
Submitted:
20 July 2023
Posted:
20 July 2023
You are already at the latest version
Abstract
Lodging is one of the important factors affecting the high and stable yield of wheat worldwide. Solid-stemmed wheat has higher stem strength and lodging resistance than hollow-stemmed wheat. There are many solid stemmed varieties, landraces and old varieties of durum wheat. However, the transfer of solid stem genes from durum wheat is suppressed by the suppressor gene located on chromosome 3D in common wheat, and only hollow-stemmed lines have been created. However, synthetic hexaploid wheat can serve as a bridge to transfer solid stem genes from tetraploid wheat to common wheat. In this study, the F1, F2, and F2:3 generations of the cross between solid-stemmed Syn-SAU-119 and semisolid-stemmed Syn-SAU-117 were developed. A single dominant gene, tentatively designated Su-TdDof, was identified in synthetic hexaploid wheat Syn-SAU-117 by genetic analysis, which suppresses stem solidity. Using bulked segregant RNA-seq (BSR-seq) analysis, Su-TdDof was mapped to chromosome 7DS and flanked by markers KASP-669 and KASP-1055 within a 4.53 cM genetic interval corresponding to 3.86 Mb and 2.29Mb physical region in the Chinese Spring (IWGSC RefSeq v1.1) and Ae. tauschii (AL8/78 v4.0) genome, respectively, in which three genes related to solid stem development were annotated. Su-TdDof differed from a previously reported solid stem suppressor gene based on its origin and position. Su-TdDof would provide a valuable example for research on the suppression phenomenon. The flanking markers developed in this study would be useful for screening Ae. tauschii accessions with no suppressor gene (Su-TdDof) to develop more synthetic hexaploid wheat lines for wheat lodging resistance breeding and further cloning the suppressor gene Su-TdDof.
Keywords:
semisolid
; gene mapping
; Aegilops tauschii
; synthetic hexaploid wheat
; lodging resistance
1. Introduction
Wheat is the largest cereal crop in the world, accounting for 220 million hectares with annual global production of 772 million tons [1]. The world's population is expected to increase by nearly 2 billion over the next 30 years [2], and there will be a greater demand for global wheat production [3]. Therefore, we must strive to increase wheat yield. However, with the continuous improvement of wheat yield and the increase in fertilizer application, lodging has become an important factor affecting the high and stable yield of wheat [4,5,6,7,8]. Plant dwarfing can effectively alleviate the lodging damage of wheat [9]; however, severe dwarfism leads to inadequate biomass accumulation and lower yield potential [10]. Another potential strategy is to breed for increased mechanical strength [11]. Solid-stemmed wheat plays an important role in lodging resistance breeding. The ratio of stem wall thickness to outer stem diameter and mechanical tissue content of solid-stemmed wheat were significantly higher than those of common wheat [12,13,14]. Therefore, solid-stemmed wheat has higher stem strength and lodging resistance than common wheat [13].
To date, two candidate genes, TraesCS3B01G608800 and TRITD3BV1G280530, have been reported for stem solidity in hexaploid wheat and durum wheat, respectively. The differentially expressed gene TraesCS3B01G608800 (KAF7034036.1) showed copy number variation associated with stem solidity in different hexaploid wheat cultivars [15]. However, TRITD3BV1G280530 was confirmed as a candidate gene in SSt1 in durum wheat, and the copy number of TRITD3BV1G280530 in solid-stemmed durum wheat and hollow-stemmed durum wheat is different. The protein encoded by TRITD3BV1G280530 is a zinc finger protein, and its physical location is 829.1 Mb on chromosome 3BL of durum wheat genome v1.0 [16]. Liu et al. [17] found that a QTL for pith thickness in wheat was previously discovered on 3BL in a double haploid population of ‘Westonia’ × ‘Kauz’. A putative vacuolar processing enzyme gene TaVPE3cB was screened out as a potential pith-thickness candidate gene in Australian ‘Westonia’ wheat [17].
There are many solid-stemmed varieties, landraces and old varieties in durum wheat (Triticum turgidum L. ssp. durum, AABB, 2n = 4x = 28) [18]. Durum wheat has higher stem firmness and more stable genetic characteristics than common wheat varieties [19,20,21]. Beginning in the 1940s, breeders tried to transfer the solid stem gene from Golden Ball to hexaploid wheat by crossing, but the solid stem trait was suppressed, and only hollow-stemmed lines were created [19,22,23]. Then, it was found that the expression of the solid stem gene was suppressed, and the suppressor gene was presumed to be located on chromosome 3D in common wheat [19]. Suppression is a common phenomenon in nature. For example, suppression of disease resistance is especially frequent in resistance gene expression to fungal plant pathogens causing stem rust, leaf rust, and powdery mildew [24,25,26,27,28,29,30]. Inhibition often occurs, especially during the transfer of foreign genes from diploid and tetraploid ancestors to hexaploid wheat [24,25,26,29,31,32].
In our previous study, it was found that the D genome of Ae. tauschii AS92 suppressed the expression of the solid stem gene from the 3B chromosome in Syn-SAU-117 [33]. The objective of this study was to identify and map the solid stem suppressor gene in Syn-SAU-117 using bulked segregant RNA-seq (BSR-seq) analysis.
2. Results
2.1. Differential Expression of the TdDof Gene in Different Materials
The gene expression levels of TdDof were determined in two synthetic wheat cultivars, Syn-SAU-117 and Syn-SAU-119, and two durum wheat cultivars, Ma (solid) and Cocorit (hollow), as solid and hollow-stemmed contrasts during early internode elongation (Zadoks stage 32 and Zadoks stage 34). The expression level of TdDof was higher in Syn-SAU-119 than in Syn-SAU-117 in both periods (Figure 1 a, b).
2.2. Genetic Analysis of the Solid Stem Suppressor Gene Su-TdDof
According to Pauw et al. [34], synthetic hexaploidy wheat Syn-SAU-119 was solid-stemmed (solidity = 5.0; on 1-5 scale) (Figure 2a), and Syn-SAU-117 was semisolid-stemmed (solidity = 4.2; on 1-5 scale) (Figure 2c). Syn-SAU-117 and Syn-SAU-119 were hybridized to obtain F1, F2 and F2:3 populations for the genetic analysis of solid stem inhibition in Syn-SAU-117. The solidity of stems of all F1 plants was similar to that of Syn-SAU-117 plants (solidity = 4.0) (Figure 2b). The F2 population segregated into 36 solid-stemmed (solidity = 1) and 120 semisolid-stemmed (1 < solidity < 5) plants, fitting a 1S: 3SS ratio (χ2 = 0.308, p = 0.579) (Figure 2e, 2f) (Table 1), indicating that the inhibition of solid stems was conferred by a dominant single gene tentatively designated Su-TdDof. At Zadoks stage 34, the anatomical structure of the stalks of synthetic hexaploid wheat showed that the Syn-SAU-119 parenchyma was complete (Figure 3a), and the Syn-SAU-117 and Syn-SAU-117 × Syn-SAU-119 F1 parenchyma showed a similar degree of apoptosis (Figure 3c, 3b). In the Syn-SAU-117 × Syn-SAU-119 F2 population, the parenchyma of some plants was complete (Figure 3d), while that of other plants showed apoptosis (Figure 3e). The segregation rate of the F2:3 population composed of 134 families was 35 (homozygous solid): 67 (heterozygous): 32 (homozygous semisolid) (χ1:2:12 = 0.134, p = 0.935), which is consistent with the separation results of the F2 population (Table 1).
2.3. BSR-Seq Analysis of the RNA of Bulks with Contrasting Stem Solidity
The RNA samples of the solid bulk and the semisolid bulk were subjected to RNA-seq analysis, which generated 56,746,340 and 82,341,7507 raw reads, respectively. After quality control, 56,740,782 and 82,334,750 high-quality reads from the solid bulk and semisolid bulk were uniquely mapped to the Chinese Spring genome (IWGSC RefSeq v1.1), respectively. A total of 8581 SNPs (p < 1e-8 and AFD > 0.6) were identified from these reads using GATK software (Figure. 4). One hundred and twenty-three SNPs was located in an 8 Mb genomic interval (4-11 Mb) on the short arm of chromosome 7D in the Chinese Spring genome (IWGSC RefSeq v1.1), which were regarded as candidate SNPs linked to Su-TdDof (Figure 5).
2.4. Molecular Mapping of Su-TdDof
Fifty-one out of the 123 clustered SNPs on 7DS were chosen to develop KASP markers. Four of them were successfully converted into KASP markers (KASP-533, KASP-669, KASP-1055, KASP-1166) (Table 2) and scored reliably on the parents as well as the solid and semisolid bulks. All tested markers exhibited identical haplotypes between Syn-SAU-117 and AS92 but were distinct from those of Syn-SAU-119 and AS96 (Table 3). Subsequently, these KASP markers were used to genotype 134 F2 plants derived from the cross between solid-stemmed Syn-SAU-119 and semisolid-stemmed Syn-SAU-117 plants. Linkage analysis indicated that KASP-669 was potentially mapped 1.88 cM distal and KASP-1055 was placed 2.65 cM proximal to Su-TdDof (Figure 6).
2.5. Gene Analysis of the Su-TdDof Genomic Region
The sequences of the closely linked markers KASP-669 and KASP-1055 were blasted against the Chinese Spring genome and the Ae. tauschii genome to obtain their physical positions. Su-TdDof was physically mapped to a 3.86 Mb region between the 6.69 Mb to 10.55 Mb regions of the Chinese Spring 7DS chromosome (IWGSC RefSeq v1.1) and between the 6.58 Mb to 8.87 Mb regions (2.29 Mb) in the Ae. tauschii AL8/78 7DS chromosome (Ae. tauschii AL8/78 v4.0) (Figure 6). There were 180 and 125 predicted genes in the target physical regions in Chinese Spring and Ae. tauschii AL8/78, respectively (IWGSC RefSeq v1.1; Ae. tauschii AL8/78 v4.0, Supplementary Tables S1 and S2). In the Chinese Spring genome, ten genes may be associated with the growth and development of plant stems, including six zinc finger protein-related genes (TraesCS7D02G015800, TraesCS7D02G019600LC, TraesCS7D02G022000LC, TraesCS7D02G023300LC, TraesCS7D02G023400LC, TraesCS7D02G024900LC), two biofunction inhibitor genes (TraesCS7D02G020000, TraesCS7D02G020100), one pectin lyase-like superfamily protein gene (TraesCS7D02G016300), and one homeobox-like protein BEL1 gene (TraesCS7D02G019800). Five genes, including two zinc finger protein-related genes (AET7Gv20034800, AET7Gv20042400), one 36.4 kDa proline-rich protein gene (AET7Gv20040700), one transcription factor gene (AET7Gv20036400), and one homeobox-like protein BEL1 gene, were found in the Ae. tauschii genome, which had a good collinear relationship with those of Chinese Spring (Figure S1).Transcriptome analysis revealed a total of 43362 differentially expressed genes, including 341 upregulated genes, 227 downregulated genes and 42794 nondifferentially expressed genes (Figure 7). There were 12 and 7 significant differentially expressed genes in the target physical regions in the Chinese Spring genome and Ae. tauschii genome (Supplementary Tables S3 and S4, Figure S1). Among them, the annotations of TraesCS7D02G016300, AET7Gv20040000 and AET7Gv20040700 were probably associated with the growth and development of plant pith [16,44,45,46,47,48,49,50]. TraesCS7D02G016300 was a gene encoding pectin lyase superfamily proteins (PG), acting on pectin and lignin of the cell wall and promoting cell wall degradation and shedding, thus promoting cell apoptosis (Supplementary Table S3). The homologous genes of TraesCS7D02G016300 were two polygalacturonase genes (AET7Gv20034200 and AET7Gv20034900) in the Ae. tauschii genome (Ae. tauschii AL8/78 v4.0), and their gene annotations were not correlated with the inhibition of solid stems (Supplementary Tables S3 and S4). AET7Gv20040700 and AET7Gv20040000 in the Ae. tauschii genome (Ae. tauschii AL8/78 v4.0) had a good collinear relationship with those of Chinese Spring (Figure S1). AET7Gv20040000 was annotated as a homeobox-like protein BEL1 gene (Supplementary Table S4). The homologous gene of AET7Gv20040000 in Chinese Spring is TraesCS7D02G019800, and its functional annotation was a homeobox-like protein BEL1 gene (Supplementary Table S1), the same as AET7Gv20040000 in Ae. tauschii AL8/78 genome. AET7Gv20040700 was annotated as a 36.4 kDa proline-rich protein gene (Supplementary Table S4). The homologous gene of AET7Gv20040700 in Chinese Spring is TraesCS7D02G020100, and its functional annotation is bifunctional (protease/α-amylase) inhibitor/plant lipid transfer protein/seed storage helical domain (Supplementary Table S1).
3. Discussion
To date, lodging is still a problem in wheat-growing regions worldwide, despite scientists having made great efforts to solve it for many years. The selection of excellent germplasms with alternative semidwarf genes or good stem mechanical strength may be an effective way to solve this problem [51]. Solid-stemmed wheat has strong lodging resistance due to its higher stalk strength [13,35]. Durum wheat has many solid-stemmed varieties, landraces, and old varieties [18]. However, attempts to transfer solid stem genes to hexaploid wheat by direct cross have been unsuccessful because the expression of solid stem genes is suppressed by the suppressor gene on chromosome 3D in common wheat [12,22,23]. In this study, a new solid stem suppressor gene Su-TdDof was identified in synthetic hexaploid wheat Syn-SAU-117 and mapped on chromosome arm 7DS flanked by markers KASP-669 and KASP-1055 within a 4.53 cM genetic interval corresponding to 3.86 Mb physical region in the Chinese Spring genome (IWGSC RefSeq v1.1). In addition, the expression of solid stem gene TdDof in Syn-SAU-117 was lower than that in Syn-SAU-119, confirming the existence of the solid stem suppressor gene Su-TdDof.
In common wheat, the existence of many suppressor genes affects the normal expression of some important genes and the utilization of excellent foreign genes [30,35,52,53,54,55,56]. To date, many disease resistance genes and corresponding suppressor genes have been found in the D genome of common wheat and the D genome donor Ae. tauschii [35,55,57,58], such as the leaf rust suppressor gene Su-Lr23 on chromosome 2DS [56] and the stem rust suppressor gene SuSr-D1 on chromosome 7DL of hexaploid wheat cultivar 'Canthatch' (CTH) [60,61,62,63]. A recent study showed that the gene SuSr-D1 encoded Med15, a subunit of the Mediator complex that suppressed the expression of stem rust resistance [30]. In the present study, Su-TdDof was from Ae. tauschii, which was different from the suppressor gene presumed to be located on chromosome 3D in common wheat found by Larson et al. [12].
Su-TdDof was physically mapped to the region between 6.58 Mb to 8.87 Mb (2.29 Mb) in the Ae. tauschii AL8/78 7DS chromosome (Ae. tauschii AL8/78 v4.0) (Figure. 6). Based on gene functional annotation and screening of differentially expressed genes in the transcriptome, there were two protein-coding genes, AET7G20040000 and AET7Gv20040700, in the target physical regions in the Ae. tauschii genome (Supplementary Tables S3, S4, Figure S1). AET7Gv20040000 was annotated as a homeobox-like protein BEL1 gene, and the homologous gene of AET7Gv20040000 in Chinese Spring is TraesCS7D02G019800. Its functional annotation was the same as that of AET7Gv20040000. BEL1 protein family genes play an important role in the growth and development of plant stems, leaves, flowers and other organs [45,47]. For example, in Arabidopsis thaliana, the specific interaction between the BEL1 protein-like family BLH6 and KNAT7 inhibits the transcription factor REVOLUTA (REV), affecting growth and development in the stem of Arabidopsis inflorescences, thereby regulating secondary cell wall development [46]. AET7Gv20040700 was annotated as a 36.4 kDa proline-rich protein gene, and the homologous gene of AET7Gv20040700 in Chinese Spring is TraesCS7D02G020100. Its functional annotation is bifunctional (protease/α-amylase) inhibitor/plant lipid transfer protein/seed storage helical domain. Studies have shown that the formation of pith in the stem is related to starch (16,44,48). AET7Gv20040700 may inhibit the hydrolysis of starch and affect the formation of pith. These two genes, AET7Gv20040000 and AET7Gv20040700, will be cloned and sequenced in future studies to further develop markers for verification.
During the introduction of foreign genes into common wheat, with the increase in ploidy, the expression of superior genes decreased or was completely inhibited because of the existence of suppressor genes [35,52]. Therefore, exploring new suppressor genes, screening accessions without suppressor genes or carrying out artificial mutation of suppressor genes can enable breeders to break through this restriction and provide beneficial help for the introduction of foreign genes into common wheat [64]. The flanking markers KASP-669 and KASP-1055 developed in this study could be used as molecular markers to screen recombinant heterozygous plants, construct secondary F2 populations and develop markers, and further narrow the location interval to fine map and clone Su-TdDof. The flanking markers KASP-669 and KASP-1055 were also used to screen Ae. tauschii accessions with no suppressor gene (Su-TdDof) to develop more synthetic hexaploid wheat lines with solid stems for lodging-resistance breeding. Solid-stemmed synthetic hexaploid wheat can be used as a bridge to cross with elite wheat cultivars [65]. Combined with molecular marker-assisted selection, the transfer of solid stem genes from tetraploid wheat into common wheat cultivars and breeding new wheat cultivars with solid stems will provide new materials for wheat lodging resistance breeding.
4. Materials and Methods
4.1. Plant Materials
Two synthetic hexaploid wheat lines (Syn-SAU-117 and Syn-SAU-119), two different durum wheats (Ma and Cocorit), and two different Ae. tauschii (2n = 2x = 14, DD) accessions (AS92 and AS96) were used in this study. Syn-SAU-117 and Syn-SAU-119 were generated by natural chromosome doubling of Ma × AS92 F1 and Ma × AS96 F1, respectively. Syn-SAU-117 and Syn-SAU-119 were identified by FISH using oligonucleotide probes Oligo-pSc119.2-1 and Oligo-pTa535-1 [33]. Plants with 42 chromosomes were used in this study. Durum wheats Ma (solid stem) and Cocorit (hollow stem) were supplied by George Fedak at the Ottawa Research and Development Centre in Agriculture and Agri-Food Canada. The lines with the code AS are stored in our institute. All materials used in this study were kept at the Triticeae Research Institute of Sichuan Agricultural University.
4.2. Population Construction and Phenotypic Investigation
Two synthetic hexaploid wheat lines were sown in the greenhouse in July 2020, and Syn-SAU-119/Syn-SAU-117 F1 plants were subsequently generated. These F1 seeds were sown in a greenhouse in March 2021. Syn-SAU-119/Syn-SAU-117 F2 seeds were sown in the greenhouse in July 2021. Syn-SAU-119/Syn-SAU-117 F2:3 plants were sown in the field in November 2021. Each plant was 10 cm apart within rows, 30 cm apart between rows, and 1.5 m in length. The stems were sampled according to Kong et al. [13]. More than ten stems from the main tiller were randomly selected after flowering and were cross-sectionally cut at the center of each internode. The level of stem solidity was rated as 1-5 (1 for hollow and 5 for solid) following Pauw et al. [34].
4.3. Observation of the Anatomical Structures of Stems
The internodes were numbered consecutively from the base to the top of the stem. At the jointing stage, the main tiller was selected. The center of the second internode of the wheat stem base was cut into 1 cm pieces and then soaked in FAA fixative for more than 24 h [35]. The samples were sent to Wuhan CVI Biotechnology Co., Ltd. (https://www.servicebio.cn/) for the preparation of paraffin sections. CaseViewer 2.3 (https://www.3dhis tech. com/solut ions/casev iewer/, 3DHISTECH, Hungary) was used to view the results of the paraffin section analysis.
4.4. Solid Stem Gene Expression Analysis
A quantitative reverse transcription polymerase chain reaction (qRT‒PCR) system (Bio-Rad) was used to analyze the gene expression of TdDof [15]. The D1 probe primers (D1_F: GTTCCTGCACGCCATGGAC; D1_R: TCCCCCATCGTCGCCATTA) were designed specifically to distinguish differences in expression levels between different plants, and the housekeeping gene GAPDH was used as a reference for gene expression analysis. The main stems of three plants were sampled at Zadoks Stage 32 and Zadoks Stage 34 when the first two and four nodes were present on the stem. Approximately 0.5 cm of the stem was sampled, measuring from the bottom of the lowermost node toward the uppermost node. The samples were immediately placed in 1.5 mL microcentrifuge tubes, flash-frozen in liquid nitrogen, and stored at −80 °C before RNA extraction. The stem tissue was ground in liquid nitrogen with a sterilized mortar and pestle. Total RNA extraction was performed using the Tiangen DP441 RNA prep Pure RNA Extraction Kit according to the manufacturer's protocol. The quality of RNA was assessed by polyacrylamide gel electrophoresis, and RNA reverse transcription was performed using the Fermentas K1622 RT Reverse Transcription Kit (Thermo Scientific).
4.5. Bulked Segregant RNA-Seq (BSR-Seq)
Phenotypically contrasting F2 generations showing different stem solidity in the greenhouse were used to construct solid and semisolid RNA pools for RNA-Seq. Equal amounts of RNA from 20 homozygous solid-stemmed and 20 semisolid-stemmed generations were pooled to conduct bulked segregant analysis [36]. The RNA samples were sequenced on the platform of Chengdu Tiancheng Future Technology Co., Ltd. (https://www.tcuni.com/). Sequence quality control was performed using the software fastp v0.19.5 [37]. RNA reads of the solid stem and semisolid stem bulks were aligned to the reference genome sequence of Chinese Spring v1.1 [15] and Ae. tauschii AL8/78 v4.0 using STARv2.5.1b software [38]. The unique and confident alignments were applied to call SNP variants using GATK v3.6 software [39]. The SNP variants with P values of Fisher’s exact test (FET) < 1e−8 and allele frequency difference (AFD) > 0.6 were considered associated with solid stem suppression and further used as templates to develop SNP markers [36].
4.6. Kompetitive Allele-Specific PCR (KASP) Assays
The solidity-related SNPs and the 500 bp flanking sequences served to design the KASP primers and tested polymorphisms on the parental lines, the solid and semisolid stem DNA bulks. Polymorphic markers that could be reliably scored were genotyped on the F2 segregation population of Syn-SAU-119 × Syn-SAU-117. For each KASP assay, a 10 µl reaction volume containing 5 µl of 2 KASP master mix (Biosearch Technologies), 1.4 µl primer mix (mixture of 0.168 µM each forward A1 and A2 primers, and 0.42 µM of reverse primer), 100 ng of genomic DNA and 2.6 µl of ddH2O was prepared. The CFX96Touch™ real-time PCR detection system (Bio-Rad, USA) was used for amplification under the following conditions: 15 min at 94 °C, 10 touchdown cycles of 20 s at 94 °C, 60 s at 65-57 °C (decreasing by 0.8 °C per cycle), and 32 cycles of 20 s at 94 °C, 60 s at 57 °C.
4.7. Data Analysis
Chi-square (χ2) tests were used to determine the goodness of fit for the observed segregation and expected ratios of the F2 and F2:3 populations. Linkage analysis was performed using MAPMAKER/EXP v3.0b [40]. The Kosambi function was used to convert recombination values to genetic distances [41]. A logarithmic odds (LOD) ratio of 3.0 and a maximum distance of 50.0 cM were set as thresholds for the declaration of linkage. The genetic linkage map was drawn using Mapdraw v2.1 software [42].
4.8. Candidate Gene Analysis
The corresponding sequences of the markers KASP-669 and KASP-1055 linked to Su-TdDof were used to BLAST against the genomes of common wheat cv. Chinese Spring [15] and Ae. tauschii AL8/78 v4.0 [43]. Gene annotations between the flanking markers of the two genomes were retrieved from the online databases Ensembl Plants (http://plants.ensembl.org/index.html) and Swiss-Prot (http://www.gpm-aw.com/html/swi-ss-prot.html). Furthermore, the differentially expressed genes within the interval were screened and analyzed based on the results of RNA-seq, with the screening criteria of FDR<0.05 and |LogFC|>1. Collinearity analysis was performed on the differentially expressed genes related to the function of solid stems among parents and mixed pools in the Chinese Spring reference genome and the Ae. tauschii reference genome.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions statement
L.Z., D.L., and H.W. designed the research and supervised the study. H. L., X.L., J.Z., L.C., M.Z, Y.M., P.M., X.C., and X.C. performed the experiments. H. L., L.Z., D.L., H.W., M.H., B.J., S.N., L.H., and Z.W. discussed results and wrote the paper. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
This research was supported by Major Program of National Agricultural Science and Technology of China (NK20220607), the Key Research and Development Program of Sichuan Province, China (2021YFYZ0002) and Open Fund of Key Laboratory of Wheat Biology and Genetic Improvement on Southwestern China (ZWS2022001).
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAOSTAT. Food and agricultural organization of the United Nations, FAOSTAT https:// www. fao. org/ faost at/en/. Accessed 28 January 2022.
- UN (United Nations, Department of Economic and Social Affairs, Population Division). World Population Prospects: The 2015 Revision, Key Findings and Advance Tables. 2015.
- Zhang, C.Z.; Huang, L.; Zhang, H.F.; Hao, Q.Q.; Lyu, B.; Wang, M.N.; Lynn, E.; Liu, M.; Kou, C.L.; Qi, J.; et al. An ancestral NB-LRR with duplicated 3’ UTRs confers stripe rust resistance in wheat and barley. Nat. Commun. 2019, 10, 4023. [CrossRef]
- Keller M, Karutz C, Schmid J E, et al. Quantitative trait loci for lodging resistance in a segregating wheat× spelt population. Theor. Appl. Genet. 1999, 98, 1171–1182. [CrossRef]
- Islam, M.S; Peng, S.B.; Visperas, R.; Ereful, N.; Bhuiya, M.S. Julfiquar, A.W. Lodging-related morphological traits of hybrid rice in a tropical irrigated ecosystem. Field Crop Res. 2007, 101, 240–248. [CrossRef]
- Acreche, M.M.; Slafer, G.A. Lodging yield penalties as affected by breeding in Mediterranean wheats. Field Crop Res. 2011, 122, 40–48. [CrossRef]
- Berry, P.M.; Spink, J. Predicting yield losses caused by lodging in wheat. Field Crop Res. 2012, 137, 19–26. [CrossRef]
- Peake, A.S.; Huth, N.I.; Carberry, P.S.; Raine, S.R.; Smith, R.J. Quantifying potential yield and lodging-related yield gaps for irrigated spring wheat in sub-tropical Australia. Field Crop Res. 2014, 158, 1–14. [CrossRef]
- Flintham, J.E.; Börner, A.; Worland, A.J.; GALE, M.D. Optimizing wheat grain yield: effects of Rht (gibberellin-insensitive) dwarfing genes. J. Agr. Sci. 1997, 128, 11–25. [CrossRef]
- Hirano, K.; Ordonio, R.L.; Matsuoka, M. Engineering the lodging resistance mechanism of post-Green Revolution rice to meet future demands. P. Jpn. Acad. B-Phys. 2017, 93(4), 220–233. [CrossRef]
- Keller, M.; Karutz, C.; Schmid, J.E.; Stamp, P.; Winzeler, M.; Keller, B. Messmer, M.M. Quantitative trait loci for lodging resistance in a segregating wheat × spelt population. Theor. Appl. Genet. 1999, 98, 1171–1182. [CrossRef]
- Larson, R.I.; MacDonald, M.D.; Inheritance of the type of solid stem in Golden Ball (Triticum durum). III. The effect of selection for solid stem beyond F5 in hexaploid segregates of the hybrid Rescue (T. aestivum) × Golden Ball. Can. J. Genet. Cytol. 1963, 5, 437–444.
- Kong, E.Y.; Liu, D.C.; Guo, X.L.; Yang, W.L.; Sun, J.Z.; Li, X.; Zhan, K.H.; Cui, D.Q.; Lin, J.X.; Zhang, A.M. Anatomical and chemical characteristics associated with lodging resistance in wheat. Crop J., 2013, 1, 43–49. [CrossRef]
- Beres, B.L.; Cárcamo, H.A.; Byers, J.R.; Clarke, F. R.; Pozniak C.J.; Basu, S.K.; DePauw, R.M. Host plant interactions between wheat germplasm source and wheat stem sawfly Cephus cinctus Norton (Hymenoptera: Cephidae) I. Commercial cultivars. Can. J. Plant Sci. 2013, 93, 607–617. [CrossRef]
- International Wheat Genome Sequencing Consortium (IWGSC).; Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.; Choulet, F.; Distelfeld, A.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science, 2018, 361, eaar7191. [CrossRef]
- Nilsen, K.T.; Walkowiak, S.; Xiang D.Q.; Gao, P.; Quilichini, T.D.; Willick, L.R.; Byrns, B.; Diaye, A.N.; Ens, J.; Wiebe, K.; et al. Copy number variation of TdDof controls solid-stemmed architecture in wheat. Proc. Natl. Acad. Sci. 2020, 117, 28708–28718. [CrossRef]
- Liu, Q.; Zhao, Y.; Rahman, S.J.; She, M.; Zhang, J.J.; Yang, R.C.; Islam, S.; O’Hara, G.; Varshney, R.; Liu, H.; et al. The putative vacuolar processing enzyme gene TaVPE3cB is a candidate gene for wheat stem pith-thickness. Theor. Appl. Genet. 2023, 136(6): 138. [CrossRef]
- Damania, A.B.; Pecetti, L.; Qualset, C.O.; Humeid, B. Diversity and geographic distribution of stem solidness and environmental stress tolerance in a collection of durum wheat landraces from Turkey. Genet. Resour. Crop Evol. 1997, 44, 101–108. [CrossRef]
- Platt, A.; Farstad, C. W.; Breeding spring wheats for resistance to wheat stem sawfly attack. Proc. 7th Pac. Sci. Cong. 1949, 4, 215–220.
- Clarke, F.R.; Clarke, J.M.; Knox, R.E.; Inheritance of stem solidness in eight durum wheat crosses. Can. J. Plant Sci. 2002, 82, 661–664. [CrossRef]
- Singh, A.K.; Clarke, J.M.; Knox, R.E.; Depauw, R.M.; McCaig, T.N.; Cuthbert, R.D.; Clarke, F.R.; Fernandez, M.R. AAC Raymore durum wheat. Can. J. Plant Sci. 2014, 94, 1289–1296. [CrossRef]
- Larson, R.I. Inheritance of the type of solid stem in Golden Ball (Triticum durum): I. early generations of a hybrid with Rescue (T. aestivum). Can. J. Bot. 1959, 37, 889–896. [CrossRef]
- Larson, R.I. Inheritance of the type of solid stem in Golden Ball (Triticum durum): II. Cytogenetics of the relation between solid stem and other morphological characters in hexaploid F5 lines of a hybrid with Rescue (T. aestivum). Can. J. Bot. 1959, 37, 1207–1216.
- Baker, E.P. Basic studies relating to the transference of genetic characters from Triticum monococcum L. to hexaploid wheat. Aust. J. Biol. Sci. 1975, 28, 189–200. [CrossRef]
- Bai, D.; Knott, D.R. Suppression of rust resistance in bread wheat (Triticum aestivum L.) by D-genome chromosomes. Genome 1992, 35, 276–282.
- Innes, R.L.; Kerber, E.R. Resistance to wheat leaf rust and stem rust in Triticum tauschii and inheritance in hexaploid wheat of resistance transferred from T. tauschii. Genome 1994, 37, 813–822.
- Kema, G.H.J.; Lange, W.; Van Silfhout, C.H. Differential suppression of stripe rust resistance in synthetic wheat hexaploids derived from Triticum turgidum subsp. dicoccoides and Aegilops squarrosa. Phytopathology. 1995, 85, 425–429.
- Hanušová, R.; Hsam, S.L.K.; Bartoš, P.; Zeller, F.J. Suppression of powdery mildew resistance gene Pm8 in Triticum aestivum L. (common wheat) cultivars carrying wheat-rye translocation T1BL·1RS. Heredity 1996, 77, 383–387. [CrossRef]
- Nelson, J.C.; Singh, R.P.; Autrique, J.E.; Sorrells, M.E. Mapping genes conferring and suppressing leaf rust resistance in wheat. Crop Sci. 1997, 37, 1928–1935. [CrossRef]
- Hiebert, C.W.; Moscou, M.J.; Hewitt, T.; Steuernagel, B.; Hernández-Pinzón, I.; Green, P.; Pujol, V.; Zhang, P.; Rouse, M.N.; Jin, Y.; et al. Stem rust resistance in wheat is suppressed by a subunit of the mediator complex. Nat. Commun. 2020, 11, 1123. [CrossRef]
- Assefa, S.; Fehrmann, H. Evaluation of Aegilops tauschii Coss. for resistance to wheat stem rust and inheritance of resistance genes in hexaploid wheat. Genet. Resour. Crop Evol. 2004, 51, 663–669. [CrossRef]
- Liu, W.; Danilova, T.V.; Rouse, M.N.; Bowden, B.L.; Friebe, B.; Gill, B.S.; Pumphrey, M.O. Development and characterization of a compensating wheat-Thinopyrum intermedium Robertsonian translocation with Sr44 resistance to stem rust (Ug99). Theor. Appl. Genet. 2013, 126, 1167–1177. [CrossRef]
- Liang, D.Y.; Zhang, M.H.; Liu, X.; Li, H.; Jia, Z.J.; Wang, D.H.; Peng, T.; Hao, M.; Liu, D.C.; Jiang, B.; et al. Development and identification of four new synthetic hexaploid wheat lines with solid stems. Sci. Rep. 2022, 12, 4898. [CrossRef]
- Pauw, R.M.D.; Read, D.W.L. The effect of nitrogen and phosphorus on the expression of stem solidness in Canuck wheat at four locations in southwestern Saskatchewan. Can. J. Plant Sci. 1982, 62, 593–598. [CrossRef]
- Chen, H.H.; Li, J.; Wan, H.S.; Wang, L.L.; Peng, Z.S.; Yang, W.Y. Microsatellite markers for culm wall thickness and anatomical features of solid stem wheat 86-741. Acta Agron. Sin. 2008, 34, 1381–1385.
- Li, Y.H.; Shi, X.H.; Hu, J.H.; Wu, P.P.; Qiu, D.; Qu, Y.F.; Xie, J.Z.; Wu, Q.H.; Zhang, H.J.; Yang, L.; et al. Identification of a recessive gene PmQ conferring resistance to powdery mildew in wheat landrace Qingxinmai using BSR-Seq analysis. Plant Dis. 2020, 104, 743–751. [CrossRef]
- Chen, S.; Zhou, Y.; Chen,Y, Gu, J Fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890.
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: ultrafast universal RNA-Seq aligner. Bioinformatics 2013, 29, 15–21. [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K,; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabrielet, S.; Daly, M.; et al. The genome analysis toolkit: a mapreduce framework for analyzing next-generation DNA sequencing data. Genome Re. 2010, 20, 1297–1303. [CrossRef]
- Lander, E.S.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.J.; Lincoln, S.E.; Newburg, L. MAPMAKER: an interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1987, 1, 174–181. [CrossRef]
- Kosambi, D.D. The estimation of map distances from recombination values. In Selected Works in Mathematics and Statistics; Springer: New Delhi, India, 2016; pp. 125–130.
- Liu, R.H.; Meng, J.L. MapDraw: a microsoft excel macro for drawing genetic linkage maps based on given genetic linkage data. Hereditas 2003, 25, 317–321.
- Luo, M.C.; Gu, Y.Q.; Puiu, D.; Wang, H.; Twardziok, S.V.; Deal, K.R.; Huo, N.X.; Zhu, T.T.; Wang, L.; Wang, Y.; et al. Genome sequence of the progenitor of the wheat D genome Aegilops tauschii. Nature 2017, 551, 498–502. [CrossRef]
- Oiestad, A.J.; Martin, J.M.; Cook, J.; Giroux, M.J. Identification of candidate genes responsible for stem pith production using expression analysis in solid-stemmed wheat. Plant Genome 2017, 10, plantgenome2017.2002.0008. [CrossRef]
- Rutjens, B.; Bao, D.; Van, Eck-Stouten E.V.; Brand, M.; Smeekens, S.; Proveniers, M. Shoot apical meristem function in Arabidopsis requires the combined activities of three BEL1-like homeodomain proteins. Plant J. 2009, 58, 641–654. [CrossRef]
- Tao, Y.; Chen, M.; Shu, Y.J.; Zhu, Y.J.; Wang, S.; Huang, L.Y.; Yu, X.W.; Wang, Z.K.; Qian, P.P.; Gu, W.H.; Ma, H. Identification and functional characterization of a novel BEL1-LIKE homeobox transcription factor GmBLH4 in soybean. Plant Cell Tiss. Org. 2018, 134, 331–344. [CrossRef]
- Smith, H.M.S.; Ung, N.; Lal, S.; Courtier, J. Specification of reproductive meristems requires the combined function of SHOOT MERISTEMLESS and floral integrators FLOWERING LOCUS T and FD during Arabidopsis inflorescence development. J Exp. Bot. 2011, 62, 583–593. [CrossRef]
- Fujimoto, M.; Sazuka, T.; Oda, Y.; Kawahigashi, H.; Wu, J.Z.; Takanashi, H.; Ohnishi, T.; Yoneda, J.; Ishimori, M.; Kajiya-Kanegae, H.; et al. Transcriptional switch for programmed cell death in pith parenchyma of sorghum stems. Proc. Natl. Acad. Sci. 2018, 115, E8783–E8792. [CrossRef]
- Ma, Q.H. The expression of caffeic acid 3-O-methyltransferase in two wheat genotypes differing in lodging resistance. J Exp. Bot. 2009, 60, 2763–2771. [CrossRef]
- Tu, Y.; Rochfort, S.; Liu, Z.Q.; Ran, Y.D.; Griffith, M.; Badenhorst P.; Louie, G.V.; Bowman, M.E.; Smith, K.F.; Noel, J.P.; et al. Functional analyses of caffeic acid O-methyltransferase and cinnamoyl-CoA-reductase genes from perennial ryegrass (Lolium perenne). Plant Cell 2010, 22, 3357–3373.
- Miller, C.N.; Harper, A.L.; Trick, M.; Werner, P.; Waldron, K.; Bancroft, I. Elucidation of the genetic basis of variation for stem strength characteristics in bread wheat by Associative Transcriptomics. BMC Genomics. 2016, 17, 500. [CrossRef]
- Knott, D.R.; Ramanujam, S. Transfer of genes for rust resistance to wheat from related species. Therm Sci. 1978, 16, 513–525.
- Hurni, S.; Brunner, S.; Stirnweis, D.; Herren, G.; Peditto, D.; McIntosh, R.A.; Keller, B. The powdery mildew resistance gene Pm8 derived from rye is suppressed by its wheat ortholog Pm3. Plant J. 2014, 79, 904–913. [CrossRef]
- Wu, L.; Xia, X.C.; Rosewarne, G.M.; Zhu, H.Z.; Li, S.Z.; Zhang, Z.Y.; He, Z.H. Stripe rust resistance gene Yr18 and its suppressor gene in Chinese wheat landraces. Plant Breed. 2015, 134, 634–640. [CrossRef]
- Athiyannan, N.; Long, Y.; Kang, H.; Chandramohan, S.; Bhatt, D.; Zhang, Q.J.; Klindworth, D.L.; Rouse, M.N.; Friesen, T.L.; McIntosh, R.; et al. Haplotype variants of Sr46 in Aegilops tauschii, the diploid D genome progenitor of wheat. Theor. Appl. Genet. 2022, 135, 2627–2639. [CrossRef]
- Jin, H.L.; Zhang, H.P.; Zhao, X.Y.; Long, L.; Guan, F.N.; Wang, Y.P.; Huang, L.Y.; Zhang, X.Y.; Wang, Y.Q.; Li, H.; et al. Identification of a suppressor for the wheat stripe rust resistance gene Yr81 in Chinese wheat landrace Dahongpao. Theor. Appl. Genet. 2023, 136, 67. [CrossRef]
- Huang, L.; Brooks, S.A.; Li, W.L.; Feller, J.P.; Trick, H.N.; Gill, B.S. Map-based cloning of leaf rust resistance gene Lr21 from the large and polyploid genome of bread wheat. Genetics 2003, 164, 655–664. [CrossRef]
- Arora, S.; Steuernagel, B.; Gaurav, K.; Chandramohan, S.; Long, Y.M.; Matny, O.; Johnson, R.; Enk, J.; Periyannan, S.; Singh, N.; et al. Resistance gene cloning from a wild crop relative by sequence capture and association genetics. Nat. Biotechnol. 2019, 37, 139–143. [CrossRef]
- Dyck, P.L. Genetic inhibition of expression of resistance gene Lr23 in wheat to Puccinia recondita. Can. J. Plant Sci. 1982, 62, 219–220. [CrossRef]
- Kerber, E.R.; Green, G.J. Suppression of stem rust resistance in the hexaploid wheat cv. Canthatch by chromosome 7DL. Can. J. Bot. 1980, 58, 1347–1350. [CrossRef]
- Kerber, E.R.; Stem-rust resistance in 'Canthatch' hexaploid wheat induced by a nonsuppressor mutation on chromosome 7DL. Genome 1991, 34, 935–939. [CrossRef]
- Williams, N.D.; Miller, J.D.; Klindworth, D.L. Induced mutations of a genetic suppressor of resistance to wheat stem rust. Crop Sci. 1992, 32, 612–616. [CrossRef]
- Talajoor, M.; Jin, Y.; Wan, A.; Chen, X.M.; Bhavani, S.; Tabe, L.; Lagudah, E.; Huang, L. Specificity of a rust resistance suppressor on 7DL in the spring wheat cultivar Canthatch. Phytopathology 2015, 105, 477–481. [CrossRef]
- Mclntosh, R.A. Genetics of resistance to pathogens and pests: Recent developments. Proc. 8th Int. Wheat Genet. Symp. 1995, 2, 889–896.
- Hao, M.; Zhang, L.Q.; Zhao, L.B.; Dai, S.F.; Li, A.L.; Yang, W.Y.; Xie, D.; Li, Q.C.; Ning, S.Z.; Yan, Z.H.; et al. A breeding strategy targeting the secondary gene pool of bread wheat: introgression from a synthetic hexaploid wheat. Theor. Appl. Genet. 2019, 132, 2285–2294. [CrossRef]
Figure 1.
The expression differences of TdDof gene in Ma, Syn-SAU-119, Syn-SAU-117 and Cocorit at Zadoks Stage 32 and Zadoks Stage 34: (a) Expression differences of TdDof gene at the Zadoks Stage 32; (b) Expression differences of TdDof gene at the Zadoks Stage 34.
Figure 1.
The expression differences of TdDof gene in Ma, Syn-SAU-119, Syn-SAU-117 and Cocorit at Zadoks Stage 32 and Zadoks Stage 34: (a) Expression differences of TdDof gene at the Zadoks Stage 32; (b) Expression differences of TdDof gene at the Zadoks Stage 34.
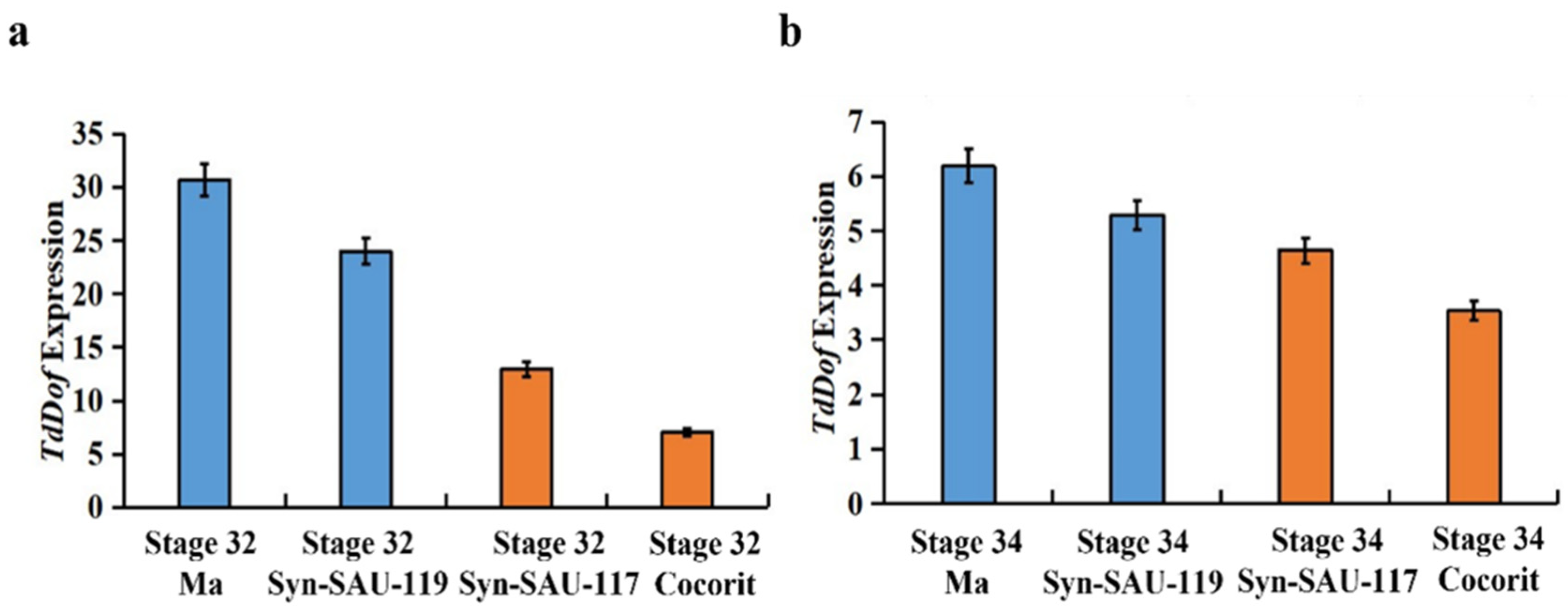
Figure 2.
The stem solidity of Syn-SAU-119 and Syn-SAU-117 and F1、F2 individual plants in the greenhouse: (a) Syn-SAU-119; (b) Syn-SAU-117 × Syn-SAU-119 F1; (c) Syn-SAU-117; (d) Solid F2 plant; (e)Semisolid F2 plan.
Figure 2.
The stem solidity of Syn-SAU-119 and Syn-SAU-117 and F1、F2 individual plants in the greenhouse: (a) Syn-SAU-119; (b) Syn-SAU-117 × Syn-SAU-119 F1; (c) Syn-SAU-117; (d) Solid F2 plant; (e)Semisolid F2 plan.
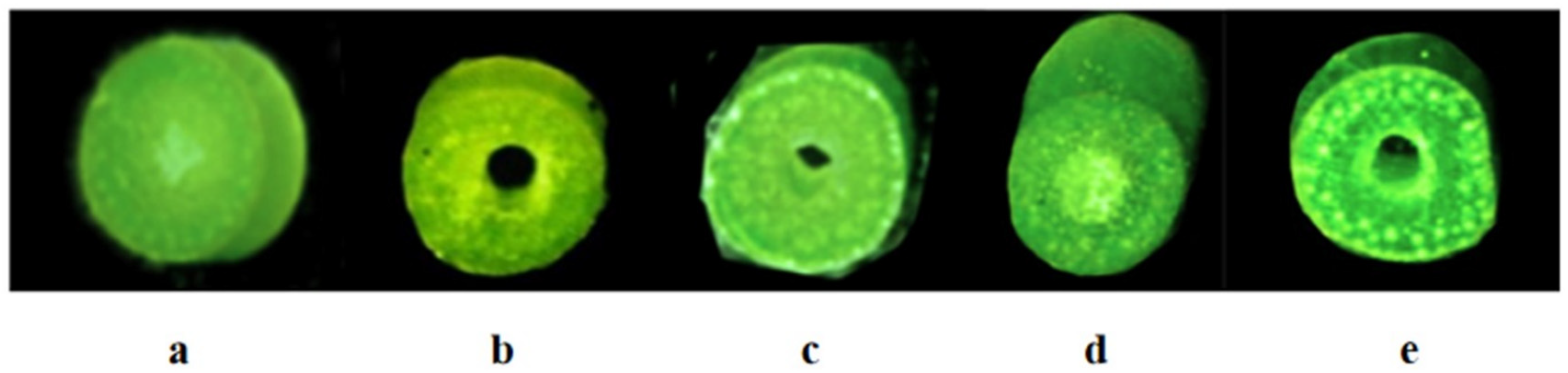
Figure 3.
Anatomical structure of stems of Syn-SAU-119 and Syn-SAU-117 and F1、F2 individual plants in the greenhouse: (a) Syn-SAU-119; (b)Syn-SAU-117 × Syn-SAU-119 F1; (c) Syn-SAU-117; (d) Solid F2 plant; (e) Semisolid F2 plant.
Figure 3.
Anatomical structure of stems of Syn-SAU-119 and Syn-SAU-117 and F1、F2 individual plants in the greenhouse: (a) Syn-SAU-119; (b)Syn-SAU-117 × Syn-SAU-119 F1; (c) Syn-SAU-117; (d) Solid F2 plant; (e) Semisolid F2 plant.
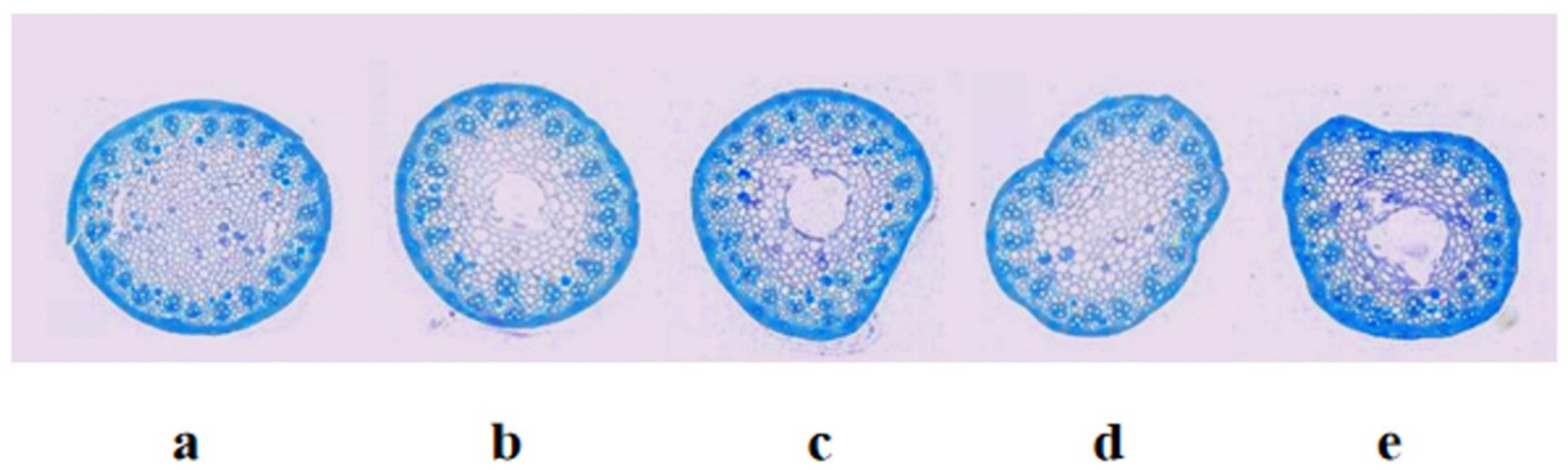
Figure 4.
Distribution of SNPs (AFD > 0.6, P-value<1e-8) on 21 chromosomes.
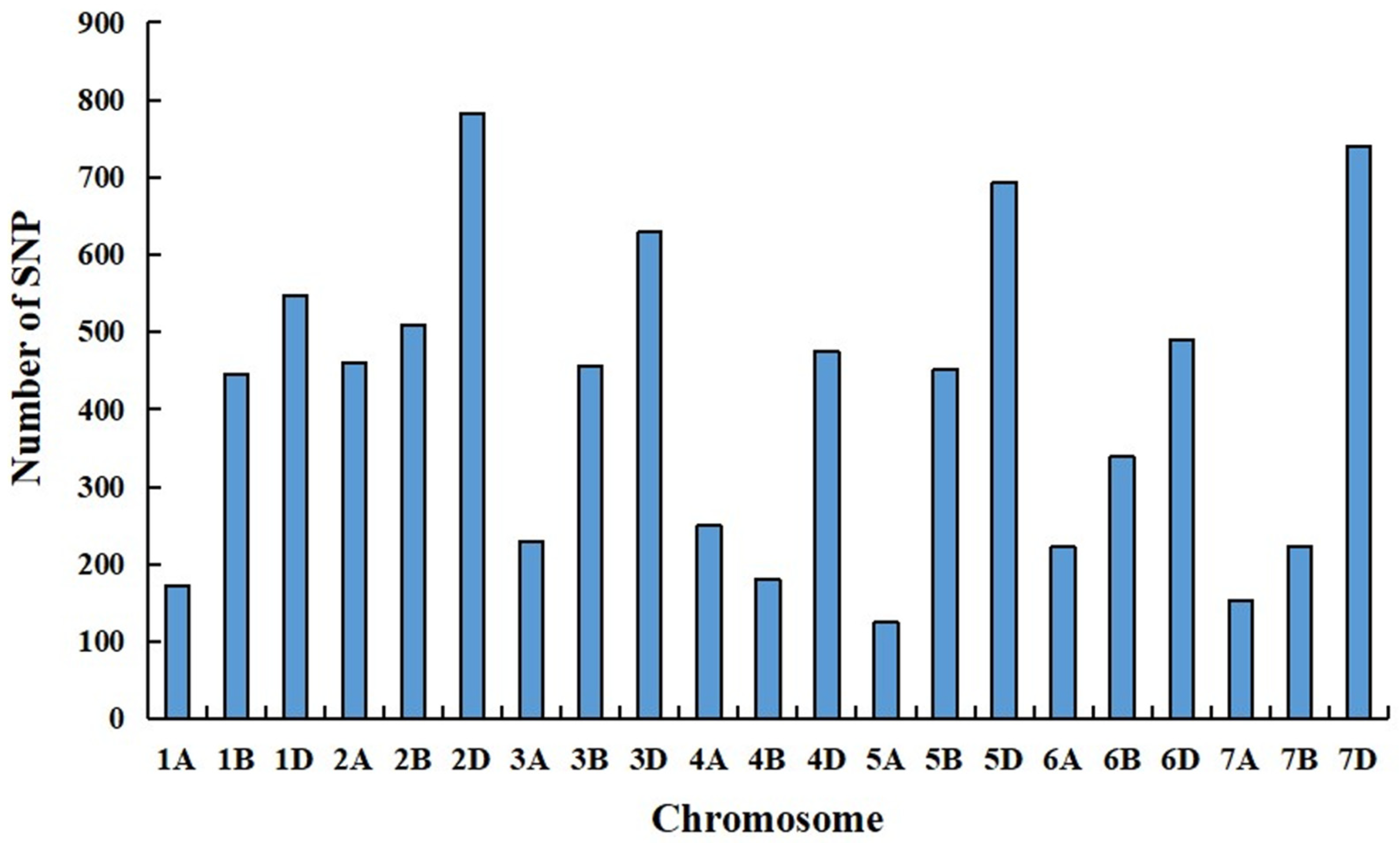
Figure 5.
The enrichment of SNPs within 1 Mb window size on wheat chromosomes.

Figure 6.
Genetic linkage map of Su-TdDof gene on chromosome 7DS showing physical location of Su-TdDof: (a) Linkage map of Su-TdDof; (b)The physical interval (blue part) where the four KASP markers linked to Su-TdDof anchored in Chinese Spring, orange dot with dotted lines indicating the physical positions of each marker; (c)Physical intervals anchored by markers linked to Su-TdDof in Aegilops tauschii.
Figure 6.
Genetic linkage map of Su-TdDof gene on chromosome 7DS showing physical location of Su-TdDof: (a) Linkage map of Su-TdDof; (b)The physical interval (blue part) where the four KASP markers linked to Su-TdDof anchored in Chinese Spring, orange dot with dotted lines indicating the physical positions of each marker; (c)Physical intervals anchored by markers linked to Su-TdDof in Aegilops tauschii.
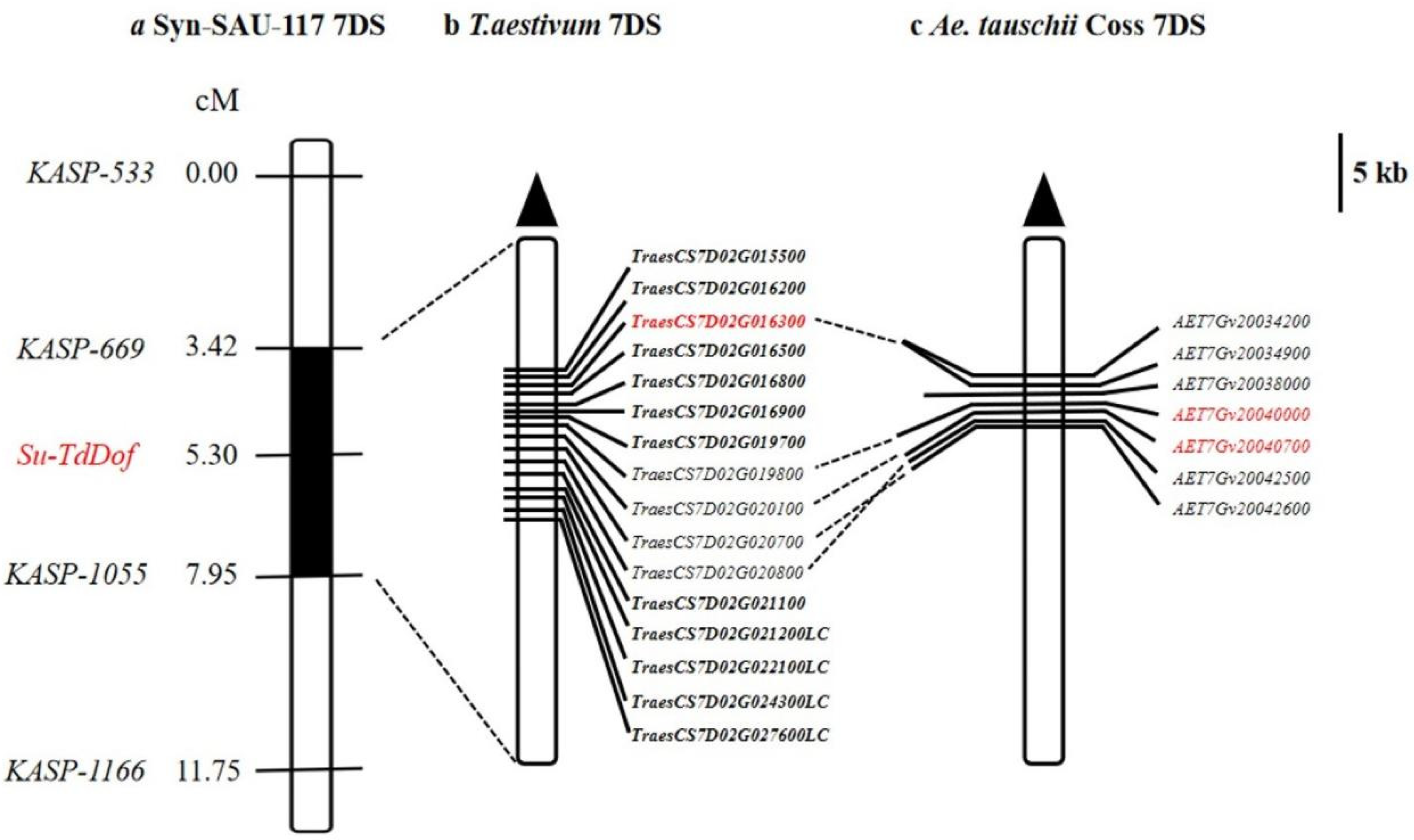
Figure 7.
Volcano map of differentially expressed genes.
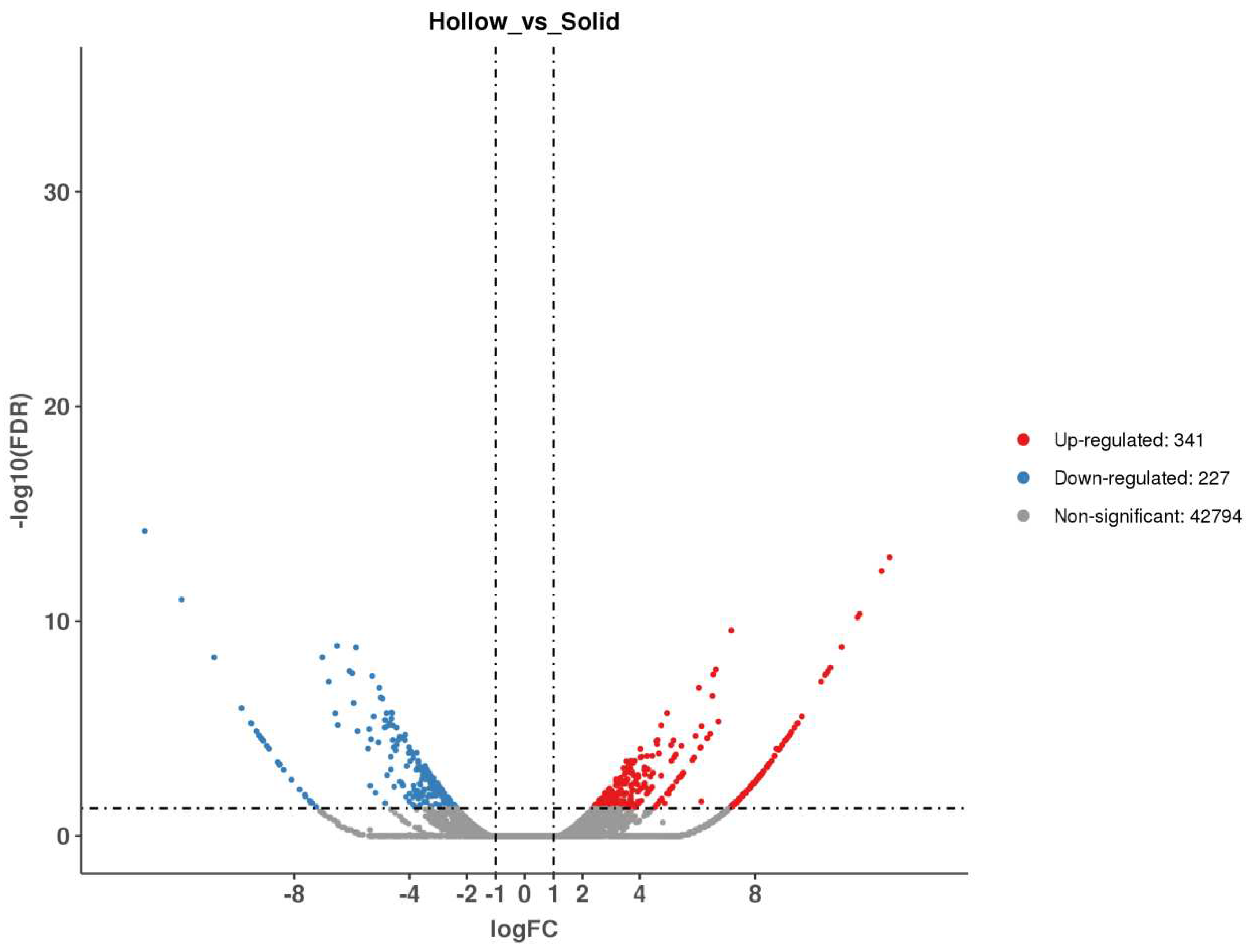

Table 1.
Genetic analysis of solid stem suppressor genes in F1, F2, and F2:3 families of Syn-SAU-119 × Syn-SAU-117.
Table 1.
Genetic analysis of solid stem suppressor genes in F1, F2, and F2:3 families of Syn-SAU-119 × Syn-SAU-117.
| Parents and cross | Generationa | No. of plants/families | Observed ratiob | Actual ratio | Expected ratio | χ2 | P-value | ||
| S | Seg | Ss | |||||||
| Syn-SAU-117 | PS | 20 | 20 | ||||||
| Syn-SAU-119 | PH | 20 | 20 | ||||||
| PSs × PS | F1 | 20 | 20 | ||||||
| F2 | 156 | 36 | 120 | 0.9:3 | 1:3 | 0.308 | 0.579 | ||
| F2:3 | 134 | 35 | 67 | 32 | 1.04:2:0.96 | 1:2:1 | 0.134 | 0.935 | |
aPS: solid parent Syn-SAU-119; and PSs: semisolid parent Syn-SAU-117. bS: homozygous solid; Seg: segregating within F2:3 families; Ss: homozygous semisolid.
Table 2.
Primer sequences of KASP markers used for genetic mapping of Su-TdDof.
| Marker | Physical position (bp) | Allele 1 primera | Allele 2 primerb | Common/reverse primer |
|---|---|---|---|---|
| KASP-533 | 5336907 | TCAGCTTCAATTTCGGCAGC | TCAGCTTCAATTTCGGCAGT | AGAAGCTGAACGTGCGGAAG |
| KASP-669 | 6695986 | GTCGGATTCGGTTACTTTGAC | GTCGGATTCGGTTACTTTGAT | AGAGGTGCATGGTGTCGT |
| KASP-1055 | 10558194 | TCTTTCTCCTTCAGCCTCTTA | TCTTTCTCCTTCAGCCTCTTG | GCCTGATTGTAGTACATTATG |
| KASP-1166 | 11664145 | AACGAGGTCCCGCGCTCCTCCC | AACGAGGTCCCGCGCTCCTCCG | GTGTGAAGAGCGCTTCTGC |
a A1 primer labelled with FAM: GAAGGTGACCAAGTTCATGCT. b A2 primer labelled with HEX: GAAGGTCGGAGTCAACGGATT.
Table 3.
Genotyping of AS92, Syn-SAU-117, Syn-SAU-119, and AS96 using KASP markers linked to Su-TdDof.
Table 3.
Genotyping of AS92, Syn-SAU-117, Syn-SAU-119, and AS96 using KASP markers linked to Su-TdDof.
| Parents | Marker genotypea | |||
|---|---|---|---|---|
| KASP-533 | KASP-669 | KASP-1055 | KASP-1166 | |
| AS92 | CC | CC | AA | CC |
| Syn-SAU-117 | CC | CC | AA | CC |
| Syn-SAU-119 | TT | TT | GG | GG |
| AS96 | TT | TT | GG | GG |
a AA, CC, GG, and TT represent the haplotype results of SNP genotyping.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



