
Submitted:
20 July 2023
Posted:
21 July 2023
You are already at the latest version
Abstract
Vanadium is a well-known essential trace element, which usually exists in oxidation states in form of vanadate cation intracellularly. The pharmacological study of vanadium begins at the discovery of its unexpected inhibitory effect on ATPase. Thereafter, the protective effects on cells and the abilities in glucose metabolism regulation were observed from vanadium compound, leading to the application of vanadium compounds in clinical trials for curing diabetes. Alzheimer’s dis-ease (AD) is the most common dementia disease in elderly people. However, there is still no efficient agents for treating AD safely to date. This is mainly because of the complexity of the pathology, which are characterized by the senile plaques composed by amyloid-beta (Aβ) protein in the parenchyma of brain and the neurofibrillary tangles (NFTs) derived from hyperphosphorylated tau protein in neurocyte, along with mitochondrial damage, and eventually the central nervous system (CNS) atrophy. AD was also illustrated as type-3 diabetes, because of the observations of insulin deficiency and the high level of glucose in cerebrospinal fluid (CSF), as well as the im-paired insulin signaling in brain. In this review, we summarized the advance of applicating vanadium compound on AD treatment in experimental research and pointed out the limitation of the current study on using vanadium compounds in AD treatment. We hope it will help the future study in this field.
Keywords:
Vanadium
; Alzheimer’s disease
; diabetes
; insulin resistance
; mitochondrial
; oxidative phosphorylation
1. Introduction
Vanadium is an essential trace element which plays important role in the metabolism of cholesterol and triglyceride, as well as the oxidation of glucose and the synthesis of glycogen [1]. Vanadium usually existed in form of vanadate anion (VO3−) extra-cellularly, and in form of vanadyl cation (VO2+) intracellularly in body, respectively [2]. Since the ATPase inhibitory effects of vanadate were observed by accident [3], it had been well documented that vanadate was similar with phosphate in size and charge, which gave vanadium the ability of irreversibly prohibiting the conformation-al variety of dephosphorylate enzyme [4]. Later on, vanadyl ions showed insulin-like feature in rat adipocytes [5]. Further studies revealed that peroxovanadates inhibited the function of protein tyrosine phosphatase (PTPase) [6], which was involved in the dephosphorylation of insulin receptor and arrest the insulin signaling. It was also demonstrated that vanadyl bisacetylacetonate exerted an antilipolytic influence via activating Akt (protein kinase B, PKB) [7, 8], a key kinase that downstream of insu-lin-PI3K (phosphatidylinositol-3-kinase) signaling pathway. Moreover, it was reported that the protein level of PPARγ (peroxisome proliferator-activated receptor gamma) , a transcriptional factor which was shown to reduce insulin resistance [9] upon activation in -pancreas cells [10] and adipocytes [11], was modulated by vanadyl bisacety-lacetonate.
Though the biological functions of vanadium were well documented, the toxicity was also seen in animal studies [12, 13]. Clinical study revealed that the consumption of vanadium at 125 mg/day was safe for adults [14]. However, rats would all die when they received vanadyl sulfate more than 2 mM/kg body weight [15]. The biological effects are different according to the species of vanadium compounds [16], the toxicities of vanadium in different oxidation state were divergent as well. Studies illustrated that the highest oxidated valence (+5) of vanadium was the most toxic state [17]. Be-cause in this state, the strong prooxidant property of vanadium would severely aggrandize oxidative stress [18] and perturb mitochondria [19]. In pharmacological studies, many vanadium compounds, such as, bis(maltolato) oxovanadium(IV) (BMOV) [20], bis(2-ethyl-3-hydroxy-4-pyronato) oxovanadium (IV) (BEOV) [21] , N,N-dimethylphenylenediamine-derivatized nitrilotriacetic acid vanadyl complexes (VO(dmada)) [22], vanadyl complex of p-hydroxyl aminophenol derivative (VOphpa-da) [23], and graphene quantum dots(GQD)-VO(p-dmada) [24], have been synthesized to improve the affordability and stability of inorganic vanadium salts. It was shown that oral uptake of BEOV increased the absorbance of vanadium 2-3 times in most tis-sues than VOSO4 [25].
Interestingly, it was reported that the level of vanadium was declined in the plasma of Alzheimer’s disease (AD) patients [26, 27], indicating that this trace element may get involved in AD pathology. Recently, the protective effects of vanadium com-pounds on AD pathology were observed in different AD mice models. In this article, we reviewed the emerging role of vanadium compounds in AD treatment and the un-derlying mechanisms of these agents.
2. The Deficits of Insulin Signal in AD
2.1. The Role of APP in Glucose Metabolism
The extracellular deposits of amyloid-beta (Aβ) plaques and the intracellular neurofibrillary tangles (NFT) formed by hyperphosphorylated tau are well known his-topathological characters of AD, which are accompanied by damaged mitochondrial in neuron and the severe atrophy of central nervous system (CNS).
It seems that the level of Aβ correlated with onset of AD, as indicated by the fa-milial AD (FAD) patients, those who carrying the mutations on amyloid precursor protein (APP) [28], a disintegrin and metalloproteinase 10 (ADAM10, an -secretase) [29, 30] and/or presenilin-1/2 (PS1/2, the components of -secretase) [31] which give rise to overproduction of Aβ, probably suffer AD during their younger life. The typical neuropathology of AD seen in Down’ syndrome also emphasized the toxicant of Aβ The gene encoding APP is located on chromosome 21. The trisomy 21 patients harbor-ing 3 copies of APP exhibited abundant diffused Aβ plaques in their CNS and invaria-bly get AD pathologies after certain age. Aβ is produced by the cleavage of APP, which is a type 1 transmembrane protein, by -secretase (BACE1) and -secretase to generate 3 fragments including a soluble APP ectodomain, an Aβ domain, and an APP intra-cellular domain (AICD). However, when APP is hydrolyzed by -secretase, it will gen-erate the APP ectodomain, which is longer than APP ectodomain, without produc-ing the Aβ fragment. Thus, this is not an amyloidogenic process.
The toxicities of Aβ have been intensively studied. It was observed that Aβ oligo-mers could assemble to form pores on cell membranes for ion transportation and im-pair the appropriate permeability of membranes [32], which resulted in the depolari-zation of microglia and neuron [33]. The soluble Aβ could also impair the synaptic plasticity through over-activating NMDA receptor [34], which resulted in mitochon-dria damage [35, 36]. In addition, Aβ oligomers was demonstrated to induce inflam-matory reactions through toll-like receptor [37] and perturb the blood-brain barrier [38]. Although, Aβ overproduction is considered as the most pivotal risk factor for AD development, it was observed that many elderly non-dementia persons also carrying Aβ plaques in their brains. Recently, it has been confirmed that the severity of demen-tia is dependent on the NFT burden but not the level of Aβ senile depositions [39]. Therefore, many scientists suggested that AD should be considered as a secondary tau pathology. This idea is also supported by the discovery that two persons who carrying PS1-E280A mutation, which usually resulted in typical AD before 50 years old, did not get dementia before age 70. They all had severe Aβ plaques burden in their brains, but they did not develop tau pathology in brains as other PS1-E280A mutation carriers did. One of them is a APOE3-R136S homozygote [40], the other one is a RELN-H3447R mutation carrier [41].
Is the function of APP aimed to produce Aβ which is a toxicant for brain? The answer must be not. It has been found that APP played important roles in glucometa-bolic. For example, the App knockout mice had reduced plasma glucose than the wild-types (WT) [42]. When mice were treated with glucose or a membrane-permeant cAMP, the insulin secretion in App knockout mice was increased much higher than that in WT [43]. More interestingly, the APP deficiency resulted in mice being resistant to diet-induced obesity and having higher energy expenditure at night [44]. Meanwhile, the level of insulin was lower in brains of App ablated mice, because of the increase of insulin-degrading enzyme (IDE), and the synaptosomes prepared form App ablated mice showed diminished insulin receptor phosphorylation compared with WT mice [45]. On the other hand, the APP fragment of APP, which is generated by -secretase hydrolyzation, also modified the phosphorylation of Akt [46], indicating that APP it-self is involved in glucometabolic.
2.2. The Influence of Tau on Insulin Signal
Despite the terrible toxicity of Aβ seen in vivo and in vitro, a great many elder people bearing Aβ plaques in their brains did not exhibit dementia symptom until tau pathology appeared [47]. This may due to the sequestration of Aβ plaques by microglia [48]. Tau is a microtubule-associated protein which was believed to stabilize microtu-bule facilitate cargo transport. It is encoded by MAPT on chromosome 17. In human brain, the exons 2, 3, and 10 of MAPT can be alternatively spliced, the former two en-coding two N-terminal repeats (N), while the later one encoding a microtu-bule-binding repeat (R) domain. There are 4 microtubule-binding repeats in total. Therefore, alternative splicing of MAPT will produce 6 distinct tau isoforms, which are 0N3R, 1N3R, 2N3R, 0N4R, 1N4R, 2N4R. All of them could be detected in the paired helical filaments of AD.
A great many efforts have been made to disclose how Aβ can trigger tau patholo-gy, thus to integrate the conventional Aβ cascade hypothesis of AD pathophysiology. It was found that Aβ oligomer activated Fyn through prion protein (PrP) [49], leading to the hyperphosphorylation of tau [50]. It was also demonstrated that oligomeric Aβ overstimulated N-methyl-D-aspartate receptor (NMDAR), which in turn triggered cyclin dependent kinases 5 (CDK5) activation and tau phosphorylation [51] (Figure 1). In addition, it was shown that Aβ was able to attenuate insulin signaling and activate glycogen synthase kinase -3 (GSK-3β, which resulted in tau phosphorylation [52]. Moreover, Aβ was found to increase tau proteolysis at Asp421 and exacerbate the rate and extent of tau filament assembly in vitro [53].
Notably, there are many other tau pathologies besides AD, such as, Pick’s disease (PiD), chronic traumatic encephalopathy (CTE), argyrophilic grain disease (AGD), cor-ticobasal degeneration (CBD), progressive supranuclear palsy (PSP), and a subclass of frontotemporal dementia with Parkinsonism linked to chromosome 17 (FTDP-17tau) [54]. Tau is hyperphosphorylated not only in the NFT of AD but also in other tau pa-thologies. There are many kinases involved in tau hyperphosphorylation, including death-associated protein kinase 1 (DAPK) which is also associated with late-onset of AD [55] [56], Ca2+/Calmodulin-dependent protein kinase II (CAMKII) which is in-volved in LTP formation [57], and Fyn, CDK5, GSK-3β as forementioned above. On the other side, the disfunction of protein phosphatase 2A (PP2A) was also shown to be re-sponsible for intensive phosphorylation of tau [58]. The hyperphosphorylation of tau resulted in the dissociation of tau and microtubules [59, 60]. However, it seems that the NFT itself was not sufficient to cause cognitive decline or neuronal death alone [61].
The acetylated tau was also seen in tauopathies due to the dysregulation of both p300 acetyltransferase and sirtuin 1 (SIRT1) deacetylase [62]. The acetylation of tau inhibited chaperone mediated clearance of tau and promoted tau propagation in mice [63]. Inhibition of p-300 induced tau acetylation by salsalate reduced tau level and prevented hippocampal atrophy [64]. Attractively, it was found that the acetylation of tau was significantly enhanced in high glucose treated cells. In contrast, the activation of AMP-activated protein kinase (AMPK) ameliorated acetylation of tau and rescue memory impairments in a SIRT1 dependent manner in mice model [65]. AMPK is in-volved in glucose metabolism. Upon activating by liver kinase B1 (LKB1), transforming growth factor -activated kinase 1 (TAK1), AMPK can regulate the level of peroxisome proliferator activated receptor gamma coactivator 1 α (PGC-1α[66].
Before asking the neurotoxicity of tau phosphorylation and/or aggregation, one may want to know the basic functions of tau itself. Indeed, except for binding to mi-crotubule, tau is involved in regulating insulin signaling as well. It was found that tau interacted with tension homologue on chromosome 10 (PTEN) and exert an inhibitory effect on its lipid phosphatase activity. Knockout of tau resulted in the activation of PTEN, and the dephosphorylation of PtdIns(3,4,5), thus impaired the hippocampal re-sponse to insulin induced LTD in brain slides [67]. It was also reported that tau abla-tion in mice lead to pancreatic cell dysfunction and glucose intolerance [68]. Besides, tau knockdown increased basal insulin level, but perturbed glucose-stimulated insulin secretion [69]. Interestingly, it was also observed that the phosphorylation of tau re-sulted in intraneuronal accumulation of insulin oligomers and insulin signaling defi-cits [70]. However, in streptozotocin (STZ) induced type 1 diabetes model mice, tau knockout attenuated the cognitive impairment triggered by insulin deficiency [71]. Whereas, in the same conditions human tau transgenic mice showed severe impair-ments in learning and memory [72]. In addition, in P301L mutation knock-in male mice but not female mice, the high-fat diet triggered higher insulinemia and glucose intolerance by comparing with wild type littermates [73]. These studies suggested that tau is closely correlated with insulin signaling and glucometabolic instead of only par-ticipating in microtubule stabilization.
Aβ overproduction resulted in Ca2+ influx through NMDAR, which in turn activated CDK5 via calpain mediate cleavage of p35 into p25. CDK5 subsequently phosphorated tau and suppressed the activity of PPARγ. Tau phosphorylation and truncation will impair the functions of mitochondria and increase the level of ROS. However, vana-dium can activate PPARγ, which is involved in facilitating insulin secretion and main-taining insulin receptor activation through upregulating GIP receptor and IRS, thus restraining the hyperphosphorylation of tau, on the other hand, the activation of PPARγ by vanadium may protect mitochondria from the accumulation of ROS by downregulating the level of cyclooxygenase-2 (COX2) and inducible nitric oxide syn-thase (iNOS).
2.3. The Impaired Insulin Signal in AD
The “Type 3 diabetes” was first used to describe AD by Steen, E.; et al., [74] for the abnormal levels of insulin, and glucose in CSF [75], as well as the insulin resistance that were found in brains of AD patients [76]. Type 3 diabetes is not a medical ap-proved term though, it has been demonstrated that Ab oligomer interrupted the acti-vation of PI3K and abolished the suppression of insulin on GSK-3β, which is involved in triggering the hyperphosphorylation of tau besides the energy metabolism [77]. In addition, the IDE is able to decompose both insulin and Aβ [78]. In IDE deficient mice, the level of endogenous soluble Aβ was elevated brain n the contrary, overexpression of IDE in the neuron of APP transgenic mice significantly reduced the level of soluble Aβ and postponed the formation of amyloid plaque. Interestingly, in brain of those who carrying apolipoprotein E-epsilon 4 (APOE4), the most significant genetic risk factor for sporadic AD, the protein level of IDE was reduced by approximately 50% [79]. However, in the blood-brain barrier of AD with cerebral amyloid angiopathy (CAA) the level of IDE was enhanced [80], which may impair the transportation of insulin from periphery to CNS. Moreover, when insulin was depleted in mice, both of tau phosphorylation and tau filaments were reinforced in brains [81]. In line with these observations, depleting insulin by STZ also triggered tau phosphorylation and NFT formation [82]. Moreover, when insulin receptor substrate 2 (IRS2) was lost, the phosphorylation of tau had been promoted [83]. Taken together, these evidences coin-cidentally demonstrated that insulin signaling pathway disfunction may play a pivotal role between Aβ overproduction and tau pathology.
In brains, insulin can either derived from in situ de novo synthesis [84] or from the peripheral plasma. Insulin can pass across the capillary endothelial cells of BBB in saturable, selective, receptor dependent manners [85, 86]. Through stimulating insulin receptor (IR) and/or insulin-like growth factor 1 receptor (IGF1R), insulin facilitated the phosphorylation of insulin receptor substrate (IRS), and subsequently activated PI3K and AKT (Figure 1). As a result, the glucose transporter 4 (GLUT4) in cytosol were translocated onto the plasma membrane to enhance the glucose uptake [87]. In-sulin triggered translocation of GLUT4 is very critical in the process of hippocampal dependent memory consolidation [88]. Of note, the insulin signaling pathway was reg-ulated by negative feedback. Except for stimulating Rho GTPase to facilitate the transportation of GLUT4, the activation of Akt also induced the functioning of mam-malian target of rapamycin complex 1 (mTORC1), which is sensitive to Rapamycin. mTORC1 can further stimulate ribosomal protein S6 kinase (S6K), which will inhibit the activity of IRS1, thus silencing the insulin-PI3K-Akt signal. mTORC1 is also in-volved in regulating some other cellular process, including autophagy and mitochon-drial oxidative respiration. Upon binding to its receptor, insulin can also trigger the ac-tivation of growth factor receptor-bound protein 2 (Grb2), which will further stimulate Ras, Raf and mitogen-activated protein kinases (MAPK) [89]. Notably, hyperactivation of mTORC1 was spotted in early to mid-stage of AD brains [90]. In terms of MAPK, except from being stimulated by insulin signal, the overreaction of p38 was also impli-cated in Aβ induced toxicity [91].
The dysregulation of insulin signal were also seen in APOE4 carrier, it was found that the insulin receptor were trapped in the endosomes in primary neurons treated by APOE4 [92]. In addition, knockout of triggering receptor expressed on myeloid cells 2 (TREM2), which is a great genetic risk factor following APOE4, also exacerbated insu-lin resistance [93]. Interestingly, the insulin resistance upregulated the expression of GCN5, a histone acetyltransferase, which resulted in the increase of CDK5 and tau phosphorylation [94]. These data indicated that the Aβ overproduction and genetic risk factors of AD can directly and indirectly impair insulin signal, therefore triggering tau phosphorylation. On the other hand, the disfunction of tau may further induce in-sulin resistance and/or insulin deficiency in AD brain.
3. The Effects of Vanadium on Curing AD
It was shown that the administration of insulin can reduce the ratio of tau-phosphorylated tau-181/Aβ42 in plasma and maintained the volume of AD brains [95]. However, the long-term insulin administration probably triggers insulin re-sistance. In contrast, the intranasal administration allowed insulin arriving in CNS bypasses the periphery and prevents the risks associated with hypoglycemia [96]. The study on rats indicated that intranasal insulin supply was able to improve memory and inhibit the inflammation in AD [97]. The clinical study also indicated that verbal memory of MCI and AD patients without APOE4 were improved immediately after 40 IU intranasal insulin supply, without perturbing the plasma levels of insulin and glu-cose [98]. Another trial showed that 40 IU/day intranasal insulin administration for 21 days significantly ameliorated the working verbal memory and working visuospatial memory [99]. Nevertheless, these effects were affected by the APOE alleles. Insulin administration alleviated insulin resistance only in APOE4 carriers but not in APOE3 or APOE2 carriers [100] . Nonetheless, a recently study reported that the intranasal insulin administration exhibited no benefits on cognitive functions in a randomized clinical trial including 289 adults with mild cognitive impairment or AD [101]. How-ever, this study had a profound limitation, which is the device that used in this study for intranasal insulin administration had not been tested in before. Therefore, further researches on this field are still needed to illustrate the effects of insulin on AD curing and the underling mechanism.
In other studies, the effects of insulin sensitizers, which showed benefits on curing type-2 diabetes, were tested in curing AD on different mice models or clinical trials. The peroxisome proliferator-activated receptor (PPAR-γ) agonists, such as, rosiglita-zone [102] pioglitazone [103], showed great benefits on AD pathologies. 6 months of rosiglitazone administration for 4 mg/day significantly improved the selective atten-tion and delayed recall of AD patients. In addition, 6 months of pioglitazone supplica-tion for 10-30 mg/day decreased fasting plasma insulin levels of AD patients who also suffered from type2 diabetes mellitus. Meanwhile, the plasma Aβ levels of these sub-jects were declined by comparing with the AD patients in control group who received placebo [102, 104]. Another study showed that 24 weeks of rosiglitazone administra-tion at 8 mg/day significantly ameliorated the performance of the APOE4 negative AD patients [105]. Nonetheless, a phase 3 trial demonstrated that rosiglitazone had no ef-fects on the cognitive functions of AD, regardless of APOE type [106]. In addition, it was found that metformin increased the IDE level in transgenic AD mice [107], and prevented amyloid plaque deposition and memory impairment [108]. In vitro study also revealed that metformin induced dephosphorylation of tau through PP2A [109]. Clinically, the use of metformin showed protective effects on brain volumes in non-demented elderly individuals with diabetes [110]. For mild cognitive impairment or mild dementia due to AD, metformin also improved executive functioning [111].
Vanadium (IV) compound could rescue cholinergic neurons in the medial septum of bilateral olfactory bulbectomy mouse in a dose dependent manner. The impaired long-term potentiation (LTP) of these mice was also prevented by bis(1-N-oxide-pyridine-2-thiolato)oxovanadium(IV) (VO-(OPT)) [112]. However, the mechanisms have not been studied deeply. The vanadyl (IV) acetylacetonate (VAC) was found to attenuate neuron loss in APP/PS1 transgenic AD model mice, and pre-served cognitive functions. It up-regulated the expression of glucose-regulated protein 75 (Grp75), thus suppressed p53-mediated neuronal apoptosis without reducing Aβ plaques in the mice brain. Furthermore, the neuroprotective ability of VAC is corre-lated the activation of PPARγ and AMPK signaling [113]. Another vanadium com-pound, BEOV significantly reduced the phosphorylation of tau, and inhibited the Aβ induced inflammation by inhibiting NF-κB signal both in vitro and in vivo [114]. BEOV also blocked the neurotoxicity induced by endoplasmic reticulum (ER) stress through inhibiting Bip and p-eIF2α [115], and ameliorated the spatial learning and memory in AD mouse models [116]. More importantly, we found that the biological benefits of BEOV on AD pathologies are dependent on PPARγ [115, 117, 118], which resembled the functions of bis (5-hydroxy-4-oxo-4H-pyran-2-hydroxy-benzoatato) oxovanadium (IV) (BSOV) [119].
PPARγ is a member of nuclear hormone receptor family of ligand-inducible tran-scription factors, which plays a pivotal role in lipid and glucose homeostasis. The ac-tivity of PPARγ could be inhibited by CDK5 and MAPK [120] (Figure 1). It was report-ed that the activation of PPARγ is involved in upregulating the level of glu-cose-dependent insulinotropic polypeptide (GIP) receptor [121], GLUT4, and pyruvate carboxylase [122, 123], which are corelated with the insulin sensitization. In addition, the activation of PPARγ is able to repress the NF-κB-dependent transcription of iNOS and COX2 [124], which are involved in the generation of ROS. Molecular docking analysis revealed that the binding energy of BEOV with PPARγ was ~8.1 kcal mol-1, indicating that BEOV interacts quite well with PPARγ and may be a agonist for PPARγ[115]. Though the vanadium compound showed great protective effects on transgenic AD model mice, it is still unknown whether these anti-diabetes agents are valid in later stages of AD which is featured by severe neurotrophy accompanied with the propagation of prion-liked tau.
4. The Potential Mechanisms of Vanadium in Curing AD for Future Study
Tau is localized in mitochondria besides associating with microtubule [125]. In neuron expressing mutated tau that found in FTLD, the hyperphosphorylated tau im-paired the function of mitochondria by breaking down the complex I of electron transport chain [126, 127]. In addition, it was demonstrated that the hyperphosphory-lated tau also promoted mitochondrial fission and morphology change through inter-acting with dynamin-related GTPase (Drp1) [128]. Moreover, it was shown that the cleavage of tau promoted the formation of NFTs [129]. Importantly, the cleaved tau perturbed the mitochondrial dynamics when the intracellular calcium level was in-creased by thapsigargin treatment as well [130, 131]. On the other hand, the reduction of tau has been found to protect neuron from loss of mitochondrial membrane poten-tial loss [132], excitotoxicity [133] and axonal transport inhibition [134] that induced by Aβ[135].
The functions of tau on mitochondrial metabolism and homeostasis attracted more and more attentions these days. It has been shown overexpression of human tau resulted in mitochondrial elongation and accumulation, along with the reduced ubiq-uitination of mitofusion 2 (MFN2) [136]. Moreover, in the mutated human tau (P301) transgenic mice [137], 3x transgenic AD mice [138] and AD patients [139], the level of MFN2 was reduced [140]. In fly, the overexpression of tau affected the expression of drp1 and Marf (the homologous to human MFN2) [141]. The level of MFN1/2 were re-duced in APOE4 carriers [142]. By analyzing genotypes and allele frequencies in Ko-rean AD population, the rs1042837 polymorphism in MFN2 is involved in the patho-genesis of AD [143, 144]. On the other hand, the forced overexpression of MFN2 in P301S human tau transgenic mice suppressed tau pathology induced neurodegenera-tion and cognitive decline [145]. It has also been reported that in tau knockout mice, the protein level of nuclear factor-erythroid-2-related factor 2 (Nrf2) was reduced, while the expression of MFN2 and PGC-1αwas significantly increased [146] . MFN2 is a guanosine triphosphatase (GTPase) on the outer membrane of mitochondria, which is involved in mitochondrial fusion. MFNs form dimers in a GTP dependent manner to facilitate the membrane tethering ability [147]. MFN1/2 are critical for glu-cose-stimulated insulin secretion (GSIS) through regulating the mtDNA expression via Tfam [148]. The trafficking of mitochondrial induced by 3,4-methylenedioxymeth-amphetamine (MDMA) is dependent on tau and MFN2/ Drp1 [149].
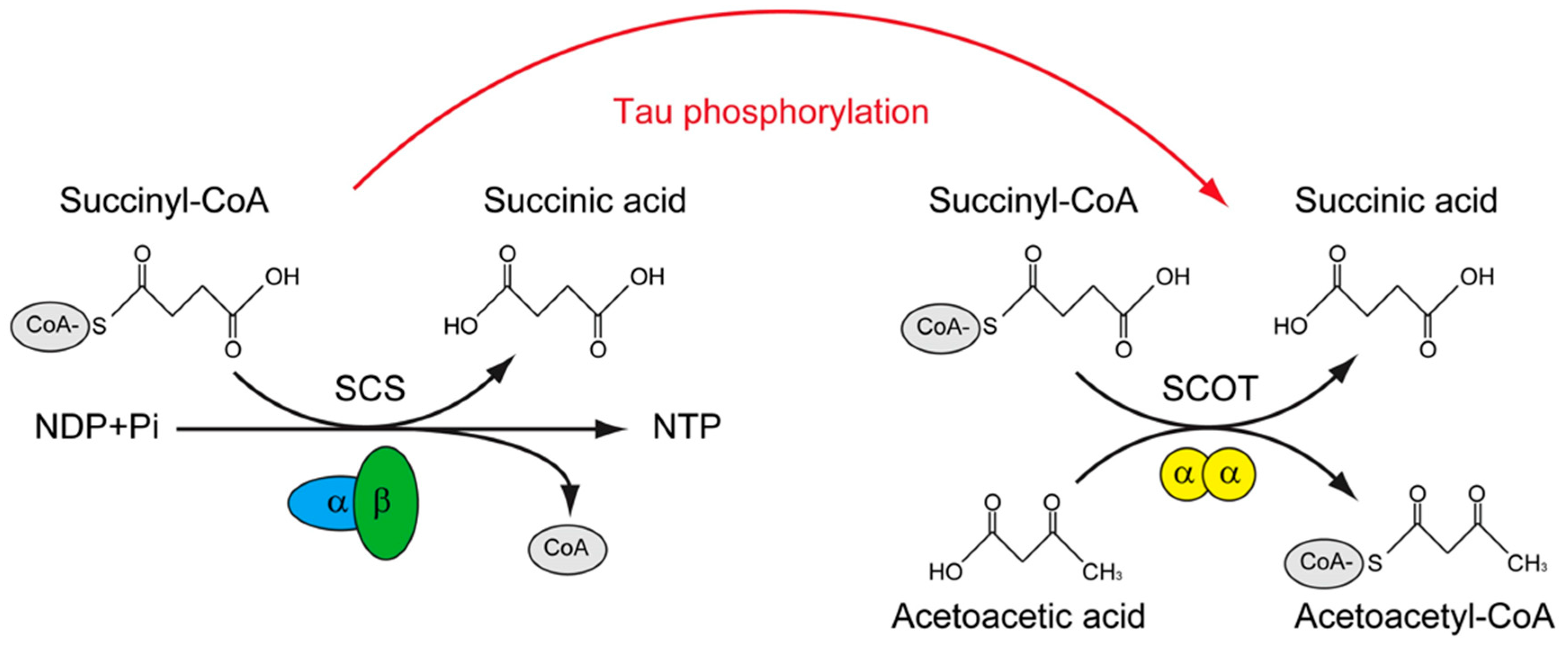
The tau interactome revealed that except for microtubule, tau could interact with presynaptic vesicle proteins and mitochondria proteins. More importantly, the FTD related mutations of tau impair the interaction of tau with mitochondria proteins, in-cluding SUCLG1, SUCLG2, SLC25A6, CYCS, et., al. [150]. In contrast, in the phosphor-ylated tau interactome that derived from NFT of AD, many of these mitochondrial proteins were not found, instead, novel phosphorylated tau interactors were presented, including OXCT1, COX5B, VDAC2, for example [151]. Among these tau interacting mitochondrial proteins, Oxct1 has been identified as a p-tau interacting protein [151] and a therapeutic target of AD [152], SUCLG2 has been recognized as a determinator of CSF Aβ1-42 levels and [153] and promising AD signature protein [154]. Interesting-ly, these two proteins are involved in a similar biological process, which is the transfer Co-A from Succinyl-CoA. The difference of them is that, SUCLG1/2 catalyzes the only substrate-level phosphorylation in the tricarboxylic acid cycle, and the transfer of CoA is accompanied with the production of GTP in mammal [155], however, OXCT1 cata-lyzes the reversible transfer of CoA from succinyl-CoA to acetoacetic acid without the production of GTP [156]. (Figure 2)
Succinyl-CoA synthetase (SCS) catalyzes the only substrate-level phosphorylation in the tricarboxylic acid cycle, the transfer of CoA is accompanied with the production of ATP/GTP. The SCS is a heterodimer, which is composed of SUCLG1, and either SU-CLG2A (specific for ATP production) or SUCLG2G (specific for GTP production). Suc-cinyl-CoA: 3-ketoacid-CoA transferase catalyzes (SCOT/OXCT1) is a mitochondrial homodimer, which catalyzes the reversible transfer of CoA from succinyl-CoA to acetoacetic acid without the production of ATP/GTP. The levels of ATP/GTP are criti-cal for mitochondrial dependent glucose stimulated insulin secretion. SCS was found to interact with tau, whereas SCOT/OXCT1 was found to interact with phosphorylated tau. Whether tau phosphorylation is involved in regulating the substrate level phos-phorylation in mitochondria is of interest to know.
GTP level determines cell fate through regulating Bcl2/Bax expression and activa-tion of caspase-3 [157] and p53 [158]. Importantly, the Bax has been found to positive-ly regulate mitochondrial fusion through MFN2 [159, 160]. In addition, Bak was in-volved in regulating mitochondrial morphology and pathology during apoptosis by interacting with MFNs [161]. Whether tau pathology will perturb the level of GTP is unknown. While, the experimental result indicated that phosphorylation of tau may perturb the substrate level phosphorylation and mitochondrial dependent GSIS. As also evidenced by the observation that tau knockout prevent neurotoxicity induced by Aβ peptide [135, 162, 163] and stress induced dendritic atrophy [164] and type 1 dia-betes induced cognitive impairment [71]. In addition, the tau ablation also improved mitochondrial function through increasing the levels of MFN2 and increased ATP production in the hippocampus [146]. Vanadium compound BEOV was found to sig-nificantly inhibit tau phosphorylation at Ser396 and Ser404 in primary neuron and brain of transgenic AD mice model, and improved the spatial learning activity of these mice [165]. However, whether vanadium had any influence on mitochondria functions is unknown. In the future study, the function of mitochondrial need to be recruit into this field.
5. Conclusions and Perspectives
Numerous studies demonstrated that tau is not only localized within mitochon-dria [125], but also exert pivotal functions in mitochondrial metabolisms. Apart from inducing mitochondrial abnormalities by hyperphosphorylated tau [140], the caspase 3 cleaved tau also impaired mitochondrial dynamics in AD [131] [166]. Meanwhile, the acetylation of tau also found in the brains of AD patients, which not only resulted in disability of mitochondria fission by decreasing mitofusion proteins, but also impaired mitochondrial biogenesis via reducing the level of PGC-1α [167]. Collectively, these studies indicated that tau is intimately corelated with mitochondrial dependent glu-cose metabolisms and insulin signaling in brain.
Many theories of AD etiology had been devised, such as, the amyloid cascade hy-pothesis [168], mitochondrial hypothesis [169], cholinergic hypothesis [170], neuroin-flammatory hypothesis [171], oxidative stress hypothesis [172], insulin resistance hy-pothesis [173], calcium hypothesis [174]. They are all supported by substantial clinical researches and experimental data. In the current paper, we try to put together the data that corelated with insulin signal, Aβ overproduction, and tau phosphorylation, to il-lustrate a chain of evidence future pharmacological study in this field.
As the evidence accumulated, we proposed that insulin signaling pathway play an important role in AD pathologies. Farther more, the impairment of substrate level of phosphorylation may be involved in the hyperphosphorylated and truncated tau in-duced mitochondrial damage. In earlier stage of AD, anti-diabetes agents such as va-nadium compounds was able to prevent or postpone the initiation of tau pathology though modulating insulin signaling pathway. However, further study is needed to investigate whether the vanadium compounds have any protective function on mito-chondria.
Author Contributions
J.Y. and N.L. wrote the manuscript. Z.H. and G.Y. draw the figures. Q.L. and N.L. revised the manuscript. All authors reviewed and concurred with the final manuscript.
Funding
National Natural Science Foundation of China (31700919; 21877081; 21771126); Shen-zhen-Hong Kong Institute of Brain Science-Shenzhen Fundamental Research Institutions (2021SHIBS0003).
Conflicts of Interest
The authors declare that they have no competing interests.
Abbreviations
| Aβ | amyloid-beta |
| AD | Alzheimer’s disease |
| ADAM10 | a disintegrin and metalloproteinase 10 |
| AGD | argyrophilic grain disease |
| AICD | APP intracellular domain |
| APOE | apolipoprotein E |
| APP | amyloid precursor protein |
| BEOV | bis(2-ethyl-3-hydroxy-4-pyronato) oxovanadium (IV) |
| BMOV | bis(maltolato) oxovanadium(IV) |
| CAA | cerebral amyloid angiopathy |
| CBD | corticobasal degeneration |
| CDK5 | cyclin-dependent kinase 5 |
| COX2 | cyclooxygenase-2 |
| CTE | chronic traumatic encephalopathy |
| CTF-83 | C-terminal fragment |
| CNS | central nervous system |
| Drp1 | dynamin-related GTPase |
| FAD | familial AD |
| FTDP-17 | frontotemporal dementia with Parkinsonism linked to chromosome 17 |
| GIP | glucose-dependent insulinotropic polypeptide |
| GLUT4 | glucose transporter 4 |
| GSIS | glucose-stimulated insulin secretion |
| GSK-3β | 3glycogen synthase kinase -3 |
| IDE | insulin degrading enzyme |
| IGF | insulin-like growth factor |
| iNOS | inducible nitric oxide synthase |
| IR | insulin receptor |
| IRS2 | insulin receptor substrate |
| LTP | long-term potentiation |
| MFN | mitofusion |
| NFTs | neurofibrillary tangles |
| Nrf2 | nuclear factor-erythroid-2-related factor 2 |
| PGC-1α | proliferator activated receptor gamma coactivator 1 α |
| PI3K | phosphatidylinositol-3-kinase |
| PiD | Pick’s disease |
| PP2A | protein phosphatase 2A |
| PPARγ | proliferator-activated receptor gamma |
| PS1/2 | presenilin-1/2 |
| PSP | progressive supranuclear palsy |
| PTEN | phosphatase and tension homologue on chromosome 10 |
| PTPase | protein tyrosine phosphatase |
| SCS | Succinyl-CoA synthetase |
| SCOT/OXCT1 | Succinyl-CoA: 3-ketoacid-CoA transferase catalyzes |
| STZ | streptozotocin |
References
- Harland, B.F.; Harden-Williams, B.A. Is vanadium of human nutritional importance yet? J Am Diet Assoc 1994, 94, 891–894. [Google Scholar] [CrossRef] [PubMed]
- Zaporowska, H.; Wasilewski, W. Hematological Effects of Vanadium on Living Organisms. Comparative Biochemistry and Physiology C-Pharmacology Toxicology & Endocrinology 1992, 102, 223–231. [Google Scholar]
- Beauge, L.A.; Glynn, I.M. A modifier of (Na+ + k+) atpase in commercial ATP. Nature 1977, 268, 355–356. [Google Scholar] [CrossRef] [PubMed]
- Beauge, L.A.; Glynn, I.M. Commercial ATP containing traces of vanadate alters the response of (Na+ + K+) ATPase to external potassium. Nature 1978, 272, 551–552. [Google Scholar] [CrossRef] [PubMed]
- Shechter, Y.; Karlish, S.J. Insulin-like stimulation of glucose oxidation in rat adipocytes by vanadyl (IV) ions. Nature 1980, 284, 556–558. [Google Scholar] [CrossRef]
- Huyer, G.; et al. Mechanism of inhibition of protein-tyrosine phosphatases by vanadate and pervanadate. J Biol Chem 1997, 272, 843–851. [Google Scholar] [CrossRef]
- Liu, J.C.; et al. Bis(acetylacetonato)-oxovanadium(IV), bis(maltolato)-oxovanadium(IV) and sodium metavanadate induce antilipolytic effects by regulating hormone-sensitive lipase and perilipin via activation of Akt. Metallomics 2013, 5, 813–820. [Google Scholar] [CrossRef]
- Wu, J.X.; Hong, Y.H.; Yang, X.G. Bis(acetylacetonato)-oxidovanadium(IV) and sodium metavanadate inhibit cell proliferation via ROS-induced sustained MAPK/ERK activation but with elevated AKT activity in human pancreatic cancer AsPC-1 cells. J Biol Inorg Chem 2016, 21, 919–929. [Google Scholar] [CrossRef]
- Montaigne, D.; Butruille, L.; Staels, B. PPAR control of metabolism and cardiovascular functions. Nature Reviews Cardiology 2021, 18, 809–823. [Google Scholar] [CrossRef]
- Zhao, P.; Yang, X.D. Vanadium compounds modulate PPAR gamma activity primarily by increasing PPAR gamma protein levels in mouse insulinoma NIT-1 cells. Metallomics 2013, 5, 836–843. [Google Scholar] [CrossRef]
- Wu, Y.L.; et al. Vanadyl acetylacetonate upregulates PPAR gamma and adiponectin expression in differentiated rat adipocytes. Journal of Biological Inorganic Chemistry 2013, 18, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Ladagu, A.D.; et al. Novel NMDA-receptor antagonists ameliorate vanadium neurotoxicity. Naunyn Schmiedebergs Arch Pharmacol 2020, 393, 1729–1738. [Google Scholar] [CrossRef] [PubMed]
- Colin-Barenque, L.; et al. Neuroprotective effect of carnosine in the olfactory bulb after vanadium inhalation in a mouse model. Int J Exp Pathol 2018, 99, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; et al. Vanadium in the detection, prevention and treatment of cancer: the in vivo evidence. Cancer Lett 2010, 294, 1–12. [Google Scholar] [CrossRef]
- Llobet, J.M.; Domingo, J.L. Acute toxicity of vanadium compounds in rats and mice. Toxicol Lett 1984, 23, 227–231. [Google Scholar] [CrossRef]
- Yang, X.G.; Wang, K. Chemical, biochemical, and biological behaviors of vanadate and its oligomers. Prog Mol Subcell Biol 2013, 54, 1–18. [Google Scholar] [PubMed]
- Scibior, A.; et al. Vanadium: Risks and possible benefits in the light of a comprehensive overview of its pharmacotoxicological mechanisms and multi-applications with a summary of further research trends. J Trace Elem Med Biol 2020, 61, 126508. [Google Scholar] [CrossRef] [PubMed]
- Scibior, A.; Kurus, J. Vanadium and Oxidative Stress Markers - In Vivo Model: A Review. Curr Med Chem 2019, 26, 5456–5500. [Google Scholar] [CrossRef]
- Zhao, Y.B.; et al. Vanadium compounds induced mitochondria permeability transition pore (PTP) opening related to oxidative stress. Journal of Inorganic Biochemistry 2010, 104, 371–378. [Google Scholar] [CrossRef]
- Thompson, K.H.; Orvig, C. Vanadium compounds in the treatment of diabetes. Metal Ions in Biological Systems 2004, 41, 221–252. [Google Scholar]
- Mcneill, J.H.; et al. Bis(Maltolato)Oxovanadium(Iv) Is a Potent Insulin Mimic. Journal of Medicinal Chemistry 1992, 35, 1489–1491. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; et al. Synthesis, characterization and anti-diabetic therapeutic potential of novel aminophenol-derivatized nitrilotriacetic acid vanadyl complexes. J Inorg Biochem 2015, 152, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Yang, J.; Yang, X. Synthesis and anti-diabetic activity of new N,N-dimethylphenylenediamine-derivatized nitrilotriacetic acid vanadyl complexes. J Inorg Biochem 2017, 177, 291–299. [Google Scholar] [CrossRef]
- Du, J.; et al. Vanadium coordination compounds loaded on graphene quantum dots (GQDs) exhibit improved pharmaceutical properties and enhanced anti-diabetic effects. Nanoscale 2020, 12, 9219–9230. [Google Scholar] [CrossRef] [PubMed]
- Setyawati, I.A.; et al. Kinetic analysis and comparison of uptake, distribution, and excretion of 48V-labeled compounds in rats. J Appl Physiol (1985) 1998, 84, 569–575. [Google Scholar] [CrossRef]
- Gerhardsson, L.; et al. Metal concentrations in plasma and cerebrospinal fluid in patients with Alzheimer's disease. Dement Geriatr Cogn Disord 2008, 25, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Szabo, S.T.; et al. Comparison of Metal Levels between Postmortem Brain and Ventricular Fluid in Alzheimer's Disease and Nondemented Elderly Controls. Toxicological Sciences 2016, 150, 292–300. [Google Scholar] [CrossRef]
- Busche, M.A.; Hyman, B.T. Synergy between amyloid-beta and tau in Alzheimer's disease. Nature Neuroscience 2020, 23, 1183–1193. [Google Scholar] [CrossRef]
- Muller, U.C.; Deller, T.; Korte, M. Not just amyloid: physiological functions of the amyloid precursor protein family. Nat Rev Neurosci 2017, 18, 281–298. [Google Scholar] [CrossRef]
- Kim, M.; et al. Potential late-onset Alzheimer's disease-associated mutations in the ADAM10 gene attenuate alpha-secretase activity. Human Molecular Genetics 2009, 18, 3987–3996. [Google Scholar] [CrossRef]
- Aguero, P.; et al. alpha-Secretase nonsense mutation (ADAM10 Tyr167*) in familial Alzheimer's disease. Alzheimers Res Ther 2020, 12, 139. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.J.; Chen, Y.R. The coexistence of an equal amount of Alzheimer's amyloid-beta 40 and 42 forms structurally stable and toxic oligomers through a distinct pathway. Febs Journal 2014, 281, 2674–2687. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; et al. Mechanisms for the Insertion of Toxic, Fibril-like beta-Amyloid Oligomers into the Membrane. Journal of Chemical Theory and Computation 2013, 9, 822–833. [Google Scholar] [CrossRef] [PubMed]
- Morkuniene, R.; et al. Small A beta(1-42) Oligomer-Induced Membrane Depolarization of Neuronal and Microglial Cells: Role of N-Methyl-D-Aspartate Receptors. Journal of Neuroscience Research 2015, 93, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Li, S.M.; et al. Soluble A beta Oligomers Inhibit Long-Term Potentiation through a Mechanism Involving Excessive Activation of Extrasynaptic NR2B-Containing NMDA Receptors. Journal of Neuroscience 2011, 31, 6627–6638. [Google Scholar] [CrossRef]
- Wang, X.L.; et al. Amyloid-beta overproduction causes abnormal mitochondrial dynamics via differential modulation of mitochondrial fission/fusion proteins. Proceedings of the National Academy of Sciences of the United States of America 2008, 105, 19318–19323. [Google Scholar] [CrossRef]
- Du, H.; et al. Early deficits in synaptic mitochondria in an Alzheimer's disease mouse model. Proceedings of the National Academy of Sciences of the United States of America 2010, 107, 18670–18675. [Google Scholar] [CrossRef]
- He, Y.; et al. Soluble oligomers and fibrillar species of amyloid beta-peptide differentially affect cognitive functions and hippocampal inflammatory response. Biochemical and Biophysical Research Communications 2012, 429, 125–130. [Google Scholar] [CrossRef]
- Zhang, Y.L.; et al. The relationship between amyloid-beta and brain capillary endothelial cells in Alzheimer's disease. Neural Regen Res 2022, 17, 2355–2363. [Google Scholar]
- Jansen, W.J.; et al. Prevalence of cerebral amyloid pathology in persons without dementia: a meta-analysis. JAMA 2015, 313, 1924–1938. [Google Scholar] [CrossRef]
- Pallo, S.P.; Johnson, G.V.W. Tau facilitates A beta-induced loss of mitochondrial membrane potential independent of cytosolic calcium fluxes in mouse cortical neurons. Neuroscience Letters 2015, 597, 32–37. [Google Scholar] [CrossRef]
- Roberson, E.D.; et al. Reducing endogenous tau ameliorates amyloid beta-induced deficits in an Alzheimer's disease mouse model. Science 2007, 316, 750–754. [Google Scholar] [CrossRef] [PubMed]
- Vossel, K.A.; et al. Tau reduction prevents Abeta-induced defects in axonal transport. Science 2010, 330, 198. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, M.; et al. Tau is essential to beta-amyloid-induced neurotoxicity. Proceedings of the National Academy of Sciences of the United States of America 2002, 99, 6364–6369. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, E.; et al. Tau deletion promotes brain insulin resistance. Journal of Experimental Medicine 2017, 214, 2257–2269. [Google Scholar] [CrossRef]
- Lei, P.; et al. Tau deficiency induces parkinsonism with dementia by impairing APP-mediated iron export. Nat Med 2012, 18, 291–295. [Google Scholar] [CrossRef]
- Abbondante, S.; et al. Genetic Ablation of Tau Mitigates Cognitive Impairment Induced by Type 1 Diabetes. American Journal of Pathology 2014, 184, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Trujillo-Estrada, L.; et al. Tau underlies synaptic and cognitive deficits for type 1, but not type 2 diabetes mouse models. Aging Cell 2019, 18. [Google Scholar] [CrossRef] [PubMed]
- Steen, E.; et al. Impaired insulin and insulin-like growth factor expression and signaling mechanisms in Alzheimer's disease - is this type 3 diabetes? Journal of Alzheimers Disease 2005, 7, 63–80. [Google Scholar] [CrossRef]
- Frolich, L.; et al. Brain insulin and insulin receptors in aging and sporadic Alzheimer's disease. J Neural Transm (Vienna) 1998, 105, 423–438. [Google Scholar] [CrossRef]
- de la Monte, S.M.; et al. Early-Stage Alzheimer's Disease Is Associated with Simultaneous Systemic and Central Nervous System Dysregulation of Insulin-Linked Metabolic Pathways. J Alzheimers Dis 2019, 68, 657–668. [Google Scholar] [CrossRef]
- Confettura, A.D.; et al. Neddylation-dependent protein degradation is a nexus between synaptic insulin resistance, neuroinflammation and Alzheimer's disease. Translational Neurodegeneration 2022, 11. [Google Scholar] [CrossRef]
- Ma, Q.L.; et al. Beta-amyloid oligomers induce phosphorylation of tau and inactivation of insulin receptor substrate via c-Jun N-terminal kinase signaling: suppression by omega-3 fatty acids and curcumin. J Neurosci 2009, 29, 9078–9089. [Google Scholar] [CrossRef] [PubMed]
- Farris, W.; et al. Insulin-degrading enzyme regulates the levels of insulin, amyloid beta-protein, and the beta-amyloid precursor protein intracellular domain in vivo. Proc Natl Acad Sci U S A 2003, 100, 4162–4167. [Google Scholar] [CrossRef]
- Cook, D.G.; et al. Reduced hippocampal insulin-degrading enzyme in late-onset Alzheimer's disease is associated with the apolipoprotein E-epsilon 4 allele. American Journal of Pathology 2003, 162, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Morelli, L.; et al. Insulin-degrading enzyme in brain microvessels: proteolysis of amyloid {beta} vasculotropic variants and reduced activity in cerebral amyloid angiopathy. J Biol Chem 2004, 279, 56004–56013. [Google Scholar] [CrossRef] [PubMed]
- Schechter, R.; Beju, D.; Miller, K.E. The effect of insulin deficiency on tau and neurofilament in the insulin knockout mouse. Biochemical and Biophysical Research Communications 2005, 334, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Clodfelder-Miller, B.J.; et al. Tau is hyperphosphorylated at multiple sites in mouse brain in vivo after streptozotocin-induced insulin deficiency. Diabetes 2006, 55, 3320–3325. [Google Scholar] [CrossRef]
- Schubert, M.; et al. Insulin receptor substrate-2 deficiency impairs brain growth and promotes tau phosphorylation. Journal of Neuroscience 2003, 23, 7084–7092. [Google Scholar] [CrossRef]
- Duarte, A.I.; Moreira, P.I.; Oliveira, C.R. Insulin in central nervous system: more than just a peripheral hormone. J Aging Res 2012, 2012, 384017. [Google Scholar] [CrossRef]
- Banks, W.A. The source of cerebral insulin. European Journal of Pharmacology 2004, 490, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A.; Owen, J.B.; Erickson, M.A. Insulin in the brain: There and back again. Pharmacology & Therapeutics 2012, 136, 82–93. [Google Scholar]
- Grillo, C.A.; et al. Insulin-stimulated translocation of GLUT4 to the plasma membrane in rat hippocampus is PI3-kinase dependent. Brain Research 2009, 1296, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Pearson-Leary, J.; Mcnay, E.C. Novel Roles for the Insulin-Regulated Glucose Transporter-4 in Hippocampally Dependent Memory. Journal of Neuroscience 2016, 36, 11851–11864. [Google Scholar] [CrossRef]
- Radhakrishnan, Y.; et al. Insulin-like growth factor-I stimulates Shc-dependent phosphatidylinositol 3-kinase activation via Grb2-associated p85 in vascular smooth muscle cells. Journal of Biological Chemistry 2008, 283, 16320–16331. [Google Scholar] [CrossRef]
- Rodriguez-Rodriguez, P.; et al. Tau hyperphosphorylation induces oligomeric insulin accumulation and insulin resistance in neurons. Brain 2017, 140, 3269–3285. [Google Scholar] [CrossRef]
- Wijesekara, N.; et al. Tau ablation in mice leads to pancreatic beta cell dysfunction and glucose intolerance. Faseb Journal 2018, 32, 3166–3173. [Google Scholar] [CrossRef]
- Zhao, N.; et al. Apolipoprotein E4 Impairs Neuronal Insulin Signaling by Trapping Insulin Receptor in the Endosomes. Neuron 2017, 96, 115–+. [Google Scholar] [CrossRef]
- Craft, S.; et al. Effects of Regular and Long-Acting Insulin on Cognition and Alzheimer's Disease Biomarkers: A Pilot Clinical Trial. J Alzheimers Dis 2017, 57, 1325–1334. [Google Scholar] [CrossRef]
- Born, J.; et al. Sniffing neuropeptides: a transnasal approach to the human brain. Nature Neuroscience 2002, 5, 514–516. [Google Scholar] [CrossRef]
- Adzovic, L.; et al. Insulin improves memory and reduces chronic neuroinflammation in the hippocampus of young but not aged brains. J Neuroinflammation 2015, 12, 63. [Google Scholar] [CrossRef] [PubMed]
- Reger, M.A.; et al. Effects of intranasal insulin on cognition in memory-impaired older adults: modulation by APOE genotype. Neurobiol Aging 2006, 27, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Reger, M.A.; et al. Intranasal insulin improves cognition and modulates beta-amyloid in early AD. Neurology 2008, 70, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Claxton, A.; et al. Long Acting Intranasal Insulin Detemir Improves Cognition for Adults with Mild Cognitive Impairment or Early-Stage Alzheimer's Disease Dementia. J Alzheimers Dis 2015, 45, 1269–1270. [Google Scholar] [CrossRef]
- Craft, S.; et al. Safety, Efficacy, and Feasibility of Intranasal Insulin for the Treatment of Mild Cognitive Impairment and Alzheimer Disease Dementia: A Randomized Clinical Trial. JAMA Neurol 2020, 77, 1099–1109. [Google Scholar] [CrossRef]
- Sato, T.; et al. Efficacy of PPAR-gamma agonist pioglitazone in mild Alzheimer disease. Neurobiol Aging 2011, 32, 1626–1633. [Google Scholar] [CrossRef]
- Watson, G.S.; et al. Preserved cognition in patients with early Alzheimer disease and amnestic mild cognitive impairment during treatment with rosiglitazone: a preliminary study. Am J Geriatr Psychiatry 2005, 13, 950–958. [Google Scholar] [CrossRef]
- Liu, J.; Wang, L.N.; Jia, J.P. Peroxisome Proliferator-Activated Receptor-Gamma Agonists for Alzheimer's Disease and Amnestic Mild Cognitive Impairment: A Systematic Review and Meta-Analysis. Drugs & Aging 2015, 32, 57–65. [Google Scholar]
- Risner, M.E.; et al. Efficacy of rosiglitazone in a genetically defined population with mild-to-moderate Alzheimer's disease. Pharmacogenomics Journal 2006, 6, 246–254. [Google Scholar] [CrossRef]
- Harrington, C.; et al. Rosiglitazone Does Not Improve Cognition or Global Function when Used as Adjunctive Therapy to AChE Inhibitors in Mild-to-Moderate Alzheimer's Disease: Two Phase 3 Studies. Current Alzheimer Research 2011, 8, 592–606. [Google Scholar] [CrossRef]
- Lu, X.Y.; et al. Metformin Ameliorates A beta Pathology by Insulin-Degrading Enzyme in a Transgenic Mouse Model of Alzheimer's Disease. Oxidative Medicine and Cellular Longevity 2020. [Google Scholar]
- Ou, Z.R.; et al. Metformin treatment prevents amyloid plaque deposition and memory impairment in APP/PS1 mice. Brain Behavior and Immunity 2018, 69, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Kickstein, E.; et al. Biguanide metformin acts on tau phosphorylation via mTOR/protein phosphatase 2A (PP2A) signaling. Proceedings of the National Academy of Sciences of the United States of America 2010, 107, 21830–21835. [Google Scholar] [CrossRef] [PubMed]
- Nabizadeh, F.; et al. Metformin use and brain atrophy in nondemented elderly individuals with diabetes. Exp Gerontol 2022, 166, 111890. [Google Scholar] [CrossRef]
- Koenig, A.M.; et al. Effects of the Insulin Sensitizer Metformin in Alzheimer Disease: Pilot Data From a Randomized Placebo-controlled Crossover Study. Alzheimer Dis Assoc Disord 2017, 31, 107–113. [Google Scholar] [CrossRef]
- Han, F.; et al. The vanadium (IV) compound rescues septo-hippocampal cholinergic neurons from neurodegeneration in olfactory bulbectomized mice. Neuroscience 2008, 151, 671–679. [Google Scholar] [CrossRef]
- Dong, Y.; et al. Anti-diabetic vanadyl complexes reduced Alzheimer's disease pathology independent of amyloid plaque deposition. Sci China Life Sci 2019, 62, 126–139. [Google Scholar] [CrossRef]
- He, Z.; et al. Bis(ethylmaltolato)oxidovanadium (IV) attenuates amyloid-beta-mediated neuroinflammation by inhibiting NF-kappaB signaling pathway via a PPARgamma-dependent mechanism. Metallomics 2021, 13. [Google Scholar]
- He, Z.J.; et al. Bis(ethylmaltolato)oxidovanadium (IV) mitigates neuronal apoptosis resulted from amyloid-beta induced endoplasmic reticulum stress through activating peroxisome proliferator-activated receptor gamma. Journal of Inorganic Biochemistry 2020, 208. [Google Scholar] [CrossRef]
- He, Z.J.; et al. The Protective Effect of Vanadium on Cognitive Impairment and the Neuropathology of Alzheimer's Disease in APPSwe/PS1dE9 Mice. Frontiers in Molecular Neuroscience 2020, 13. [Google Scholar] [CrossRef]
- He, Z.J.; et al. Bis(ethylmaltolato)oxidovanadium (IV) alleviates neuronal apoptosis through regulating peroxisome proliferator-activated receptor gamma in a triple transgenic animal model of Alzheimer's disease. Journal of Biological Inorganic Chemistry 2021, 26, 551–568. [Google Scholar] [CrossRef] [PubMed]
- He, Z.J.; et al. Bis(ethylmaltolato)oxidovanadium(iv) inhibited the pathogenesis of Alzheimer's disease in triple transgenic model mice (vol 71, pg 521, 2020). Metallomics 2020, 12, 631–631. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; et al. Is the hypoglycemic action of vanadium compounds related to the suppression of feeding? Biol Trace Elem Res 2014, 157, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Spires-Jones, T.L.; et al. Tau pathophysiology in neurodegeneration: a tangled issue. Trends Neurosci 2009, 32, 150–159. [Google Scholar] [CrossRef]
- Prokopovich, D.V.; et al. Impact of Phosphorylation and Pseudophosphorylation on the Early Stages of Aggregation of the Microtubule-Associated Protein Tau. Journal of Physical Chemistry B 2017, 121, 2095–2103. [Google Scholar] [CrossRef]
- Chang, E.; et al. Pseudophosphorylation of tau protein directly modulates its aggregation kinetics. Biochimica Et Biophysica Acta-Proteins and Proteomics 2011, 1814, 388–395.
- SantaCruz, K.; et al. Tau suppression in a neurodegenerative mouse model improves memory function. Science 2005, 309, 476–481. [Google Scholar] [CrossRef]
- Cieri, D.; et al. Tau localises within mitochondrial sub-compartments and its caspase cleavage affects ER-mitochondria interactions and cellular Ca2+ handling. Biochimica Et Biophysica Acta-Molecular Basis of Disease 2018, 1864, 3247–3256. [Google Scholar] [CrossRef]
- David, D.C.; et al. Proteomic and functional analyses reveal a mitochondrial dysfunction in P301L Tau transgenic mice. Journal of Biological Chemistry 2005, 280, 23802–23814. [Google Scholar] [CrossRef]
- Esteras, N.; et al. Mitochondrial hyperpolarization in iPSC-derived neurons from patients of FTDP-17 with 10+16 MAPT mutation leads to oxidative stress and neurodegeneration. Redox Biology 2017, 12, 410–422. [Google Scholar] [CrossRef]
- Manczak, M.; Reddy, P.H. Abnormal interaction between the mitochondrial fission protein Drp1 and hyperphosphorylated tau in Alzheimers disease neurons: implications for mitochondrial dysfunction and neuronal damage. Human Molecular Genetics 2012, 21, 2538–2547. [Google Scholar] [CrossRef]
- Gamblin, T.C.; et al. Caspase cleavage of tau: Linking amyloid and neurofibrillary tangles in Alzheimer's disease. Proceedings of the National Academy of Sciences of the United States of America 2003, 100, 10032–10037. [Google Scholar] [CrossRef]
- de Calignon, A.; et al. Caspase activation precedes and leads to tangles. Nature 2010, 464, 1201–U123. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla, R.A.; et al. Caspase-cleaved Tau Expression Induces Mitochondrial Dysfunction in Immortalized Cortical Neurons IMPLICATIONS FOR THE PATHOGENESIS OF ALZHEIMER DISEASE. Journal of Biological Chemistry 2009, 284, 18754–18766. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.J.; et al. Caspase-Cleaved Tau Impairs Mitochondrial Dynamics in Alzheimer's Disease. Molecular Neurobiology 2018, 55, 1004–1018. [Google Scholar] [CrossRef] [PubMed]
- Li, X.C.; et al. Human wild-type full-length tau accumulation disrupts mitochondrial dynamics and the functions via increasing mitofusins. Scientific Reports 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Kandimalla, R.; et al. Reduced dynamin-related protein 1 protects against phosphorylated Tau-induced mitochondrial dysfunction and synaptic damage in Alzheimer's disease. Human Molecular Genetics 2016, 25, 4881–4897. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; et al. The Protective Effect of Icariin on Mitochondrial Transport and Distribution in Primary Hippocampal Neurons from 3 x Tg-AD Mice. International Journal of Molecular Sciences 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; et al. Impaired Balance of Mitochondrial Fission and Fusion in Alzheimer's Disease. Journal of Neuroscience 2009, 29, 9090–9103. [Google Scholar] [CrossRef] [PubMed]
- Kandimalla, R.; et al. Hippocampal phosphorylated tau induced cognitive decline, dendritic spine loss and mitochondrial abnormalities in a mouse model of Alzheimer's disease. Human Molecular Genetics 2018, 27, 30–40. [Google Scholar] [CrossRef]
- Abtahi, S.L.; Masoudi, R.; Haddadi, M. The distinctive role of tau and amyloid beta in mitochondrial dysfunction through alteration in Mfn2 and Drp1 mRNA Levels: A comparative study in Drosophila melanogaster. Gene 2020, 754. [Google Scholar] [CrossRef]
- Yin, J.X.; et al. Effect of ApoE isoforms on mitochondria in Alzheimer disease. Neurology 2020, 94, E2404–E2411. [Google Scholar] [CrossRef]
- Song, J.Y.; et al. The association between MFN2 (mitofusin 2) gene polymorphism and late-onset Alzheimer's disease in Korean population. European Neuropsychopharmacology 2015, 25, S169–S169. [Google Scholar] [CrossRef]
- Kim, Y.J.; et al. Association between Mitofusin 2 Gene Polymorphisms and Late-Onset Alzheimer's Disease in the Korean Population. Psychiatry Investigation 2017, 14, 81–85. [Google Scholar] [CrossRef]
- Wang, L.W.; et al. Mitochondrial Fusion Suppresses Tau Pathology-Induced Neurodegeneration and Cognitive Decline. Journal of Alzheimers Disease 2021, 84, 1057–1069. [Google Scholar] [CrossRef]
- Jara, C.; et al. Genetic ablation of tau improves mitochondrial function and cognitive abilities in the hippocampus. Redox Biology 2018, 18, 279–294. [Google Scholar] [CrossRef]
- Li, Y.J.; et al. Structural insights of human mitofusin-2 into mitochondrial fusion and CMT2A onset. Nature Communications 2019, 10. [Google Scholar] [CrossRef]
- Sidarala, V.; et al. Mitofusin 1 and 2 regulation of mitochondrial DNA content is a critical determinant of glucose homeostasis. Nature Communications 2022, 13. [Google Scholar] [CrossRef]
- Barbosa, D.J.; et al. MDMA impairs mitochondrial neuronal trafficking in a Tau- and Mitofusin2/Drp1-dependent manner. Archives of Toxicology 2014, 88, 1561–1572. [Google Scholar] [CrossRef]
- Tracy, T.E.; et al. Tau interactome maps synaptic and mitochondrial processes associated with neurodegeneration. Cell 2022, 185, 712–+. [Google Scholar] [CrossRef]
- Drummond, E.; et al. Phosphorylated tau interactome in the human Alzheimer's disease brain. Brain 2020, 143, 2803–2817. [Google Scholar] [CrossRef]
- Zhang, F.; et al. Identification of Potential Therapeutic Targets of Alzheimer's Disease By Weighted Gene Co-Expression Network Analysis. Chin Med Sci J 2020, 35, 330–341. [Google Scholar]
- Ramirez, A.; et al. SUCLG2 identified as both a determinator of CSF A beta(1-42) levels and an attenuator of cognitive decline in Alzheimer's disease. Human Molecular Genetics 2014, 23, 6644–6658. [Google Scholar] [CrossRef]
- Wang, H.; et al. Integrated analysis of ultra-deep proteomes in cortex, cerebrospinal fluid and serum reveals a mitochondrial signature in Alzheimer's disease. Molecular Neurodegeneration 2020, 15. [Google Scholar] [CrossRef]
- Huang, J.; Fraser, M.E. The structure of succinyl-CoA synthetase bound to the succinyl-phosphate intermediate clarifies the catalytic mechanism of ATP-citrate lyase. Acta Crystallogr F Struct Biol Commun 2022, 78 Pt 10 Pt 10, 363–370. [Google Scholar] [CrossRef]
- Alghamdi, M.A.; et al. Clinical variability and outcome of succinyl-CoA:3-ketoacid CoA transferase deficiency caused by a single OXCT1 mutation: Report of 17 cases. JIMD Rep 2021, 62, 91–96. [Google Scholar] [CrossRef]
- Meshkini, A.; Yazdanparast, R.; Nouri, K. Intracellular GTP level determines cell's fate toward differentiation and apoptosis. Toxicol Appl Pharmacol 2011, 253, 188–196. [Google Scholar] [CrossRef]
- Dagher, P.C. Apoptosis in ischemic renal injury: Roles of GTP depletion and p53. Kidney International 2004, 66, 506–509. [Google Scholar] [CrossRef]
- Cleland, M.M.; et al. Bcl-2 family interaction with the mitochondrial morphogenesis machinery. Cell Death Differ 2011, 18, 235–247. [Google Scholar] [CrossRef]
- Hoppins, S.; et al. The soluble form of Bax regulates mitochondrial fusion via MFN2 homotypic complexes. Mol Cell 2011, 41, 150–160. [Google Scholar] [CrossRef]
- Brooks, C.; et al. Bak regulates mitochondrial morphology and pathology during apoptosis by interacting with mitofusins. Proc Natl Acad Sci U S A 2007, 104, 11649–11654. [Google Scholar] [CrossRef]
- Pallo, S.P.; Johnson, G.V. Tau facilitates Abeta-induced loss of mitochondrial membrane potential independent of cytosolic calcium fluxes in mouse cortical neurons. Neurosci Lett 2015, 597, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla, R.A.; et al. Phosphorylated tau potentiates Abeta-induced mitochondrial damage in mature neurons. Neurobiol Dis 2014, 71, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Lopes, S.; et al. Tau Deletion Prevents Stress-Induced Dendritic Atrophy in Prefrontal Cortex: Role of Synaptic Mitochondria. Cerebral Cortex 2017, 27, 2580–2591. [Google Scholar] [CrossRef]
- He, Z.; et al. An Adequate Supply of Bis(ethylmaltolato)oxidovanadium(IV) Remarkably Reversed the Pathological Hallmarks of Alzheimer's Disease in Triple-Transgenic Middle-Aged Mice. Biol Trace Elem Res 2022, 200, 3248–3264. [Google Scholar] [CrossRef]
- Fasulo, L.; Ugolini, G.; Cattaneo, A. Apoptotic effect of caspase-3 cleaved tau in hippocampal neurons and its potentiation by tau FTDP-mutation N279K. Journal of Alzheimers Disease 2005, 7, 3–13. [Google Scholar] [CrossRef]
- Ho, K.H.; et al. Glucose Regulates Microtubule Disassembly and the Dose of Insulin Secretion via Tau Phosphorylation. Diabetes 2020, 69, 1936–1947. [Google Scholar] [CrossRef]
- Benderradji, H.; et al. Impaired Glucose Homeostasis in a Tau Knock-In Mouse Model. Frontiers in Molecular Neuroscience 2022, 15. [Google Scholar]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer's disease at 25 years. EMBO Mol Med 2016, 8, 595–608. [Google Scholar] [CrossRef]
- Swerdlow, R.H.; Burns, J.M.; Khan, S.M. The Alzheimer's disease mitochondrial cascade hypothesis. J Alzheimers Dis 2010, 20 (Suppl 2), S265–279. [Google Scholar] [CrossRef] [PubMed]
- Bartus, R.T.; et al. The cholinergic hypothesis of geriatric memory dysfunction. Science 1982, 217, 408–414. [Google Scholar] [CrossRef]
- McGeer, P.L.; Rogers, J.; McGeer, E.G. , Neuroimmune mechanisms in Alzheimer disease pathogenesis. Alzheimer Dis Assoc Disord 1994, 8, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; et al. Alzheimer's disease: the two-hit hypothesis. Lancet Neurol 2004, 3, 219–226. [Google Scholar] [CrossRef]
- de la Monte, S.M. Insulin resistance and Alzheimer's disease. BMB Rep 2009, 42, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer's Association Calcium Hypothesis, W. Calcium Hypothesis of Alzheimer's disease and brain aging: A framework for integrating new evidence into a comprehensive theory of pathogenesis. Alzheimers Dement 2017, 13, 178–182.e17. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The deductive effluence of vanadium compounds on insulin signal in AD pathology.
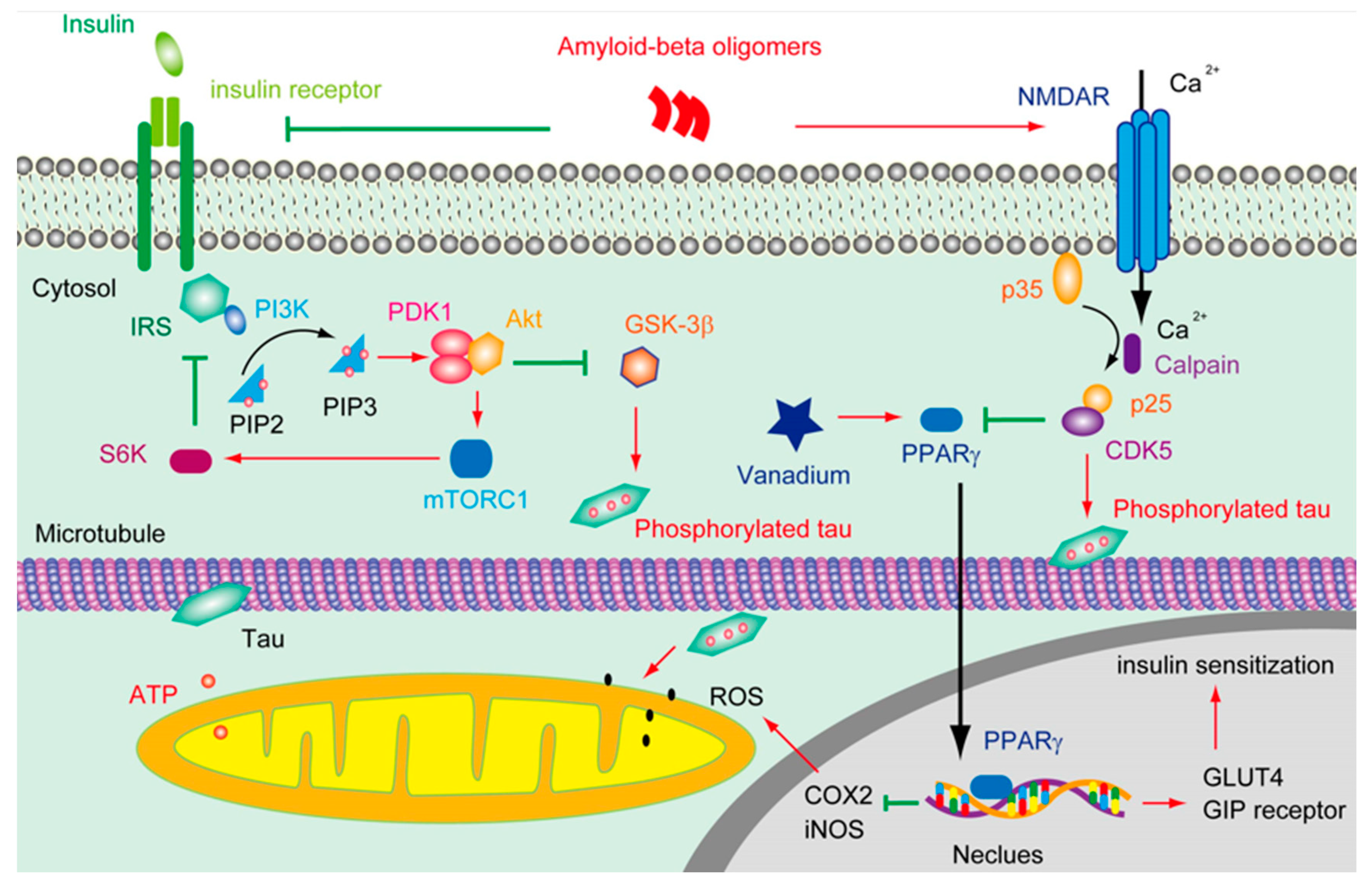
Figure 2.
The putative role of tau phosphorylation on substrate level phosphorylation of mito-chondrial.
Figure 2.
The putative role of tau phosphorylation on substrate level phosphorylation of mito-chondrial.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



