
Submitted:
18 July 2023
Posted:
20 July 2023
Read the latest preprint version here
Abstract
A strong correlation between karyotype diversity and species richness in mammals was first reported over forty years ago: in mammalian phylogenetic clades, standard deviation of karyotype diversity (KD) closely corresponds to the standard deviation of species richness (SR). These initial studies, however, did not control for phylogenetic signal, raising the possibility that the correlation was due to phylogenetic relatedness among species in a clade. Accordingly, karyotype diversity trivially reflects species diversity simply as a passive consequence of adaptive radiations. A more recent study in mammals has controlled for phylogenetic signal and established the correlation as phylogenetically independent, suggesting that species diversity cannot, in itself, explain the observed corresponding karyotype diversity. The correlation is therefore remarkable because the mechanisms contributing to karyotype diversity are evolutionarily independent of the mechanisms contributing to species diversity. Recently, it was shown in salamanders that the two processes generating genetic diversity and species diversity are indeed independent and operate in parallel, suggesting a potential non-adaptive and non-causal relationship between the two fundamental variables involved in adaptive radiations. The following will examine the relationship between KD and SR in the context of Motoo Kimura’s theory of non-adaptive radiation.
Keywords:
karyotype diversity
; genome stability
; species richness
; species eveneness
; non-adaptive radiation
1. Introduction: formulating the question
Genetic diversity depends on the continuous and apparently random supply of mutations and genome transformations that ultimately result in reproductively incompatible karyotypes (reproductive isolation). The generation of genetic diversity takes place though varying modes of genetic change, including single base pair point mutations, gene rearrangements, as well as gene, chromosomal and whole genome duplications (1, 2, 3). Various mechanisms are believed to drive these genetic changes, including DNA replication errors, error prone DNA repair systems (eg. Non-homologous End-joining), transposon activity, recombination generating DNA polymorphisms, and environmental gene toxins causing DNA damage, as well as miscellaneous missrepair/missreplication events (ionizing radiation, heavy metals, the translesion Y-family of DNA polymerases, etc.) (4, 5, 6)
In contrast, species diversity is believed to occur primarily through natural selection acting on phenotypic diversity: species diversity is contingent upon existing gene/allele diversity (DNA polymorphisms) in a population and the ecological factors that favor the reproductive fitness of the corresponding phenotype. The ecological factors through which natural selection operates include climatic and edaphic changes, habitat and niche availability and diversity, sexual selection, etc. In terms of evolutionary time, the Neo-Darwinian adaptive theory of evolution is based on a changing environment acting positively on a changing phenotype, which allows for a given random mutation in an individual to proceed to “fixation”, or substitution, at the population level. Selection from standing genetic variation in a population might therefore be the typical response to an environmental shift (7, 8, 9, 10, 11).
The relationship between species diversity and the underlying molecular diversity among species has long raised the question of whether or not a causal mechanism somehow links these two forms of diversity during the speciation process (7, 8). Does the rate and level of the input of mutations drive the rate of speciation, or does the rate and level of environmental changes in ecological factors determine speciation rates? Presumably, higher rates of mutational input in a given lineage compared to lineages with slower rates of genetic change will generate a higher level of lineage specific genetic diversity, phenotype diversity and therefore species diversity (diversity within a taxonomic clade).
The rate at which speciation occurs, however, is expected, according to the Neo-Darwinian theory, to be largely contingent on the rate of niche change and the rate of change in a lineage’s geographic range (habitat diversity in association with geographic isolation of populations of the same species). In other words, do environmental factors constrain and limit rates of speciation, or are rates of speciation constrained and limited by molecular factors in the species’ germline? In the former case, the underlying genetic diversity would simply be a reflection of a clade’s species diversity: higher levels of species diversity mean trivially correspondingly higher levels of genetic diversity, and therefore adaptive evolution would not be mutation-limited but limited by selection (7).
In quantitative terms the question becomes: is the distribution in substitution rates statistically set by the distribution of mutation rates, and therefore causally correlated with it (genetic drift); or is it statistically set by the distribution of rates of adaptation (differential selection), and therefore not causally correlated with mutation rates? Are non-silent site base substitutions in codons, for example, correlated or uncorrelated with silent site base substitutions in genes across species and phylogenetic taxonomic groups? Some evidence suggests that the two types of base substitution in genes are indeed correlated, but if and how they might be causally correlated remains to be determined (12, 13, 14).
2. The Neutral Theory of Evolution and the Hypothesis of Non-Adaptive Radiation
A central tenet of Motoo Kimura’s Neutral Theory of Evolution is based on the principle that, for selectively neutral mutations, the rate of substitution in a population should equal the rate of mutation in an individual (mutation/substitution balance) (15). Kimura qualifies that “selectively neutral” more accurately means “selectively equivalent” mutations: many different mutations can fulfill the same function or maintain the same level of fitness. Consequently, most mutations that do not negatively affect fitness will “do the job equally well in terms of survival and reproduction of the individuals possessing them” (15). Expressed otherwise, most mutations not adversely affecting a species’ fitness can be equivalently selected (or have an equivalent likelihood of being positively selected) because they confer the same fitness value on the individual’s survival and reproductive success. Genetic diversity thus reduces to a type of “molecular fitness diversity” among those potential mutations that do not negatively affect the performance or functioning of the phenotype.
Kimura’s “four stage hypothesis” (Non-Adaptive Radiation) proposes a potential solution to the above conundrum of whether rates of mutation constrain, or limit, rates of speciation or whether rates of substitution, or selection and environmental change, account for rates of speciation and the corresponding levels of species richness in a clade. Although Kimura did not specifically refer to “rates of speciation,” his proposal does relate to the striking differences of species richness and levels of species unevenness between closely related taxonomic groups (clades) in the same lineage. In the mammalian lineage, for example, the taxonomic clade Rodentia comprises some 2,500 different species (among the over 5,500 defined mammalian species), while the Monotreme clade comprises just five species (16, 17). Is this difference explained because Monotremes inhabit different environments, exposing them to higher rates of extinction, or because they have slower rates of molecular evolution that are less adaptive or, conversely, more maladaptive?
2.1. Kimura’s hypothesis
According to Kimura’s proposal, macro-evolution consists of four stages:
1). Liberation from the preexisting selective constraint.
2). Sudden increase, or boom, of neutral variants under relaxed selection that are then fixed in the population by random genetic drift.
3). Realization of latent selective potential: some of the accumulated neutral mutants become useful at the phenotypic level in a new environment, which the species is then able to exploit.
4). Intergroup competition, as well as individual selection, leads to extensive adaptive evolution creating a radically different taxonomic group adapted to a newly opened ecological niche.
Kimura’s hypothesis would seem to suggest that genetic variation is increased in response to “liberation” from a previous selective constraint that resisted genetic change (negative selection). Genetic variants then accumulate as a result of genetic drift until they encounter a new selective constraint (environmental resistance). Competition between populations results in the elimination of maladaptive variants (competitive exclusion). Adaptive radiations then occur when new niches become available and more accessible due to the new phenotypes/adaptations that emerged during genetic drift and prior to adaptation.
Kimura believed that this proposal specifically addressed—should it turn out to be correct—the criticism that if most genetic changes are genuinely neutral then, by definition, they cannot be concerned with adaptation: if they are truly neutral, the reasoning goes, how then can they possibly contribute significantly to adaptation and speciation? And therefore “the neutral theory is biologically not very important.” A later modification of the neutral theory proposed that most mutations are “nearly neutral” but are maladaptive for the most part. When they are not eliminated by negative selection they can become fixed in a population if the effective population size is small enough for genetic drift to “overwhelm” selection.
Randomly occurring neutral or nearly neutral mutations should, according to the above criticism, show little or no relationship biologically to either speciation rates or to levels of species richness and unevenness. The limiting factor in speciation is the environmentally determined substitution rate and not the molecularly determined mutation rate. The extreme species richness and species uneveness in Rodentia, for example, must be due to highly fluctuating habitats and niches rather than to elevated mutation rates in the species belonging to that lineage.
In Kimura’s scenario, the question of whether mutational or ecological processes principally drive speciation becomes moot, if not irrelevant. His hypothesis implies that the two different processes must occur in parallel (mutation/substitution balance), and must be mutually contingent on each other in order to explain how the two independent mechanisms interact to contribute to speciation. What might be the mechanisms governing the balance between mutation and substitution in the process of speciation? Kimura’s hypothesis does not specifically address that question.
3. Peto’s paradox
Peto’s paradox is the observation that the evolution of a large body size in vertebrates does not apparently incur higher rates of whole organism DNA damage, mutation and genetic diseases such as cancer (18, 19, 20, 21). Assuming that every single cell in an organism has the same mutation rate, multicellular long-lived organisms should be at a higher risk for DNA damage and its negative (or positive) consequences, since more cells, and hence more DNA, provide a larger target for mutations. Paradoxically, however, vertebrates with large body sizes, for the most part, are even less prone to DNA damage related diseases such as cancer, and consequently they tend to have significantly longer maximum lifespans (MLS) than smaller body vertebrates.
Peto’s paradox has usually been explained in terms of the relationship between body size and basal metabolic rate (BMR). According to Kleiber’s law, basal metabolic rate decays with body mass, hence large animals have relatively lower BML per mass than smaller animals. One resolution of the paradox—among many other hypotheses—has proposed that species with larger body sizes produce less reactive oxygen species (ROS), which damage DNA, proteins and lipids. Genome size, however, might also play a role across broad taxonomic groups that range from short lived invertebrates, which have among the smallest animal genomes, to long lived vertebrates, which have, comparatively speaking, substantially larger genomes (22). Larger genomes result in larger cells that are consequently expected to have lower metabolic rates.
BMR set by body size and genome size is therefore a crucial determinant of MLS. Relatively short-lived mice, for example, have high metabolic rates compared to larger mammals that have significantly lower metabolic rates but similarly sized genomes. Large body size within a taxonomic group, and larger genome sizes between taxonomic groups thus correlate, in each case, with lower metabolic rates and lower levels of ROS: whales have a larger body size compared to mice but similar genome sizes, while salamanders have much larger genomes but mostly smaller body sizes compared to mammals. Thus genome size might be the crucial factor in explaining Peto’s paradox across taxonomic groups, while body size might be the crucial factor explaining Peto’s paradox within taxonomic groups. Recent theoretical and experimental analyses, for example, have shown that organisms with larger genomes are more prone to DNA replication fork stalling and so require enhanced DNA repair systems (ref. Blow)
4. The DNA Damage Response (DDR) system mediates the rate of mutation input
The mechanisms that maintain genome stability and determine mutation rates have been extensively studied. The DNA Damage Response (DDR) is a complex damage detection and DNA repair system that evolved in eukaryotes in response to DNA damage and the (non-random) invasion of transposons and retroviruses into the genome (23, 24, 25). The DDR therefore governs the rate of mutation in individuals before mutations can spread through the population either by drift or selection (26, 27). In other words, the DDR controls the level of genetic diversity in an individual while, as Kimura would argue, genetic drift mainly but not exclusively governs the level of genetic diversity in a population: mutation rates limit genetic diversity in populations and species. The Neo-Darwinians would claim, conversely, that positive selection in a fluctuating environment alone would determine the level of genetic diversity observed in a given population or species. In either scenario, the DDR thus mediates, via mutation input and/or selection, the level of genetic diversity within a species or population.
4.1. The fidelity and efficiency of the DDR varies significantly across taxonomic groups
The longer lifespans of large body animals can be understood then as a consequence of a more robust DDR in those species. The mechanisms involved in the positive correlation between maximum life span (MLS) and body size (BS) in vertebrates appear to be quite diverse, but at the genome level they all converge, either directly or indirectly, on the DDR in a manner that promotes, or enhances, genome stability: K-specialists and vertebrates with large bodies and long lifespans tend to have more proficient DDRs than do small body, short lifespan r-specialists and invertebrates both within and among diverse lineages (28).
Elephants, for example, have a more proficient DDR than do smaller mammals. This is attributed to the fact that elephant cells have multiple copies of the tumor suppressor gene p53, the “guardian of the genome” (29, 30, 31). TP53 orchestrates a complex network of factors that results in cell cycle arrest (checkpoint activation), DNA repair, senescence or apoptosis depending on the amount of DNA damage that the cell has experienced. Elephants consequently benefit from a stronger resistance to cancer compared to other mammals, because they rely on an enhanced branch of the DDR that reduces DNA damage by eliminating damaged cells. Other studies have revealed that immune system-related genes also increase with increasing body size, thus reducing the risk of cancer in larger animals (32). Apparently these genes promote the elimination of cancer cells that have escaped the cell-based tumor suppressor pathways.
More recently, it has been shown that the bowhead whale relies on more accurate and efficient DNA repair systems to preserve genome integrity. The bowhead has a lifespan of over 200 years, which is 100 fold more than the two to three year lifespan of the mouse (Mus musculus)—a difference of two orders of magnitude. The bowhead whale thus benefits from a long lifespan because its cells are more proficient at repairing DNA damage. The study identified two proteins (CIRBP and RPA2) that are present at high levels in fibroblasts and are known to increase the fidelity and efficiency of DNA double strand break (DSB) repair (33).
Other examples of species with a high resistance to cancer include sharks and salamanders (34, 35, 36). The mechanisms are unknown for the longevity and low rates of DNA turnover (DNA damage/genome alterations) found in these particular taxonomic groups, but both groups are known to have low levels of intra-specific heterozygosity, inter-specific genetic diversity, substitutions, single nucleotide variants (SNV), karyotype diversity (KD), etc. Among Amphibia, for example, salamanders have lower rates of genetic change compared to frogs (37, 38).
The same pattern is observed across taxonomic groups belonging to the Mammalia: different rates of genetic change among different phylogenetic groups, including mouse, rat, hamster and human, correlate positively with MLS. The elevated frequency of mutations in mouse was attributed in these studies to the mutagenic DNA translesion polymerase eta (39). Taken together, these observations explain, at the molecular level, why different taxonomic groups might have different levels of genetic diversity, and might reflect Kimura’s claim that most mutations are indeed neutral or nearly neutral: mutation rates are equivalent, or directly proportional, to substitution rates (mutation/substitution balance). One would expect then that varying lineage specific karyotype diversity would likewise reflect varying lineage specific genome stability: if mutation rates vary between lineages then karyotype diversity should also vary.
4.2. Sirtuin 6 and the Naked Mole Rat (NMR): a more proficient DDR promotes a longer lifespan independent of body size.
Perhaps the most intensively studied example of a species with a very long lifespan compared to its body mass/size is the naked mole rat (40, 41). The naked mole rat is a eusocial species of rodent, with a relatively small body size compared to other rodents. Again, in this species the relatively long lifespan was found to correlate with genome sequence integrity (genome stability) while deviating significantly from the expected correlation between MLS (per mass) and body size (42, 43). The Capybara, for example, has a body mass of 55,000 grams and a lifespan of only 15 years compared to the NMR with a body mass of just 35 grams but a lifespan of 32 years. In contrast, the house mouse, which has a body mass of about 40 grams, lives only about 4 years (42, 43).
In this model organism (NMR), longevity has been shown to correlate with the efficiency of the error prone Non-Homologous End Joining (NHEJ) and error free Homologous Recombination (HR) double strand break (DSB) repair pathways. Interestingly, longevity does not correlate with nucleotide excision repair (42), which mediates point mutations. The proteins involved in the DSB repair pathways are more highly expressed in the NMR compared to other related species. The enhanced repair efficiency is also related to the chromatin organizer SIRT6, which has been implicated in mediating the DNA repair of both single-strand and double strand DNA breaks. SIRT6 also plays an important role in maintaining heterochromatin and repressing transposon activity, both of which have been show to be related to aging (43). Hence, the correlations between cancer rates, the BMR and MLS are directly related to more proficient DDRs independent of body mass, at least in the naked mole rat and other long living small-bodied mammals, for example squirrels, whose longevity has been associated with a higher level of genome stability and integrity ().
5. Adaptive Evolution: mutation limited or selection limited, or both?
The above review of the relationship between DNA damage, body mass and MLS suggests that selection acts on different populations having different levels of genetic diversity, which depends directly on their different levels of DDR proficiency. Early studies have shown that genome stability is correlated with species richness in a taxonomic group or clade. In 1980, Bengsston measured standard deviations in karyotype diversity (KD) in mammals, and found that the standard deviations of KD in the different taxonomic groups correspond positively with their respective species richness (SR). Based on these observations, Bengsston hypothesized that the “properties of stable or unstable karyotypes may indicate that the cytological factors of importance are all of a submicroscopic nature.” One might propose that Bengsston’s submicroscopic cytological factors include the components of the DDR, and relate to the lineage varying DDR proficiencies that underlie stable or unstable karyotypes (44)
Other investigators have made similar observations. Bush, for example, examined 225 different vertebrate species in 1977 and found that rates of speciation are correlated with rates of chromosomal evolution, indicating that high KD reflects rapid speciation rates in species rich genera (45, 46). None of these studies, however, controlled for phylogenetic relatedness, and hence the findings might represent artifacts due to phylogenetic signal. A more recent investigation of Mammalia that did control for phylogenetic signal confirmed and extended the earlier observations: SR is proportional to and positively correlated with KD independently of species, or taxonomic, relatedness (Figure 1; 47).
Another study controlling for phylogenetic signal revealed a similar correlation between genome size (C-value) variation and species richness in Urodela. C-value variation, which is a proxy variable for KD in salamanders because of the large range in C-values, was found to correlate strongly with SR (Figure 2). Sister Families with pronounced differences in C-values had correspondingly pronounced differences in SR. Related findings reveal a close negative correlation between heterozygosity and C-value in the different salamander lineages (48, add ref. to Pierce and Mitton) Whether or not these correlations are due to a causal relationship between genome stability and rates of speciation, or to life history traits and effective population sizes, or some other variable(s), remains an open question.
Perhaps more remarkable than the correlation between KD and SR is the correlation found between KD and species unevenness observed in the mammalian phylogenetic tree. Species uneveness here refers to the abundance of species in a taxonomic group such as genus, family or order. At the order level, the mammalian phylogenetic tree is highly skewed, whereas the tree is much less skewed at the genus and family levels (Figure 3). Presumably the difference in unevenness at different taxonomic levels can be attributed to different rates of speciation and/or extinction, since different taxonomic levels in a lineage represent older (group) and younger (genus) radiations. Why such a close correlation between KD and SR unevenness would exist is unclear if selection is not limited by mutation or genome stability. A mutation-limited model of adaptation would seem to be the most parsimonious explanation, but such an explanation still remains to be confirmed.
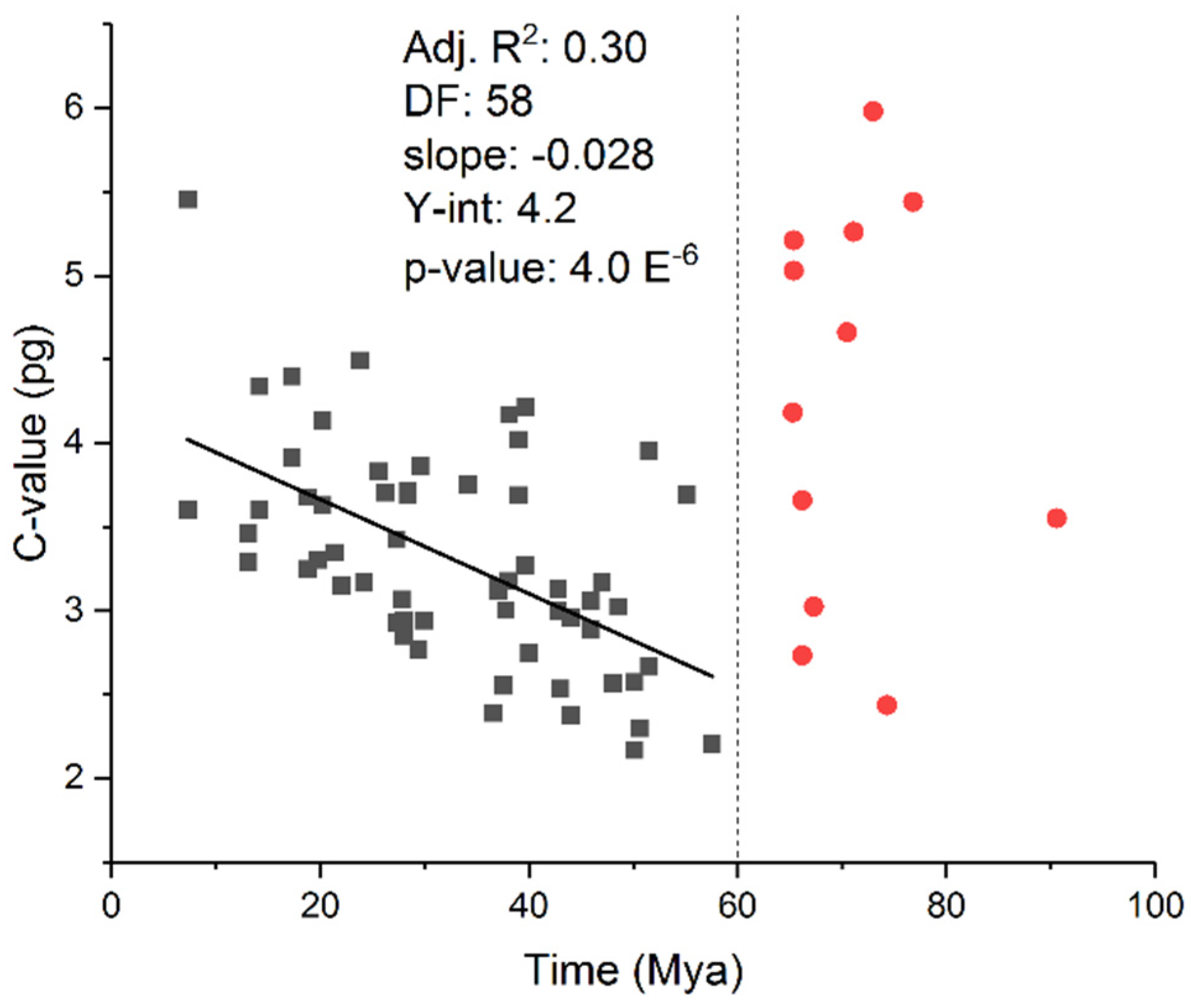
In contrast, path analysis in the salamander study did not reveal a causal relationship between C-value diversity and species richness, but instead showed that variation in C-value more likely occurred in parallel with variation in SR (49). In other words, the two variables (mutation and substitution/selection) are evolving simultaneously in a manner that determines rates of speciation: differing sizes of adaptive radiation might coincide with and might be contingent on differing rates of change in genome size and structure. Moreover, an abrupt transition in C-value size distributions in mammals was found to occur about 65 million years ago at the Cretaceous-Paleogene (K-Pg) boundary (Figure 4). It is commonly believed that the extinction of the dinosaurs resulted in the adaptive radiation of mammals.
Both of these observations would be consistent with Kimura’s hypothesis of non-adaptive radiation: 1) extinction of the dinosaurs “liberated” mammals from a selective constraint; 2) the small effective population sizes of mammalian species at the time of that event led to a “boom” of variants under “relaxed selection” (genetic drift); 3) some of the “neutral variants” became useful in the new unoccupied niches, habitats and environments (natural selection); competition (competitive exclusion) between groups resulted in adaptive radiations and created “radically different taxonomic groups.”
6. Other questions
What does it mean that the two speciation variables (mutation and substitution/selection) are coevolving independently but simultaneously? Selection can act only on mutations that improve (or degrade) phenotypes, and only at a rate corresponding to the available genetic variability present at the time of an ecological or environmental shift (such as the “sudden” extinction of the dinosaurs 65 million years ago). The larger the genetic diversity in a population, the faster will be the rate of speciation—necessarily, because of positive selection. Likewise, the faster the mutation rate, the larger will be the level of genetic diversity on which natural selection can act—though not necessarily, because of the differential effects of negative selection compounded with genetic drift, which can produce a positive, a negative or no correlation at all.
The correlation between species diversity and genetic diversity has been extensively studied at the community level, with genetic diversity within species showing both positive, negative and null relationships with species diversity in the respective community (50, 51, 52, 53, 54). Since the mechanisms governing mutation inputs and substitutions within a lineage are biologically unrelated and occur at different scales (germline vs. organism), there seems to be no reason then to expect, a priori, that KD and SR would be so consistently and so uniformly correlated across diverse mammalian lineages (Figure 3). Although that point might still be open to debate, it is nonetheless difficult to understand in statistical terms why any two taxonomic groups in Mammalia having the same SR would have exactly, or nearly, the same levels of KD, that is if selection, rather than drift, is the “typical response” to randomly fluctuating environments.
Transposon elements (TEs) appear to be another factor associated with varying mutation rates and levels of genome stability across different taxonomic groups (55). TE activation is not only related to aging, but also has been associated with adaptive radiations. Transposon activation, for example, is positively related to speciation in mammals (56). High rates of TE activity occur in the genome during the aging process as the DDR is weakened and repressive heterochromatin is lost (43). It would seem reasonable then that loss of repressive heterochromatin as a result of an environmental or some other form of stress might also occur during rapid adaptive radiations in response to a weakened, or “relaxed”, DDR (57).
Are the bursts in TE activity that coincide with recent adaptive radiations due to changes in the proficiencies of the DDR either due to genetic drift (58), or due to selection for less (or more) active DDRs? Whether or not these transposition events are causes or consequences of speciation remains unclear (59); but TEs probably reach fixation during speciation events via genetic drift rather than natural selection (60), which would be consistent with Kimura’s hypothesis. The epigenome, and heterochromatin in particular, would seem then to play a crucial role during either adaptive or non-adaptive radiations.
Negative selection reduces and limits genetic variability at the population level, and purifying selection is the preponderant form of natural selection. Genetic diversity is expected to increase in growing populations with small effective population sizes, but a small effective population size also implies a low amount of genetic diversity and heterozygosity. On the other hand, elevated mutation rates are maladaptive and result in genome instability, shorter lifespans and, potentially, higher rates of extinction. The balance between mutation and substitution thus seems to be set by MLS, body size and reproductive rate, and indeed there is some evidence in support of that proposal: propagule size, in accordance with a nucleotypic effect regulating cell size and generation time, correlates negatively with genome size and species richness (61). Other ecological factors such as a subterranean or cave dwelling habitat, for example, in some rodent and salamander lineages, certainly play important roles in speciation. What mechanisms might mediate these intriguing correlations remain to be more fully investigated.
7. Conclusion
This review has examined the evidence in support of a non-causal but biologically meaningful relationship between the vertebrate DDR (rate of mutation input) and adaptive radiations (substitution/selection rates) in the context of Kimura’s non-adaptive radiation hypothesis and Peto’s paradox: a larger supply of mutations due to a correspondingly weaker DDR will result in a wider spectrum of the genetic diversity on which positive/negative selection can act, and consequently a wider/narrower spectrum of species diversity among different taxonomic groups and lineages.
The issue then is to better elucidate the molecular and physiological origins of the wide and varying adaptive potential evident in the phylogenetic trees of diverse metazoan lineages, both in vertebrates and in invertebrates, in plants and in animals. As the evidence currently stands, the DDR proposal for explaining the correlated levels of KD and SR within and across metazoan lineages remains a hypothesis complementary to Kimura’s hypothesis that non-adaptive radiation constitutes the principle driving force behind macro-evolution.
Conflicts of Interest
The author declares no conflict of interest.
References
- Ayala, F.J; and Coluzzi, M. Chromosome speciation: Humans, Drosophila, and mosquitoes, Proc. Nat. Acad. Sci. 2005, 102 (suppl_1), 6535-6542.
- Presgraves, D.C. The molecular evolutionary basis of species formation Nature Reviews Genetics, 2010, 11, 175–180.
- Graphodatsky, A.S.; Trifonov, V.A.; Stanyon, R. The genome diversity and karyotype evolution of mammals. Mol Cytogenet. 2011, 12, 4:22. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Zhou, P.K. DNA damage repair: historical perspectives, mechanistic pathways and clinical translation for targeted cancer therapy. Signal Transduct Target Ther. 2021, 6, 254. [Google Scholar] [CrossRef] [PubMed]
- Scully, R.; Panday, A.; Elango, R.; Willis, N.A. DNA double-strand break repair-pathway choice in somatic mammalian cells. Nat Rev Mol Cell Biol. 2019, 20, 698–714. [Google Scholar] [CrossRef] [PubMed]
- Friedberg, E.C. A brief history of the DNA repair field. Cell Res. 2008, 18, 3–7. [Google Scholar] [CrossRef]
- Kern, A.D.; Hahn, M.W. The Neutral Theory in Light of Natural Selection. Mol Biol Evol. 2018, 35, 1366–1371. [Google Scholar] [CrossRef]
- Jensen, J.D.; Payseur, B.A.; Stephan, W.; Aquadro, C.F.; Lynch, M.; Charlesworth, D.; Charlesworth, B. The importance of the Neutral Theory in 1968 and 50 years on: A response to Kern and Hahn 2018. Evolution. 2019, 73, 111–114. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, J. Antagonistic pleiotropy conceals molecular adaptations in changing environments. Nat Ecol Evol. 2020, 4, 461–469. [Google Scholar] [CrossRef]
- Rousselle, M.; Simion, P.; Tilak, M.K.; Figuet, E.; Nabholz, B.; Galtier, N. Is adaptation limited by mutation? A timescale-dependent effect of genetic diversity on the adaptive substitution rate in animals. PLoS Genet. 2020, 16, e1008668. [Google Scholar] [CrossRef]
- Gerrish, P.J.; Colato, A.; Sniegowski, P.D. Genomic mutation rates that neutralize adaptive evolution and natural selection. J R Soc Interface. 2013, 10, 20130329. [Google Scholar] [CrossRef]
- Alvarez-Valin, F.; Jabbari, K.; Bernardi, G. Synonymous and nonsynonymous substitutions in mammalian genes: intragenic correlations. J Mol Evol. 1998, 46, 37–44. [Google Scholar] [CrossRef]
- Cameron, J.M.; Kreitman, M. The correlation between synonymous and nonsynonymous substitutions in Drosphila: mutation, selection or relaxed constraints? Genetics, 1998, 150, 767–775. [Google Scholar] [CrossRef]
- Wyckoff, G. J.; Malcom, C. M.; Vallender, E. J.; Lahn, B. T. A highly unexpected strong correlation between fixation probability of nonsynonymous mutations and mutation rate, Genome Analysis, 2005, 21, 381-385.
- Kimura, M. The neutral theory of molecular evolution: a review of recent evidence. Jpn J Genet. 1991, 66, 367–86. [Google Scholar] [CrossRef] [PubMed]
- Jones, K. E.; Safi, K. Ecology and evolution of mammalian biodiversity. Philos Trans R Soc Lond B Biol Sci. 2011, 366, 2451–61. [Google Scholar] [CrossRef] [PubMed]
- Burgin, C.J.; Colella, J.P.; Kahn, P.L.; Upham, N.S. . How many species of mammals are there? Journal of Mammalogy, 2018, 99, 1–14. [Google Scholar] [CrossRef]
- Aleah, F.; Caulin and Carlo, C.; Maley, C.C. Peto’s Paradox: Evolution’s Prescription for Cancer Prevention. Trends Ecol Evol. 2011, 26, 175–182. [Google Scholar]
- Callier, V. Solving Peto’s Paradox to better understand cancer. Proc. Natl. Acad. Sci., 2019, 116, 1825–1828. [Google Scholar] [CrossRef]
- Dart, A. Peto’s paradox put to the test. Nature Reviews Cancer 2022, 22, 129. [Google Scholar] [CrossRef] [PubMed]
- Nery, M. F.; Rennó, M.; Picorelli, A.; Ramos, E. A phylogenetic review of cancer resistance highlights evolutionary solutions to Peto's Paradox. Genet Mol Biol. 2022, 45(3 Suppl 1), e20220133. [Google Scholar] [CrossRef]
- Maciak, S. Cell size, body size and Peto's paradox. BMC Ecol Evol. 2022, 22, 142. [Google Scholar] [CrossRef]
- Feschotte, C.; Pritham, E.J. DNA Transposons and the Evolution of Eukaryotic Genomes. Ann Rev Genet. 2007, 41, 331–368. [Google Scholar] [CrossRef]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvák, Z.; Levin, H.L.; Macfarlan, T.S.; Mager, D.L.; Feschotte, C. Ten things you should know about transposable elements. Genome Biol. 2018, 19, 199. [Google Scholar] [CrossRef] [PubMed]
- Platt, R.N. 2nd; Vandewege, M.W.; Ray, D.A. 2nd; Vandewege, M. W.; Ray, D.A. Mammalian transposable elements and their impacts on genome evolution. Chromosome Res. 2018, 26, 25–43. [Google Scholar]
- Uphoff, S. Real-time dynamics of mutagenesis reveal the chronology of DNA repair and damage tolerance responses in single cells. Proc Natl Acad Sci U S A. 2018, 115, E6516–E6525. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, L.A.; Besenbacher, S.; Zheng, J.; Li, P.; Bertelsen, M.F.; Quintard, B.; Hoffman, J.I.; Li, Z.; St Leger, J.; Shao, C.; Stiller, J.; Gilbert, M.T.P.; Schierup, M.H.; Zhang, G. Evolution of the germline mutation rate across vertebrates. Nature. 2023, 615, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Seluanov, A.; Gladyshev, V.N.; Vijg, J.; Gorbunova, V. Mechanisms of cancer resistance in long-lived mammals. Nat Rev Cancer. 2018, 18, 433–441. [Google Scholar] [CrossRef]
- Abegglen, L.M.; Caulin, A.F.; Chan, A.; Lee, K.; Robinson, R.; Campbell, M.S.; Kiso, W.K.; Schmitt, D.L.; Waddell, P.J.; Bhaskara, S.; Jensen, S.T.; Maley, C.C.; Schiffman, J.D. Potential Mechanisms for Cancer Resistance in Elephants and Comparative Cellular Response to DNA Damage in Humans. JAMA 2015, 314, 1850–60. [Google Scholar] [CrossRef]
- Sulak, M.; Fong, L.; Mika, K.; Chigurupati, S.; Yon, L.; Mongan, N.P.; Emes, R.D.; Lynch, V.J. TP53 copy number expansion is associated with the evolution of increased body size and an enhanced DNA damage response in elephants. Elife 2016, 5, e11994. [Google Scholar] [CrossRef]
- Callaway, E. How Elephants Avoid Cancer. Nature 2015. [Google Scholar] [CrossRef]
- Takemoto, K.; Ii, M.; Nishizuka, S.S. Importance of metabolic rate to the relationship between the number of genes in a functional category and body size in Peto's paradox for cancer. R Soc Open Sci. 2016, 2016 3, 160267. [Google Scholar] [CrossRef]
- Firsanov, D.; Zacher, M.; Tian, X.; Zhao, Y.; George, J.C.; Sformo, T. L.; Trombline, G.; Biashed, A.; Gilman, A.: Hamilton, N.; Patel, A.; Straight, A.; Lee, I. S.; Lu, Y J.: Haseljic, E. Williams, A.: Miller, N.; Gladyshev, V. N.; Zhang, Z. Vijg, J. Seluanove, A.: Gorbunova, V. DNA repair and anti-cancer mechanisms in the longest-living mammal: the bowhead whale. bioRxiv, 2023. [CrossRef]
- Marra, N.J.; Richards, V.P.; Early, A. Comparative transcriptomics of elasmobranchs and teleosts highlight important processes in adaptive immunity and regional endothermy. BMC Genomics 2017, 18, 87. [Google Scholar] [CrossRef]
- Cohen A., A.; Deelen, J.; Jones, O. R. Editorial: Mechanisms and Pathways Contributing to the Diversity of Aging Across the Tree of Life. Front Cell Dev Biol. 2022, 10, 854700. [Google Scholar] [CrossRef]
- Yun, M.H. Salamander Insights Into Ageing and Rejuvenation. Front Cell Dev Biol. 2021, 9, 689062. [Google Scholar] [CrossRef]
- Mohlhenrich, E.R.; Mueller, R.L. Genetic drift and mutational hazard in the evolution of salamander genomic gigantism. Evolution 2016, 70, 2865–2878. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, H.C.; Gower, D.J.; Wilkinson, M.; Gomez-Mestre, I. Macroevolutionary shift in the size of amphibian genomes and the role of life history and climate. Nat Ecol Evol. 2018, 2, 1792–1799. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dong, X.; Tian, X.; Lee, M.; Ablaeva, J.; Firsanov, D.; Lee, S.G.; Maslov, A.Y.; Gladyshev, V.N.; Seluanov, A.; Gorbunova, V.; Vijg, J. Maintenance of genome sequence integrity in long- and short-lived rodent species. Sci Adv. 2021, 7, eabj3284. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.N.; Mele, J.; Hornsby, P.J.; Buffenstein, R. Stress resistance in the naked mole-rat: the bare essentials - a mini-review. Gerontology. 2012, 58, 453–62. [Google Scholar] [CrossRef]
- Munro, D.; Baldy, C.; Pamenter, M.E.; Treberg, J.R. The exceptional longevity of the naked mole-rat may be explained by mitochondrial antioxidant defenses. Aging Cell. 2019, 18, e12916. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Firsanov, D.; Zhang, Z.; Cheng, Y.; Luo, L.; Tombline, G.; Tan, R.; Simon, M.; Henderson, S.; Steffan, J.; Goldfarb, A.; Tam, J.; Zheng, K.; Cornwell, A.; Johnson, A.; Yang, J.N.; Mao, Z.; Manta, B.; Dang, W.; Zhang, Z.; Vijg, J.; Wolfe, A.; Moody, K.; Kennedy, B.K.; Bohmann, D.; Gladyshev, V.N.; Seluanov, A.; Gorbunova, V. SIRT6 Is Responsible for More Efficient DNA Double-Strand Break Repair in Long-Lived Species. Cell 2019, 177, 622–638. [Google Scholar] [CrossRef]
- Gorbunova, V.; Seluanov, A.; Mita, P.; McKerrow, W.; Fenyö, D.; Boeke, J.D.; Linker, S.B.; Gage FH, Kreiling, J. A.; Petrashen, A.P.; Woodham, T.A.; Taylor, J.R.; Helfand, S.L.; Sedivy, J.M. The role of retrotransposable elements in ageing and age-associated diseases. Nature 2021, 596, 43–53. [Google Scholar] [CrossRef]
- Bengtsson, B.O. Rates of karyotype evolution in placental mammals. Hereditas 1980, 92, 37–47. [Google Scholar] [CrossRef]
- Bush, G.L.; Case, S.M.; Wilson, A.C.; Patton, J.L. Rapid speciation and chromosomal evolution in mammals. Proc Natl Acad Sci U S A. 1977, 74, 3942–6. [Google Scholar] [CrossRef] [PubMed]
- Maxson, L. E. R.; Wilson, A. C. RATES OF MOLECULAR AND CHROMOSOMAL EVOLUTION IN SALAMANDERS. Evolution 1979, 33, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Herrick, J.; Sclavi, B. Genome diversity and species richness in mammals bioRxiv 2019,. [CrossRef]
- Nevo, E.; Beiles, A. (1991) Genetic diversity and ecological heterogeneity in amphibian evolution. Copeia 1991, 565–592. [Google Scholar] [CrossRef]
- Sclavi, B.; Herrick, J. Genome size variation and species diversity in salamanders. J Evol Biol. 2019, 32, 278–286. [Google Scholar] [CrossRef]
- Leffler, E.M.; Bullaughey, K.; Matute, D.R.; Meyer, W.K.; Ségurel, L.; Venkat, A.; Andolfatto, P.; Przeworski, M. Revisiting an old riddle: what determines genetic diversity levels within species? PLoS Biol. 2012, 10, e1001388. [Google Scholar] [CrossRef]
- Vellend, M. Species diversity and genetic diversity: parallel processes and correlated patterns. Am Nat. 2005, 166, 199–215. [Google Scholar] [CrossRef]
- Vellend, M.; Geber, M.A. Connections between species diversity and genetic diversity. Ecology letters, 2005, 8, 762–781. [Google Scholar] [CrossRef]
- Petersen, H.C.; Hansen, B.W.; Knott, K.E.; Banta, G.T. Species and genetic diversity relationships in benthic macroinvertebrate communities along a salinity gradient. BMC Ecol Evol. 2022, 22, 125. [Google Scholar] [CrossRef]
- Marchesini, A.; Vernesi, C.; Battisti, A.; Ficetola, G.F. Deciphering the drivers of negative species-genetic diversity correlation in Alpine amphibians. Mol Ecol. 2018, 27, 4916–4930. [Google Scholar] [CrossRef]
- Osmanski, A,B.; Paulat, N.S.; Korstian, J.; Grimshaw, J.R.; Halsey, M.; Sullivan, K.A.M.; Moreno-Santillán, D.D.; Crookshanks, C.; Roberts, J.; Garcia, C. Johnson, M.G.; Densmore, L.D.; Stevens, R.D.; Zoonomia Consortium; Rosen, J, Storer, J.M.; Hubley, R.; Smit, A.F.A.; Dávalos, L.M.; Karlsson, E.K.; Lindblad-Toh, K.; Ray, D.A. Insights into mammalian TE diversity through the curation of 248 genome assemblies. Science. 2023, 380, eabn1430. 1430.
- Ricci, M.; Peona, V.; Guichard, E.; Taccioli, C.; Boattini, A. Transposable Elements Activity is Positively Related to Rate of Speciation in Mammals. J Mol Evol. 2018, 86, 303–310. [Google Scholar] [CrossRef]
- Wang, J.; Jia, S.T.; Jia, S. New Insights into the Regulation of Heterochromatin. Trends Genet. 2016, 32, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Sung, W.; Ackerman, M.S.; Miller, S.F.; Doak, T.G.; Lynch, M. Drift-barrier hypothesis and mutation-rate evolution. Proc Natl Acad Sci U S A. 2012, 109, 18488–92. [Google Scholar] [CrossRef] [PubMed]
- Serrato-Capuchina, A.; Matute, D.R. The Role of Transposable Elements in Speciation. Genes 2018, 9, 254. [Google Scholar] [CrossRef] [PubMed]
- Jurka, J.; Bao, W.; Kojima, K.K. Families of transposable elements, population structure and the origin of species. Biol. Direct 2011, 19, 44. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. Genome Size Covaries More Positively with Propagule Size than Adult Size: New Insights into an Old Problem. Biology 2021, 10, 270. [Google Scholar] [CrossRef]
- ADDED REFERENCE: Roscoe, S.; Stone, G.; Garcia, M.; Lutz, F. Reciprocal chromosome painting shows that squirrels, unlike murid rodents, have a highly conserved genome organization. Genomics 2003, 82, 745–749. [Google Scholar]
Figure 1.
Phlylogenetic Least Squares (PGLS) analysis reveals that Karyotype Diversity (KD) and Species Richness (SR) in Mammalia are strongly correlated at the taxonomic Order level, thus confiming Bengtsson’s orginal observation that SR correlates with KD.
Figure 1.
Phlylogenetic Least Squares (PGLS) analysis reveals that Karyotype Diversity (KD) and Species Richness (SR) in Mammalia are strongly correlated at the taxonomic Order level, thus confiming Bengtsson’s orginal observation that SR correlates with KD.
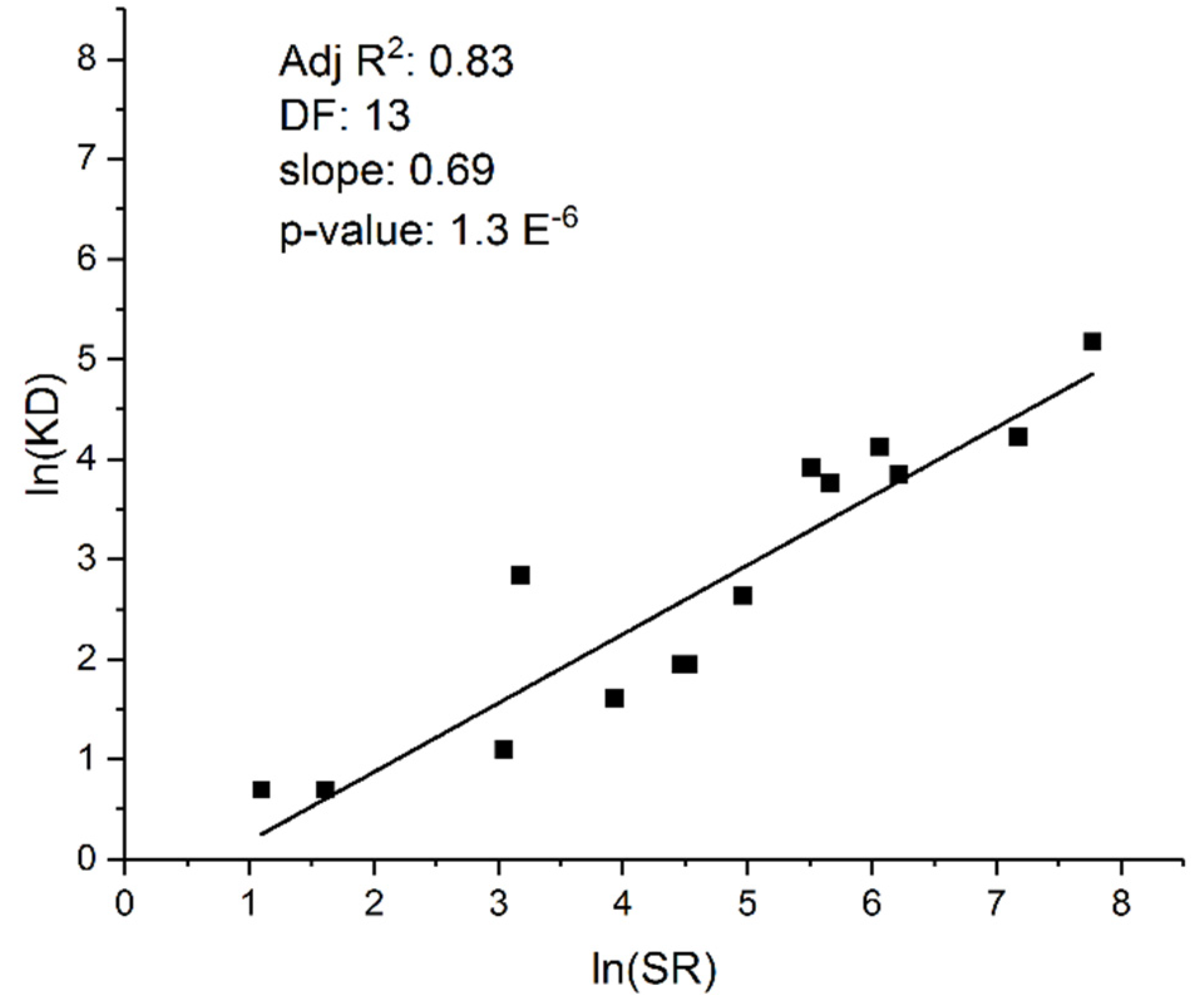
Figure 2.
Regression analysis reveals a correlation in salamanders between Species Richness and genome size (C-value: haploid genome size measured in picograms). A) Ordinary Least Squares (OLS) regression analysis at the taxonomic Family level. The letters refer to the different salamander families examined (inset); B) Phylogenetic Independent Contrasts (PIC) confirm a non-phylogeneticlly determined relationship between SR and C-value. C-value is considered a proxy variable for karyotype and genome diversity because of the highly variable salamander genome sizes across the Urodela (C-value ranges from 10 to up to 120 pg). Note that controlling for phylogenetic signal substantially improves the correlation, whereas OLS, which does not control for phylogenic signal, reveals two distinct groups depending on life history traits: paedomorphs (Families a, e, f, i) vs. metamorphic and direct developing Families).
Figure 2.
Regression analysis reveals a correlation in salamanders between Species Richness and genome size (C-value: haploid genome size measured in picograms). A) Ordinary Least Squares (OLS) regression analysis at the taxonomic Family level. The letters refer to the different salamander families examined (inset); B) Phylogenetic Independent Contrasts (PIC) confirm a non-phylogeneticlly determined relationship between SR and C-value. C-value is considered a proxy variable for karyotype and genome diversity because of the highly variable salamander genome sizes across the Urodela (C-value ranges from 10 to up to 120 pg). Note that controlling for phylogenetic signal substantially improves the correlation, whereas OLS, which does not control for phylogenic signal, reveals two distinct groups depending on life history traits: paedomorphs (Families a, e, f, i) vs. metamorphic and direct developing Families).
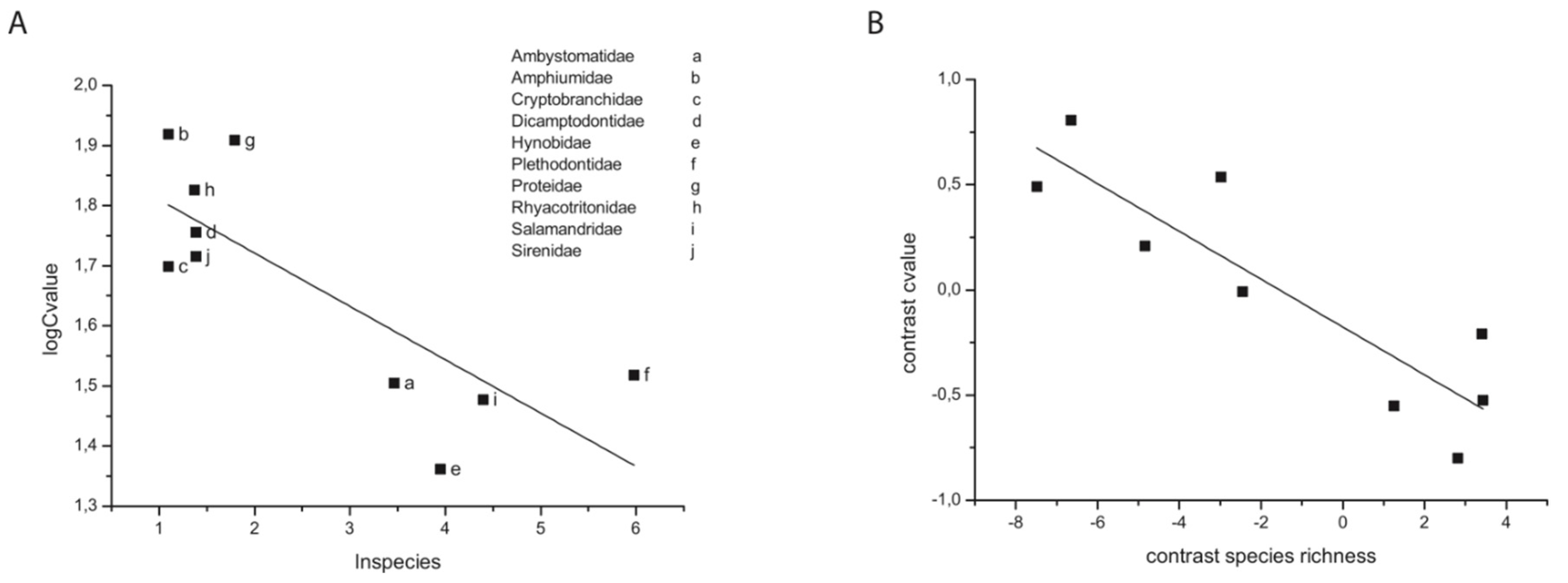
Figure 3.
Unevenness in species richness and unevenness in karyotype diversity. Left y-axis: number of different karyotypes per taxonomic order. Right y-axis: number of species per taxonomic order. The findings extend Bengsston’s original observation revealing a consistency and uniformity of unevenness between species richness and karyotype diversity in the different mammalian orders.
Figure 3.
Unevenness in species richness and unevenness in karyotype diversity. Left y-axis: number of different karyotypes per taxonomic order. Right y-axis: number of species per taxonomic order. The findings extend Bengsston’s original observation revealing a consistency and uniformity of unevenness between species richness and karyotype diversity in the different mammalian orders.
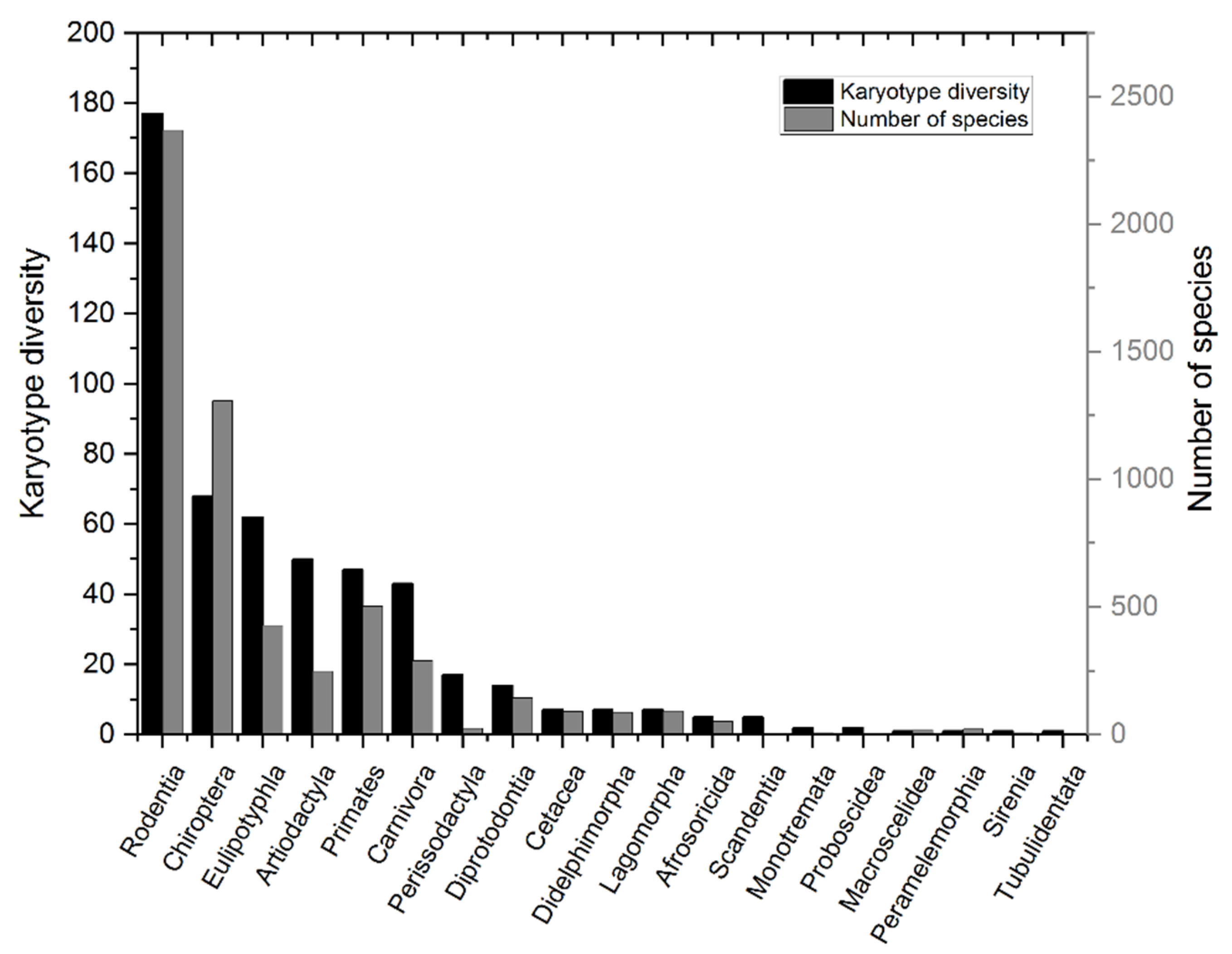
Figure 4.
Genome size evolution in Mammalia. OLS anaylsis was used to regress the clade stem age (origin in time of the appearance of a taxonomic group in the mammalian Tree of Life) on to clade average C-values at both family and order level clades. Two distinct populations are identifiable: clades with origins before the Pg-K extinction event (to right of 60 Mya), and clades that emerged after the Pg-K extinction event (to the left of 60 Mya). Note that clades extant before the extinction event have widely varying C-values, and no correlation exists between stem age and C-value. In contrast, clades that emerged after the event exhibit a clear and significant correlation between stem age and C-value revealing that later emerging clades (more recent) have increasingly larger C-values (R2 = 0.30; p = 4 x 10-6; slope = 0.028 pg/Mya; y-intercept = 4.2 pg). Mammalian population sizes presumably increased dramatically after the K-Pg mass extinction due to “liberation” from a previous selective constraint (environmental resistance such as predation, niche availability etc.). The respective populations then appear to diversify at a fairly constant rate according to a genome size evolutionary “molecular clock”, which might reflect a balance between the forces of genetic drift and natural selection.
Figure 4.
Genome size evolution in Mammalia. OLS anaylsis was used to regress the clade stem age (origin in time of the appearance of a taxonomic group in the mammalian Tree of Life) on to clade average C-values at both family and order level clades. Two distinct populations are identifiable: clades with origins before the Pg-K extinction event (to right of 60 Mya), and clades that emerged after the Pg-K extinction event (to the left of 60 Mya). Note that clades extant before the extinction event have widely varying C-values, and no correlation exists between stem age and C-value. In contrast, clades that emerged after the event exhibit a clear and significant correlation between stem age and C-value revealing that later emerging clades (more recent) have increasingly larger C-values (R2 = 0.30; p = 4 x 10-6; slope = 0.028 pg/Mya; y-intercept = 4.2 pg). Mammalian population sizes presumably increased dramatically after the K-Pg mass extinction due to “liberation” from a previous selective constraint (environmental resistance such as predation, niche availability etc.). The respective populations then appear to diversify at a fairly constant rate according to a genome size evolutionary “molecular clock”, which might reflect a balance between the forces of genetic drift and natural selection.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



