
Submitted:
21 July 2023
Posted:
21 July 2023
You are already at the latest version
Abstract
Pharmacogenetics and DNA methylation influence therapeutic outcomes and provide insights into potential therapeutic targets for brain-related disorders. To understand the effect of genetic polymorphisms on drug response and disease risk, we analyzed the relationship between global DNA methylation, drug-metabolizing enzymes, transport genes, and pathogenic gene phenotypes in serum samples from two groups of patients: Group A, which showed increased 5-methylcytosine (5mC) levels during clinical follow-up, and Group B, which exhibited no discernible change in 5mC levels. We identified specific SNPs in several metabolizing genes, including CYP1A2, CYP2C9, CYP4F2, GSTP1, and NAT2 that were associated with differential drug responses. Specific SNPs in CYP had a significant impact on enzyme activity, leading to changes in phenotypic distribution between the two patient groups. Group B, which contained a lower frequency of normal metabolizers and a higher frequency of ultra-rapid metabolizers compared to patients in Group A, did not show an improvement in 5mC levels during follow-up. Furthermore, there were significant differences in phenotype distribution between patient Groups A and B for several SNPs associated with transporter genes (ABCB1, ABCC2, SLC2A9, SLC39A8, and SLCO1B1) and pathogenic genes (APOE, NBEA, and PTGS2). These findings appear to suggest that the interplay between pharmacogenomics and DNA methylation has important implications for improving treatment outcomes in patients with brain-related disorders.
Keywords:
Epigenetics
; DNA methylation
; Pharmacogenetics
; Neurological Disorders
1. Introduction
Neurological disorders (NDs) are increasingly prevalent and represent the leading cause of disability-adjusted life years [1]. They account for over 6% of the global disease burden [2], are the second most common cause of death worldwide with nine million deaths annually, and are exacerbated by severe health disparities. For example, approximately 80% of the 50 million people with epilepsy reside in low- and middle-income countries [3]. NDs can be classified into six major categories that include age-related neurodegenerative disorders (e.g., Alzheimer’s disease, Huntington’s disease, and Parkinson’s disease), mental disorders (e.g., depression, psychosis), neurotoxic disorders (e.g., alcoholism), cerebrovascular disorders (e.g., stroke), neurodevelopmental disorders (e.g., autism), and other complex disorders (e.g., epilepsy). The pathogenesis of CNS disorders is complex and involves a combination of genetic and environmental factors.
There is currently a lack of pre-symptomatic biomarkers to facilitate the early detection of and intervention for NDs. Treating NDs often involves prolonged symptomatic management, carries significant costs, and has an increased risk of adverse drug reactions (ADRs). Moreover, developing effective treatments requires a comprehensive approach that considers each disorder’s unique characteristics as well as drug-drug interactions (DDIs) and the risk of ADRs in polypharmacy. To this, most patients with NDs require multifactorial treatment. However, this carries the risk of ADRs and DDIs due to comorbidity with other conditions such as hypertension, obesity, diabetes and cardiovascular disorders [4]. ADRs and DDIs are becoming a major health concern worldwide in subjects undergoing treatment [5,6] and rank among the top ten leading causes of death and illness in developed countries[6]. In the United States alone, approximately half a million ADRs are reported annually, with associated direct costs that exceed $150 billion per year [7]. Furthermore, ADRs increase hospital admissions, lengthen hospital stays, raise mortality rates and healthcare costs; they may also lead to drug withdrawal from the market [8]. A focus on precision medicine could improve patient outcomes and reduce the burden of NDs on individuals, families, and society.
Pharmacogenomics, the study of the influence of genetic variability on drug response and toxicity, is a vital component of precision medicine. By providing insight into how patients will respond to specific therapies, pharmacogenomics guides prescription and drug dose decisions. This helps reduce the risk, occurrence and severity of ADRs while optimizing drug efficacy [9]. Over 50% of drugs currently have a known pharmacogenomic profile that can be used to optimize efficacy and prevent adverse effects [6]. The pharmacogenomic machinery integrates drug and gene interactions through two pathways: the pharmacogenomic-pharmacokinetic pathway, which occurs during the absorption, distribution, metabolism and excretion (ADME) processes, and the pharmacogenomic-pharmacodynamic pathway, which occurs at the drug-target level [10]. Proper functioning of these pathways is essential for drug efficacy, and deficiencies or dysfunctions can cause ADRs or toxicity [11].
Pharmacogenetics accounts for approximately 80% of variability in drug safety and efficacy [9]. Rare variants make up 50% of the functional variability reported in more than 140 clinically relevant pharmagenes. Over 400 genes, including their encoded enzymes/proteins, influence drug efficacy and safety, and approximately 240 pharmagenes are associated with ADRs [12]. Pharmacogenetic outcome is influenced by various classes of genes, including pathogenic, mechanistic, metabolic, transporter and pleiotropic genes that comprise the pharmacogenetic machinery. These genes are regulated by epigenetic factors such as DNA methylation, chromatin/histone modifications, and miRNAs [4,13]. The enzymes and transporters that are integral to metabolic pathways exhibit a considerable degree of polymorphism. The presence of polymorphisms in the genes that encode these molecular components cause significant variability in interindividual drug responses [13,14]. Cytochrome P450 oxidases (CYPs) play a crucial role in regulating drug efficacy and toxicity. CYP1A2, CYP3A4/5, CYP2C9, CYP2C19 and CYP2D6 are among the most important CYPs involved in drug metabolism [15].
Genomics and epigenetics play a crucial role in the development and progression of neurodegenerative diseases. Epigenetics refers to hereditary changes in phenotype or gene expression that result from chromatin-based mechanisms, which do not involve alterations in the DNA sequence. Both biological and environmental factors modulate epigenetic modifications, which consequently impact gene expression and phenotype [16]. The accumulation of various epigenetic alterations over the lifespan may contribute to neurodegenerative disorders [17,18]. DNA methylation, the most extensively studied epigenetic mark, is a reversible mechanism catalyzed by DNA methyltransferases (DNMTs) that transfer a methyl group from the cofactor SAM (S-adenosyl-l- methionine) to the C5 position of cytosine found in CpG dinucleotides [19]; this results in the conversion of cytosines to 5-methylcytosines (5mC). DNA methylation changes the stability and accessibility of DNA, which regulates gene expression [18]. It is most commonly associated with gene silencing [20] that attracts other silencing elements such as methyl-CpG-binding proteins [21]. There are three families of DNMT proteins: DNMT1, DNMT2, and DNMT3, and all are expressed in neurons [22]. DNMT1 is responsible for maintaining methylation patterns after cell division, which allows for the inheritance of methylation marks [23]. DNMT3a and DNMT3b are responsible for de novo methylation [24]. Decreased global DNA methylation levels detected in blood samples of patients with neurodegenerative disorders could potentially function as a diagnostic biomarker [25,26,27,28]. Epigenetic-based therapies may potentially restore 5mC and other epigenetic modifications, providing a novel approach that could delay or reduce disease progression and improve the quality of life of patients.
Our recent study identified two categories of patients: Group A, in which 5mC levels were higher during the follow-up than during the initial visit, and Group B, in which patients had lower or similar 5mC levels during the follow-up than during the initial visit [29]. Given that pharmacogenetics can account for more than 80% variability in drug pharmacokinetics and pharmacodynamics, we conducted a retrospective study to investigate the influence of genetic factors on global DNA methylation levels in patients with NDs. More specifically, we investigated the influence of metabolic (CYP1A1, CYP1A2, CYP1B1, CYP2A6, CYP2B6, CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4, CYP3A5, CYP4F2, CES1, CHAT, COMT, GSTM1, GSTP1, GSTT1, NAT2, SOD2, TPMT, UGT1A1), transporter (ABCB1, ABCC2, ABCG2, SLC2A2, SLC2A9, SLC6A2, SLC6A3, SLC6A4, SLC39A8, SLCO1B1), and pathogenic (NBEA, PTGS2, APOE) gene variants on global DNA methylation levels, with respect to the therapeutic outcome during the initial and follow-up clinical assessments in both groups of patients.
Our aim in the current study was to provide new insights into the complex interplay between genetic and epigenetic determinants in modulating temporal DNA methylation patterns in patients with NDs to help us better understand disease susceptibility, treatment outcomes, and develop precision medicine strategies.
2. Materials and Methods
2.1. Subjects
In this retrospective study, a total of 98 patients ranging in age from 42 to 84 years old (mean age: 56 ± 1.61 years) were recruited from the CIBE Database at EuroEspes International Center of Neuroscience and Genomic Medicine (C000925, 21 October 2013, EuroEspes Biomedical Research Center). The study was conducted in accordance with the Helsinki Declaration, Spanish law (Organic Law on Biomedical Research, 14 July 2007), and with the approval of the Ethics/Research Committee of the EuroEspes Biomedical Research Center (Epibiomarkers EE0620). Blood samples were collected from patients during their first visit and then the follow-up visit at the EuroEspes Biomedical Research Center. Informed consent was obtained from all patients and/or legal caregivers. Patients were diagnosed according to globally accepted diagnostic criteria, following a comprehensive clinical and genetic examination. The clinical protocol for patients included a genomic analysis of single nucleotide polymorphisms (SNPs) linked to Parkinson’s disease (PD), Alzheimer’s disease (AD), or vascular risk, as well as psychological tests, brain mapping, and neuroimaging.
Patients were classified into two groups based on their 5mC levels. Group A consisted of patients with higher 5mC levels during the follow-up visit than during the initial visit, while Group B included those who had the same or lower 5mC levels during the follow-up visit than during the initial visit. This classification was done manually, and only differences between 5mC levels greater than 0.2% were considered for the separation of patients into Groups A and B.
2.2. Sample Collection and Analysis
Peripheral venous blood samples were collected from fasting individuals in a supine position. Blood was collected into EDTA-coated tubes and centrifuged at 3000 rpm for 10 min at 4 °C to isolate the buffy coat, which was then stored at -40 °C until DNA extraction. These tubes were allowed to clot for 30 min at room temperature before being centrifuged at 1500 rpm for 10 min at 4 °C. The resulting supernatant (serum) was removed and stored at -80 °C.
2.3. DNA Extraction
Peripheral blood lymphocyte DNA was isolated using the QIAcube robotic workstation together with the QIAamp DNA Mini Kit (Qiagen) following the manufacturer’s recommended protocol. The purity and concentration of the extracted DNA were determined using a microplate spectrophotometer (Epoch, BioTek Instruments, Vermont). Only DNA samples exhibiting 260/280 and 260/230 ratios greater than 1.8 were included in this study.
2.4. Quantification of Global DNA Methylation (5mC)
The quantification of global 5mC levels was performed colorimetrically with the MethylFlash Methylated DNA Quantification Kit (EpiGentek, New York, USA) with 50 ng of DNA per sample according to the manufacturer’s instructions. Absorbance was measured at 450 nm using a microplate reader. A standard curve was constructed using linear regression (Microsoft Excel) to determine the absolute quantity of methylated DNA. The amount and percent of 5mC were calculated using the following formulas:
5 mC (ng) = (Sample OD − Blank OD)/(Slope × 2)
5 mC (%) = 5 mC (ng)/sample DNA (ng) × 100
2.5. Genotyping
Our aim was to investigate the effect of polymorphisms in ND-related genes on their corresponding mRNA expression levels. We genotyped SNPs using qPCR with TaqMan assays and the Step one Plus Real Time PCR System (Life Technology) and TaqMan OpenArray DNA microchips for the QuantStudioTM 12K Flex Real-Time PCR System. The resulting genotyping data were analyzed using Genotyper software (Thermo Fisher Scientific).
3. Results
3.1. Genetic Variability in Drug Metabolizing Enzymes Influences 5mC Levels
We first investigated the influence of genetic variability in drug metabolizing enzymes, which are involved in regulating drug efficacy and toxicity [30,31], on the levels of 5-methylcytosine (5mC). Here, we analyzed the frequency of different phenotypes of cytochrome P450 (CYP) enzymes in two distinct groups (A and B) of patients based on their 5mC levels during their initial and follow-up visits. Phase I metabolism, which converts lipophilic drugs into more polar molecules by oxidation, reduction, or hydrolysis reactions, plays a key role in drug elimination [32], and over 2000 mutations have been described in CYP genes, some of which have a substantial impact on CYP activity [33]. More specifically, we investigated the impact of SNPs on the activity of CYP enzymes, including CYP1A1, CYP1A2, CYP1B1, CYP2A6, CYP2B6, CYP2C9, CYP2C19, CYP2D6, CYP3A4, CYP3A5, and CYP4F2.
Based on previous findings demonstrating the influence of genetic variability in drug metabolizing enzymes on 5mC levels, we further analyzed the frequency of different CYP phenotypes in two distinct patient groups: those whose 5mC levels increased during follow-up (Group A) and those whose levels decreased or remained similar to the initial visit (Group B) [29]. For CYP1A1 rs1378942, Group A showed a distribution of 20% abnormal metabolizers, 59% deficient metabolizers, and 21% normal metabolizers, while Group B had 28% abnormal, 59% deficient, and 21% normal metabolizer phenotypes (Figure 1A). For CYP1A2, the percentage of normal metabolizers decreased from 75% to 40% in Group B compared to Group A, and the percentage of ultra-rapid metabolizers increased from 25% to 60% (Figure 1B). The SNPs analyzed were rs2069514, rs35694136, and rs762551. In the case of CYP1B1 rs1056836 phenotypes, a rapid metabolizer was detected in 92% (Group A) and 96% (Group B), while the normal metabolizer was observed in 8% and 4% of cases, respectively (Figure 1C). For rs28399433 CYP2A6 phenotype analysis, a normal metabolizer incidence of 92% in Group A and 96% in Group B was observed (Figure 1D). The intermediate metabolizer phenotype showed an incidence of 8% in Group A and 4% in Group B (Figure 1D). No changes were detected in rs3745274 CYP2B6 phenotypes between Group A and B, with 68% and 65% normal metabolizers, 29% and 31% intermediate metabolizers, and 3% and 4% poor metabolizers, respectively (Figure 1E).
For CYP2C9, the percentage of normal metabolizers increased from 68% to 88% in Group B compared to Group A, and the percentage of intermediate metabolizers decreased from 29% to 12% (Figure 1F). The poor metabolizer phenotype was only detected in Group A (3%) (Figure 1F). The SNPs analyzed were rs1057910, rs1799853, rs28371685, rs28371686, rs7900194, and rs9332131. In the case of CYP2C19, the percentage of normal metabolizers increased from 57% to 64% in Group B compared to Group A, and the percentage of intermediate metabolizers increased from 14% to 24% (Figure 1G). The percentage of ultrarapid metabolizers decreased from 29% to 12% (Figure 1G).
We next examined the effects of specific SNPs on CYP2D6, CYP3A4, CYP3A5, and CYP4F2 phenotypes in patients in Groups A and B. Analysis of the rs12248560 and rs4244285 SNPs in CYP2D6 showed that the percentage of ultrarapid metabolizers decreased from 29% to 12% in Group B (Figure 1G). After analyzing the SNPs rs28371725, rs35742686, rs3892097, and rs5030655, there were no significant changes in the phenotypic distribution between Groups A and B in CYP2D6, with 48% and 54% of normal metabolizers, 37% and 32% of intermediate metabolizers, and 15% and 14% of ultrarapid metabolizers, respectively (Figure 1H). Similarly, there were no significant changes in the phenotypic distribution of the CYP3A4 SNPs rs2242480 and rs35599367 between Group A and B (66% and 76% normal metabolizers, 23% and 16% intermediate metabolizers, and 11% and 8% poor metabolizers, respectively) (Figure 1J). In contrast, the rs776746 SNP in CYP3A5 showed an increase in the percentage of normal metabolizers from 69.7% to 84% in Group B, while the percentage of rapid metabolizers decreased from 30.3% to 12% (Figure 1K). Furthermore, only Group B had intermediate metabolizers (4%) (Figure 1K). Analysis of the rs2108622 SNP in CYP4F2 showed an increase in the percentage of normal metabolizers from 51% to 84% in Group B, a decrease in intermediate metabolizers from 37% to 12%, and a decrease in poor metabolizers from 12% to 4% (Figure 1L).
After examining the effects of specific SNPs on CYP phenotypes in patient Groups A and B, we proceeded to analyze SNPs that are associated with phase II metabolism. This process involves conjugation reactions that increase drug solubility in water [32]. Our analysis of the rs71647871 SNP in the CES gene revealed that all cases in Group A were poor metabolizers. In contrast, 96% of cases in Group B were poor metabolizers and 4% were intermediate metabolizers (Figure 2A). For the rs2177369 SNP in the CHAT gene, there was a decrease in normal metabolizers from 35% to 24% and an increase in intermediate metabolizers from 27% to 48% in Group B compared to Group A. The percentage of poor metabolizers decreased from 38% to 28% (Figure 2B). When analyzing the rs4680 SNP in the COMT gene, there was a decrease in normal metabolizers from 29% to 20%, a decrease in poor metabolizers from 24% to 12%, and an increase in intermediate metabolizers from 47% to 68% (Figure 2C). For the Intel SNP in the GSTM1 gene, there was an increase in normal metabolizers from 63% to 72% in Group B, a decrease in poor metabolizers from 29% to 20%, and no significant change for intermediate metabolizers between Groups A and B (Figure 2D). Finally, our analysis of rs1138272 and rs1695 SNPs in GSTP1 showed an increase of normal metabolizer phenotype from 47% to 76% (Group B) while intermediate and poor metabolizer phenotypes decreased respectively from 38% to 16% and 15% to 8% (Figure 2E).
By studying multiple SNPs in different genes, we next aimed to identify additional genetic factors that influence drug metabolism and inter-individual differences in patient drug responses. Therefore, to better understand the genetic variations that contribute to drug metabolism in our patient populations, we analyzed SNPs in GSTT1, NAT2, SOD2, TPMT, and UGT1A1. The GSTT1 gene encodes a member of the glutathione S-transferase (GST) family of enzymes, which play an important role in the detoxification of xenobiotics, including drugs by conjugating them with glutathione [32]. By examining the insertion and deletion (indel) variant (SNP) of the GSTT1 gene, we assessed whether this polymorphism had an effect on the overall drug metabolism in our patient cohorts, and whether it contributed to the observed differences in metabolizer phenotypes between patients in Groups A and B. Our analysis of the Indel SNP of the GSTT1 gene revealed an increase in the proportion of normal metabolizers from 21% to 28%, while intermediate metabolizers showed a slight increase from 50% to 56%, and poor metabolizers decreased from 29% to 16% (Figure 2F). Additionally, we analyzed seven different NAT2 SNPs (rs1041983, rs1208m, rs1700020, rs1799930, rs1799931m, rs1801279, and rs1801280) and found that the incidence of slow acetylators decreased from 46% to 36% in Group B, whereas fast acetylators increased from 3% to 24% (Figure 2G). The incidence of intermediate acetylators decreased from 51% to 40%.
Our analysis of the rs4880 SNP in the SOD2 gene sought to investigate its potential impact on drug metabolism in our patient cohorts. This SNP has been associated with various diseases and oxidative stress and may also influence enzyme activity [34]. Therefore, we examined its distribution in Groups A and B to further understand its relationship with drug metabolism. For the rs4880 SOD2 SNP, we observed a similar distribution of patients with normal (32% and 28% in Groups A and B, respectively) as comparing with patients showing an abnormal activity (67% and 72%) (Figure 2H).
The TPMT gene encodes the thiopurine S-methyltransferase enzyme, which is responsible for the metabolism of thiopurine drugs widely used for the treatment of leukemia and autoimmune disorders [35]. Genetic polymorphisms within TPMT significantly impact the activity of the TPMT enzyme, causing interindividual variability in response to thiopurine treatment [36]. In the current study, to determine the distribution of TPMT SNPs in Groups A and B, which have differential responses to drug treatment, we analyzed four specific SNPs, rs1142345, rs1800460, rs1800462, and rs1800584. Our aim was to investigate whether genetic differences in TPMT contribute to the observed differences in drug response between the two groups. Analysis of the TPMT SNPs rs1142345, rs1800460, rs1800462 and rs1800584 showed that most patients exhibited a normal activity phenotype, with only 4% being identified as patients with abnormal activity in Group B (Figure 2I).
The UGT1A1 gene is implicated in the development of several cancer subtypes, including colon, breast, and prostate cancer. The UGT1A1 enzyme is involved in the metabolism of various drugs, including anticancer agents [37]. The UGT1A1 gene is significantly polymorphic, and several SNPs impact UGT1A1 enzyme activity. Certain UGT1A1 variants have been associated with a heightened risk of toxicity in response to particular drugs, including the chemotherapy drug irinotecan [38,39]. In the current study, we analyzed the distribution of UGT1A1 SNPs in patient groups A and B to evaluate their potential role in drug metabolism and toxicity in these population cohorts. Analysis of the distribution of UGT1A1 SNPs rs35350960, rs14124874, rs4148323, and rs887829 revealed a slight increase in the proportion of normal metabolizers from 63% (Group A) to 72% (Group B) (Figure 2J). The incidence of poor metabolizers decreased from 29% (Group A) to 20% (Group B), while the incidence of intermediate metabolizers remained unchanged in both groups (8%).
3.2. Modification of DNA Methylation Levels by Transport Gene Phenotypes
Our next objective was to identify genetic factors that could influence patient response to treatment by analyzing transport gene phenotypes. To achieve this, we determined the frequencies of normal, deficient, and abnormal response phenotypes for each gene in patient groups A and B. By examining the genetic profiles of these patients, we aimed to elucidate the mechanisms underlying treatment outcomes and provide insights into precision medicine. Specifically, we investigated whether specific SNPs could modify DNA methylation levels and alter phenotypic responses to treatment. We analyzed phenotypic changes in response to treatment for several SNPs in the ABCB1, ABCC2, ABCG2, SLC2A2, SLC2A9, SLC6A2, and SLC6A3 genes.
Analysis of the ABCB1 SNPs rs1128503, rs1032582.01, rs2032582.02 and rs1045642 revealed a significant decrease in abnormal response phenotype from 60% to 32% in Group B (Figure 3A). Normal and deficient response phenotypes increased from 23% to 40%, and from 17% to 28%, respectively (Figure 3A). ABCC2 SNPs rs717620, rs2273697, rs17222723 and rs3740066 showed an increase in normal response phenotype from 26% to 36% in Group B, whereas deficient response phenotype increased from 53% to 60 % (Figure 3B). A significant decrease from 21% to 4% in abnormal response phenotype was observed in Group B (Figure 3B).
ABCG2 SNP rs2231142 showed a similar frequency of the normal response phenotype in Groups A (85%) and B (84%) (Figure 3C). The principal difference between the two groups was the presence of an abnormal response phenotype (3%) in Group A (Figure 3C). Analysis of the rs5400 SNP in the SLC2A2 gene showed only minor differences in phenotype distribution between the two groups (Figure 3D). In contrast, normal response phenotypes were found in 9% of Group A patients and in 12% of Group B patients, while deficient response phenotypes were observed in 26% and 36% of Group A and B patients, respectively (Figure 3D). In groups A and B, abnormal responses were observed in 65% and 52% of patients respectively. The normal response phenotypes were observed in 9% of Group A patients and in 12% of Group B patients. Deficient response phenotypes were observed in 26% of Group A patients and 36% of Group B patients (Figure 3D).
Analysis of the SLC2A9 SNP rs16890979 revealed a significant reduction in the deficit response phenotype in Group B, decreasing from 41% in Group A to 28% in Group B (Figure 3E). The normal response phenotype increased from 56% (Group A) to 64% (Group B), while the altered response increased slightly from 3% in Group A to 8% in Group B (Figure 3E). Analysis of the rs5569 SNP in the SLC6A2 gene indicated minor differences in the frequency of the various phenotypes (Figure 3F). Specifically, the normal response phenotype increased from 9% in Group A to 12% in Group B, while the deficient response phenotype increased from 26% in Group A to 36% in Group B. The abnormal response decreased from 65% (Group A) to 52% (Group B) (Figure 3F). Analysis of the SLC6A3 SNP rs460000 revealed no change in phenotype distribution between Groups A and B (Figure 3G). The frequency of normal response phenotypes was 66% and 64% for Groups A and B respectively. For the deficient response phenotype, it was 31% and 32%, and for the abnormal response phenotype, it was 3% and 4%, for Groups A and B respectively (Figure 3G). The SLC6A4 SNP rs2020936 showed an increase in normal response phenotypes from 50% in Group A to 64% in Group B (Figure 3H). Both abnormal and deficient phenotypes decreased from 18% to 12% and from 32% to 24%, in Groups A and B, respectively (Figure 3H). The SLC39A8 SNP rs13107325 showed a marked reduction in the frequency of normal response phenotypes from 85% in Group A to 59% in Group B (Figure 3I). Abnormal response phenotypes were observed in 15% and 27% of patients in Groups A and B, respectively, whereas the deficient response phenotype was only observed in 14% of Group B subjects (Figure 3I). Analysis of the SLCO1B1 SNPs rs2306283, rs4149015 and rs1419056 showed a strong increase in the frequency of the normal response phenotype from 49% in Group A to 92% in Group B, with only minor changes in the deficient response phenotype observed in 11% and 8% of patients in Groups A and B, respectively (Figure 3J). Interestingly, an abnormal response phenotype was only present in Group A, with a frequency of 40% (Figure 3J).
3.3. Pathological Gene Phenotypes Alter DNA Methylation Levels
Altered DNA methylation levels caused by pathological gene phenotypes have been implicated in numerous diseases, including AD, for which apolipoprotein E4 (APOE4) is a major risk factor [40]. The E2 allele of the APOE gene is protective against AD [41]. To gain insight into the role of genetic factors in modifying DNA methylation levels and their potential impact on disease risk and drug response, we next analyzed the frequency of different genotypes in Groups A and B and found significant differences in the distribution of APOE, neurobeachin (NBEA), and prostaglandin-endoperoxide synthase 2 (PTGS2) genotypes.
The frequency of the APOE 3.3 genotype decreased from 73% in Group A to 56% in Group B (Figure 4). Furthermore, the frequency of the pathological APOE 4.4 genotype increased from 2.7% in Group A to 8% in Group B. The APOE 2.4 genotype was only present in Group B at a frequency of 4%.
Neurobeachin (NBEA) is a kinase-anchoring protein that contributes to synapse maturation and development of the nervous system [42]. The NBEA SNP rs1779800 is associated with differential therapeutic responses to acetylcholinesterase inhibitors (AChEIs) [43]. In the current study, there was a substantial decrease in the normal response phenotype from 64% (Group A) to 35% (Group B), and a corresponding increase in the deficient response phenotype from 27% (Group A) to 55% (Group B) (Figure 5). However, the abnormal response phenotype showed similar values in both groups A and B (9% and 10%, respectively).
The emergence of behavioral disorders in AD may be associated with certain genetic polymorphisms, metabolic dysfunctions, and cerebrovascular risk factors. The symptomatology of behavioral disorders in different forms of dementia is not uniform, but depression and apathy are common manifestations that often necessitate drug intervention. The reported frequency of depression in AD varies widely, ranging from 5% to over 40% [44]. Nonetheless, after apathy, depression is identified consistently as the second most prevalent psychiatric symptom in AD, and its incidence increases in advanced stages of the disease [45]. The PTGS2 SNP rs5275 has been implicated in the development of major depressive disorder (MDD) [46] [47]. Our analysis of this SNP revealed an increase in the normal response phenotype from 46% in Group A to 67% in Group B, accompanied by a substantial decrease in the frequency of the abnormal response phenotype from 15% (Group A) to 1% (Group B) (Figure 6). These data provide insight into the genetic factors that influence patient response to treatment and have important implications for the development of precision medicine strategies for NDs.
4. Discussion
DNA methylation has emerged as a promising biomarker for brain-related disorders as it provides insights into gene expression patterns and potential therapeutic targets [25,26]. In a recent study, we examined the correlation between hypovitaminosis, psychometric parameters, DNA methylation, and NDs, and found two distinct patient categories based on changes in 5mC levels during clinical follow-up: Group A, which showed increased 5mC levels during the follow-up, and Group B, which exhibited no discernible change in 5mC levels [29]. In the current study, we investigated, in those two groups of patients, how different pharmacogenetic patient phenotypes, encoded by metabolizing, transporter- and pathogenic genes, affect DNA methylation during clinical follow-up. Our major finding was that variations in genotype-phenotype interactions cause different responses to treatments and are associated with differential changes in 5mC levels over time.
Our first analysis examined the impact of genetic variability in drug metabolizing enzymes on 5mC levels. We focused on different phenotypes of CYP enzymes in the two groups of patients and found that specific SNPs had a significant impact on CYP enzyme activity, leading to changes in phenotypic distribution between the two groups of patients. In particular, there were no significant changes in the phenotypic distribution for some SNPs, while others showed a shift in the percentage of different metabolizing phenotypes.
To identify additional genetic factors that influence drug metabolism and inter-individual differences in patient drug responses, we next analyzed the effects of specific SNPs on CYP and phase II metabolism phenotypes in patient Groups A and B. We found that genetic polymorphisms in CES, CHAT, COMT, GSTM1, GSTP1, GSTT1, NAT2, SOD2, TPMT, and UGT1A1 contributed to the observed differences in drug response between the two groups. Furthermore, there was an increase in normal metabolizers in some genes, whereas others showed a decrease in poor metabolizers and an increase in intermediate metabolizers.
By analyzing transport gene phenotypes, we then assessed whether specific SNPs could modify DNA methylation levels and alter phenotypic responses to treatment. Specifically, we examined the frequencies of normal, deficient, and abnormal response phenotypes for each gene in patient groups A and B. There were significant differences in phenotype distribution between the two patient groups for several SNPs associated with ABCB1, ABCC2, ABCG2, SLC2A2, SLC2A9, SLC6A2, SLC6A3, SLC6A4, SLC39A8, and SLCO1B1. SLCO1B1 SNPs had a strong effect on patient response to treatment, with a marked increase in the frequency of the normal response phenotype in Group B compared to Group A.
Finally, to understand the role of pathogenic gene phenotypes in modifying DNA methylation levels and their impact on disease risk and drug response, we then analyzed the distribution of different genotypes in the two groups of patients and found substantial differences in the frequencies of APOE, NBEA, and PTGS2 genotypes.
CYP enzymes constitute a diverse group of drug-metabolizing enzymes that regulate the metabolism of the majority of xenobiotic substances [48]. CYPs are responsible for approximately 80% of oxidative metabolism and 50% of drug elimination for currently used drugs [6]. The normal metabolizer (NM), intermediate metabolizer (IM), poor metabolizer (PM), and ultra-rapid metabolizer (UM) phenotypes, linked to different gene variants, determine drug efficacy and toxicity [6,49,50]. CYP1A2, for example, metabolizes various drugs, including phenacetin, caffeine, clozapine, tacrine, propranolol, and mexiletine [51]. In particular, a strong correlation exists between coffee consumption and PD among slow metabolizers of caffeine homozygous for CYP1A2 polymorphisms [52]. In patients with schizophrenia, clozapine dosage is influenced by CYP1A2 activity, and tacrine metabolism is primarily dependent on the activity of CYP1A2 and CYP3A4 enzymes [53,54]. Tacrine, a reversible cholinesterase inhibitor [55], is a major substrate of CYP1A2 and CYP3A4 [56]. Propanolol is commonly prescribed for migraine prophylaxis and anxiety treatment [57,58]. Our findings suggest that Group B, in which patients did not show an improvement in 5mC levels during follow-up, contained a lower frequency of subjects that were normal metabolizers (40%, compared to 75% in Group A) and a higher frequency of patients that were ultra-rapid metabolizers. CYP1A2 is important for the dosing of several antipsychotics. Ultra-rapid metabolizers are resistant to clozapine treatment, and improved outcomes are achieved by co-administration of the CYP1A2 inhibitor fluvoxamine and by increasing clozapine dosage [59]. The higher frequency of ultra-rapid metabolizers in Group B patients may therefore explain the poor response to treatment and lack of improvement in 5mC levels during the clinical follow-up.
CYP2E1 is involved in the metabolism of fatty acids, which are abundant in the brain [60], and in the biotransformation of exogenous compounds [61]. These compounds include ethanol, nicotine, acetaminophen, acetone, aspartame, chloroform, chlorzoxazone, tetrachloride, and antiepileptic drugs such as phenobarbital. In transgenic (APP/PS1) AD mice, chlorzoxazone is neuroprotective by attenuating neuroinflammation and neurodegeneration [62]. Our results indicate that in Group B, the frequency of the normal metabolizer phenotype was lower (76% vs. 89%) and the frequency of the intermediate metabolizer phenotype was higher (24% vs. 11%) compared to Group A. Intermediate metabolizers display reduced enzymatic activity and increased side effects because of incomplete drug metabolism compared to normal metabolizers, indicating that a lower dose may be required [63]. The increase in the frequency of intermediate metabolizers in Group B subjects may contribute to the observed increase in toxicity issues and lack of improvement in 5mC levels during the patient follow-up period.
CYP4F2 is involved in the metabolism of fatty acids and vitamin E [64]. Vitamin E is associated with decreased DNMT expression [65]. Our current findings showed that Group B had a higher frequency of the normal metabolizer phenotype (82%) compared to Group A (52%). Moreover, the frequencies of intermediate and poor metabolizers were lower in Group B. This suggests that patients in Group B may be able to metabolize vitamin E more effectively than patents in Group A, which could potentially decrease DNA methylation.
Variants in the CHAT gene may influence the response to AChEIs [66]. In our study, the frequency of the intermediate metabolizer phenotype increased from 27% to 48% in Group B. Intermediate metabolizers have reduced enzymatic activity and increased side effects, which could explain the lack of response to treatment and the absence of improvement in 5mC levels in Group B patients. Similarly, COMT analysis showed an increase in the frequency of the intermediate metabolizer phenotype from 47% (Group A) to 68% in Group B. COMT enzyme activity is linked to various psychiatric and neurological disorders [67]. COMT is involved in Phase II metabolism and transfers a methyl group from S-adenosylmethionine (SAM) to a hydroxyl group on the catechol ring of endogenous and xenobiotic catechol substrates. During COMT-catalyzed methylation, SAM is converted to a competitive inhibitor, S-adenosylhomocysteine (SAH), resulting in a negative feedback loop [67]. Endogenous substrates of COMT include dopamine, norepinephrine, and epinephrine [68] and there is a significant association between the COMT rs4680 SNP and the response to antidepressant treatment [67].
Although previous studies found no significant correlations between NAT2 genotypes and AD or PD [69,70], our present study found a higher frequency of rapid acetylators in Group B (24%) than in Group A (35%) patients. This increased frequency of rapid acetylators could contribute to the stable levels of 5mC. AtreMorine, a novel compound obtained from the Vicia faba L. plant using a non-denaturing biotechnological process, increases DNA methylation in patients with PD by providing L-DOPA, a dopamine precursor commonly used to treat PD [71,72]. Pharmacogenomic analysis of NAT2 phenotypes in response to AtreMorine show that the increase in DNA methylation is not statistically significant in patients with the fast acetylator phenotype [72].
Polymorphisms in drug transporter genes affect drug pharmacokinetics and ultimately drug concentration in plasma and target tissues [73]. Among these transporters, ABCB1 plays a critical role in the brain [74]. In PD, the frequency of ABCB1 phenotypes is 24.76% for low responders (LR), 32.67% for intermediate responders (IR), and 42.57% for high responders (HR), with no significant differences in response to AtreMorine among the phenotypes [75]. ABCB1 also transports 42% of antiepileptic drugs and 16% of benzodiazepines [43]. ABCB1 is a major transporter (55%) of antidepressants [76] and benzodiazepines (16%); antidepressants are substrates (25%) and inducers (3%) of ABCB1 [76]. Our findings showed a substantial decrease in the high responder phenotype in Group B, from 60% to 32%. Furthermore, SLC39A8 is involved in the transport of a variety of drugs used to treat anxiety, panic attacks, sleep disorders, agitation, and behavioral anomalies [43]. In our study, the normal response SLCOB1 phenotype increased from 49% (Group A) to 92% in Group B patients, while only an intermediate response phenotype was observed in Group A.
ApoE is the major carrier of lipids and cholesterol in the CNS [77]. The expression of the APOE4 genotype is a significant risk factor for developing late-onset AD [77]. The severity of synucleinopathies is associated with the APOE4 variant, independent of concomitant AD severity [78,79]. While APOE4 is not a risk factor for PD, it increases the risk of developing dementia and cognitive decline [77]. The APOE genotype also affects the age of onset and severity of stroke, and individuals with the APOE4 allele exhibit delayed recovery of verbal memory function [80] and an increased risk of developing stroke-associated dementia [81]. We and others have shown that APOE expression decreases in venous blood and plasma samples in AD patients, suggesting its potential as a diagnostic biomarker for the disease [82,83]. APOE mRNA expression is also lower in E4 carriers than in individuals with APOE 2.3 and -3.3 genotypes [29]. The incidence of the APOE4 allele was higher in Group B (20% 3.4, 8% 4.4, and 4% 2.4) than in Group A (16% 3.4 and 3% 4.4), which may explain the lack of improvement in 5mC levels during the follow-up.
In patients with AD, the NBEA SNP rs17798800 is a potential predictor of response to treatment with AChEIs [84]. In patients with AD and depression, the observed phenotype frequency of this SNP for AChEI responders (AR), AChEI intermediate responders (DR), and AChEI non-responders (NR) phenotypes are 8.72%, 22.56%, and 68.72%, respectively [85], which is similar to the phenotype distribution in Group A of our patient cohort. However, in the current study, we observed a different frequency distribution of the NBEA phenotype in Group B patients, with a significant decrease in the normal response frequency (10% AR, 55% DR, and 35% NR). This difference in frequency distribution suggests that the NBEA phenotype may play a role in the changes in 5mC levels observed during the follow-up period.
The present study proposes that analyzing genotypes related to metabolism, transporters, and pathological pharmacogenetics can be used to analyze the evolution of global DNA methylation levels (Table 1). Other studies have highlighted the correlation between neurodegenerative diseases and decreased levels of sirtuin activity, neurodegenerative gene expression, and global DNA methylation [25,26,82]. Furthermore, DNA methylation has shown potential as a biomarker for monitoring disease outcome in patients with neurological disorders [29].
5. Conclusions
Genetic polymorphisms significantly impact drug response and disease risk, as demonstrated by our analysis of the relationships between global DNA methylation and drug-metabolizing enzymes, transport genes, and pathogenic gene phenotypes. We identified specific SNPs in CYP, CES, CHAT, COMT, GSTM1, GSTP1, GSTT1, NAT2, SOD2, TPMT, UGT1A1, ABCB1, ABCC2, ABCG2, SLC2A2, SLC2A9, SLC6A2, SLC6A3, SLC6A4, SLC39A8, and SLCO1B1 genes that were associated with differential drug responses. There was also a higher frequency of ultra-rapid metabolizers in patient Group B, which may explain their poor response to treatment and lack of improvement in 5mC levels. Our study highlights the potential of DNA methylation as a biomarker for brain-related disorders and emphasizes the significance of incorporating pharmacogenomics into personalized treatment plans for brain disorders, which may improve patient treatment outcomes.
Author Contributions
Conceptualization, O.M.I..; methodology, O.M.I, V.N, I.C. Y J.C.C..; validation, O.M..; formal analysis, O.M.I..; resources, N.C..; writing—original draft preparation, O.M.I and V.N..; writing—review and editing, O.M.I., V.N., J.C.C., N.C and R.C..; supervision, O.M.I., R.C. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
The study was conducted in accordance with the Helsinki Declaration, Spanish law (Organic Law on Biomedical Research, 14 July 2007), and following the approval of the Ethics/Research Committee of the EuroEspes Biomedical Research Center (Epibiomarkers EE0620).
Informed Consent Statement
Written informed consent has been obtained from the patient(s) to publish this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- A., R.; M.M., R. Potential therapeutics against neurological disorders: Natural products-based drugs. Front Pharmacol 2022, 13, 950457. [CrossRef] [PubMed]
- A., F. Global, Regional, and Netional burden of 12 Mental Disorders in 204 countries and territories, 1990-2019: A systematic analysis for the Global Burden of Disease Study. Lancet Psychiatry 2022, 9, 137–150. [CrossRef] [PubMed]
- Singh, G.; Sander, J.W. The global burden of epilepsy report: Implications for low- and middle-income countries. Epilepsy & behavior 2020, 105, 106949. [Google Scholar] [CrossRef]
- R., C.; V., N.; O., M.-I.; L., C.; N., C.; R., P.; J.C., C. Pharmacogenomics of Alzheimer´s Disease: Novel strategies for drug utilization and development. Pharmacogenomics in Drug Discov and Development 2022, 275–387. [CrossRef]
- O., O.; M., P.; A.K., D. Pharmacogenetics of adverse drug reactions. Adv Pharmacol 2018, 83, 155–190.
- R., C.; V., N.; L., C.; N., C.; J.C., C. Genophenotypoc factors and Pharmacogenomics in Adverse Drug Reactions. Int J Mol Sci 2021, 22, 13302. [CrossRef]
- J.C., E.; R., A. Pharmacogenomics: What the Doctor ordered? Mo Med 2019, 116, 217–225.
- L., B. Pharmacogenomics of adverse drug reactions: Practical applications and prespectives. Pharmacogenomics 2009, 10, 961–969. [CrossRef] [PubMed]
- R., C. Pharmacogenomi of drugs to treat brain disorders. Expert Rev Prec Med Drug Dev 2020, 5, 181–234. [CrossRef]
- J., J.; J., K. PG-path: Modeling and personalizing phramacogenomis-based pathways. PLoS ONE 2020, 15, e0230950. [CrossRef]
- R.A., W.; D.M., R.; J.H., M. Combinatorial pharmacogenetics. Nat Rev Drug Discovery 2005, 4, 911–918. [CrossRef] [PubMed]
- Kozyra, M.; Ingelman-Sundberg, M.; Lauschke, V.M. Rare genetic variants in cellular transporters, metabolic enzymes, and nuclear receptors can be important determinants of interindividual differences in drug response. Genet. Med. 2017, 19, 20–29. [Google Scholar] [CrossRef]
- R., C.; N., C.; J.C., C. The role of pharmacogenomics in adverse drug reactions. Expert Rev clin Pharmacol 2019, 12, 407–442. [CrossRef] [PubMed]
- Z, D.; DA., F. Pharmacogenetics of drug metabolism. Clin Transl Sci 2017, 327–345.
- IS, L.; D., K. Polymorphic metabolism by functional alterations of human cytochrome P450 enzymes. Arch Pharm Res 2011, 34, 1799–1816. [CrossRef]
- J.D., S. The emerging field of neuroepigenetics. Neuron 2013, 80, 624–632. [CrossRef]
- B., M.; D.K., L. Epigenetics of dementia:understanding the disease as a transformation rather than a state. Lancet Neurol 2016, 15, 760–774. [CrossRef]
- A., B. The essentials of DNA methylation. Cell 1992, 70, 5–8. [CrossRef]
- A., H.; H., G.; A., J. Biochemistry and biology of mammal DNA methyltransferases. Cell Mol Life Sci 2004, 61, 2571–2587. [CrossRef] [PubMed]
- J., G.; I.M., M. Epigenetic codes in cognition and behaviour. Behav Brain Res 2008, 192, 70–87. [CrossRef]
- X., N.; F., V.; A., B. MeCP2 is a transcriptional repressor with abundant binding sites in genomic chromatin. cell 1997, 88, 4471–4481. [CrossRef]
- J.U., G.; Y., S.; J.H., S.; J., S.; H., L.; al, e. Distribution recognition and regulation of non-CpG methylation in the adult mamamlian brain. Nat Neurosci 2013, 17, 215–222. [CrossRef]
- A., H.; R., G.; A., J. The Dnmt1 DNA-cytosine-c5-methyltransferase methylates DNA processively with high preference for hemimethylated target sites. J Boil Chem 2004, 279, 48350–48359. [CrossRef]
- M., O.; S., X.; E., L. Cloning and characterization of a family of novel mammalian DNMA (cytosine-5) methyltransferases. Nat genet 1998, 19, 219–220. [CrossRef] [PubMed]
- O., M.-I.; I., C.; J.C., C.; L., F.-N.; N., C.; R., C. DNA methylation in neurodegenrative and cerebrovascular disorders. Int J Mol Sci 2020, 21, 2220. [CrossRef]
- O., M.-I.; V., N.; N., C.; R., C. Epigenetic biomarkers as diagnostic tools for Neurodegenrative Disorders. Int J Mol Sci 2022, 23, 13. [CrossRef]
- E., M.; W., D.; D, G.; P., D. Distinctive patterns of DNA methylation associated with Parkinson disease: Identification of concordant epigenetic changes in brain and peripheral blood leukocytes. Epigenetics 2013, 1030–1038. [CrossRef]
- a., D.F.; B., A.; A., F.; M., D.B.; M., K.; al, e. Global changes in DNA methylation in Alzheimer´s Disease peripheral blood mononuclear cells. Brain Behav Immun 2015, 45, 139–144. [CrossRef] [PubMed]
- O., M.-I.; V., N.; L., C.; R., P.; S., S.; S., R.; M., A.; A., M.; N., C.; R., C. DNA methylation as a biomarker for monitoring disease outcome in patients with hypovitaminosis and Neurological Disorders. Genes 2023, 14, 365. [CrossRef] [PubMed]
- S., C.; N., P.; M., M. Pharmaciogenetics of pase I and phase II drug metabolism. Curr Pharm Des 2010, 16, 204–219. [CrossRef]
- M., M. Role of CYO pharmacogenetics and drug-drug interactions in the efficacy and safety of atypical and other antipsychotic agents. J Pharmacy & Pharmacology 2010, 58, 871–885. [CrossRef]
- S., P.-L.; V., L. Biochemistry, Biotransformation. StatPearls 2022.
- S., P.; M., H.; R., P.; M., D.; A., G.; S., P. Polymorphic Citochrome P450 enzymes (CYPs) and their role in personalized therapy. PLoS ONE 2013. [CrossRef]
- Siokas, V.; Stamati, P.; Pateraki, G.; Liampas, I.; Aloizou, A.M.; Tsirelis, D.; Nousia, A.; Sgantzos, M.; Nasios, G.; Bogdanos, D.P.; et al. Analysis of SOD2 rs4880 Genetic Variant in Patients with Alzheimer's Disease. Current issues in molecular biology 2022, 44, 4406–4414. [Google Scholar] [CrossRef] [PubMed]
- Abaji, R.; Krajinovic, M. Thiopurine S-methyltransferase polymorphisms in acute lymphoblastic leukemia, inflammatory bowel disease and autoimmune disorders: Influence on treatment response. Pharmacogenomics and personalized medicine 2017, 10, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Relling, M.V.; Schwab, M.; Whirl-Carrillo, M.; Suarez-Kurtz, G.; Pui, C.H.; Stein, C.M.; Moyer, A.M.; Evans, W.E.; Klein, T.E.; Antillon-Klussmann, F.G.; et al. Clinical Pharmacogenetics Implementation Consortium Guideline for Thiopurine Dosing Based on TPMT and NUDT15 Genotypes: 2018 Updat. Clinical pharmacology and therapeutics 2019, 105, 1095–1105. [Google Scholar] [CrossRef]
- EEB, P.; LPC, L.; RB, A.; AAC, M.; BM, F.; RMR, B.; PP, A.; MR, F.; JF, G.; SEBD, S.; et al. UGT1A1 Gene Polymorphism Contributes as a Risk Factor for Lung Cancer: A Pilot Study with Patients from the Amazon. Genes 2022, 11, 493. [Google Scholar] [CrossRef]
- X, Z.; R, M.; X, M.; G., Y. Association of UGT1A1*6 polymorphism with irinotecan-based chemotherapy reaction in colorectal cancer patients: A systematic review and a meta-analysis. Biosci Rep 2020, 40, BSR20200576. [CrossRef] [PubMed]
- Y, Y.; M, Z.; M, H.; Y, C.; Q, Z.; L, L.; F., H. UGT1A1*6 and UGT1A1*28 polymorphisms are correlated with irinotecan-induced toxicity: A meta-analysis. Asia Pac J Clin Oncol 2018, 14, e479–e489. [CrossRef]
- J.T., Y.; L., T.; J., H. Apolipoprotein E in Alzheiemr´s Disease: An update. Ann Rev Neurosci 2014, 37, 79–100. [CrossRef] [PubMed]
- E., A.; Y., p.; S., P.; C., P. APOE genotype and Alzheimer´s disease: The influence of lifestyle and environmental factors. ACS Chem Neurosci 2021, 12, 2749–2764. [CrossRef]
- F., M.-B.; G., G.; G., M.; G., B.; E., C.; al, e. Pharmacogenomics in Alzheimer´s disease: A genome-wide association study of response to cholinesterase inhibitors. Neurobiol Aging 2013, 34, e7–e13. [CrossRef]
- R., C. Pharmacogenomics of cognitive and neuropsychiatric disorders in Dementia. Int J Mol Sci 2020, 21, 3059. [CrossRef]
- Kuring, J.K.; Mathias, J.L.; Ward, L. Prevalence of Depression, Anxiety and PTSD in People with Dementia: A Systematic Review and Meta-Analysis. Neuropsychol. Rev. 2018, 28, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Cacabelos, R. Pharmacogenomics of Cognitive Dysfunction and Neuropsychiatric Disorders in Dementia. Int J Mol Sci 2020, 21, 3059. [Google Scholar] [CrossRef] [PubMed]
- Z., C.; J., X.; C., W.; L., Q.; Y., F.; al., e. Population genetic differnece of pharmacigenomic VIP gene variants in the Lisu population from Yunnan Province. Medicine 2018, 97, e13674. [CrossRef]
- K., B.; P., C.; C., W.; P., W.; M., T.; al, e. Novel association between TGFA, TGFB1, IRF1, PTSG2 and IKBKB single-nucleotide polymorphisms and occurrence, severity and treatment response of major depressive disorder. Peer J 2020, 8, e8676. [CrossRef] [PubMed]
- X., T.; S., C. Epigenetic regulation of Cytochrome P450 enzymed and clinical implication. Curr Drug Metab 2015, 16, 86–96. [CrossRef] [PubMed]
- Y., S.; C., L.; G., L.; Y., C.; W., L.; al, e. Drug-metabolizing cytochrome P450 enzymes have multifarious influences on treatment outcomes. Clin Pharm 2021, 60, 585–601. [CrossRef] [PubMed]
- U.A., M. Pharmacogenetics and adverse drug reactions. Lancet 2000, 356, 1667–1671. [CrossRef] [PubMed]
- J., G.; X., Z.; S., B.; A., I.; Z., L.; C., X.; X., W. Metabolism and mechanism of human Cytochrome P450 Enzyme 1A2. Curr Drug Metab 2021, 22, 40–49. [CrossRef]
- R.A., P.; S.K., V.D.E.; C.M., T.; F., K.; D.M., U.; al, e. Coffee, ADORA2A, and CYP1A2: The caffeine connection in Parkinson´s disease. Eur J Neurology 2011, 18, 756–765. [CrossRef]
- L., v.T.; R., K.; H., V.; H-J., G. CYP1A2 activity is an important determinant of clozapine dosage in schizophrenic patients. Eur J Pharm Sci 2003, 20, 451–457. [CrossRef]
- L., D.; M., K. Clozapine therapy and CYP genotype. Med Genet Summaries 2021.
- R.C., G. Tacrine. Encyclopedia of Toxicology 2014, 466–467.
- R., C. Pharmacogenetic considerations when prescribing cholinesterase inhibitors for the treatment of Alzheimer´s disease. Expert Opin Drug Metab % Toxicology 2020, 16. [CrossRef]
- K., L.; K., R. Propanolol for migraine prophylasis. Cochrane Datanase Syst Rev 2017. [CrossRef]
- S., S.; A., v.W.; G., v.d.H.; R., v.W.; J., s.L.; A., d.J. Propanolol for the treatment of anxiety disorders: Systematic review and meta-analysis. J Phsychopharmacol 2016, 30, 128–139. [CrossRef]
- C., T.; E., A.; T., K.; R., A. Pharma GKB summary: Very important pharmacogene information for CYP1A2. Pharmacogenet Genom 2013, 22, 73–77.
- C.S., L. Microsomal ethanol-oxidizing system (MEOS): The first 30 years (1968-1998)- a review. Alcoholism: Clin % Exp Res 1999, 23, 991–1007. [CrossRef]
- W.A., G.-S.; L.A., R.-C.; M., R.-O.; M., C.-V.; J.A., A.-M.; J., G.; D., S.-A. The role of CYP2E1 in the drug metabolism or bioactivation in the brain. Oxid Med Cell Longev 2017, 2017, 4680732. [CrossRef]
- Y., B.; X., M. Chorzoxazone exhibits neuroprotection against AD by attenuating neuroinflammation and neurodegenration in vitro and in vivo. Int Immunopharmacol 2020, 88, 106790. [CrossRef] [PubMed]
- S., K.; B., S.; A., S.; S., T.-G.; K., Z.; U., S. hepatic. extrahepatic and extracellular vesicle Cytochrome P450 2E1 in alcohol and acetaminophen-mediated adverse intreactions and potential treatment options. Cells 2022, 11, 2620. [CrossRef]
- A., D. Pharmacogenomics of Warfarin. Stratified Medicine 2014, 14, 497–507.
- M., B.; S., H.-Z.; K., S.; K., L.; D., K.; H., O.H.; B., S. Dietary antioxidants remodel DNA methylation patterns in chronid disease. Br J Pharmacol 2019, 177, 1382–1408. [CrossRef]
- H., U.; W., M.; S.W., L.; H.S., K.; S., K.; H.H., W.; B.J., C.; D.K., K. association of the cjoline acetyltransferase gene with responsiveness to acetylcholinesterase inhibitors in Alzheimer´s disease. Pharmacopsych 2015, 48. [CrossRef]
- K., H.; J., L.; T., K. System pharmacogenomics-gene, disease, drug and placebo interactions: A case study in COMT. Pharmacogenomics 2019, 20, 529–551. [CrossRef] [PubMed]
- J., A.; R., T. Enzymmatic O-methylation of epinephrine and other catechol. J Biol Chem 1958, 233, 702–705. [CrossRef]
- N., J.; P., B.; V., J.; M., B.; E., S. NAT gene polymorphisms and susceptibility to Alzheimer´s disease: Identification of a novel NAT1 allelic variant. BMC Med Genet 2004, 5, 6. [CrossRef]
- Borlak J; S., R.-B. N-acetyltransferase 2 (NAT2) gene polymorphisms in Parkinson´s disease. BMC Med Genet 2006, 7, 30. [CrossRef]
- R, C.; L, F.-N.; R, A.; al, e. EPodoFavalin-15999 (Atremorine ® )-induced dopamine response in Parkinson’s Disease: Pharmacogenetics-related effects. J Genomic Med Pharmacogenomics 2016, 1, 1–26.
- O., M.-I.; V., N.; J.C., C.; I., C.; L., C.; al, e. AtreMorine Treatment Regulates DNA Methylation in Neurodegenerative Disorders: Epigenetic and Pharmacogenetic Studies. Curr Pharmacogen & Pers Medicine 2021. [CrossRef]
- S., A.; Z., Z.; J., Z.; S-Q., C. Pharmacogenomics of drug metabolizing enzymes and transporters: Relevance to precision medicine. Genomics, Proteomics & Bioinformatis 2016, 14, 298–313.
- R., C.; C., T.; I., C. Opportinities in pharmacogenomics for the treatment of Alzheimer´s disease. Future Neurol 2015, 10, 229–252. [CrossRef]
- R., C.; I., C.; O., M.; R., A.; L., F.-N.; al, e. Atremorine in Parkisnon´s disease: From dopaminergic neuroprotection to pharmacogenomics. Medic Res Rev 2021, 1–46.
- R., C.; J.C., C.; L., C.; L., F.-N.; R., P.; al, e. Influence of pathogenic and metabolic genes on the pharmacogenetic of mood disorders in Alzheimer´s disease. Pharmaceuticals 2021, 14, 366. [CrossRef] [PubMed]
- R., F.-C.; S., K.; J., F.-R.; J., G.-R.; L., C.-F.; al, e. APOE in the bullseye of neurodegenrative diseases: Impact of the APOE genotype in Alzheimer´s disease pathology and brain disease. Mol Neurodegeneration 2022, 17, 62. [CrossRef]
- DW, D. ; al, e. E4 is associated with severity of Lewy body pathology independent of Alzheiemr pathology. Neurology 2018, 91, e1182–e1195. [CrossRef] [PubMed]
- N., Z.; al, e. APOE4 exacerbates a-synuclein pathology and related toxicity independent of amyloid. Sci Transl Med 2020, 12, eaay1809. [CrossRef]
- E., W.; al, e. APOE varepsilon4 carriers show delayed recovery of verbal memory and smaller enthrinal volume in the fisrt year after ischemic stroke. J Alzheim Dis 2019, 71, 245–259. [CrossRef]
- ST., P.; al, e. APOE-epsilon4 genotype and dementia before and after transient ischemic attack and stroke: Population based cohort study. Stroke 2020, 51, 751–758. [CrossRef] [PubMed]
- O., M.-I.; V., N.; J.C., C.; S., S.; N., C.; R., C. Gene expression profiling as a novel diagnostic tool for Neurodegenrative Disorders. Int J Mol Sci 2023, 24, 5746. [CrossRef] [PubMed]
- K., R.; al, e. Plasma levels of apolipoprotein E and risk of dementia in general population. Ann Neurol 2014, 77, 301–311.
- F., P.; E., C.; I., A.; C., F.; L., T. Predictors of response to acetylcholinesterase inhibitors: A systematic review. Front Neurosci 2022, 16, 998224. [CrossRef] [PubMed]
- R., C.; J.C., C.; L., C.; R., P.; N., C.; al, e. Pharmacogenetics of anxiety and depression in Alzheimer´s disease. Future Med 2023, 24.
Figure 1.
Distribution of CYP phenotypes in patient groups A and B based on changes in 5mC levels. The impact of genetic variability in drug metabolizing enzymes on 5mC levels was investigated by analyzing the frequency of different CYP phenotypes in two patient groups: those whose 5mC levels increased during follow-up (Group A) and those whose levels decreased or remained similar to the initial visit (Group B). The distribution of CYP phenotypes was analyzed for each SNP. (A) CYP1A1 rs1378942, (B) CYP1A2 rs2069514, rs35694136, and rs762551, (C) CYP1B1 rs1056836, (D) CYP2A6 rs28399433, (E) CYP2B6 rs3745274, (F) CYP2C9 rs1057910, rs1799853, rs28371685, rs28371686, rs7900194, and rs9332131, (G) CYP2C19 rs12248560 and rs4244285, (H) CYP2D6 rs28371725, rs35742686, rs3892097, and rs5030655, (J) CYP3A4 rs2242480 and rs35599367, (K) CYP3A5 rs776746, and (L) CYP4F2 rs2108622 SNP. Percentages of normal, intermediate, fast, deficient, and abnormal metabolizer phenotypes are indicated for each SNP.
Figure 1.
Distribution of CYP phenotypes in patient groups A and B based on changes in 5mC levels. The impact of genetic variability in drug metabolizing enzymes on 5mC levels was investigated by analyzing the frequency of different CYP phenotypes in two patient groups: those whose 5mC levels increased during follow-up (Group A) and those whose levels decreased or remained similar to the initial visit (Group B). The distribution of CYP phenotypes was analyzed for each SNP. (A) CYP1A1 rs1378942, (B) CYP1A2 rs2069514, rs35694136, and rs762551, (C) CYP1B1 rs1056836, (D) CYP2A6 rs28399433, (E) CYP2B6 rs3745274, (F) CYP2C9 rs1057910, rs1799853, rs28371685, rs28371686, rs7900194, and rs9332131, (G) CYP2C19 rs12248560 and rs4244285, (H) CYP2D6 rs28371725, rs35742686, rs3892097, and rs5030655, (J) CYP3A4 rs2242480 and rs35599367, (K) CYP3A5 rs776746, and (L) CYP4F2 rs2108622 SNP. Percentages of normal, intermediate, fast, deficient, and abnormal metabolizer phenotypes are indicated for each SNP.
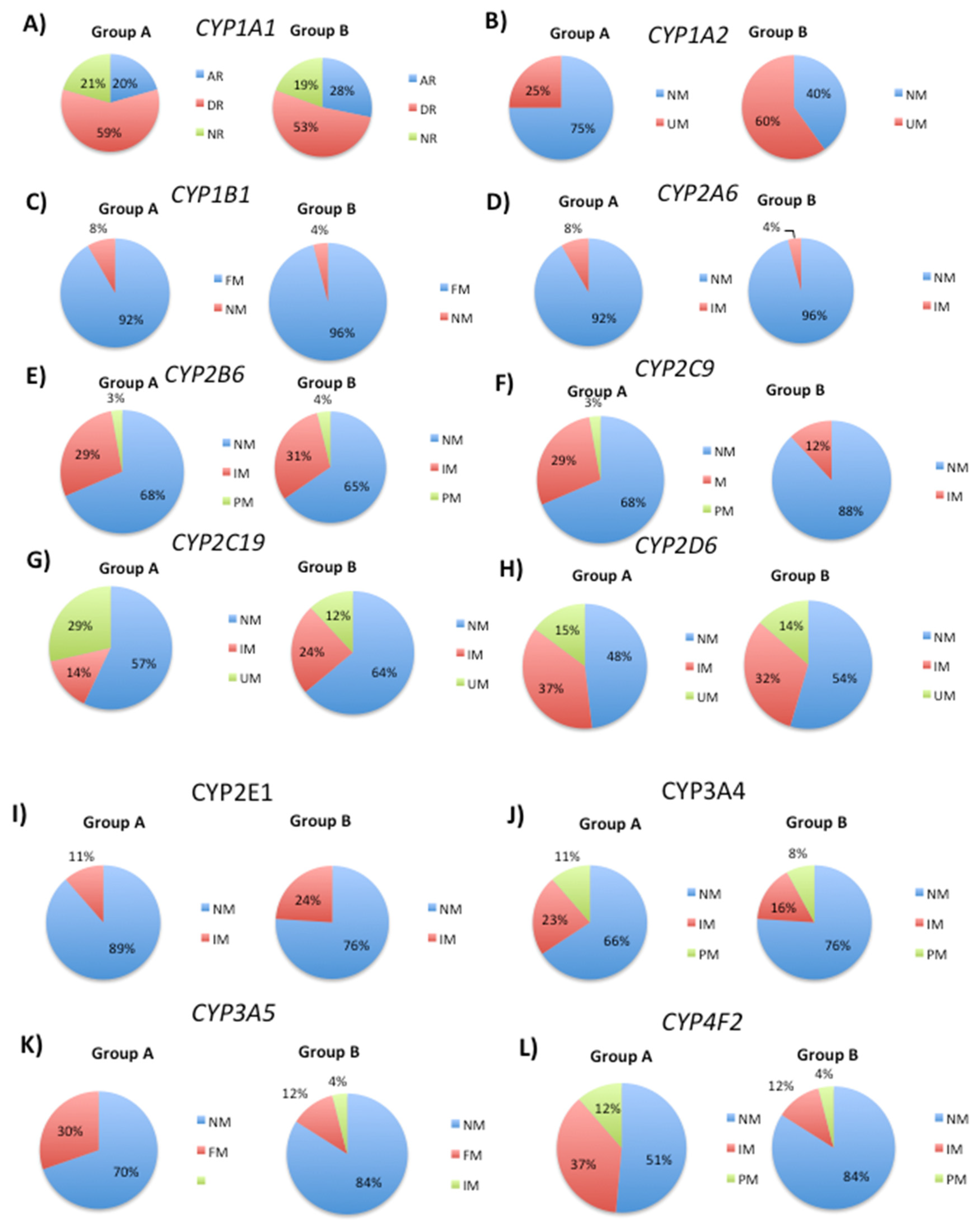
Figure 2.
Distribution of different metabolizer phenotypes in patient groups A and B based on the analysis of several SNPs.
Figure 2.
Distribution of different metabolizer phenotypes in patient groups A and B based on the analysis of several SNPs.
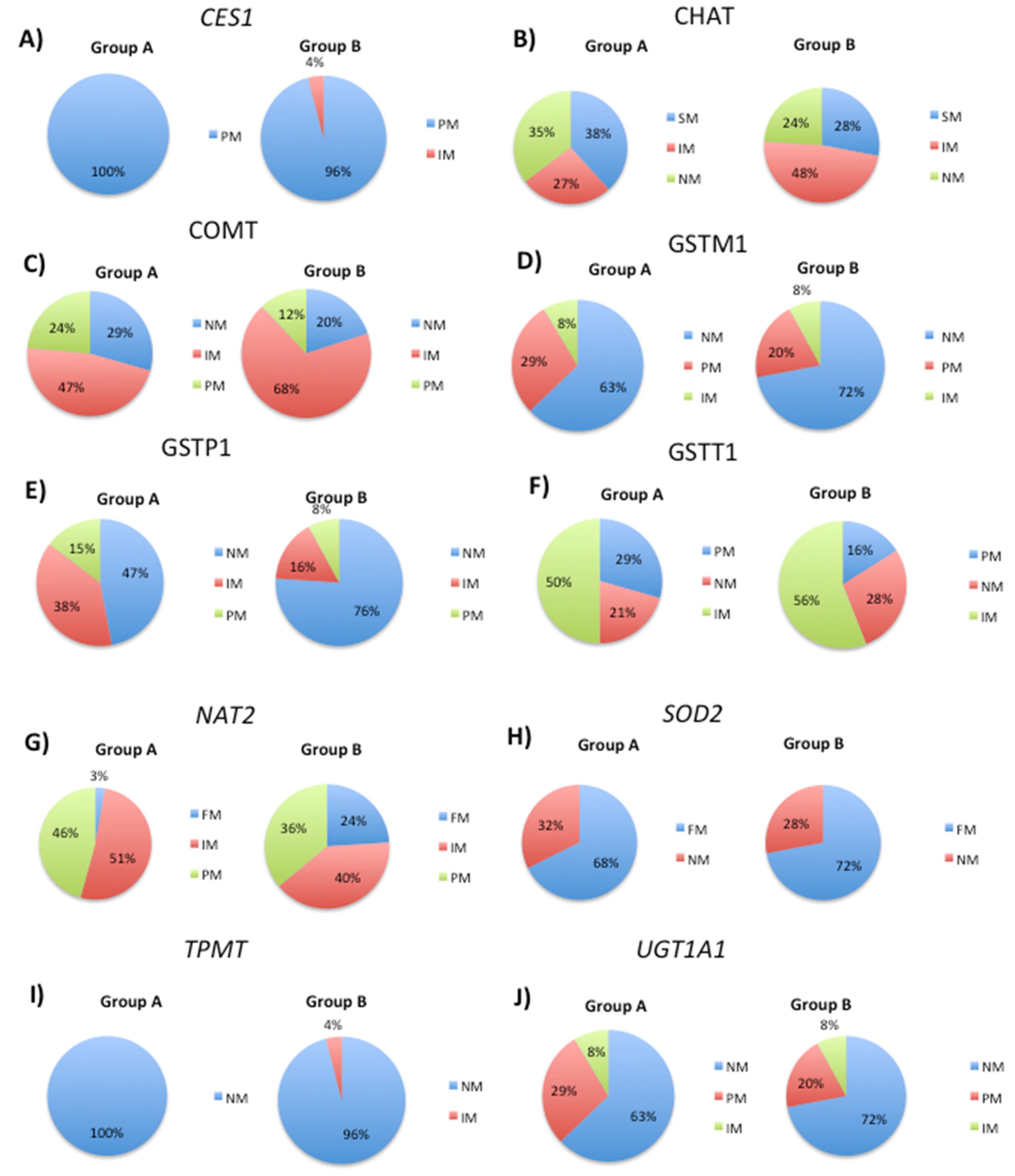
Figure 3.
Pie charts showing the distribution of transport gene phenotypes in patient groups A and B. The frequencies of normal, deficient, and abnormal response phenotypes were determined for each gene, and specific SNPs were analyzed to investigate their effect on phenotypic responses to treatment.
Figure 3.
Pie charts showing the distribution of transport gene phenotypes in patient groups A and B. The frequencies of normal, deficient, and abnormal response phenotypes were determined for each gene, and specific SNPs were analyzed to investigate their effect on phenotypic responses to treatment.
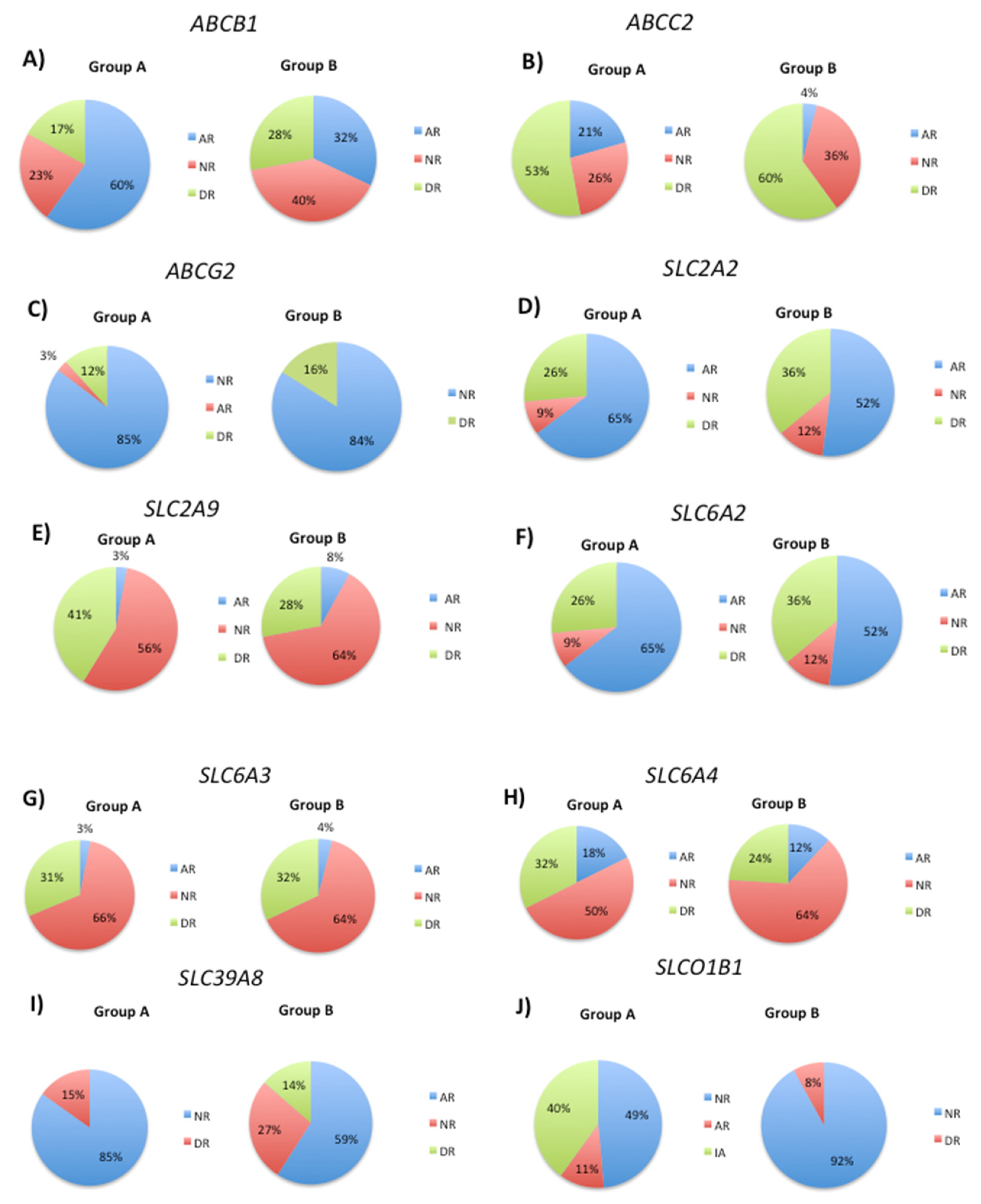
Figure 4.
Distribution of APOE genotypes in Group A and Group B patients.
Figure 5.
Frequency of different response phenotypes to acetylcholinesterase inhibitors (AChEIs) in Group A and Group B patients.
Figure 5.
Frequency of different response phenotypes to acetylcholinesterase inhibitors (AChEIs) in Group A and Group B patients.
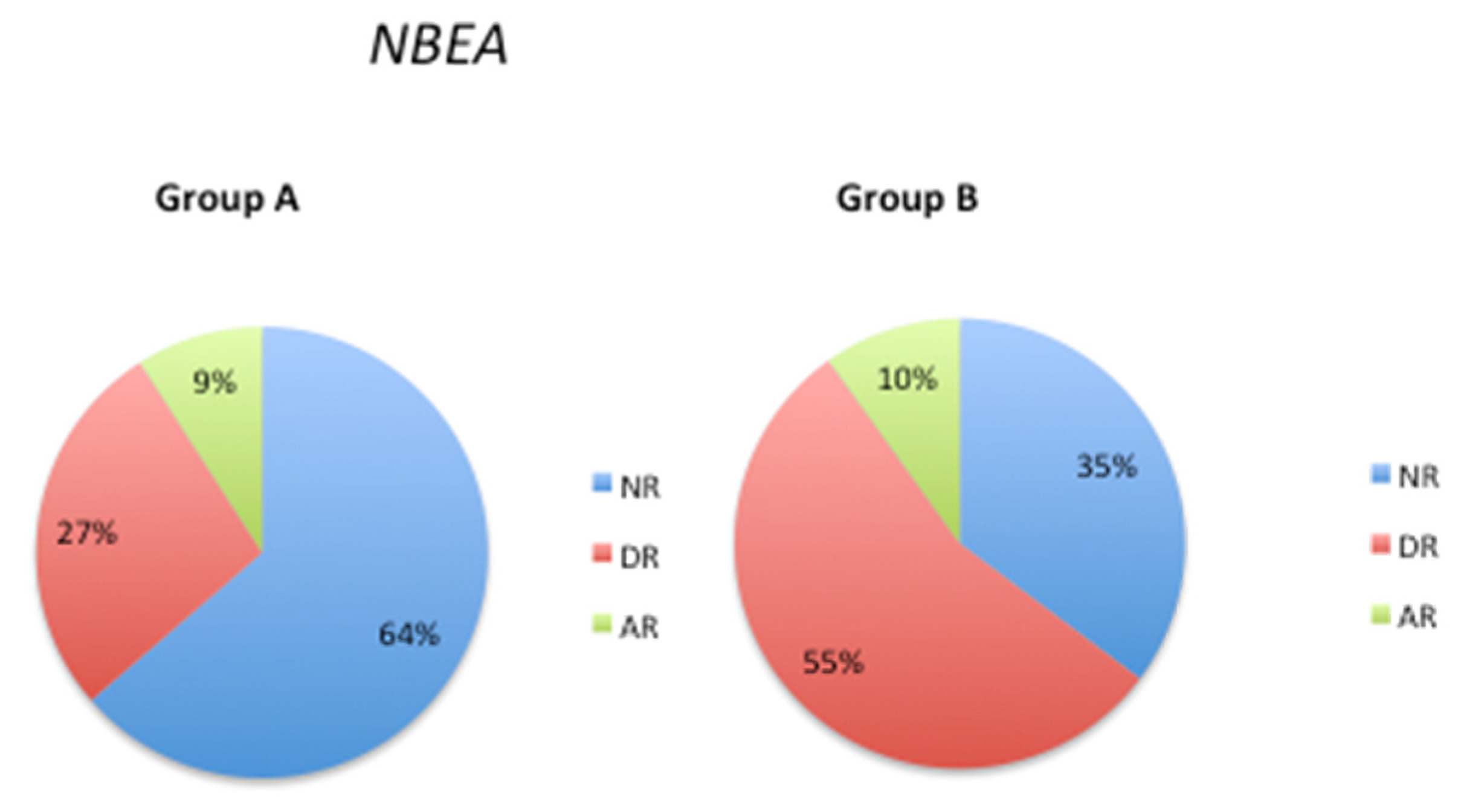
Figure 6.
Distribution of PTGS2 SNP rs5275 response phenotypes in Group A and Group B patients.
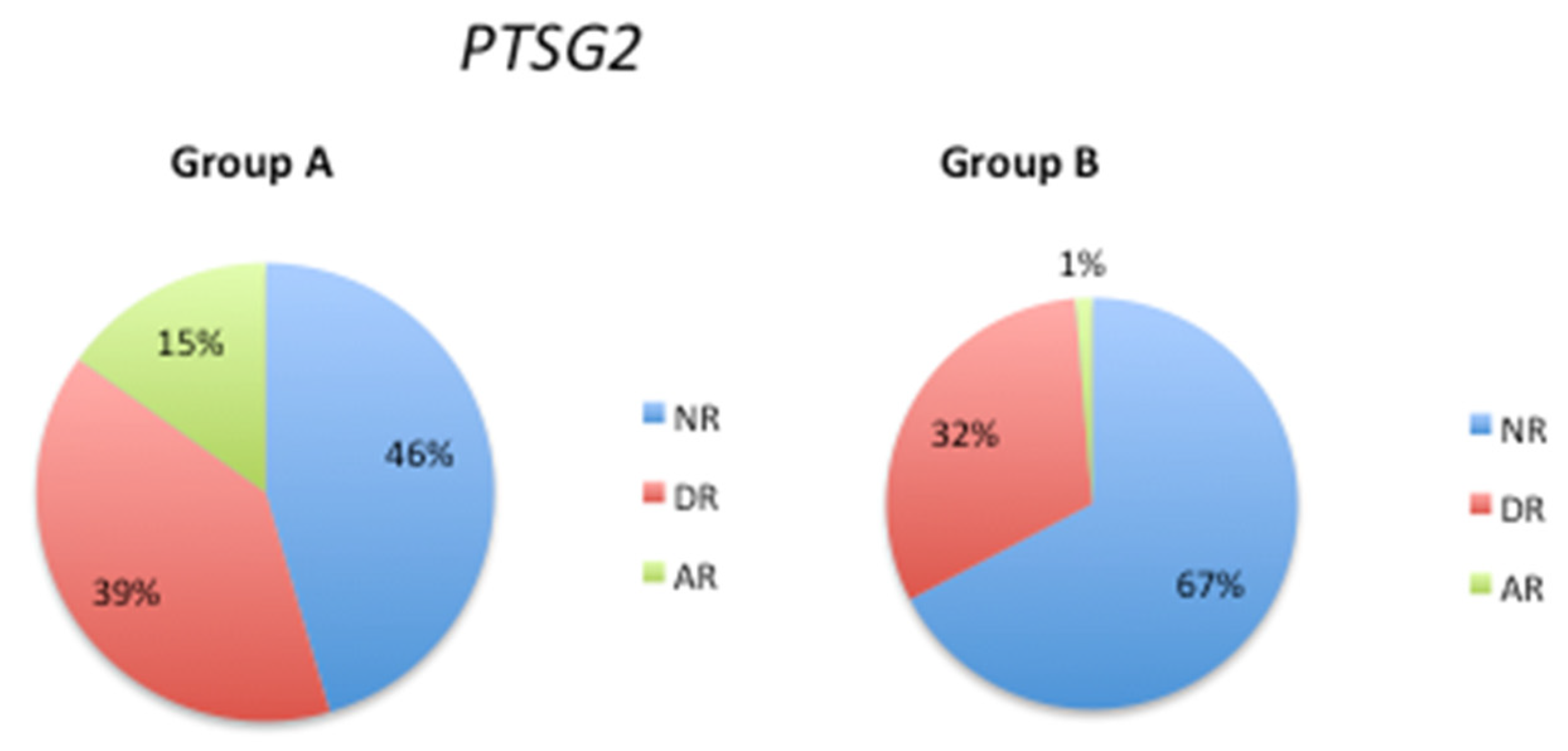

Table 1.
Summary of the pharmacogenetic genes that influence global DNA methylation levels during patient follow-up.
Table 1.
Summary of the pharmacogenetic genes that influence global DNA methylation levels during patient follow-up.
| Metabolice (Phase I) | Metabolic (Phase II) | Transporter | Pathogenic |
|---|---|---|---|
| CYP1A2 | GSTP1 | ABCB1 | APOE |
| CYP2C9 | NAT2 | ABCC2 | NBEA |
| CYP4F2 | SLC2A9 | PTSG2 | |
| SLC39A8 | |||
| SLCO1B1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



