Submitted:
20 July 2023
Posted:
25 July 2023
You are already at the latest version
Abstract
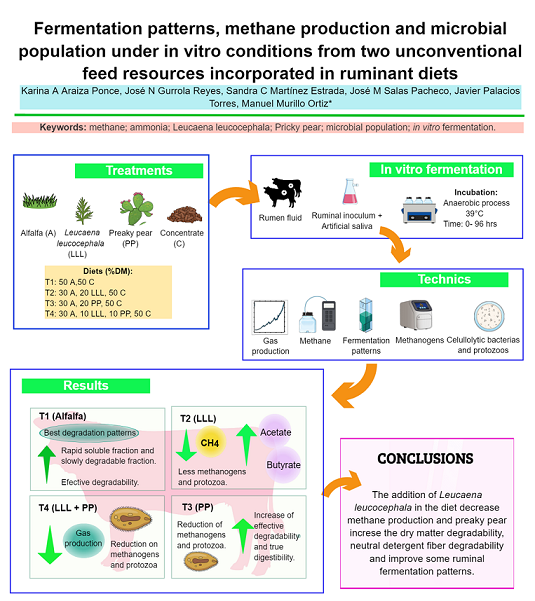
In this study, four experimental treatments were evaluated: (T1) alfalfa hay + concentrate, (50:50%, DM); (T2) alfalfa hay + Leucaena leucocephala + concentrate, (30:20:50%, DM); (T3) alfalfa hay + prickly pear + concentrate, (30:20:50%, DM); and (T4) alfalfa hay + Leucaena leucocephala + prickly pear + concentrate, (30:10:10:50%, DM). NH3-N concentrations in T2 and T4 decreased when replaced alfalfa hay in 20 and 10 %, respectively. Treatments did not affect the concentration of total volatile fatty acids (TVFA) between T3 and T4 (p>0.05); while the concentrations among T1 and T2 were different (P<0.05). T2 showed a reduction of 25.5 % in the methane production when compared to T1(p < 0.05).The lowest concentrations of protozoa were observed in T2 and T4, which contained Leucaena leucocephala (T2) and Leucaena leucocephala + prickly pear (T4) (p<0.05). The highest concentration of total methanogens was recorded in T1 and were different to T2, T3 and T4 (p<0.05). Leucaena leucocephala at a inclusion percentage of 20 % decreased the methane when compared to T1; whereas prickly pear there was not a positive effect the methane production.
Keywords:
methane
; ammonia
; Leucaena leucocephala
; Pricky pear
; microbial population
; in vitro fermentation
1. Introduction
The growing world population demands food that includes proteins of animal origin such as beef and milk. The intensive production of meat and milk worldwide requires the supply of fodder and energy concentrates to livestock. However, ruminal fermentation of forages and concentrates produces high amounts of methane (CH4). Therefore, considerable efforts have been devoted to find alternative sources of forage that contribute to satisfying the nutritional requirements of ruminants and that contribute to the reduction of methane production in the rumen [1]. The CH4 is produced from carbon dioxide and hydrogen as a by-product in rumen fermentation. Agriculture accounts for about 47% to 56% of total anthropogenic methane emissions [2]. It is known that from the agricultural sector, dairy, bovine, caprine and ovine livestock contribute substantially to the increase in CH4 production through the continuous process of ruminal fermentation. Hence, livestock activity contributes to the emission of greenhouse gases (GHG), contributing almost 30% of the total anthropogenic methane emissions into the atmosphere [3]. It has recently been recognized that due to their secondary metabolite content, some plants, leaves, fruits and roots reduce methane production in the rumen [4]. In vitro studies have shown that secondary metabolites such as tannins have antimethanogenic activity, either directly by inhibiting methanogens or indirectly by attacking protozoa [5]. Plants with bioactive compounds (saponins and tannins) to modify fermentation and rumen inhibition of methanogenesis are one option and are generally safe, cheap, and readily available [6]. In this way, some plants such as LLL and PP can be used as alternative sources of forage in ruminant feeding. The LLL is a highly available legume species commonly used as forage for ruminant feeding [7]. Also, PP has recently been introduced into diets and supplements to maintain ruminant body condition during dry periods. [8]. Although numerous studies on the effects of plants with a high tannin content such as LLL on rumen fermentation have found reductions in enteric methane production, the mechanisms of how the tannin content of these plants reduce methane production in vitro are still unclear. In accordance with the above, it is hypothesized that the incorporation of LLL and PP into ruminant diets decreases methane production in vitro and alters the rumen microbial population. Therefore, the objective of this study was to evaluate the effects of incorporating LLL and PP in ruminant diets on gas and methane production, rumen fermentation patterns and the amount of methanogenic bacteria during in vitro fermentation.
2. Materials and Methods
2.1. Location of area study and ethical procedure
The experiment was carried out in the animal metabolic unit and nutrition laboratory of the Faculty of Veterinary Medicine and Animal Science of the Juárez University in Durango (Mexico). Surgical procedures and management of rumen fistulated steers that were used to obtain rumen fluid were performed in accordance with the guidelines established by the Animal Protection Committee of the State of Durango (Mexico) and in accordance with the Official Mexican Standard NOM-062-ZOO-2019.
2.2. Plant collection, sampling and chemical analysis of Leucaena leucocephala leaves and prickly pear
Samples of LLL and PP were collected in a silvopastoral pasture and in medium-sized arbosufrutescent rangeland, respectively. In general, the climate of this region is semi-arid where the harsh conditions of drought are caused by the lack of rain. The lowest temperature is observed in winter around 5 °C; while in summer it fluctuates between 25 – 35 °C. The average annual rainfall is about 350 mm, distributed over a period of 60 rainy days during the summer. The LLL and PP leaf samples were dried at 40°C in a forced-air oven for 72 h and ground through a 1-mm mesh prior to chemical analysis and in vitro assays. The chemical composition of forage resources is shown in Table 1.
2.3. Chemical composition of the forage sources and experimental treatments
In each experimental diet, alfalfa hay, LLL and PP were used as a forage source. The chemical composition of the forage sources are shown in Table 1.
Four treatments were evaluated: (T1) alfalfa hay + concentrate, (50:50%, DM); (T2) alfalfa hay + LLL + concentrate, (30:20:50%, DM); (T3) alfalfa hay + PP + concentrate, (30:20:50%, DM); and (T4) alfalfa hay + LLL + PP + concentrate (30:10:10:50%, DM).
2.4. -Nutritional composition of the experimental treatments
The nutritional composition of the experimental treatments is shown in Table 2.Samples from each experimental treatment were analyzed in triplicate for dry matter (DM), organic matter (OM), crude protein (CP) and ether extract (EE) [9].
Analysis of neutral detergent fiber (NDF), acid detergent fiber (ADF), and acid detergent lignin (ADL) were determined using the filter bag technique with a fiber analyzer (ANKOM Technology, Fairport, NY, EE. UU.). The total carbohydrate contents (TCH) were calculated according to the equation proposed by Sniffen et al. [10]: %TCH = 100 – (% CP+ % EE+ % ash); while the content of non-fibrous carbohydrates (NFC) was calculated from the difference between %TCH and % NDF . The true in vitro digestibility of dry matter (TIVDMD) was determined using a Daisy incubator (ANKOM Technology, Fairport, NY, EE. UU.). Total tannins were calculated as the difference between total phenols and non-tannin phenols. The condensed tannins were measured by the HCL-butanol method [11].
2.5. In vitro gas and methane production parameters
In vitro gas production was measured using the ANKOM gas production system. Rumen fluid was collected approximately 3 h after morning feeding from two steers with rumen fistula. Rumen fluid was immediately squeezed through four layers of gauze and transported to the laboratory in a sealed thermos. The resulting ruminal fluid was purged with deoxygenated CO2 before using it as inoculum. Approximately 1 g of dried and ground samples from each treatment were weighed and placed in glass modules. Rumen fluid buffered with McDougall's buffer (20 ml) was pipetted into each module. Gas production was recorded after 2, 4, 6, 8, 12, 16, 24, 36, 48, 72, and 96 h of incubation. The accumulated production of gas (Y) in time (t) was adjusted to the model proposed by McDonald, [12]: GP= a + b x (1- exp -Kdx(t-L); where GP= gas production, a = Gas production from the rapidly soluble fraction, b = Gas production from the slowly degradable fraction, (a + b) = potential gas production, Kd = Constant rate of gas production, t = incubation time, L = delay time. Gas relative production (GRP) was estimated with the following model: GRP (ml/g MS) = a + (bc/ (Kd + Kp )) e-KpL [13], where: a, b, Kd and t were previously defined. The passage rate (Kp) was estimated from the model proposed by Haugen et al. [14]: Kp (%/h)= 0.07 IVDMD (%) – 0.20. To measure CH4 and CO2 production, once the incubation period is over at 24 h, The pressure release valve of each glass module was opened for 2 seconds and the gas released in each module was passed through a tube and connected to a portable gas analyzer to measure CH4 according to procedures proposed by the equipment manufacturer (GEMTM5000, LANDTEC, USA).
2.6. Dry matter intake and in vitro degradability parameters of dry matter and neutral detergent fiber.
Dry matter intake (DMI) was estimated according to Khazaal et al. [15] using the following model: DMI (g DM/kg LW75) = 18,9 + 0,23 (a + b) + 687 (Kd) + 0,11 (CP); while the kinetics of DM and NDF degradation were performed in the Daisy II incubator (ANKOM, Technology Corp., Fairport, NY, EE. UU.). Bags with 2.0 g of each treatment (3 replicates) were incubated in a module (21 modules/flask) containing ruminal fluid combined with a buffer solution (1:4, vol/vol). The ruminal fluid was obtained from two steers cannulated in the rumen, which were fed with a diet containing 60% oat hay and 40% concentrate. Degradation patterns were recorded after 0, 4, 8, 12, 24, 30, 48, 72, and 96 h of incubation. The bags were removed from each module at the defined incubation times, then washed with cold water and processed in the ANKOM fiber analyzer (Fibertec 2010 (Tecator Comp)) for the determination of dry matter and neutral detergent fiber (NDF). The degradation curves of DM and NDF at the different incubation times were adjusted to the following model proposed by McDonald [12]: Dt= a + b * (1- exp(-Kd*(t-L); where Dt= degradability; a = rapidly soluble fraction; b = slowly degradable fraction, Kd = degradation rate constant, t = incubation time, L = latency time. The effective degradability (ED) was estimated as: ED= a + b*(Kd ⁄ (Kd+Kp) [16]; whereas mean retention time in the rumen (MRTR) from the following model: MRTR (h)= [ (1/kp) + 10] * 0.75 [14].
2.7. True digestibility in vitro of dry matter, rumen fermentation patterns and synthesis of microbial biomass of the experimental treatments
True digestibility in vitro of dry matter (TDIVDM) was determined using a Daisy incubator (ANKOM Technology, Fairport, NY, EE. UU.). Bags with 2.0 g of each experimental treatment (3 replicates) were incubated in a module (3 bags/module) containing ruminal fluid combined with a buffer solution (1:4, vol/vol). Rumen fluid was obtained from two steers cannulated in the rumen fed a diet containing 60% oat hay and 40% concentrate. The DM digestibility was recorded after 48 h of incubation. Percent weight loss was determined and recorded as the TDIVDM. After 24 h of incubation, two samples (5 ml) were taken of the liquid of the glass modules. The first subsample was acidified with 0.3 mL of 50% H2SO4 and the second subsample with 2.5 mL of 25% metaphosphoric acid. Both subsamples were immediately frozen at -40°C and then analyzed for ammonia nitrogen (NH3-N) and total volatile fatty acids (TVFA), respectively [17]. The microbial biomass synthesis yield (MBS) and partition factor (PF) were calculated using the TDIVDM (mg) and the volume of gas registered at 24 h (GP24) as follows manner: MBS (mg-1g DM) = TDIVDM (GP24 × 2.25); PF= TDIVDM/GP24 [18].
2.8. Rumen microbial population
For the extraction of DNA from each experimental treatment, rumen fluid was obtained from two steers with rumen fistulation, fed with alfalfa hay and concentrate in a 50:50 ratio, which were fed twice a day at 08:00 and 16:00 h. Rumen fluid was collected 4 h after morning feeding in thermos and transported directly to the laboratory. Rumen fluid was filtered through four layers of cheesecloth and kept at 39°C in a CO2 atmosphere. In glass modules of the ANKOM gas production system, 1 g of the ground samples of each treatment were introduced. Immediately, 125 ml of ruminal fluid and a buffer solution prepared according to Menke and Steingass were added [19]. All the glass modules containing the incubation medium and the treatments samples were incubated at 39°C for 24 h. After the 24 h incubation was complete, 50 mL of liquid was collected from each glass module and placed in tubes to centrifuge at 20,000 xg for 30 min. The supernatant was discarded and 0.5 g of the residue was immediately taken for deoxyribonucleic acid extraction.
2.8.1. Extraction of rumen microbial DNA.
Deoxyribonucleic acid extraction was performed using the method described by Rojas et al. [20]. DNA concentration was calculated using a NanoDrop 2000 (Thermo Scientific, Waltham, MA, EE. UU.) and DNA integrity was confirmed by agarose gel electrophoresis. The DNA samples obtained were stored at −80 ◦C until the quantitative analysis of microbial DNA. The YATP (g mole microbial cells -1 ATP) was calculated according to Czerkawski [21].
2.8.2. Quantitative analysis of real-time PCR populations
Microbial DNA was amplified from total DNA with specific primers for each population. The sequence of the primers used for the detection of bacteria and total methanogens is shown in Table 3. The primers used for the detection of total bacteria and methanogens were 16S rRNA and mcrA, respectively [22]. The specificity of the primers was verified with the conventional PCR technique using the Multigene Labnet 96-well thermal cycler (Labnet Corporation, Inc, Global). The number of copies was calculated from the formula proposed by Marconell [23]; while the absolute quantification was obtained with the equation proposed by Angarita et al. [24].
2.9. Protozoa and cellulolytic bacteria
The estimation of the population of protozoa in the ruminal fluid was carried out by diluting 8 ml of ruminal fluid with 16 ml of formal saline solution (1 part of 37% formalin and 9 parts of 0.9% saline solution) and counting the protozoa under an optical microscope (10x) using a Neubauer camera [25]. To assess the bacterial population, ruminal fluid samples were diluted 1:3 in formal saline and again diluted 103 in formal saline. Crystal violet (20 ml) was added to 200 ml of this solution and the stained bacteria were read under light microscopy (40x) using a Neubauer chamber [26].
2.10. Statistical analysis
All data were submitted to a completely randomized design and the significance of the differences between means was determined using Tukey's multiple range test. Differences at p<0.05 were considered statistically significant. All analyzes were performed using SAS [27], from the following statistical model:
where Yij is the response variable, µ is the overall mean, ti is the treatment effect, and eij is the error due to the j-th replicate of the i-th normally distributed treatment with zero mean and constant variance.
Yij = µ + ti + eij
3. Results
3.1. In vitro gas and methane production
Fractions "a" and "b" were different between treatments (p< 0.05), being higher for control treatment (T1) in relation to T2, T3 and T4 (Table 4). The Kd value was affected by the treatments (p<0.05).
Lower Kd value for T2 (4.0 ml-1h) , T3 (6.0 ml-1h) and T4 (4.5 ml-1h) was observed and the highest Kd was obtained with the control treatment(8.0 ml-1h) (p < 0.05). Despite, the nutrients supply by LLL leaves and PP in T2, T3 and P4, the potential gas production gas (PGP) and relative gas production (RGP) values were lower than the control treatment where alfalfa hay was a main source of forage (p< 0.05) . There were differences among treatments in the of methane (CH4) and carbon dioxide CO2) productions (p<0.05). Treatment with LLL (T2) showed a reduction of 25.5 % in the methane production (p < 0.05) when compared to control treatment (T1); whereas carbon dioxide production showed a increase of 28.6 % with T2 when compared to control treatment (T1) (p < 0.05). Moreover, there were differences between T1 and T2 in the CO2:CH4 ratio (p<0.05). The CO2:CH4 ratio showed a increase of 44.4 % with T2 when compared to control treatment (T1) (p < 0.05).
3.2.-. Dry matter intake, in vitro degradability parameters of dry matter and neutral detergent fiber.
Dry matter intake (DMI) was higher for T1 compared to the other treatments (p<0.05) (Table 5). There were differences between treatments in the values of rapidly degradable
fraction of dry matter (aDM), slowly degradable fraction of dry matter (bDM), potential degradability of the dry matter ( PDDM), effective degradability of dry matter (EDDM) and degradation rate constant of dry matter (kdDM) (p<0.05), except to rate passage of dry matter (kpDM) in T2, T3 and T4 (p>0.05). Treatment control (T1) showed a increase of 37.0 % in PDDM and 68.7 % in EDDM (p<0.05), respectively. There were differences between treatments in the values of rapidly degradable fraction of neutral detergent fiber (aNDF) (p<0.05). Furthermore, the slowly degradable fraction of neutral detergent fiber (bNDF), potential degradability of neutral detergent fiber (PDNDF) and effective degradability of neutral detergent fiber (EDNDF) values, were affected by treatments (p<0.05). The highest value of degradation rate constant of neutral detergent fiber (KdNDF) was recorded in T1 and the lowest in T2 (p<0,05). LLL and PP did not induce any effect on the rate passage of neutral detergent fiber (KpNDF) (p>0.05). Likewise, the highest value of KpNDF was recorded in T1 and the lowest in T2 and T4 (p<0,05). The longer values of mean rumen retention time of dry matter (MRRTDM) and mean rumen retention time of neutral detergent fiber (MRRTNDF) were observed in T2 and the slowest values in T1 (p<0,05).
3.3. True digestibility in vitro of dry matter (TDIVDM)), ruminal fermentation patterns and microbial biomass synthesis.
There were differences among treatments in the TDIVDM values (p<0,05) (Table 6). The highest values of TDIVDM were recorded in T1 and the lowest in T2 (p<0,05). At the same time, the concentrations of NH3-N were significantly affected by the treatments (p<0.05). In our study, the concentrations of NH3-N in T2 and T4 decreased when LLL replaced alfalfa hay by 20 and 10 %, respectively. Likewise, the concentration of total volatile fatty acids (TVFA) in the rumen liquor was statistically similar between T3 and T4 (p>0.05); while the concentrations between T1 and T2 were different (P<0.05). Acetate and propionate ruminal concentrations of were affected by the treatments (p<0.05). Acetate concentrations decreased when LLL replaced alfalfa hay in 20% (T2) and 10% (T4). However, propionate concentrations increased with both treatments. The highest values of microbial biomass synthesis (MBS) were recorded in T1 and the lowest in T2 (p<0,05). Control treatment (T1) showed a increase of 23.0 % 12.0 % and 14.0 % in relation with T2, T3 and T4, respectively (p<0,05). Partition factor (PF) values were statistically similar between T2, T3 and T4 (p>0.05); but different to T1 (p<0.05).
3.4. Rumen microbial population after in vitro incubation with rumen fluid
There were no differences between treatments in the total number of bacteria and celulolytic bacteria (p>0.05) (Table 7). However, there were differences between treatments in the population of protozoa (p< 0.05). The lowest concentrations of protozoa were observed in T2 and T4, which contained LLL (T2) and LLL + PP (T4) (p<0.05). The highest concentration of total methanogens was recorded in T1 and were different to T2, T3 and T4 (p<0.05). YATP values were not affected by treatments (p>0.05).
4. Discussion
4.1. In vitro gas and methane production
Low in vitro gas production parameters observed in T2 and T4 could be partly explained by the negative effect of tannins on ruminal fermentation [28]. The values of the in vitro gas production parameters obtained in the current study are in partial agreement with the findings of Khazaal et al. [29] and Torres et al. [30], who evaluated in vitro conditions phenolic compounds and alfalfa hay in addition to concentrates in a 50:50 range, respectively. Despite the nutrient supply of LLL leaves and PP in T2, T3 and T4 the "a", "b", "GPP" and "Kd" values were lower than the control treatment where alfalfa hay was a main source of forage, which could be due to the fact that rumen microbes were stimulated, as well as the digestibility of the incubated substrate, resulting in better gas production kinetics [31]. As regards to the decrease in CH4 production recorded in T2 and T4, could be attributed to the content of condensed tannins (CT) Beauchemin et al. [32]. This suggests that CT are at least partially responsible for this effect. According to Soltan et al. [33], LLL inhibits CH4 production both in vitro and in vivo conditions. The reduction in CH4 production is attributable not only to CT, but could be partly due to differences in other components of the diets, mainly cell wall components [34]. There are two mechanisms to reduce enteric CH4 production in ruminants through CT supplementation: a) indirectly through reduced fiber digestion, decreasing H and methane production through the CO2 pathway, and b) directly by inhibiting the growth of methanogens [35]. Highest structural carbohydrate contents were recorded at T2 and T4, hence the better CO2 production efficiency registered in T2 and T4 compared to the other treatments, could be explained from the degradation of the structural carbohydrates of both diets. Cellulolytic bacteria that hydrolyze the structural carbohydrates of the cell wall, generate acetate and CO2 as final products [36]. Greater production of acetate by ruminal fermentation causes a greater availability of CO2 [37].
4.2.- Dry matter intake, in vitro degradability parameters of dry matter and neutral detergent fiber.
Differences observed between treatments in DMI could be attributed to the NDF contents of the experimental diets [34]. Our results do not agree with Paengkoum [38], who found higher values of DMI when supplementing diets based on corn silage with LLL. Highest values of aDM, bDM, DPDM EDDM and KdDM recorded in T1, may suggest the availability of nutrients provided by carbohydrates and proteins from alfalfa hay [39]. Therefore, the higher values obtained for the aDM, bDM and EDDM fractions will indicate a better nutrient availability for rumen microorganisms. Furthermore, the slower value of KdMS recorded in T2 indicates that the amount of energy that can be extracted from the diets during the time it remains in the rumen is low. Our results are in partial agreement with the DM degradability parameters reported by Mohammadabadi et al. [40] who investigated the effect of replacing alfalfa hay with L. leucocephala leaves in proportions of 25, 50 and 100%. EDDM values registered in T1 could be associated with the high contents of NDF and TC, which become severe limitations to improve the ruminal digestion of the nutrients contained in the treatments and negatively affect the dry matter degradability. MRRTDM higher values and lower KpDM values observed in T2 and T4 can also be explained by the NDF contents. Minson [41] have pointed out that when the content of the cell walls increases, the KpDM decreases and the MRRTDM increases proportionally, causing the cellulose and hemicellulose to ferment slowly and this causes the physical filling of the rumen [42]. The highest kdNDF value was recorded in T1 and the lowest in T2 (p<0.05). This suggests that T2 was being broken down at a slower rate. In the present study, aNDF, bNDF, EDNDF, and kdNDF decreased when LLL replaced alfalfa hay by 20%. This may be due to the presence of anti-nutritional factors such as saponins and tannins in LLL [43]. KpNDF value was higher in the treatment with alfalfa hay as a forage source (T1) (p<0.05); while the other three were similar (p > 0.05). The decrease in the in vitro degradability parameters of NDF observed in T2 (aNDF, bNDF, EDNDF, and kdNDF) does not agree with the results obtained by Barros et al. [44], who found higher values including LLL at 20 % of the diet. These differences between both studies, can be attributed to differences in the methods to determine degradability [45].
4.3. True digestibility in vitro of dry matter, ruminal fermentation patterns and microbial biomass synthesis.
TDIVDM value in T1 could be attributed to a high synthesis of microbial biomass [46]; while in T2 it could be explained by the lower contribution of ammonia and non-fibrous carbohydrates (NFC) for microbial growth [47]. The decrease in ammonia concentrations in T2 and T4 could be explained from the tannin content of the diets. There is general agreement that tannins decrease the degradation of proteins provided by the diets, mainly through the formation of tannin-protein complexes, which helps to decrease the concentration of NH3-N [48]. Despite this trend, the NH3-N values observed in this study are within the optimal range to maximize microbial growth in the rumen, which is reported between 5 and 10 mg/dl [49]. Ruminal NH3-N concentrations are consistent with the results found by Kang et al. [50] when evaluating LLL in ruminant diets. The acetate decrease in T2 and T4 and increases in propionate concentrations in both treatments, can be attributed to the contents of structural carbohydrates (NDF) as well as non-fibrous carbohydrates (NFC) supplied by the experimental diets [51]. In fact, previous studies have consistently reported a decrease in the molar ratio of acetate and increases in the ratio of propionate under in vitro conditions using high-starch concentrates [52] and high fiber forages [53]. Generally, the results of the in vitro fermentation patterns obtained in this study, are consistent with the findings of previous studies in which alfalfa hay, LLL and PP were evaluated as sources of forage in ruminant diets [54]. MBS values recorded in T1 could be attributed to a greater supply of NH3-N by the experimental diet. TDIVDM estimates and MBS obtained in this study, do not agree with those found by Albores et al. [55], who found higher values in TDIVMD and MBS when including various levels of LLL in ruminant diets. PF is regularly used as an indicator of substrate degradation rate, as well as in vitro gas production efficiency and microbial biomass. In this study PF value are consistent with what was reported by Abdallah et al. [56], and were higher than the theoretically possible maximum value of 4.41 mg TDIVMD/ml of gas [57]. The increase in PF could indicate a lower partition of nutrients for the synthesis of microbial proteins [58].
4.4. Microbial population after in vitro incubation with rumen fluid
Regarding the number of total bacteria, the resulted obtained, agree with Pilajun and Wanapat [59], who reported that supplementation with tannin-rich plants did not change the total number of bacteria. However, other studies have shown that plants rich in tannins reduce the number of bacteria [60, 61]. Also, these results agree with Longo et al. [62], who found that the diversity indices of the methanogenic community did not change when LLL or other tannin-rich plants were supplied. Pineiro et al. [63], when evaluating the LLL in heifers fed with low-quality forage, did not observe changes in the concentration of protozoa. However, Barros et al. [44] reported that rumen protozoa decreased when ewes were fed 20% and 40% LLL. YATP values recorded in this study, are within the established ranges for different diets supplied to ruminants. For a mixed-species microbial population, the estimated YATP (grams dry weight of cells formed/mole ATP spent) is 29 to 30 for growth on rich media containing preformed monomers and from 20 to 29 for growth in simple media containing carbohydrates and inorganic salts [64].
5. Conclusions
In this experiment, the results revealed that the addition of alfalfa hay to diets as a forage source (control treatment) produced the best results, although the addition of LLL (T2) in the diet decreased methane production. However, the values in the in vitro gas production parameters, dry matter degradability, neutral detergent fiber degradability, ruminal fermentation patterns and protozoa population were higher in T3 (PP) than when compared with T2 (LLL) and T4 (LLL + PP). The results obtained in this research also indicate that both unconventional forage sources can be used in ruminant diets.
Author Contributions
Conceptualization, K.A.A-P. and M.M-O.; methodology,. J.N.G-R; software, J.P-T; validation, S.C.M-E and M.M-O.; formal analysis, J.N.G-R.; investigation, J.M.S-P.; resources, K.A.A-P and J.P-T; data curation, S.C.M-E; writing—original draft preparation, K.A.A-P; writing—review and editing, M.M-O. and J.N.G-R; visualization, S.C.M-E. and J.P-T; supervision, J.M.S-P; project administration, J.N.G-R; funding acquisition, M.M.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Durango Regional Livestock Union (Project DRLU-A-888).
Institutional Review Board Statement
Surgical procedures and management of rumen fistulated steers that were used to obtain rumen fluid were performed in accordance with the guidelines established by the State Committee for the Promotion and Protection of Livestock of the State of Durango (Mexico) and in accordance with the Official Mexican Standard NOM-062-ZOO-2019.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ku, V.J.C.; Castelan, O.O.A.; Galindo, M.F.A.; Arango, J.; Chirinda, N.; Jiménez, O.R.; Valencia, S.S.S.; Flores, S.E.J.; Montoya, F.M.D.; Molina, B.I.C.; Piñeiro, V.A.T.; Arceo, C.J.I.; Aguilar, P.C.F.; Aviles, R.L.; Solorio, S.F.J. Review: Strategies for enteric methane mitigation in cattle fed tropical forages. Animal. 2020, 14, 453–463. [Google Scholar]
- Jackson, R.B.; Saunois, M.; Bousquet, P.; Canadell, J.G.; Poulter, B.; Stavert, A.R.; Bergamaschi, P.; Niwa, Y.; Segers, A.; Tsuruta, A. Increasing anthropogenic methane emissions arise equally from agricultural and fossil fuel sources. Environ. Res. Lett. 2020, 15, 071002. [Google Scholar] [CrossRef]
- Hernández, M.J.H. El metano y la ganadería bovina en México: ¿Parte de la solución y no del problema? . Agro Productividad. 2018, 11, 2, 46–51. [Google Scholar]
- Ibrahim, T.A.; Hassen, A.; Apostolides, Z. The Antimethanogenic Potentials of Plant Extracts: Their Yields and Phytochemical Compositions as Affected by Extractive Solvents. Plants 2022, 11, 3296. [Google Scholar] [CrossRef]
- Aboagye, I.A.; Beauchemin, K.A. Potential of Molecular Weight and Structure of Tannins to Reduce Methane Emissions from Ruminants: A Review. Animals 2019, 9, 856. [Google Scholar] [CrossRef]
- Srinivas, B.; Krishnamoorthy, U. Panoply of microbial protein production in ruminants. A review Indian. J. Anim. Sci. 2013, 83, 4, 331–346. [Google Scholar]
- Montoya, F.M.D.; Botero, M.I.C.; Arango; J.; Muñoz, R.J.J.; Solorio, S.F.J.; Aguilar, P.C.F.; Ku, V.J.C. Effect of dried leaves of Leucaena Leucocephala on rumen fermentation, rumen microbial population, and enteric methane production in crossbred heifers. Animals, 2020, 10, 300.
- Pastorelli, G.; Serra, V.; Vannuccini, C.; Attard, E. Opuntia spp. as Alternative Fodder for Sustainable Livestock Production. Animals 2022, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- AOAC International, 2019. Official Methods of Analysis of AOAC International. 21th Edition. Gaithersburg, MD.
- Sniffen, C.J.; O'Connor, J.D.; Van Soest, P.J.; Fox, D.G.; Russell, J.B. A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci. 1992, 70, 3562–3577. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.S. Quantification of tannins in tree foliage. IAEA. (FAO/IAEA Working Document). 2000. Vienna, Austria.
- McDonald, I. A revised model for the estimation of protein degradability in the rumen. J. Agric. Sci. 1981, 96, 251–252. [Google Scholar] [CrossRef]
- Edmunds, B.; Südekum, K.-H.; Spiekers, H.; Schuster, M.; Schwarz, F. Estimating utilisable crude protein at the duodenum, a precursor to metabolisable protein for ruminants, from forages using a modified gas test. Anim. Feed. Sci. Technol. 2012, 175, 106–113. [Google Scholar] [CrossRef]
- Haugen, H.L.; Ivan, S.K.; MacDonald, J.C.; Klopfenstein, T.J. Determination of undegradable intake protein digestibility of forages using the mobile nylon bag technique1. J. Anim. Sci. 2006, 84, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Khazaal, K.; Dentinho, M.T.; Ribeiro, J.M.; Ørskov, E.R. Prediction of apparent digestibility and voluntary intake of hays fed to sheep: comparison between using fibre components, in vitro digestibility or characteristics of gas production or nylon bag degradation. Anim. Sci. 1995, 61, 527–538. [Google Scholar] [CrossRef]
- rskov, ER.; Mcdonald, I. The estimate of protein degradability in the rumen from incubation measurements weighed according to rate of passage. J. Agri. Sci. 1979, 92, 499–503. [Google Scholar] [CrossRef]
- Galyean, M.L. Laboratory Procedures in Animal Nutrition Research. Department of Animal and Food Sciences Texas Tech. 2010. University, Lubbock. USA.
- Blümmel, M.; Steingass, H.; Becker, K. The relationship between in vitro gas production, in vitro microbial biomass yield and 15N incorporation and its implications for the prediction of voluntary feed intake of roughages. Brit. J. Nut. 1997, 77, 911–92. [Google Scholar] [CrossRef] [PubMed]
- Menke, K.H.; Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Develop. 1988, 28, 7–55. [Google Scholar]
- Rojas, H.R.; Narváez, Z.J.; Zamudio, M.M.; Mena, M.M.E. A simple silica-based method for metagenomic DNA extraction from soil and sediments. Molecular Biotech. 2008, 40, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Czerkawski, J. Reassessment of Efficiency of Synthesis of Microbial Matter in the Rumen. J. Dairy Sci. 1978, 61, 1261–1273. [Google Scholar] [CrossRef]
- Denman, S.E.; Tomkins, N.W.; McSweeney, C.S. Quantitation and diversity analysis of ruminal methanogenic populations in response to the antimethanogenic compound bromochloromethane. FEMS Microbiol. Ecol. 2007, 62, 313–322. [Google Scholar] [CrossRef]
- Marconell A., C. Estudio de la dinámica poblacional de arqueas metanogénicas en un reactor anaerobio de membranas mediante PCR cuantitativa y métodos microscópicos y su relación con los parámetros operacionales. 2017. Available online: http://hdl.handle.net/10251/90999.
- Angarita, E.; Molina, I.; Villegas, G.; Mayorga, O.; Chará, J.; Rosales, R.B. Quantitative analysis of rumen microbial populations by qPCR in heifers fed on Leucaena leucocephala in the Colombian Tropical Dry Forest. Acta Sci. Anim. Sci. 2015, 37, 135–142. [Google Scholar] [CrossRef]
- Ley De Coss, A.; Cobos, P.M.A.; Hernández. S.D.; Medina, G.E. Formulation of an anaerobic culture medium of rumen ciliate protozoa and in vitro evaluation in the defaunant capacity of the extract of plants. Revista Científica, FCV-LUZ. 2011, 21, 43 – 49.
- Londoño, Z.A.F.; Fernandez, C.J.; Molina, G.L.P.; Polanco, E.D.; Gutierrez, B.L.A. Quantification of anaerobic cellulolytic bacteria from the rumen of cattle: comparison of three techniques. Hechos Microb. 2011, 2, 51–59. [Google Scholar]
- SAS Institute, Inc. Statistical analysis system, Windows version 9.1.2. 2003. SAS Institute, Inc., Cary, NC.
- Kelln, B.; Penner, G.B.; Acharya, S.N.; McAllister, T.A.; Lardner, H.A. Impact of condensed tannin-containing legumes on ruminal fermentation, nutrition, and performance in ruminants: a review. Can. J. Anim. Sci. 2021, 101, 210–223. [Google Scholar] [CrossRef]
- Khazaal, K.; Boza, J.; Ørskov, E. Assessment of phenolics-related antinutritive effects in Mediterranean browse: a comparison between the use of the in vitro gas production technique with or without insoluble polyvinylpolypyrrolidone or nylon bag. Anim. Feed. Sci. Technol. 1994, 49, 133–149. [Google Scholar] [CrossRef]
- Torres, F.K.; Páez, L.J.; Pàmanes, C.G.; Herrera, T.E.; Carrete, C.F.; Murillo, O.M. Substitution of garlic leaves to alfalfa hay and its effect in vitro ruminal fermentation. Abanico Veterinario. 2020, 10, 1–11. [Google Scholar]
- Phesatcha, K.; Phesatcha, B.; Wanapat, M.; Cherdthong. A. Mitragyna speciosa korth leaves supplementation on feed utilization, rumen fermentation efficiency, microbial population, and methane production. Fermentation. 2022, 8, 2-11.
- Beauchemin, K.A.; McGinn, S.M.; Martinez, T.F.; McAllister, T.A. Use of condensed tannin extract from quebracho trees to reduce methane emissions from cattle1. J. Anim. Sci. 2007, 85, 1990–1996. [Google Scholar] [CrossRef]
- Soltan, Y.A.; Morsy, A.S.; Sallam, S.M.; Lucas, R.C.; Louvandini, H.; Kreuzer, M.; Abdalla, A.L. Contribution of condensed tannins and mimosine to the methane mitigation caused by feedingLeucaena leucocephala. Arch. Anim. Nutr. 2013, 67, 169–184. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional Ecology of the Ruminant; Cornell University Press: Ithaca, NY, USA, 1994; p. 476. [Google Scholar] [CrossRef]
- Tavendale, M.H.; Meagher, L.P.; Pacheco, D.; Walker, N.; Attwood, G.T.; Sivakumaran, S. Methane production from in vitro rumen incubations with Lotus pedunculatus and Medicago sativa, and effects of extractable condensed tannin fractions on methanogenesis. Anim. Feed. Sci. Technol. 2005, 123-124, 403–419. [Google Scholar] [CrossRef]
- Cai, S.; Li, J.; Ze, H.F.Z.; Zhang, K.;. Lou, Y,; Janto, B.; Boissy, R.; Ehrlich, G.E.; Dong, X. Cellulosilyticum ruminicola, a newly described rumen bacterium that possesses redundant fibrolytic-protein-encoding genes and degrades lignocellulose with multiple carbohydrate borne fibrolytic enzymes. Appl. Environ. Microb. 2010, 76, 3818–3824.
- Billman, E.D.; Leanne, S. D:, Roca, F.A.I.; Soder, K.J. Supplementation of oilseeds to an herbage diet high in condensed tannins affects methane production with minimal impact on ruminal fermentation in continuous culture. Fermentation. 2022, 8, 3, 109.
- Paengkoum, P.; Paengkoum, S. ORIGINAL ARTICLE: Effects of supplementing rice straw with Leucaena (Leucaena leucocephala) and Madras thorn (Pithecellobium dulce) foliages on digestibility, microbial N supply and nitrogen balance of growing goats. J. Anim. Physiol. Anim. Nutr. 2010, 94, e59–e65. [Google Scholar] [CrossRef]
- Shahriari, Z.; Mohammadabadi, T.; Vakili, S. T.; Chaji, M.; Sari, M. Effect of replacing alfalfa with subabul (Leucaena leucocephala) pod on digestibility, in vitro fermentation and in situ degradability in cow and buffalo. J. Anim. Prod Res. 2017, 6, 63–72. [Google Scholar]
- Mohammadabadi, T.; Chaji, M.; Direkvandi, E.; Alqaisi, O. Effect of replacing alfalfa hay with Leucaena leucocephala (L. Leucocephala) leaves on in vitro gas production, digestibility and in situ degradability in buffalo Acta Scientiarum. Anim. Sci. 2021, 43, 521-529.
- Minson, D.J. Forage in Nutrition Ruminant. 1990. Academic Press, INC. United Kingdom. Edition.
- Wang, J.; Zhang, Z.; Liu, H.; Xu, J.; Liu, T.; Wang, C.; Zheng, C. Evaluation of Gas Production, Fermentation Parameters, and Nutrient Degradability in Different Proportions of Sorghum Straw and Ammoniated Wheat Straw. Fermentation 2022, 8, 415. [Google Scholar] [CrossRef]
- Kakengi, A.M.; Shem, M.N.; Mtengeti, E.P.; Otsyina, R. Leucaena leucocephala leaf meal as supplement to diet of grazing dairy cattle in semiarid Western Tanzania. Agrofor. Syst. 2001, 52, 73–82. [Google Scholar] [CrossRef]
- Barros, R.M.A.; Solorio, S.F.J.; Sandoval, C,A.F.; Klieve, A.; Rojas, H.R.A.; Briceño, P.E.G.; Ku, V.J.C. Rumen function in vivo and in sheep fed Leucaena Leucocephala. Trop. Anim. Health. Prod. 2015, 47, 757–764.
- Spanghero, M.; Gruber, L.; Zanfi, C. Precision and accuracy of the NDF rumen degradability of hays measured by the Daisy fermenter. Ital. J. Anim. Sci. 2007, 6, 363–365. [Google Scholar] [CrossRef]
- E Griswold, K.; Hoover, W.H.; Miller, T.K.; Thayne, W.V. Effect of form of nitrogen on growth of ruminal microbes in continuous culture. J. Anim. Sci. 1996, 74, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Tu, Y.; Zhang, N.F.; Deng, K.D.; Diao, Q.Y. Effect of the Ratio of Non-fibrous Carbohydrates to Neutral Detergent Fiber and Protein Structure on Intake, Digestibility, Rumen Fermentation, and Nitrogen Metabolism in Lambs. Asian-Australasian J. Anim. Sci. 2015, 28, 1419–1426. [Google Scholar] [CrossRef] [PubMed]
- Rira, M.; Morgavi, D.P.; Archimède, H.; Marie-Magdeleine, C.; Popova, M.; Bousseboua, H.; Doreau, M. Potential of tannin-rich plants for modulating ruminal microbes and ruminal fermentation in sheep1. J. Anim. Sci. 2015, 93, 334–347. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekharaiah, M.; Thulasi, A.; Suresh, K.P.; Sampath, K.T. Rumen degradable nitrogen requirements for optimum microbialprotein synthesis and nutrient utilization in sheep fed on finger milletstraw (Eleucine coracana) based diet. Anim. Feed Sci. Technol. 2011, 163, 130–135. [Google Scholar] [CrossRef]
- Kang, S.; Wanapat, M.; Pakdee, P.; Pilajun, R.; Cherdthong, A. Effects of energy level and Leucaena leucocephala leaf meal as a protein source on rumen fermentation efficiency and digestibility in swamp buffalo. Anim. Feed. Sci. Technol. 2012, 174, 131–139. [Google Scholar] [CrossRef]
- Wei, Z.; Xie, X.; Xue, M.; Valencak, T.G.; Liu, J.; Sun, H. The Effects of Non-Fiber Carbohydrate Content and Forage Type on Rumen Microbiome of Dairy Cows. Animals 2021, 11, 3519. [Google Scholar] [CrossRef]
- Wang, K.; Xiong, B.; Zhao, X. Could propionate formation be used to reduce enteric methane emission in ruminants? Sci. Total. Environ. 2023, 855, 158867. [Google Scholar] [CrossRef]
- Krueger, N.A.; Adesogan, A.T.; Staples, C.R.; Krueger, W.K.; Kim, S.C.; Littell, R.C.; Sollenberger, L.E. Effect of method of applying fibrolytic enzymes or ammonia to Bermudagrass hay on feed intake, digestion, and growth of beef steers1. J. Anim. Sci. 2008, 86, 882–889. [Google Scholar] [CrossRef]
- Araiza, P.K.; Murillo, OM.; Herrera, T.E.; Valencia,V.R.; Carrete, C.F.; Pámanes, C.G. Leucaena Leucocephala and Opuntia ficus-indica reduce the ruminal methane production in vitro. 2020, Abanico Veterinario, 10: 1-13.
- Albores, M.S.; Alayón, G.J.A.; Mirana, R.L.A.; Alarcón, Z.B.; Jiménez, F.G.; Ku, V.J.C.; Piñero, V.A.T. Effect of tree foliage supplementation of tropical grass diet on in vitro digestibility and fermentation, microbial biomass synthesis and enteric methane production in ruminants. Trop. Anim. Health. Prod. 2019, 51, 893–904. [Google Scholar] [CrossRef]
- Abdalla, A.L.; Louvandini, H.; Sallam, S.M.A.H.; Bueno, I.C.d.S.; Tsai, S.M.; Figueira, A.V.d.O. In vitro evaluation, in vivo quantification, and microbial diversity studies of nutritional strategies for reducing enteric methane production. Trop. Anim. Heal. Prod. 2012, 44, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Blümmel, M.; Cone, J.W.; Van Gelder, A.H.; Nshalai, I.; Umunna, N.N.; Makkar, H.P.S.; Becker, K. Prediction of forage intake using in vitro gas production methods Comparison of multiphase fermentation kinetics measured in an automated gas test, and combined gas volume and substrate degradability measurements in a manual syringe system. Anim. Feed Sci. Technol. 2005, 123–124. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Sen, S.; Blümmel, M.; Becker, K. Effects of fractions containing saponins from Yucca schidigera, Quillaja saponaria and Acacia auriculoformis on rumen fermentation. J. Agric. Food. Chem. 1998, 46, 4324–4328. [Google Scholar] [CrossRef]
- Pilajun, R.; Wanapat, M. Microbial population in the rumen of swamp buffalo (Bubalus bubalis) as influenced by coconut oil and mangosteen peel supplementation. J. Anim. Physiol. Anim. Nutr. 2013, 97, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Saxena, J. Dietary phytochemicals as rumen modifiers: a review of the effects on microbial populations. Antonie van Leeuwenhoek 2009, 96, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Anantasook, N.; Wanapat, M.; Cherdthong, A.; Gunun, P. Changes of Microbial Population in the Rumen of Dairy Steers as Influenced by Plant Containing Tannins and Saponins and Roughage to Concentrate Ratio. Asian-Australasian J. Anim. Sci. 2013, 11, 1583–1591. [Google Scholar] [CrossRef]
- Longo, C.; Abdalla, A.; Liebich, J.; Janzik, I.; Hummel, J.; Correa, P.; Südekum, K.-H.; Burauel, P. Evaluation of the effects of tropical tanniferous plants on rumen microbiota using qRT PCR and DGGE analysis. Czech J. Anim. Sci. 2013, 3, 106–116. [Google Scholar] [CrossRef]
- Piñeiro, V.A.T.; Canul, S.J.R.; Jimenez, F.G.O.; Alayon, G.J.A.; Chay, C.A.J.; Ayala, B.A.J.; Aguilar, P.C.F.; Ku, V.J.C. Effect of condensed tannins from Leucaena Leucocephala on rumen fermentation, methane production and population of rumen protozoa in heifers fed low-quality forage. Asian. Australas. J. Anim. Sci. 2018, 31, 11,1738–1746. [Google Scholar]
- Hespell, R.B.; Bryant, M.P. Efficiency of rumen microbial growth: influence of some theoretical and experimental factors of YATP. J. Anim. Sci. 1979, 49, 1640-1659.Author 1, A.; Author 2, B. Title of the chapter. In Book Title, 2nd ed.; Editor 1, A., Editor 2, B., Eds.; Publisher: Publisher Location, Country, 2007; Volume 3, pp. 154–196.

Table 1.
Chemical compositions of the three forage sources (g Kg-1 DM).
| Alfalfa hay | LLL | PP | |
| DM | 897 | 895 | 900 |
| OM | 871 | 915 | 720 |
| CP | 167 | 213 | 53 |
| NDF | 450 | 429 | 483 |
| Lignin | 81 | 53 | 48 |
| TDIVMD | 557 | 457 | 515 |
| NFC | 234 | 670 | 649 |
| TPC g tannic acid eq/kg DM | 96.5 | 119.6 | 101.2 |
| CT mg /g DM | 0.40 | 0.98 | 0.51 |
DM= Dry matter; OM= Organic matter; CP= Crude protein; NDF= Neutral detergent fiber; TPC= Total phenolic compounds; CT= condensed tannins; TDIVMD = True digestibility in vitro of dry matter; NFC= Nonfibrous carbohydrate.
Table 2.
Nutritional composition of experimental treatments.
| Treatments | ||||
| Ingredient (g Kg-1 DM). | T1 | T2 | T3 | T4 |
| Alfalfa hay | 500 | 300 | 300 | 300 |
| Leucaena Leaves | 0 | 200 | 0 | 100 |
| Prickly pear | 0 | 0 | 200 | 100 |
| Corn milled | 350 | 370 | 280 | 340 |
| Cottonseed | 140 | 120 | 210 | 150 |
| Minerals | 10 | 10 | 10 | 10 |
| Chemical composition (g Kg-1 DM). | ||||
| DM | 883 | 879 | 803 | 898 |
| OM | 904 | 915 | 874 | 883 |
| CP | 140 | 146 | 142 | 148 |
| EE | 31 | 27 | 21 | 24 |
| NDF | 422 | 473 | 424 | 461 |
| Lignin | 41 | 58 | 45 | 55 |
| NFC | 292 | 218 | 273 | 245 |
| TPC g tannic acid eq/kg DM | 105.4 | 122.5 | 95.9 | 106.0 |
| CT mg /g DM | 0.305 | 3.34 | 0.360 | 1.96 |
| ME Mcal/ Kg-DM* | 3.8 | 3.1 | 3.4 | 3.0 |
DM= Dry matter; OM= Organic matter; CP= Crude protein; EE= ether extract; matter; NDF= Neutral detergent fiber; ADF=Acid detergent fiber; TPC= Total phenolic compounds; CT= condensed tannins; NFC= Nonfibrous carbohydrate; *Estimated from the equation ME (Mcal kg-1 DM) = 2.20 + 0.136 Gas production24h + 0.057 CP + 0.0029 ether extract2/4.184 [19].
Table 3.
Primer sequences used to quantify total bacteria by qPCR.
| Primer sequences used to quantify total bacteria by qPCR. | |||
| Gene 16S rRNA |
Sequence (5´- 3´) | Extension | |
| Forward | 5´CGGCAACGAGCGCAACCC3´ | 130 bp | |
| Reverse | 5´CCATTGTAGCACGTGTGTAGCC3´ | ||
| Primer sequences used to quantify total methanogens by qPCR | |||
| Gene mcrA |
Sequence (5´- 3´) | Extension | |
| Forward | 5´TTCGGTGGATCDCARAGRGC3´ | 128 bp | |
| Reverse | 5´ GBARGTCGWAWCCGTAGAATCC 3´ | ||
mcrA = Methyl-coenzyme M reductase; bp= base pairs.
Table 4.
In vitro gas parameters and methane production of experimental treatments.
| Treatments | SEM | p< value | |||||
| T1 | T2 | T3 | T4 | ||||
| a (ml 200 mg-1 DM) | 16.1a | 7.1d | 12.3b | 9.4c | 1.8 | 0.001 | |
| b (ml 200 mg-1 DM); | 102.0a | 85.7d | 98.3b | 92.1c | 2.3 | 0.01 | |
| PGP (ml 200 mg-1 DM) | 118.1a | 92.8c | 110.6b | 101.5b | 1.1 | 0.05 | |
| RGP (ml 200 mg-1 DM) | 110.1a | 85.8d | 101.6b | 93.5c | 1.7 | 0.003 | |
| Kd (ml-1h) | 8.0a | 4.0d | 6.0b | 4.5c | 0.01 | 0.01 | |
| L (h); | 2.5 | 3.3 | 3.1 | 3.1 | 0.33 | 0.120 | |
| CH4 (ml g-1 DM); | 13.7b | 10.2d | 15.8a | 12.8c | 3.3 | 0.01 | |
| CO2 (ml g-1 DM). | 74.8d | 96.2a | 87.5c | 91.1b | 1.2 | 0.001 | |
| CO2:CH4: ratio | 5.4b | 7.8a | 5.5b | 7.1a | 0.004 | 0.310 | |
abcd Means within the same row with various superscripts are significantly different (p<0.05). a = Gas production from quickly soluble fraction; b = Gas production from insoluble fraction; PGP = Potential gas production; RGP= Relative gas production; Kd = Gas production rate; L= Discrete lag time prior to gas production; CH4= Methane; CO2= Carbon dioxide; SEM = Standard error of mean.
Table 5.
Intake and dry matter in vitro degradability parameters and neutral detergent fiber.
| Treatments | SEM | p< value | |||||
| T1 | T2 | T3 | T4 | ||||
| DMI (g -1 LW0.75) | 73.0a | 41.2d | 62.2b | 48.6c | 2.9 | 0.001 | |
| aDM (mg g-1 DM) | 27.4a | 17.6d | 22.1b | 19.7c | 2.1 | 0.002 | |
| aNDF (mg g-1 NDF) | 12.4a | 10.5c | 11.7b | 11.1b | 1.8 | 0.04 | |
| bDM (mg g-1 DM) | 58.2a | 44.9d | 51.3b | 48.3c | 2.0 | 0.004 | |
| bNDF (mg g-1 NDF) | 71.1a | 62.6d | 69.8b | 66.3c | 2.3 | 0.01 | |
| PDDM (mg g-1 DM) | 85.6a | 62.5d | 73.4b | 68.0c | 1.5 | 0.005 | |
| PDNDF (mg g-1 NDF) | 83.1a | 73.1d | 81.5b | 77.4c | 2.1 | 0.05 | |
| EDDM (mg g-1 DM) | 65.8a | 39.0d | 54.6b | 47.0c | 1.1 | 0.004 | |
| EDNDF (mg g-1 NDF) | 45.3a | 28.3d | 42.0b | 40.5c | 1.6 | 0.05 | |
| KdDM (mg-1h) | 8.2a | 3.3d | 7.1b | 5.2c | 0.005 | 0.005 | |
| KdNDF (mg-1h) | 5.0a | 2.0c | 4.0b | 4.0b | 0.002 | 0.01 | |
| LDM (h) | 2.0 | 2.3 | 2.1 | 2.0 | 0.98 | 0.05 | |
| LNDF (h) | 3.3 | 4.0 | 3.5 | 3.8 | 1.7 | 0.18 | |
| KpDM (mg-1h) | 4.2a | 3.6b | 4.1a | 4.0a | 0.002 | 0.05 | |
| KpNDF (mg-1h) | 5.8a | 5.0b | 5.2b | 5.0b | 0.007 | 0.05 | |
| MRRTDM (h) | 23.8c | 27.7a | 24.3c | 25.0b | 1.1 | 0.002 | |
| MRRTNDF (h) | 12.5c | 18.2a | 14.6b | 14.9b | 2.5 | 0.03 | |
SEM = Standard error of mean.
Table 6.
True degradability in vitro dry matter, fermentation ruminal patterns and microbial biomass synthesis of experimental treatments.
Table 6.
True degradability in vitro dry matter, fermentation ruminal patterns and microbial biomass synthesis of experimental treatments.
| Treatments | SEM | p< value | ||||
| T1 | T2 | T3 | T4 | |||
| TDIVMD48h, (mg -1 100 mg DM) | 667a | 622d | 654b | 642c | 2.4 | 0.030 |
| pH | 6.60a | 6.6a | 6.6a | 6.5a | 0.017 | 0.854 |
| N-NH3, (mg dL-1) | 12.6a | 8.7d | 11.5b | 9.3c | 0.152 | 0.024 |
| TVFA, (mM/L) | 10.6a | 6.5.d | 7.9b | 7.5b | 0.281 | 0.148 |
| Volatile fatty acids (molar%) | ||||||
| Acetate | 66.8d | 72.0a | 67.7c | 70.5b | 0.161 | 0.741 |
| Propionate | 24.4a | 18.2d | 22.3b | 19.3c | 0.674 | 0.033 |
| Butyrate | 5.7b | 9.1a | 9.3a | 9.0a | 0.247 | 0.027 |
| A:P ratio | 2.7 | 3.9 | 3.0 | 3.6 | 0.143 | 0.911 |
| MBS (mg -1 g DM) | 165.2a | 132.2d | 147.4b | 145.1c | 1.13 | 0.007 |
| PF (mg TDMD/mL gas) | 6.0a | 6.5b | 6.5b | 6.3b | 1.05 | 0.050 |
abcdMeans within the same row with various superscripts are significantly different (P<0.05). TDMD48h= True degradability dry matter; TVFA= Total volatile fatty acids MBS= Microbial biomass synthesis; PF= Partition factor. SEM = Standard error of mean.
Table 7.
Ruminal microbial population of the experimental treatments after in vitro incubation with rumen fluid.
Table 7.
Ruminal microbial population of the experimental treatments after in vitro incubation with rumen fluid.
| Treatments | SEM | p< value | ||||
| T1 | T2 | T3 | T4 | |||
| Total bacteria1 | 14.7 | 14.9 | 15.2 | 15.4 | 0.030 | 0.22 |
| Celulolytic bacteria4 | 7.6 | 7.6 | 7.6 | 5.0 | 0.076 | 0.98 |
| Protozoa3 | 16.6a | 7.3d | 13.3b | 10.2c | 0.082 | 0.04 |
| Total methanogens2 | 14.2a | 13.5b | 13.8b | 13.6b | 0.066 | 0.05 |
| Methanogen:bacteria ratio | 0.95 | 0.91 | 0.91 | 0.88 | 0.091 | 0.88 |
| YATP (g microbial cells mol-1 ATP) | 24.0 | 22.2 | 24.0 | 23.3 | 0.61 | 0.930 |
abcValues with different letters in the same row are statistically different (P<0.05);1,2Log [ngDNAg-1 RC]; 3(x104 CFU -1.ml); 4(x106 CFU -1ml); RC= Ruminal content; CFU= Colony forming units. SEM = Standard error of mean.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



