
Submitted:
18 July 2023
Posted:
20 July 2023
You are already at the latest version
Abstract
Extracellular vesicles (EVs) are emerging mediators of intracellular and inter-organ communications in cardiovascular diseases (CVDs), especially in the pathogenesis of heart failure through the transference of EV-containing bioactive substances. microRNAs (miRNAs) are contained in EV cargo and are involved in the progression of heart failure. Over the past several years, a growing body of evidence has suggested that the biogenesis of miRNAs and EVs are tightly regulated, and the sorting of miRNAs into EVs is highly selective and tightly controlled. Extracellular miRNAs, in particular circulating EV-miRNAs, have shown promising potential as prognostic and diagnostic biomarkers for heart failure and as therapeutic targets. In this review, we summarize the latest progress concerning the role of EV-miRNAs in HF and their application in a therapeutic strategy development for heart failure.
Keywords:
Extracellular vesicles
; MicroRNAs
; Sorting mechanism
; Biomarkers
; Heart failure
1. Introduction
Cardiovascular disease, including chronic heart failure (HF), is the number one cause of mortality and morbidity in the United States. The 2021 statistical update from the American Heart Association estimated more than 6 million Americans are living with HF. This equates to approximately 1.8% of the total US population and this number will rise by 46% over the next several years to reach approximately 8 million by 2030 [1]. Accumulating evidence suggests that HF is a vicious cycle and neurohormonal dysregulation including sympathetic nervous system excitation, the renin-angiotensin system activation and others contribute to the pathogenesis of HF [2-8].
It has also been well documented that clinically relevant signaling pathways (e.g. those activating angiotensin and adrenergic pathways) are well-coordinated during the development and progression of HF through a signature pattern of miRNA expression highly associated with cardiac hypertrophy and fibrosis in both mouse and human HF [9,10]. Importantly, these studies suggest that individual cardiac stress-inducible miRNAs (e.g. miR-208 and miR-195) are sufficient to provoke HF.
miRNAs are a class of non-coding RNAs and well-known as fundamental regulators of gene expression by binding the 3′ UTR of targeting mRNAs or other regions, including the 5′ UTR of mRNA, coding sequence, and gene promoters to induce mRNA degradation or translational repression [11-14]. Despite the involvement of miRNAs in the pathophysiology of maladaptive remodeling in HF ([15-18], extracellular miRNAs, especially those carried in extracellular vesicles (EVs), are promising biomarkers and paracrine mediators of intra- and inter-organ communication as well as potential therapeutic targets. This review will focus on the roles of EV miRNAs in intercellular and inter-organ communication in the progression of HF and summarize the current progress in miRNA selectively packaged into EVs. We also focus on utilizing and targeting extracellular miRNAs as diagnostic biomarkers and therapy in HF.
2. miRNA Biogenesis and Extracellular Vesicle Selection
miRNAs as a dominating class of small ncRNAs are approximately 22 nucleotides in length and are produced by Drosha and Dicer, two RNase III proteins (see Figure 1).
miRNAs mediate RNA silencing by targeting most protein-coding transcripts to modulate pathological processes associated with HF. Extracellular vesicles are natural membrane-bound nanoparticles released by all cells, have a wide range of diameters and are produced by different intracellular pathways. A growing body of literature shows that EVs are important mediators of intercellular and intra-organ communication in the pathogenesis of HF by transferring bioactive materials, including miRNAs. To better understand the various roles of EV miRNAs in the progression of HF it will be necessary to understand the biogenesis of miRNAs and EVs and the selection mechanisms of extracellular miRNAs.
2.1. miRNAs and Extracellular Vesicle Biogenesis
miRNA biogenesis is tightly controlled at all levels including transcription, processing, modification and Argonaute (AGO) protein loading as well as RNA decay [22]. Increasing evidence supports the view that dysregulation of miRNA biogenesis is associated with human HF [23-26]. miRNA biogenesis can be regulated either by the canonical pathway (Figure 1A) or non-canonical pathways (Figure 1B,C,E). The canonical pathway comprises a series of steps: First, miRNA transcription is carried out by RNA polymerase II (Pol II) and Pol II-associated transcription factors and epigenetic regulators to pri-miRNAs [27-30], followed by nuclear processing of pri-miRNAs by Drosha and DGCR8 (microprocessor complex) to pre-miRNAs [31,32]. Second, following nuclear events, nuclear export of pre-miRNAs is mediated by a transport complex comprising exportin 5 (XPO5) and GTP-binding RAN (Ras-related nuclear protein) in the cytoplasm [33], where pre-miRNAs are processed by Dicer, one RNase III-type enzyme, and TRBP (HIV-1 transactivating response (TAR) RNA-binding protein) to miRNA-duplex [34,35]. Third, selective miRNA-duplexes are loaded onto AGO proteins to form an RNA-induced silencing complex (RISC), unwinding the miRNA-duplexes. Theoretically, if one miRNA strand is selectively loaded onto an AGO protein to form the mature RISC, which further cleaves target mRNA or represses mRNA translation by binding to 3’-UTR of mRNA [36,37], the other strand will be ejected from the RISC and subjected to degradation. However, increasing evidence not only suggests that miRNAs can bind to other regions of targets in addition to 3’-UTR including 5’-UTR, coding area and promoter regions to activate translation or regulate transcription [38-40], but also both miRNA strands were found to functionally co-exist and participate in the pathogenesis of HF [41,42].
In addition to the canonical pathway, there are several non-canonical pathways involved in miRNA biogenesis. These non-canonical pathways are generally divided into two groups: Drosha/DGCR8-independent (Figure 1B,C) and Dicer-independent pathways (Figure 1E). Mirtron is one type of pre-miRNAs produced from introns of mRNAs by splicing [43]. The 7-methylguanosine (m7G)-capped pre-miRNAs are also produced by Drosha/DGCR8-independent pathway [44]. Both pre-miRNAs are still dependent on Dicer-mediated cytoplasmic maturation, but they differ in nuclear export. Mirtron is exported by XPO5, whereas m7G-capped pre-miRNA is exported by XPO1 [22,44]. In addition, some miRNAs are made through Dicer-independent pathways (e.g. miR-451). The product of pri-miRNA-451 cleavage by Drosha is too short to be processed by Dicer, and pre-miRNA-451 will be directly loaded onto AGO2 and then sliced into an AGO-cleaved pre-miRNA-451 (ac-pre-miR-451) which will be further matured by poly (A)-specific ribonuclease PARN-mediated trimming [45].
Extracellular vesicles (EVs), as one type of membrane-enclosed nanoparticle, are attractive mediators of intercellular and inter-organ communication in various diseases including HF [46-49]. Currently, EVs are divided into three types of vesicles in terms of their origin and mechanisms of biogenesis [50] (see Figure 1): (1) Exosomes (EXOs) (50-160 nm); (2) Microvesicles (MVs) (100 nm-1000 nm) and (3) Apoptotic bodies (APO-EVs) (1-5 μm). EXOs are a type of EV with an endosomal origin made by sequential invagination of the cell membrane resulting in the formation of multivesicular bodies (MVBs). The MVBs will ultimately generate exosomes by fusing with the plasma membrane and undergoing exocytosis [51,52]. Increasing evidence suggests that the biogenesis of EXOs is tightly regulated not only by the endosomal sorting complex required for transport (ESCRT) machinery, Syndecan-syntenin-ALIX (Apoptosis-linked gene 2-interacting protein X), Tetraspanins and ceramides during the formation of MVBs, but also by cytoskeletal elements, molecular motors and Ras-associated binding GTPases (RABs) during the transport/docking of MVBs from cytoplasm to cell membrane. In addition, the final fusion of MVBs with the plasma membrane and the secretion of EXOs are driven by the soluble N-ethylmaleimide-sensitive factor attachment protein receptors (SNAREs), sGTPases and calcium [50,52].
MVs are membrane-bound vesicles with a size range of 100 nm to 1000 nm, produced by direct budding and pinching of the plasma membrane. Although the biogenesis mechanisms are not as well understood, increasing evidence suggests that the formation of MVs are in addition to being highly related to the regulation of cytoskeletal elements by small GTPases, such as Rho (RAS homolog) family and ADP-ribosylation factors (ARFs), they are also associated with the recruitment of the Tumor susceptibility gene 101 (TSG101) by Arrestin domain-containing protein 1 (ARRDC1) to the plasma membrane facilitating the shedding and release of MVs [53,54].
Other than EXOs and MVs, apoptotic bodies are also EVs which are generated by dying cells during apoptosis and generally recognized and engulfed by phagocytes [55]. However, recent studies suggested that apoptotic bodies are generated by budding from the plasma membrane. The formation of apoptotic bodies is regulated by apoptotic cell disassembly via several molecular regulators including ROCK, Pannexin-1 and Plexin-B2 (well-summarized in [56]). In addition, another novel beads-on-a-string membrane structure is also involved in the formation of apoptotic bodies [57]. Increasing evidence suggests that apoptotic bodies not only facilitate the clearance and degradation of apoptotic materials but also contain other biomolecular cargos (e.g. miRNAs, DNA, protein and lipids) to mediate intercellular communication [58,59].
Although these membrane-bound vesicles differ in size distribution and mechanism of biogenesis, they are secreted by cells under various pathophysiological conditions into the extracellular space where they mediate intercellular and inter-organ communication via EV bioactive substances including proteins, RNAs, DNAs and lipids. Accumulating evidence suggests that packaging of cargos into EVs is highly selective and tightly regulated. In this review, we will also focus on the selective packaging of miRNAs into EVs.
2.2. Mechanisms of miRNA Selection into EVs
Extracellular miRNAs (Ex-miRs) have been found to be stably transported by ribonucleoproteins (RNPs), lipoproteins and neutrophil extracellular traps [60-62]. However, it has become attractive to view EV function as intercellular and inter-organ mediators conveying their cargos including non-coding RNAs, in particular, miRNAs. Interestingly, the existence of EV-miRs in the circulation of patients with cardiovascular diseases (CVDs), including HF, has raised the possibility that EV-miRs serve as prognostic and diagnostic markers and potential therapeutic targets [63-65]. However, the underlying mechanisms by which miRNAs are selectively secreted remain unclear.
In 2010, a study by Kosaka et al. [21] revealed that secretory miRNAs are regulated by the neutral sphingomyelinase 2 (nSMase 2) which regulates the biogenesis of ceramide and triggers exosome secretion, rather than by the ESCRT system, while the members of ESCRT regulate the biogenesis of EXOs. Increasing studies further suggest that the sorting of miRNA into EVs and their secretion are controlled by specific sequence motifs present in miRNAs that are recognized by RNA-binding Proteins (Figure 1G) including Heterogeneous nuclear ribonucleoprotein A2B1 (hnRNPA2B1) [66] and hnRNPU [67], Y-box protein 1 (YBX1) [68,69], Synaptotagmin-binding cytoplasmic RNA-interacting protein (SYNCRIP) [70,71], ELAV-like protein 1 (or HuR, human antigen R) [72,73], HSP90AB1, XPO5 and major vault protein (MVP) [72,74] and Serine/arginine splicing factor 1 (SRSF1) [75]. Recently, another RNA-binding protein, Lupus La, has been identified to mediate the selective sorting of miRNAs into EVs, and in particular, it selectively sorts miRNA-122 through the specific motifs located at the 3’ end of miRNA-122 [76]. Further studies by analyzing either EV-enriched (EXOmotifs) or Cell-enriched miRNA sequence (CELLmotifs) and structure revealed that the sorting sequences (EXOmotifs) present in miRNAs determine their secretion by EVs. Two novel RNA-binding proteins, Alyref and Fus, have also been identified to function as at least two RNA-binding proteins responsible for EXOmotif recognition and miRNA export into EVs [19].
Additionally, the post-translational modifications of some RNA-Binding Proteins, such as SUMOylation [66], oligomerization and ubiquitination [73], Liquid-liquid phase separation (LLPS) [77] and O-GlcNAcylation [78], also control the sorting of miRNAs into EVs. Interestingly, cell activation-dependent alterations of miRNA targets promote the sorting of miRNAs into EVs [79], and modifications of miRNAs also determine the distribution of miRNA in EVs, such as 3’-end uridylation rather than adenylation [20]. In addition, the ALG-2-interacting protein X (Alix), an accessory protein of ESCRT, is also involved in the miRNA sorting to EVs by interacting with Ago2 and miRNAs during the EV biogenesis [80]. Caveolin-1 (Cav-1) was identified as the first membrane protein to be involved in the selective sorting of miRNAs to EVs by directly interacting with hnRNPA2B1, and the phosphorylation of Cav-1 at Y14 not only promotes the O-GlcNAcylation of hnRNPA2B1 but also enhances the interaction between Cav-1 and O-GlcNAcylated hnRNPA2B1, subsequently facilitating the trafficking of the Cav-1/hnRNPA2B1/miRNAs complex into MVs [78]. The studies and mechanisms cited above strongly support the conclusion that the sorting of miRNAs into EVs is highly selective and that EV secretion is tightly controlled, which may contribute to pathophysiological alterations in human diseases, including HF. For example, these EV sorting mechanisms of miRNAs have been potentially involved in the biogenesis of EV-miRNAs in heart failure (See Table 1).
3. EV miRNAs in the Pathogenesis of Heart Failure
To better understand the role of EV-miRNAs in cardiac homeostasis and pathology, recent large-scale single-cell sequencing data revealed that the cellular composition of the adult human heart is heterogenous and shows transcriptional and cellular diversity [91,92]. The human heart is composed of nine major cell types, including cardiomyocytes, cardiac fibroblasts, endothelial cells, macrophages, etc., and 20 sub-clusters of cell types within the heart [91]. Cardiac homeostasis is normally maintained by dynamic cell-cell and cell-extracellular matrix interactions [93]. When this cardiac homeostasis is disrupted, such as in response to cardiac disease or cardiac damage, the heart will undergo remodeling which includes fibrosis and hypertrophy - both hallmarks of the HF state [94,95]. Extracellular vesicles are involved in both intercellular and inter-organ cross-talk via the effective transfer of bioactive substances into recipient cells, in particular miRNAs, which are key players in the pathogenesis of HF [96-98] (See Table 2).
3.1. EV-miRNA in the Cardiac Hypertrophy
Under cardiac stress, the heart undergoes left ventricular hypertrophy, which is an initial compensatory mechanism. Although cardiac fibroblast-derived EVs play a critical role in inducing cardiac hypertrophy by activating the Renin Angiotensin System in cardiomyocytes [120], the EV-enriched miRNAs secreted by cardiac fibroblasts in response to cardiac stress, such as miRNA-21-3p [41] and miRNA-27a-5p [42], can also be taken up by cardiomyocytes, resulting in the cardiac hypertrophy via translational inhibition of both SORBS2 and PDLIM5 [41] or PDLIM5 [42]. In addition, studies from our group also suggest that in response to cardiac stress, cardiac fibroblasts secrete EVs abundant with three miRNAs including miRNA-27a, miRNA-28-3p and miRNA-34a. They are taken up by cardiomyocytes where they mediate the oxidative stress response by targeting Nrf2/ARE signaling, leading to cardiac hypertrophy [99]. In addition to cardiac fibroblasts, adipocytes increase miR-200a expression and secretion by EVs in response to selective activation of PPARγ signaling, and the uptake of EV-miR-200a by cardiomyocytes results in decreased tuberous sclerosis complex (TSC1) and subsequent mTOR activation, leading to cardiomyocyte hypertrophy [100].
3.2. EV-miRNA in the Cardiac Fibrosis
Cardiac fibrosis is another hallmark of chronic HF, characterized by extracellular matrix degradation and collagen accumulation. As one of the cardiac-specific miRNAs, miRNA-208a can also be secreted via EVs to mediate the intercellular communication between cardiomyocytes and cardiac fibroblasts and further facilitate cardiac fibroblast proliferation and differentiation into myofibroblasts [104], being transcribed by an intron of alpha-myosin heavy chain (Myh6) in cardiomyocytes, and contributes to cardiac hypertrophy and conduction defects in the heart [101,102]. This molecular mechanism further revealed that EV-miRNA-208a promotes cardiac fibrosis by targeting Dyrk2 (dual-specificity tyrosine phosphorylation-regulated kinase 2) to promote NFAT (nuclear factor of activated T cells) dephosphorylation and nuclear translocation, which triggers fibrosis. In addition, miRNA-217 was found to be elevated in hearts of patients with chronic HF, and exogenous overexpression of miRNA-217 in cardiomyocytes in the thoracic aortic constriction (TAC)-induced HF model demonstrated enhanced pressure overload-induced cardiac dysfunction and cardiac remodeling (cardiac fibrosis and hypertrophy). Interestingly, miRNA-217 not only directly regulates cardiac hypertrophy but also indirectly contributes to cardiac fibrosis via cardiomyocyte-derived EVs by targeting PTEN in both cells [103]. Moreover, under cardiac stress, such as mechanical stretch and pressure overload, cardiomyocyte-specific Peli1 (Pellino E3 Ubiquitin Protein Ligase 1) has been involved in the enhanced transcription of miRNA-494-3p in cardiomyocytes via regulating NF-ĸB/AP-1 activation, and promotes the secretion of EV-enriched miRNA-494-3p into cardiac fibroblasts to activate cardiac fibroblasts by targeting PETN to enhance the phosphorylation of AKT, ERK and SMAD2/3[105]. Furthermore, HF patients with familial dilated cardiomyopathy also demonstrate late-stage cardiac fibrosis. A recent study using human iPSCs-derived cardiomyocytes as a disease model suggested that EVs derived from Ang II-stimulated DCM highly exhibit increased cardiac fibrosis and impaired cardiac function in vitro and in vivo compared to control cardiomyocytes via EV-enriched miRNA-218-5p targeting TNFAIP3 to activate TGF-β signaling [106]. Other than cardiomyocytes, Ang II-stimulated adipocytes can also secrete EV-enriched miRNA-23a-3p, which transforms cardiac fibroblasts into myofibroblasts and promotes collagen accumulation by targeting RAP1 (Ras-related protein 1) [107]. In addition, cardiac infiltration of CD4+ T cells is implicated in the healing process post-myocardial infarction (MI), contributing to cardiac fibrosis and dysfunction [121]. A mechanistic study further revealed that cardiac activated CD4+ T cells can transport excessive miR-142-3p via EVs into cardiac fibroblasts in which miR-143-3p activates the WNT signaling pathway by targeting APC (Adenomatous Polyposis Coli) to transform cardiac fibroblasts into activated myofibroblasts boosting post-ischemic ventricular remodeling in the progression of HF [108]. It has been well-documented that miRNA-21 is a central regulator of cardiac fibrosis in HF and shows therapeutic potential as a target for HF treatment [109-112]. Recently, using the TAC-induced HF model combined with single-cell sequencing, bioinformatics analyses revealed that EV-enriched miR-21 also determines macrophage-fibroblast cross-talk and promotes the transition from cardiac fibroblasts to activated myofibroblasts, leading to cardiac fibrosis [113].
3.3. EV-miRNA in Cardiac Angiogenesis during Heart Failure
In response to cardiac injury, intercellular communication is fundamental for maintaining homeostasis and integrity. The aforementioned studies have demonstrated that EV-enriched miRNAs derived from other types of cardiac cells including macrophages, T cells and adipocytes contribute to cardiac fibrosis and/or hypertrophy in HF. EVs secreted from cardiomyocytes also play an important role in anti-angiogenesis through miRNA transfer in HF. Recently, miRNA-200c-3p has been found to be one of the most enriched miRNAs in hypertrophic cardiomyocyte-derived EVs under pressure overload, and was transported into endothelial cells (ECs) in which it functions as a detrimental anti-angiogenic factor to impair endothelial function including proliferation, migration and tube formation [114]. It has also been shown that hypertrophic cardiomyocytes induced by Ang II also release miRNA-29a via EVs to inhibit the proliferation, migration and angiogenic ability of cardiac microvascular ECs [115]. Other than hypertrophic cardiomyocytes, activated cardiac fibroblasts can also secrete EVs abundant with miRNA-200a-3p, which mediates the cross-talk between cardiac endothelial cells and induces endothelial dysfunction by targeting the ETS1/VEGF-A signaling axis [116].
3.3. Cardiac EV miRNA-Mediated Inter-Organ Communication in Heart Failure
Increasing evidence suggests that heart-brain communication at the miRNA level contributes to neuronal dysfunction in the brain mediated by EVs in the HF state [49,117]. miRNA-1 is a cardiac miRNA abundantly expressed in the myocardium [102]. Circulating miRNA-1 was significantly increased in patients with acute myocardial infarction (AMI) [102,122]. Interestingly, cardiac-derived miRNA-1 can be transported from the infarcted heart into the hippocampus via EVs where cardiac miRNA-1 causes neuronal microtubular damage. This is independent of brain hypoperfusion induced by MI [117], suggesting a novel mechanism by which the damaged heart contributes to brain dysfunction. A study from our group also demonstrated that cardiac-derived miRNAs can be sorted into EVs and circulate into the rostral ventrolateral medulla (RVLM) of the brain stem where these miRNAs evoke sympathetic excitation by targeting Nrf2/ARE signaling to induce oxidative stress [49]. Moreover, it is well-documented that neutrophils promote the progression of acute MI by releasing ROS, granular components and extracellular traps to aggravate inflammation [123,124]. Clinical evidence also shows that the numbers of peripheral blood neutrophils and plasma EVs correlate closely with the extent of acute MI, including infarct size, mortality and HF development [48,125,126]. Interestingly, miRNA analysis of human plasma EVs from patients with AMI demonstrated twelve significantly enriched miRNAs, and two out of twelve, including miRNA-126-3p and -5p, were highly regulated and secreted by ECs post- acute MI and are responsible for cell adhesion and chemotaxis [118]. Importantly, recent studies from the same group revealed that cardiac ECs in response to cardiac injury rapidly release VCAM-1+ EVs containing miRNA-126, which preferentially mediate heart-spleen communication. These VCAM-1+ EVs rapidly and selectively recruit splenic neutrophils to peripheral blood following myocardial injury. Mechanistic studies further suggest that VCAM-1+ EVs-enriched miRNA-126 induce the transcriptional activation of neutrophils in the spleen before they arrive at the ischemic myocardium, contributing to local inflammation and chemokine production [48,119]. In addition, genetic deletion of VCAM-1 from EVs by the CRISPR-Cas9 system or silencing miRNA-126 in vivo by antagomir significantly blocked the mobilization of splenic neutrophils to ischemic myocardium and reduced myocardial infarct size in a LAD ligation mouse model, suggesting that specific surface proteins such as VCAM-1 in EVs will determine their organotropism, and functionalities of EVs.
4. Extracellular Vesicle miRNA-Based Prognosis, Diagnosis and Therapeutics of Heart Failure
Although natriuretic peptides and cardiac troponins are currently the most widely employed biomarkers for the prognosis and diagnosis of HF, and next-generation biomarkers including soluble source of tumorigenicity 2 (sST2), Proenkephalin, Growth differentiation factor-15 (GDF-15) and Galectin-3 (Gal-3) have also been well-established as promising biomarkers for HF diagnosis and prognosis [127,128], the discovery of altered levels of circulating miRNAs in patients with HF provide the possibilities to utilize circulating miRNAs, in particular, EV-enriched miRNAs as biomarkers for HF [63,64,129]. Recently, circulating EV-miRNAs have been emerging as non-invasive prognostic and diagnostic biomarkers for HF, and their potential clinical applications have been well-summarized [130]. For example, some circulating EV-miRNAs, including miR-92-5p, miR-146a, miR-181c and miR-495, showed diagnostic potential for HF, and other EV-enriched miRNAs such as miR-192, -194, -34a, -425 and miR-744 represent promising prognostic biomarkers for HF. Moreover, two circulating EV-enriched miRNAs associated with HF including, miR-30d-5p and miR-126a-5p, have shown promising potential as biomarkers for HFpEF (Heart Failure with preserved Ejection Fraction) in diabetes mellitus because their down-regulations in circulating EVs and left ventricle remain consistently correlated with decreased cardiac output [131]. In addition, circulating miR-30d, especially EV-miR-30d, has shown high potential as a biomarker to evaluate left ventricular remodeling and clinical outcome for patients with HF [132,133].
Other than serving as prognostic and diagnostic biomarkers, EV-miRNAs have been used as therapeutic targets for HF. Some miRNAs such as miR-126, 146a, miR-125a-5p, miR-125b-5p, miR-29b, miR-98-5p, miR-30e and miR-30d have shown protective effects on cardiac function. Extracellular vesicle-based delivery of these miRNAs shows promising clinical applications in HF therapy [134-140]. A recent study showed the beneficial effects of adipose tissue-derived mesenchymal stem cell (ADSC)-derived EVs on acute MI-induced cardiac injury by EV-enriched miR-205, suggesting a promising therapeutic potential [141]. Alternatively, targeting injured cardiac-secreted EVs by exogenously pre-loading miRNA inhibitors (i.e. antagomirs) into EVs, such as miR-27a, miR-28 and miR-34a [49] as well as miR-126 [48] may represent another therapeutic strategy.
5. Perspectives and Future Directions
Extracellular vesicle miRNAs are attractive candidates as diagnostic and prognostic biomarkers as well as therapeutic targets. However, using EV-miRNAs for these purposes still face great challenges in becoming well-accepted biomarkers for prognosis and diagnosis of HF. First, rigorous standards for performing EV research are imperative and are time-consuming processes, including EV categorization, purification and separation technology development and characterization. Uniform standard guidelines are urgently needed in the field. Second, the pathogenesis of HF is complicated and cardiac function is also influenced by systemic communication with other organs. Selective criteria are needed to evaluate the potential of EV-miRNAs for use as diagnostic and prognostic biomarkers for HF. In addition, the cell origins and categories of circulating EVs are heterogenous and the exploration of cell origin and sorting mechanisms of circulating EV-miRNAs will be of interest to better understand the pathophysiological functions of circulating EV-miRNAs in HF. Third, the sample size used for evaluating circulating EV-miRNAs as biomarkers for HF is still relatively small, which is insufficient to support their use as sensitive biomarkers at this time.
Although stem cell-derived EV-miRNAs have shown promising therapeutic potentials for HF management, it remains questionable concerning the beneficial contributions of other EV components, such as proteins, lipids and various metabolites to cardiac functions in HF treatment. Moreover, an interesting new therapeutic direction relates to the effects of stem cell-derived EVs in HF by engineering EVs with heart homing peptides and genetically also used therapeutically for the delivery of miRNA-21 to effectively restore cardiac function after MI [142]. For example, hypoxia-conditioned BM-MSCs (Bone Marrow Mesenchymal Stem Cells) secreted EVs abundant with miRNA-125b-5p, a cardioprotective miRNA, demonstrated high specificity to the ischemic myocardium in a mouse acute MI model when miR-125-enriched EVs were conjugated with an ischemic myocardium-targeted peptide and then administered by intravenous injection [134]. A cardiac homing peptide-guided delivery system has been used for the treatment of MI [143,144]. Given cardiac-derived EVs also contribute to neuronal dysfunction in the brain in HF [49,117], engineered EVs derived from dendritic cells expressing Lamp2b (an exosomal membrane protein), fused to the neuron-specific RVG peptide [145], may be therapeutically used for HF management. In addition, the next generation of “Hybrid EVs” [146] produced by stem cell-derived EVs fused with modified liposomes may represent an innovative therapeutic strategy for HF treatment. The resulting hybrid EVs will not only possess the intrinsic immunomodulatory effects and blood barrier penetrating ability of original stem cell-derived EVs but also highly enrich some cardioprotective miRNAs by pre-loading them into parental liposomes. Importantly, the parental partners can be modified by either engineering them with organotropic peptides or integrating multimodal imaging capability for clinical tracking, guiding and optimizing. Taken together, miRNAs are differentially regulated in response to heart injury and selectively sorted into EVs, contributing to the pathogenesis of HF via intercellular and/or inter-organ communications, which make it possible to become a promising type of prognostic and diagnostic biomarker and therapeutic target for HF.
Author Contributions
C.T. and I.H.Z conceived of the subject matter of this review. C.T summarized the published results and drafted the manuscript. C.T., J.N.Z and I.H.Z revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Institution of Health Grant R01HL153176 to IHZ/CT and P01 HL62222 to IHZ; American Heart Association (AHA) Career Development Award (19CDA34520004) to CT, and University of Kentucky start-up funding to C.T.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Roger, V.L. Epidemiology of Heart Failure: A Contemporary Perspective. Circ Res 2021, 128, 1421–1434. [Google Scholar] [CrossRef]
- Wollert, K.C.; Drexler, H. The renin–angiotensin system and experimental heart failure. Cardiovascular Research 1999, 43, 838–849. [Google Scholar] [CrossRef]
- Verbrugge, F.H.; Tang, W.H.W.; Mullens, W. Renin-Angiotensin-Aldosterone System Activation During Decongestion in Acute Heart Failure. JACC: Heart Failure 2015, 3, 108–111. [Google Scholar] [CrossRef]
- Jia, G.; Aroor, A.R.; Hill, M.A.; Sowers, J.R. Role of Renin-Angiotensin-Aldosterone System Activation in Promoting Cardiovascular Fibrosis and Stiffness. Hypertension 2018, 72, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Florea, V.G.; Cohn, J.N. The autonomic nervous system and heart failure. Circ Res 2014, 114, 1815–1826. [Google Scholar] [CrossRef]
- Kishi, T. Heart failure as an autonomic nervous system dysfunction. Journal of Cardiology 2012, 59, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Anderson, A.S. The sympathetic nervous system and heart failure. Cardiol Clin 2014, 32, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Booth, L.C.; May, C.N.; Yao, S.T. The role of the renal afferent and efferent nerve fibers in heart failure. Front Physiol 2015, 6, 270. [Google Scholar] [CrossRef] [PubMed]
- van Rooij, E.; Sutherland, L.B.; Liu, N.; Williams, A.H.; McAnally, J.; Gerard, R.D.; Richardson, J.A.; Olson, E.N. A signature pattern of stress-responsive microRNAs that can evoke cardiac hypertrophy and heart failure. Proc Natl Acad Sci U S A 2006, 103, 18255–18260. [Google Scholar] [CrossRef] [PubMed]
- van Rooij, E.; Sutherland, L.B.; Qi, X.; Richardson, J.A.; Hill, J.; Olson, E.N. Control of stress-dependent cardiac growth and gene expression by a microRNA. Science 2007, 316, 575–579. [Google Scholar] [CrossRef]
- Younger, S.T.; Corey, D.R. Transcriptional gene silencing in mammalian cells by miRNA mimics that target gene promoters. Nucleic Acids Res 2011, 39, 5682–5691. [Google Scholar] [CrossRef]
- Lytle, J.R.; Yario, T.A.; Steitz, J.A. Target mRNAs are repressed as efficiently by microRNA-binding sites in the 5’ UTR as in the 3’ UTR. Proc Natl Acad Sci U S A 2007, 104, 9667–9672. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Zhang, J.; Thomson, A.M.; Lim, B.; Rigoutsos, I. MicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiation. Nature 2008, 455, 1124–1128. [Google Scholar] [CrossRef]
- Gu, W.; Xu, Y.; Xie, X.; Wang, T.; Ko, J.H.; Zhou, T. The role of RNA structure at 5’ untranslated region in microRNA-mediated gene regulation. Rna 2014, 20, 1369–1375. [Google Scholar] [CrossRef]
- Kumarswamy, R.; Thum, T. Non-coding RNAs in cardiac remodeling and heart failure. Circ Res 2013, 113, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Melman, Y.F.; Shah, R.; Das, S. MicroRNAs in heart failure: is the picture becoming less miRky? Circ Heart Fail 2014, 7, 203–214. [Google Scholar] [CrossRef]
- Gholaminejad, A.; Zare, N.; Dana, N.; Shafie, D.; Mani, A.; Javanmard, S.H. A meta-analysis of microRNA expression profiling studies in heart failure. Heart Fail Rev 2021, 26, 997–1021. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.N.; Wang, J.L.; Fu, Y.P. The microRNA Expression Profiling in Heart Failure: A Systematic Review and Meta-Analysis. Front Cardiovasc Med 2022, 9, 856358. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martin, R.; Wang, G.; Brandão, B.B.; Zanotto, T.M.; Shah, S.; Kumar Patel, S.; Schilling, B.; Kahn, C.R. MicroRNA sequence codes for small extracellular vesicle release and cellular retention. Nature 2022, 601, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Koppers-Lalic, D.; Hackenberg, M.; Bijnsdorp, I.V.; van Eijndhoven, M.A.J.; Sadek, P.; Sie, D.; Zini, N.; Middeldorp, J.M.; Ylstra, B.; de Menezes, R.X.; et al. Nontemplated nucleotide additions distinguish the small RNA composition in cells from exosomes. Cell Rep 2014, 8, 1649–1658. [Google Scholar] [CrossRef]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and intercellular transfer of microRNAs in living cells. J Biol Chem 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nature Reviews Molecular Cell Biology 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Takagi, M.; Ono, S.; Kumaki, T.; Nishimura, N.; Murakami, H.; Enomoto, Y.; Naruto, T.; Ueda, H.; Kurosawa, K. Complex congenital cardiovascular anomaly in a patient with AGO1-associated disorder. Am J Med Genet A 2023, 191, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Ranjan, P.; Suleiman, Z.G.; Goswami, S.K.; Li, J.; Prasad, R.; Verma, S.K. mRNA modifications in cardiovascular biology and disease: with a focus on m6A modification. Cardiovasc Res 2022, 118, 1680–1692. [Google Scholar] [CrossRef]
- Zhu, M.; Gao, J.; Lin, X.J.; Gong, Y.Y.; Qi, Y.C.; Ma, Y.L.; Song, Y.X.; Tan, W.; Li, F.Y.; Ye, M.; et al. Novel roles of an intragenic G-quadruplex in controlling microRNA expression and cardiac function. Nucleic Acids Res 2021, 49, 2522–2536. [Google Scholar] [CrossRef]
- da Costa Martins, P.A.; Bourajjaj, M.; Gladka, M.; Kortland, M.; van Oort, R.J.; Pinto, Y.M.; Molkentin, J.D.; De Windt, L.J. Conditional dicer gene deletion in the postnatal myocardium provokes spontaneous cardiac remodeling. Circulation 2008, 118, 1567–1576. [Google Scholar] [CrossRef]
- Varela, M.A.; Roberts, T.C.; Wood, M.J. Epigenetics and ncRNAs in brain function and disease: mechanisms and prospects for therapy. Neurotherapeutics 2013, 10, 621–631. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. Embo j 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat Rev Genet 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Davis-Dusenbery, B.N.; Hata, A. Mechanisms of control of microRNA biogenesis. J Biochem 2010, 148, 381–392. [Google Scholar] [CrossRef]
- Lee, Y.; Jeon, K.; Lee, J.T.; Kim, S.; Kim, V.N. MicroRNA maturation: stepwise processing and subcellular localization. Embo j 2002, 21, 4663–4670. [Google Scholar] [CrossRef] [PubMed]
- Gregory, R.I.; Yan, K.P.; Amuthan, G.; Chendrimada, T.; Doratotaj, B.; Cooch, N.; Shiekhattar, R. The Microprocessor complex mediates the genesis of microRNAs. Nature 2004, 432, 235–240. [Google Scholar] [CrossRef]
- Lund, E.; Güttinger, S.; Calado, A.; Dahlberg, J.E.; Kutay, U. Nuclear export of microRNA precursors. Science 2004, 303, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Heo, I.; Tian, Y.; Simanshu, D.K.; Chang, H.; Jee, D.; Patel, D.J.; Kim, V.N. Dicer recognizes the 5’ end of RNA for efficient and accurate processing. Nature 2011, 475, 201–205. [Google Scholar] [CrossRef]
- Benoit, M.P.; Imbert, L.; Palencia, A.; Pérard, J.; Ebel, C.; Boisbouvier, J.; Plevin, M.J. The RNA-binding region of human TRBP interacts with microRNA precursors through two independent domains. Nucleic Acids Res 2013, 41, 4241–4252. [Google Scholar] [CrossRef]
- Mourelatos, Z.; Dostie, J.; Paushkin, S.; Sharma, A.; Charroux, B.; Abel, L.; Rappsilber, J.; Mann, M.; Dreyfuss, G. miRNPs: a novel class of ribonucleoproteins containing numerous microRNAs. Genes Dev 2002, 16, 720–728. [Google Scholar] [CrossRef] [PubMed]
- Kawamata, T.; Tomari, Y. Making RISC. Trends Biochem Sci 2010, 35, 368–376. [Google Scholar] [CrossRef]
- Ni, W.J.; Leng, X.M. miRNA-Dependent Activation of mRNA Translation. Microrna 2016, 5, 83–86. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Frontiers in Endocrinology 2018, 9. [Google Scholar] [CrossRef]
- Lee, S.; Vasudevan, S. Post-transcriptional stimulation of gene expression by microRNAs. Adv Exp Med Biol 2013, 768, 97–126. [Google Scholar] [CrossRef]
- Bang, C.; Batkai, S.; Dangwal, S.; Gupta, S.K.; Foinquinos, A.; Holzmann, A.; Just, A.; Remke, J.; Zimmer, K.; Zeug, A.; et al. Cardiac fibroblast-derived microRNA passenger strand-enriched exosomes mediate cardiomyocyte hypertrophy. J Clin Invest 2014, 124, 2136–2146. [Google Scholar] [CrossRef]
- Tian, C.; Hu, G.; Gao, L.; Hackfort, B.T.; Zucker, I.H. Extracellular vesicular MicroRNA-27a* contributes to cardiac hypertrophy in chronic heart failure. J Mol Cell Cardiol 2020, 143, 120–131. [Google Scholar] [CrossRef]
- Ruby, J.G.; Jan, C.H.; Bartel, D.P. Intronic microRNA precursors that bypass Drosha processing. Nature 2007, 448, 83–86. [Google Scholar] [CrossRef]
- Xie, M.; Li, M.; Vilborg, A.; Lee, N.; Shu, M.D.; Yartseva, V.; Šestan, N.; Steitz, J.A. Mammalian 5’-capped microRNA precursors that generate a single microRNA. Cell 2013, 155, 1568–1580. [Google Scholar] [CrossRef] [PubMed]
- Cheloufi, S.; Dos Santos, C.O.; Chong, M.M.; Hannon, G.J. A dicer-independent miRNA biogenesis pathway that requires Ago catalysis. Nature 2010, 465, 584–589. [Google Scholar] [CrossRef]
- Michel, L.Y.M. Extracellular Vesicles in Adipose Tissue Communication with the Healthy and Pathological Heart. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Yuan, Y.; Mei, Z.; Qu, Z.; Li, G.; Yu, S.; Liu, Y.; Liu, K.; Shen, Z.; Pu, J.; Wang, Y.; et al. Exosomes secreted from cardiomyocytes suppress the sensitivity of tumor ferroptosis in ischemic heart failure. Signal Transduct Target Ther 2023, 8, 121. [Google Scholar] [CrossRef]
- Akbar, N.; Braithwaite, A.T.; Corr, E.M.; Koelwyn, G.J.; van Solingen, C.; Cochain, C.; Saliba, A.E.; Corbin, A.; Pezzolla, D.; Møller Jørgensen, M.; et al. Rapid neutrophil mobilization by VCAM-1+ endothelial cell-derived extracellular vesicles. Cardiovasc Res 2023, 119, 236–251. [Google Scholar] [CrossRef]
- Tian, C.; Gao, L.; Rudebush, T.L.; Yu, L.; Zucker, I.H. Extracellular Vesicles Regulate Sympatho-Excitation by Nrf2 in Heart Failure. Circ Res 2022, 131, 687–700. [Google Scholar] [CrossRef]
- Rädler, J.; Gupta, D.; Zickler, A.; Andaloussi, S.E. Exploiting the biogenesis of extracellular vesicles for bioengineering and therapeutic cargo loading. Mol Ther 2023, 31, 1231–1250. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367. [Google Scholar] [CrossRef]
- Nabhan, J.F.; Hu, R.; Oh, R.S.; Cohen, S.N.; Lu, Q. Formation and release of arrestin domain-containing protein 1-mediated microvesicles (ARMMs) at plasma membrane by recruitment of TSG101 protein. Proc Natl Acad Sci U S A 2012, 109, 4146–4151. [Google Scholar] [CrossRef]
- Clancy, J.W.; Schmidtmann, M.; D’Souza-Schorey, C. The ins and outs of microvesicles. FASEB Bioadv 2021, 3, 399–406. [Google Scholar] [CrossRef]
- Segawa, K.; Nagata, S. An Apoptotic ’Eat Me’ Signal: Phosphatidylserine Exposure. Trends Cell Biol 2015, 25, 639–650. [Google Scholar] [CrossRef]
- Ozkocak, D.C.; Phan, T.K.; Poon, I.K.H. Translating extracellular vesicle packaging into therapeutic applications. Front Immunol 2022, 13, 946422. [Google Scholar] [CrossRef]
- Atkin-Smith, G.K.; Tixeira, R.; Paone, S.; Mathivanan, S.; Collins, C.; Liem, M.; Goodall, K.J.; Ravichandran, K.S.; Hulett, M.D.; Poon, I.K. A novel mechanism of generating extracellular vesicles during apoptosis via a beads-on-a-string membrane structure. Nat Commun 2015, 6, 7439. [Google Scholar] [CrossRef]
- Holmgren, L.; Szeles, A.; Rajnavölgyi, E.; Folkman, J.; Klein, G.; Ernberg, I.; Falk, K.I. Horizontal transfer of DNA by the uptake of apoptotic bodies. Blood 1999, 93, 3956–3963. [Google Scholar] [CrossRef] [PubMed]
- Bergsmedh, A.; Szeles, A.; Henriksson, M.; Bratt, A.; Folkman, M.J.; Spetz, A.L.; Holmgren, L. Horizontal transfer of oncogenes by uptake of apoptotic bodies. Proc Natl Acad Sci U S A 2001, 98, 6407–6411. [Google Scholar] [CrossRef]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nature Cell Biology 2011, 13, 423–433. [Google Scholar] [CrossRef]
- Linhares-Lacerda, L.; Temerozo, J.R.; Ribeiro-Alves, M.; Azevedo, E.P.; Mojoli, A.; Nascimento, M.T.C.; Silva-Oliveira, G.; Savino, W.; Foguel, D.; Bou-Habib, D.C.; et al. Neutrophil extracellular trap-enriched supernatants carry microRNAs able to modulate TNF-α production by macrophages. Scientific Reports 2020, 10, 2715. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Ansel, K.M.; Bitzer, M.; Breakefield, X.O.; Charest, A.; Galas, D.J.; Gerstein, M.B.; Gupta, M.; Milosavljevic, A.; McManus, M.T.; et al. The Extracellular RNA Communication Consortium: Establishing Foundational Knowledge and Technologies for Extracellular RNA Research. Cell 2019, 177, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Tijsen, A.J.; Creemers, E.E.; Moerland, P.D.; de Windt, L.J.; van der Wal, A.C.; Kok, W.E.; Pinto, Y.M. MiR423-5p as a circulating biomarker for heart failure. Circ Res 2010, 106, 1035–1039. [Google Scholar] [CrossRef]
- Jansen, F.; Yang, X.; Proebsting, S.; Hoelscher, M.; Przybilla, D.; Baumann, K.; Schmitz, T.; Dolf, A.; Endl, E.; Franklin, B.S.; et al. MicroRNA expression in circulating microvesicles predicts cardiovascular events in patients with coronary artery disease. J Am Heart Assoc 2014, 3, e001249. [Google Scholar] [CrossRef]
- Chang, Y.J.; Wang, K.C. Therapeutic perspectives of extracellular vesicles and extracellular microRNAs in atherosclerosis. Curr Top Membr 2021, 87, 255–277. [Google Scholar] [CrossRef]
- Villarroya-Beltri, C.; Gutiérrez-Vázquez, C.; Sánchez-Cabo, F.; Pérez-Hernández, D.; Vázquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sánchez-Madrid, F. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat Commun 2013, 4, 2980. [Google Scholar] [CrossRef] [PubMed]
- Zietzer, A.; Hosen, M.R.; Wang, H.; Goody, P.R.; Sylvester, M.; Latz, E.; Nickenig, G.; Werner, N.; Jansen, F. The RNA-binding protein hnRNPU regulates the sorting of microRNA-30c-5p into large extracellular vesicles. J Extracell Vesicles 2020, 9, 1786967. [Google Scholar] [CrossRef] [PubMed]
- Shurtleff, M.J.; Yao, J.; Qin, Y.; Nottingham, R.M.; Temoche-Diaz, M.M.; Schekman, R.; Lambowitz, A.M. Broad role for YBX1 in defining the small noncoding RNA composition of exosomes. Proc Natl Acad Sci U S A 2017, 114, E8987–e8995. [Google Scholar] [CrossRef]
- Shurtleff, M.J.; Temoche-Diaz, M.M.; Karfilis, K.V.; Ri, S.; Schekman, R. Y-box protein 1 is required to sort microRNAs into exosomes in cells and in a cell-free reaction. Elife 2016, 5. [Google Scholar] [CrossRef]
- Santangelo, L.; Giurato, G.; Cicchini, C.; Montaldo, C.; Mancone, C.; Tarallo, R.; Battistelli, C.; Alonzi, T.; Weisz, A.; Tripodi, M. The RNA-Binding Protein SYNCRIP Is a Component of the Hepatocyte Exosomal Machinery Controlling MicroRNA Sorting. Cell Rep 2016, 17, 799–808. [Google Scholar] [CrossRef]
- Hobor, F.; Dallmann, A.; Ball, N.J.; Cicchini, C.; Battistelli, C.; Ogrodowicz, R.W.; Christodoulou, E.; Martin, S.R.; Castello, A.; Tripodi, M.; et al. A cryptic RNA-binding domain mediates Syncrip recognition and exosomal partitioning of miRNA targets. Nat Commun 2018, 9, 831. [Google Scholar] [CrossRef]
- Statello, L.; Maugeri, M.; Garre, E.; Nawaz, M.; Wahlgren, J.; Papadimitriou, A.; Lundqvist, C.; Lindfors, L.; Collén, A.; Sunnerhagen, P.; et al. Identification of RNA-binding proteins in exosomes capable of interacting with different types of RNA: RBP-facilitated transport of RNAs into exosomes. PLoS One 2018, 13, e0195969. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, K.; Ghoshal, B.; Ghosh, S.; Chakrabarty, Y.; Shwetha, S.; Das, S.; Bhattacharyya, S.N. Reversible HuR-microRNA binding controls extracellular export of miR-122 and augments stress response. EMBO Rep 2016, 17, 1184–1203. [Google Scholar] [CrossRef]
- Teng, Y.; Ren, Y.; Hu, X.; Mu, J.; Samykutty, A.; Zhuang, X.; Deng, Z.; Kumar, A.; Zhang, L.; Merchant, M.L.; et al. MVP-mediated exosomal sorting of miR-193a promotes colon cancer progression. Nat Commun 2017, 8, 14448. [Google Scholar] [CrossRef]
- Xu, Y.F.; Xu, X.; Gin, A.; Nshimiyimana, J.D.; Mooers, B.H.M.; Caputi, M.; Hannafon, B.N.; Ding, W.Q. SRSF1 regulates exosome microRNA enrichment in human cancer cells. Cell Commun Signal 2020, 18, 130. [Google Scholar] [CrossRef] [PubMed]
- Temoche-Diaz, M.M.; Shurtleff, M.J.; Nottingham, R.M.; Yao, J.; Fadadu, R.P.; Lambowitz, A.M.; Schekman, R. Distinct mechanisms of microRNA sorting into cancer cell-derived extracellular vesicle subtypes. Elife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.M.; Ma, L.; Schekman, R. Selective sorting of microRNAs into exosomes by phase-separated YBX1 condensates. Elife 2021, 10. [Google Scholar] [CrossRef]
- Lee, H.; Li, C.; Zhang, Y.; Zhang, D.; Otterbein, L.E.; Jin, Y. Caveolin-1 selectively regulates microRNA sorting into microvesicles after noxious stimuli. J Exp Med 2019, 216, 2202–2220. [Google Scholar] [CrossRef]
- Squadrito, M.L.; Baer, C.; Burdet, F.; Maderna, C.; Gilfillan, G.D.; Lyle, R.; Ibberson, M.; De Palma, M. Endogenous RNAs modulate microRNA sorting to exosomes and transfer to acceptor cells. Cell Rep 2014, 8, 1432–1446. [Google Scholar] [CrossRef]
- Iavello, A.; Frech, V.S.; Gai, C.; Deregibus, M.C.; Quesenberry, P.J.; Camussi, G. Role of Alix in miRNA packaging during extracellular vesicle biogenesis. Int J Mol Med 2016, 37, 958–966. [Google Scholar] [CrossRef]
- Liu, Y.; Song, J.W.; Lin, J.Y.; Miao, R.; Zhong, J.C. Roles of MicroRNA-122 in Cardiovascular Fibrosis and Related Diseases. Cardiovasc Toxicol 2020, 20, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Hosen, M.R.; Goody, P.R.; Zietzer, A.; Xiang, X.; Niepmann, S.T.; Sedaghat, A.; Tiyerili, V.; Chennupati, R.; Moore, J.B.t.; Boon, R.A.; et al. Circulating MicroRNA-122-5p Is Associated With a Lack of Improvement in Left Ventricular Function After Transcatheter Aortic Valve Replacement and Regulates Viability of Cardiomyocytes Through Extracellular Vesicles. Circulation 2022, 146, 1836–1854. [Google Scholar] [CrossRef]
- Liu, X.; Xu, Y.; Deng, Y.; Li, H. MicroRNA-223 Regulates Cardiac Fibrosis After Myocardial Infarction by Targeting RASA1. Cell Physiol Biochem 2018, 46, 1439–1454. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Long, B.; Liu, F.; Wang, J.X.; Liu, C.Y.; Zhao, B.; Zhou, L.Y.; Sun, T.; Wang, M.; Yu, T.; et al. A circular RNA protects the heart from pathological hypertrophy and heart failure by targeting miR-223. Eur Heart J 2016, 37, 2602–2611. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, Y.; Li, M.; Yuan, J.; Yu, Y.; Bi, X.; Hong, H.; Ye, J.; Liu, P. MicroRNA-34c-5p provokes isoprenaline-induced cardiac hypertrophy by modulating autophagy via targeting ATG4B. Acta Pharm Sin B 2022, 12, 2374–2390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, Q.; Feng, Y.; Chen, X.; Yang, L.; Xu, M.; Wang, X.; Li, W.; Niu, X.; Gao, D. MicroRNA-26a Protects the Heart Against Hypertension-Induced Myocardial Fibrosis. J Am Heart Assoc 2020, 9, e017970. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Guo, M.; Ma, G.; Zhang, N.; Pan, F.; Fan, X.; Wang, R. MicroRNA-30c-5p protects against myocardial ischemia/reperfusion injury via regulation of Bach1/Nrf2. Toxicol Appl Pharmacol 2021, 426, 115637. [Google Scholar] [CrossRef]
- Danielson, L.S.; Park, D.S.; Rotllan, N.; Chamorro-Jorganes, A.; Guijarro, M.V.; Fernandez-Hernando, C.; Fishman, G.I.; Phoon, C.K.; Hernando, E. Cardiovascular dysregulation of miR-17-92 causes a lethal hypertrophic cardiomyopathy and arrhythmogenesis. Faseb j 2013, 27, 1460–1467. [Google Scholar] [CrossRef]
- Nair, N.; Kumar, S.; Gongora, E.; Gupta, S. Circulating miRNA as novel markers for diastolic dysfunction. Mol Cell Biochem 2013, 376, 33–40. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, L.; Li, Z.; Fu, G. miR-1231 exacerbates arrhythmia by targeting calciumchannel gene CACNA2D2 in myocardial infarction. Am J Transl Res 2017, 9, 1822–1833. [Google Scholar]
- Tucker, N.R.; Chaffin, M.; Fleming, S.J.; Hall, A.W.; Parsons, V.A.; Bedi, K.C., Jr.; Akkad, A.D.; Herndon, C.N.; Arduini, A.; Papangeli, I.; et al. Transcriptional and Cellular Diversity of the Human Heart. Circulation 2020, 142, 466–482. [Google Scholar] [CrossRef] [PubMed]
- Litviňuková, M.; Talavera-López, C.; Maatz, H.; Reichart, D.; Worth, C.L.; Lindberg, E.L.; Kanda, M.; Polanski, K.; Heinig, M.; Lee, M.; et al. Cells of the adult human heart. Nature 2020, 588, 466–472. [Google Scholar] [CrossRef]
- Howard, C.M.; Baudino, T.A. Dynamic cell-cell and cell-ECM interactions in the heart. J Mol Cell Cardiol 2014, 70, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.P.; Zhang, Y.S.; Xu, X.; Zhou, Q.; Li, J.D.; Yan, C. Vinpocetine Attenuates Pathological Cardiac Remodeling by Inhibiting Cardiac Hypertrophy and Fibrosis. Cardiovasc Drugs Ther 2017, 31, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Travers, J.G.; Kamal, F.A.; Robbins, J.; Yutzey, K.E.; Blaxall, B.C. Cardiac Fibrosis: The Fibroblast Awakens. Circ Res 2016, 118, 1021–1040. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.G.; Lee, P.; Gordon, R.E.; Sahoo, S.; Kho, C.; Jeong, D. Analysis of extracellular vesicle miRNA profiles in heart failure. J Cell Mol Med 2020, 24, 7214–7227. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Wang, C.; Mang, G.; Xu, X.; Fu, S.; Chen, J.; Wang, X.; Wang, W.; Li, H.; Zhao, P.; et al. Extracellular vesicle-packaged mitochondrial disturbing miRNA exacerbates cardiac injury during acute myocardial infarction. Clin Transl Med 2022, 12, e779. [Google Scholar] [CrossRef]
- Das, S.; Halushka, M.K. Extracellular vesicle microRNA transfer in cardiovascular disease. Cardiovasc Pathol 2015, 24, 199–206. [Google Scholar] [CrossRef]
- Tian, C.; Gao, L.; Zimmerman, M.C.; Zucker, I.H. Myocardial infarction-induced microRNA-enriched exosomes contribute to cardiac Nrf2 dysregulation in chronic heart failure. Am J Physiol Heart Circ Physiol 2018, 314, H928–h939. [Google Scholar] [CrossRef]
- Fang, X.; Stroud, M.J.; Ouyang, K.; Fang, L.; Zhang, J.; Dalton, N.D.; Gu, Y.; Wu, T.; Peterson, K.L.; Huang, H.D.; et al. Adipocyte-specific loss of PPARγ attenuates cardiac hypertrophy. JCI Insight 2016, 1, e89908. [Google Scholar] [CrossRef]
- Callis, T.E.; Pandya, K.; Seok, H.Y.; Tang, R.H.; Tatsuguchi, M.; Huang, Z.P.; Chen, J.F.; Deng, Z.; Gunn, B.; Shumate, J.; et al. MicroRNA-208a is a regulator of cardiac hypertrophy and conduction in mice. J Clin Invest 2009, 119, 2772–2786. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. Cardiac-specific miRNA in cardiogenesis, heart function, and cardiac pathology (with focus on myocardial infarction). J Mol Cell Cardiol 2016, 94, 107–121. [Google Scholar] [CrossRef]
- Nie, X.; Fan, J.; Li, H.; Yin, Z.; Zhao, Y.; Dai, B.; Dong, N.; Chen, C.; Wang, D.W. miR-217 Promotes Cardiac Hypertrophy and Dysfunction by Targeting PTEN. Mol Ther Nucleic Acids 2018, 12, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, X.; Xue, F.; Li, Y.; Liu, W.; Zhang, S. Exosomes derived from cardiomyocytes promote cardiac fibrosis via myocyte-fibroblast cross-talk. Am J Transl Res 2018, 10, 4350–4366. [Google Scholar] [PubMed]
- Tang, C.; Hou, Y.X.; Shi, P.X.; Zhu, C.H.; Lu, X.; Wang, X.L.; Que, L.L.; Zhu, G.Q.; Liu, L.; Chen, Q.; et al. Cardiomyocyte-specific Peli1 contributes to the pressure overload-induced cardiac fibrosis through miR-494-3p-dependent exosomal communication. Faseb j 2023, 37, e22699. [Google Scholar] [CrossRef]
- Fu, X.; Mishra, R.; Chen, L.; Arfat, M.Y.; Sharma, S.; Kingsbury, T.; Gunasekaran, M.; Saha, P.; Hong, C.; Yang, P.; et al. Exosomes mediated fibrogenesis in dilated cardiomyopathy through a MicroRNA pathway. iScience 2023, 26, 105963. [Google Scholar] [CrossRef]
- Su, M.; Li, W.; Yuan, Y.U.E.; Liu, S.; Liang, C.; Liu, H.E.; Zhang, R.; Liu, Y.; Sun, L.I.; Wei, Y.; et al. Epididymal white adipose tissue promotes angiotensin II-induced cardiac fibrosis in an exosome-dependent manner. Translational Research 2022, 248, 51–67. [Google Scholar] [CrossRef]
- Cai, L.; Chao, G.; Li, W.; Zhu, J.; Li, F.; Qi, B.; Wei, Y.; Chen, S.; Zhou, G.; Lu, X.; et al. Activated CD4(+) T cells-derived exosomal miR-142-3p boosts post-ischemic ventricular remodeling by activating myofibroblast. Aging (Albany NY) 2020, 12, 7380–7396. [Google Scholar] [CrossRef]
- Hinkel, R.; Ramanujam, D.; Kaczmarek, V.; Howe, A.; Klett, K.; Beck, C.; Dueck, A.; Thum, T.; Laugwitz, K.L.; Maegdefessel, L.; et al. AntimiR-21 Prevents Myocardial Dysfunction in a Pig Model of Ischemia/Reperfusion Injury. J Am Coll Cardiol 2020, 75, 1788–1800. [Google Scholar] [CrossRef]
- Ramanujam, D.; Sassi, Y.; Laggerbauer, B.; Engelhardt, S. Viral Vector-Based Targeting of miR-21 in Cardiac Nonmyocyte Cells Reduces Pathologic Remodeling of the Heart. Mol Ther 2016, 24, 1939–1948. [Google Scholar] [CrossRef]
- Thum, T.; Chau, N.; Bhat, B.; Gupta, S.K.; Linsley, P.S.; Bauersachs, J.; Engelhardt, S. Comparison of different miR-21 inhibitor chemistries in a cardiac disease model. J Clin Invest 2011, 121, 461–462. [Google Scholar] [CrossRef]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Ramanujam, D.; Schön, A.P.; Beck, C.; Vaccarello, P.; Felician, G.; Dueck, A.; Esfandyari, D.; Meister, G.; Meitinger, T.; Schulz, C.; et al. MicroRNA-21-Dependent Macrophage-to-Fibroblast Signaling Determines the Cardiac Response to Pressure Overload. Circulation 2021, 143, 1513–1525. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, L.; Juni, R.P.; de Abreu, R.C.; Sansonetti, M.; Sampaio-Pinto, V.; Halkein, J.; Hegenbarth, J.C.; Ring, N.; Knoops, K.; Kocken, J.M.M.; et al. Intercellular transfer of miR-200c-3p impairs the angiogenic capacity of cardiac endothelial cells. Mol Ther 2022, 30, 2257–2273. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Qiu, Z.; Li, C.; Zhao, R.; Zhang, Y.; Shen, C.; Liu, W.; Long, X.; Zhuang, S.; Wang, Y.; et al. Exosomal MiR-29a in Cardiomyocytes Induced by Angiotensin II Regulates Cardiac Microvascular Endothelial Cell Proliferation, Migration and Angiogenesis by Targeting VEGFA. Current Gene Therapy 2022, 22, 331–341. [Google Scholar] [CrossRef]
- Ranjan, P.; Kumari, R.; Goswami, S.K.; Li, J.; Pal, H.; Suleiman, Z.; Cheng, Z.; Krishnamurthy, P.; Kishore, R.; Verma, S.K. Myofibroblast-Derived Exosome Induce Cardiac Endothelial Cell Dysfunction. Front Cardiovasc Med 2021, 8, 676267. [Google Scholar] [CrossRef]
- Sun, L.L.; Duan, M.J.; Ma, J.C.; Xu, L.; Mao, M.; Biddyut, D.; Wang, Q.; Yang, C.; Zhang, S.; Xu, Y.; et al. Myocardial infarction-induced hippocampal microtubule damage by cardiac originating microRNA-1 in mice. J Mol Cell Cardiol 2018, 120, 12–27. [Google Scholar] [CrossRef]
- Akbar, N.; Digby, J.E.; Cahill, T.J.; Tavare, A.N.; Corbin, A.L.; Saluja, S.; Dawkins, S.; Edgar, L.; Rawlings, N.; Ziberna, K.; et al. Endothelium-derived extracellular vesicles promote splenic monocyte mobilization in myocardial infarction. JCI Insight 2017, 2. [Google Scholar] [CrossRef]
- Panda, R.; Kubes, P. Extracellular vesicles selectively mobilize splenic neutrophils. Cardiovasc Res 2023, 119, 1–2. [Google Scholar] [CrossRef]
- Lyu, L.; Wang, H.; Li, B.; Qin, Q.; Qi, L.; Nagarkatti, M.; Nagarkatti, P.; Janicki, J.S.; Wang, X.L.; Cui, T. A critical role of cardiac fibroblast-derived exosomes in activating renin angiotensin system in cardiomyocytes. J Mol Cell Cardiol 2015, 89, 268–279. [Google Scholar] [CrossRef]
- Savvatis, K.; Pappritz, K.; Becher, P.M.; Lindner, D.; Zietsch, C.; Volk, H.D.; Westermann, D.; Schultheiss, H.P.; Tschöpe, C. Interleukin-23 deficiency leads to impaired wound healing and adverse prognosis after myocardial infarction. Circ Heart Fail 2014, 7, 161–171. [Google Scholar] [CrossRef]
- Kuwabara, Y.; Ono, K.; Horie, T.; Nishi, H.; Nagao, K.; Kinoshita, M.; Watanabe, S.; Baba, O.; Kojima, Y.; Shizuta, S.; et al. Increased microRNA-1 and microRNA-133a levels in serum of patients with cardiovascular disease indicate myocardial damage. Circ Cardiovasc Genet 2011, 4, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Carbone, F.; Nencioni, A.; Mach, F.; Vuilleumier, N.; Montecucco, F. Pathophysiological role of neutrophils in acute myocardial infarction. Thromb Haemost 2013, 110, 501–514. [Google Scholar] [CrossRef]
- Helseth, R.; Shetelig, C.; Andersen, G.; Langseth, M.S.; Limalanathan, S.; Opstad, T.B.; Arnesen, H.; Hoffmann, P.; Eritsland, J.; Seljeflot, I. Neutrophil Extracellular Trap Components Associate with Infarct Size, Ventricular Function, and Clinical Outcome in STEMI. Mediators Inflamm 2019, 2019, 7816491. [Google Scholar] [CrossRef] [PubMed]
- Arruda-Olson, A.M.; Reeder, G.S.; Bell, M.R.; Weston, S.A.; Roger, V.L. Neutrophilia predicts death and heart failure after myocardial infarction: a community-based study. Circ Cardiovasc Qual Outcomes 2009, 2, 656–662. [Google Scholar] [CrossRef]
- Dogan, I.; Karaman, K.; Sonmez, B.; Celik, S.; Turker, O. Relationship between serum neutrophil count and infarct size in patients with acute myocardial infarction. Nucl Med Commun 2009, 30, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Magnussen, C.; Blankenberg, S. Biomarkers for heart failure: small molecules with high clinical relevance. J Intern Med 2018, 283, 530–543. [Google Scholar] [CrossRef] [PubMed]
- Siranart, N.; Laohasurayotin, K.; Phanthong, T.; Sowalertrat, W.; Ariyachaipanich, A.; Chokesuwattanaskul, R. Proenkephalin as a Novel Prognostic Marker in Heart Failure Patients: A Systematic Review and Meta-Analysis. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Creemers, E.E.; Tijsen, A.J.; Pinto, Y.M. Circulating microRNAs: novel biomarkers and extracellular communicators in cardiovascular disease? Circ Res 2012, 110, 483–495. [Google Scholar] [CrossRef]
- Zhou, R.; Wang, L.; Zhao, G.; Chen, D.; Song, X.; Momtazi-Borojeni, A.A.; Yuan, H. Circulating exosomal microRNAs as emerging non-invasive clinical biomarkers in heart failure: Mega bio-roles of a nano bio-particle. IUBMB Life 2020, 72, 2546–2562. [Google Scholar] [CrossRef]
- Huang, J.P.; Chang, C.C.; Kuo, C.Y.; Huang, K.J.; Sokal, E.M.; Chen, K.H.; Hung, L.M. Exosomal microRNAs miR-30d-5p and miR-126a-5p Are Associated with Heart Failure with Preserved Ejection Fraction in STZ-Induced Type 1 Diabetic Rats. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Melman, Y.F.; Shah, R.; Danielson, K.; Xiao, J.; Simonson, B.; Barth, A.; Chakir, K.; Lewis, G.D.; Lavender, Z.; Truong, Q.A.; et al. Circulating MicroRNA-30d Is Associated With Response to Cardiac Resynchronization Therapy in Heart Failure and Regulates Cardiomyocyte Apoptosis: A Translational Pilot Study. Circulation 2015, 131, 2202–2216. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Salvador, A.M.; Li, G.; Valkov, N.; Ziegler, O.; Yeri, A.; Yang Xiao, C.; Meechoovet, B.; Alsop, E.; Rodosthenous, R.S.; et al. Mir-30d Regulates Cardiac Remodeling by Intracellular and Paracrine Signaling. Circ Res 2021, 128, e1–e23. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.P.; Tian, T.; Wang, J.Y.; He, J.N.; Chen, T.; Pan, M.; Xu, L.; Zhang, H.X.; Qiu, X.T.; Li, C.C.; et al. Hypoxia-elicited mesenchymal stem cell-derived exosomes facilitates cardiac repair through miR-125b-mediated prevention of cell death in myocardial infarction. Theranostics 2018, 8, 6163–6177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wei, Q.; Liu, X.; Zhang, T.; Wang, S.; Zhou, L.; Zou, L.; Fan, F.; Chi, H.; Sun, J.; et al. Exosomal microRNA-98-5p from hypoxic bone marrow mesenchymal stem cells inhibits myocardial ischemia-reperfusion injury by reducing TLR4 and activating the PI3K/Akt signaling pathway. Int Immunopharmacol 2021, 101, 107592. [Google Scholar] [CrossRef]
- Yuan, J.; Yang, H.; Liu, C.; Shao, L.; Zhang, H.; Lu, K.; Wang, J.; Wang, Y.; Yu, Q.; Zhang, Y.; et al. Microneedle Patch Loaded with Exosomes Containing MicroRNA-29b Prevents Cardiac Fibrosis after Myocardial Infarction. Adv Healthc Mater 2023, e2202959. [Google Scholar] [CrossRef]
- Pu, L.; Kong, X.; Li, H.; He, X. Exosomes released from mesenchymal stem cells overexpressing microRNA-30e ameliorate heart failure in rats with myocardial infarction. Am J Transl Res 2021, 13, 4007–4025. [Google Scholar]
- Yan, F.; Cui, W.; Chen, Z. Mesenchymal Stem Cell-Derived Exosome-Loaded microRNA-129-5p Inhibits TRAF3 Expression to Alleviate Apoptosis and Oxidative Stress in Heart Failure. Cardiovasc Toxicol 2022, 22, 631–645. [Google Scholar] [CrossRef]
- Shafei, S.; Khanmohammadi, M.; Ghanbari, H.; Nooshabadi, V.T.; Tafti, S.H.A.; Rabbani, S.; Kasaiyan, M.; Basiri, M.; Tavoosidana, G. Effectiveness of exosome mediated miR-126 and miR-146a delivery on cardiac tissue regeneration. Cell Tissue Res 2022, 390, 71–92. [Google Scholar] [CrossRef]
- Gao, L.; Qiu, F.; Cao, H.; Li, H.; Dai, G.; Ma, T.; Gong, Y.; Luo, W.; Zhu, D.; Qiu, Z.; et al. Therapeutic delivery of microRNA-125a-5p oligonucleotides improves recovery from myocardial ischemia/reperfusion injury in mice and swine. Theranostics 2023, 13, 685–703. [Google Scholar] [CrossRef]
- Wang, T.; Li, T.; Niu, X.; Hu, L.; Cheng, J.; Guo, D.; Ren, H.; Zhao, R.; Ji, Z.; Liu, P.; et al. ADSC-derived exosomes attenuate myocardial infarction injury by promoting miR-205-mediated cardiac angiogenesis. Biol Direct 2023, 18, 6. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, C.; Zhang, J.; Jiao, Z.; Dong, N.; Wang, G.; Wang, Z.; Wang, L. Localized injection of miRNA-21-enriched extracellular vesicles effectively restores cardiac function after myocardial infarction. Theranostics 2019, 9, 2346–2360. [Google Scholar] [CrossRef] [PubMed]
- Vandergriff, A.; Huang, K.; Shen, D.; Hu, S.; Hensley, M.T.; Caranasos, T.G.; Qian, L.; Cheng, K. Targeting regenerative exosomes to myocardial infarction using cardiac homing peptide. Theranostics 2018, 8, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Won, Y.W.; McGinn, A.N.; Lee, M.; Bull, D.A.; Kim, S.W. Targeted gene delivery to ischemic myocardium by homing peptide-guided polymeric carrier. Mol Pharm 2013, 10, 378–385. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol 2011, 29, 341–345. [Google Scholar] [CrossRef]
- Moholkar, D.N.; Kandimalla, R.; Gupta, R.C.; Aqil, F. Advances in lipid-based carriers for cancer therapeutics: Liposomes, exosomes and hybrid exosomes. Cancer Lett 2023, 565, 216220. [Google Scholar] [CrossRef]
Figure 1.
The biogenesis and potential sorting mechanisms of EV-miRNAs. The biogenesis of miRNAs is regulated by canonical (A) and non-canonical pathways (B and C), respectively. The latter is composed of Drosha/DGCR8-independent (B and C) and Dicer-independent pathways (E); The sorting sequences or 3’ modifications of miRNAs determine their incorporation in EVs (EXOmotifs or 3’-Uridylation) or cellular retention (CELLmotifs or 3’-Adenylation) [19,20]. The miRNAs with CELLmotifs or 3’-A’s are combined with AGO proteins to form the mature RISC, and then mediate either the mRNA degradation or translational repression by binding to 3’ UTR, and also regulate the translational activation by binding to 5’ UTR (G); The miRNAs with EXOmotifs or 3’-U’s are specifically recognized by RNA binding proteins (RNPs), and are then sorted into microvesicles (MVs) by binding with group 2 RNPs (K), or into Exosomes (EXOs) by binding with group 1 RNPs (I). In addition, the secretion of miRNAs is also controlled by neutral sphingomyelinase 2 (nSMase2) (K) [21]. Apoptotic extracellular vesicles (APO-EVs); ALG-2-interacting protein X (Alix); Caveolin-1 (Cav-1); DiGeorge syndrome critical region 8 (DGCR8); ELAV-like protein 1 (or HuR, human antigen R); Exosomes (EXOs); Exportin 1 (XPO1); Exportin 5 (XPO5); Heat shock protein HSP 90-beta (HSP90AB1); Heterogeneous nuclear ribonucleoprotein A2B1 (hnRNPA2B1); Heterogeneous nuclear ribonucleoprotein U (hnRNPU); HIV-1 transactivating response (TAR) RNA-binding protein (TRBP); Liquid-liquid phase separation (LLPS); Major vault protein (MVP); Microvesicles (MVs); Multivesicular bodies (MVBs); Ras-related nuclear protein (RAN); RNA binding proteins (RNPs); RNA-induced silencing complex (RISC); RNA polymerase II (Pol II); Serine/arginine-rich splicing factor 1 (SRSF1); Synaptotagmin-binding cytoplasmic RNA-interacting protein (SYNCRIP); Y-box protein 1 (YBX1).
Figure 1.
The biogenesis and potential sorting mechanisms of EV-miRNAs. The biogenesis of miRNAs is regulated by canonical (A) and non-canonical pathways (B and C), respectively. The latter is composed of Drosha/DGCR8-independent (B and C) and Dicer-independent pathways (E); The sorting sequences or 3’ modifications of miRNAs determine their incorporation in EVs (EXOmotifs or 3’-Uridylation) or cellular retention (CELLmotifs or 3’-Adenylation) [19,20]. The miRNAs with CELLmotifs or 3’-A’s are combined with AGO proteins to form the mature RISC, and then mediate either the mRNA degradation or translational repression by binding to 3’ UTR, and also regulate the translational activation by binding to 5’ UTR (G); The miRNAs with EXOmotifs or 3’-U’s are specifically recognized by RNA binding proteins (RNPs), and are then sorted into microvesicles (MVs) by binding with group 2 RNPs (K), or into Exosomes (EXOs) by binding with group 1 RNPs (I). In addition, the secretion of miRNAs is also controlled by neutral sphingomyelinase 2 (nSMase2) (K) [21]. Apoptotic extracellular vesicles (APO-EVs); ALG-2-interacting protein X (Alix); Caveolin-1 (Cav-1); DiGeorge syndrome critical region 8 (DGCR8); ELAV-like protein 1 (or HuR, human antigen R); Exosomes (EXOs); Exportin 1 (XPO1); Exportin 5 (XPO5); Heat shock protein HSP 90-beta (HSP90AB1); Heterogeneous nuclear ribonucleoprotein A2B1 (hnRNPA2B1); Heterogeneous nuclear ribonucleoprotein U (hnRNPU); HIV-1 transactivating response (TAR) RNA-binding protein (TRBP); Liquid-liquid phase separation (LLPS); Major vault protein (MVP); Microvesicles (MVs); Multivesicular bodies (MVBs); Ras-related nuclear protein (RAN); RNA binding proteins (RNPs); RNA-induced silencing complex (RISC); RNA polymerase II (Pol II); Serine/arginine-rich splicing factor 1 (SRSF1); Synaptotagmin-binding cytoplasmic RNA-interacting protein (SYNCRIP); Y-box protein 1 (YBX1).
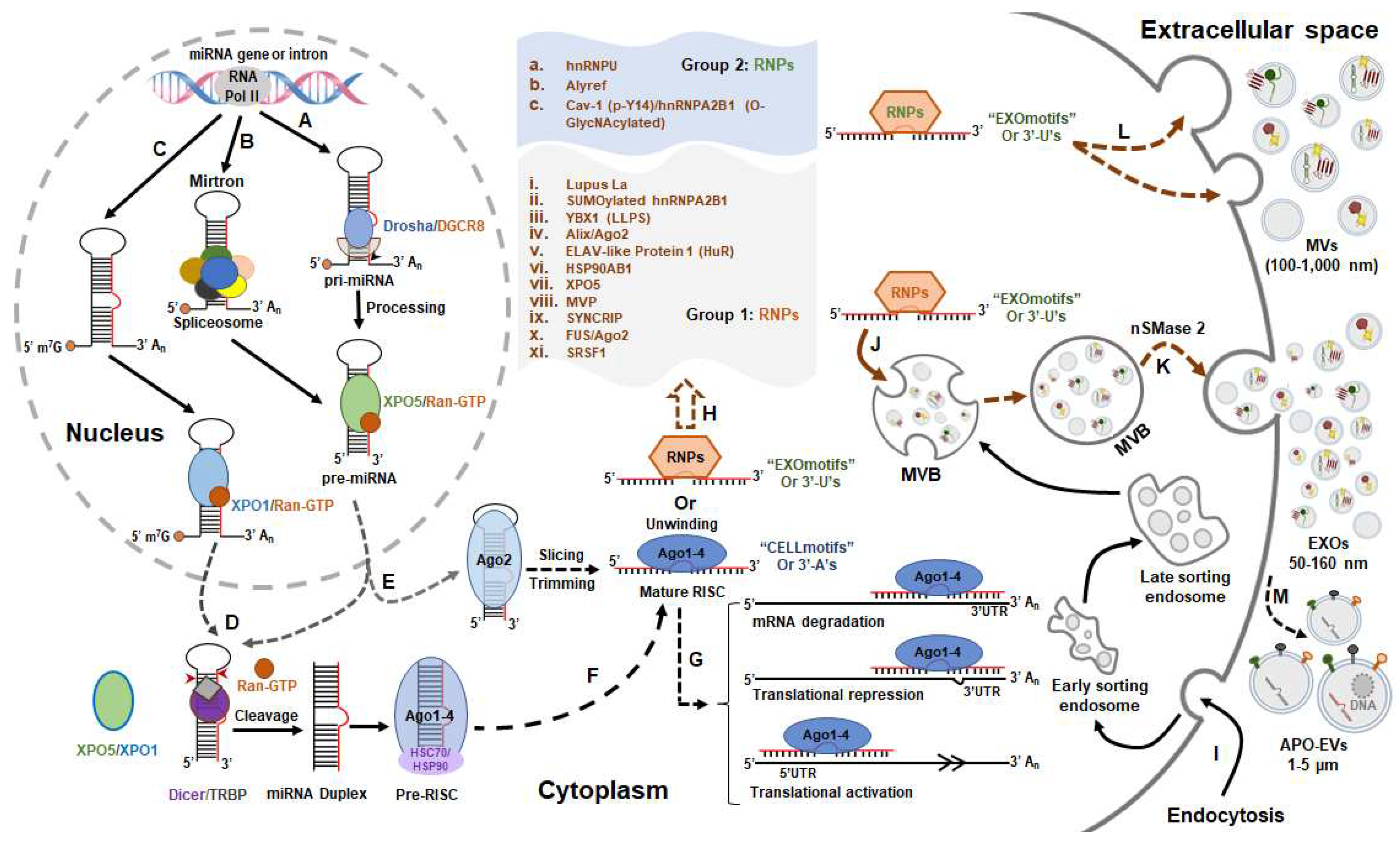

Table 1.
The potential EV-sorting mechanisms of some HF-associated miRNAs.
| miRNAs | Sorting mechanism | Functions in HF | Ref. |
|---|---|---|---|
| miR-122 | The binding of Lupus La protein, hnRNPU and/or HuR to miR122 controls extracellular export | Promote apoptosis, inflammation, fibrosis, pathological hypertrophy and remodeling | [73,76,81,82] |
| miR-223 | Selective sorting of miR-223 into EXOs by phase-separated YBX1 condensates | Promote cardiac fibrosis and hypertrophy | [69,77,83,84] |
| miR-34c-5p | The binding of Alyref and/or Fus to the CGGGAG motif at the 3’ end of miR-34c | Cardiac hypertrophy | [19,85] |
| miR-26a | The binding of Alyref and/or Fus to the CGGGAG motif at the 3’ end of miR-34c; alternatively, 3’-end uridylation of miR-26a | Protects the Heart Against Hypertension-Induced Myocardial Fibrosis | [19,20,86] |
| miR-30c-5p | The binding of hnRNPU to the AAMRUGCU motif of miR-30c-5p | Protects against myocardial ischemia/reperfusion injury | [67,87] |
| miR-17/92 | The binding of cav-1/hnRNPA2B1 complex to miR-17/92 regulates its MV sorting | Hypertrophic and arrhythmogenic cardiomyopathy | [78,88] |
| miR-1246 | The binding of SRSF1 to miR-1246 regulates its exosomal enrichment | Upregulated in diastolic dysfunction | [75,89] |
| miR-1231 | The binding hnRNPA2B1 to the GGAG EXOmotif at the 3’ end of miR-1231 | Induction of arrhythmias in ischemic hearts | [70,90] |
Table 2.
The EV-miRNAs in the pathogenesis of HF.
| Pathological phenotype | miRNA | Cell source | Target cell | Potential functional mechanism | Ref. |
|---|---|---|---|---|---|
| Cardiac hypertrophy | miRNA-21-3p | CF | CM | Translational inhibition of both SORBS2 and PDLIM5 | [41] |
| miRNA-27a-5p | CF | CM | Translational inhibition of PDLIM5 | [42] | |
| miRNA-27a-3p, miRNA-28-3p miRNA-34a | CF | CM | Dysregulation of Nrf2/ARE signaling and oxidative stress | [99] | |
| miR-200a | Adipocyte | CM | Selective activation of PPARγ signaling, and decreased TSC1 and subsequent mTOR activation | [100] | |
| miRNA-208a | CM | CM | Repression of Thrap1 and myostatin expression | [101,102] | |
| miRNA-217 | CF | CM | Targeting PTEN | [103] | |
| Cardiac fibrosis | miRNA-208a | CM | CF | Targeting Dyrk2 to promote NFAT dephosphorylation and nuclear translocation | [104] |
| miRNA-217 | CM | CF | Targeting PTEN | [103] | |
| miRNA-494-3p | CM | CF | Targeting PETN to enhance the phosphorylation of AKT, ERK and SMAD2/3 | [105] | |
| miRNA-218-5p | CM | CF | Targeting TNFAIP3 to activate TGF-β signaling | [106] | |
| miRNA-23a-3p | Adipocyte | CF | Targeting RAP1 | [107] | |
| miR-142-3p | Activated CD4+ T cell | CF | Targeting APC to activate the WNT signaling pathway | [108] | |
| miRNA-21 | MP and/or CM | CF | Targeting Spry1 to augment ERK-MAP kinase activity | [109-113] | |
| Angiogenesis | miRNA-200c-3p | CM | EC | Impaired endothelial migration and tube formation as well as a lower proliferation capacity. | [114] |
| miRNA-29a | CM | EC | Inhibiting the proliferation, migration and angiogenic ability of cardiac microvascular ECs | [115] | |
| miRNA-200a-3p | Activated CF | EC | Targeting ETS1/VEGF-A signaling axis | [116] | |
| Inter-organ communications | miRNA-1 | CM | Neuron | Targeting TPPP/p25 to disturb the stability of neuronal microtubules | [102,117] |
| miRNA-27a-3p, miRNA-28-3p and miRNA-34a | CM and/or CF | Neuron | Targeting Nrf2/ARE signaling to induce oxidative stress and subsequently elicit sympathetic excitation | [49] | |
| miRNA-126 | EC | NEUT | Transcriptional activation of NEUTs, and contribution to cardiac inflammation and chemokine production | [48,118,119] |
Abbreviations: Adenomatous Polyposis Coli (APC), Cardiac fibroblast (CF), Cardiac myocyte (CM), Dual-specificity tyrosine phosphorylation-regulated kinase 2 (Dyrk2), Endothelial cell (EC), ETS Proto-Oncogene 1 (ETS1), Macrophages (MP), Neutrophils (NEUTs), Nuclear factor of activated T cells (NFAT), Ras-related protein 1 (RAP1), Sprouty homologue 1 (Spry1), Thyroid hormone-associated protein 1 (Thrap1), Tuberous sclerosis complex (TSC1), Tubulin polymerization promoting protein (TPPP/p25).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



