Submitted:
24 July 2023
Posted:
24 July 2023
You are already at the latest version
Abstract
Pregnancy associated plasma protein-A (PAPP-A) plays an integral role in breast cancer (BC), especially triple negative breast cancer (TNBC). This subtype accounts for the most aggressive BC, possesses high tumor heterogeneity, is least responsive to standard treatments and has the poorest clinical outcomes. There is a critical need to address the lack of effective targeted therapeutic options available. PAPP-A is a protein that is highly elevated during pregnancy. Frequently, higher PAPP-A expression is detected in tumors than in healthy tissues. The increase in expression coincides with increased rates of aggressive cancers. In BC, PAPP-A has been demonstrated to play a role in tumor initiation, progression, metastasis including epithelial-mesenchymal transition (EMT), as well as acting as a biomarker for predicting patient outcomes. In this review, we present the role of PAPP-A, with specific focus on TNBC. The structure and function of PAPP-A, belonging to the pappalysin subfamily, and its proteolytic activity are assessed. We highlight the link of BC and PAPP-A with respect to the IGFBP/IGF axis, EMT, the window of susceptibility and the impact of pregnancy. Importantly, the relevance of PAPP-A as a TNBC clinical marker is reviewed and its influence on immune-related pathways are explored. The relationship and mechanisms involving PAPP-A reveal the potential for more treatment options that can lead to successful immunotherapeutic targets and the ability to assist with better predicting clinical outcomes in TNBC.
Keywords:
pregnancy associated plasma protein-A
; breast cancer
; triple negative breast cancer
; epithelial-mesenchymal transition
; cancer therapy
; cancer biomarker
; immunotherapy
1. Introduction
Pregnancy associated plasma protein-A (PAPP-A) is a protein originally detected in 1974 in high quantities in the plasma of third-trimester pregnant women. Along with three other placental antigens identified, PAPP-A was found to be susceptible to proteinases and the molecular weight was estimated to be 750,000 MW at the time of initial discovery [1]. The function of PAPP-A remained unknown till its role in proteolytic activity of insulin-like growth factor (IGF) -dependent insulin-like growth factor-binding proteins (IGFBP) was revealed for the first time in cell culture media conditioned by human fibroblasts; where PAPP-A was detected and sequenced through mass spectroscopy and identified as the protein causing IGFBP-4 proteolysis [2]. It has since been found that during pregnancy, the placental syncytiotrophoblasts synthesize and secrete PAPP-A into the plasma and maintain its high levels (>10,000 fold in humans [3]) throughout gestation [4,5]. Subsequently, in addition to conditions not related to pregnancy, PAPP-A has been discovered to be expressed during injury and repair responses for wound healing, cardiovascular and developmental diseases, and several malignancies, albeit in smaller concentrations than the placenta (Figure 1) [5,6]. PAPP-A is a metalloproteinase with IGFBP-4 proteolytic activity now demonstrated in breast, ovarian, lung, smooth muscle, bone, kidney, thymus, adipose, cardiovascular and immune cells; and its deletion has been implicated in promoting longevity and reducing tumor burden [7,8,9,10,11]. PAPP-A cleaves three of the six IGFBPs known, such as IGFBP-4, leading to increased bioavailability of IGFs, which in turn mediates cell proliferation, migration and survival [12]. High quantities of IGFs and the enhanced type 1 insulin-like growth factor receptor (IGF-IR) signaling pathways are heavily implicated in tumor initiation and progression [6,13,14]. Indeed, the link between PAPP-A and breast cancer (BC) was reported even prior to the discovery of PAPP-A function and its involvement in the IGF system; wherein independent of estrogen receptor (ER) status in stage I BC, PAPP-A was found to be a clinically significant predictor of early recurrence [15]. It has been reported that PAPP-A is overexpressed in more than 70% of BC [16]. This review will cover the role of PAPP-A in BC, focusing on the aggressive subtype of triple negative breast cancer (TNBC), with specific emphasis on the functional regulation of PAPP-A that causes TNBC progression and metastasis, its relation to the immune system, as well as its clinical relevance.
One in seven women are at the risk of BC in their lifetime and it is one of the leading causes of cancer-related mortality and morbidity in women [17]. Globally, it is the most commonly diagnosed cancer and is the fifth leading cause of death [18]. BC is sub-divided into four subtype categories according to the hormone receptor status: Luminal A (ER+, progesterone receptor (PR)+, and human epidermal receptor 2 (HER2)-); Luminal B (ER+ and/or PR+, HER2+); HER2 overexpressing (ER-, PR- and HER2+); and triple negative (ER-, PR- and HER2-) [19]. Among these subtypes, TNBC is the most aggressive, is frequently diagnosed at earlier ages and has median survival of <14 months once metastasized [20,21]. For non-TNBC subtypes, the 5-year survival rate is 99% and women diagnosed with this disease have several successful therapies such as hormonal and anti-HER2 targeted treatments [22]. However, TNBCs, comprising 15-20% of all BC cases, lack targeted therapy, have much poorer clinical outcomes, show increased local and distant recurrences, and have significantly lower survival rates [23,24]. TNBC phenotype is highly prevalent in women diagnosed with BC during pregnancy and is associated with a poor prognosis [19]. This aggressiveness of BC is reported to link with the highly elevated levels of PAPP-A throughout pregnancy. Increased PAPP-A expression in BC cells have been shown to correlate with tumor progression through epithelial-mesenchymal transition (EMT), and clinically tends towards worse overall survival [25]. However, while emerging studies make it clear that the IGFBP cleaving enzyme PAPP-A is heavily implicated in breast tumorigenesis, a few differential results report PAPP-A as having a more tumor suppressive role. For instance, PAPP-A silencing has led to an increase in BC aggressiveness; and a reduction in maternal serum concentrations of PAPP-A was found to correlate with elevated BC risk [26,27]. Such studies, explored in detail further in the review, provide insights that PAPP-A needs to be more thoroughly investigated. Tumor staging and cancer subtypes account for the conflicting observations in part. Nonetheless, detailed assessments are necessary to resolve the mechanisms and impacts of PAPP-A action in mammary tumorigenesis in order to fully realize its promising potential as a therapeutic and/or diagnostic target in TNBC.
Functionally, PAPP-A proteolyzes three IGFBPs, causing highly specific, regulated and efficient cleavage of IGFBPs-4, -5 and -2 at single sites. IGFBPs-4 and -2 cleavages require prior IGF-I or IGF-II binding before PAPP-A activity, and IGFBP-5 degradation is IGF independent [7]. PAPP-A interacts and binds with the cell surface glycosaminoglycans to become active for proteolysis of the IGFBPs [28]. Following proteolysis, the free IGFs bind to their respective receptors and initiate downstream signaling leading to enhanced proliferation, metastasis and survival [29,30]. Some regulators of PAPP-A have been reported, such as inhibition by proform of the eosinophil major basic protein (proMBP) and stanniocalcins STC1 and STC2 [31,32,33]. Furthermore, PAPP-A overexpression has recently been shown to significantly promote migration, invasion and EMT, especially in BC cells [25,34]. This review highlights the current research that lays the groundwork for evaluating PAPP-A as a biomarker, treatment modality or diagnostic molecule in BC – elucidating its involvement specifically in TNBC, highlighting the structure and function of PAPP-A, its regulators, and links to tumor progression. Importantly, we provide insights on its correlations with the immune system, its clinical applications for BC prognosis and potential in therapeutic targeting.
2. PAPP-A: Structure, Function, and Regulation
2.1. Structure
The gene for PAPP-A is located on chromosome 9q33.1 in humans, comprising 22 exons and 21 introns [35]. Highly conserved among species, PAPP-A shares more than 90% homology between the human and murine proteins [36]. Interestingly though, unlike humans, placenta of rats and mice do not produce PAPP-A [37]. The preprocessed protein is a 1627-residue polypeptide that undergoes maturation to yield a 1547 kDA protein [38]. In non-pregnancy related states and tissues, PAPP-A is a 400 kDA homodimer [39]. In pregnancy plasma, PAPP-A was initially thought to be a tetramer comprising approximately 200 kDA subunits [39]. However, it has since been shown to be a heterotetrametric complex made of two separate equimolar chains linked with disulfide bridges [40]. One chain is comprised of two PAPP-A subunits and the other chain is comprised of two disulfide bridged subunits of proMBP [39]. The PAPP- A/proMBP subunits are glycoproteins consisting of carbohydrates, glycosaminoglycans, zinc binding motifs, metal chelation columns and heparin binding sites [41]. Free proMBP is the precursor of eosinophil major basic protein (MBP) and is cleaved to release cytotoxic MBP during maturation of the eosinophil precursor cells [42]. However, there are no reports observed for proMBP of the PAPP-A/proMBP complex to go through a similar process. In the complexed form, proMBP functions as an inhibitor of the proteolytic activity of PAPP-A, reducing PAPP-A proteinase activity by more than 100-fold. Trace amounts of less than 1% uncomplexed PAPP-A are also present in pregnancy plasma and serum [40].
The PAPP-A subunit contains a 250-residue laminin G-like module at the N-terminus, the function of which is so far unknown [43]. It is followed by the elongated zinc binding protein motif consensus sequence HEXXHXXGXXH (His-482-His-492) of 350-residues [44]. Presence of this consensus motif places PAPP-A in the metzincin superfamily of metalloproteinases [45], where it is the founding member of the subgroup called pappalysins. A homologue of PAPP-A has been identified, termed PAPP-A2, that is also a pappalysin and is estimated to have similar functions in growth regulation as PAPP-A [46]. Three linear notch repeat (LNR) sequences are contained in the subunit, two of which span the proteolytic domain of PAPP-A close to the N-terminus and the third is towards the C terminus [47]. The proteolytic domain consists of roughly 350 residues and the Glu-483 is critical for catalytic activity [44]. Five complement control protein (CCP) modules are present close to the C terminus that contain glycosaminoglycan binding sequences to facilitate PAPP-A cell surface attachment [28,48].
During human pregnancy, serum PAPP-A/proMBP levels continually increase up to parturition [49]. The syncytiotrophoblasts are the main source of PAPP-A and extravillous X cells or cytotrophoblasts are the source of proMBP [4]. At term, up to 50 mg/L of serum PAPP-A circulates as PAPP-A/proMBP [50]. This covalent complex is established in the extracellular environment and accounts for nearly 99% of circulating PAPP-A. However, in the first trimester, up to approximately 30% can be in the form of uncomplexed PAPP-A [51]. Regardless of cancer, since the early 90s, aberrant levels of PAPP-A have been known to indicate unfavorable pregnancy outcomes such as Down’s syndrome, low birth weights, preeclampsia, growth retardations and other chromosomal abnormalities [52,53,54]. In males, immunohistochemistry (IHC) studies have revealed PAPP-A presence in Leydig cells, epididymis, testes and seminal vesicles as well as semen. Abnormally high levels of circulating PAPP-A levels correlated with prostate and testicular cancers, and levels decreased following orchidectomy or prostatectomy [55].
2.2. Function
The primary function of PAPP-A is proteolysis (Table 1), despite the presence of the metalloproteinase domain [30]. The first demonstration of PAPP-A proteolysis was IGFBP-4 cleavage [2]. Antibodies against PAPP-A blocked IGFBP-4 degradation in media conditioned with human fibroblasts. Pregnancy serum PAPP-A cleaved IGFBP-4 and secreted PAPP-A proteins were isolated from fibroblasts and osteoblasts [2]. Since then, PAPP-A mediated IGFBP-4 proteolysis has been seen in ovarian, lung, smooth muscle and endometrial stromal cells [56]. A distinctive feature of IGFBP-4 cleavage is the requirement of IGF-I or IGF-II to be present [57]. IGF-II is reported to be more efficient than IGF-I, and their binding to IGFBP-4 enhances sensitivity to PAPP-A. PAPP-A subsequently cleaves the IGFBP-4 protein at the sites Met-135 and Lys-136, thus releasing the bound IGFs and increasing their bioavailability for activating IGF receptor specific downstream signaling pathways [57]. Such PAPP-A activity can proceed in an autocrine or paracrine manner [28]. A small proportion of IGF-II independent IGFBP-4 cleavage by PAPP-A is also recorded, possibly at the same site, although proceeding at much lower rates of kinetic efficiency than in the presence of IGFs [57]. A comparison study utilizing a recombinant PAPP-A protein expressed from human embryonic kidney 293 T cells identified a single nucleotide polymorphism (SNP) in the PAPP-A allele (rs7020782; serine < tyrosine) that effects the proteolytic cleavage of IGFBP-4 [58]. The SNP with the serine variant was found to be significantly less efficient for cleaving IGFBP-4 as compared to the tyrosine variant. However, impact of the SNP on other IGFBP substrates such as IGFBP-5 and IGFBP-2 was not significant [58].
IGFBP-5 is another substrate for PAPP-A. Like IGFBP-4, proteolysis occurs roughly in the middle of the protein sequence (between Ser-143 and Lys-144 of IGFBP-5) to yield similar sized degraded fragments. But unlike IGFBP-4, PAPP-A mediated IGFBP-5 cleavage requires the presence of a 25-amino acid anchor peptide, and it is not only IGF independent but also found to be inhibited in the presence of IGFs by nearly three-fold [59,60,62]. PAPP-A mediates cleavage of IGFBP-2 as well [61]. IGFBP-2 is less susceptible than IGFBP-4 to PAPP-A, but its hydrolysis is also IGF-dependent, occurring between Gln-165 and Met-166. Similar to activity with IGFBP-4, IGFs are not considered as directly interacting with PAPP-A, rather, IGF binding to the IGFBPs-2 or -4 render conformational changes that enhance the degradation process [61]. Once IGFs are liberated from the IGF/IGFBP complexes, the IGFs bind to IGF receptors and cause initiation of multiple signaling pathways, leading to enhancement in cell proliferation and migration capacities, as well as reduced cell death and apoptosis [63]. This fosters a tumorigenic environment and promotes carcinogenesis of tissues, with PAPP-A therefore playing a critical role in establishment of malignancy.
2.3. Regulation
Several regulators of PAPP-A activity are emerging (Table 2). The first known inhibitor of PAPP-A is proMBP [40]. proMBP covalently binds PAPP-A and almost completely abrogates any proteolytic activity [64]. So far, all reported proteolytic function of PAPP-A is found in tissues where it is present free of proMBP [64]. As the levels of serum PAPP-A as PAPP-A/proMBP complex are increased by more than 10,000-fold during pregnancy, it stands to reason the strong inhibitory action of proMBP on PAPP-A protease activity has a major role in preventing PAPP-A mediated tumorigenesis during normal physiology. Stanniocalcins (STC) 1 and 2 are mammalian glycoprotein hormones that also potently inhibit PAPP-A, and both are thought to be proteinases specific to pappalysins [32,33,65]. STC1 is abundantly synthesized in multiple organs including heart, lung, liver, kidney, adrenal gland, ovary and prostate. Although a consensus role of STC1 is lacking, it is proposed to be associated with tumorigenesis as well as other physiological pathways of chondrogenesis and adipogenesis [32]. STC1 has been shown to interact with PAPP-A with strong affinity, but not with covalent binding, and inhibits PAPP-A proteolysis of IGFBP-4 and is antagonistic towards PAPP-A facilitated phosphorylation of IGF-IR [32]. The potent inhibitory effect of STC2 on PAPP-A activity is reported to occur through covalent binding of the cysteine-120 residue of STC2 with PAPP-A [33]. This leads to prevention of IGFBP-4 proteolysis by PAPP-A and the subsequent increased bioavailability of IGFs in tissues. Interestingly, mice overexpressing wild-type STC2 showed growth retardation, whereas mice overexpressing mutated STC2 that could not inhibit PAPP-A did not show any growth retardation [33]. Similarly, PAPP-A knockout mice are smaller in size compared to their wildtype littermates and have longer life span [14]. Such results suggest that PAPP-A inhibition can reduce cell growth in vivo.
Cyclic adenosine monophosphate (cAMP) is another reported regulator of PAPP-A expression. Cloning and sequencing of the PAPP-A cDNA revealed a long 5’ untranslated region (5’ UTR) that bears a cAMP inducible region, and PAPP-A protein synthesis could be induced in vitro in the presence of cAMP [66]. PAPP-A has also been reported to be progesterone dependent, where in vitro treatment with progesterone antagonists depleted PAPP-A secretion and subsequent progesterone treatment recovered PAPP-A expression in humans and monkeys [67,68]. Hormonal regulation of PAPP-A during the estrous cycle has been seen in a comparison study of human PAPP-A with the mouse ovarian PAPP-A cDNA [69]. Sequencing of PAPP-A from the two species demonstrated 88% match at the nucleotide level, 89% at the amino acid level and 93% similarity score for conservative amino acid substitutions. While PAPP-A transcripts in mice ovaries could not be detected to the same extent as human placenta and human fibroblasts, injection with pregnant mare serum gonadotropin (PMSG) led to transient increase in levels of PAPP-A transcripts in the mouse ovarian tissues at specific locations in granulosa and follicles. Subsequent treatment of the above mice with human chorionic gonadotropin (hCG) reintroduced sustained PAPP-A expression from ovulation to luteinization [69]. PMSG, as well as follicle stimulating hormone (FSH) also induce PAPP-A mRNA expression in granulocyte cells of rats in a spatiotemporal manner [70]. The oocyte growth factor, bone morphogenetic protein (BMP)-15, is reported to inhibit PAPP-A expression that has been enhanced by FSH [70]. Such PAPP-A expression is noted to occur at significantly differential patterns in rhesus monkeys [71].
Pro-inflammatory cytokines including tumor necrosis factor (TNF)-α, interleukin (IL)-1β and IL-6 have been seen as strong inducers of PAPP-A expression in human dermal fibroblasts and human coronary artery endothelial and smooth muscle cells; with IL-6 mediated PAPP-A upregulation seen only in the coronary artery smooth muscle cells [72]; and TNF-α, IL-1β, IL-4 and transforming growth factor-β (TGF-β) induced expression is seen in human osteoblasts as well [72]. In line with such effects of pro-inflammatory cytokines, a Kunitz-type protease inhibitor called Bikunin, responsible for suppression of pro-inflammatory cytokines, has been reported to suppress PAPP-A expression. Bikunin overexpression in human ovarian cancer cell line could decrease cancer invasion and metastasis, and acted as a suppressor of the PAPP-A gene with a nearly 9-fold reduction in PAPP-A RNA. In the study, knockdown of PAPP-A led to decrease in invasiveness of the ovarian cancer cells [73]. Resveratrol, another anti-inflammatory molecule, has also been shown to reduce cytokine-mediated PAPP-A expression in coronary artery smooth muscle cells, and interferon-γ (INF-γ), another cytokine with both pro and anti-inflammatory roles, has reduced PAPP-A expression in human fibroblasts [74]. PAPP-A thus appears to be involved more in promotion of inflammation than inflammation suppression responses. However, such a feature is not conclusive, as some of the cytokines have dual impacts on inflammation. In addition to cytokines, growth factors such as the epidermal growth factor (EGF) are reported to induce PAPP-A expression. Importantly for BC, in the TNBC cell line MDA-MB-468, treatment with EGF increased PAPP-A expression. The study highlighted the role of EGF as a potent activator of EMT and the resultant PAPP-A expression correlated strongly with increase in mesenchymal markers [25]. As discussed before, in a study in human osteoblasts, TGF-β also induced PAPP-A expression [72]. TGF-β is another well know inducer of EMT, potentiating further indications of the link between PAPP-A and EMT; explored in more details in the review in Section 3.2. The reported role of the tumor suppressor p53 as a regulator of PAPP-A in BC is also explored further in Section 3.1. The wild type protein could lead to PAPP-A suppression in TNBC cell line MDA-MB-157, but in human fibroblasts, p53 appeared to cause overexpression of PAPP-A [75,76,77].
Furthermore, microRNA (miRNA) mediated alteration of PAPP-A expression have also been reported. miRNA-214, a metastasis-linked mediator, is a potential suppressor of PAPP-A. Knockdown of miRNA-214 in non-small cell lung carcinoma cell lines upregulated PAPP-A expression, and overexpression of miRNA-214 decreased PAPP-A expression [78]. In human coronary artery smooth muscle cells, miR-490-3p has been demonstrated to target PAPP-A and inhibit its upregulation. The resultant downregulated PAPP-A led to decrease in IGFBP-4 proteolysis. Conversely, inhibition of miR-490-3p upregulated PAPP-A expression and increased its proteolytic activity on IGFBP-4 [79]. In vascular smooth muscle cells, miR-141 is reported to repress PAPP-A expression by directly inhibiting its translation. Interestingly, PAPP-A protein, and not PAPP-A mRNA, appeared to be significantly reduced following overexpression of miR-141 [80]. Importantly for BC, in BC cell lines MDA-MB-231 and MCF-7, miR-497-5p is found to be a negative regulator of PAPP-A. MiR-497-5p could significantly reduce PAPP-A expression in BC cell lines and serum of pregnancy associated BC patients as compared to non-cancer tissues [34].
To summarize, it is evident that not all regulators of PAPP-A have been discovered or validated in BC. It must be noted that, to the best of our knowledge, not many similar studies have been carried out to assess for PAPP-A regulation specifically in human mammary glands. The presence of species-based and tissue-based differences have clearly emerged, and bear consideration while assessing the biological role of PAPP-A.

Table 2.
Regulators of PAPP-A activity.
| Regulator | Role | Mode of PAPP-A interaction | Organism | Reference |
|---|---|---|---|---|
| proMBP | Inhibitor | Covalently binds PAPP-A to abrogate its proteolytic activity | Human serum, HEK 293T Cells | [40,64] |
| STC1 | Inhibitor | High affinity binding to PAPP-A rather than covalent binding | HEK 293T Cells | [32] |
| STC2 | Inhibitor | Covalent binding of PAPP-A through Cys-120 residue of STC2 | HEK 293T Cells, transgenic mice, mouse embryonic fibroblasts | [32,33] |
| cAMP | Inducer | cAMP inducible region in 5’ UTR of PAPP-A cDNA | Human placental choriocarcinoma cell line JAR cells | [66] |
| Progesterone antagonist (RU486) | Inhibitor | Inhibition of PAPP-A production rate; PAPP-A production recovered by addition of progesterone | Human trophoblastic and decidual explants, cynomolgus monkey | [67,68] |
| PMSG | Inducer | Transient increase in PAPP-A transcripts | Mouse ovary | [68] |
| hCG | Inducer | Sustained increase in PAPP-A expression after PMSG treatment | Mouse ovary | [68] |
| FSH | Inducer | Increased PAPP-A mRNA expression | Rat granulocytes | [70] |
| BMP-15 | Inhibitor | Reduced PAPP-A expression following FSH stimulation | Rat granulocytes | [70] |
| p53 | Inhibitor or inducer | PAPP-A suppression in TNBC; PAPP-A overexpression in human fibroblasts. | TNBC cell line MDA-MB-157; BJ/ET cell line |
[75,76,77]. |
| Bikunin | Inhibitor | Early suppression of PAPP-A mRNA in response to bikunin treatment | Ovarian cancer cell line HRA | [73] |
| TNF-α, IL-1β, IL-6, IL-4, TGF-β | Inducer | Upregulation of PAPP-A expression | TNF-α and IL-1β: human dermal fibroblasts and human coronary artery endothelial and smooth muscle cells; IL-6: coronary artery smooth muscle cells; TNF-α, IL-1β, IL-4, TGF-β: human osteoblasts |
[72] |
| EGF | Inducer | Upregulation of PAPP-A expression | TNBC cell lines | [25] |
| INF-γ | Inhibitor | Suppression of PAPP-A expression | Human fibroblasts | [74] |
| Resveratrol | Inhibitor | Reduction in cytokine-mediated PAPP-A expression | Coronary artery smooth muscle cells | [74] |
| miRNA-214 | Inhibitor | Targeted suppression of PAPP-A mRNA | NSCLC cell lines U-1810 or H23 | [78] |
| miR-497-5p | Inhibitor | Negative regulator of PAPP-A mRNA | Pregnancy-associated BC tissues and serum, normal breast tissues, BC cell lines MDA-MB-231 and MCF-7 | [34] |
| miR-490-3p | Inhibitor | Targeted suppression of PAPP-A expression | Human coronary artery smooth muscle cells | [79] |
| miR-141 | Inhibitor | Suppression of PAPP-A protein | Vascular smooth muscle cells | [80] |
3. PAPP-A in BC
PAPP-A is overexpressed in nearly all subtypes of malignant BC, with luminal B correlating with higher expression than luminal A subtypes [16]. A landmark study on formalin-fixed paraffin-embedded (FFPE) tumor tissues from 46 patients with invasive BC revealed positive PAPP-A expression in 98% of the samples, with significantly higher detection in luminal B than luminal A, and nearly complete absence of expression in non-tumor tissues [16]. PAPP-A is considered to have oncogenic activity in BC due to its impact on proteins of the IGFBP family (Table 3). With some exceptions, the binding proteins are regarded as generally tumor suppressive due to their inhibition of the mitogenic IGFs [81]. Numerous studies report increased IGFBPs lead to reduced aggressiveness of various cancers such as those of bladder, melanoma, lung and gastric in addition to BC. For instance, in bladder cancer, transcriptomic analysis of 200 patients and five normal samples revealed IGFBP-5 as a tumor sensitivity predictor of anti-IGF therapy that inversely corelated with IGF phosphorylation pathways [82]. In melanoma, an analysis of 54 melanoma cell lines by whole genome microarray expression profiling revealed upregulation of IGF-receptors (IGF-IR and IGF-IIR) and IGF-R substrates (IRS1 and IRS2) [83]. IGFBP-4 and-6 were also seen to be differentially expressed, with higher levels corresponding to more mesenchymal phenotypes. In the same study, 47 patient tumor samples were assessed, and PAPP-A expression was noted in 87% of metastatic tumor cases [83]. In melanoma cell lines and xenograft models, overexpression of IGFPB-5 also led to significant inhibition of malignancy through suppression of EMT, reduction of IGF signaling and decreased stem cell markers [84]. IGFBP-5 has been shown to promote p53 upregulation and resultant tumor suppression in gastric cancers. In the mammary gland, the IGFBP/IGF axis is heavily implicated in development and physiology of the breast; with its disruption playing a causal role in breast tumorigenesis [85,86]. Out of the six IGFBPs known, IGFBP-5 is of particular relevance in BC [87,88]. For instance, SNPs of IGFBP-5 is linked to increased risk of mammary tumors [89]. The multifunctional role of IGFBP-5 in apoptosis, migration, growth, cellular interactions and trafficking has been reviewed to reveal significant involvement in BC [87]. At present, proteolytic modulation of the IGFBP-5, -4 and -2 axis forms the basis of the known functions of PAPP-A activity in tumorigenesis (Figure 2). In the following sections, we provide a comprehensive overview focusing on BC, especially the aggressive form of TNBC, with respect to the interplay between PAPP-A and the IGFBPs, where its role in promoting metastasis through EMT, impact on the window of susceptibility (WOS) as well as its influence in pregnancy associated oncogenesis is examined. PAPP-A mediated immunomodulation is emerging. However, while its immune relationship specifically in BC has not yet been studied, promising results from other cancers provide insights for BC cases. The clinical relevance of PAPP-A is subsequently reviewed, with several reports demonstrating it as a biomarker for disease progression, as well as a possible therapeutic target, and thus, the potential of targeting PAPP-A in providing an overall beneficial outcome is critically assessed and revealed (Figure 2).
3.1. PAPP-A in TNBC: Proteolysis of IGFBPs
Proteolysis of the IGFBPs by PAPP-A is a critical component of BC development and progression. The clinical correlation between BC cases and PAPP-A is demonstrated in case-control studies on individuals enrolled in a BC prevention program [90]. In the study, serum concentrations of total PAPP-A, IGF-I and IGFBP-4 were found to be possible indicators of BC prevalence in a normal population, with PAPP-A displaying remarkable differences between the control group and chemotherapy treated group [90]. Mechanistically, in breast tissue, the cleavage of IGFBP-4 by PAPP-A was revealed for the first time in bovine mammary tissues through in vitro immunoblotting assays. PAPP-A was identified as the protease which degrades IGFBP-4 to increase IGF-I availability in the bovine mammary fibroblast (BMF) cells, but not in bovine mammary epithelial (BME) cells, as none of the IGFBP regulatory components studied (IGF-I and IGF-II) could be detected in the BME cells [91]. Such variant expression could arise from differential regulation of IGFBPs in the bovine fibroblasts as compared to epithelial cells. In an orthotopic model of 4T1.2 BCs, it was seen that PAPP-A resistant IGFBP-4 retained their tumor suppressive role of sequestering IGFs even in the presence of PAPP-A, representing a novel avenue of IGF-blocking strategies for BC therapeutics [92]. This observation was further validated by utilizing a recombinant PAPP-A resistant IGFBP-4 in 4T1.2 cells to achieve reduction in cell migration and invasion through IGF-I suppression [93]. The recombinant IGFBP-4 could not be cleaved by PAPP-A and thus acted as an IGF-I reservoir to reduce bioavailability of the mitogen. Notably, direct intra-tumoral injection of the PAPP-A resistant IGFBP-4 decreased angiogenesis and lung metastasis in the 4T1.2 mice BC models [93].
The exceptionally high incidences of p53 mutations associated with aggressive BCs such as TNBC characterize one pathway leading to aberrant PAPP-A expression that can promote tumorigenic architecture in the breast. PAPP-A has been revealed to be the link between IGFBP-4 proteolysis and p53 defects in BC [75]. Skp2B is an F-box protein reported to be overexpressed in BC [94]. In transgenic mice with BC xenografts that overexpress Skp2B, a 4–5-fold increase in PAPP-A levels were seen in their breasts as compared to non-transgenic mice. This increase in PAPP-A correlated with reduced or loss in levels of IGFBP-4 during pregnancy and lactation in the mice mammary glands; indicating IGFBP-4 degradation. Furthermore, sequence analysis of the PAPP-A gene revealed putative p53 binding sites in intron 1 of the PAPP-A gene. Interestingly, overexpression of p53 in the p53-negative cell line MDA-MB-157 revealed that mutated p53 binds to the intron 1 site to activate PAPP-A transcription, whereas wild type p53 binding leads to PAPP-A suppression. Mutant p53 thus binds to activate PAPP-A which in turn cleaves IGFBP-4 to release IGFs that play a causal role in cancer etiology [75]. Such findings demonstrate the oncogenic activity of PAPP-A in BC that is mediated via the tumor suppressor p53. However, ChIP-seq analysis in other cell lines such as BJ/ET (human fibroblast foreskin cell line BJ immortalized by human telomerase reverse transcriptase) could not detect similar p53 binding sites in the PAPP-A gene. Instead, it revealed an overexpression in PAPP-A transcription by p53 mediated by other signaling pathways such as rat sarcoma (RAS) expression [76,77]. Differences in cell type behaviors and the multifunctional mode of actions of p53 and PAPP-A, both of which require further elucidation, are major factors in accounting for the paradoxical effects seen with PAPP-A. PAPP-A is predominantly reported to possess oncogenic capacity [6], however that is also a matter of contention. The contradictory role is illustrated in BC cell lines and human mammary epithelial cells in which PAPP-A was found to have a tumor suppressive role [26]. Mitotic studies of FFPE specimens from a cohort of invasive BC patients as well as several BC cell lines revealed that a high proportion of luminal, HER2 and basal-like cases had PAPP-A silenced. Normal breast had much reduced silencing of PAPP-A and loss of function correlated with increasing progression of malignancy and higher invasiveness of BC [26]. In humans, a population-based cohort study including more than 600,000 pregnancies revealed that lower maternal serum concentration of PAPP-A in the first trimester appeared to bestow a higher long term BC risk [27]. Prenatal concentrations of alpha-fetoprotein (AFP), hCG, unconjugated estriol (uE3), PAPP-A, and dimeric inhibin-A (DIA) were also assessed. Only hCG, AFP, or PAPP-A were seen to indicate a slight elevation in future risk of hormone-dependent cancers such as BC [27]. However, the strongest outcome of the study was an overall lack in the future cancer predictive capacity of any of the assessed prenatal serum proteins, highlighting the strong need for further research for clarifying the mechanisms underlying some of the counterintuitive results seen with PAPP-A.
The pathogenic role of PAPP-A with respect to other IGFBPs such as IGFBP-2 has been noted but the underlying biological mechanisms with all IGFBPs are currently unknown. In a clinical evaluation of 301 females with BC and 531 non-cancer healthy controls, high PAPP-A level was predictive of worse prognosis for recurrence free survival (RFS) [95]. Serum levels of PAPP-A and IGFBP-2 independently prognosticated RFS and overall survival (OS), but no substantial increase in predictive value was observed. However, baseline quantities of PAPP-A and IGFBP-2 had no significant differences between cancer and non-cancer individuals. Interestingly, elevation of both PAPP-A and IGFBP-2 in serum was seen; suggesting IGFBP-2 proteolysis by PAPP-A occurs at a slower rate than IGFBPs-4 or 5 [95].
Table 3.
Role of PAPP-A and IGFBP/IGF axis in BC.
| IGFBP/IGF | Role | PAPP-A expression | Organism | References |
|---|---|---|---|---|
| IGFBP-4 proteolysis | Leads to increase in IGF-I | Secreted along with IGF-I | Bovine mammary fibroblast cells | [91] |
| PAPP-A resistant IGFBP-4 | Leads to sequestering of IGF-I | Retained tumor suppression; decreased angiogenesis and lung metastasis | 4T1.2, orthotopic model of 4T1.2 BCs | [92,93] |
| IGFBP-4 proteolysis | Due to increased PAPP-A levels from Skp2B overexpression | Putative p53 binding sites in PAPP-A gene | During pregnancy and lactation in the mice mammary glands | [75] |
| IGFBP-4 | Co-expression with PAPP-A | Co-expression with IGFBP-4 | HCC70, MDA-MB-468 and MDA-MB-231 cells | [25] |
| IGF-IR | Co-expression with PAPP-A | Co-expression with IGF-IR | HCC70, MDA-MB-468, HCC1954 and MDA-MB-231 | [25] |
| IGF-IIR | Independent of PAPP-A expression | Expression with and without IGF-IIR | With PAPP-A: MDA-MB-468, HCC1954 and MDA-MB-231 Without PAPP-A: MCF-7, BT474, SKBR3, HCC1569, MDA-MB-453 |
[25] |
| IGFBP-5 proteolysis | Due to increased PAPP-A levels from increased collagen deposition | Higher in parous mice breast than in nulliparous mice | Transgenic mice overexpressing PAPP-A in the mammary gland and MCF-7 BC cells | [96,97] |
3.2. PAPP-A in TNBC: Role in Epithelial-Mesenchymal Transition (EMT)
Epithelial-Mesenchymal Transition (EMT) is a vital cellular reprogramming process adopted by cancer cells for enhancing motility and migration in order to aid metastasis [98,99]. PAPP-A/IGFBP/IGF axis is found to play a significant role in motility and EMT of TNBC. In a study evaluating PAPP-A mRNA expression through performing qRT-PCR assays in a panel of eight TNBC cell lines, high expression was reported in four cell lines (HCC70, MDA-MB-468, HCC1954 and MDA-MB-231) [25]. Of the cell lines expressing PAPP-A, three expressed IGFBP-4 (HCC70, MDA-MB-468 and MDA-MB-231), all four expressed IGF-IR, and eight cell lines expressed IGF-IIR independent of PAPP-A expression (MDA-MB-468, HCC1954 and MDA-MB-231, MCF-7, BT474, SKBR3, HCC1569, and MDA-MB-453). The role of PAPP-A/IGFBP axis in impacting the migratory ability of TNBC cells was subsequently revealed. Anti-PAPP-A antibodies were found to significantly reduce migration in MDA-MB-231 cells, as did anti-IGFPB-4 antibody. Co-culturing with anti-PAPP-A and anti-IGFBP-4 antibodies further reduced invasiveness; underscoring the pro-migratory impacts of PAPP-A and its involvement in EMT. The connection to EMT was validated and further explored by analyzing The Cancer Genome Atlas (TCGA) dataset of BC (n = 1,105), as well as RNA sequencing, where PAPP-A significantly linked with co-expression of several canonical mesenchymal markers. Another dataset of 51 human BC cell lines revealed increased PAPP-A expression in basal B subtypes (corresponding to highest mesenchymal expressions) as compared to the more epithelial subtypes; thus, highlighting the tendency of PAPP-A to present with aggressive mesenchymal phenotypes of BC such as TNBC [25].
In another study, high PAPP-A expression was seen in BC cell lines including TNBC (T47D, MCF-7, BT549, and MDA-MB-231 and MDA-MB-468) as compared to normal breast epithelial cells MCF-10A through qRT-PCR and western blot assays [34]. Overexpression of PAPP-A in the cell lines MDA-MB-231 and MCF-7 significantly increased cellular proliferation, higher cell numbers in S phase of the cell cycle, and increased wound healing, migration and invasive capacity. Overexpression also led to higher protein levels of mesenchymal markers N-cadherin and vimentin and reduced epithelial marker E-cadherin. On culturing the above cell lines with PAPP-A rich serum collected from pregnancy associated BC patient samples, similar results were observed as those from PAPP-A overexpression. Conversely, knock down of PAPP-A using PAPP-A targeting siRNAs in the cell lines BT549 and MDA-MB-468 reduced proliferation, S phase cell numbers, wound healing, migration and invasive capacity; as well as reduced N-cadherin and vimentin and increased E-cadherin. miRNA regulation of PAPP-A was investigated, with miR-497-5p appearing to be a negative regulator of PAPP-A. The tumor promoting role of PAPP-A was also seen in vivo, where injection of PAPP-A recombinant protein in mice xenograft models led to significantly increased lung metastases [34].
IHC studies on the mammary glands of PAPP-A overexpressing transgenic mice, at the time of involution or postpartum, has identified a significant upregulation in Snail nuclear expression [97]. Snail oncogene is widely recognized as a key driver of EMT, with its nuclear expression being associated with the more aggressive malignant phenotypes [100]. On corroborating in MCF-7 cells overexpressing PAPP-A, significantly higher levels of Snail were similarly recorded, along with significant increase in invasive capacity, as compared to the non-PAPP-A overexpressing cells. Interestingly, the presence of collagen in the culture media further enhanced Snail expression in the PAPP-A overexpressing cells, as well as increasing levels of phosphorylated discoidin domain receptor 2 (DDR2), a known mediator of cancer progression that is a member of the collagen activation pathway [97,101].
3.3. PAPP-A in BC: Role in the Window of Susceptibility (WOS)
The role of PAPP-A in WOS, i.e., time periods spanning childhood, adolescence and young adulthood for susceptibility to initiating events for long term BC, is emerging [102]. The mammary carcinoma susceptibility 5c (Mcs5c) locus on rat chromosome 5 shares homology with mice and humans [103]. PAPP-A regulation has been demonstrated in mammary glands of Mcs5c Wistar-Kyoto homozygous congenic rats and rat BC cell line LA7 and has been identified to be associated with the WOS leading to increased risk of BC development [104,105]. Using chromosome conformation capture, the Mcs5c locus was found to bear a temporal control element that physically interacts with the neighboring PAPP-A gene located over 517kb away in mammary epithelial cells. This interaction is genotype-independent and happens through an intriguing chromatin looping mechanism. A methylation interaction is also reported in addition to the looping, conversely occurring in a genotype dependent manner in vivo. Importantly, differential PAPP-A expressions were noted for two different WOS periods in the lifetime. Adolescent WOS was marked by the chromatin looping between Mcs5c locus and PAPP-A as well as significant methylation differences in the CpG regions which bear the PAPP-A looping fragment. In contrast, sexually immature WOS lacked the looping interactions and had no significant differences in the methylation patterns. The age-dependent changes of PAPP-A expression are of important note, indicating the significance of proteomic interactions occurring at specific developmental points; with decrease in PAPP-A levels during adolescence appearing to reduce susceptibility to BC [105].
3.4. PAPP-A in TNBC: Impact of Pregnancy
Pregnancy, a condition featuring highly elevated levels of PAPP-A, marks another state that is linked with a short-term increase in risk of BC, and pregnancy related BCs present with higher rates of TNBC [106,107]. In a population study of maternal and infant birth weights including more than 400 women, increased serum concentrations of PAPP-A and ratios of estriol/anti-estrogen alpha-fetoprotein were seen in females delivering heavier babies and was associated with a greater risk of BC [108]. Although in normal pregnancy, increased serum PAPP-A is mainly in the form of PAPP-A/proMBP complex, the elevated hormonal concentrations along with increase in bioavailable IGFs mediated by free PAPP-A levels presents the biological plausibility of PAPP-A creating a milieu favoring tumor development and growth.
The parallel between increased BC risk and PAPP-A expression during pregnancy is mechanistically linked. Involution of the breast, a key phenomenon in pregnancy and post-partum state, highly involves IGFBPs-4 and -5 and the altered breast architecture is rich in collagen [109,110,111,112]. Independent of the breast and/or pregnancy, PAPP-A is known to colocalize with newly synthesized collagen during wound healing of the skin [113]. Thus, the relationship between PAPP-A, collagen and BC is further striking. The oncogenic potential of PAPP-A was found to be pregnancy and collagen dependent in a study including transgenic mice overexpressing PAPP-A in the mammary gland and MCF-7 BC cells [96]. Collagen deposition significantly upregulated PAPP-A expression during involution which led to increased IGFBP-5 proteolysis. The resultant elevation in IGFs signaled additional collagen deposition. PAPP-A also upregulated the La ribonucleoprotein domain family member 6 (LARP6) which further drove up collagen deposition [97]. Consequently, an oncogenic feedback loop between collagen and IGF signaling pathways via aberrant PAPP-A expression has been found. The loop was absent in mammary glands of virgin mice as they did not show PAPP-A mediated IGFBP-5 cleavage, indicating PAPP-A becomes tumor inducing only during pregnancy [96,97]. IHC on tumor samples from 46 patients with BC revealed parous females (corresponding to 79%) had higher positive PAPP-A expression as compared to nulliparous females. Notably, this correlated with the parous group presenting with an increased incidence of TNBC as compared to the nulliparous group. Furthermore, multiple pregnancies increased the oncogenic capacity of PAPP-A [96]. This indicates the accumulation of PAPP-A associated factors following every pregnancy event – leading to increase in risk window of BC. On the other hand, prolonged breast-feeding reduced BC risk by increasing the accumulation of PAPP-A inhibitory glycoproteins from the ovary, STC1 and STC2 [96].
Studying the tumor-associated collagen signature (TACS) of mice overexpressing PAPP-A in the mammary gland revealed that postpartum breasts have higher collagen signatures than virgin breasts, with the collagen being anti-tumorigenic in nature [97]. Of note was a particular orientation of collagen called TACS-3 that is associated with more aggressive forms of BC [114,115,116]. TACS-3 levels reduced and gave way to less aggressive collagen forms (TACS-1 and -2) in postpartum involution in non-transgenic mice, however in PAPP-A overexpressing transgenic mice, TACS-3 levels did not show reduction in postpartum involution [97]. The overexpression of PAPP-A post-partum thus transforms the collagen during involution from tumor inhibiting to tumor promoting TACS-3. In normal human breasts, PAPP-A is periodically expressed rather than constitutively [97]. The study reports that in human postpartum BC driven by PAPP-A, IGF signaling and TACS-3 collagen formation is constitutive. Furthermore, both in vivo transgenic mice and in vitro cell line studies in MCF-7 cells showed that one of the impacts of PAPP-A overexpression is on the DDR2/Snail pathways. PAPP-A activated DDR2 phosphorylation and Snail expression through promoting collagen production in an IGFBP proteolytic dependent manner. As explored in Section 3.2 of this review, elevated phosphorylated DDR2 and Snail are known drivers of cancer progression and EMT; and experimentally correlated with significantly higher invasion and migration. In line with this, a CRISPR mediated deletion of DDR2 led to elimination of the invasion-promoting effects of PAPP-A. Furthermore, gene set expression analysis for screening human BC datasets revealed that BC patients showing positive PAPP-A, Snai1 and Col1A1 signatures presented with shorter OS and increased metastasis. Out of 13 significantly dysregulated pathways in the above genotype signature, EMT, extracellular matrix (ECM) architecture and collagen formation represented the most elevated pathways [97].
4. Immunological Relevance of PAPP-A in BC
The immune system plays an indispensable role in cancer progression and resistance to therapy, with immune evasion being a common feature of tumor survival and metastasis in cancers such as those of the breast [117,118,119]. It is a multi-step process that is largely linked to the tumor-immune microenvironment, a suppressive cytokine milieu and the diminished ability of the innate and adaptive immune systems to effectively detect and eliminate tumor cells [120,121]. These factors are key reasons why therapeutic strategies against cancers, especially TNBCs, remain troublesome. Increasingly, immunotherapy is becoming one of the most clinically relevant strategies for effective treatment in BC [122,123,124,125]. In the neonatal setting, PAPP-A maintains immune homeostasis. However, in oncogenesis, PAPP-A is associated with tumor initiation, migration and invasion – pathways that comprise the molecular and cellular events of EMT, and this reprogramming process is well known to be strongly related to immune evasion [8,16,34,43,83,126,127]. Importantly, PAPP-A levels have been reported to be directly elevated during EMT [128,129]. Nevertheless, to the best of our knowledge, very little is known about the overlap between cancer, PAPP-A and immune evasion, with the link being unreported in BC. An initial correlation has so far been shown in Ewing sarcoma, followed by liver cancer, though the mechanisms remain unidentified.
As PAPP-A expression is high in Ewing sarcoma, a study investigating the profile of T cell receptor (TCR) transgenic T cells against PAPP-A revealed enhanced T cell targeting in A673 Ewing sarcoma cells and mice xenografts [126]. The study showed that the isolated T cell clone PAPP-A-2G6, which recognizes a specific PAPP-A peptide, was able to target A673 cells, and further, PAPP-A-specific T cells also lysed the cancer cells [126]. Functionally, the PAPP-A specific T cells harbored both a central memory and effector memory phenotype. Central memory phenotype facilitates T cell homing to secondary lymphoid organs and effector memory harbors a more cytolytic phenotype [130]. The findings were validated in vivo where immune deficient Rag2-/- γc-/- mice on a BALB/c background, inoculated with A673 tumor cells to form xenografts, were injected with a combination of PAPP-A-2G6 TCR transgenic T cells and CD8+ depleted peripheral blood mononuclear cells (PBMC), along with relevant controls [126]. The results indicated an enhanced tumor elimination process, lower tumor burden, greater accumulation of infiltrating CD8+ T cells in the tumor, while paired with no adverse effects in mice given PAPP-A-2G6 TCR transgenic T cells. Notably, IHC showed that PAPP-A expression in the A673 xenografts was higher compared to adjacent normal murine tissue, determining a correlation between PAPP-A expression, PAPP-A-2G6 TCR transgenic T cells and CD8+ T cell infiltration [126].
An RNA-seq study in Ewing sarcoma cell lines with PAPPA knockout demonstrated the induction of immune related genes and the downregulation of reactive oxygen species, DNA repair and the endoplasmic reticulum unfolded protein response [8]. The immune related genes included those that are part of the complement system, allograft rejection, inflammatory response, acute-phase response, tumor necrosis factor alpha signaling, and interferon responses [8]. Moreover, from this data set, important antigen processing/presentation pathway genes such as peptide loading complex TAP-1/TAP-2/TAPBP, the proteasome components low molecular mass polypeptides 2 and 7 (LMP2 and LMP7), proteasome activator complex subunits (PSME) 1 and 2, and the endoplasmic reticulum aminopeptidase 1 (ERAP1) were also found to be upregulated in PAPP-A knockout cells. Downregulation of antigen processing/presentation pathways are yet another way tumor cells evade detection by the immune system [126]. Of particular interest, genes associated with the subunits of major histocompatibility complex (MHC) class I molecules (beta2-microglobulin and classical MHC class I human leukocyte antigens A, B and C) were enriched, indicating that PAPP-A could potentially impact antigen processing and presentation machinery and the adaptive immunity as MHC molecules are heavily involved in the adaptive arm of the immune system [131]. MHC class I molecules are expressed on all nucleated cells and are essential for presenting foreign peptide antigens to activate cytotoxic T cells function to directly target and kill tumor cells [131,132]. Further to this, in the absence of PAPP-A (EW8 cell line with PAPP-A knockout), allogenic T cells had enhanced killing capacity, supporting the notation that PAPP-A may support tumor progression via depleting the cytotoxic functions of T cells [8]. The use of such TCR transgenic approach targeting PAPP-A is thus suggestive of a potentially beneficial route of novel therapeutics in BC.
PAPP-A knockout profiles are not unique to Ewing sarcoma and may represent a general feature of cancer physiology. The downregulation of immune related genes reported by Heitzeneder et al. was also seen in livers of BL\6 PAPP-A overexpressed mice [133]. TCGA data was analyzed from a cohort of 361 individuals with hepatocellular carcinomas in order to determine the link between PAPP-A signature and immune evasion in humans. Consistent with the general theme of PAPP-A expression and the downregulation of immune profiles, there was a strong association of PAPP-A signature and a subgroup of hepatocellular carcinomas with an exhausted immune phenotype [133]. Both males and females were included in the cohort analyzed indicating that the PAPP-A signature is independent of gender. Overall, while the mechanistic basis remains uncertain, these studies suggest PAPP-A may alter the expression profile and phenotype of immune-related pathways leading to immune evasion.
5. Clinical Relevance of PAPP-A in TNBC
PAPP-A is an important component of regulating IGF availability through the IGFBP receptor axis; with several of the IGFs and IGF receptors acting as clinical biomarkers [134,135,136,137]. The potential of PAPP-A itself as a clinical marker is gradually emerging as highly promising, and is of particular focus in this review (Table 4). The co-incidence of PAPP-A with BC cases was revealed to be of clinical relevance initially in predicting recurrence in stage II BC patients [138]. In 30 cases treated with low or standard chemotherapy doses, immunostaining and assessment for 25 clinicopathological features revealed that 82% of the cases with early recurrence were PAPP-A positive. Subsequently, 40 cases with stage I ER negative BC were assessed for a range of 33 clinicopathological features including PAPP-A expression [139]. PAPP-A staining (along with necrosis, nuclear atypia, and mitoses) significantly correlated with tumor recurrence in pairwise correlations. PAPP-A also emerged as a significant independent predictor in 56% of the recurrent patients. In combination with extensive necrosis as the only other significant independent predictor found in the study, PAPP-A positivity was seen in 81% of the recurrent cases [139]. The study was extended to analyse 30 ER positive cases at stage I, assessed for a range of 25 clinicopathological features in addition to PAPP-A expression [15]. On immunostaining patient samples, PAPP-A positivity again emerged as significantly correlating with both ER positive (30%) or negative (40%) cases, as well as a significant independent predictor of early recurrence in ER positive (13.3%) or negative (20%) cases. PAPP-A thus possessed recurrence predictive capacity independent of ER status. Furthermore, comparison frequencies revealed no correlation of PAPP-A recurrence predictive capacity between ER or progesterone receptor (PR) status [15].
An intensive study comparison has been carried out for several IGFs, IGFBPs and PAPP-A levels between 301 patients (with early BC treated by surgery, with and without adjuvant treatments) and 531 non-cancer individuals as controls [95]. For some treatment regimens, statistical significance was seen in IGF-I, IGFBP-3 and PAPP-A between the cancer and non-cancer individuals, but biological relevance was reported as low. On evaluating RFS and OS of the BC groups, elevation in circulating IGFBP-2 and PAPP-A quantities in the serum, determined at start of treatment, was found to be independently associated with worse prognosis in females with long-term follow up subsequent to treatment. Interestingly, in contrast to previous studies with PAPP-A tissue staining, the study concluded serum PAPP-A as independently prognostic for RFS and OS in the long-term, but not in early recurrence of BC [95]. While such contrasting observations could arise from low sample sizes that are limitations of the studies, it is possible that the biological role of PAPP-A may be varied and differential in both cellular and temporal contexts of BC, further standing to reason serum, tissue and stage dependent correlations with PAPP-A.
The IGFBP-4/ IGF-I/PAPP-A axis along with follistatin-like (FSTL)-3 has been shown to be surrogate markers for BC in a study involving 100 females with benign tumors, 145 females with malignant tumors (including treatment naïve and chemotherapy treated) and 100 disease-free control individuals [90]. In adjusted comparisons, serum PAPP-A (along with FSTL-3, IGFBP-4, and IGF-I) was reported to have substantial elevation across groups between treated and non-treated samples, but statistical significance was not observed. Positive PAPP-A association was also seen with serum activin A, serum activin B, total IGFBP-4, and a correlation with total IGF-I. Interestingly, negative PAPP-A association was seen repeatedly with total cholesterol and triglycerides. Importantly, PAPP-A (as well as total IGF-I, total and intact IGFBP-4) was shown to be a predictor of the presence of malignancy and total IGFBP-4 independently positively corelated with PAPP-A [90].
The immunoreactivity of PAPP-A and its association with clinicopathological characteristics in BC has been evaluated in a study cohort of 45 BC patients [25]. IHC on tissue microarrays (TMA) of BC samples, comprising 80% cases as TNBC, revealed 57% as positive for cytoplasmic and membranous PAPP-A expression; with majority of PAPP-A expression seen for tumor tissues corresponding to T2 staging. Furthermore, elevated PAPP-A expression strongly tended towards high-grade tumors and correlated with involvement of lymph node status. While PAPP-A expression did not correlate with OS in the limited sample size, it strongly trended with worse prognosis (median survival of 25 months) as compared with PAPP-A negative expression (median survival of 69 months). Importantly, increasing the sample size to include nine datasets revealed elevated PAPP-A expression to link with high risk of disease recurrence, and significantly correlated with poor OS in grade 3 BC as compared to grades 1 or 2 [25].
In Section 3.4, we review the association of collagen with PAPP-A in promoting aggressive BC. In line with the results discussed, the study analyzing the link between PAPP-A, collagen, and Snail used the gene expression panel (PAPP-A/SNAI1/COL1A1) to assess clinical outcomes of patients from a dataset of 327 cases with primary BC. In the survival analysis, the population scoring high in the panel significantly associated with distant metastases as compared to the low scoring group [97]. While majority of studies have found PAPP-A expression or elevation to coincide with BC progression, some contradictions have been reported. A study investigating epigenetic silencing of FFPE cancer and non-cancer tissues found that PAPP-A is strongly silenced through promoter hypermethylation in 46% (80/173 cases) of invasive BC. The PAPP-A promoter was non-methylated in 90% (27/30 cases) of normal mammary tissues [26], indicating PAPP-A silencing to be associated with oncogenesis. In another report, the results from a population-based cohort study that looked at 677,247 pregnancies to assess long-term risk of cancer in females found lower levels of serum PAPP-A in the first trimester to correlate with greater risk of BC in the long-term [27]. However, the risk no longer appeared significant after adjusting for covariates. Greater risk was also seen with abnormally reduced PAPP-A levels in parous females as compared to nulliparous [27]. As is evident, further research on PAPP-A is required to tease out the underpinnings in order to reach a unified consensus. Nonetheless, utilizing PAPP-A in the clinical setting for BC exists with great promise. Importantly for TNBC that has limited therapeutic options, the results with PAPP-A elucidate its potential that may provide novel, improved or adjuvant strategies to improve outcomes.
6. Conclusion
TNBC represents the most aggressive form of BC with the least favourable outcomes, are naturally recurrent, have poor prognosis and there is a steady increase in incidence rates [23]. Unlike other BC subtypes, dearth of efficient and optimal therapeutic options remain a major cause behind the dismal survival rates (approximately 10 months) seen in patients [23]. PAPP-A represents a promising clinical strategy for utilizing as a treatment modality and/or biomarker that ought to be exploited to address such critical unmet needs. Currently, the most evident method of PAPP-A mediated TNBC therapy appears to involve the IGFBP/IGF axis. Direct inhibition of IGF-I receptor utilizing antibodies have shown significant benefits only in early phase clinical trials, with a lack of similar efficiency seen in phase II–III trials due to insufficient specificity [140,141]. Indirect blocking of the IGF receptors, utilizing therapies targeting PAPP-A, may thus be conceptualized as more selective or tissue specific than targeting IGF receptors. The role of PAPP-A in addition to proteolysis is as of yet unknown, but cannot be ruled out. Furthermore, it is clear that PAPP-A is heavily involved in p53 mediated pathways, collagen deposition, EMT and ECM tissue remodeling of BCs including TNBC – representing a clear benefit of PAPP-A targeting to impact such tumorigenic pathways. Majority of research till date presents PAPP-A as a cancer promoting antigen. However, the possibility of PAPP-A correlating with tumor suppression has been reported, highlighting the need for further research. Immunotherapy has been of great benefit in several cancers that have limited therapeutic targets such as TNBC, and PAPP-A expression has been shown to influence immune cells and immune related pathways. As TNBC is a difficult cancer to target due to its lack of ER, PR and HER2 expression, shedding light on the relationship and mechanisms between PAPP-A and the immune system may potentially lead to successful immunotherapeutic targets. In addition, quantifying PAPP-A levels can potentially contribute to positive patient outcomes by influencing patient selection and directing more accurate therapy regimens. In cases with aberrantly overexpressed PAPP-A, intervention at the prevention stage may also be a viable option. Through more focused research, a comprehensive and integrated understanding of the PAPP-A mediated impacts in TNBC is thus needed, representing a critical step forward in providing favorable outcomes and reliable clinical benefits.
7. List of Abbreviations
| 5’ UTR | 5’ untranslated region |
| AFP | Alpha-fetoprotein |
| BC | Breast cancer |
| BME | Bovine mammary epithelial |
| BMF | Bovine mammary fibroblast |
| BMP | Bone morphogenetic protein |
| cAMP | Cyclic adenosine monophosphate |
| CCP | Complement control protein |
| ChIP-seq | Chromatin immunoprecipitation followed by sequencing |
| DDR2 | Discoidin domain receptor 2 |
| DIA | Dimeric inhibin-A |
| ECM | Extracellular matrix |
| EGF | Epidermal growth factor |
| EMT | Epithelial-mesenchymal transition |
| ER | Estrogen receptor |
| ERAP1 | Endoplasmic reticulum aminopeptidase 1 |
| FFPE | Formalin-fixed paraffin-embedded |
| FSH | Follicle stimulating hormone |
| FSTL | Follistatin-like |
| hCG | Human chorionic gonadotropin |
| HER2 | Human epidermal receptor 2 |
| IGF | Insulin-like growth factor |
| IGFBP | Insulin-like growth factor dependent insulin-like growth factor-binding proteins |
| IGF-IR | Enhanced type I insulin-like growth factor receptor |
| IGF-IIR | Enhanced type II insulin-like growth factor receptor |
| IHC | Immunohistochemistry |
| IL-1β | Interleukin 1 beta |
| LARP6 | La ribonucleoprotein domain family member 6 |
| LMP | Low molecular mass polypeptides |
| LNR | Linear notch repeat |
| Mcs5c | Mammary carcinoma susceptibility 5c |
| MHC | Major histocompatibility complex |
| miR | microRNA |
| OS | Overall survival |
| PAPP-A | Pregnancy associated plasma protein-A |
| PBMC | Peripheral blood mononuclear cells |
| PMSG | Pregnant mare serum gonadotropin |
| PR | Progesterone receptor |
| proMBP | Proform of the eosinophil major basic protein |
| PSME | Proteasome activator complex subunits |
| qRT-PCR | Real-time quantitative reverse transcription pcr |
| RAS | Rat sarcoma |
| RFS | Recurrence free survival |
| SNP | Single nucleotide polymorphism |
| STC | Stanniocalcins |
| TACS | Tumor-associated collagen signature |
| TCGA | The cancer genome atlas |
| TCR | T cell receptor |
| TGF-β | Transforming growth factor beta |
| TMA | Tissue microarrays |
| TNBC | Triple negative breast cancer |
| TNF-α | Tumor necrosis factor alpha |
| uE3 | Unconjugated estriol |
| WOS | The Window of Susceptibility |
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors have read and agreed to the submitted version of the manuscript.
Availability of data and materials
Not applicable.
Competing Interests
The authors declare there are no competing interests.
Author Contributions
Data curation, A.P.; writing—original draft preparation, A.P.; writing—original draft preparation, F.A.; writing— review and editing, S.R.R., R.S., G.K., P.P. and A.J.; conceptualization, P.P., A.J.; writing—original draft preparation, P.P., A.J. All authors read and approved the final manuscript.
Funding
This manuscript was supported by the Fiona Elsey Cancer Research Institute and the Perpetual’s IMPACT Philanthropy Program.
Acknowledgements
The authors acknowledge support provided by the team at Fiona Elsey Cancer Research Institute towards this research.
References
- Lin TM, Galbert SP, Kiefer D, Spellacy WN, Gall S. Characterization of four human pregnancy-associated plasma proteins. Am J Obstet Gynecol. 1974;118(2):223-36. [CrossRef]
- Lawrence, J.B.; Oxvig, C.; Overgaard, M.T.; Sottrup-Jensen, L.; Gleich, G.J.; Hays, L.G.; Yates, J.R., 3rd; Conover, C.A. The insulin-like growth factor (IGF)-dependent IGF binding protein-4 protease secreted by human fibroblasts is pregnancy-associated plasma protein-A. Proc Natl Acad Sci U S A 1999, 96, 3149–3153. [Google Scholar] [CrossRef]
- Qin, Q.P.; Christiansen, M.; Oxvig, C.; Pettersson, K.; Sottrup-Jensen, L.; Koch, C.; Nørgaard-Pedersen, B. Double-monoclonal immunofluorometric assays for pregnancy-associated plasma protein A/proeosinophil major basic protein (PAPP-A/proMBP) complex in first-trimester maternal serum screening for Down syndrome. Clin Chem 1997, 43, 2323–2332. [Google Scholar] [CrossRef]
- Bonno, M.; Oxvig, C.; Kephart, G.M.; Wagner, J.M.; Kristensen, T.; Sottrup-Jensen, L.; Gleich, G.J. Localization of pregnancy-associated plasma protein-A and colocalization of pregnancy-associated plasma protein-A messenger ribonucleic acid and eosinophil granule major basic protein messenger ribonucleic acid in placenta. Lab Invest 1994, 71, 560–566. [Google Scholar]
- Overgaard, M.T.; Oxvig, C.; Christiansen, M.; Lawrence, J.B.; Conover, C.A.; Gleich, G.J.; Sottrup-Jensen, L.; Haaning, J. Messenger ribonucleic acid levels of pregnancy-associated plasma protein-A and the proform of eosinophil major basic protein: expression in human reproductive and nonreproductive tissues. Biol Reprod 1999, 61, 1083–1089. [Google Scholar] [CrossRef]
- Guo, Y.; Bao, Y.; Guo, D.; Yang, W. Pregnancy-associated plasma protein a in cancer: expression, oncogenic functions and regulation. Am J Cancer Res 2018, 8, 955–963. [Google Scholar]
- Jenkins, E.C.; Brown, S.O.; Germain, D. The Multi-Faced Role of PAPP-A in Post-Partum Breast Cancer: IGF-Signaling is Only the Beginning. J Mammary Gland Biol Neoplasia 2020, 25, 181–189. [Google Scholar] [CrossRef]
- Heitzeneder, S.; Sotillo, E.; Shern, J.F.; Sindiri, S.; Xu, P.; Jones, R.; Pollak, M.; Noer, P.R.; Lorette, J.; Fazli, L.; et al. Pregnancy-Associated Plasma Protein-A (PAPP-A) in Ewing Sarcoma: Role in Tumor Growth and Immune Evasion. J Natl Cancer Inst 2019, 111, 970–982. [Google Scholar] [CrossRef]
- D'Elia, P.; Ionta, V.; Chimenti, I.; Angelini, F.; Miraldi, F.; Pala, A.; Messina, E.; Giacomello, A. Analysis of pregnancy-associated plasma protein A production in human adult cardiac progenitor cells. Biomed Res Int 2013, 2013, 190178. [Google Scholar] [CrossRef]
- Hjortebjerg, R. IGFBP-4 and PAPP-A in normal physiology and disease. Growth Horm IGF Res 2018, 41, 7–22. [Google Scholar] [CrossRef]
- Conover, C.A.; Bale, L.K. Loss of pregnancy-associated plasma protein A extends lifespan in mice. Aging Cell 2007, 6, 727–729. [Google Scholar] [CrossRef]
- Oxvig, C. The role of PAPP-A in the IGF system: location, location, location. J Cell Commun Signal 2015, 9, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Bowers, L.W.; Rossi, E.L.; O'Flanagan, C.H.; deGraffenried, L.A.; Hursting, S.D. The Role of the Insulin/IGF System in Cancer: Lessons Learned from Clinical Trials and the Energy Balance-Cancer Link. Front Endocrinol (Lausanne) 2015, 6, 77. [Google Scholar] [CrossRef]
- Conover, C.A.; Oxvig, C. PAPP-A and cancer. J Mol Endocrinol 2018, 61, T1–t10. [Google Scholar] [CrossRef] [PubMed]
- Kuhajda, F.P.; Eggleston, J.C. Pregnancy-associated plasma protein A. A clinically significant predictor of early recurrence in stage I breast carcinoma is independent of estrogen receptor status. Am J Pathol 1985, 121, 342–348. [Google Scholar] [PubMed]
- Mansfield, A.S.; Visscher, D.W.; Hart, S.N.; Wang, C.; Goetz, M.P.; Oxvig, C.; Conover, C.A. Pregnancy-associated plasma protein-A expression in human breast cancer. Growth Horm IGF Res 2014, 24, 264–267. [Google Scholar] [CrossRef]
- Gadaleta, E.; Thorn, G.J.; Ross-Adams, H.; Jones, L.J.; Chelala, C. Field cancerization in breast cancer. J Pathol 2022, 257, 561–574. [Google Scholar] [CrossRef]
- Azamjah, N.; Soltan-Zadeh, Y.; Zayeri, F. Global Trend of Breast Cancer Mortality Rate: A 25-Year Study. Asian Pac J Cancer Prev 2019, 20, 2015–2020. [Google Scholar] [CrossRef]
- Poddar, A.; Rao, S.R.; Prithviraj, P.; Kannourakis, G.; Jayachandran, A. Crosstalk between Immune Checkpoint Modulators, Metabolic Reprogramming and Cellular Plasticity in Triple-Negative Breast Cancer. Curr Oncol 2022, 29, 6847–6863. [Google Scholar] [CrossRef]
- Emens, L.A.; Cruz, C.; Eder, J.P.; Braiteh, F.; Chung, C.; Tolaney, S.M.; Kuter, I.; Nanda, R.; Cassier, P.A.; Delord, J.P.; et al. Long-term Clinical Outcomes and Biomarker Analyses of Atezolizumab Therapy for Patients With Metastatic Triple-Negative Breast Cancer: A Phase 1 Study. JAMA Oncol 2019, 5, 74–82. [Google Scholar] [CrossRef]
- Ensenyat-Mendez, M.; Llinas-Arias, P.; Orozco, J.I.J.; Iniguez-Munoz, S.; Salomon, M.P.; Sese, B.; DiNome, M.L.; Marzese, D.M. Current Triple-Negative Breast Cancer Subtypes: Dissecting the Most Aggressive Form of Breast Cancer. Front Oncol 2021, 11, 681476. [Google Scholar] [CrossRef]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. Jama 2019, 321, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Almansour, N.M. Triple-Negative Breast Cancer: A Brief Review About Epidemiology, Risk Factors, Signaling Pathways, Treatment and Role of Artificial Intelligence. Front Mol Biosci 2022, 9, 836417. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Zhu, C.; Wang, G.; Gu, J. Treatment for Triple-Negative Breast Cancer: An Umbrella Review of Meta-Analyses. Int J Gen Med 2022, 15, 5901–5914. [Google Scholar] [CrossRef] [PubMed]
- Prithviraj, P.; Anaka, M.; Thompson, E.W.; Sharma, R.; Walkiewicz, M.; Tutuka, C.S.A.; Behren, A.; Kannourakis, G.; Jayachandran, A. Aberrant pregnancy-associated plasma protein-A expression in breast cancers prognosticates clinical outcomes. Sci Rep 2020, 10, 13779. [Google Scholar] [CrossRef]
- Loddo, M.; Andryszkiewicz, J.; Rodriguez-Acebes, S.; Stoeber, K.; Jones, A.; Dafou, D.; Apostolidou, S.; Wollenschlaeger, A.; Widschwendter, M.; Sainsbury, R.; et al. Pregnancy-associated plasma protein A regulates mitosis and is epigenetically silenced in breast cancer. J Pathol 2014, 233, 344–356. [Google Scholar] [CrossRef]
- Park, A.L.; Huang, T.; Meschino, W.S.; Iqbal, J.; Ray, J.G. Prenatal Biochemical Screening and a Woman's Long-Term Risk of Cancer: A Population-Based Cohort Study. JNCI Cancer Spectr 2020, 4, pkz077. [Google Scholar] [CrossRef]
- Laursen, L.S.; Overgaard, M.T.; Weyer, K.; Boldt, H.B.; Ebbesen, P.; Christiansen, M.; Sottrup-Jensen, L.; Giudice, L.C.; Oxvig, C. Cell surface targeting of pregnancy-associated plasma protein A proteolytic activity. Reversible adhesion is mediated by two neighboring short consensus repeats. J Biol Chem 2002, 277, 47225–47234. [Google Scholar] [CrossRef]
- Brahmkhatri, V.P.; Prasanna, C.; Atreya, H.S. Insulin-like growth factor system in cancer: novel targeted therapies. Biomed Res Int 2015, 2015, 538019. [Google Scholar] [CrossRef]
- Kobberø, S.D.; Gajhede, M.; Mirza, O.A.; Kløverpris, S.; Kjær, T.R.; Mikkelsen, J.H.; Boesen, T.; Oxvig, C. Structure of the proteolytic enzyme PAPP-A with the endogenous inhibitor stanniocalcin-2 reveals its inhibitory mechanism. Nat Commun 2022, 13, 6084. [Google Scholar] [CrossRef]
- Glerup, S.; Kløverpris, S.; Laursen, L.S.; Dagnaes-Hansen, F.; Thiel, S.; Conover, C.A.; Oxvig, C. Cell surface detachment of pregnancy-associated plasma protein-A requires the formation of intermolecular proteinase-inhibitor disulfide bonds and glycosaminoglycan covalently bound to the inhibitor. J Biol Chem 2007, 282, 1769–1778. [Google Scholar] [CrossRef]
- Kløverpris, S.; Mikkelsen, J.H.; Pedersen, J.H.; Jepsen, M.R.; Laursen, L.S.; Petersen, S.V.; Oxvig, C. Stanniocalcin-1 Potently Inhibits the Proteolytic Activity of the Metalloproteinase Pregnancy-associated Plasma Protein-A. J Biol Chem 2015, 290, 21915–21924. [Google Scholar] [CrossRef] [PubMed]
- Jepsen, M.R.; Kløverpris, S.; Mikkelsen, J.H.; Pedersen, J.H.; Füchtbauer, E.M.; Laursen, L.S.; Oxvig, C. Stanniocalcin-2 inhibits mammalian growth by proteolytic inhibition of the insulin-like growth factor axis. J Biol Chem 2015, 290, 3430–3439. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, Y.; Li, L.; Nian, Y.; Chen, Y.; Shen, R.; Ma, X. Pregnancy-associated plasma protein-A (PAPPA) promotes breast cancer progression. Bioengineered 2022, 13, 291–307. [Google Scholar] [CrossRef]
- Silahtaroglu, A.N.; Tümer, Z.; Kristensen, T.; Sottrup-Jensen, L.; Tommerup, N. Assignment of the human gene for pregnancy-associated plasma protein A (PAPPA) to 9q33.1 by fluorescence in situ hybridization to mitotic and meiotic chromosomes. Cytogenet Cell Genet 1993, 62, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Sexton, C.; Byun, D.; Strong, D.D.; Baylink, D.J.; Mohan, S. Differential regulation of pregnancy associated plasma protein (PAPP)-A during pregnancy in human and mouse. Growth Horm IGF Res 2002, 12, 359–366. [Google Scholar] [CrossRef]
- Søe, R.; Overgaard, M.T.; Thomsen, A.R.; Laursen, L.S.; Olsen, I.M.; Sottrup-Jensen, L.; Haaning, J.; Giudice, L.C.; Conover, C.A.; Oxvig, C. Expression of recombinant murine pregnancy-associated plasma protein-A (PAPP-A) and a novel variant (PAPP-Ai) with differential proteolytic activity. Eur J Biochem 2002, 269, 2247–2256. [Google Scholar] [CrossRef]
- Bischof, P. Purification and characterization of pregnancy associated plasma protein A (PAPP-A). Arch Gynecol 1979, 227, 315–326. [Google Scholar] [CrossRef]
- Oxvig, C.; Sand, O.; Kristensen, T.; Gleich, G.J.; Sottrup-Jensen, L. Circulating human pregnancy-associated plasma protein-A is disulfide-bridged to the proform of eosinophil major basic protein. J Biol Chem 1993, 268, 12243–12246. [Google Scholar] [CrossRef]
- Overgaard, M.T.; Haaning, J.; Boldt, H.B.; Olsen, I.M.; Laursen, L.S.; Christiansen, M.; Gleich, G.J.; Sottrup-Jensen, L.; Conover, C.A.; Oxvig, C. Expression of recombinant human pregnancy-associated plasma protein-A and identification of the proform of eosinophil major basic protein as its physiological inhibitor. J Biol Chem 2000, 275, 31128–31133. [Google Scholar] [CrossRef]
- Oxvig, C.; Sand, O.; Kristensen, T.; Kristensen, L.; Sottrup-Jensen, L. Isolation and characterization of circulating complex between human pregnancy-associated plasma protein-A and proform of eosinophil major basic protein. Biochim Biophys Acta 1994, 1201, 415–423. [Google Scholar] [CrossRef]
- Weyer, K.; Glerup, S. Placental regulation of peptide hormone and growth factor activity by proMBP. Biol Reprod 2011, 84, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Boldt, H.B.; Glerup, S.; Overgaard, M.T.; Sottrup-Jensen, L.; Oxvig, C. Definition, expression, and characterization of a protein domain in the N-terminus of pregnancy-associated plasma protein-A distantly related to the family of laminin G-like modules. Protein Expr Purif 2006, 48, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Boldt, H.B.; Overgaard, M.T.; Laursen, L.S.; Weyer, K.; Sottrup-Jensen, L.; Oxvig, C. Mutational analysis of the proteolytic domain of pregnancy-associated plasma protein-A (PAPP-A): classification as a metzincin. Biochem J 2001, 358, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Stöcker, W.; Grams, F.; Baumann, U.; Reinemer, P.; Gomis-Rüth, F.X.; McKay, D.B.; Bode, W. The metzincins--topological and sequential relations between the astacins, adamalysins, serralysins, and matrixins (collagenases) define a superfamily of zinc-peptidases. Protein Sci 1995, 4, 823–840. [Google Scholar] [CrossRef] [PubMed]
- Barrios, V.; Chowen, J.A.; Martín-Rivada, Á.; Guerra-Cantera, S.; Pozo, J.; Yakar, S.; Rosenfeld, R.G.; Pérez-Jurado, L.A.; Suárez, J.; Argente, J. Pregnancy-Associated Plasma Protein (PAPP)-A2 in Physiology and Disease. Cells 2021, 10. [Google Scholar] [CrossRef]
- Boldt, H.B.; Kjaer-Sorensen, K.; Overgaard, M.T.; Weyer, K.; Poulsen, C.B.; Sottrup-Jensen, L.; Conover, C.A.; Giudice, L.C.; Oxvig, C. The Lin12-notch repeats of pregnancy-associated plasma protein-A bind calcium and determine its proteolytic specificity. J Biol Chem 2004, 279, 38525–38531. [Google Scholar] [CrossRef]
- Weyer, K.; Overgaard, M.T.; Laursen, L.S.; Nielsen, C.G.; Schmitz, A.; Christiansen, M.; Sottrup-Jensen, L.; Giudice, L.C.; Oxvig, C. Cell surface adhesion of pregnancy-associated plasma protein-A is mediated by four clusters of basic residues located in its third and fourth CCP module. Eur J Biochem 2004, 271, 1525–1535. [Google Scholar] [CrossRef]
- Leguy, M.C.; Brun, S.; Pidoux, G.; Salhi, H.; Choiset, A.; Menet, M.C.; Gil, S.; Tsatsaris, V.; Guibourdenche, J. Pattern of secretion of pregnancy-associated plasma protein-A (PAPP-A) during pregnancies complicated by fetal aneuploidy, in vivo and in vitro. Reprod Biol Endocrinol 2014, 12, 129. [Google Scholar] [CrossRef]
- Bischof, P.; DuBerg, S.; Herrmann, W.; Sizonenko, P.C. Pregnancy-associated plasma protein-A (PAPP-A) and hCG in early pregnancy. Br J Obstet Gynaecol 1981, 88, 973–975. [Google Scholar] [CrossRef]
- Gyrup, C.; Christiansen, M.; Oxvig, C. Quantification of proteolytically active pregnancy-associated plasma protein-A with an assay based on quenched fluorescence. Clin Chem 2007, 53, 947–954. [Google Scholar] [CrossRef]
- Shiefa, S.; Amargandhi, M.; Bhupendra, J.; Moulali, S.; Kristine, T. First Trimester Maternal Serum Screening Using Biochemical Markers PAPP-A and Free β-hCG for Down Syndrome, Patau Syndrome and Edward Syndrome. Indian J Clin Biochem 2013, 28, 3–12. [Google Scholar] [CrossRef]
- Antsaklis, P.; Fasoulakis, Z.; Theodora, M.; Diakosavvas, M.; Kontomanolis, E.N. Association of Low Maternal Pregnancy-associated Plasma Protein A with Adverse Perinatal Outcome. Cureus 2019, 11, e4912. [Google Scholar] [CrossRef] [PubMed]
- Kirkegaard, I.; Uldbjerg, N.; Oxvig, C. Biology of pregnancy-associated plasma protein-A in relation to prenatal diagnostics: an overview. Acta Obstet Gynecol Scand 2010, 89, 1118–1125. [Google Scholar] [CrossRef] [PubMed]
- Bischof, P.; Mégevand, M. Pregnancy-associated plasma protein-A concentrations in men with testicular and prostatic tumors. Arch Androl 1986, 16, 155–160. [Google Scholar] [CrossRef]
- Boldt, H.B.; Conover, C.A. Pregnancy-associated plasma protein-A (PAPP-A): a local regulator of IGF bioavailability through cleavage of IGFBPs. Growth Horm IGF Res 2007, 17, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Byun, D.; Mohan, S.; Kim, C.; Suh, K.; Yoo, M.; Lee, H.; Baylink, D.J.; Qin, X. Studies on human pregnancy-induced insulin-like growth factor (IGF)-binding protein-4 proteases in serum: determination of IGF-II dependency and localization of cleavage site. J Clin Endocrinol Metab 2000, 85, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Bøtkjær, J.A.; Noer, P.R.; Oxvig, C.; Andersen, C.Y. Author Correction: A common variant of the pregnancy-associated plasma protein-A (PAPPA) gene encodes a protein with reduced proteolytic activity towards IGF-binding proteins. Sci Rep 2019, 9, 17523. [Google Scholar] [CrossRef]
- Laursen, L.S.; Overgaard, M.T.; Søe, R.; Boldt, H.B.; Sottrup-Jensen, L.; Giudice, L.C.; Conover, C.A.; Oxvig, C. Pregnancy-associated plasma protein-A (PAPP-A) cleaves insulin-like growth factor binding protein (IGFBP)-5 independent of IGF: implications for the mechanism of IGFBP-4 proteolysis by PAPP-A. FEBS Lett 2001, 504, 36–40. [Google Scholar] [CrossRef]
- Gaidamauskas, E.; Gyrup, C.; Boldt, H.B.; Schack, V.R.; Overgaard, M.T.; Laursen, L.S.; Oxvig, C. IGF dependent modulation of IGF binding protein (IGFBP) proteolysis by pregnancy-associated plasma protein-A (PAPP-A): multiple PAPP-A-IGFBP interaction sites. Biochim Biophys Acta 2013, 1830, 2701–2709. [Google Scholar] [CrossRef]
- Monget, P.; Mazerbourg, S.; Delpuech, T.; Maurel, M.C.; Manière, S.; Zapf, J.; Lalmanach, G.; Oxvig, C.; Overgaard, M.T. Pregnancy-associated plasma protein-A is involved in insulin-like growth factor binding protein-2 (IGFBP-2) proteolytic degradation in bovine and porcine preovulatory follicles: identification of cleavage site and characterization of IGFBP-2 degradation. Biol Reprod 2003, 68, 77–86. [Google Scholar] [CrossRef]
- Judge, R.A.; Sridar, J.; Tunyasuvunakool, K.; Jain, R.; Wang, J.C.K.; Ouch, C.; Xu, J.; Mafi, A.; Nile, A.H.; Remarcik, C.; et al. Structure of the PAPP-A(BP5) complex reveals mechanism of substrate recognition. Nat Commun 2022, 13, 5500. [Google Scholar] [CrossRef]
- Grimberg, A. Mechanisms by which IGF-I may promote cancer. Cancer Biol Ther 2003, 2, 630–635. [Google Scholar] [CrossRef]
- Overgaard, M.T.; Glerup, S.; Boldt, H.B.; Rodacker, V.; Olsen, I.M.; Christiansen, M.; Sottrup-Jensen, L.; Giudice, L.C.; Oxvig, C. Inhibition of proteolysis by the proform of eosinophil major basic protein (proMBP) requires covalent binding to its target proteinase. FEBS Lett 2004, 560, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Oxvig, C.; Conover, C.A. The Stanniocalcin-PAPP-A-IGFBP-IGF Axis. J Clin Endocrinol Metab 2023, 108, 1624–1633. [Google Scholar] [CrossRef] [PubMed]
- Haaning, J.; Oxvig, C.; Overgaard, M.T.; Ebbesen, P.; Kristensen, T.; Sottrup-Jensen, L. Complete cDNA sequence of the preproform of human pregnancy-associated plasma protein-A. Evidence for expression in the brain and induction by cAMP. Eur J Biochem 1996, 237, 159–163. [Google Scholar] [CrossRef]
- Bischof, P.; Sizonenko, M.T.; Herrmann, W.L. Trophoblastic and decidual response to RU486: effects on human chorionic gonadotrophin, human placental lactogen, prolactin and pregnancy-associated plasma protein-A production in vitro. Hum Reprod 1986, 1, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Sinosich, M.J.; Lee, J.; Wolf, J.P.; Williams, R.F.; Hodgen, G.D. RU 486 induced suppression of placental neutrophil elastase inhibitor levels. Placenta 1989, 10, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Hourvitz, A.; Kuwahara, A.; Hennebold, J.D.; Tavares, A.B.; Negishi, H.; Lee, T.H.; Erickson, G.F.; Adashi, E.Y. The regulated expression of the pregnancy-associated plasma protein-A in the rodent ovary: a proposed role in the development of dominant follicles and of corpora lutea. Endocrinology 2002, 143, 1833–1844. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.; Sonntag, B.; Hwang, S.S.; Byerly, T.; Hourvitz, A.; Adashi, E.Y.; Shimasaki, S.; Erickson, G.F. Pregnancy-associated plasma protein-a production in rat granulosa cells: stimulation by follicle-stimulating hormone and inhibition by the oocyte-derived bone morphogenetic protein-15. Endocrinology 2004, 145, 3686–3695. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, J.; Penny, D.; Monget, P.; Arraztoa, J.A.; Fogelson, L.J.; Bondy, C.A. Insulin-like growth factor binding protein 4 expression parallels luteinizing hormone receptor expression and follicular luteinization in the primate ovary. Biol Reprod 2003, 69, 22–29. [Google Scholar] [CrossRef]
- Conover, C.A. Key questions and answers about pregnancy-associated plasma protein-A. Trends Endocrinol Metab 2012, 23, 242–249. [Google Scholar] [CrossRef]
- Suzuki, M.; Kobayashi, H.; Tanaka, Y.; Hirashima, Y.; Kanayama, N.; Takei, Y.; Saga, Y.; Suzuki, M.; Itoh, H.; Terao, T. Bikunin target genes in ovarian cancer cells identified by microarray analysis. J Biol Chem 2003, 278, 14640–14646. [Google Scholar] [CrossRef]
- Conover, C.A.; Bale, L.K.; Harrington, S.C.; Resch, Z.T.; Overgaard, M.T.; Oxvig, C. Cytokine stimulation of pregnancy-associated plasma protein A expression in human coronary artery smooth muscle cells: inhibition by resveratrol. Am J Physiol Cell Physiol 2006, 290, C183–188. [Google Scholar] [CrossRef] [PubMed]
- Chander, H.; Halpern, M.; Resnick-Silverman, L.; Manfredi, J.J.; Germain, D. Skp2B overexpression alters a prohibitin-p53 axis and the transcription of PAPP-A, the protease of insulin-like growth factor binding protein 4. PLoS One 2011, 6, e22456. [Google Scholar] [CrossRef] [PubMed]
- Melo, C.A.; Drost, J.; Wijchers, P.J.; van de Werken, H.; de Wit, E.; Oude Vrielink, J.A.; Elkon, R.; Melo, S.A.; Léveillé, N.; Kalluri, R.; et al. eRNAs are required for p53-dependent enhancer activity and gene transcription. Mol Cell 2013, 49, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Drost, J.; Mantovani, F.; Tocco, F.; Elkon, R.; Comel, A.; Holstege, H.; Kerkhoven, R.; Jonkers, J.; Voorhoeve, P.M.; Agami, R.; et al. BRD7 is a candidate tumour suppressor gene required for p53 function. Nat Cell Biol 2010, 12, 380–389. [Google Scholar] [CrossRef]
- Salim, H.; Arvanitis, A.; de Petris, L.; Kanter, L.; Hååg, P.; Zovko, A.; Özata, D.M.; Lui, W.O.; Lundholm, L.; Zhivotovsky, B.; et al. miRNA-214 is related to invasiveness of human non-small cell lung cancer and directly regulates alpha protein kinase 2 expression. Genes Chromosomes Cancer 2013, 52, 895–911. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, D.; Cao, L.; Zhang, R.; Zhou, J.; Chen, H.; Li, Y.; Li, M.; Cao, J.; Wang, Z. MiR-490-3p modulates the proliferation of vascular smooth muscle cells induced by ox-LDL through targeting PAPP-A. Cardiovasc Res 2013, 100, 272–279. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, B.; Ming, L.; Qin, H.; Zheng, L.; Yue, Z.; Cheng, Z.; Wang, Y.; Zhang, D.; Liu, C.; et al. MicroRNA-141 inhibits vascular smooth muscle cell proliferation through targeting PAPP-A. Int J Clin Exp Pathol 2015, 8, 14401–14408. [Google Scholar]
- Yu, H.; Rohan, T. Role of the insulin-like growth factor family in cancer development and progression. J Natl Cancer Inst 2000, 92, 1472–1489. [Google Scholar] [CrossRef]
- Neuzillet, Y.; Chapeaublanc, E.; Krucker, C.; De Koning, L.; Lebret, T.; Radvanyi, F.; Bernard-Pierrot, I. IGF1R activation and the in vitro antiproliferative efficacy of IGF1R inhibitor are inversely correlated with IGFBP5 expression in bladder cancer. BMC Cancer 2017, 17, 636. [Google Scholar] [CrossRef] [PubMed]
- Prithviraj, P.; Anaka, M.; McKeown, S.J.; Permezel, M.; Walkiewicz, M.; Cebon, J.; Behren, A.; Jayachandran, A. Pregnancy associated plasma protein-A links pregnancy and melanoma progression by promoting cellular migration and invasion. Oncotarget 2015, 6, 15953–15965. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ding, N.; Li, Y.; Cheng, H.; Wang, D.; Yang, Q.; Deng, Y.; Yang, Y.; Li, Y.; Ruan, X.; et al. Insulin-like growth factor binding protein 5 (IGFBP5) functions as a tumor suppressor in human melanoma cells. Oncotarget 2015, 6, 20636–20649. [Google Scholar] [CrossRef] [PubMed]
- Kleinberg, D.L. Role of IGF-I in normal mammary development. Breast Cancer Res Treat 1998, 47, 201–208. [Google Scholar] [CrossRef]
- Christopoulos, P.F.; Msaouel, P.; Koutsilieris, M. The role of the insulin-like growth factor-1 system in breast cancer. Mol Cancer 2015, 14, 43. [Google Scholar] [CrossRef] [PubMed]
- Akkiprik, M.; Feng, Y.; Wang, H.; Chen, K.; Hu, L.; Sahin, A.; Krishnamurthy, S.; Ozer, A.; Hao, X.; Zhang, W. Multifunctional roles of insulin-like growth factor binding protein 5 in breast cancer. Breast Cancer Res 2008, 10, 212. [Google Scholar] [CrossRef]
- Dittmer, J. Biological effects and regulation of IGFBP5 in breast cancer. Front Endocrinol (Lausanne) 2022, 13, 983793. [Google Scholar] [CrossRef]
- Stacey, S.N.; Manolescu, A.; Sulem, P.; Rafnar, T.; Gudmundsson, J.; Gudjonsson, S.A.; Masson, G.; Jakobsdottir, M.; Thorlacius, S.; Helgason, A.; et al. Common variants on chromosomes 2q35 and 16q12 confer susceptibility to estrogen receptor-positive breast cancer. Nat Genet 2007, 39, 865–869. [Google Scholar] [CrossRef]
- Panagiotou, G.; Papakonstantinou, E.; Vagionas, A.; Polyzos, S.A.; Mantzoros, C.S. Serum Levels of Activins, Follistatins, and Growth Factors in Neoplasms of the Breast: A Case-Control Study. J Clin Endocrinol Metab 2019, 104, 349–358. [Google Scholar] [CrossRef]
- Fleming, J.M.; Leibowitz, B.J.; Kerr, D.E.; Cohick, W.S. IGF-I differentially regulates IGF-binding protein expression in primary mammary fibroblasts and epithelial cells. J Endocrinol 2005, 186, 165–178. [Google Scholar] [CrossRef]
- Ryan, A.J.; Napoletano, S.; Fitzpatrick, P.A.; Currid, C.A.; O'Sullivan, N.C.; Harmey, J.H. Expression of a protease-resistant insulin-like growth factor-binding protein-4 inhibits tumour growth in a murine model of breast cancer. Br J Cancer 2009, 101, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Smith, Y.E.; Toomey, S.; Napoletano, S.; Kirwan, G.; Schadow, C.; Chubb, A.J.; Mikkelsen, J.H.; Oxvig, C.; Harmey, J.H. Recombinant PAPP-A resistant insulin-like growth factor binding protein 4 (dBP4) inhibits angiogenesis and metastasis in a murine model of breast cancer. BMC Cancer 2018, 18, 1016. [Google Scholar] [CrossRef]
- Radke, S.; Pirkmaier, A.; Germain, D. Differential expression of the F-box proteins Skp2 and Skp2B in breast cancer. Oncogene 2005, 24, 3448–3458. [Google Scholar] [CrossRef] [PubMed]
- Espelund, U.; Renehan, A.G.; Cold, S.; Oxvig, C.; Lancashire, L.; Su, Z.; Flyvbjerg, A.; Frystyk, J. Prognostic relevance and performance characteristics of serum IGFBP-2 and PAPP-A in women with breast cancer: a long-term Danish cohort study. Cancer Med 2018, 7, 2391–2404. [Google Scholar] [CrossRef] [PubMed]
- Takabatake, Y.; Oxvig, C.; Nagi, C.; Adelson, K.; Jaffer, S.; Schmidt, H.; Keely, P.J.; Eliceiri, K.W.; Mandeli, J.; Germain, D. Lactation opposes pappalysin-1-driven pregnancy-associated breast cancer. EMBO Mol Med 2016, 8, 388–406. [Google Scholar] [CrossRef]
- Slocum, E.; Craig, A.; Villanueva, A.; Germain, D. Parity predisposes breasts to the oncogenic action of PAPP-A and activation of the collagen receptor DDR2. Breast Cancer Res 2019, 21, 56. [Google Scholar] [CrossRef]
- Jayachandran, A.; Dhungel, B.; Steel, J.C. Epithelial-to-mesenchymal plasticity of cancer stem cells: therapeutic targets in hepatocellular carcinoma. J Hematol Oncol 2016, 9, 74. [Google Scholar] [CrossRef]
- Shrestha, R.; Bridle, K.R.; Crawford, D.H.G.; Jayachandran, A. Immune checkpoint molecules are regulated by transforming growth factor (TGF)-β1-induced epithelial-to-mesenchymal transition in hepatocellular carcinoma. Int J Med Sci 2021, 18, 2466–2479. [Google Scholar] [CrossRef]
- Muqbil, I.; Wu, J.; Aboukameel, A.; Mohammad, R.M.; Azmi, A.S. Snail nuclear transport: the gateways regulating epithelial-to-mesenchymal transition? Semin Cancer Biol 2014, 27, 39–45. [Google Scholar] [CrossRef]
- Valiathan, R.R.; Marco, M.; Leitinger, B.; Kleer, C.G.; Fridman, R. Discoidin domain receptor tyrosine kinases: new players in cancer progression. Cancer Metastasis Rev 2012, 31, 295–321. [Google Scholar] [CrossRef]
- Totzkay, D.; Silk, K.J.; Thomas, B.; Walling, B.M.; Smith, S.W. Women's Understanding of Windows of Susceptibility and the Role of the Environment in Breast Cancer Risk. J Cancer Educ 2023, 38, 115–126. [Google Scholar] [CrossRef]
- Veillet, A.L.; Haag, J.D.; Remfert, J.L.; Meilahn, A.L.; Samuelson, D.J.; Gould, M.N. Mcs5c: a mammary carcinoma susceptibility locus located in a gene desert that associates with tenascin C expression. Cancer Prev Res (Phila) 2011, 4, 97–106. [Google Scholar] [CrossRef]
- Samuelson, D.J.; Haag, J.D.; Lan, H.; Monson, D.M.; Shultz, M.A.; Kolman, B.D.; Gould, M.N. Physical evidence of Mcs5, a QTL controlling mammary carcinoma susceptibility, in congenic rats. Carcinogenesis 2003, 24, 1455–1460. [Google Scholar] [CrossRef]
- Henning, A.N.; Haag, J.D.; Smits, B.M.; Gould, M.N. The Non-coding Mammary Carcinoma Susceptibility Locus, Mcs5c, Regulates Pappa Expression via Age-Specific Chromatin Folding and Allele-Dependent DNA Methylation. PLoS Genet 2016, 12, e1006261. [Google Scholar] [CrossRef]
- Lambe, M.; Hsieh, C.; Trichopoulos, D.; Ekbom, A.; Pavia, M.; Adami, H.O. Transient increase in the risk of breast cancer after giving birth. N Engl J Med 1994, 331, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Asztalos, S.; Pham, T.N.; Gann, P.H.; Hayes, M.K.; Deaton, R.; Wiley, E.L.; Emmadi, R.; Kajdacsy-Balla, A.; Banerji, N.; McDonald, W.; et al. High incidence of triple negative breast cancers following pregnancy and an associated gene expression signature. Springerplus 2015, 4, 710. [Google Scholar] [CrossRef] [PubMed]
- Bukowski, R.; Chlebowski, R.T.; Thune, I.; Furberg, A.S.; Hankins, G.D.; Malone, F.D.; D'Alton, M.E. Birth weight, breast cancer and the potential mediating hormonal environment. PLoS One 2012, 7, e40199. [Google Scholar] [CrossRef] [PubMed]
- Wallace, T.R.; Tarullo, S.E.; Crump, L.S.; Lyons, T.R. Studies of postpartum mammary gland involution reveal novel pro-metastatic mechanisms. J Cancer Metastasis Treat 2019, 5. [Google Scholar] [CrossRef]
- Allan, G.J.; Beattie, J.; Flint, D.J. The role of IGFBP-5 in mammary gland development and involution. Domest Anim Endocrinol 2004, 27, 257–266. [Google Scholar] [CrossRef]
- Watson, C.J. Involution: apoptosis and tissue remodelling that convert the mammary gland from milk factory to a quiescent organ. Breast Cancer Res 2006, 8, 203. [Google Scholar] [CrossRef]
- Jena, M.K.; Jaswal, S.; Kumar, S.; Mohanty, A.K. Molecular mechanism of mammary gland involution: An update. Dev Biol 2019, 445, 145–155. [Google Scholar] [CrossRef]
- Chen, B.K.; Leiferman, K.M.; Pittelkow, M.R.; Overgaard, M.T.; Oxvig, C.; Conover, C.A. Localization and regulation of pregnancy-associated plasma protein a expression in healing human skin. J Clin Endocrinol Metab 2003, 88, 4465–4471. [Google Scholar] [CrossRef]
- Lyons, T.R.; O'Brien, J.; Borges, V.F.; Conklin, M.W.; Keely, P.J.; Eliceiri, K.W.; Marusyk, A.; Tan, A.C.; Schedin, P. Postpartum mammary gland involution drives progression of ductal carcinoma in situ through collagen and COX-2. Nat Med 2011, 17, 1109–1115. [Google Scholar] [CrossRef]
- Conklin, M.W.; Eickhoff, J.C.; Riching, K.M.; Pehlke, C.A.; Eliceiri, K.W.; Provenzano, P.P.; Friedl, A.; Keely, P.J. Aligned collagen is a prognostic signature for survival in human breast carcinoma. Am J Pathol 2011, 178, 1221–1232. [Google Scholar] [CrossRef]
- Provenzano, P.P.; Inman, D.R.; Eliceiri, K.W.; Knittel, J.G.; Yan, L.; Rueden, C.T.; White, J.G.; Keely, P.J. Collagen density promotes mammary tumor initiation and progression. BMC Med 2008, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Drake, C.G.; Jaffee, E.; Pardoll, D.M. Mechanisms of immune evasion by tumors. Adv Immunol 2006, 90, 51–81. [Google Scholar] [CrossRef] [PubMed]
- Vinay, D.S.; Ryan, E.P.; Pawelec, G.; Talib, W.H.; Stagg, J.; Elkord, E.; Lichtor, T.; Decker, W.K.; Whelan, R.L.; Kumara, H.; et al. Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin Cancer Biol 2015, 35 Suppl, S185–S198. [Google Scholar] [CrossRef]
- Fang, Y.; Wang, L.; Wan, C.; Sun, Y.; Van der Jeught, K.; Zhou, Z.; Dong, T.; So, K.M.; Yu, T.; Li, Y.; et al. MAL2 drives immune evasion in breast cancer by suppressing tumor antigen presentation. J Clin Invest 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Dersh, D.; Yewdell, J.W. Immune MAL2-practice: breast cancer immunoevasion via MHC class I degradation. J Clin Invest 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Cho, S.W. The Evasion Mechanisms of Cancer Immunity and Drug Intervention in the Tumor Microenvironment. Front Pharmacol 2022, 13, 868695. [Google Scholar] [CrossRef]
- Southall, P.J.; Boxer, G.M.; Bagshawe, K.D.; Hole, N.; Bromley, M.; Stern, P.L. Immunohistological distribution of 5T4 antigen in normal and malignant tissues. Br J Cancer 1990, 61, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Jungbluth, A.A.; Silva, W.A., Jr.; Iversen, K.; Frosina, D.; Zaidi, B.; Coplan, K.; Eastlake-Wade, S.K.; Castelli, S.B.; Spagnoli, G.C.; Old, L.J.; et al. Expression of cancer-testis (CT) antigens in placenta. Cancer Immun 2007, 7, 15. [Google Scholar] [PubMed]
- Silva, W.A., Jr.; Gnjatic, S.; Ritter, E.; Chua, R.; Cohen, T.; Hsu, M.; Jungbluth, A.A.; Altorki, N.K.; Chen, Y.T.; Old, L.J.; et al. PLAC1, a trophoblast-specific cell surface protein, is expressed in a range of human tumors and elicits spontaneous antibody responses. Cancer Immun 2007, 7, 18. [Google Scholar]
- Salanti, A.; Clausen, T.M.; Agerbaek, M.O.; Al Nakouzi, N.; Dahlback, M.; Oo, H.Z.; Lee, S.; Gustavsson, T.; Rich, J.R.; Hedberg, B.J.; et al. Targeting Human Cancer by a Glycosaminoglycan Binding Malaria Protein. Cancer Cell 2015, 28, 500–514. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, A.; Thiede, M.; Grunewald, T.G.; Alba Rubio, R.; Richter, G.H.; Kirchner, T.; Busch, D.H.; Burdach, S.; Thiel, U. Pappalysin-1 T cell receptor transgenic allo-restricted T cells kill Ewing sarcoma in vitro and in vivo. Oncoimmunology 2017, 6, e1273301. [Google Scholar] [CrossRef]
- Nakasato, M.; Kohsaka, H.; Mizutani, T.; Watanabe, G.; Taya, K.; Nagaoka, K. Pregnancy-associated plasma protein (PAPP)-A expressed in the mammary gland controls epithelial cell proliferation and differentiation. Endocrine 2013, 43, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Prithviraj, P.; Shrestha, R.; Sharma, R.; Anaka, M.; Bridle, K.R.; Kannourakis, G.; Crawford, D.H.G.; Jayachandran, A. Prognostic Role of Immune Checkpoint Regulators in Cholangiocarcinoma: A Pilot Study. J Clin Med 2021, 10. [Google Scholar] [CrossRef]
- Shrestha, R.; Prithviraj, P.; Anaka, M.; Bridle, K.R.; Crawford, D.H.G.; Dhungel, B.; Steel, J.C.; Jayachandran, A. Monitoring Immune Checkpoint Regulators as Predictive Biomarkers in Hepatocellular Carcinoma. Front Oncol 2018, 8, 269. [Google Scholar] [CrossRef]
- Sallusto, F.; Lenig, D.; Forster, R.; Lipp, M.; Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 1999, 401, 708–712. [Google Scholar] [CrossRef]
- Wieczorek, M.; Abualrous, E.T.; Sticht, J.; Alvaro-Benito, M.; Stolzenberg, S.; Noe, F.; Freund, C. Major Histocompatibility Complex (MHC) Class I and MHC Class II Proteins: Conformational Plasticity in Antigen Presentation. Front Immunol 2017, 8, 292. [Google Scholar] [CrossRef]
- Leone, P.; Shin, E.C.; Perosa, F.; Vacca, A.; Dammacco, F.; Racanelli, V. MHC class I antigen processing and presenting machinery: organization, function, and defects in tumor cells. J Natl Cancer Inst 2013, 105, 1172–1187. [Google Scholar] [CrossRef]
- Chattopadhyay, M.; Jenkins, E.C.; Lechuga-Vieco, A.V.; Nie, K.; Fiel, M.I.; Rialdi, A.; Guccione, E.; Enriquez, J.A.; Sia, D.; Lujambio, A.; et al. The portrait of liver cancer is shaped by mitochondrial genetics. Cell Rep 2022, 38, 110254. [Google Scholar] [CrossRef]
- Lodhia, K.A.; Tienchaiananda, P.; Haluska, P. Understanding the Key to Targeting the IGF Axis in Cancer: A Biomarker Assessment. Front Oncol 2015, 5, 142. [Google Scholar] [CrossRef]
- Pohlman, A.W.; Moudgalya, H.; Jordano, L.; Lobato, G.C.; Gerard, D.; Liptay, M.J.; Seder, C.W.; Borgia, J.A. The role of IGF-pathway biomarkers in determining risks, screening, and prognosis in lung cancer. Oncotarget 2022, 13, 393–407. [Google Scholar] [CrossRef]
- Zhou, Q.; Mao, Y.Q.; Jiang, W.D.; Chen, Y.R.; Huang, R.Y.; Zhou, X.B.; Wang, Y.F.; Shi, Z.; Wang, Z.S.; Huang, R.P. Development of IGF signaling antibody arrays for the identification of hepatocellular carcinoma biomarkers. PLoS One 2012, 7, e46851. [Google Scholar] [CrossRef]
- Douglas, J.B.; Silverman, D.T.; Pollak, M.N.; Tao, Y.; Soliman, A.S.; Stolzenberg-Solomon, R.Z. Serum IGF-I, IGF-II, IGFBP-3, and IGF-I/IGFBP-3 molar ratio and risk of pancreatic cancer in the prostate, lung, colorectal, and ovarian cancer screening trial. Cancer Epidemiol Biomarkers Prev 2010, 19, 2298–2306. [Google Scholar] [CrossRef]
- Kuhajda, F.P.; Abeloff, M.D.; Eggleston, J.C. Pregnancy-associated plasma protein A: a clinically significant predictor of early recurrence in stage II breast carcinoma. Hum Pathol 1985, 16, 228–235. [Google Scholar] [CrossRef]
- Kuhajda, F.P.; Eggleston, J.C. Pregnancy-associated plasma protein A and extensive necrosis. Clinically significant predictors of early recurrence in stage I estrogen receptor-negative breast carcinoma. Lab Invest 1985, 53, 101–107. [Google Scholar]
- Ekyalongo, R.C.; Yee, D. Revisiting the IGF-1R as a breast cancer target. npj Precision Oncology 2017, 1, 14. [Google Scholar] [CrossRef]
- Hamilton, N.; Austin, D.; Márquez-Garbán, D.; Sanchez, R.; Chau, B.; Foos, K.; Wu, Y.; Vadgama, J.; Pietras, R. Receptors for Insulin-Like Growth Factor-2 and Androgens as Therapeutic Targets in Triple-Negative Breast Cancer. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef]
Figure 1.
Pathological expression of pregnancy associated plasma protein-A (PAPP-A) correlates with triple negative breast cancer (TNBC).
Figure 1.
Pathological expression of pregnancy associated plasma protein-A (PAPP-A) correlates with triple negative breast cancer (TNBC).
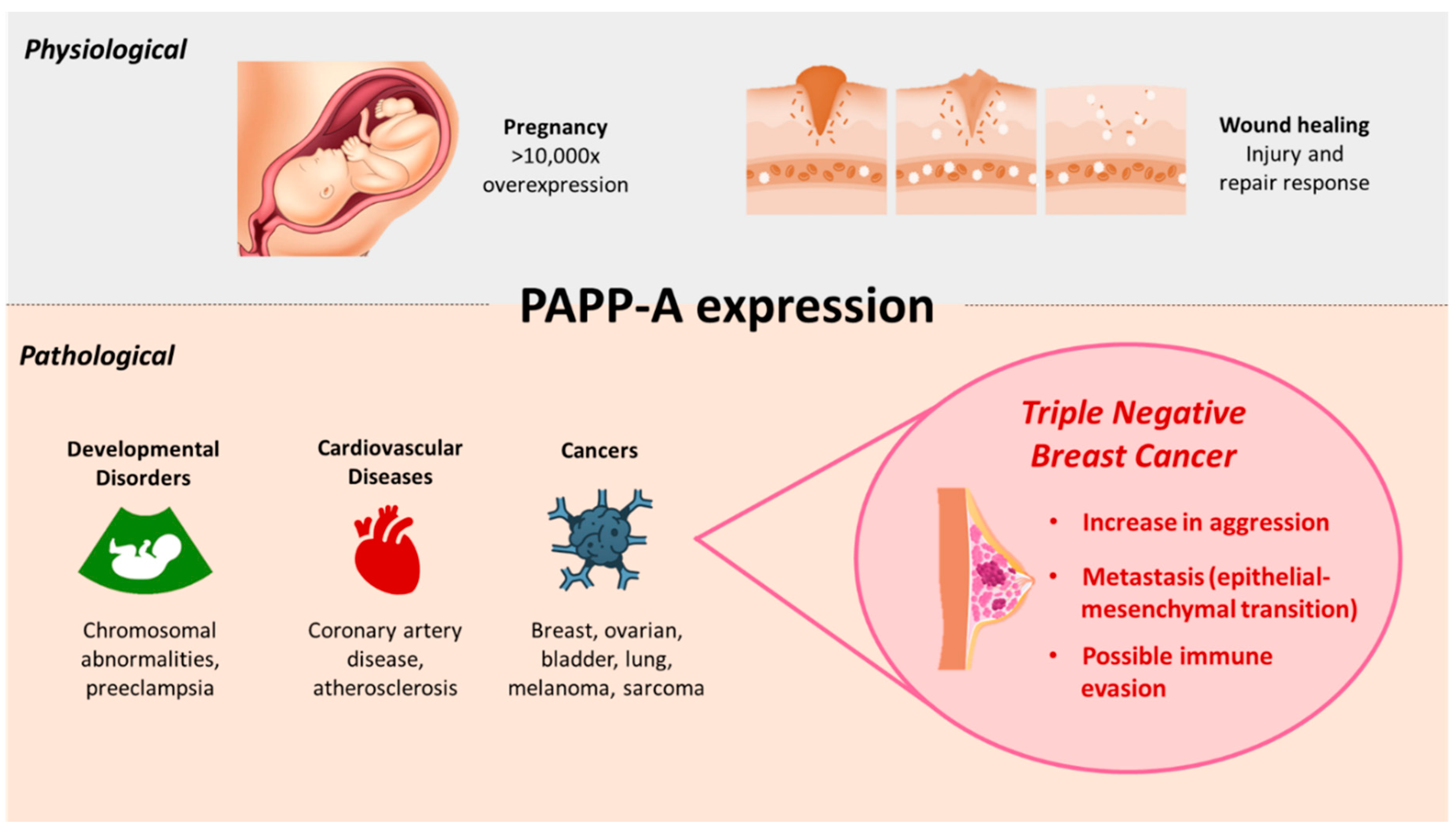
Figure 2.
PAPP-A impacts pregnancy, epithelial-mesenchymal-transition, window of susceptibility and immunomodulation – representing potential clinical targeting of TNBC.
Figure 2.
PAPP-A impacts pregnancy, epithelial-mesenchymal-transition, window of susceptibility and immunomodulation – representing potential clinical targeting of TNBC.
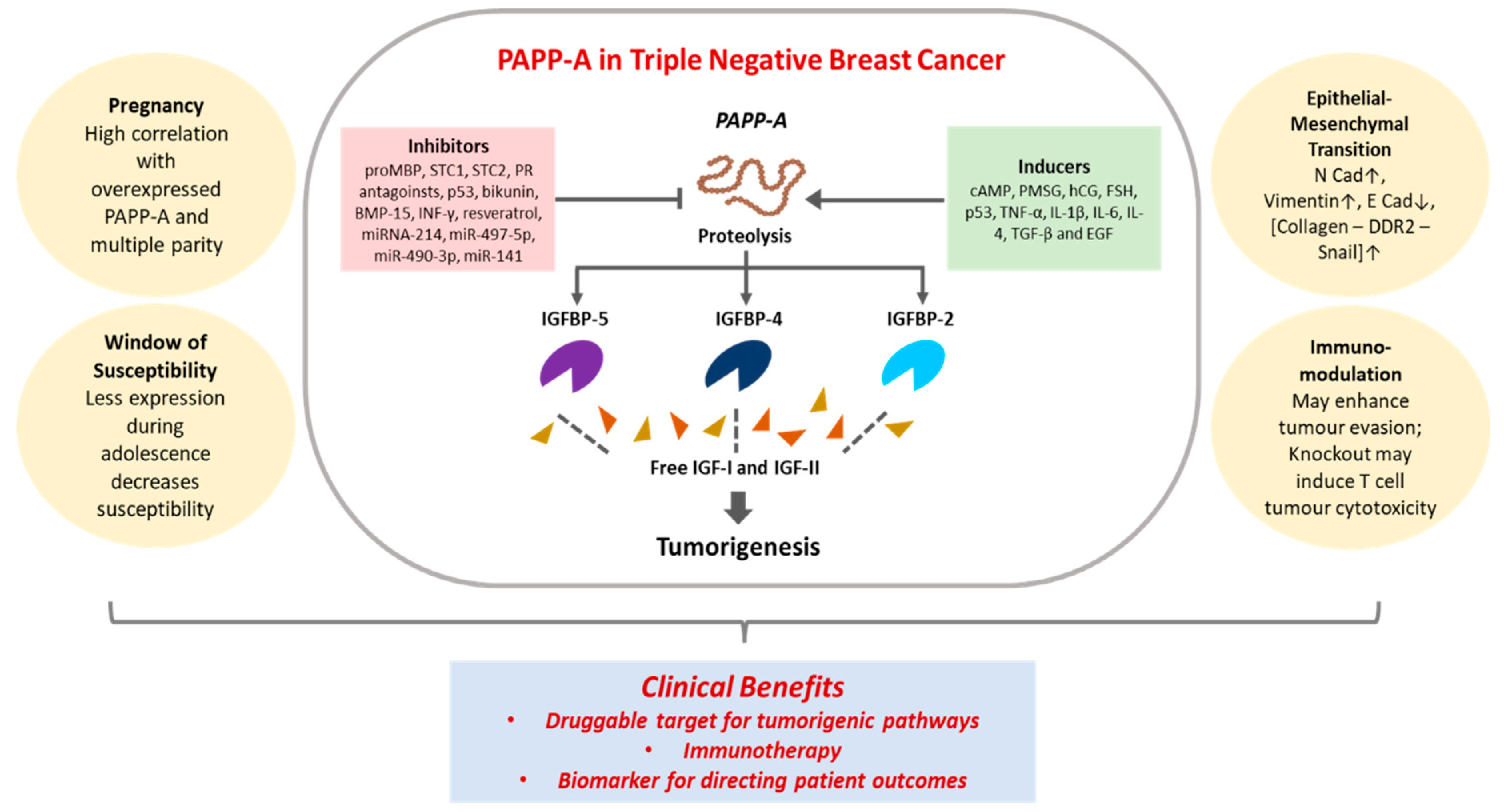
Table 1.
Known PAPP-A mediated IGFBP proteolysis.
| IGFBP | IGF requirement | Role of PAPP-A | Kinetic efficiency | Reference |
|---|---|---|---|---|
| IGFBP-4 | IGF-II (more efficient) or IGF-I | Cleavage at Met-135 and Lys-136 | High | [57] |
| IGFBP-4 | Not required | *Same site | Very low | [57] |
| IGFBP-5 | Not required; inhibited by IGF presence | Cleavage at Ser-143 and Lys-144 | High | [59,60] |
| IGFBP-2 | IGF dependent | Cleavage at Gln-165 and Met-166 | Less than IGFBP-4 | [61] |
Table 4.
PAPP-A as a clinical biomarker for BC.
| Clinical indicator | Sample Size | Analysis method | References |
|---|---|---|---|
| Independent predictor of early recurrence | Stage II BC: 30 cases (treated with low or standard chemotherapy) | Immunostaining and clinicopathology | [138] |
| Independent predictor of early recurrence | Stage I ER negative: 40 cases | Immunostaining and clinicopathology | [139] |
| Independent predictor of early recurrence; independent of ER status | Stage I ER positive: 30 cases | Immunostaining and clinicopathology | [15] |
| Worse prognosis seen in elevated PAPP-A; Independently prognostic for RFS and OS in the long-term | Early BC (with and without treatment): 301 cases, Non-cancer: 531 cases |
Serum assays on patient samples | [95] |
| Predictor of malignancy presence; positive association with serum activin A, serum activin B, total IGFBP-4, and correlation with total IGF-I; negative association with total cholesterol and triglycerides | Benign tumors: 100 cases, Malignant BC (treatment naïve and chemotherapy): 145 cases Non-cancer: 100 cases |
Serum assays on patient samples | [90] |
| High expression correlated with lymph node metastasis and high-grade tumor; worse prognosis, disease recurrence and poor OS in high-grade BC | BC: 45 cases (with 80% TNBC) | IHC on TMA | [25] |
| PAPP-A/SNAI1/COL1A1 expression panel: High score correlates with distant metastases | Primary BC: dataset of 327 cases | Gene set analysis | [97] |
| Silencing links with distant metastases | Invasive BC: 173 cases Normal breast: 30 cases |
DNA methylation analysis on FFPE | [26] |
| Low serum level in first trimester: greater long-term BC risk | 677,247 pregnancies | Biochemical screening | [27] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



