
Submitted:
18 July 2023
Posted:
19 July 2023
You are already at the latest version
Abstract
Cymbidium goeringii, locally known as the spring orchid in Korea, is one of the most important and popular horticultural species in the family Orchidaceae. C. goeringii cultivars were originated from plants with rare phenotypes in wild mountains where pine trees commonly grow. This study aimed to determine the cultivar-specific combined genotypes (CGs) of short sequence repeats (SSRs) by analyzing multiple samples per cultivar of C. goeringii. In this study, we collected more than 4,000 samples from 67 cultivars and determined the genotypes of 12 SSRs. Based on the most frequent combined genotypes (CG1s), the average observed allele number and combined matching probability were 11.8 per marker and 3.118×10-11, respectively. Frequencies of the CG1 in 50 cultivars (n ≥ 10) ranged from 40.9% to 100.0%, with an average of 70.1%. Assuming that individuals with the CG1 are genuine in the corresponding cultivars, approximately 30% of C. goeringii on the farms and markets may be not genuine. The dendrogram of the phylogenetic tree and principal coordinate analysis largely divided the cultivars into three groups according to their countries of origin; however, the genetic distances were not distant among the cultivars. In conclusion, this dataset of C. goeringii cultivar-specific SSR profiles could be used for ecogenetic studies and forensic authentication. This study suggests that genetic authentication should be introduced for the sale of expensive C. goeringii cultivars. We believe that this study will help establish a genetic method for the forensic authentication of C. goeringii cultivars.
Keywords:
goeringii
; cultivar authentication
; microsatellite
; simple sequence repeats (SSRs)
; spring orchid
1. Introduction
Cymbidium goeringii, belonging to the family Orchidaceae is one of the most important and popular horticultural species in East Asia [1,2]. C. goeringii is locally known as the spring orchid in Korea because it blooms in early spring. C. goeringii cultivars are classically divided into two types: the “flower-variant cultivar” showing characteristic phenotypes of flower color and shape and the “leaf-variant cultivar” showing characteristic phenotypes of leaf color or variegation pattern and shape (Figure 1). Plants showing characteristic phenotypes in both leaf and flower are usually called “double-variant cultivar” [3]. In Korea, thousands of C. goeringii cultivars have been registered by two orchid registration organizations: the Korea Orchid Registration Association (KORA; http://www.koreso.com/) and the Registration Committee of the Korea Orchid Union (RCKOU; http://www.kour.or.kr/).
C. goeringii cultivars with horticulturally rare phenotypes are actively traded commercially at high prices through direct sales between sellers and purchasers or online systems. Depending on the cultivar, prices vary widely from a few to hundreds of thousands. Orchid cultivators and purchasers frequently worry about non-genuine cultivars, in which plants belonging to different cultivars are provided instead of the real cultivars.
C. goeringii cultivars originally grew in wild mountains, where pine trees commonly grow. Each spring orchid showing an unusual unique phenotype was selected by orchid collectors and registered as a specific cultivar. Cymbidium species are usually reproduced through self-pollination in wild fields [4-5]; however, the individual number of each cultivar was increased through asexual vegetative propagation as a method of separating the shoots of an individual plant. Theoretically, all individuals originating from a particular cultivar strain are genetically identical when mutations that have occurred after cultivar fixation are ignored. Therefore, forensic discrimination can be to determine whether two C. goeringii plants originate from a common cultivar, using microsatellite profiling [3,6]. This application is similar to microsatellite genotyping and the comparison of genetic profiles between suspects and the evidence(s) collected at crime scenes.
When a new C. goeringii cultivar was registered, whole orchid photograph(s) and phenotypic features were provided to the orchid registration organizations (KORA or RCKOU); however, neither tissue sample nor genetic information was deposited. Therefore, tracking the exact origin of a particular cultivar is difficult. In addition, determining which of the plants with similar phenotypes but different genetic profiles are from the original cultivar, is challenging. In the case of the leaf-variant types, originality could be predicted from the differences in leaf shape, color, and variegation patterns, but it is difficult to predict originality from the phenotypes of the flower-variant types. This is because blooming is observed only in spring and the subtle features of flower colors and shapes differ depending on the cultivation conditions. A recent study suggested that approximately 40% of the purchased spring orchids (with tagged names) among the 10 flower-variant cultivars in Korea were not genuine cultivars [3].
Microsatellites, also known as simple sequence repeats (SSRs) in plant studies, were first reported in C. goeringii by Huang et al and Hyun et al [7,8], then several studies have examined microsatellite markers in molecular genetic studies of Cymbidium species [3,6,9-13]. In particular, Lee et al suggested the potential usefulness of microsatellite combined genotypes as a forensic authentication tool for discriminating among C. goeringii cultivars [3]. Lee et al determined microsatellite combined genotypes using a multiplex PCR for ten cultivars of Korea C. goeringii [3]. Nam et al analyzed the genetic kinship among cultivars using combined genotypes in four closely related cultivar groups [6].
This study aimed to determine the cultivar-specific combined genotypes (CGs) of SSRs by analyzing multiple samples per cultivar in C. goeringii. We collected more than 4,000 samples from 67 cultivars and determined the genotypes of 12 SSRs. In addition, we analyzed the phylogenetic relationships among the examined cultivars.
2. Materials and Methods
2.1. Collection of C. goeringii samples
In this study, 4,046 C. goeringii samples were collected from 66 cultivars (Table 1). Among these, 61 cultivars (n = 3957) were originally collected from the mountains of Korea, whereas five cultivars (n = 86) originated from Japan. Most samples were flower-variant cultivars; some were leaf- or double-variant cultivars. The 61 Korean cultivars were Hwanggeumso, Gwaneum, Boreumdal, Deungdae, Manmool, Cheonhwangso, Daehongbo, Cheonsoo, Cheonjong, Hongdaewang, Cheonunso, Hwansaeng, Wonmyoung, Irwolhwa, Taehongso, Munsubong, Agassi, Geummaek, Haeoreum, Okro, Sagye, Chaeun, Cheongeumso, Seongyeong, Wonhongseol, Dongryeon, Jinna, Jeonju-Hwangu, Cheonbulso, Bulkkot, Hallasan, Cheonsa, Susa, Hongwon, Geumhaso, Honggwiin, Hwangmae, Migeum, Dongcheon, Unmuryeong, Gollyongpo, Sacheonwang, Songjeong, Wangjungwang, Hojeong, Saebyeok, Taegeukseon, Youngchoonso, Geumsusan, Jinjusu, Sinmun, Sebo, Silla, Ilchulhwa, Cheongoksan, Hongun, Jangdan, Miryeon, Hojin, Hyangsu, Namhaeso, and Juhongseong. Six cultivars from Japan and China were collected for genotype comparisons and phylogenetic analyses. Five C. goeringii cultivars of Chanbo, Jilbugeum, Changseongjihwa, Hobakjeon, and Sumunsan were from Japan, In addition to the established cultivar samples, 155 wild C. goeringii samples were collected from Korean mountains. The Chinese cultivar, Hwanguhajeong belonging to C. forestii was sampled as an outgroup (n = 5).
2.2. DNA Purification
Leaves or roots of the orchid plants were cut into 0.5~1 cm long fragments and powdered using a TissueLyser II (Qiagen, Hilden, Germany). Genomic DNA was purified from the powder using the DNeasy Plant Mini Kit (Qiagen). DNA concentration was determined using a NanoDrop 2000 (Thermo Fisher Scientific, Wilmington, NC, USA).
2.3. Multiplex PCR for 12 SSR Markers
The 12 SSR markers were amplified using two multiplex PCR systems. The multiplex system 1 included seven markers: CG415 (HQ842905.1), CG709 (HQ842922.1), CG722 (HQ842923.1), CG787 (HQ842926.1), CG1023 (HQ842937.1), CG1210 (HQ842946.1), and CG1281 (HQ842953.1) and the system 2 included five markers: CG649 (HQ842919.1), CG1028 (HQ842938.1), CG1085 (HQ842942.1), CG1320 (HQ842954.1), and CG1400 (HQ842958.1), respectively. The forward primers were labeled with one of the four fluorescent dyes: VIC, FAM, PET, and NED. The primer sequences, concentrations, and repeat units of the microsatellite markers were as described by Nam et al [6]. PCR mixture was prepared to 10 µL including 20 ng of genomic DNA, primer mixture, and 5 µL of 2 × PCR Master Solution of AccuPower Multiplex PCR Master Mix (Bioneer, Daejeon, Korea). The thermal cycling conditions were as follows: 10 min predenaturation at 95°C, 35 cycles of 30 s at 95°C, 30 s at 58°C, and 1 min at 72°C, and final extension of 30 min at 72°C using a SimpliAmp Thermal Cycler (Applied Biosystems-Applied Biosystems, Foster city, CA, USA). The PCR products were resolved using a SeqStudio Genetic Analyzer (Thermo Fisher-Applied Biosystems), and the genotypes were determined using the Gene Mapper program (NT, Ver. 6.1) (Thermo Fisher-Applied Biosystems). The nomenclatures of the alleles were principally determined by the repeat numbers of the microsatellite motifs according to the recommendation of the DNA Commission of the International Society for Forensic Hemogenetics (ISFH) [14] and based on the information from Hyun et al [8].
2.4. Phylogenetic and Sibling Analysis
Genetic distances among cultivars were determined from similarities in CGs using a GenAlEx (v6.5) [15]. A dendrogram of the phylogenetic tree was constructed from the dissimilarity matrix using the unweighted pair-group method with arithmetic averages. In addition, the dissimilarity matrix was used to perform principal coordinate analysis (PCoA), which graphically represents the genetic relationships among C. goeringii cultivars. The sibling probability using CG profiles was determined using Bayes theory [16].
2.5. Statistical Analysis
Allele frequencies were calculated by counting the number of observed alleles in all examined samples. Reference allele frequencies were obtained from a wild Korean C. goeringii population (n = 155). The combined matching probability (CMP) for the CGs of the 12 SSRs was calculated using the PowerStatsV12 program (Promega, Madison, WI, USA). A simple program based on MATLAB (MathWorks, Natick, MA, USA) was designed to search for similar CG from a pool of several thousand CGs.
3. Results
3.1. Determination of Combined Genotypes
This study determined the CGs of 12 microsatellite markers for 61 Korean cultivars (3,957 samples) and six Japanese and Chinese cultivars (91 samples). In principle, alleles were named by repeating the number of SSR core units, as described earlier [3,6,8]. Most of the markers were analyzed well, but genotyping of some samples failed for markers such as CG649, CG787, CG1023, and CG1085. The failed markers, even when retested from the same plants, were not amplified or were poorly amplified by PCR, suggesting the possibility of variations in the primer sequences.
The most and second most frequent CGs (indicated by CG1 and CG2, respectively) and their CMPs are shown in Table S1. Based on CG1, the average observed allele number was 11.8 per marker with the highest number (16) in CG649 and CG709 and the lowest number (8) in CG1320. The average CMP was 3.118×10-11, ranging from 8.890×10-10 for Daehongbo to 4.496×10-36 for Cheongoksan. This powerful discrimination made it possible to determine whether a C. goeringii individual labelled as a certain cultivar was genuine.
3.2. Determination of Cultivar-Specific Combined Genotypes
Only limited information, such as short phenotypic descriptions and photographs, is available for registered orchid cultivars in Korea. Therefore, it is difficult to determine which genotype is genuine for two samples with different genotype profiles. If the CGs were the same in the two samples, they might have originated from identical plants; otherwise, they might have originated from different plants. For a specific cultivar, the most frequent CG (CG1) among the samples that belonged to the specific cultivar was determined as the representative genotype profile of the corresponding cultivar [3].
In this study, 50 Korean cultivars with a sample size of 10 or more were analyzed for the frequencies of CG1s (Table 2). The frequencies of the CG1s were higher than 50% in most examined cultivars, except for cultivars of Cheonhwangso (41.0%), Hallasan (47.5%), Sacheonwang (40.9%), and Geumsusan (45.5%). In particular, the second frequent CGs (CG1B) were observed at relatively high frequencies of 29.8% in Cheonhwangso and 30.0% in Hallasan. When the most and second frequent CGs were compared in the two cultivars (Cheonhwangso: CG1A: 11–19/14–15/17–34/12–12/24–24/13–13/17–17/15–15/12–12/16–16/12–12/20–20 vs. CG1B: 11–11/15–29.1/17–34/12–20/24–24/13–25/17–17/16–16/17–17/16–16/7–12/13.1–20, and Hallasan: CG1A: 11-15/15-15/17-17/17-17/24-24/13-13/12-12/14-14/17-17/10-16/10-10/13.1-13.1 vs. CG1B: 11-11/15-15/17-17/17-17/24-24/13-13/17-17/14-14/15-17/16-16/10-10/13.1-13.1; matching alleles are underlined), they were considerably similar to each other with high sibling probabilities of 71.016% and 99.997%, suggesting that the plant individuals with both pairs were genetically close relatives. The horticultural phenotypes of plants with CG1A and CG1B were indistinguishable between the two cultivars. Therefore, we concluded that the individuals with either CG1A or CG1B were genuine for the corresponding cultivars. The sum of the frequencies of CG1A and CG1B was 70.8% in Cheonhwangso and 77.5% in Hallasan.
Among the 50 cultivars (n ≥ 10), the frequency of the CG1 ranged from 40.9% (Sacheonwang) to 100.0% (Jinjusu) with an average of 70.1% (Figure 2). Assuming that individuals with the CG1 are genuine to the corresponding cultivars, approximately 30% of C. goeringii on the farms and markets may be not genuine. The average frequency of the second frequent combined genotype (CG2) was 7.3%. For the CG1s observed in 12 cultivars with fewer than 10 samples, they were considered insufficient for assigning as representative SSR profiles for those cultivars (Table S1). Therefore, we considered them as “probable CGs”.
3.3. Tracing Cultivar Origin for Samples Assumed to Be Non-Genuine
This study traced the actual cultivars in 29.9% samples that were suggested to be non-genuine. Most of them did not belong to the 66 C. goeringii cultivars examined in this study; however, some were determined to be CG1 of other cultivars (Table 3).
In the cultivar Hwanggeumso, CG1s were detected in seven cultivars (Gwaneum, Youngchoonso, Namhaeso, Cheonhwangso, Geumhaso, Chanbo, and Hobakjeon). The second most frequent CG2 in Hwanggeumso was identical to the CG1 in Gwaneum (50 times observed). In addition, CG1s of Youngchoonso (seven times) and Namhaeso (five times) were frequently observed in the samples as Hwanggeumso. In the samples collected from the cultivar Gwaneum, five types of CGs were identical to the CG1s of different cultivars (Youngchoonso, Cheongeumso, Geumhaso, Hobakjeon, and Chanbo). Among these, Youngchoonso (30 times) and Hobakjeon (5 times) were frequently observed. CG1 in Gwaneum was frequently identified (6 times) in the cultivar Cheonhwangso. In the samples of cultivars Cheonsoo and Cheonsa, the CG1 of the Japanese cultivar Chanbo was frequently observed 10 times and five times, respectively. In the samples of the cultivar Agassi, CG1 of Jinjusu was identified seven times. In the samples of the cultivar Hongdaewang, CG1 of Jangdan was identified eight times. In the samples of cultivar Munsubong, the CG1 of Hyangsu was identified six times.
The CG1s of the Japanese cultivars Chanbo and Hobakjeon were observed quite frequently in the samples of several different cultivars: Chanbo in Hwanggeumso (three times), Gwaneum (one time), Cheonsoo (ten times), Cheonsa (five times), Hobakjeon in Hwanggeumso (one time), and Gwaneum (five times). For Korean cultivars, the CG1 of Youngchoonso was observed in the samples from three cultivars (Hwanggeumso, Gwaneum, and Cheongeumso); Gwaneum, in two cultivars (Hwanggeumso and Cheongeumso); and Geumhaso, in 2 cultivars (cultivars (Hwanggeumso and Gwaneum). Interestingly, a Chinese cultivar Hwanguhajung which belongs to the C. forestii was once observed in the samples collected by the cultivar Wonmyoung.
3.4. Phylogenetic Analysis among Cultivars
A dendrogram of the phylogenetic tree was prepared from the genetic distance matrix of the CG1s from the 67 cultivars (Figure 3). The dendrogram roughly divided the cultivars into three groups according to their origin. Cultivars of Korean origin were separated from Japanese cultivars (Hobakjeon, Changseongjihwa, Sumunsan, Jilbugeum, and Chanbo) and the C. forestii cultivar (Hwanguhajeong) originating from China.
The C. goeringii cultivars were also separated by the principal coordinate analysis (PCoA). The cumulative percentage variances of three principal components (cum %) accounted for 11.83%, 18.52%, and 25.05% (Figure 4). With features similar to those in the phylogenetic dendrogram, the PCoA roughly divided them into three groups according to their origin. However, the Korean cultivar, Hwansaeng was included in the cluster of Japanese cultivars, and some Korean cultivars, such as Cheonunso and Jinna, were located near the Japanese cultivars. In both the phylogenetic tree and PCoA, the cultivars were poorly separated by variant type.
4. Discussion
Each established cultivar of C. goeringii is asexually propagated by dividing the shoots of a single plant; therefore, individuals constituting a cultivar are genetically identical clones in principle. In this study, we determined the CGs of 12 SSR markers for more than 60 cultivars of C. goeringii using multiplex PCR and subsequent CG profiling. We previously reported CGs of 10 Korean flower-variant cultivars [3]. The present study determined the CGs and their phylogenetic distribution in large-scale samples, including more than 4 thousand samples. This study revealed a very strong power of discrimination and polymorphic information, with a powerful average CMP of 3.118×10-11; this implies that the possibility of the exactly same genotype profile in two randomly chosen C. goeringii samples is less than 1 in 20 billion. Therefore, we believe that profiling the 12 SSR markers is an excellent forensically-applicable method to discriminate cultivars (or individuals) and analyze phylogenetic relationships.
The allele names of microsatellites are usually expressed as the relative repeat numbers of short sequence units in the given samples in the phylogenetic and pedigree construction or linkage analyses. This study named alleles based on the absolute values of repeat numbers, similar to that used in forensic genetics and criminal investigations [14]. Therefore, the C. goeringii SSR dataset from this study will enable comparative research by ensuring data compatibility among research groups; in addition it will serve as a standardized reference SSR database. For all examined cultivars, the most common SSR profiles (SG1) and the second most frequent profiles (CG2), with allele names indicated by repeat numbers are presented in Table S1.
The most frequent CGs (CG1s) were observed in 2664 samples from the 50 cultivars with 10 or more samples (n = 3,923). The mean of the obtained CG1 ratio for each cultivar showed a similar value of 70.1%. If we assume that only samples with CG1 are genuine to the corresponding cultivars, approximately 30% of the spring orchid cultivars in the Korean markets and farms may not be genuine.
Non-genuine sales of spring orchids usually involve cultivars showing similar horticultural phenotypes but different prices. The cultivar Hwanggeumso is subject to frequent non-genuine sales. The cultivar Hwanggeumso has a superior phenotype among the class of yellow flowers with a non-anthocyanin white lip, and its price has remained steady at approximately 5000 USD. The most-frequently observed non-genuine cultivar of Hwanggeumso is Granum. The price of Gwaneum, which has yellow flowers with a non-anthocyanin white lip, is approximately 1000 USD. In addition, Youngchoonso is frequently observed as a non-genuine cultivar of Hwanggeumso and Gwaneum. Youngchoonso belongs to the class of yellow flowers with non-anthocyanin white lips. However, its price is approximately 300 USD, which is much cheaper than that of Hwanggeumso and Gwaneum. In particular, two Japanese cultivars (Chanbo and Hobakjeon) are sold as Korean cultivars. These two Japanese cultivars are far cheaper than their Korean counterparts, such as Hwanggeumso, Gwaneum, Cheonsoo, and Cheonsa.
The frequencies of the second frequent CGs were generally low, below 15%; however, they were high in two cultivars Cheonhwangso (29.8%) and Hallasan (30.0%). In the cultivars of Cheonhwangso and Hallasan, the most common CG (CG1A) and the second most common CG (CG1B) showed similar profiles with high sibling probabilities. In addition, the horticultural phenotypes of both plants having these different CGs were so similar that they are indistinguishable. Therefore, these were a pair of genetically close sister cultivars. Orchid cultivars collected from nearby wild locations that exhibit similar phenotypes are traditionally called as sister cultivars. Nam et al determined the genetic kinship between four groups of closely related sister cultivars of C. goeringii [6]. If individual plants with CG1A or CG1B in Cheonhwangso and Hallasan were sister cultivars, it would be difficult to determine which plant was the original. This gives rise to the question of whether plants with both CG1A and CG1B should be considered genuine cultivars of Cheonhwangso or Hallasan.
The phylogenetic tree based on SSR genotypes divided the cultivars into three groups according to their country of origin. The 61 Korea-origin cultivars were largely separated from the five Japan-origin cultivars and the China-origin C. forestii cultivar in the phylogenetic tree. These results were consistent with those of a previous study [5]. However, the genetic distances among the C. goeringii cultivars seemed to be close; even C. forestii from China was not far from the cultivars of C. goeringii. In the PCoA, C. forestii was located between the Korean and Japanese cultivar groups but was closer to the Japanese group. Several Korean cultivars were located around a cluster of Japanese cultivars.
The complete nuclear genome sequence of C. goeringii is not available; however, sequences of the full chloroplast genome (cpDNA) of C. goeringii and a hybrid of C. goringii and C. sinense is available [17,18]. In addition, the recent RNA sequencing and transcriptomic analyses have suggested molecular mechanisms for phenotyping leaf color, floral patterning, and scent [19-22]. If genetic information on cpDNA variation and RNA expression is added to the dataset of SSR profile in future studies, a much more reliable cultivar discrimination in spring orchids can be performed.
5. Conclusions
We examined 61 Korean cultivars of C. goeringii by genotyping 12 SSR markers for forensic genetic discrimination. The established dataset for C. goeringii cultivar-specific SSR profiles could be used for ecogenetic studies and forensic authentication. This study revealed that almost 30% of C. goeringii in the market may not be genuine. Therefore, we suggest that genetic authentication should be introduced for the sale of expensive C. goeringii cultivars. In addition, we suggest the preparation of guidelines for the DNA deposition and profiling of SSR genotypes in newly registered C. goeringii cultivars. We believe that this study will help establish a genetic method for the forensic authentication of C. goeringii cultivars.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Combined genotypes of 12 SSRs observed in C. goeringii cultivars.
Author Contributions
Conceptualization, K.W.C.; methodology, D.E.N., and Y.D.; validation, D.E.N., Y.D., and S.G.K.; formal analysis, M.J.C., Y.D.K., and M.A.; writing—original draft preparation, K.W.C., and Y.D.; writing—review and editing, D.E.N, Y.D., and S.G.K.; project administration, K.W.C. All authors have read and agreed to the manuscript.
Funding
This study was supported by the grant of National Research Foundation in Korea (grant number 2022R1I1A2068995).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
“Not applicable.
Acknowledgments
We would like to thank all the persons who helped with the collection of samples and the Korea Orchid Registration Association for providing photographs of spring orchids.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- DuPuy, D.; Cribb, P.J. The genus Cymbidium. Richmond. Kew Publishing: UK, 2007. [Google Scholar]
- Lawler, L.J. Ethnobotany of Orchidaceae, p. 30-116. In: J. Arditti (ed.). Orchid Bioiogy: Reviews and perspectives. Comstock Publishing Associate. Cornell University Press, Ithaca, NY. 1994; Volume 3. [Google Scholar]
- Lee, H.J.; Park, H.R.; Lee, A.J.; Nam, D.E.; Lee, D.G.; Do, Y.; Chung, K.W. Genetic authentication of cultivars with flower-variant types using SSR markers in spring orchid, Cymbidium goeringii. Hortic Environ Biotechnol 2020, 61, 577–590. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, B.M.; Yu, S.O.; Ryu, K.H. Analysis of phenotypic and genetic polymorphism of self-pollinated seedings of Korean native Cymbidium goeringii. Kor J Hort Sci Technol 2004, 22, 486–490. [Google Scholar]
- Suetsugu, K. Autonomous self-pollination and insect visitors in partially and fully mycoheterotrophic species of Cymbidium (Orchidaceae). J Plant Res 2015, 128, 115–125. [Google Scholar] [CrossRef]
- Nam, D.E.; Yu, J.S.; Noh, S.W.; Lee, K.S.; Hwang, J.H.; Lee, D.G.; Chung, K.W. Genetic kinship and discrimination between the cultivars consisting a sister cultivar group in spring orchid Cymbidium goeringii. Hort Sci Technol 2021, 39, 106–121. [Google Scholar]
- Huang, J.L.; Zeng, C.X.; Li, H.T.; Yang, J.B. Isolation and characterization of 15 microsatellite markers from the spring orchid (Cymbidium goeringii) (Orchidaceae). Am J Bot 2011, 98, e76–e77. [Google Scholar] [CrossRef]
- Hyun, Y.S.; Kim, J.; Chung, K.W. Development of polymorphic microsatellite markers for Cymbidium goeringii (Orchidaeceae). Am J Bot 2012, 99, e193–e198. [Google Scholar] [CrossRef] [PubMed]
- Moe, K.T.; Zhao, W.; Song, H.-S.; Kim, Y.-H.; Chung, J.-W.; Cho, Y.-I.; Park, P.H.; Park, H.-S.; Chae, S.-C.; Park, Y.-J. Development of SSR markers to study diversity in the genus Cymbidium. Biochem Syst Ecol 2010, 38, 585–594. [Google Scholar] [CrossRef]
- Lee, D.G.; Koh, J.C.; Chung, K.W. Determination and application of simple sequence repeats (SSR) DNA ID for cultivars of Cymbidium goeringii. Kor J Hort Sci Technol 2012, 30, 78–285. [Google Scholar]
- Tang, M.; Zeng, C.X.; Bi, Y.F.; Yang, J.B. Microsatellite markers for the Chinese endangered and endemic orchid Cymbidium tortisepalum (Orchidaceae). Am J Bot 2012, 99, e11–e13. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jin, F.; Jin, L.; Jackson, A.; Huang, C.; Li, K.; Shu, X. Development of Cymbidium ensifolium genic-SSR markers and their utility in genetic diversity and population structure analysis in cymbidiums. BMC Genet 2014, 5, 124. [Google Scholar]
- Noh, S.W.; Park, J.-K.; Yu, J.S.; Nam, D.E.; Do, Y.; Chung, K.W. Genetic diversity and population structure of the spring orchid Cymbidium goeringii in Korean distant islands. Diversity 2020, 12, 486. [Google Scholar] [CrossRef]
- Bär, W.; Brinkmann, B.; Budowle, B.; Carracedo, A.; Gill, P.; Lincoln, P.; Mayr, W.; Olaisen, B. DNA recommendations-Further report of the DNA Commission of the ISFH regarding the use of short tandem repeat systems. Int J Legal Med 1997, 110, 175–176. [Google Scholar] [PubMed]
- Evett, I.W.; Weir, B.S. Interpreting DNA evidence: statistical genetics for forensic scientists., Sunderland, MA, Sinauer Associates, USA, 1998, Volume 244.
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Park, S.Y.; Lee, A.R.; Jang, S.G.; Im, D.E.; Jun, T.H.; Lee, J.; Chung, J.W.; Ham, T.H.; Kwon, S.W. Next-generation sequencing yields the complete chloroplast genome of C. goeringii acc. smg222 and phylogenetic analysis. Mitochondrial DNA B Resour 2018, 3, 215–216. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.I.; Lyu, J.I.; Lee, H.O.; Kim, J.B.; Kim, S.H. Complete chloroplast genome sequence of an orchid hybrid Cymbidium sinense (♀) × C. goeringii (♂). Mitochondrial DNA B Resour 2020, 5, 3802–3803. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhu, G.; Wang, Z.; Liu, H.; Xu, Q.; Huang, D.; Zhao, C. Integrated mRNA and microRNA transcriptome variations in the multi-tepal mutant provide insights into the floral patterning of the orchid Cymbidium goeringii. BMC Genomics 2017, 18, 367. [Google Scholar] [CrossRef] [PubMed]
- Ramya, M.; Park, P.H.; Chuang, Y.C.; Kwon, O.K.; An, H.R.; Park, P.M.; Baek, Y.S.; Kang, B.C.; Tsai, W.C.; Chen, H.H. RNA sequencing analysis of Cymbidium goeringii identifies floral scent biosynthesis related genes. BMC Plant Biol. 2019, 19, 337. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, S.W.; Lim, G.H.; Lyu, J.I.; Choi, H.I.; Jo, Y.D.; Kang, S.Y.; Kang, B.C.; Kim, J.B. Transcriptome analysis to identify candidate genes associated with the yellow-leaf phenotype of a Cymbidium mutant generated by γ-irradiation. PLoS One 2020, 5, e0228078. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Li, H.; Chen, X.; Zhang, Y.; Lu, L.; Li, S.; Tao, X.; Zhu, W.; Wang, J.; Ma, L. Insight into the molecular mechanisms of leaf coloration in Cymbidium ensifolium. Front Genet 2022, 13, 923082. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Cultivars of Cymbidium goeringii. (a) Flower-variant types. (b) Leaf-variant types.

Figure 2.
Frequencies of combined genotypes of 12 SSR markers in 50 C. goeringii cultivars with samples of 10 or more (CG1: most frequent combined genotypes, CG2: second frequent combined genotypes). The red dotted line represents the mean frequency of CG1.
Figure 2.
Frequencies of combined genotypes of 12 SSR markers in 50 C. goeringii cultivars with samples of 10 or more (CG1: most frequent combined genotypes, CG2: second frequent combined genotypes). The red dotted line represents the mean frequency of CG1.
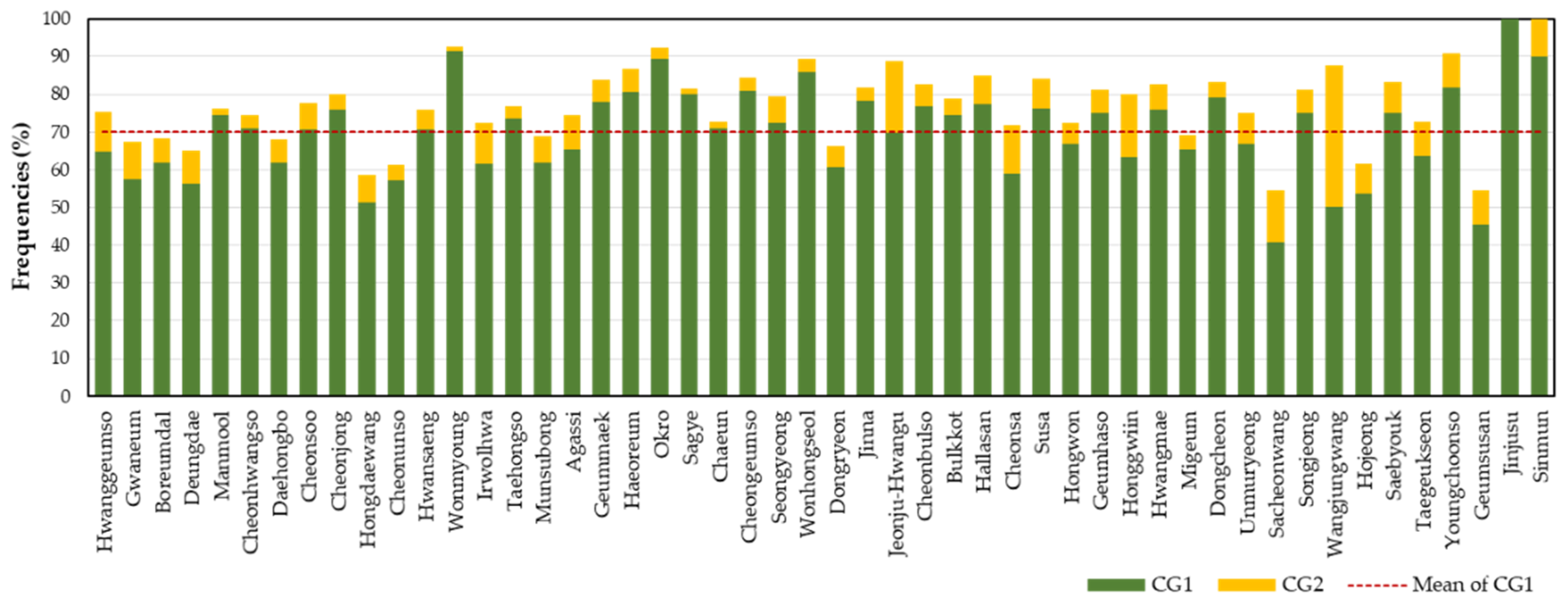
Figure 3.
Phylogenetic tree of 66 cultivars in C. goeringii based on the genetic distance measured from genotypes of 12 SSR loci using the unweighted pair group method with arithmetic average (UPGMA) as the cluster method. The five cultivars within a yellow box are Japan-origin C. goeringii, while the Hwanguhajeong within a blue box is China-origin C. forestii.
Figure 3.
Phylogenetic tree of 66 cultivars in C. goeringii based on the genetic distance measured from genotypes of 12 SSR loci using the unweighted pair group method with arithmetic average (UPGMA) as the cluster method. The five cultivars within a yellow box are Japan-origin C. goeringii, while the Hwanguhajeong within a blue box is China-origin C. forestii.
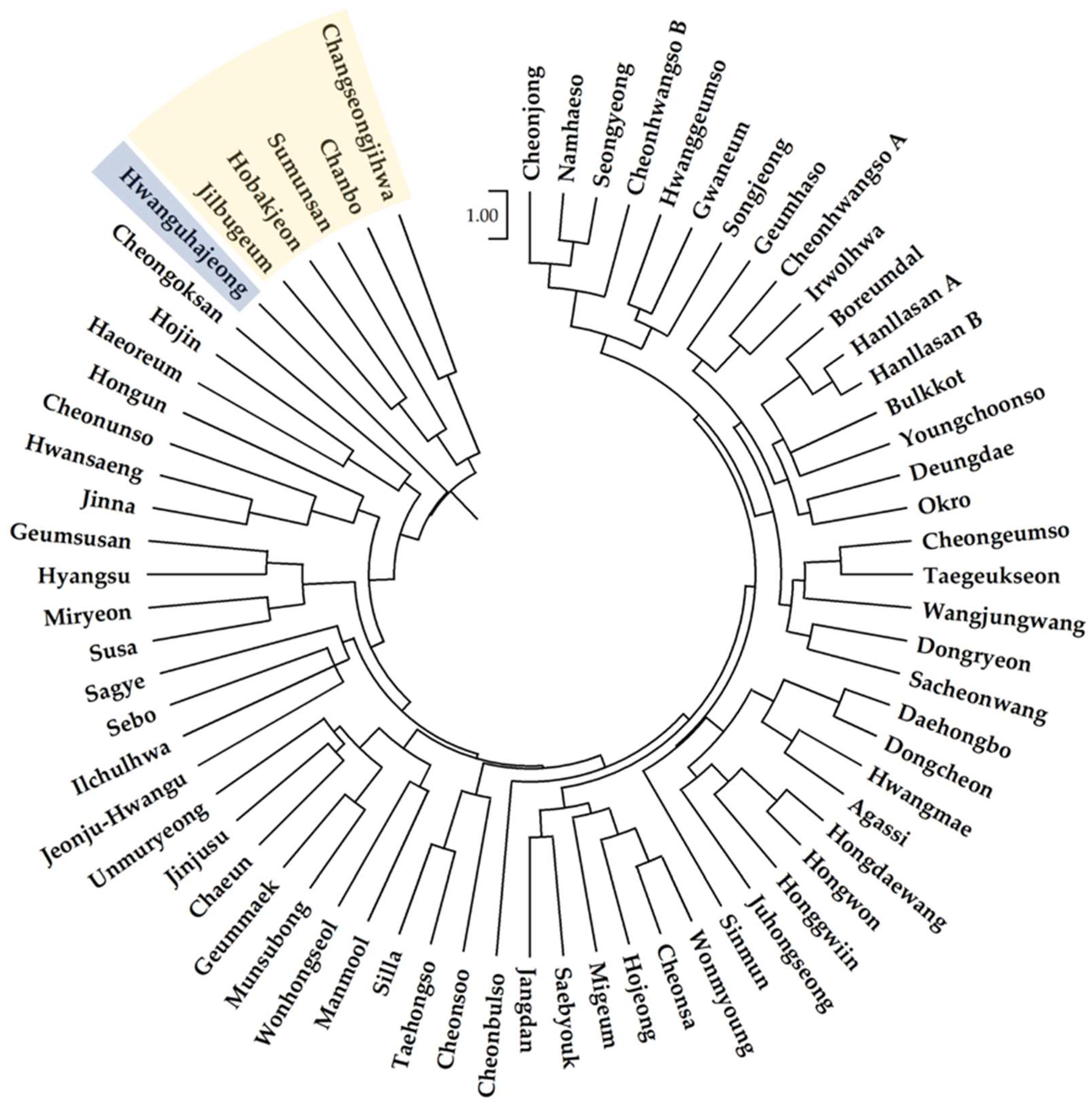
Figure 4.
Principal coordinate analyses (PCoA) of 67 cultivars of C. goeringii or C. forestii. The five cultivars within the yellow box are Japan-origin C. goeringii, while Hwanguhajeong within the blue box is a China-origin C. forestii. The cumulative variability captured by the three principal component axes is 25.1 % (PC1: 11.8 %; PC2: 6.7 %; PC3: 6.5 %).
Figure 4.
Principal coordinate analyses (PCoA) of 67 cultivars of C. goeringii or C. forestii. The five cultivars within the yellow box are Japan-origin C. goeringii, while Hwanguhajeong within the blue box is a China-origin C. forestii. The cumulative variability captured by the three principal component axes is 25.1 % (PC1: 11.8 %; PC2: 6.7 %; PC3: 6.5 %).
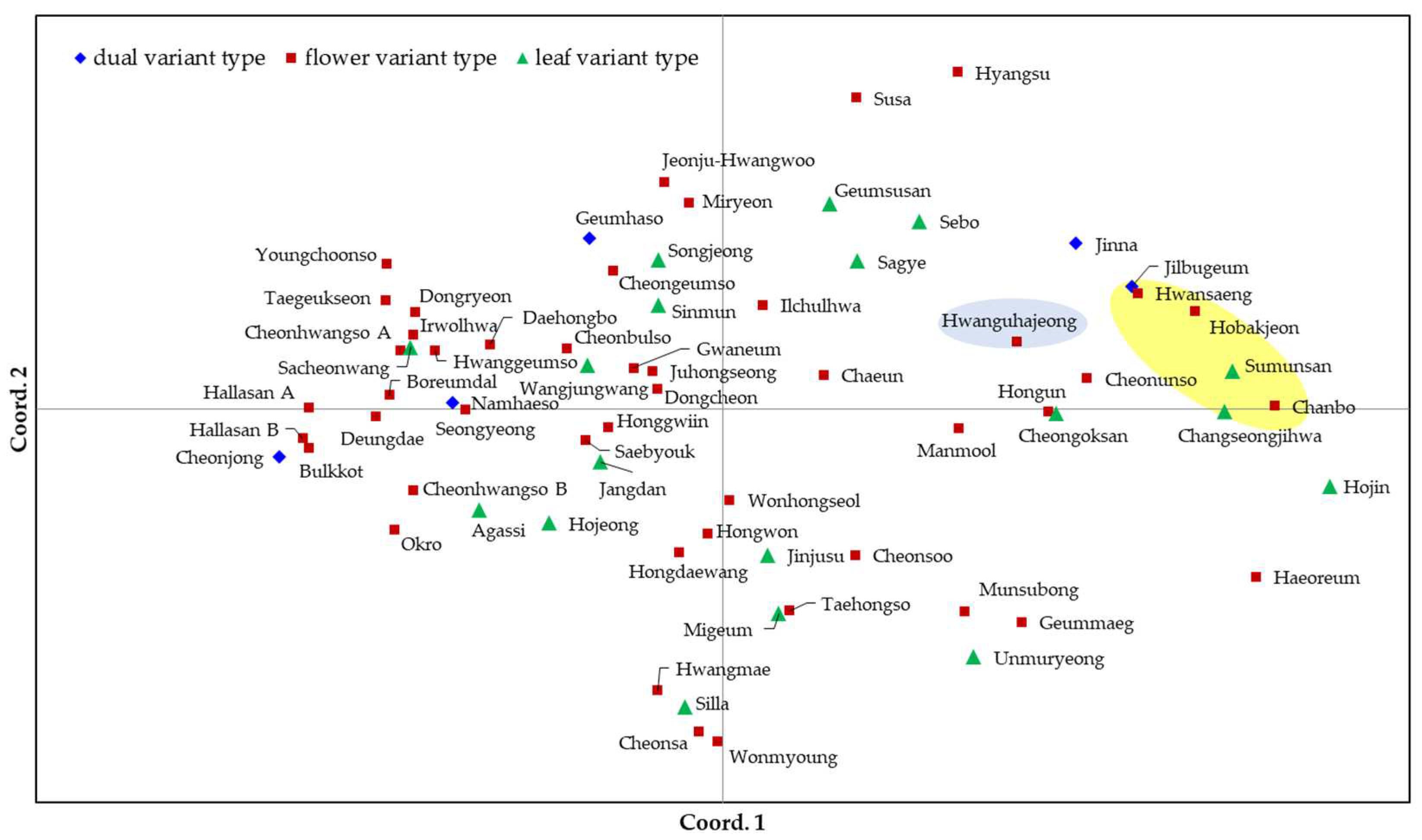

Table 1.
List of C. goeringii cultivars.
| Cultivar name | Type1 | Main phenotype | Sample number |
|---|---|---|---|
| Cultivars originated from Korea | |||
| Hwanggeumso | F | Yellow flower with non-anthocyanin lip | 468 |
| Gwaneum | F | Yellow flower with non-anthocyanin lip | 301 |
| Boreumdal | F | Yellow flower with non-anthocyanin lip | 250 |
| Deungdae | F | Orange-yellow flower with non-anthocyanin lip | 194 |
| Manmool | F | Orange-yellow flower | 184 |
| Cheonhwangso | F | Yellow flower with non-anthocyanin lip | 161 |
| Daehongbo | F | Red flower | 144 |
| Cheonsoo | F | Orange-yellow flower with non-anthocyanin lip | 143 |
| Cheonjong | D | Short leaf and round flower both with yellow variegation | 125 |
| Hongdaewang | F | Red flower | 109 |
| Cheonunso | F | Flower with mixed colors and non-anthocyanin lip | 98 |
| Hwansaeng | F | Yellow flower with non-anthocyanin lip | 95 |
| Wonmyoung | F | Round yellow flower | 93 |
| Irwolhwa | F | Small round flower with non-anthocyanin lip | 91 |
| Taehongso | F | Orange-yellow flower with non-anthocyanin lip | 91 |
| Munsubong | F | Flower with mixed colors | 84 |
| Agassi | L | Leaf with narrow yellow stripes | 78 |
| Geummaek | F | Small round yellow flower | 68 |
| Haeoreum | F | Red flower | 67 |
| Okro | F | Flower with mixed colors and orange bordering | 66 |
| Sagye | L | Short leaf with yellow variegation | 65 |
| Chaeun | F | Orange-yellow flower with non-anthocyanin lip | 62 |
| Cheongeumso | F | Yellow flower with non-anthocyanin lip | 58 |
| Seongyeong | F | Leaf with yellow stripes and flower with non-anthocyanin lip | 58 |
| Wonhongseol | F | Purple small round flower | 57 |
| Dongryeon | F | Leaf with short yellow stripes and small round flower | 56 |
| Jinna | D | Leaf and flower with yellow variegation | 55 |
| Jeonju-Hwangu | F | Yellow flower | 53 |
| Cheonbulso | F | Red flower with non-anthocyanin lip | 52 |
| Bulkkot | F | Red flower | 47 |
| Hallasan | F | Small round flower | 40 |
| Cheonsa | F | Orange-yellow flower with non-anthocyanin lip | 39 |
| Susa | F | Red flower | 38 |
| Hongwon | F | Round red flower | 36 |
| Geumhaso | D | Leaf and flower with yellow stripe | 32 |
| Honggwiin | F | Orange-yellow flower with non-anthocyanin lip | 30 |
| Hwangmae | F | Pale yellow round flower | 29 |
| Migeum | L | Leaf with short multiple yellow stripes and spots | 26 |
| Dongcheon | F | Orange-yellow flower with non-anthocyanin lip | 24 |
| Unmuryeong | L | Overall yellowish leaf | 24 |
| Sacheonwang | L | Leaf with yellow stripes | 22 |
| Songjeong | L | Leaf with narrow yellow stripes | 16 |
| Wangjungwang | L | Short leaf with yellow stripes | 16 |
| Hojeong | L | Short leaf with yellow stripes | 13 |
| Saebyeok | F | Red flower with non-anthocyanin lip | 12 |
| Taegeukseon | F | Flower with yellow and green mixed colors | 11 |
| Youngchoonso | F | Yellow flower with non-anthocyanin lip | 11 |
| Geumsusan | L | Overall yellowish leaf | 11 |
| Jinjusu | L | Leaf with yellow stripes | 10 |
| Sinmun | L | Leaf with yellow stripes | 10 |
| Sebo | L | Leaf with yellow stripes | 6 |
| Silla | L | Short leaf with yellow bordering | 5 |
| Ilchulhwa | F | Small round flower | 4 |
| Cheongoksan | L | Leaf with multiple yellow spots | 3 |
| Hongun | F | Red flower | 3 |
| Jangdan | F | Red flower | 3 |
| Miryeon | F | Round flower with pale anthocyanin lip | 3 |
| Hojin | L | Leaf with yellow variegation | 2 |
| Hyangsu | F | Green flower with yellow bordering | 2 |
| Namhaeso | D | Leaf and flower with white bordering | 2 |
| Juhongseong | F | Flower with mixed colors and orange bordering | 1 |
| Cultivars originated from Japan | |||
| Chanbo | F | Yellow flower with non-anthocyanin lip | 40 |
| Jilbugeum | D | Leaf and flower with multiple yellow stripes and spots | 14 |
| Changseongjihwa | L | Leaf showing leopard print | 13 |
| Hobakjeon | F | Orange-yellow flower with non-anthocyanin lip | 11 |
| Sumunsan | L | Leaf with yellowish variegation | 8 |
| Cultivar originated from China | |||
| Hwanguhajeong2 | F | Floral fragrance | 5 |
| Total | - | - | 4048 |
1 F: flower-variant type, L: leaf-variant type, D: dual-variant type. 2 It belongs to C. forestii.
Table 2.
Combined genotypes of 12 SSRs observed in C. goeringii cultivars.
| Cultivars | CG1 % | Genotype of SSR markers1 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CG415 | CG649 | CG709 | CG722 | CG787 | CG1023 | CG1028 | CG1085 | CG1210 | CG1281 | CG1320 | CG1400 | ||
| Hwanggeumso | 64.7 | 11-11 | 15-30.1 | 17-18 | 12-20 | 24-29 | 13-13 | 15-17 | 14-15 | 29-29 | 16-16 | 10-13 | 13.1-20 |
| Gwaneum | 57.4 | 11-13 | 21-30.1 | 18-18 | 12-20 | 24-24 | 13-13 | 15-15 | 14-19 | 15-29 | 16-16 | 10-13 | 13.1-20 |
| Boreumdal | 62.0 | 11-20 | 15-15 | 17-25 | 17-17 | 24-24 | 13-13 | 6-7 | 14-14 | 6-6 | 16-16 | 10-10 | 15-20 |
| Deungdae | 56.2 | 11-15 | 16-16 | 17-17 | 12-12 | 24-24 | 13-13 | 7-19 | 14-38 | 15-29 | 16-16 | 12-12 | 20-21 |
| Manmool | 74.5 | 13-13 | 15-29.1 | 23-23 | 12-17 | 18-18 | 13-20 | 15-17 | 19-19 | 17-17 | 9-9 | 12-12 | 20-20 |
| Cheonhwangso | 41.0 (A) | 11-19 | 14-15 | 17-34 | 12-12 | 24-24 | 13-13 | 17-17 | 15-15 | 12-12 | 16-16 | 12-12 | 20-20 |
| 29.8 (B) | 11-11 | 15-29.1 | 17-34 | 12-20 | 24-24 | 13-25 | 17-17 | 16-16 | 17-17 | 16-16 | 7-12 | 13.1-20 | |
| Daehongbo | 61.8 | 13-22 | 15-15 | 17-18 | 20-20 | 24-24 | 13-13 | 17-17 | Null | 15-15 | 16-16 | 7-7 | 19-19 |
| Cheonsoo | 70.6 | 11-13 | 14-15 | 18-18 | 17-20 | 24-24 | 20-20 | 17-17 | 14-14 | 15-15 | 10-16 | 7-12 | 20-20 |
| Cheonjong | 76.0 | 11-11 | 15-19 | 17-17 | 12-20 | 24-24 | 13-13 | 6-30 | 14-14 | 15-15 | 9-9 | 10-15 | 13.1-20 |
| Hongdaewang | 51.4 | 13-13 | 15-33.1 | 17-22 | 12-20 | 24-24 | 19-19 | 30-30 | 14-14 | 6-15 | 16-16 | 6-7 | 19-20 |
| Cheonunso | 57.1 | 13-13 | 15-15 | 18-18 | 30-30 | Null | 26-26 | 17-17 | 14-14 | 15-15 | 16-16 | 7-7 | 10.1-13.1 |
| Hwansaeng | 70.5 | 13-13 | 26.1-30.1 | 18-18 | 11-11 | Null | 13-13 | 6-12 | 14-15 | 17-29 | 16-16 | 7-9 | 20-20 |
| Wonmyoung | 91.4 | 11-13 | 15-15 | 22-22 | 11-12 | 24-24 | 13-20 | 17-17 | 15-15 | 15-15 | 16-16 | 12-12 | 13.1-13.1 |
| Irwolhwa | 61.5 | 11-19 | 15-15 | 17-34 | 12-17 | 22-24 | 13-13 | 15-15 | 14-14 | 15-33 | 9-9 | 7-7 | 19-19 |
| Taehongso | 73.6 | 11-13 | 15-20 | 17-17 | 12-20 | 22-22 | 20-20 | 6-6 | 14-14 | 15-28 | 9-16 | 12-12 | 15-15 |
| Munsubong | 61.9 | 13-13 | 25.1-29.1 | 17-22 | 12-12 | 22-24 | 20-20 | 15-17 | 14-19 | 12-15 | 16-16 | 7-12 | 13.1-18 |
| Agassi | 65.4 | 13-22 | 15-15 | 17-17 | 12-20 | 24-24 | 13-20 | 15-17 | 16-16 | 6-15 | 10-17 | 7-12 | 19-19 |
| Geummaek | 77.9 | 13-13 | 15-29.1 | 17-22 | 11-12 | 22-25 | 13-20 | 6-12 | 19-19 | 12-15 | 10-16 | 10-12 | 19-20 |
| Haeoreum | 80.6 | 11-13 | 14-14 | 25-25 | 11-11 | 25-25 | 20-20 | 17-17 | 14-14 | 17-17 | 9-9 | 7-7 | 13.1-13.1 |
| Okro | 89.4 | 11-13 | 15-20 | 17-17 | 12-22 | 24-24 | 13-13 | 7-17 | 14-15 | 12-15 | 16-16 | 6-12 | 13.1-13.1 |
| Sagye | 80.0 | 13-15 | 15-26.1 | 18-26 | 12-12 | 16-22 | 13-25 | 17-30 | 14-15 | 6-17 | 9-16 | 7-12 | 15-15 |
| Chaeun | 71.0 | 13-13 | 29.1-29.1 | 16-16 | 12-12 | 22-24 | 13-13 | 15-15 | 14-14 | 17-17 | 9-16 | 7-13 | 13.1-20 |
| Cheongeumso | 81.0 | 13-19 | 15-29.1 | 17-25 | 12-17 | 24-25 | 13-13 | 17-29 | 14-24 | 15-15 | 16-16 | 7-7 | 13.1-19 |
| Seongyeong | 72.4 | 11-11 | 15-29.1 | 13-17 | 12-20 | 17-24 | 13-13 | 17-17 | 14-14 | 15-29 | 9-16 | 6-6 | 20-20 |
| Wonhongseol | 86.0 | 13-13 | 16-34.1 | 13-17 | 12-17 | 24-24 | 13-20 | 7-17 | 14-19 | 11-17 | 9-9 | 7-7 | 13.1-20 |
| Dongryeon | 60.7 | 15-15 | 15-15 | 17-25 | 12-20 | 22-24 | 13-13 | 15-15 | 14-14 | 15-15 | 16-16 | 12-12 | 20-20 |
| Jinna | 78.2 | 10-13 | 14-29.1 | 18-18 | 11-17 | Null | 13-13 | 7-7 | 14-14 | 15-15 | 16-16 | 7-7 | 19-19 |
| Jeonju-Hwangu | 69.8 | 13-15 | 15-15 | 34-34 | 12-20 | Null | 13-13 | 7-24 | 14-14 | 12-12 | 16-16 | 7-7 | 15-19 |
| Cheonbulso | 76.9 | 11-19 | 23-26.1 | 25-34 | 20-20 | 24-24 | 13-13 | 30-30 | 14-14 | 15-15 | 16-16 | 12-12 | 20-20 |
| Bulkkot | 74.5 | 10-11 | 15-15 | 17-17 | 12-12 | 24-24 | 13-13 | 17-17 | Null | 12-13 | 10-16 | 12-12 | 19-20 |
| Hallasan | 47.5 (A) | 11-15 | 15-15 | 17-17 | 17-17 | 24-24 | 13-13 | 12-12 | 14-14 | 17-17 | 10-16 | 10-10 | 13.1-13.1 |
| 30.0 (B) | 11-11 | 15-15 | 17-17 | 17-17 | 24-24 | 13-13 | 17-17 | 14-14 | 15-17 | 16-16 | 10-10 | 13.1-13.1 | |
| Cheonsa | 59.0 | 11-13 | 15-25.1 | 17-18 | 11-12 | 24-24 | 13-20 | 17-17 | 14-14 | 6-15 | 9-16 | 7-12 | 19-20 |
| Susa | 76.3 | 13-22 | 14-20 | 13-22 | 12-17 | 15-22 | 13-13 | 17-17 | 14-15 | 12-15 | 10-16 | 7-7 | 13.1-19 |
| Hongwon | 66.7 | 13-13 | 15-15 | 19-22 | 12-20 | 24-24 | 19-19 | 30-30 | 14-14 | 15-29 | 16-16 | 7-7 | 19-19 |
| Geumhaso | 75.0 | 11-19 | 14-34.1 | 18-26 | 12-17 | 24-24 | 13-13 | 6-17 | 14-19 | 15-17 | 16-16 | 7-13 | 13.1-20 |
| Honggwiin | 63.3 | 13-13 | 15-34.1 | 22-22 | 12-20 | 24-24 | 13-13 | 7-12 | 14-14 | 17-17 | 16-16 | 10-10 | 13.1-13.1 |
| Hwangmae | 75.9 | 13-13 | 15-15 | 17-17 | 11-22 | 24-24 | 13-20 | 17-30 | 13-14 | 15-15 | 16-16 | 7-7 | 20-23.1 |
| Migeum | 65.4 | 11-19 | 15-15 | 22-25 | 11-11 | 19-24 | 13-20 | 6-6 | 14-14 | 17-17 | 10-10 | 7-7 | 19-20 |
| Dongcheon | 79.1 | 13-13 | 15-15 | 18-18 | 20-20 | 24-24 | 13-13 | 12-17 | 14-14 | 15-15 | 16-16 | 7-7 | 13.1-13.1 |
| Unmuryeong | 66.7 | 13-13 | 15-29.1 | 16-17 | 11-11 | 17-24 | 13-20 | 12-12 | 14-14 | 12-12 | 16-16 | 10-10 | 19-19 |
| Sacheonwang | 40.9 | 15-15 | 15-15 | 13-13 | 12-12 | 24-24 | 13-13 | 6-7 | 14-14 | 15-15 | 16-16 | 7-13 | 20-20 |
| Songjeong | 75.0 | 11-12 | 19-30.1 | 17-18 | 12-20 | 17-18 | 13-13 | 17-17 | 24-25 | 29-30 | 16-16 | 13-13 | 13.1-20 |
| Wangjungwang | 56.3 | 13-15 | 15-15 | 18-25 | 12-22 | 24-24 | 13-29 | 7-17 | 15-19 | 15-17 | 17-17 | 10-13 | 13.1-20 |
| Hojeong | 53.8 | 11-11 | 15-23 | 13-22 | 11-12 | 24-24 | 13-13 | 16-16 | 14-22 | 15-15 | 9-9 | 12-13 | 13.1-20 |
| Saebyeok | 75.0 | 11-15 | 15-32.1 | 17-18 | 11-22 | 22-24 | 13-13 | 30-30 | 14-14 | 15-15 | 16-16 | 12-12 | 19-20 |
| Taegeukseon | 63.6 | 13-15 | 15-15 | 17-25 | 12-17 | 24-24 | 13-13 | 17-17 | 14-14 | 15-15 | 16-16 | 6-7 | 19-20 |
| Youngchoonso | 81.8 | 15-15 | 14-15 | 17-17 | 12-17 | 24-24 | 13-13 | 17-17 | 15-15 | 15-17 | 8-8 | 7-12 | 13.1-13.1 |
| Geumsusan | 45.5 | 13-22 | 14-14 | 17-17 | 17-20 | 25-25 | 13-13 | 17-17 | 15-15 | 15-17 | 9-9 | 7-7 | 19-19 |
| Jinjusu | 100.0 | 12-13 | 15-29.1 | 12-17 | 11-12 | 22-24 | 13-13 | 15-17 | 14-16 | 17-27 | 9-16 | 7-10 | 19-20 |
| Sinmun | 90.0 | 13-15 | 15-29.1 | 18-18 | 17-23 | 24-24 | 13-13 | 6-17 | 14-14 | 15-17 | 16-16 | 7-7 | 17-20 |
1 Allele were named by the repeat numbers of SSR motif according to the guideline of the ISFH. Abbreviations: CG: combined genotype, CMP: combined matching probability, SSR: simple sequence repeats.
Table 3.
This is a table. Tables should be placed in the main text near to the first time they are cited.
Table 3.
This is a table. Tables should be placed in the main text near to the first time they are cited.
| Cultivar names mentioned by suppliers | Genetically matching cultivars1 |
|---|---|
| Hwanggeumso (n=468) | Gwaneum (50), Youngchoonso (7), Namhaeso (5), Cheonhwangso (1), Geumhaso (1), Chanbo (3)2, Hobakjeon (1)2 |
| Gwaneum (n=301) | Youngchoonso (30), Cheongeumso (2), Geumhaso (1), Hobakjeon (5)2, Chanbo (1)2 |
| Boreumdal (n=250) | Saebyeok (1) |
| Cheonhwangso (n=161) | Gwaneum (6), Youngchoonso (1) |
| Daehongbo (n=144) | Wonmyoung (1) |
| Cheonsoo (n=143) | Chanbo (10)2 |
| Cheonjong (n=125) | Sebo (3), Hwansaeng (1) |
| Hongdaewang (n=109) | Jangdan (8) |
| Cheonunso (n=98) | Taegeukseon (4) |
| Hwansaeng (n=95) | Deungdae (1) |
| Wonmyoung (n=93) | Hwanguhajeong (1)3 |
| Irwolhwa (n=91) | Miryeon (1), Cheonjong (1) |
| Munsubong (n=84) | Hyangsu (6) |
| Agassi (n=78) | Jinjusu (7), Sinmun (3), Sebo (1) |
| Geummaek(n=68) | Ilchulhwa (4) |
| Chaeun (n=62) | Cheonsa (1) |
| Cheongeumso (n=58) | Wonhongseol (1) |
| Cheonbulso (n=52) | Hwanggeumso (1), Sacheonwang (1) |
| Bulkkot (n=47) | Juhongseong (2) |
| Cheonsa (n=39) | Chaeun (1), Chanbo (5)2 |
| Hongwon (n=36) | Hongun (1) |
| Sacheonwang (n=22) | Sinmun (3) |
| Songjeong (n=16) | Sebo (1) |
| Taegeukseon (n=11) | Songjeong (1) |
1 Numbers in parentheses are observed numbers. 2 Japan-origin cultivar. 3China-origin cultivar of C. forestii.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



