). We speculate that concurrent inhibition of IFNα, TNFα, and IFNγ promotes the ability of these cells to respond to the other cytokines that are upregulated secondary to MYXV inoculation (including IFNβ and IL-4). This disruption of the balance of intracellular cytokines could lead to a more effective anti-viral immune response. Additional work will need to be done to see if this is true.
3.4. Post-Operative MYXVΔSERP2 Treatement in Dogs with Soft Tissue Sarcoma
3.4.1. Safety of Combination Oclacitinib and MYXVΔSERP2 Treatment in Dogs
The safety profile of post-operative MYXVΔSERP2 treatment in 5 dogs was previously reported [18]. Similar findings were observed in 4 dogs treated with O+MYXVΔSERP2. No viral shedding was detected in urine, feces, or blood collected on Days 0, ~14, and ~28, and periodically up to 62 days post-operatively (Figure S1). Also, no clinically significant changes in physical examination, CBC, serum biochemistry analysis, or urinalysis were detected in dogs treated with O+MYXVΔSERP2 (Table S1).
3.4.2. Immune Response to Combination Oclacitinib and MYXVΔSERP2 Treatment in Dogs
Immune responses were evaluated with virus neutralization assays, flow cytometric analysis of peripheral blood, and measurement of serum cytokine concentrations. It was previously reported that dogs treated with MYXV∆SERP2 alone did not have pre-existing antibodies that neutralized MYXV (Day 0) nor did they seroconvert on Day 14; however, 2 of 5 dogs seroconverted by Day 28 [18]. Likewise, dogs treated with O+MYXV∆SERP2 did not have anti-MYXV antibodies on Day 0 or 14 but 2 of 4 dogs seroconverted by Day 28 (Figure 12). Flow cytometric evaluation of peripheral blood leukocytes indicated no significant abnormalities in 5 of 5 dogs treated with MYXVΔSERP2 [18] and 3 of 4 dogs treated with O+MYXVΔSERP2. One O+MYXVΔSERP2-treated patient (Dog 6) had a low concentration of circulating CD8+ T cells (89 cells/µL, reference interval 157-965 cells/µL) on Day 14 (Figure S2). All cell counts were within reference intervals at the pre-treatment exam and on Days 0, 29, and 43 in this patient. As this finding was transient and was only observed in one patient who had no clinical signs of disease, it was determined to be of no clinical significance.
It is known that oclacitinib inhibits expression of a variety of cytokines signaled through JAK1 pathways [29]. In dogs treated with oclacitinib, the inhibition of serum IL-31 concentration is indicative of treatment success. Figure 13 compares pre- and post-treatment serum cytokine concentrations in 4 of 5 MYXV∆SERP2- and 4 of 4 O+MYXV∆SERP2-treated dogs (paired sera from Dog 4 were not available for this assay due to failure to collect a sufficient volume of blood from this patient at Day 0). Interleukin-31, IFNα, and IFNβ concentrations increased in 3 of 4 dogs treated with MYXVΔSERP2 alone. Whereas IL-31 and IFNα concentrations remained near pre-treatment concentrations or decreased in dogs treated with O+ MYXVΔSERP2. IFNβ concentrations remained near pre-treatment concentrations or increased slightly in patients treated with combination therapy, but IFNβ concentrations in all O+MYXV∆SERP2-treated dogs remained lower than IFNβ concentrations in 3 of 4 dogs treated with MYXV∆SERP2 alone. Importantly for this study, data suggest that oclacitinib successfully suppressed IL-31 and type I IFN responses in O+MYXV∆SERP2-treated dogs.
3.4.3. Clinical Response to Combination Oclacitinib and MYXVΔSERP2 Treatment in Dogs
We hypothesized that a delayed type I IFN response following MYXV∆SERP2 treatment may allow the virus more time to replicate in tumor cells thereby improving outcomes in dogs treated with O+MYXV∆SERP2 as compared to dogs treated with MYXV∆SERP2 alone. Tumor regrowth is expected in dogs with STS if the tumor is not completely resected during surgery [2,3]. In dogs treated with MYXV∆SERP2 post-operatively, tumor regrowth was documented in 2 of 4 dogs (one dog was lost to follow-up on Day 32). Similarly, in dogs treated with O+MYXV∆SERP2, tumor regrowth was observed in 3 of 4 dogs. Table 4 describes histologic findings in patient tumors and indicates the time to tumor regrowth or the last day of patient follow-up. Time to tumor regrowth was not significantly different in dogs treated with MYXV∆SERP2 and dogs treated with O+MYXV∆SERP2.
4. Discussion
One goal of this study was to determine if manipulation of intracellular cytokine concentrations alters MYXV replication in primary canine cells as it does in murine and human primary cells. When mRNA transcripts were evaluated, JAK-STAT and cytokine/chemokine pathway signaling scores increased in healthy canine fibroblasts following MYXV and MYXVΔSERP2 inoculation, but decreased in two canine cancer cell cultures (STS-1 and STS-2). The importance of JAK-STAT signaling in inhibition of viral replication is well established in human cells, including human keratinocyte susceptibility to poxvirus (vaccinia virus) infection [55]. This study suggests it is also important in canine cells. Importantly, treatment of cells with type I IFNs reduced MYXV replication in permissive canine cancer cells and blocking type I IFNs allowed for more robust MYXV infection in poorly permissive FB. Specifically, both canine sarcoma cell cultures showed a significant decrease in MYXV replication when cells were treated with IFNα or a combination of IFNα and IFNβ. This is similar to findings in murine cells where blocking type I IFNs allowed for MYXV replication in nonpermissive primary cells [23]. Canine sarcoma cell cultures also showed a significant reduction in MYXV replication in the presence of IFNα or IFNβ when combined with TNFα. The combination of IFNβ and TNFα is known to have a synergist effect to prevent MYXV infection of human primary cells [24,54]. Previous work demonstrated that MYXV replication is inhibited in canine cells that have low concentrations of phosphorylated Akt (including primary FB) [41]. Blocking IFNα signaling in canine FB more than doubled MXYV-red titers recovered from the cells at 72 hpi. These new data suggest that defective type I interferon responses in canine cancer cells also are critical in permitting MYXV replication. The similarities observed in human and canine cell culture response to cytokine alterations support the use of canine cancer patients as models for human cancer patient response to OVs.
We had found that oclacitinib treatment prolonged MYXV∆SERP2 replication in rhabdomyosarcoma allografts in mice [56]. Here we show that combination oclacitinib and MYXV∆SERP2 therapy is safe in dogs with STS. However, clinical improvements were not observed in mice or dogs when combination treatment was compared to MYXV∆SERP2 treatment alone. It might be beneficial to combine different type I IFN inhibitors with MYXV therapy. Rapamycin inhibits the mammalian target of rapamycin pathway to decrease translation of IFNs. Replication of MYXV was increased in cancer cell cultures and in mouse models of melanoma treated with rapamycin [53,57,58,59,60,61,62,63]. Ruxolitinib is a JAK1/2 inhibitor that promotes vesicular stomatitis, herpes simplex, and measles viral replication in cell culture [64,65,66,67,68,69,70]. The drug is not listed for use in dogs, however a recent study indicated that in vitro canine mast cell proliferation is inhibited by ruxolitinib treatment [71]. The decrease in proliferation of these cells was associated with ruxolitinib inhibition of JAK2/STAT5 phosphorylation, indicating there may be additional benefits to using ruxolitinib to treat canine cancers.
Use of other OVs could improve outcomes in dogs as well. Attenuated adenoviruses have been used to treat canine spontaneous soft tissue sarcomas. A dog with fibrosarcoma was treated with a modified canine adenovirus and post-treatment surgery successfully removed the tumor [11]. Using a different adenovirus construct, progressive disease was observed in 1 dog with a schwannoma but a complete response was obtained in 1 dog with a schwannoma, partial responses in 2 dogs with hemangiopericytoma, and stable disease in 2 dogs with fibrosarcomas, 2 with undifferentiated sarcomas, and 1 with a hemangiopericytoma [12].
Additional immunotherapeutics that could augment OVs include inhibitors of programed cell death protein 1 (PD-1) or cytotoxic T lymphocyte associated protein 4 interactions, monoclonal antibodies that block tumor growth, tyrosine kinase inhibitors, cytokines that stimulate inflammation, and pro-apoptotic drugs [72]. One future direction for the laboratory is evaluating the effect of concurrent MYXV therapy, PD-1/PD-L1 inhibition, and cytokine stimulation of macrophages in dogs with spontaneous tumors.
5. Conclusions
Inhibition of IFNα increases MYXV replication in healthy canine FBs. Combination treatment with oclacitinib (an inhibitor of type I IFNs) and MYXV∆SERP2 was safe in dogs with STS, however little treatment benefit was seen; 2 of 4 dogs treated with MYXV∆SERP2 and 3 of 4 dogs treated with O+MYXV∆SERP2 suffered tumor recurrence. This outcome may be slightly better than surgery alone, but larger placebo-controlled clinical trials are needed.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Agarose gel electrophoresis of polymerase chain reactions designed to detect MYXVΔSERP2 DNA. Figure S2: Flow cytometric results of peripheral blood cell analysis from Dog 6 on Day 14. Table S1: Clinical data lists.
Author Contributions
Conceptualization, A.L.M.; methodology, L.V.A. and A.L.M.; validation, L.V.A.; formal analysis, L.V.A. and A.L.M.; resources, A.L.M.; data curation, L.V.A. and A.L.M.; writing—original draft preparation, L.V.A. and A.L.M.; writing—review and editing, L.V.A., K.M.W., B.S. and A.L.M.; supervision, K.M.W., B.S. and A.L.M.; client consent, K.M.W.; patient examination, K.M.W and B.S.; surgery, B.S.; project administration, A.L.M.; funding acquisition, A.L.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Elsa U. Pardee Foundation, a Colorado Clinical & Translational Sciences Institute CSU Pilot Program Grant, a CSU College Research Council Interdisciplinary Pilot Award, and CSU Clinical Pathology Research and Development Funds.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Veterinary Clinical Studies Review Board of CSU (protocols 2016-016 and 2017-134, approved January 2018 and August 2018, respectively).
Informed Consent Statement
Informed consent was obtained from all of the dog’s owners whose pets were involved in the study. Written informed consent has been obtained from the dogs’ owners to publish this paper.
Acknowledgments
The authors would like to thank Mariah Jordan for helping assess cell viability following viral inoculation and Dr. Bryce Zietz who began the cytokine cytotoxicity studies. Additionally, we are extremely grateful to the owners who elected to enroll their beloved pets in this clinical trial.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Porrello, A.; Cardelli, P.; Spugnini, E.P. Oncology of companion animals as a model for humans. an overview of tumor histotypes. J. Exp. Clin. Cancer Res. 2006, 25, 97–105. [Google Scholar] [PubMed]
- Ehrhart, N. Soft-tissue sarcomas in dogs: A review. J. Am. Anim. Hosp. Assoc. 2005, 41, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Dernell, W.S.; Withrow, S.J.; Kuntz, C.A.; Powers, B.E. Principles of treatment for soft tissue sarcoma. Clin Tech Small Anim Pr. 1998, 13, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Hohenhaus, A.E.; Kelsey, J.L.; Haddad, J.; Barber, L. Canine Cutaneous and Subcutaneous Soft Tissue Sarcoma: An Evidence-Based Review of Case Management. J. Am. Anim. Hosp. Assoc. 2016, 52, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Liang, M. Oncorine, the World First Oncolytic Virus Medicine and its Update in China. Curr. Cancer Drug Targets 2018, 18, 171–176. [Google Scholar] [CrossRef]
- Pol, J.; Kroemer, G.; Galluzzi, L. First oncolytic virus approved for melanoma immunotherapy. Oncoimmunology 2015, 5, e1115641. [Google Scholar] [CrossRef] [PubMed]
- Cinatl Jr, J.; Cinatl, J.; Michaelis, M.; Kabickova, H.; Kotchetkov, R.; Vogel, J.U.; Doerr, H.W.; Klingebiel, T.; Driever, P.H. Potent oncolytic activity of multimutated herpes simplex virus G207 in combination with vincristine against human rhabdomyosarcoma. Cancer Res. 2003, 63, 1508–1514. [Google Scholar]
- Gentschev, I.; Adelfinger, M.; Josupeit, R.; Rudolph, S.; Ehrig, K.; Donat, U.; Weibel, S.; Chen, N.G.; Yu, Y.A.; Zhang, Q.; et al. Preclinical evaluation of oncolytic vaccinia virus for therapy of canine soft tissue sarcoma. PLoS One 2012, 7, e37239. [Google Scholar] [CrossRef]
- Kinn, V.G.; Hilgenberg, V.A.; MacNeill, A.L. Myxoma virus therapy for human embryonal rhabdomyosarcoma in a nude mouse model. Oncolytic Virotherapy 2016, 5, 59–71. [Google Scholar]
- Barton, K.N.; Paielli, D.; Zhang, Y.; Koul, S.; Brown, S.L.; Lu, M.; Seely, J.; Kim, J.H.; Freytag, S.O. Second-generation replication-competent oncolytic adenovirus armed with improved suicide genes and ADP gene demonstrates greater efficacy without increased toxicity. Mol. Ther. 2006, 13, 347–56. [Google Scholar] [CrossRef]
- Laborda, E.; Puig-Saus, C.; Rodriguez-García, A.; Moreno, R.; Cascalló, M.; Pastor, J.; Alemany, R. A pRb-responsive, RGD-modified, and hyaluronidase-armed canine oncolytic adenovirus for application in veterinary oncology. Mol. Ther. 2014, 22, 986–98. [Google Scholar] [CrossRef]
- Cejalvo, T.; Perise-Barrios, A.J.; del Portillo, I.; Laborda, E.; Rodriguez-Milla, M.A.; Cubillo, I.; Vazquez, F.; Sardon, D.; Ramirez, M.; Alemany, R.; et al. Remission of spontaneous canine tumors after systemic cellular viroimmunotherapy. Cancer Res. 2018, 78, 4891–4901. [Google Scholar] [CrossRef]
- Hwang, C.C.; Igase, M.; Sakurai, M.; Haraguchi, T.; Tani, K.; Itamoto, K.; Shimokawa, T.; Nakaichi, M.; Nemoto, Y.; Noguchi, S.; et al. Oncolytic reovirus therapy: Pilot study in dogs with spontaneously occurring tumours. Vet. Comp. Oncol. 2017, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Béguin, J.; Nourtier, V.; Gantzer, M.; Cochin, S.; Foloppe, J.; Balloul, J.M.; Laloy, E.; Tierny, D.; Klonjkowski, B.; Quemeneur, E.; et al. Safety studies and viral shedding of intramuscular administration of oncolytic vaccinia virus TG6002 in healthy beagle dogs. BMC Vet. Res. 2020, 16. [Google Scholar] [CrossRef]
- Béguin, J.; Gantzer, M.; Farine, I.; Foloppe, J.; Klonjkowski, B.; Maurey, C.; Quéméneur, É.; Erbs, P. Safety, biodistribution and viral shedding of oncolytic vaccinia virus TG6002 administered intravenously in healthy beagle dogs. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ilyinskaya, G. V.; Mukhina, E. V.; Soboleva, A. V.; Matveeva, O. V.; Chumakov, P.M. Oncolytic sendai virus therapy of canine mast cell tumors (A pilot study). Front. Vet. Sci. 2018, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Omar, N.B.; Bentley, R.T.; Crossman, D.K.; Foote, J.B.; Koehler, J.W.; Markert, J.M.; Platt, S.R.; Rissi, D.R.; Shores, A.; Sorjonen, D.; et al. Safety and interim survival data after intracranial administration of M032, a genetically engineered oncolytic HSV-1 expressing IL-12, in pet dogs with sporadic gliomas. Neurosurg. Focus 2021, 50, 1–11. [Google Scholar] [CrossRef]
- MacNeill, A.L.; Weishaar, K.M.; Séguin, B.; Powers, B.E. Safety of an oncolytic myxoma virus in dogs with soft tissue sarcoma. Viruses 2018, 10, 10–23. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, A.K.; Naik, S.; Galyon, G.D.; Jenks, N.; Steele, M.; Peng, K.-W.; Federspiel, M.J.; Donnell, R.; Russell, S.J. Safety studies on intravenous administration of oncolytic recombinant vesicular stomatitis virus in purpose-bred beagle dogs. Hum. Gene Ther. Clin. Dev. 2013, 24, 174–81. [Google Scholar] [CrossRef] [PubMed]
- Mcnab, F.; Mayer-barber, K.; Sher, A.; Wack, A.; Garra, A.O.; Europe PMC Funders Group. Type I interferons in infectious disease. Nat. Rev. Immunol. 2020, 15, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Li, S.F.; Gong, M.J.; Zhao, F.R.; Shao, J.J.; Xie, Y.L.; Zhang, Y.G.; Chang, H.Y. Type I interferons: Distinct biological activities and current applications for viral infection. Cell. Physiol. Biochem. 2018, 51, 2377–2396. [Google Scholar] [CrossRef] [PubMed]
- Klotz, D.; Baumgärtner, W.; Gerhauser, I. Type I interferons in the pathogenesis and treatment of canine diseases. Vet. Immunol. Immunopathol. 2017, 191, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Ma, Y.; Barrett, J.W.; Gao, X.; Loh, J.; Barton, E.; Virgin, H.W.; McFadden, G. Disruption of Erk-dependent type I interferon induction breaks the myxoma virus species barrier. Nat. Immunol. 2004, 5, 1266–1274. [Google Scholar] [CrossRef] [PubMed]
- Bartee, E.; McFadden, G. Human cancer cells have specifically lost the ability to induce the synergistic state caused by tumor necrosis factor plus interferon-beta. Cytokine 2009, 47, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Stanifer, M.L.; Pervolaraki, K.; Boulant, S. Differential regulation of type I and type III interferon signaling. Int. J. Mol. Sci. 2019, 20, 1–22. [Google Scholar] [CrossRef]
- Budhwani, M.; Mazzieri, R.; Dolcetti, R. Plasticity of Type I Interferon-Mediated Responses in Cancer Therapy: From Anti-tumor Immunity to Resistance. Front. Oncol. 2018, 8. [Google Scholar] [CrossRef]
- Yang, R.; Wang, L.; Sheng, J.; Huang, Q.; Pan, D.; Xu, Y.; Yan, J.; Wang, X.; Dong, Z.; Yang, M. Combinatory effects of vaccinia virus VG9 and the STAT3 inhibitor Stattic on cancer therapy. Arch. Virol. 2019, 164, 1805–1814. [Google Scholar] [CrossRef]
- Zoetis. Available online: https://www.zoetisus.com/products/dogs/apoquel/pdf/apoqueltechnicalmonograph.pdf.
- Gonzales, A.J.; Bowman, J.W.; Fici, G.J.; Zhang, M.; Mann, D.W.; Mitton-Fry, M. Oclacitinib (APOQUEL®) is a novel Janus kinase inhibitor with activity against cytokines involved in allergy. J. Vet. Pharmacol. Ther. 2014, 37, 317–324. [Google Scholar] [CrossRef]
- O’Shea, J.J. Targeting the Jak/STAT pathway for immunosuppression. Ann. Rheum. Dis. 2004, 63, 67–71. [Google Scholar] [CrossRef]
- Andrewes, C.H.; Harisijades, S. Propagation of myxoma virus in one-day old mice. Br. J. Exp. Pathol. 1955, 36, 18–21. [Google Scholar]
- Fenner, F.; Woodroffe, G.M. The pathogenesis of infectious myxomatosis: the mechanism of infection and the immunological response in the European rabbit (Oryctolagus cuniculus). Br. J. Exp. Pathol. 1953, 34, 400–411. [Google Scholar]
- Gorski, J.; Mizak, B.; Chrobocinska, M. Control of rabbit myxomatosis in Poland. Rev. Sci. Tech. 1994, 13, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Jackson, E.W.; Dorn, C.R.; Saito, J.K.; McKercher, D.G. Absence of serological evidence of myxoma virus infection in humans exposed during an outbreak of myxomatosis. Nature 1966, 211, 313–314. [Google Scholar] [CrossRef] [PubMed]
- McCabe, V.J.; Tarpey, I.; Spibey, N. Vaccination of cats with an attenuated recombinant myxoma virus expressing feline calicivirus capsid protein. Vaccine 2002, 20, 2454–2462. [Google Scholar] [CrossRef] [PubMed]
- McCabe, V.J.; Spibey, N. Potential for broad-spectrum protection against feline calicivirus using an attenuated myxoma virus expressing a chimeric FCV capsid protein. Vaccine 2005, 23, 5380–5388. [Google Scholar] [CrossRef] [PubMed]
- Pignolet, B.; Boullier, S.; Gelfi, J.; Bozzetti, M.; Russo, P.; Foulon, E.; Meyer, G.; Delverdier, M.; Foucras, G.; Bertagnoli, S. Safety and immunogenicity of myxoma virus as a new viral vector for small ruminants. J. Gen. Virol. 2008, 89, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Fenner, F. Adventures with poxviruses of vertebrates. FEMS Microbiol. Rev. 2000, 24, 123–133. [Google Scholar] [CrossRef] [PubMed]
- MacNeill, A.L.; Turner, P.C.; Moyer, R.W. Mutation of the Myxoma virus SERP2 P1-site to prevent proteinase inhibition causes apoptosis in cultured RK-13 cells and attenuates disease in rabbits, but mutation to alter specificity causes apoptosis without reducing virulence. Virology 2006, 356, 12–22. [Google Scholar] [CrossRef]
- MacNeill, A.L.; Moldenhauer, T.; Doty, R.; Mann, T. Myxoma virus induces apoptosis in cultured feline carcinoma cells. Res. Vet. Sci. 2012, 93, 1036–1038. [Google Scholar] [CrossRef]
- Urbasic, A.S.; Hynes, S.; Somrak, A.; Contakos, S.; Rahman, M.M.; Liu, J.; MacNeill, A.L. Oncolysis of canine tumor cells by myxoma virus lacking the serp2 gene. Am. J. Vet. Res. 2012, 73, 1252–1261. [Google Scholar] [CrossRef]
- Doty, R.A.; McFadden, G.; Roy, E.J.; MacNeill, A.L. Histological evaluation of intratumoral myxoma virus treatment in an immunocompetent mouse model of melanoma. Oncolytic Virotherapy 2013, 2, 1–17. [Google Scholar]
- Liu, J.; Wennier, S.; Reinhard, M.; Roy, E.; MacNeill, A.; McFadden, G. Myxoma virus expressing interleukin-15 fails to cause lethal myxomatosis in European rabbits. J. Virol. 2009, 83, 5933–5938. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Idate, R.; Lana, S.E.; Regan, D.P.; Duval, D.L. Integrated analysis of canine soft tissue sarcomas identifies recurrent mutations in TP53, KMT genes and PDGFB fusions. bioRxiv Prepr. Serv. Biol. 2023. [CrossRef] [PubMed]
- Tosic, V.; Thomas, D.L.; Kranz, D.M.; Liu, J.; McFadden, G.; Shisler, J.L.; MacNeill, A.L.; Roy, E.J. Myxoma virus expressing a fusion protein of interleukin-15 (IL15) and IL15 receptor alpha has enhanced antitumor activity. PLoS One 2014, 9, e109801. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Park, W.J.; Park, B.J.; Song, Y.J.; Lee, D.H.; Yuk, S.S.; Lee, J.B.; Park, S.Y.; Song, C.S.; Lee, S.W.; Choi, I.S. Analysis of cytokine production in a newly developed canine tracheal epithelial cell line infected with H3N2 canine influenza virus. Arch. Virol. 2015, 160, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Jassies-van der Lee, A.; Rutten, V.; Spiering, R.; van Kooten, P.; Willemse, T.; Broere, F. The immunostimulatory effect of CpG oligodeoxynucleotides on peripheral blood mononuclear cells of healthy dogs and dogs with atopic dermatitis. Vet. J. 2014, 200, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Belloni, L.; Allweiss, L.; Guerrieri, F.; Pediconi, N.; Volz, T.; Pollicino, T.; Petersen, J.; Raimondo, G.; Dandri, M.; Levrero, M. IFN-α inhibits HBV transcription and replication in cell culture and in humanized mice by targeting the epigenetic regulation of the nuclear cccDNA minichromosome. J. Clin. Invest. 2012, 122, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Choi, K.J.; Bae, S. Interferon-gamma enhances radiation-induced cell death via downregulation of Chk1. Cancer Biol. Ther. 2012, 13, 1018–1025. [Google Scholar] [CrossRef]
- O’Brien, W.J.; Heimann, T.; Tsao, L.S.; Seet, B.T.; McFadden, G.; Taylor, J.L. Regulation of nitric oxide synthase 2 in rabbit corneal cells. Invest. Ophthalmol. Vis. Sci. 2001, 42, 713–719. [Google Scholar]
- Greifenberg, V.; Ribechini, E.; Rößner, S.; Lutz, M.B. Myeloid-derived suppressor cell activation by combined LPS and IFN-γ treatment impairs DC development. Eur. J. Immunol. 2009, 39, 2865–2876. [Google Scholar] [CrossRef] [PubMed]
- Lun, X.; Alain, T.; Zemp, F.J.; Zhou, H.; Rahman, M.M.; Hamilton, M.G.; McFadden, G.; Bell, J.; Senger, D.L.; Forsyth, P.A. Myxoma virus virotherapy for glioma in immunocompetent animal models: optimizing administration routes and synergy with rapamycin. Cancer Res. 2010, 70, 598–608. [Google Scholar] [CrossRef] [PubMed]
- Bartee, E.; Mohamed, M.R.; Lopez, M.C.; Baker, H. V.; McFadden, G. The Addition of Tumor Necrosis Factor plus Beta Interferon Induces a Novel Synergistic Antiviral State against Poxviruses in Primary Human Fibroblasts. J. Virol. 2009, 83, 498–511. [Google Scholar] [CrossRef]
- Arnold, K.A.; Peterson, L.F.; Beck, L.A.; Brewer, M.G. JAK Signaling Is Critically Important in Cytokine-Induced Viral Susceptibility of Keratinocytes. Int. J. Mol. Sci. 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Ashton, L. V; Graham, B.; Afzali, M.F.; Gustafson, D.; MacNeill, A.L. <p>Treatment of an Alveolar Rhabdomyosarcoma Allograft with Recombinant Myxoma Virus and Oclacitinib</p>. Oncolytic Virotherapy 2020, Volume 9, 17–29. [Google Scholar] [CrossRef]
- Lun, X.Q.; Zhou, H.; Alain, T.; Sun, B.; Wang, L.; Barrett, J.W.; Stanford, M.M.; McFadden, G.; Bell, J.; Senger, D.L.; et al. Targeting human medulloblastoma: oncolytic virotherapy with myxoma virus is enhanced by rapamycin. Cancer Res. 2007, 67, 8818–8827. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.; Gong, W.; Ma, M.; Chu, B.; Qin, Y.; Zhang, M.; Lun, X.; McFadden, G.; Forsyth, P.; Yang, Y.; et al. Targeting gallbladder cancer: oncolytic virotherapy with myxoma virus is enhanced by rapamycin in vitro and further improved by hyaluronan in vivo. Mol. Cancer 2014, 13, 82. [Google Scholar] [CrossRef]
- France, M.R.; Thomas, D.L.; Liu, J.; McFadden, G.; MacNeill, A.L.; Roy, E.J. Intraventricular injection of myxoma virus results in transient expression of viral protein in mouse brain ependymal and subventricular cells. J. Gen. Virol. 2011, 92, 195–199. [Google Scholar] [CrossRef]
- Zemp, F.J.; McKenzie, B.A.; Lun, X.; Reilly, K.M.; Mcfadden, G.; Yong, V.W.; Forsyth, P.A. Cellular factors promoting resistance to effective treatment of glioma with oncolytic Myxoma virus. Cancer Res. 2014, 74, 7260–73. [Google Scholar] [CrossRef]
- Stanford, M.M.; Shaban, M.; Barrett, J.W.; Werden, S.J.; Gilbert, P.A.; Bondy-Denomy, J.; Mackenzie, L.; Graham, K.C.; Chambers, A.F.; McFadden, G. Myxoma virus oncolysis of primary and metastatic B16F10 mouse tumors in vivo. Mol. Ther. 2008, 16, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Stanford, M.M.; Barrett, J.W.; Nazarian, S.H.; Werden, S.; McFadden, G. Oncolytic virotherapy synergism with signaling inhibitors: Rapamycin increases myxoma virus tropism for human tumor cells. J. Virol. 2007, 81, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.L.; Doty, R.; Tosic, V.; Liu, J.; Kranz, D.M.; McFadden, G.; Macneill, A.L.; Roy, E.J. Myxoma virus combined with rapamycin treatment enhances adoptive T cell therapy for murine melanoma brain tumors. Cancer Immunol. Immunother. 2011, 60, 1461–1472. [Google Scholar] [CrossRef] [PubMed]
- Moerdyk-Schauwecker, M.; Shah, N.R.; Murphy, A.M.; Hastie, E.; Mukherjee, P.; Grdzelishvili, V.Z. Resistance of pancreatic cancer cells to oncolytic vesicular stomatitis virus: Role of type I interferon signaling. Virology 2013, 436, 221–234. [Google Scholar] [CrossRef]
- Dold, C.; Rodriguez Urbiola, C.; Wollmann, G.; Egerer, L.; Muik, A.; Bellmann, L.; Fiegl, H.; Marth, C.; Kimpel, J.; Von Laer, D. Application of interferon modulators to overcome partial resistance of human ovarian cancers to VSV-GP oncolytic viral therapy. Mol. Ther. - Oncolytics 2016, 3, 16021. [Google Scholar] [CrossRef] [PubMed]
- Adelfinger, M.; Gentschev, I.; Grimm de Guibert, J.; Weibel, S.; Langbein-Laugwitz, J.; Härtl, B.; Murua Escobar, H.; Nolte, I.; Chen, N.G.; Aguilar, R.J.; et al. Evaluation of a New Recombinant Oncolytic Vaccinia Virus Strain GLV-5b451 for Feline Mammary Carcinoma Therapy. PLoS One 2014, 9, e104337. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Zarate, D.; Liu, Y.-P.; Suksanpaisan, L.; Russell, S.; Peng, K.-W. Overcoming cancer cell resistance to VSV oncolysis with JAK1/2 inhibitors. Cancer Gene Ther. 2013, 20, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, M.; Shah, N.R.; Felt, S.A.; Grdzelishvili, V.Z. Breaking resistance of pancreatic cancer cells to an attenuated vesicular stomatitis virus through a novel activity of IKK inhibitor TPCA-1. Virology 2015, 485, 340–354. [Google Scholar] [CrossRef] [PubMed]
- Ghonime, M.G.; Cassady, K.A. Combination Therapy Using Ruxolitinib and Oncolytic HSV Renders Resistant MPNSTs Susceptible to Virotherapy. Cancer Immunol. Res. 2018, 6, 1499–1510. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, C.; Iankov, I.D.; Anderson, S.K.; Aderca, I.; Leontovich, A.A.; Maurer, M.J.; Oberg, A.L.; Schroeder, M.A.; Giannini, C.; Greiner, S.M.; et al. Constitutive interferon pathway activation in tumors as an efficacy determinant following oncolytic virotherapy. J. Natl. Cancer Inst. 2018, 110, 1123–1132. [Google Scholar] [CrossRef]
- Keller, A.; Wingelhofer, B.; Peter, B.; Bauer, K.; Berger, D.; Gamperl, S.; Reifinger, M.; Cerny-Reiterer, S.; Moriggl, R.; Willmann, M.; et al. The JAK2/STAT5 signaling pathway as a potential therapeutic target in canine mastocytoma. Vet. Comp. Oncol. 2018, 16, 55–68. [Google Scholar] [CrossRef]
- Sebio, A.; Wilky, B.A.; Keedy, V.L.; Jones, R.L. The current landscape of early drug development for patients with sarcoma in the immunotherapy era. Future Oncol. 2018, 14, 1197. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Timeline of sample collections and treatments in dogs with soft tissue sarcoma.
Figure 2.
MXYV-red growth rates in canine cells. Viral titers (foci-forming units/mL) in canine soft tissue sarcoma cells (STS-1 and STS-2) and non-cancerous cells (Fibroblasts) were measured by plaque assay. Briefly, confluent cells were inoculated with MYXV-red at 0.1 moi; cells were collected at 0, 3, 6, 12, 18, 24, 36, 48, and 72 hours post-inoculation; and the viral titer of each sample was determined in RK-13 cells. The graph plots the means and standard error of viral titers in four replicate wells. Significant MYXV-red replication was observed in STS cells.
Figure 2.
MXYV-red growth rates in canine cells. Viral titers (foci-forming units/mL) in canine soft tissue sarcoma cells (STS-1 and STS-2) and non-cancerous cells (Fibroblasts) were measured by plaque assay. Briefly, confluent cells were inoculated with MYXV-red at 0.1 moi; cells were collected at 0, 3, 6, 12, 18, 24, 36, 48, and 72 hours post-inoculation; and the viral titer of each sample was determined in RK-13 cells. The graph plots the means and standard error of viral titers in four replicate wells. Significant MYXV-red replication was observed in STS cells.
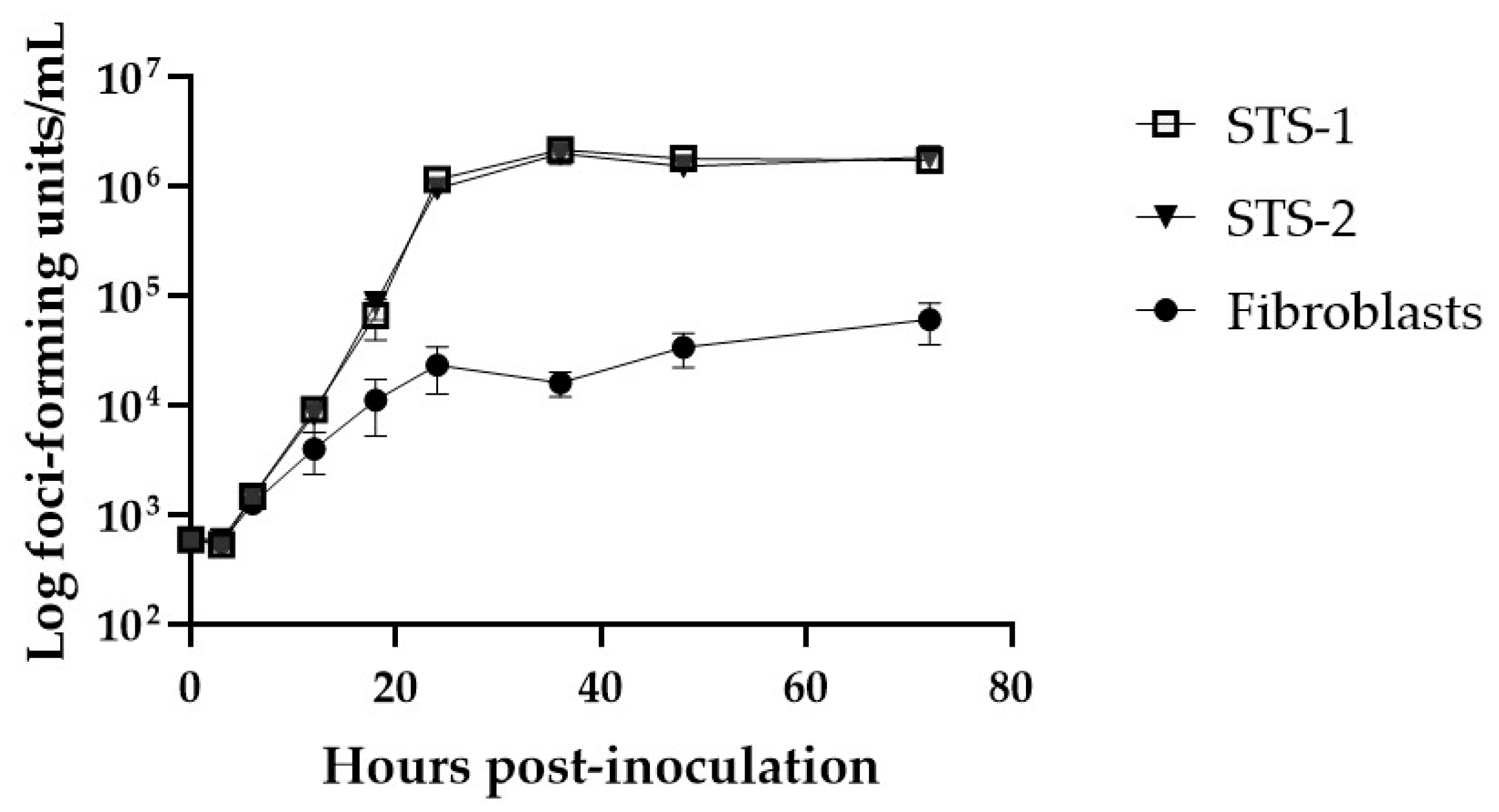
Figure 3.
Photomicrographs of representative canine cell cultures (100× magnification). Fluorescent images taken at 48 and 72 hours post-inoculation (hpi) with MYXV-red (0.1 moi) are shown for two canine soft tissue sarcoma (STS) cell isolates and canine fibroblasts. (Fibroblasts were isolated from a dog that did not have cancer.) Visible cytotoxic effects were mild even in cancer cells that expressed a significant amount of fluorescent protein.
Figure 3.
Photomicrographs of representative canine cell cultures (100× magnification). Fluorescent images taken at 48 and 72 hours post-inoculation (hpi) with MYXV-red (0.1 moi) are shown for two canine soft tissue sarcoma (STS) cell isolates and canine fibroblasts. (Fibroblasts were isolated from a dog that did not have cancer.) Visible cytotoxic effects were mild even in cancer cells that expressed a significant amount of fluorescent protein.
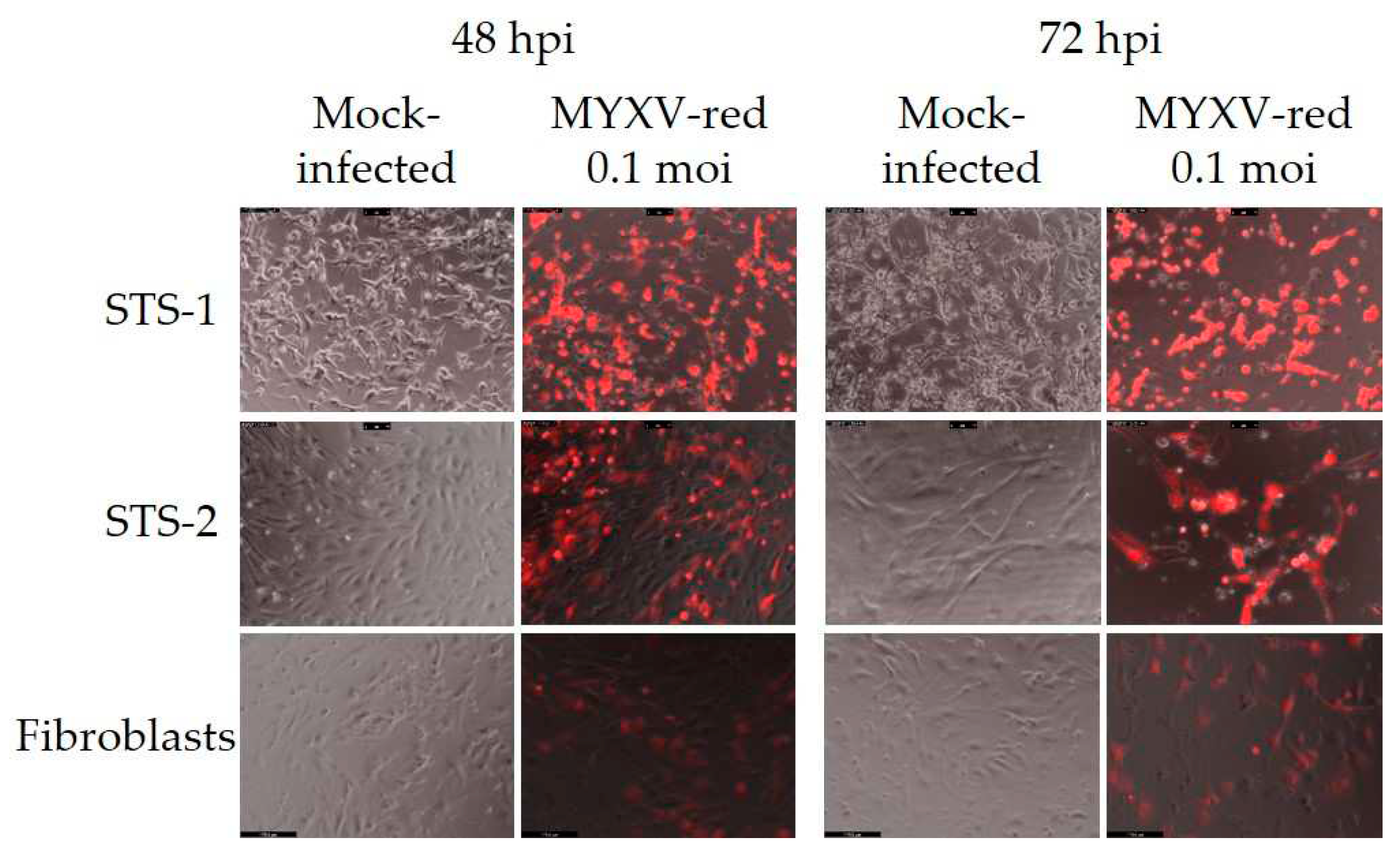
Figure 4.
Viability of mock-infected and MYXV-red-inoculated (0.1 moi) canine cells 24, 48, and 72 hours post-inoculation (hpi). A CellTIter-Glo Cell Viability Assay (Promega, Madison, WI, USA) was used to detect viable (luminescent) cells. Data were calculated as a percentage of the luminescence signal in untreated cells. Six to thirty replicate wells were averaged. Cell viability was significantly decreased in cancer soft tissue sarcoma cells (STS-1 and STS-2) as early as 24 hpi and in all three cell cultures 72 hpi (*p < .05, **p < .01).
Figure 4.
Viability of mock-infected and MYXV-red-inoculated (0.1 moi) canine cells 24, 48, and 72 hours post-inoculation (hpi). A CellTIter-Glo Cell Viability Assay (Promega, Madison, WI, USA) was used to detect viable (luminescent) cells. Data were calculated as a percentage of the luminescence signal in untreated cells. Six to thirty replicate wells were averaged. Cell viability was significantly decreased in cancer soft tissue sarcoma cells (STS-1 and STS-2) as early as 24 hpi and in all three cell cultures 72 hpi (*p < .05, **p < .01).
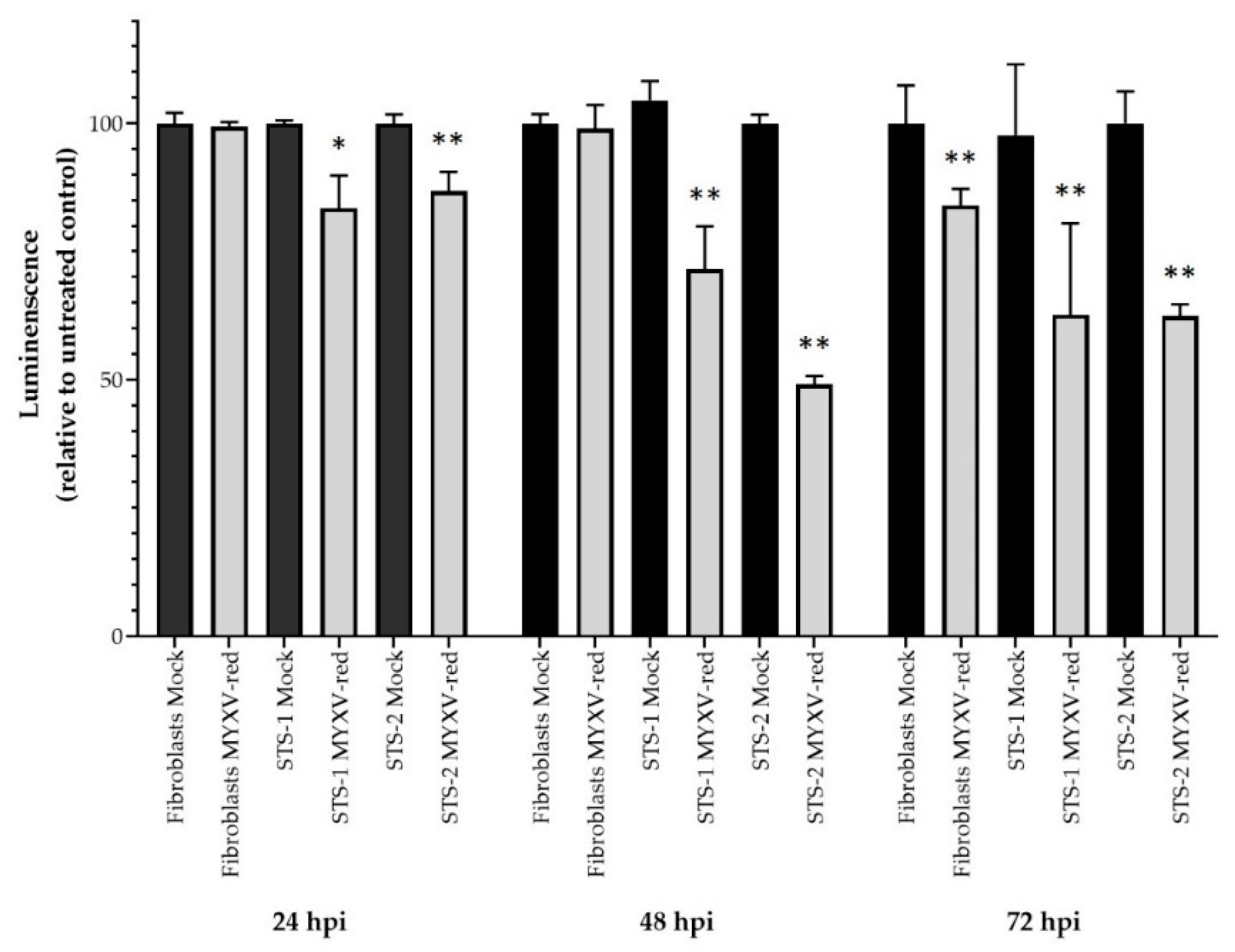
Figure 5.
Heatmaps of pathway signatures analyzed using nCounter 4.0 Advanced Analysis software. (A) Mock-infected FB, STS-1, and STS-2 cells had distinctive pathway signatures as compared to mock-infected FB at 4 hpi. Pathway signatures that were upregulated in both STS-1 and STS-2 cells are highlighted by the orange box. (B) MYXV- and MYXVΔSERP2-inoculated FB were compared to mock-infected FB at 4 hpi. Pathway signatures that were upregulated following MYXV or MYXVΔSERP2 inoculation are highlighted. (C) MYXV- and MYXVΔSERP2-inoculated STS-1 cells were compared to mock-infected STS-1 at 4 hpi. Pathway signatures that were upregulated following MYXV or MYXVΔSERP2 inoculation are highlighted. (D) MYXV- and MYXVΔSERP2-inoculated STS-2 cells were compared to mock-infected STS-2 at 4 hpi. Pathway signatures that were upregulated following MYXV or MYXVΔSERP2 inoculation are highlighted.
Figure 5.
Heatmaps of pathway signatures analyzed using nCounter 4.0 Advanced Analysis software. (A) Mock-infected FB, STS-1, and STS-2 cells had distinctive pathway signatures as compared to mock-infected FB at 4 hpi. Pathway signatures that were upregulated in both STS-1 and STS-2 cells are highlighted by the orange box. (B) MYXV- and MYXVΔSERP2-inoculated FB were compared to mock-infected FB at 4 hpi. Pathway signatures that were upregulated following MYXV or MYXVΔSERP2 inoculation are highlighted. (C) MYXV- and MYXVΔSERP2-inoculated STS-1 cells were compared to mock-infected STS-1 at 4 hpi. Pathway signatures that were upregulated following MYXV or MYXVΔSERP2 inoculation are highlighted. (D) MYXV- and MYXVΔSERP2-inoculated STS-2 cells were compared to mock-infected STS-2 at 4 hpi. Pathway signatures that were upregulated following MYXV or MYXVΔSERP2 inoculation are highlighted.
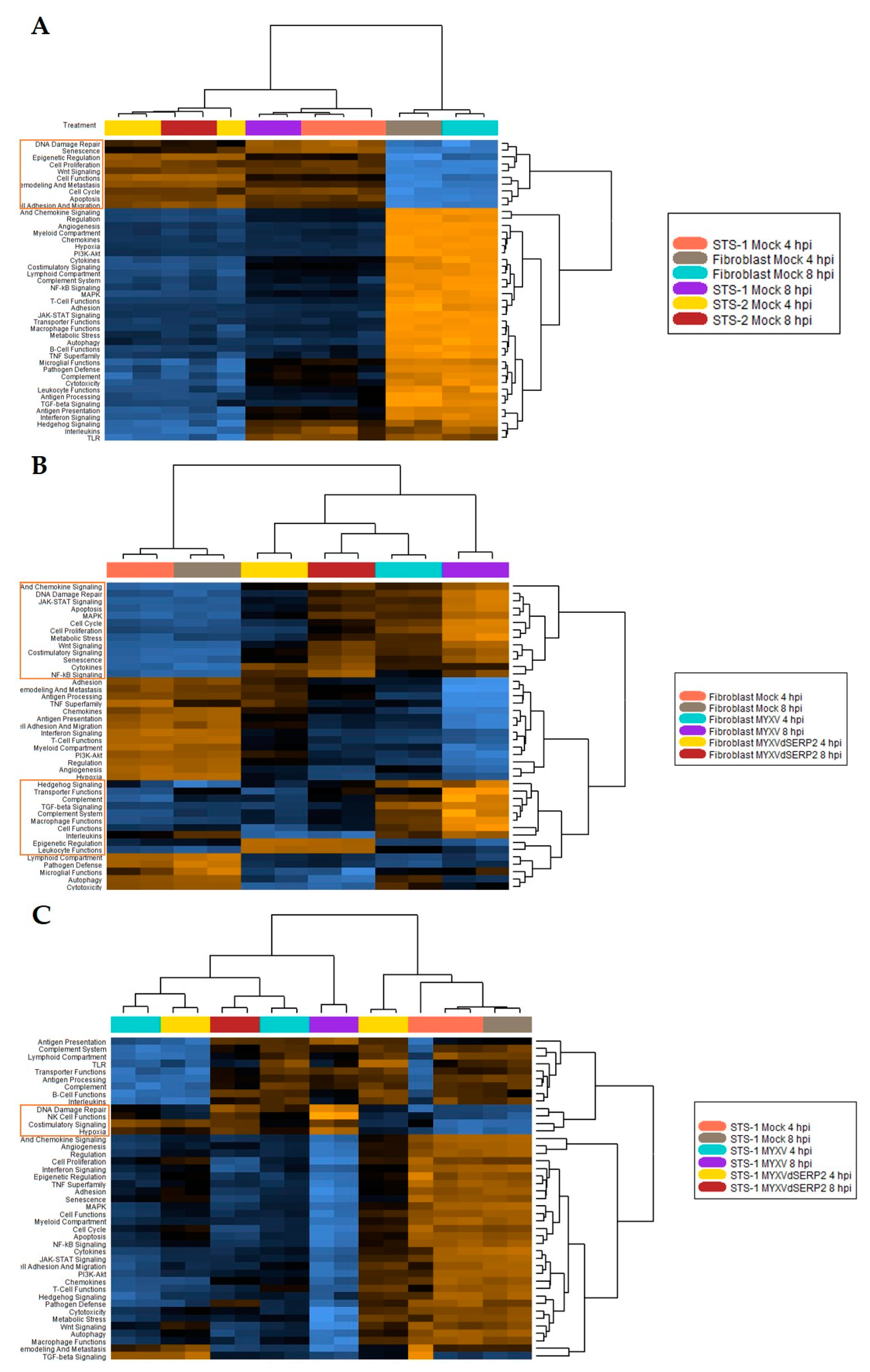

Figure 6.
Pathway signaling scores calculated using nCounter 4.0 Advanced Analysis software. MYXV- and MYXVΔSERP2-inoculated cells were compared to the same mock-infected cell type at 4 hpi. JAK-STAT and Cytokine/Chemokine Signaling scores were increased in canine FB following virus infection, but not in sarcoma cells (STS-1 and STS-2).
Figure 6.
Pathway signaling scores calculated using nCounter 4.0 Advanced Analysis software. MYXV- and MYXVΔSERP2-inoculated cells were compared to the same mock-infected cell type at 4 hpi. JAK-STAT and Cytokine/Chemokine Signaling scores were increased in canine FB following virus infection, but not in sarcoma cells (STS-1 and STS-2).
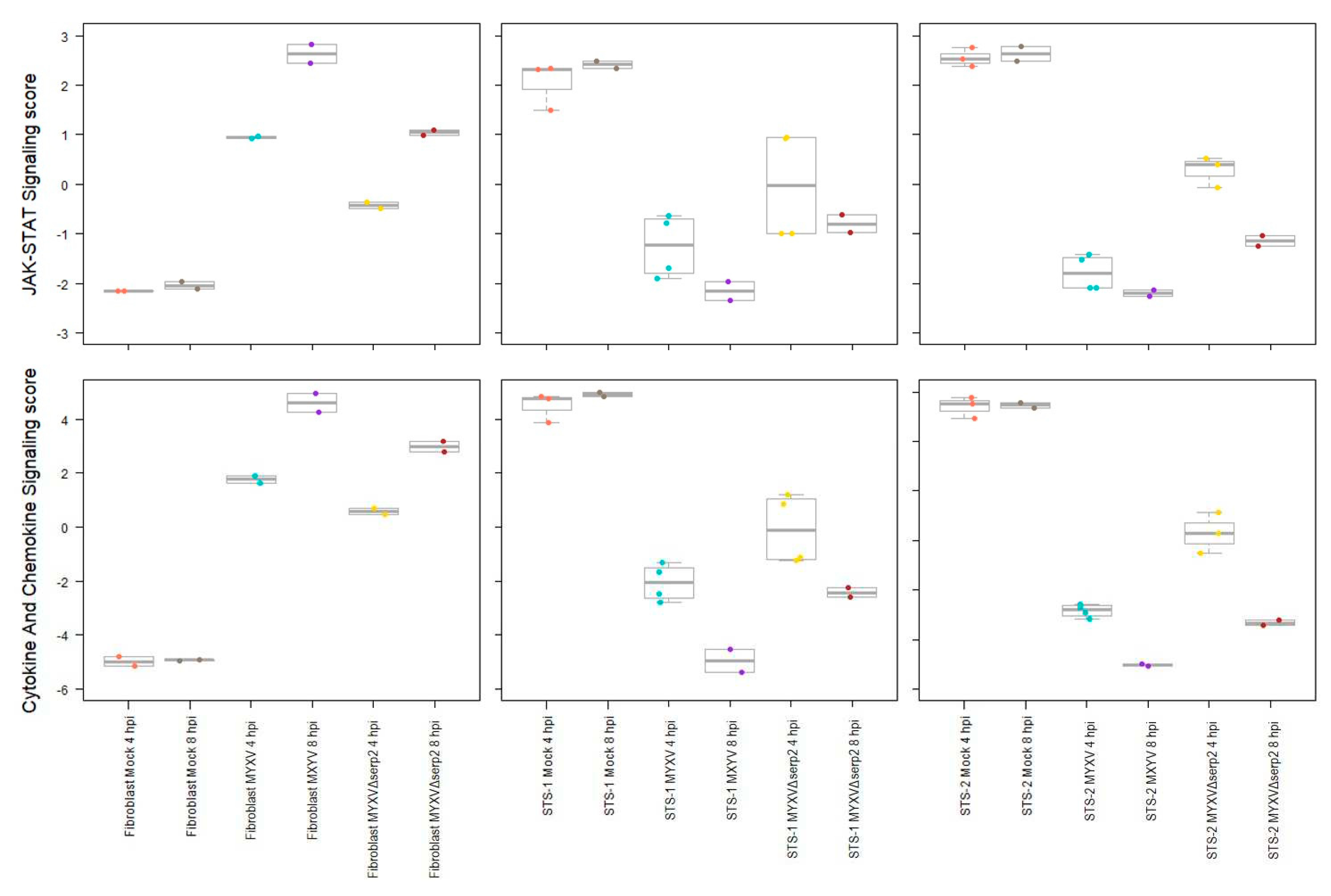
Figure 7.
Heatmaps of fold-changes in selected cytokine transcripts detected using NanoString technology. The numbers of mRNA transcripts that were detected in cells inoculated with virus were compared to mock-infected cells collected at the same time points (4 and 8 hpi). (A) MYXV-inoculated STS-1 cells and FB. (B) MXYVΔSERP2-inoculated STS-1 cells and FB.
Figure 7.
Heatmaps of fold-changes in selected cytokine transcripts detected using NanoString technology. The numbers of mRNA transcripts that were detected in cells inoculated with virus were compared to mock-infected cells collected at the same time points (4 and 8 hpi). (A) MYXV-inoculated STS-1 cells and FB. (B) MXYVΔSERP2-inoculated STS-1 cells and FB.
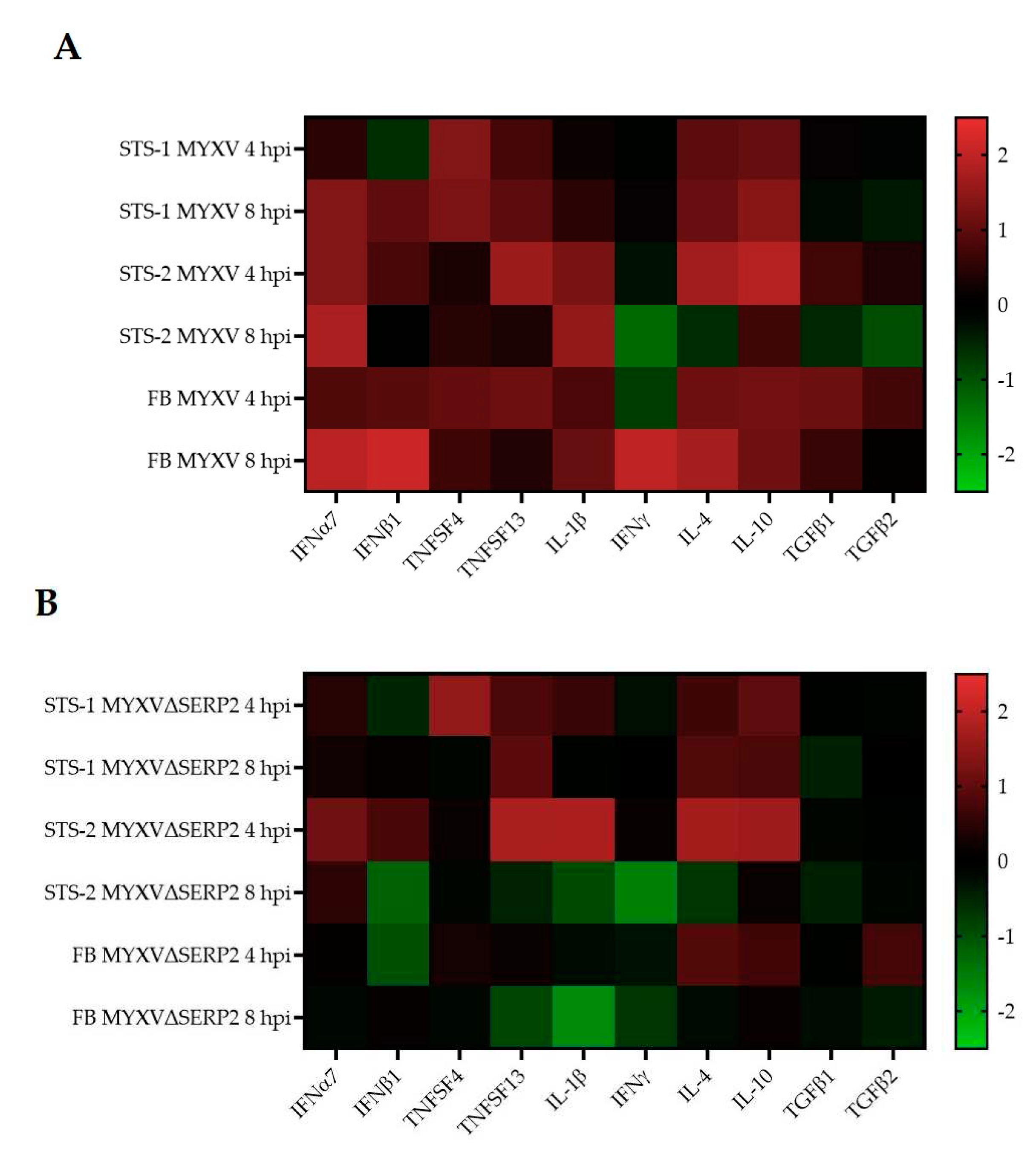
Figure 8.
Heatmaps of fold-changes in RQ values for selected cytokines detected using qPCR. RQ values from virus inoculated cells were compared to mock-infected cells collected at the same time points (4 and 24 hpi). (A) MYXV-inoculated STS-1 cells and FB. (B) MXYVΔSERP2-inoculated STS-1 cells and FB.
Figure 8.
Heatmaps of fold-changes in RQ values for selected cytokines detected using qPCR. RQ values from virus inoculated cells were compared to mock-infected cells collected at the same time points (4 and 24 hpi). (A) MYXV-inoculated STS-1 cells and FB. (B) MXYVΔSERP2-inoculated STS-1 cells and FB.
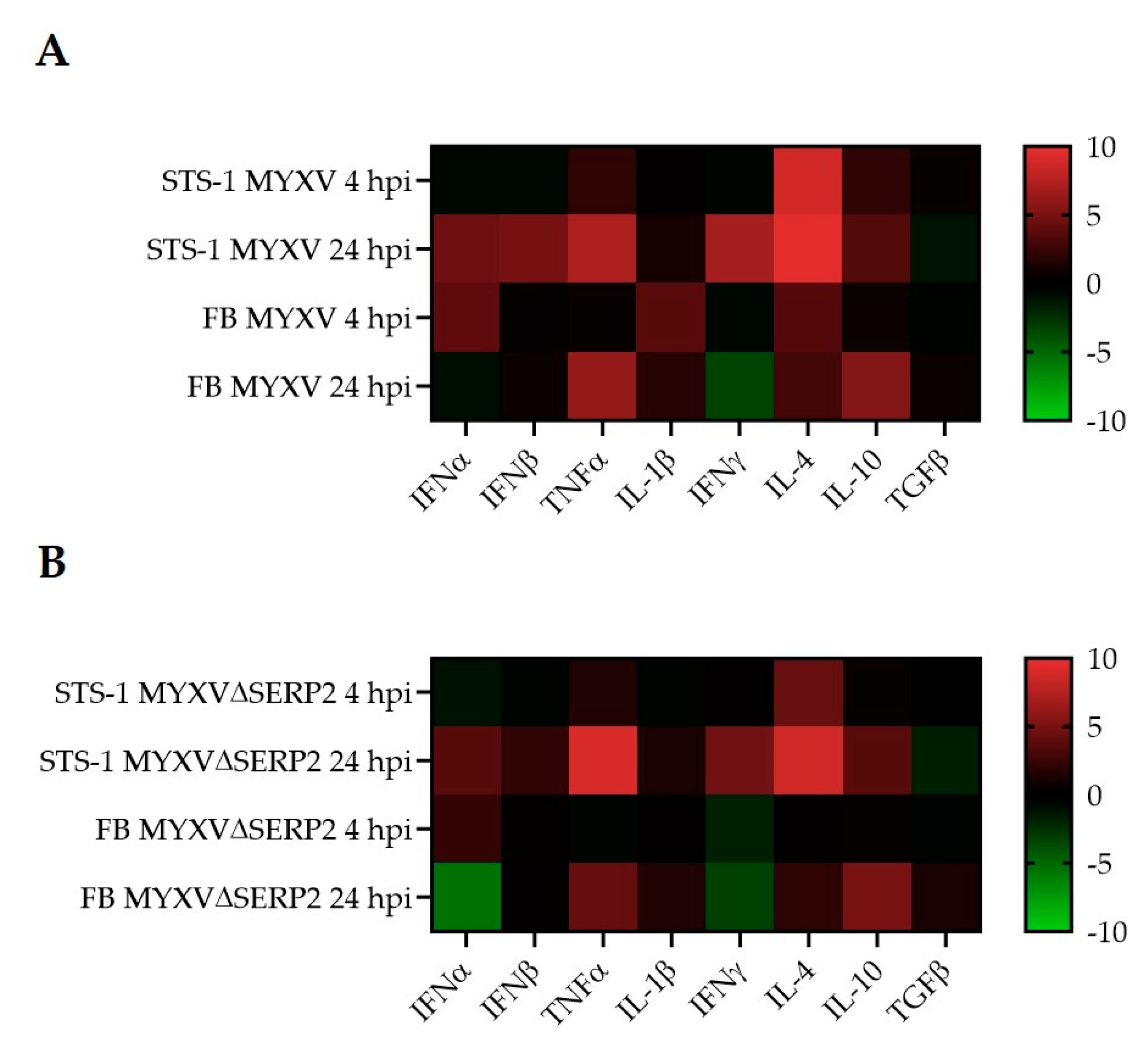
Figure 9.
Cytotoxic responses of canine sarcoma cells after 72 h exposure to cytokines [IFNα (500 U/mL), IFNβ (500 U/mL), TNFα (20 ng/mL), and IFNγ (100 U/mL)]. A Cell Titer Blue® Cell Viability Assay (Promega, Madison, WI, USA) was used to detect viable (fluorescing) cells. Data are calculated as a percentage of the fluorescence signal in untreated cells. Experiments were performed a minimum of three times for each cell culture. No statistical decreases in cell viability were observed. *A few statistically significant increases in cell viability were noted (p < .05).
Figure 9.
Cytotoxic responses of canine sarcoma cells after 72 h exposure to cytokines [IFNα (500 U/mL), IFNβ (500 U/mL), TNFα (20 ng/mL), and IFNγ (100 U/mL)]. A Cell Titer Blue® Cell Viability Assay (Promega, Madison, WI, USA) was used to detect viable (fluorescing) cells. Data are calculated as a percentage of the fluorescence signal in untreated cells. Experiments were performed a minimum of three times for each cell culture. No statistical decreases in cell viability were observed. *A few statistically significant increases in cell viability were noted (p < .05).
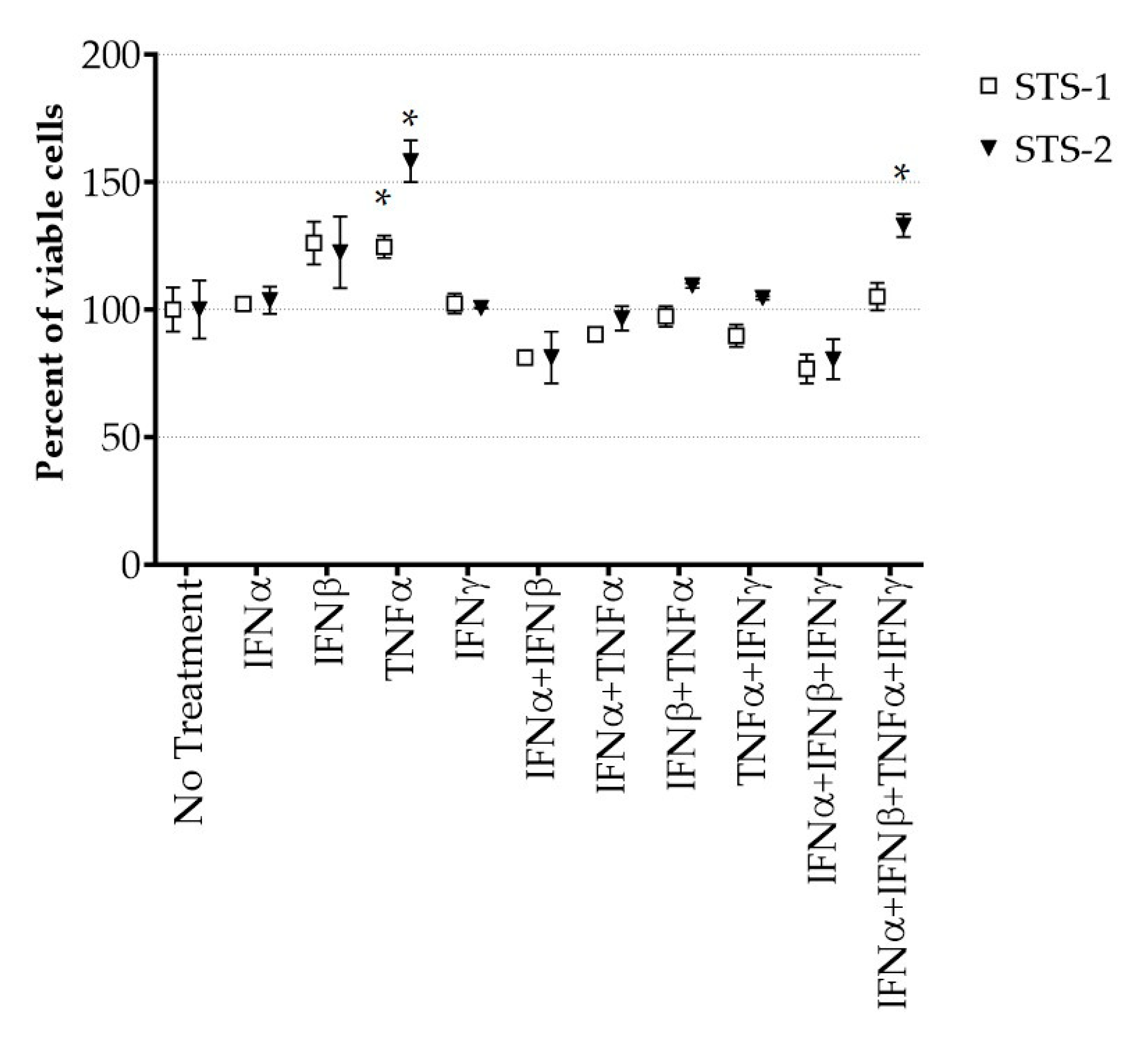
Figure 10.
Viral titers from sarcoma cells collected 72 hpi MYXV-red (0.1 moi) and treated with recombinant canine cytokines. Data columns are arranged in the following order for each cell: no treatment, IFNα (500 U/mL), IFNβ (500 U/mL), TNFα (20 ng/mL), IFNγ (100 U/mL), IFNα (500 U/mL) + IFNβ (500 U/mL), IFNα (500 U/mL) + TNFα (20 ng/mL), IFNβ (500 U/mL) + TNFα (20 ng/mL), TNFα (20 ng/mL) + IFNγ (100 U/mL), IFNα (500 U/mL) + IFNβ (500 U/mL) + IFNγ (100 U/mL), and IFNα (500 U/mL) + IFNβ (500 U/mL) + TNFα (20 ng/mL) + IFNγ (100 U/mL). *Statistically significant reductions in virus titers were observed (p < .05).
Figure 10.
Viral titers from sarcoma cells collected 72 hpi MYXV-red (0.1 moi) and treated with recombinant canine cytokines. Data columns are arranged in the following order for each cell: no treatment, IFNα (500 U/mL), IFNβ (500 U/mL), TNFα (20 ng/mL), IFNγ (100 U/mL), IFNα (500 U/mL) + IFNβ (500 U/mL), IFNα (500 U/mL) + TNFα (20 ng/mL), IFNβ (500 U/mL) + TNFα (20 ng/mL), TNFα (20 ng/mL) + IFNγ (100 U/mL), IFNα (500 U/mL) + IFNβ (500 U/mL) + IFNγ (100 U/mL), and IFNα (500 U/mL) + IFNβ (500 U/mL) + TNFα (20 ng/mL) + IFNγ (100 U/mL). *Statistically significant reductions in virus titers were observed (p < .05).
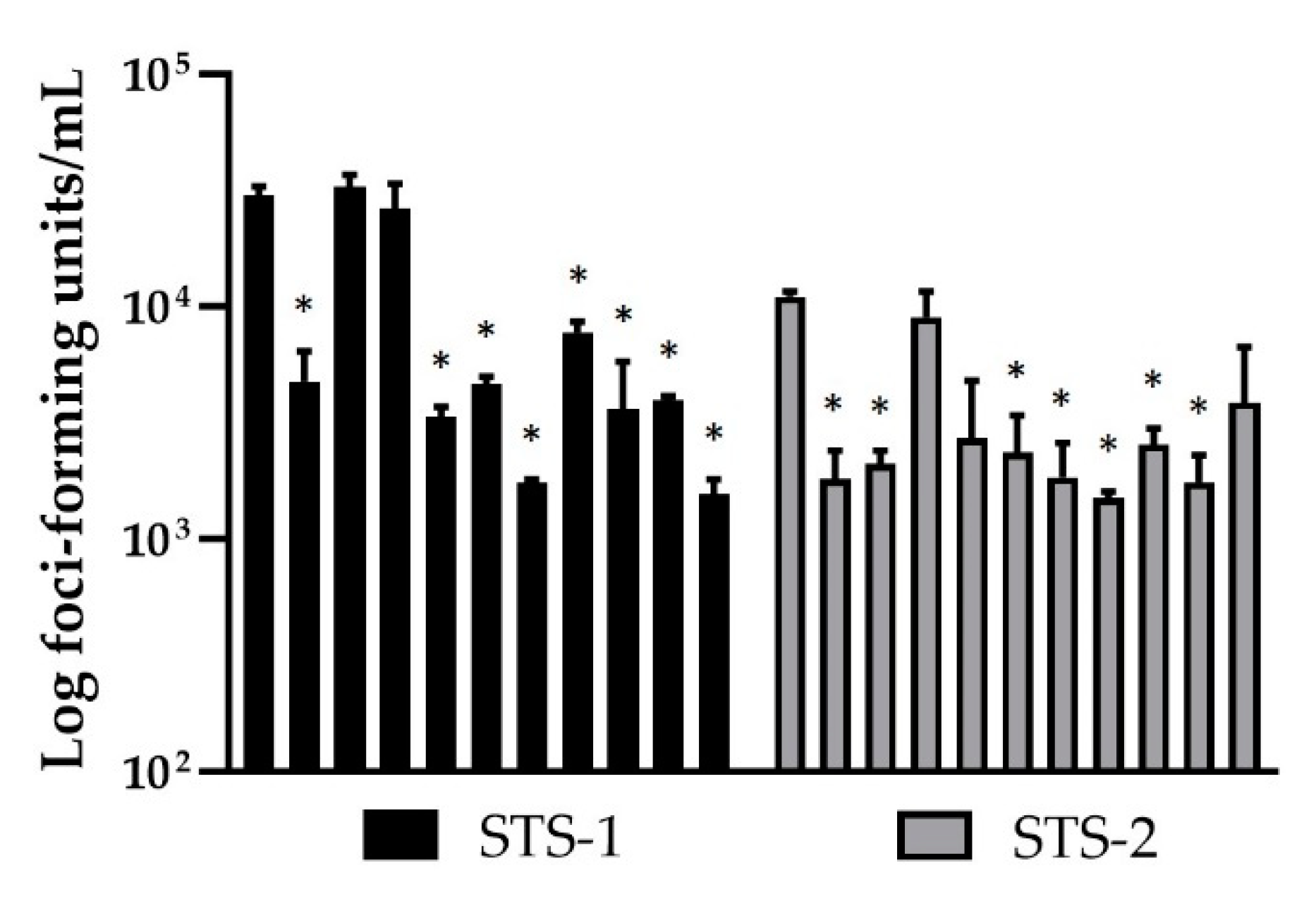
Figure 11.
MYXV-red replication (ffu/mL) in canine cells after treatment with antibodies that block cytokine function. *Antibodies (100 ng/mL) against IFNα, TNFα, and/or IFNγ that caused significant changes in MYXV-red replication at 72 hpi (0.1 moi) as compared to untreated cells (p < .05).
Figure 11.
MYXV-red replication (ffu/mL) in canine cells after treatment with antibodies that block cytokine function. *Antibodies (100 ng/mL) against IFNα, TNFα, and/or IFNγ that caused significant changes in MYXV-red replication at 72 hpi (0.1 moi) as compared to untreated cells (p < .05).
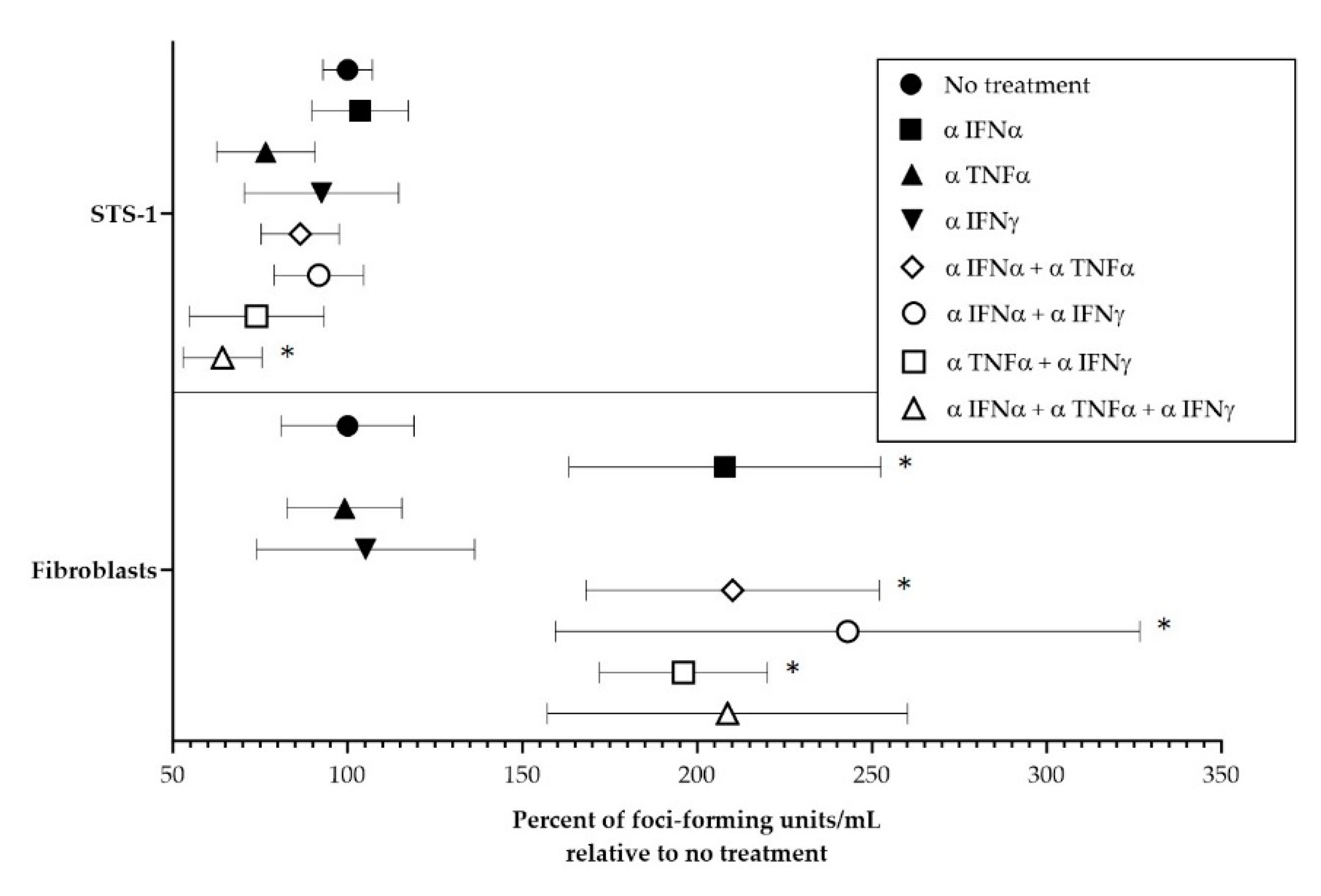
Figure 12.
Virus replication following incubation with heat-inactivated canine sera collected on Day 28. (A) Data indicating that neutralizing antibodies were not developed in Dog 9 by Day 28 after post-operative treatment with MYXVΔSERP2 was given on Days 0 and 14. (B) Data indicating stimulation of neutralizing antibodies in Dog 8 by Day 28.
Figure 12.
Virus replication following incubation with heat-inactivated canine sera collected on Day 28. (A) Data indicating that neutralizing antibodies were not developed in Dog 9 by Day 28 after post-operative treatment with MYXVΔSERP2 was given on Days 0 and 14. (B) Data indicating stimulation of neutralizing antibodies in Dog 8 by Day 28.
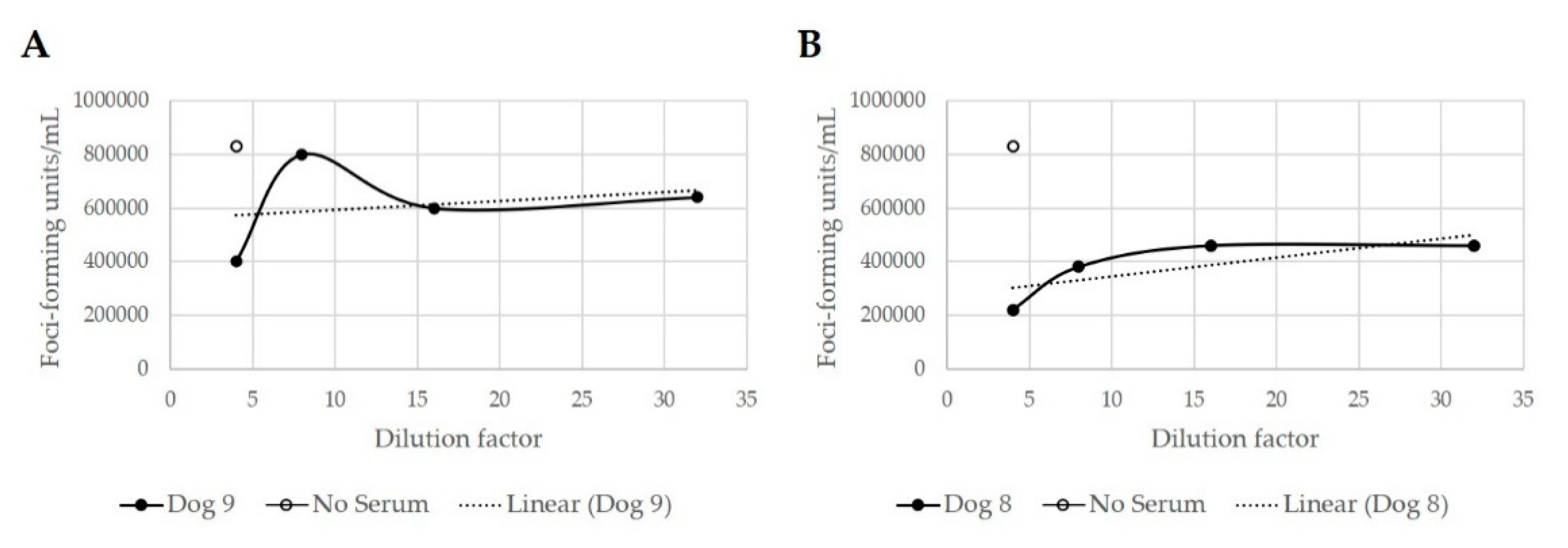
Figure 13.
Evaluation of serum cytokine concentrations in dogs treated with MYXV∆SERP2 alone or in combination with oclacitinib (O+MYXV∆SERP2). Post-treatment cytokine concentration results minus pre-treatment results are graphed. Error bars indicate data range. Note that pre-treatment samples were collected prior to surgery (Day 0) for MYXV∆SERP2-treated dogs and prior to oclacitinib treatment (~Day -7) in O+MYXV∆SERP2-treated dogs.
Figure 13.
Evaluation of serum cytokine concentrations in dogs treated with MYXV∆SERP2 alone or in combination with oclacitinib (O+MYXV∆SERP2). Post-treatment cytokine concentration results minus pre-treatment results are graphed. Error bars indicate data range. Note that pre-treatment samples were collected prior to surgery (Day 0) for MYXV∆SERP2-treated dogs and prior to oclacitinib treatment (~Day -7) in O+MYXV∆SERP2-treated dogs.
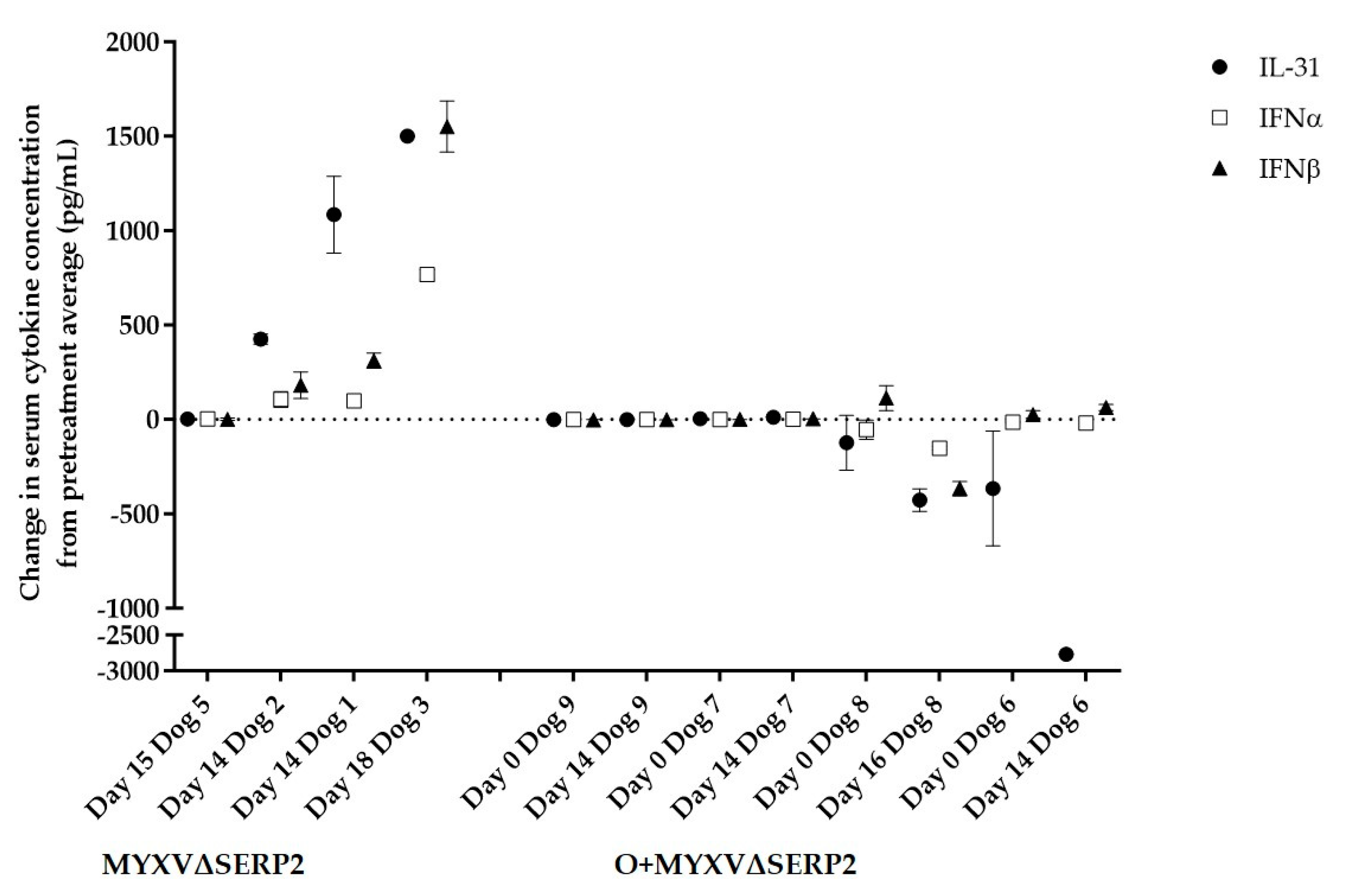

Table 1.
Primer sets used to amplify canine β-actin and cytokine cDNAs in cultured canine cells.
| Target cDNA | Forward primer 5’…3’ | Reverse primer 5’…3’ | Reference | GenBank accession number | Product size (base pairs) |
|---|---|---|---|---|---|
| β-actin | CCG CGA GAA GAT GAC CCA GA | GTG AGG ATC TTC ATG AGG TAG TCG G | [47] | Z70044 | 81 |
| IFNα | TGG GAC AGA TGA GGA GAC TCT C | GAA GAC CTT CTG GGT CAT CAC G | [48] | AB125936* | 143 |
| IFNβ | CCA GTT CCA GAA GGA GGA CA | TGT CCC AGG TGA AGT TTT CC | [47] | NM_001135787 | 200 |
| TNFα | GAG CCG ACG TGC CAA TG | CAA CCC ATC TGA CGG CAC TA | [47] | Z70046 | 79 |
| IL-1β | TCT CCC ACC AGC TCT GTA ACA A | GCA GGG CTT CAG CTT CTC | [47] | Z70047 | 80 |
| IFNγ | AGC GCA AGG CGA TAA ATG | GCG GCC TCG AAA CAG ATT | [48] | NM_001003174 | 121 |
| IL-4 | CAT CCT CAC AGC GAG AAA CG | CCT TAT CGC TTG TGT TCT TTG GA | [47] | AF054833 | 83 |
| IL-10 | CGC TGT CAC CGA TTT CTT CC | CTG GAG CTT ACT AAA TGC GCT CT | [47] | U33843 | 78 |
| TGFβ | CAA GGA TCT GGG CTG GAA GTG GA | CCA GGA CCT TGC TGT ACT GCG TGT | [48] | NM_001003309 | 113 |
*Primer sequence amplifies products for IFNα subtypes 1 (M28624), 3 (M28626), 4 (AB102731), 5 (AB125934), 6 (NM_001007128), 7 (AB125936), and 8 (AB125937).
Table 2.
Transcripts with > 10-fold changes in numbers detected using NanoString technology in MYXV-inoculated canine fibroblasts and sarcoma cells (STS-1 and STS-2) as compared to mock-infected cells collected at 8 hours post-inoculation.
Table 2.
Transcripts with > 10-fold changes in numbers detected using NanoString technology in MYXV-inoculated canine fibroblasts and sarcoma cells (STS-1 and STS-2) as compared to mock-infected cells collected at 8 hours post-inoculation.
| Canine Cells | Target | Description | Fold Change |
|---|---|---|---|
| Fibroblasts | PIK3CD | phosphatidylinositol-4,5-bisphosphate 3-kinase, catalytic subunit delta | -67.5 |
| BRCA1 | breast cancer 1, early onset | 14.8 | |
| EGR1* | early growth response 1 | 31.4 | |
| SERPINB2* | serpin peptidase inhibitor, clade B (ovalbumin), member 2 | 69.4 | |
| FOS* | FBJ murine osteosarcoma viral oncogene homolog | 82.7 | |
| STS-1 | SOX10 | SRY (sex determining region Y)-box 10 | -68.5 |
| SELE* | selectin E | 10.2 | |
| MCAM | melanoma cell adhesion molecule | 10.2 | |
| IL10RA* | interleukin 10 receptor, alpha | 12.3 | |
| CTSS | cathepsin S | 14.9 | |
| FOS* | FBJ murine osteosarcoma viral oncogene homolog | 25.8 | |
| KLRA1 | killer cell lectin-like receptor subfamily A, member 1 | 46.0 | |
| CCL20* | chemokine (C-C motif) ligand 20 | 103.5 | |
| STS-2 | PIK3CD | phosphatidylinositol-4,5-bisphosphate 3-kinase, catalytic subunit delta | -118.5 |
| CD97 | CD97 molecule | -68.0 | |
| CTSS* | cathepsin S | 32.5 |
* Transcripts with > 10-fold changes in numbers detected using NanoString technology in MYXVΔSERP2-inoculated canine fibroblasts and sarcoma cells (STS-1 and STS-2) as compared to mock-infected cells collected at 8 hours post-inoculation.
Table 3.
Effect of cytokines on MYXV-red reporter protein expression† in canine cells. Data are expressed as the mean (standard error of the mean) fluorescence units/well relative to no treatment, MYXV-red-inoculated controls.
Table 3.
Effect of cytokines on MYXV-red reporter protein expression† in canine cells. Data are expressed as the mean (standard error of the mean) fluorescence units/well relative to no treatment, MYXV-red-inoculated controls.
| Treatment | STS-1 | STS-2 | FB |
|---|---|---|---|
| No Treatment | 100.00 (1.63) | 100.00 (1.26) | 100.00 (10.28) |
| IFNα | 30.88* (7.66) | 30.10* (7.97) | 40.86* (5.11) |
| IFNβ | 50.24* (5.53) | 32.44* (8.67) | 17.97* (5.62) |
| TNFα | 30.02* (5.06) | 134.50 (15.97) | 107.05 (14.19) |
| IFNγ | 70.65 (18.42) | 97.02 (27.41) | 125.57 (6.14) |
| IFNα + IFNβ | 31.12* (8.57) | 22.18* (5.87) | 47.91* (3.40) |
| IFNα + TNFα | 19.32* (3.30) | 53.54* (9.08) | 55.72* (5.27) |
| IFNβ + TNFα | 17.69* (1.58) | 36.92* (7.86) | 46.03* (6.21) |
| TNFα + IFNγ | 23.10* (5.47) | 82.28 (21.85) | 113.67 (9.87) |
| IFNα + IFNβ + IFNγ | 24.57* (5.91) | 42.33* (8.01) | 43.71* (3.86) |
| IFNα + IFNβ + TNFα + IFNγ | 11.59* (2.13) | 27.12* (6.00) | 36.78* (12.86) |
† Fluorescence units were measured at 72 hours post-inoculation with MYXV-red at 0.1 multiplicity of infection. * Significant (p < .05) decrease in MYXV-red reporter gene expression as compared to the no treatment, MYXV-red-inoculated controls. IFNα (500 U/mL), IFNβ (500 U/mL), TNFα (20 ng/mL), IFNγ (100 U/mL). STS (soft tissue sarcoma), FB (fibroblasts).
Table 4.
Tumor histopathology and outcomes in five canine patients with spontaneous soft tissue sarcoma (STS) treated with post-operative MXYVΔSERP2 and four patients treated with a combination of oclacitinib and post-operative MXYVΔSERP2 (O+MXYVΔSERP2).
Table 4.
Tumor histopathology and outcomes in five canine patients with spontaneous soft tissue sarcoma (STS) treated with post-operative MXYVΔSERP2 and four patients treated with a combination of oclacitinib and post-operative MXYVΔSERP2 (O+MXYVΔSERP2).
| Patient | Post-operative treatment | Excised tumor histopathology | Outcome | Post-operative day |
|---|---|---|---|---|
| 1 | MXYVΔSERP2 | Grade III STS, incompletely excised | Recurrence | 42 |
| 2 | MXYVΔSERP2 | Grade III STS, incompletely excised | Recurrence | 159 |
| 3 | MXYVΔSERP2 | Grade II STS, incompletely excised | Lost to follow-up | 32 |
| 4 | MXYVΔSERP2 | Grade II STS, fibrous capsule <1 mm thick | No regrowth | 916 |
| 5 | MXYVΔSERP2 | Grade II STS, incompletely excised | No regrowth, euthanized due to unrelated causes | 231 |
| 6 | O+MXYVΔSERP2 | Grade III STS, incompletely excised | Recurrence | 188 |
| 7 | O+MXYVΔSERP2 | Grade II STS, incompletely excised | No regrowth, pyogranulomatous inflammation at incision site (Day 31) | 378 |
| 8 | O+MXYVΔSERP2 | Grade II STS, incompletely excised | Recurrence | 40 |
| 9 | O+MXYVΔSERP2 | Grade III STS, completely excised with 2 mm margins | Recurrence | 372 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



