
Submitted:
18 July 2023
Posted:
19 July 2023
You are already at the latest version
Abstract
Two isolates of Acinetobacter were isolated from the soil samples of motorbike repairing workshop floors in Vietnam, where motorcycle/scooter repairs generate various types of hazardous waste, including mainly used waste oil and fluids. Two isolates were Gram-stain-negative, coccobacilli-shaped, aerobic, non-motile, non-hemolytic, oxidase-negative, and catalase-positive, designated strains VNH17T and VNK23T. Growth of strains was inhibited by NaCl concentrations above 3% and grew at an optimal temperature of 20-35°C. Phylogenetic analysis based on 16S rRNA gene sequences revealed that two isolated strains formed a lineage within the family Moraxellaceae and clustered as members of the genus Acinetobacter. The predominant respiratory quinones of VNH17T and VNK23T are ubiquinone 9 and ubiquinone 8, and the polar lipids are mainly composed of diphosphatidyl glycerol, phosphatidylglycerol, and phosphatidylethanolamine. The major cellular fatty acids are C18:1ω9c, C16:0 and summed feature 3 (C16:1ω7c and/or C16 : 1ω6c). The DNA G+C content of VNH17T and VNK23T was 42.07% and 41.75%, respectively. In addition, the average nucleotide identity and in silico DNA-DNA hybridization relatedness values between VNH17T and VNK23T as well as between they and phylogenetically related type strains were below the threshold values used for species delineation. Based on genomic, chemotaxonomic, phenotypic and phylogenomic analyses, the isolated strains represent novel species in the genus Acinetobacter, for which the names Acinetobacter thutiue sp. nov. (type strain VNH17T = KACC 23003 = CCTCC AB 2023063), Acinetobacter kontum sp. nov. (type strain VNK23T = KACC 23002 = CCTCC AB 2023064) are proposed.
Keywords:
Acinetobacter thutiue
; Acinetobacter kontum
; oil-contaminated soil
; novel species
; Morazellaceae
1. Introduction
The genus Acinetobacter is a member of the family Moraxellaceae, first proposed by Beijerinck in 1910 [1] as Micrococcus calcoaceticus, before becoming known as Acinetobacter by Brisou and Prévot in 1954 [2]. At the time of writing, 108 different species have been assigned to the genus Acinetobacter, of which 82 with validly published and correct names have been listed (https://lpsn.dsmz.de/genus/cellulomonas; date of access: 14 July 2023) [3]. Members of this genus are Gram-negative coccobacilli, non-motile, non-spore-forming, aerobic, oxidase-negative, catalase-positive, and grow well on simple media. The genus Acinetobacter has been isolated from different sources such as a variety of animal food products like milk and meat [4-6]. Some Acinetobacter species have been isolated from clinical sources [7,8]; floral nectar [9]; others members of this genus distributed widely in soil and water [10-12]. Due to the possess versatile metabolic capabilities and degrade of various compounds, such as long-chain dicarboxylic acids and aromatics therefore some members of Acinetobacter can expand well in extreme environmental conditions such as organic solvent, and oil-contaminated soil [13-16]. In this study, we aimed to characterize and delineate the taxonomic position of two members VNH17T and VNK23T, isolated from oil-contaminated soils in Vietnam, on the basis of which they are considered to represent two novel species within the genus Acinetobacter.
2. Materials and Methods
2.1. Isolation and ecology
During a study of role verification of uncultured oil-degrading bacteria in oil-contaminated soil, a number of bacterial strains were isolated from soil samples of motorbike repairing workshop floors in Vietnam (Figure S1). Among them, two strains designated VNH17T and VNK23T were isolated from the motorbike repairing workshop at Phu Vang, Thua Thien Hue, Vietnam (17°04'16.3"N 106°59'28.4"E) and Dak Bla, Kon Tum, Vietnam (14°20'43.7"N 108°04'48.9"E), respectively by using a modified culture method with a six-transwell plate (Corning Inc., Corning, NY, US) on MSM-oil medium. The MSM-oil medium consisted of (per litre distilled water): 2.42 g KH2PO4, 5.60 g K2HPO4, 2 g (NH4)2SO4, 0.3 g MgSO4.7H2O, 0.04 g CaCl2.2H2O, 1 ml trace elements solution, and 2ml crude oil. The trace elements solution comprised (per litre distilled water): 4.5 g MnSO4.7H2O, 0.1 g CuSO4.5H2O, 0.1g FeSO4, pH adjusted to 7.0 [17]. In detail, a soil sample (3 g) was kept on the bottom of the transwell plate and 3 mL of MSM-oil was added, and then 100 µl of the soil suspension was added to the insert. The transwell plate was incubated in a shaker at 130 rpm at 28°C for 4 weeks. After incubation, 100μL of each dilution was spread on an MSM-oil agar plate. Colonies were selected and streaked separately on MSM-oil agar plates until pure colonies were grown; the strains were then subcultured on R2A and stored at −70°C in R2A broth supplemented with 20 % (v/v) glycerol. Acinetobacter junii KACC12228, Acinetobacter parvus KACC 12455, Acinetobacter tandoii KACC 12229, Acinetobacter indicus KCTC 42000, Acinetobacter bereziniae KCTC 42001 and Acinetobacter venetianus DSM 23050 were obtained from the Korean Agricultural Culture Collection (KACC), Korean Collection for Type Cultures (KCTC), and German Collection of Microorganisms (DSM) they were used as the reference organisms during this study.
2.2. Physiology and Chemotaxonomy
The colony morphology of strains VNH17T and VNK23T were observed after cultivation on R2A agar plates at 35°C for 2 days. Cell morphology grown on R2A agar for 2 days at 30°C was examined using light microscopy (BX50; Olympus) and transmission electron microscopy (Bio-TEM, Hitachi, H-7650). Gram staining was performed using Hucker’s method [18]. Endospore formation was investigated by staining with malachite green as described by Schaeffer and Fulton [19]. Production of hydrogen sulfide and indole and motility was assayed on sulfide indole motility (SIM) medium (CM0435; Oxoid) after 48-72h at 30°C and then the Kovacs’ reagent was used to check indole production [18]. Oxidase activity was assayed using 1 % (w/v) tetramethyl-p-phenylenediamine. Catalase activity was determined based on the production of bubbles after mixing a pellet of a fresh culture with a drop of 3 % (v/v) hydrogen peroxide (H2O2). Anaerobic growth was assayed using a BBL GasPak anaerobic jar with a dry anaerobic indicator strip (BD Biosciences) to indicate the presence of oxygen in the chamber. The test of growth on different agar media and temperature-dependent growth was performed by suspending fresh biomass in 0.9% (w/v) sodium chloride (NaCl) and then the cell suspension was spotted on agar media and the growth was monitored after 24, 48, and 72 hours of incubation. For the test of growth on different media, conventional media such as R2A agar (MB Cell), tryptone soya agar (TSA; MB Cell), Luria Bertani (LB) agar (Oxoid), nutrient agar (NA; Oxoid), Mueller-Hinton agar (MB Cell) and MacConkey agar (MB Cell) were used. Temperature-dependent growth was determined at different temperatures (4, 10, 15, 20, 25, 28, 30, 32, 35, 37, 39, 41, 43, and 45°C) on R2A. Growth physiology at different pH values was determined at 30°C in R2A broth adjusted to pH 3–12 (increments of 0.5 pH units) using citrate/NaH2PO4 buffer (for pH 3.0–5.5), Sorensen’s phosphate buffer (for pH 6.5–8), Tris buffer (for pH 8.5–9), carbonate buffer (for pH 9.5–10.0) and 5 M NaOH (for pH 10.5–11). NaCl tolerance tests at different NaCl concentrations were assayed in R2A supplemented with NaCl concentrations ranging from 0 to 7.0 % (0.5 % intervals) after 5 days of incubation at 30°C. Cellular growth at various pH values and salinity levels was monitored by measuring OD600 with a spectrophotometer (CARY 300, UV–Vis spectrophotometer, Varian). Hydrolysis of various substrates including hydrolyses of starch and casein was assayed as described by Tindall et al. [20], Tween 80 was assessed according to the method of Smibert and Krieg [21], the Methyl Red and Voges-Proskauer (MR-VP) test was assayed with MR-VP broth (Vaughn et al. 1939) and ADNase test was performed on DNase agar (CM0321; Oxoid), and the result was checked after 3 days of incubation at 30 °C by flooding the plates with 1 M HCl. Nitrate reduction, indole production, hydrolysis of gelatin, urease, aesculin, and other physiological and biochemical tests were tested with API 20NE strips, API 20E, and API ID 32 GN (bioMérieux) to evaluate basic chemical test and carbon source utilization. The activities of various enzymes and acid formation from sugar were further determined using API ZYM test strips (bioMérieux). The commercial kits API 20NE, API 20E, API ID 32 GN, and API ZYM were performed according to the manufacturer’s instructions. Susceptibilities of antibiotics were assayed as described by Dahal and Kim (2016b) [22]. To ascertain the presence of hemolytic activity, VNH17T, VNK23T, and reference strains were streaked on R2A agar supplemented with 5% (v/v) defibrinated sheep blood followed by incubation at 30°C for 5 days.
For the determination of respiratory quinones and polar lipids, freeze-dried cells of strains VNH17T and VNK23T that were grown on TSB plates were harvested after 2 days of growth were used. The respiratory quinones were extracted with methanol and petroleum ether from 70 mg of freeze-dried cells, the quinone purification was performed according to the method of Minnikin et al. [23] and the purified quinones were identified by a reversed-phase HPLC system [solvent MeOH/isopropanol (7: 5, v/v), flow rate 1.0 ml min−1, wavelength 270 nm]. Polar lipids were extracted according to the protocol of Minnikin et al. [23] and separated by two-dimensional TLC (Merck silica gel 60; 10×10 cm), and then identified by spraying with appropriate detection reagents including 5 % (w/v) ethanolic molybdatophosphoric acid (Sigma-Aldrich) to detect of total lipids profiles, amino lipids were stained with 0.4 % (w/v) solution of ninhydrin (Sigma Life Science) in butanol, phospholipids were detected with Zinzadze reagent (molybdenum blue spray reagent, 1.3 %; Sigma Life Science) and glycolipids were detected with α-naphthol reagent (0.5 %, w/v).
For the profiling of cellular fatty acids, cells of strains VNH17T, VNK23T, and reference strains were grown on TSB agar and they were harvested after 2 days of growth. Cellular fatty acids were extracted, saponified, and methylated following the protocol of the Sherlock Microbial Identification System version 6.3 (MIDI), analyzed by gas chromatography, and identified using the TSBA6 database of the Microbial Identification System [24].
2.3. Phylogenetic analysis based on 16S rRNA and rpoB genes
The 16S rRNA gene sequences of strains VNH17T and VNK23T were obtained using PCR with primers 27F and 1492R described by Frank et al. [25] and sequenced. Then, the 16S rRNA gene sequences were posted with the EzBioCloud database (www.ezbiocloud.net/eztaxon, last accessed 10 July 2023) [26] to find the closest phylogenetic neighbors. The 16S rRNA gene sequences of reference strains were retrieved from their depositions in the NCBI GenBank database (www.ncbi.nlm.nih.gov/, last accessed 10 July 2023). Multiple alignments of sequence data were made using the SILVA aligner “www.arb-silva.de/aligner/”. Three major phylogenetic trees (neighbor-joining (NJ), maximum-likelihood (ML), and maxi-mum-parsimony (MP)) were inferred using mega version 7.0.26 software [27].
To further investigate the taxonomic position of strains VNH17T and VNK23T in the genus Acinetobacter, comparative sequence analysis of RNA polymerase β-subunit gene (rpoB) was performed according to the method of La Scola et al. (2006) [28] and Nemec et al. (2009) [7], using two sets of primers to amplify two variable regions of the rpoB gene. Region 1 spans nucleotide positions 2916–3267 (primers Ac696F and Ac1093R), and region 2 spans nucleotide positions 3263–3773 (primers Ac1055F and Ac1598R). PCR products were purified, using the Inclone Gel & PCR purification kit (Inclone biotech), according to the supplier's instructions, and the DNA sequencing was performed by Macrogen Inc. (Seoul, Korea). The similarity between the rpoB nucleotide sequences of the isolates and the rpoB sequences of the type strains of Acinetobacter species were retrieved from their depositions in GenBank or from their whole genome sequences (http://www.ncbi.nlm.nih.gov, last accessed 10 July 2023) and then a neighbour-joining phylogenetic tree was constructed from these sequencing following the method as described above.
2.4. Genome Features
Genomic DNA of VNH17T and VNK23T was extracted using DNeasy Blood and Tissue kits (Qiagen, Hilden, Germany). The whole genome shotgun sequencing of those was performed by Macrogen (Seoul, Republic of Korea) using the Illumina HiSeq platform and assembled using SPAdes version 3.13.0 [29]. To analyze the evolutionary divergence based on whole-genome sequences, a phylogenetic tree of two isolates and 15 species within the genus Acinetobacter from the NCBI (www.ncbi.nlm.nih.gov/) was reconstructed in silico with the concatenated alignment of 92 core genes using the UBCG pipeline [30]. The DNA G+C content and average nucleotide identity (ANI) values between the whole genome sequence of two isolates and the close reference strains were calculated by using OrthoANIu (www.ezbiocloud.net/tools/ani) algorithm [31]. The digital DNA–DNA hybridization (dDDH) values between the strains VNH17T and VNK23T and between these and the type strains of genus Acinetobacter were computed on the GGDC web server (http://ggdc.dsmz.de/ggdc.php) [32]. The genome sequence of VNH17T and VNK23T was annotated by the Rapid Annotation using Subsystem Technology (RAST) server, version 2.0 [33]. To classify genes based on their function, the COG (Clusters of Orthologous Group) was performed by searching the KEGG (Kyoto Encyclopedia of Genes and Genomes) database [34].
3. Results and Discussion
3.1. Physiology and Chemotaxonomy
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn. On medium agar R2A within 48h at 30 - 35°C, the colonies color of strain VNH17T and VNK23T were white, circular, convex, and smooth surface, besides train VNK23T colonies were slightly opaque with entire margins but it was not observed in VNH17T. The colony sizes of VNH17T and VNK23T were significantly different when cultured on R2A after 48h incubation at 35 °C, the VNK23T colonies were 2-6 mm in diameter but VNH17T colonies were notably smaller 0.1 - 0.5 mm (Figure S2). They grow well on R2A, TSA, NA, LB, Mueller-Hinton, and MacConkey agar. Strains VNH17T and VNK23T had the basic features of the genus Acinetobacter, i.e. they were oxidase-negative, catalase-positive, cells were Gram-stain-negative, coccobacilli - shaped, incapable of dissimilative denitrification [35].
The two isolated strains shared a number of phenotypic characteristics tested in common consisting of cells that are 1.0–1.5 μm long and 0.8–1.0 μm wide. The hydrolysis of starch, Tween 80, casein, urease, aesculin, gelatin, and haemolysis of sheep blood were negative. Two isolated strains grew at 0–3 % (w/v) NaCl and their growth was inhibited by NaCl concentrations higher than 3 %. The pH range for growth was 6–10.0 (optimum: 6.5 -7.5). The temperature optimum for growth was 20–35 °C but the temperature rank for growth was different, VNH17T was growth from 10°C to 39°C but no growth was observed at 10°C with VNK23T. Both had no growth at temperatures above 41 °C within 14 days. In the same conditions test, no growth was observed on R2A in an anaerobic jar with VNK23T, whereas VNH17T was growth on R2A in an anaerobic jar for 5 days at 30°C. Strains VNH17T and VNK23T were sensitive to tetracycline, gentamycin, novobiocin, chloramphenicol, kanamycin, streptomycin, neomycin, rifampicin, cycloheximide, ampicillin, penicillin, tetracycline, and vancomycin. A detailed summary of the phenotypic features of two isolated strains is presented with the other closest strains (Table 1 and Table S1). All strains shared many phenotypic similarities, indicating their relatedness and placement in the genus Acinetobacter and some differential properties of two isolated strains compare with the closest strain show that two isolated strains were different from their closest neighbor.
Respiratory quinones of strain VNH17T and VNK23T were ubiquinone 9 (Q-9; 75.79% and 74.41%, respectively), ubiquinone 8 (Q-8; 22.41% and 23.67%, respectively), and a small amount of Q10 (Q-10, 1.8% and 1.92%, respectively) (Figure S4). Polar lipids included diphosphatidylglycerol (DPG), phosphatidylglycerol (PG), phosphatidylethanolamine (PE), unidentified phospholipid (PL), and no glycolipids were detected (Figure S5). Similar to those of the reference strains of members of the genus Acinetobacter, the prime fatty acid of strains VNH17T and VNK23T were C18:1ω9c (49.42% and 36.56%, respectively), C16:0 (18.96% and 22.53%, respectively), C12:03OH (3.79% and 4.18%, respectively), C12 : 0 (2.92% and 3.34%, respectively) and summed feature 3 (C16:1ω7c and/or C16 : 1ω6c), however, VNH17T had the presence of small amount of anteiso –C17:0 (0.17%), C14:1 ω5c (0.1%) and C18:3 ω6c (0.11%) which were not detected in VNK23T (Table 2). The fatty acid composition of VNH17T and VNK23T is consistence with previous results for recognized species of the genus Acinetobacter. In addition, the total fatty acid compositions of strain VNH17T showed quantity and composition differences from VNK23T showing that they could be two separate species.
3.2. Phylogenetic analysis based on 16S rRNA and rpoB genes
Analysis of the 16S rRNA gene sequences showed that VNH17T (1468 nucleotides) and VNK23T (1472 nucleotides) were separated into a cluster in a phylogenetic tree in the genus Acinetobacter of the family Moraxellaceae (Figure 1). The close relationship between the two organisms was reflected by the 100 % bootstrap support and the topology of the NJ tree was supported by those of the MP (Figure S6) and ML trees (Figure S7). Strain VNH17T was most closely related to Acinetobacter modestus (96.98%) and VNK23T was most closely related to Acinetobacter junii (97.26%). In addition, the phylogenetic tree based on rpoB gene sequence revealed that the two strains were distinct from all known Acinetobacter species (Figure 2). In rpoB gene sequence analyze, two isolates (VNH17T and VNK23T) had <95% identity to type or reference strains of all known Acientobacter species, VNH17T was most closely related to A. vivianii KJ-1 (94.65%) and A. colistiniresistens C-214 (93.45%); VNK23 was most closely related to A. vivianii KJ-1 (93.25%) and A. gyllenbergii C-214 (92.69%). It is suggested that rpoB gene sequence similarity values below 95 % may indicate distinct species in the genus Acinetobacter [28, 36]. In this study, the two isolated strains showed lower than 95% dissimilarity of rpoB with the type strains of recognized species of the genus Acinetobacter, this suggests that the isolated strains represent novel species. For verification of this proposal, we further genome features analysis.
3.3. Genome Features
The results indicated that strains VNH17T and VNK23T make clustered with strains Acinetobacter parvus DSM 16617T and Acinetobacter junii CIP 64.5T (Figure 3). The complete genome of strains VNH17T was 3873739 bp in size, the N50 of the filtered reads was 331771 kb, the genome coverage was 56.68×, and the DNA G+C content was 42.1 mol%. For VNK23T was 3947147 bp in size of the complete genome, the N50 of the filtered reads was 174689 kb, the genome coverage was 55.73×, and the DNA G+C content was 41.8 mol%. The Rast analysis revealed the presence of 3721 coding sequences, 72 RNAs, 291 subsystems in the genome in strain VNH17T and these values with VNK23T were 3926 coding sequences, 77 RNAs, and 289 subsystems. Pairwise ANI and dDDH values between strain VNH17T and VNK23T were 96.05 and 65.70%, respectively, the pairwise ANI values between them and other species of the genus Acinetobacter tested ranging from 74.41 to 91.61 and dDDH values ranging from 20.6 to 45.40% (Table S2). In light of the recommended threshold values of ANI 95–96% [37] and dDDH 70% [33] for species circumscription, these values indicate that The ANI value between VNH17T and VNK23T was 0.05% higher than generally accepted the species boundary but the dDDH value was within the accepted species boundary. These values indicate that the two isolated strains are clearly separated from all known taxa, however, two isolated strains are needed to be subjected to further genome micro restriction analysis. A total of 3586 protein-coding genes were identified for VNH17T, of which 3461 (96.5%) were assigned to COG categories, these numbers present for VNK23T were 3722 and 3525 (94.7%), respectively. The COG category assignment of genes to general function prediction of VNH17T and VNK23T was 13.5% (468 genes) and 14.0% (495 genes), and other known functions included amino acid transport and metabolism 5.0% (175 genes) and 4.7% (166 genes), inorganic ion transport and metabolism (6.1% (211 genes) and 5.3% (186 genes), and energy production and conversion (4.45% (154 genes) and 4.3% (152 genes), 28.3% and 28.5 % of genes functioned unknown into the COGs, respectively (Figure S8).
4. Conclusions
On the basis of the phylogenetic tree, strains VNH17T and VNK23T can be affiliated with the genus Acinetobacter. Both strains can be clearly differentiated from each other and from closely related Acinetobacter species on the basis of isolation source, growth characteristics, enzymic activities, assimilation patterns, and genetic analysis. Furthermore, the colonies' shape and size, rank temperature for growth, growth under anaerobic condition and the fatty acid component of strain VNH17T are clearly distinct. Thus, strains VNH17T and VNK23T represent two novel species of the genus Acinetobacter, for which the names Acinetobacter thutiue sp. nov. and Acinetobacter kontum sp. nov., respectively, are proposed. Future research should be aimed at clarifying if Acinetobacter thutiue sp. nov. and Acinetobacter kontum sp. nov. have any relevant role in oil degradation, which is intended to elucidate their importance for hydrocarbon degradation capacity, their growth properties in the oil pollution environment as well as will be necessary to elucidate their significance in oil contaminated soil ecosystem.
Description of Acinetobacter thutiue sp. nov.
Acinetobacter thutiue (thu.ti.ue. N.L. neut. adj. thutihue from Thua Thien Hue, the region of Vietnam from where the type strain was isolated).
Cells are Gram-negative, aerobic, oxidase-negative, catalase-positive, non-motile, coccobacilli shape. Colonies on R2A after 48h incubation at 35°C approximately 0.1 - 0.5 mm in diameter, circular, convex, smooth, and milk-white in colour. Cells grow at a temperature of 10–39°C (optimum, 20–35°C) and pH 6.0–10.0 (optimum pH, 6.5–7.5). Growth under anaerobic conditions was determined after incubation in an anaerobic jar on R2A after 5 days at 30°C. The hydrolyses of starch, Tween 80, casein, urease, aesculin, gelatin, and haemolysis of sheep blood were negative. Acid is not produced from d-glucose, non-gelatin hydrolyzes. For enzyme activity, positive reactions were observed with lipase, valine arylamidase, and acid phosphate whereas alkaline phosphatase, and α-glucosidase are negative. The following substrates were utilized as a sole carbon source: sodium acetate, propionic acid, capric acid, valeric acid, trisodium citrate, l-proline. The predominant respiratory lipoquinones, the major polar lipidsand the major cellular fatty acids correspond to those of the genus Acinetobacter.
The type strain is VNH17T (= KACC 23003 = CCTCC AB 2023063), isolated from oil-contaminated soil in the motorbike repairing workshop floors (Thua Thien Hue province, Vietnam). The DNA G+C content of the type strain is 42.1 mol%. GenBank accession numbers for sequences of the strain VNH17T were JAUDZE000000000 (genome) and OP727583 (16S rRNA gene nucleotide sequence).
Description of Acinetobacter kontum sp. nov.
Acinetobacter kontum (kon.tum N.L. neut. adj. kontum from Kontum, the region of Vietnam from where the type strain was isolated).
Cells are Gram-negative, strictly aerobic, oxidase-negative, catalase-positive, non-motile, coccobacilli shape. Colonies on R2A agar after 48 h incubation at 35 °C are approximately 2–6 mm in diameter, circular, convex, smooth, milk-white in colour and slightly opaque with entire margins. Growth occurs at temperatures ranging from 15 to 39 °C (optimum, 20–35°C), but not at 41 °C. Phenotypic characteristics correspond to the VNH17T i.e. the hydrolyses of starch, Tween 80, casein, urease, aesculin, gelatin, and haemolysis of sheep blood were negative. Acid is not produced from d-glucose, non-gelatin hydrolyzes, incapable of denitrification. Positive reactions were observed with lipase, valine arylamidase, and acid phosphate whereas alkaline phosphatase, and α-glucosidase are negative. The following substrates were utilized as a sole carbon source: sodium acetate, propionic acid, capric acid, valeric acid, trisodium citrate, l-proline, and no growth occurs on lactic acid, D-sorbitol, suberic acid, D-glucose, D-sorbitol, L-histidine, 3-hydroxybutyric acid.
The type strain is VNK23T (= KACC 23002 = CCTCC AB 2023064), isolated from oil-contaminated soil in the motorbike repairing workshop floors (Kon Tum province, Vietnam). The DNA G+C content of the type strain is 41.8 mol%. GenBank accession numbers for sequences of the strain VNK23T were JASXTK000000000 (genome) and OP727589 (16S rRNA gene nucleotide sequence).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Sampling site from the automobile workshops; Figure S2: Transmission electron microscopy of strains VNK23T and VNH17 T growth on R2A medium plates for 2 days at 30°C; Figure S3: Colonies of VNK23 T and VNH17 T were grown on R2A at 35 °C for 48 h; Figure S4: Quinone analysis results of strains VNH17T and VNK23T; Figure S5: Polar lipid profile of strain VNK23T and VNH17T; Figure S6: Phylogenetic tree reconstructed with the Minimum Evolution method based on 16S rRNA gene sequences of two isolated and type species of the genus Acinetobacter; Figure S7: Phylogenetic tree reconstructed with the Maximum Likelihood method based on 16S rRNA gene sequences of two isolated and type species of the genus Acinetobacter; Figure S8: COG functional classification of proteins in strains VNH17T and VNK23T genome; Table S1: Results from API ZYM, API 20NE test; Table S2: ANIb and dDDH values (%) between the genome sequences of the 16 strains in the genus Acinetobacter.
Author Contributions
N.L.T.T. conceived, designed and conducted all the experiments. N.L.T.T. and J.K. interpreted all the data, read, discussed, edited and approved the final draft of the manuscript. J.K. coordinated and supervised the study.
Funding
This work was supported by Kyonggi University’s Graduate Research Assistantship 2020.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Beijerinck, MW. Pigments as products of oxidation by bacterial action. Koninklijke Nederlandse Akademie Van Wetenschappen Proc Ser B Phy Sci, 1910, 13, 1066–77. [Google Scholar]
- Brisou J, Prevot AR. Etudes de Systematique Bacterienne. X. Revision des especes reunies dans le genre Achromobacter. Ann Inst Pasteur, 1954, 86, 722–728. [Google Scholar]
- Parte, AC. LPSN—List of prokaryotic names with standing in nomenclature (bacterio.net), 20 years on. Int J Syst Evol Microbiol, 2018, 68, 1825–1829. [Google Scholar] [CrossRef] [PubMed]
- Vaz-Moreira I, Novo A, Hantsis-Zacharov E, Lopes AR, Gomila M et al. Acinetobacter rudis sp. nov., isolated from raw milk and raw wastewater. Int J Syst Evol Microbiol, 2011, 61, 2837–2843. [Google Scholar] [CrossRef] [PubMed]
- Marí-Almirall M, Cosgaya C, Pons MJ, Nemec A, Ochoa TJ et al. Pathogenic Acinetobacter species including the novel Acinetobacter dijkshoorniae recovered from market meat in Peru. Int J Food Microbiol, 2019, 305, 108248. [Google Scholar] [CrossRef] [PubMed]
- Carvalheira A, Gonzales-Siles L, Salva-Serra F, Lindgren A, Svensson-Stadler L, Thorell K, Pineiro-Iglesias B, Karlsson R, Silva J, Teixeira P, et al. Acinetobacter portensis sp. nov. and Acinetobacter guerrae sp. nov., isolated from raw meat. Int J Syst Evol Microbiol, 2020, 70, 4544–4554. [Google Scholar] [CrossRef] [PubMed]
- Nemec A, Musilek M, Maixnerova M, De Baere T, van der Reijden TJ, Vaneechoutte M, Dijkshoorn L. Acinetobacter beijerinckii sp. nov. and Acinetobacter gyllenbergii sp. nov., haemolytic organisms isolated from humans. Int J Syst Evol Microbiol, 2009, 59, 118–124.
- Yacouba A, Sissoko S, Tchoupou Saha OF, Haddad G, Dubourg G, Gouriet F, Tidjani Alou M, Alibar S, Million M, Lagier JC, et al. Description of Acinetobacter ihumii sp. nov., Microbacterium ihumii sp. nov., and Gulosibacter massiliensis sp. nov., three new bacteria isolated from human blood. FEMS Microbiol Lett, 2022, 369, 0. [Google Scholar]
- Alvarez-Perez S, Lievens B, Jacquemyn H, Herrera CM. Acinetobacter nectaris sp. nov. and Acinetobacter boissieri sp. nov., isolated from floral nectar of wild Mediterranean insect-pollinated plants. Int J Syst Evol Microbiol, 2013, 63, 1532–1539. [CrossRef]
- Krizova L, Maixnerova M, Sedo O, Nemec A. Acinetobacter bohemicus sp. nov. widespread in natural soil and water ecosystems in the Czech Republic. Syst Appl Microbiol, 2014, 37, 467–473. [CrossRef]
- Radolfova-Krizova L, Maixnerova M, Nemec A. Acinetobacter celticus sp. nov., a psychrotolerant species widespread in natural soil and water ecosystems. Int J Syst Evol Microbiol, 2016, 66, 5392–5398. [CrossRef] [PubMed]
- Radolfova-Krizova L, Maixnerova M, Nemec A. Acinetobacter pragensis sp. nov., found in soil and water ecosystems. Int J Syst Evol Microbiol, 2016, 66, 3897–3903. [CrossRef]
- Yamahira K, Hirota K, Nakajima K, Morita N, Nodasaka Y, Yumoto I. Acinetobacter sp. strain Ths, a novel psychrotolerant and alkalitolerant bacterium that utilizes hydrocarbon. Extremophiles, 2008, 12, 729–734. [CrossRef]
- Dahal RH, Chaudhary DK, Kim J. Acinetobacter halotolerans sp. nov., a novel halotolerant, alkali tolerant, and hydrocarbon degrading bacterium, isolated from soil. Arch Microbiol, 2017, 199, 701–710. [CrossRef] [PubMed]
- Czarny J, Staninska-Pięta J, Piotrowska-Cyplik A, Juzwa W, Wolniewicz A, Marecik R, Lawniczak L, Chrzanowski L. Acinetobacter sp. as the key player in diesel oil degrading community exposed to PAHs and heavy metals. J Hazard Mater, 2020, 383, 121168. [CrossRef] [PubMed]
- Cai Y, Wang R, Rao P, Wu B, Yan L, Hu L, Park S, Ryu M, Zhou X. Bioremediation of Petroleum Hydrocarbons Using Acinetobacter sp. SCYY-5 Isolated from Contaminated Oil Sludge: Strategy and Effectiveness Study. Int J Environ Res Public Health, 2021, 19, 18(2):819. [Google Scholar]
- Xia M, Liu Y, Taylor AA, Fu D, Khan AR, Terry N. Crude oil depletion by bacterial strains isolated from a petroleum hydrocarbon impacted solid waste management site in California. Int. Biodeterior. Biodegrad, 2017, 123, 70–77. [CrossRef]
- Lányi, B. Classical and Rapid Identification Method for Medically Important Bacteria. In Method in Microbiology 19; Cowell R, Ed.; Academic Press: Oak Ridge, TN, USA, 1987; pp. 1–65. [Google Scholar]
- Schaeffer AB, Fulton M. A simplified method of staining endospores. Science, 1933, 77, 194. [Google Scholar] [CrossRef]
- Tindall BJ, Sikorski J, Smibert RA, Krieg NR. Phenotypic Characterization and the Principles of Comparative Systematics. In Methods for General and Molecular Microbiology; Reddy CA, Ed, Ed.; ASM Press: Washington, DC, USA, 2007; pp. 330–393. [Google Scholar]
- Smibert RM, Krieg NR. Phenotypic characterization. In Methods for General and Molecular Bacteriology; Gerhardt P, Murray RGE, Wood WA, Krieg NR, Ed.; American Society for Microbiology: Washington, DC, USA, 1994; pp. 607–654. [Google Scholar]
- Dahal RH, Kim J. Rhabdobacter roseus gen. nov., sp. nov., isolated from soil. Int J Syst Evol Microbiol, 2016, 66, 308–314.
- Minnikin D, O'Donnell A, Goodfellow M, Alderson G, Athalye M, Schaal A, et al. An integrated procedure for the extraction of bacterial isoprenoid quinones and polar lipids. J Microbiol Methods 1984, 2, 233–241. [CrossRef]
- Sasser, M. Identification of bacteria by gas chromatography of cellular fatty acids. Technol. Note 2001, 101, 1–6. [Google Scholar]
- Frank JA, Reich CI, Sharma S, Weisbaum JS, Wilson BA, Olsen GJ. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl Environ Microbiol, 2008, 74, 2461–2470. [Google Scholar] [CrossRef]
- Yoon SH, Ha SM, Kwon S, Lim J, Kim Y, Seo H, Chun J. Introducing EzBioCloud: A taxonomically United database of 16S rRNA gene sequences and whole-genome assemblies. Int J Syst Evol Microbiol, 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Kumar S, Stecher G, Tamura K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol, 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- La Scola B, Gundi VA, Khamis A, Raoult D. Sequencing of the rpoB gene and flanking spacers for molecular identification of Acinetobacter species. J Clin Microbiol, 2006, 44, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, Kulikov AS, Lesin VM, Nikolenko SI, Pham S, Prjibelski AD, et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol, 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Na SI, Kim YO, Yoon SH, Ha SM, Baek I, Chun J. UBCG: Up-to-date bacterial core gene set and pipeline for phylogenomic tree reconstruction. J Microbiol, 2018, 56, 280–285. [Google Scholar] [CrossRef]
- Yoon SH, Ha SM, Lim J, Kwon S, Chun J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek, 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
- Meier-Kolthoff JP, Auch AF, Klenk HP, Göker M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform, 2013, 14, 60. [Google Scholar]
- Aziz RK, Bartels D, Best AA, DeJongh M, Disz T, Edwards RA, Formsma K, Gerdes S, Glass EM, Kubal M, et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom, 2008, 9, 75. [Google Scholar]
- Kanehisa M, Goto S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res, 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Juni, E. Interspecies transformation of Acinetobacter: genetic evidence for a ubiquitous genus. J Bacteriol, 1972, 112, 917–931. [Google Scholar] [CrossRef]
- Narciso-da-Rocha C, Vaz-Moreira I, Svensson-Stadler L, et al. Diversity and antibiotic resistance of Acinetobacter spp. in water from the source to the tap. Appl Microbiol Biotechnol, 2013, 97, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Richter M, Rossello-Mora R. Shifting the genomic gold standard for the prokaryotic species definition. Proc Natl Acad Sci USA, 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The phylogenetic tree was reconstructed with the Neighbor-joining method based on 16S rRNA gene sequences of strain VNH17T, VNK23T, and type species of the genus Acinetobacter. Moraxella lacunata NCTC 7911T was used as an outgroup. Numbers at branch nodes represent confidence levels (values > 50 % are shown) from 1 000 replicate bootstrap samplings. GenBank accession numbers are shown in parentheses. Bar, 0.01 substitutions per nucleotide position.
Figure 1.
The phylogenetic tree was reconstructed with the Neighbor-joining method based on 16S rRNA gene sequences of strain VNH17T, VNK23T, and type species of the genus Acinetobacter. Moraxella lacunata NCTC 7911T was used as an outgroup. Numbers at branch nodes represent confidence levels (values > 50 % are shown) from 1 000 replicate bootstrap samplings. GenBank accession numbers are shown in parentheses. Bar, 0.01 substitutions per nucleotide position.
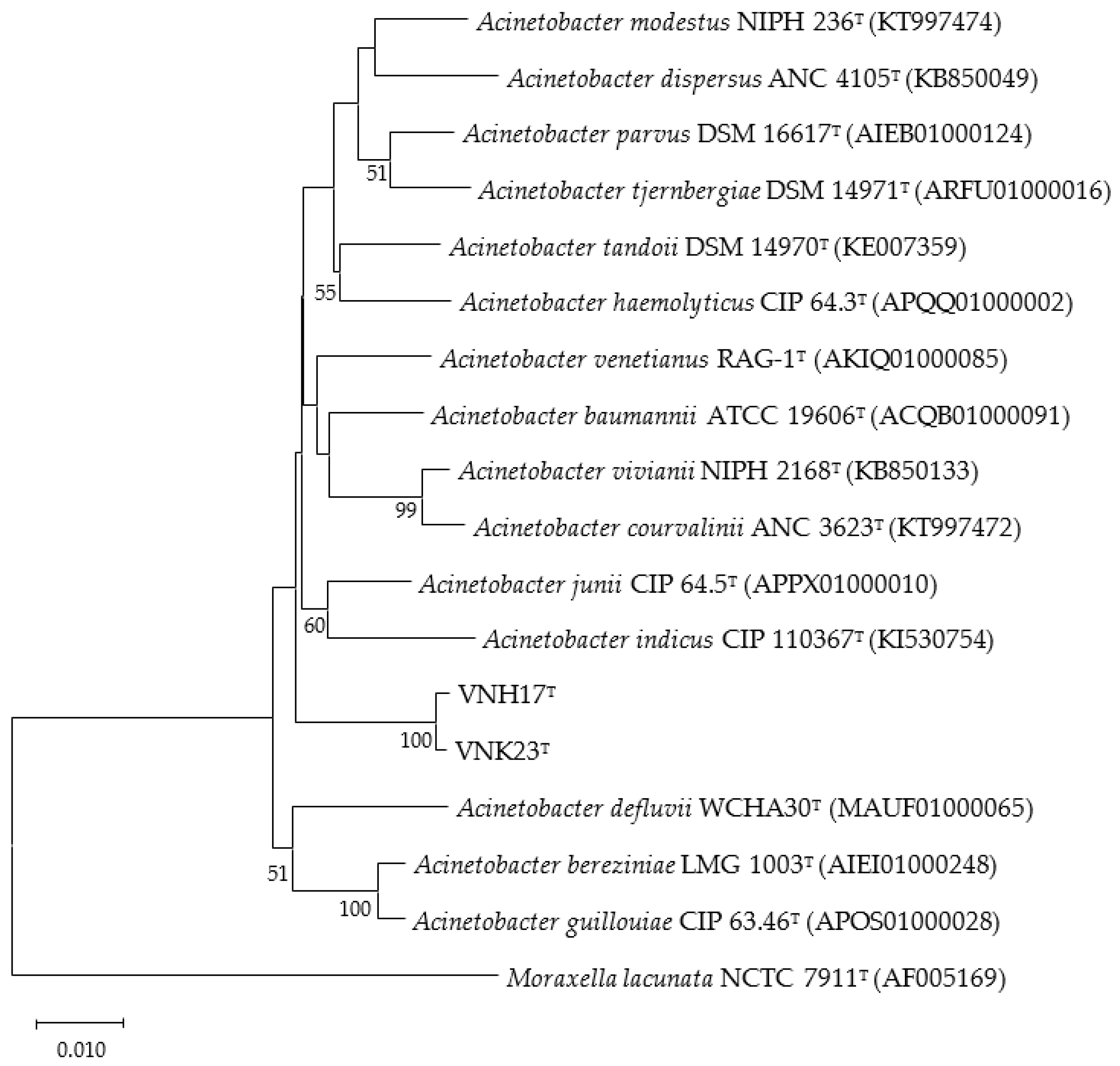
Figure 2.
Figure 1A. The phylogenetic tree was reconstructed with the Neighbor-joining method based on ropB gene sequences of strain VNH17T, VNK23T, and type species of the genus Acinetobacter. Numbers at branch nodes represent confidence levels (values > 50 % are shown) from 1 000 replicate bootstrap samplings. GenBank accession numbers are shown in parentheses. Bar, 0.01 substitutions per nucleotide position.
Figure 2.
Figure 1A. The phylogenetic tree was reconstructed with the Neighbor-joining method based on ropB gene sequences of strain VNH17T, VNK23T, and type species of the genus Acinetobacter. Numbers at branch nodes represent confidence levels (values > 50 % are shown) from 1 000 replicate bootstrap samplings. GenBank accession numbers are shown in parentheses. Bar, 0.01 substitutions per nucleotide position.
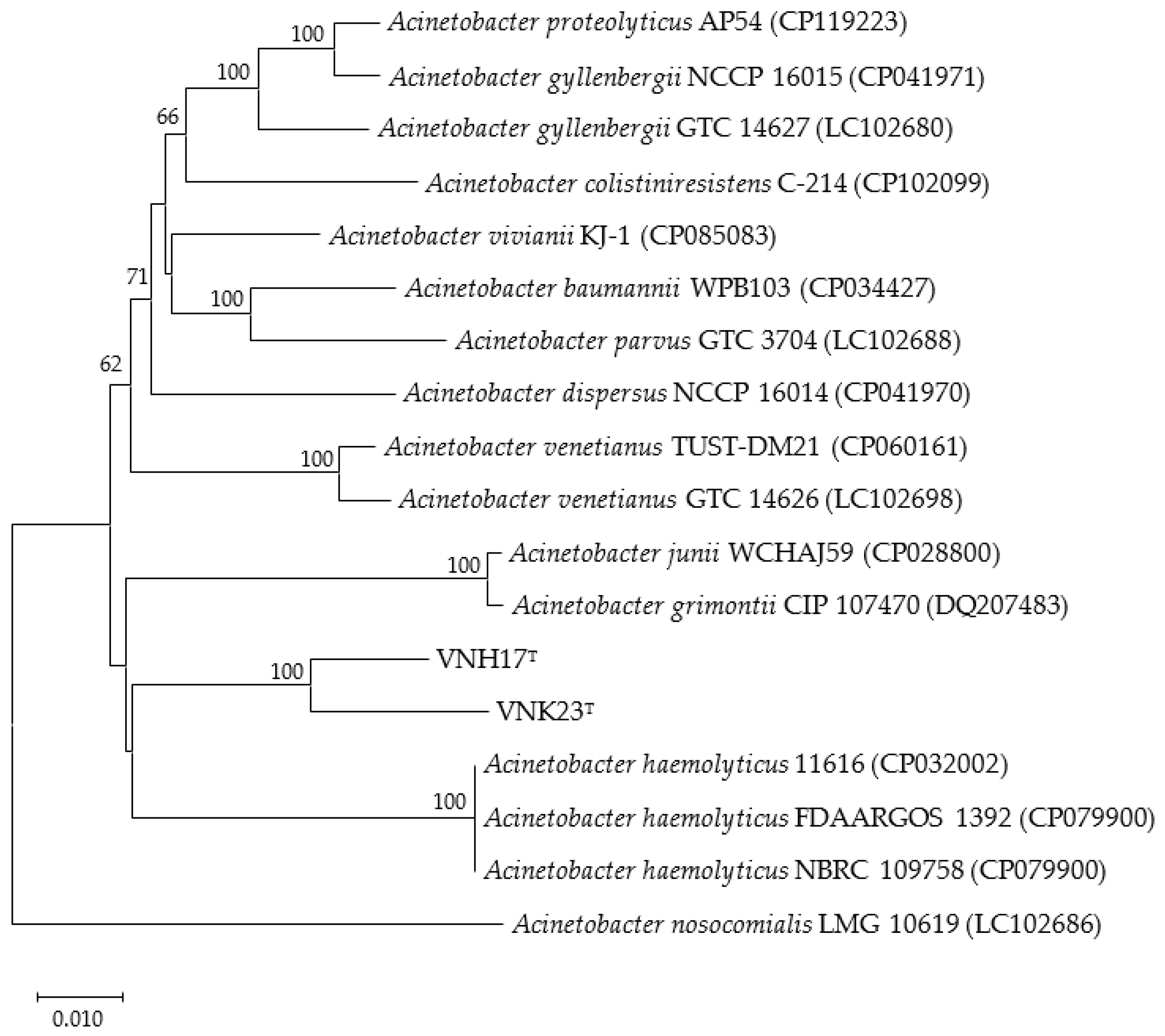
Figure 3.
The phylogenetic tree was reconstructed using the UBCG (concatenated alignment of 92 core genes) pipeline based on whole-genome sequences of strain VNH17T, VNK23T and the type species within the genus Acinetobacter. GenBank accession numbers are given in parentheses. Bar, 0.05 substitutions per nucleotide position.
Figure 3.
The phylogenetic tree was reconstructed using the UBCG (concatenated alignment of 92 core genes) pipeline based on whole-genome sequences of strain VNH17T, VNK23T and the type species within the genus Acinetobacter. GenBank accession numbers are given in parentheses. Bar, 0.05 substitutions per nucleotide position.


Table 1.
Phenotypic characteristics differentiate the novel species VNH17T and VNK23T from its closely related strains in the genus Acinetobacter. 1, VNH17T; 2, VNK23T; 3, Acinetobacter parvus KACC 12455; 4, Acinetobacter junii KACC12228; 5, Acinetobacter tandoii KACC 12229; 6, Acinetobacter indicus KCTC 42000; 7, Acinetobacter bereziniae KCTC 42001; 8, Acinetobacter venetianus DSM23050. Data are from the present study. +, positive; -, negative, +/−, weak positive or ambiguous.
Table 1.
Phenotypic characteristics differentiate the novel species VNH17T and VNK23T from its closely related strains in the genus Acinetobacter. 1, VNH17T; 2, VNK23T; 3, Acinetobacter parvus KACC 12455; 4, Acinetobacter junii KACC12228; 5, Acinetobacter tandoii KACC 12229; 6, Acinetobacter indicus KCTC 42000; 7, Acinetobacter bereziniae KCTC 42001; 8, Acinetobacter venetianus DSM23050. Data are from the present study. +, positive; -, negative, +/−, weak positive or ambiguous.
| Characteristic | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Source of isolation | Forest soil | Paddy soil | Sediment from an acidic hot spring and pyritic mine drainage | The thalli of Cladonia arbuscula and Cladonia stellaris lichens | Acidic mine drainage | Acidic northern wetlands |
| Cell shape | Ovals, Rods | Ovals, rods | Cocci, short rods | Spherical to ovoid | Rods | Coccobacilli |
| Cell size (µm) | 0.8-1.1 x1.5-3.2 | 2.0-3.0 x3.0-10 | 0.7-0.9 x0.9-1.6 | 0.8-1.6 x0.9-2.26 | 1.3-1.5 x3.2-3.3 | 0.7-1.5 x1.4-4.1 |
| Colony color | White, Mucoid | Red brown | Salmon pink | pink or salmon pink | Light yellow | White, cream |
| Motility | - | + | - | - | - | - |
| Anaerobic phototrophy | - | + | - | - | - | - |
| Bacteriochlorophyll a | - | + | + | + | - | - |
| Temperature range for growth (°C) | 20-40 | 20-40 | 20-40 | 5-30 | 20–45 | 2–30 |
| Optimal growth temperature (°C) | 30-35 | 30–35 | 20-35 | 10-15 | 37 | 18–22 |
| pH range for growth | 6-9 | 5.0–8.5 | 3.5-6.0 | 3.0–7.5 | 2.5–5 | 3.7–7.5 |
| pH optimum for growth | 7-8 | 6.0–6.5 | 4.5-5.0 | 4.5–5.5 | 4.0 | 5.0–5.7 |
| Tolerance of >2.5 (%w/v) NaCl | - | - | + | + | - | - |
| Electron donor/ carbon source | ||||||
| Acetate | - | ++ | - | - | nd | - |
| Lactate | - | + | + | - | nd | + |
| Mannose | +/- | + | - | + | - | + |
| Sorbitol | +/- | + | + | + | + | + |
| Glycerol | +/- | - | + | - | - | + |
| Asparagine | + | + | - | - | + | + |
| Aspartate | - | + | - | nd | + | - |
| Glutamate | - | ++ | - | - | nd | nd |
| Glutamine | ++ | ++ | - | - | nd | nd |
| Citrate | ++ | - | +/- | - | nd | - |
| Nitrogen fixation | - | + | nd | - | nd | nd |
| Major quinone(s) | Q-10 | Q-10, RQ-10 | Q-10 | Q-10 | Q-10 | Q-10 |
| DNA G+C content (mol%) | 68.7 | 67.8 | 69.1-69.8 | 69.1–69.8 | 65.9 | 60.5–61.9 |
Table 2.
Detailed cellular fatty acid profiles (% of totals) of strain VNH17T, VNK23T and closely related reference strains. Strains; 1, VNH17T; 2, VNK23T; 3, Acinetobacter parvus KACC12455; 4, Acinetobacter juni KACC12228; 5, Acinetobacter tondoii KACC12229; 6, Acinetobacter indicus KCTC42000; 7, Acinetobacter bereziniae KCTC42001; 8, Acinetobacter venetianus DSM23050. All data are from present study. Fatty acids that represent <0.1% of total in all strains are not shown; -, not detected.
Table 2.
Detailed cellular fatty acid profiles (% of totals) of strain VNH17T, VNK23T and closely related reference strains. Strains; 1, VNH17T; 2, VNK23T; 3, Acinetobacter parvus KACC12455; 4, Acinetobacter juni KACC12228; 5, Acinetobacter tondoii KACC12229; 6, Acinetobacter indicus KCTC42000; 7, Acinetobacter bereziniae KCTC42001; 8, Acinetobacter venetianus DSM23050. All data are from present study. Fatty acids that represent <0.1% of total in all strains are not shown; -, not detected.
| Fatty acid | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| Saturated | ||||||||
| C10:0 | 0.56 | 0.57 | 0.33 | 1.35 | 0.67 | 0.13 | - | 1.35 |
| C11:0 | - | - | - | 0.11 | - | - | - | |
| C12:0 | 2.92 | 3.34 | 6.21 | 2.80 | 5.88 | 6.55 | 9.31 | 5.18 |
| C14:0 | 0.98 | 1.19 | 0.52 | 0.57 | 1.27 | 0.81 | 0.78 | |
| C16:0 | 18.96 | 22.53 | 9.02 | 15.59 | 17.12 | 14.05 | 20.46 | 24.43 |
| N alcohol C16:0 | - | - | - | 0.14 | 1.10 | - | 0.17 | - |
| C17:0 | 0.19 | 0.19 | 0.37 | 0.92 | 0.54 | 0.22 | - | 0.29 |
| Iso C17:0 | - | 0.23 | 1.04 | 0.15 | - | - | - | |
| C18:0 | 2.12 | 0.33 | - | 1.68 | 1.31 | 1.85 | 1.19 | 0.97 |
| 10-methyl C17:0 | - | - | - | - | 0.47 | - | - | - |
| anteiso -C17:0 | 0.17 | - | - | - | - | - | - | - |
| Unsaturated | ||||||||
| C14:1 ω5c | 0.10 | - | - | - | - | - | - | - |
| C16:1 ω5c | - | - | - | 0.10 | - | - | - | - |
| C16:1 ω7c alcohol | - | - | - | - | 0.49 | - | - | - |
| C16:1 ω9c | - | - | - | 0.9 | - | - | - | - |
| C17:1 ω8c | 0.60 | 0.77 | 2.49 | 2.86 | 0.95 | 0.58 | 0.22 | 1.22 |
| C18:1 ω9c | 49.42 | 36.56 | 39.37 | 38.41 | 31.21 | 29.76 | 30.14 | 29.68 |
| C18:1 ω5c | - | - | - | - | - | - | - | 0.11 |
| C18:3 ω6c | 0.11 | - | - | 0.22 | 1.28 | - | 0.21 | - |
| Iso I C19:1 | 0.14 | 0.31 | - | - | 0.15 | - | 0.18 | 0.17 |
| Hydroxy | ||||||||
| C8:0 3OH | - | - | 0.13 | - | - | - | - | - |
| C12:0 2OH | 2.15 | 2.33 | 3.63 | 2.89 | 1.51 | 0.57 | 0.63 | 2.25 |
| C12:0 3OH | 3.79 | 4.18 | 6.73 | 4.75 | 4.68 | 3.04 | 5.59 | 5.19 |
| Summed features* | ||||||||
| Summed Feature 1 | - | - | - | 0.13 | - | - | - | - |
| Summed Feature 2 | 0.14 | 0.24 | 0.28 | 0.3 | 0.16 | 2.61 | 0.10 | 0.22 |
| Summed Feature 3 | 16.39 | 25.89 | 29.75 | 23.70 | 28.15 | 36.84 | 28.41 | 26.90 |
| Summed Feature 8 | 1.18 | 1.48 | 0.94 | 0.89 | 3.54 | 2.49 | 2.40 | 1.17 |
*Summed features represent two or three fatty acids that cannot be separated by the Microbial Indentification System. Summed feature 1 consisted of C15:1 iso H/C13:0 3OH, summed feature 2 consisted of C16:1 iso I/C14:0 3OH; summed feature 3 consist of C16:1 ω7c/C16:1 ω6c summed feature 8 consist of C18:1 ω7c/ C18:1 ω6c.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



