
Submitted:
23 July 2023
Posted:
25 July 2023
You are already at the latest version
Abstract
Large terrestrial carnivores play a crucial role in top-down control within terrestrial ecosystems, maintaining ecosystem stability and biodiversity. However, intense interspecific competition often arises among sympatric large carnivores, leading to population reductions or even extinctions. Spatial partitioning through divergent habitat selection helps mitigate such competition. In Xinlong County, Sichuan Province, we used 293 infrared cameras for monitoring from September to May 2016 and March to October 2022. By employing the Generalized Linear Model (GLM) and the Maximum Entropy Model (MaxEnt), we developed an ensemble model predicting the suitable habitat distribution of leopards (Panthera pardus) and wolves (Canis lupus). We analyzed the main environmental factors influencing habitat selection and the fragmentation of suitable habitats. We found that suitable habitat distribution differed significantly between them. Both species preferred areas with gentle slopes close to settlements. While leopards' habitat selection primarily depended on the distance from settlements, the slope was predominant for wolves. Suitable habitats displayed aggregation, yet wolves exhibited higher fragmentation and more complex patch shapes, indicating greater susceptibility to human activities. These results suggest that sympatric large carnivores, such as leopards and wolves, can reduce spatial competition intensity and promote spatial partitioning by selecting divergent suitable habitats, thereby facilitating species coexistence.
Keywords:
Interspecific competition
; Spatial ecological niche
; Ensemble model
; Habitat selection
; Xinlong County
1. Introduction
Carnivores hold a pivotal position within the food chain, crucially contributing to the preservation of ecological balance and species diversity. Notably, large carnivores (weighing more than 15 kg) exert control over populations of herbivorous animals and secondary predators through trophic cascade effects, thereby upholding ecosystem stability [1,2]. Nevertheless, escalating global anthropogenic disturbances, habitat loss, fragmentation, reduced fecundity, and climate change impacts have resulted in a rapid decline of once-abundant large carnivore populations. Currently, they are primarily concentrated in eight global hotspots [1], the southwestern mountainous area of China emerges as a significant hotspot characterized by the concentrated distribution of large carnivores. The convergence of carnivore distributions inevitably escalates interspecies competition. In line with the competitive exclusion principle, competitors with similar ecological characteristics cannot coexist within the same niche simultaneously or for an extended period. To achieve coexistence, a modification in the ecological niche of at least one of the competitors is necessary [3]. Consequently, the survival of large terrestrial carnivores is imperiled not only by external stressors but also by intense interspecific competitive interactions, prompting ecologists to focus on investigating coexistence mechanisms and conservation strategies for these magnificent predators.
In recent years, the study of carnivore competition and coexistence has predominantly revolved around three ecological niche dimensions: spatial, temporal, and nutritional dynamics [2]. Among these dimensions, exploring the spatial ecological niche forms the fundamental basis for comprehending carnivore competition and coexistence at the local scale. Notably, disparities in habitat preferences among species serve as an effective mechanism, in driving spatial niche differentiation, particularly for large carnivores. For instance, in Nepal's National Park, leopards strategically position themselves at the peripheries of tiger (Panthera tigris) territories, deliberately selecting relatively suboptimal habitats to achieve spatial avoidance from tigers [4].
Habitat selection constitutes a critical aspect of large carnivore survival, influenced by various factors encompassing individual characteristics, environmental conditions, and anthropogenic disturbances. Understanding the intricate interplay between these factors is paramount for effective conservation management. Large carnivores exhibit selective preferences driven by adaptability, behavioral characteristics, and the need to enhance resilience against external threats and interspecific competition. For instance, the tiger's distinctive orange coat with black stripes provides excellent camouflage within forest thickets, effectively blending with light and shade patterns. This distinctive coat provides effective camouflage, as certain herbivores cannot distinguish between orange and green [5]. The availability of crucial resources, including water and prey, shapes carnivore habitat selection. A study conducted in Iran highlighted the significance of ecological zones with higher vegetation cover and rainfall in providing abundant prey resources for leopards, influencing their habitat preference [6]. The interspecific competition also plays a pivotal role, as observed in South Asia, where leopards favor habitats with relatively low prey resources and vegetation cover to minimize competition with tigers [4]. Moreover, escalating anthropogenic disturbances introduce additional complexities. For instance, road construction and infrastructure edge effects have prompted tiger and leopard populations in Thai national parks to shift away from human settlements [7]. These disturbances contribute to habitat loss and fragmentation, posing significant threats to the persistence of large carnivore populations. Habitat fragmentation can impair key ecosystem functions and cause long-term, cumulative effects by reducing biodiversity [8]. Fragmentation processes result in increased numbers of habitat patches, isolation between patches, and potential habitat loss [9]. Reducing anthropogenic disturbances and enhancing habitat connectivity represent critical measures in mitigating the adverse effects of fragmentation. For instance, heterogeneous habitat connectivity has been instrumental in safeguarding the remaining cheetah populations in Iran [10]. In conclusion, the selection of suitable habitats for large carnivores necessitates considering prey availability, environmental conditions, human disturbances, and more. Analyzing habitat fragmentation is essential in guiding the development of scientifically grounded conservation management strategies.
Both leopards and wolves, the focal species of this study, are large carnivores with distinct characteristics and habitat preferences. Leopards, solitary ambush hunters, tend to favor secluded habitats for their survival. Their global distribution spans sub-Saharan Africa, Southeast Asia, and East Asia (Figure 1a). On the other hand, wolves are group-living animals known for their opportunistic predation and remarkable adaptability to various environments, including tundra, grassland, and forests [11]. Wolves have a wide-ranging global distribution (Figure 1a). There is a certain degree of spatial overlap between them, and due to their dietary preferences, coexisting wolves and leopards often engage in intense competition. Studies conducted in areas where wolves and leopards coexist have found that when wolves are absent, the survival rate of leopards can increase to 98% [12].
Currently, comparative research is scarce on habitat selection differences between leopards and wolves in both domestic and international studies on large carnivores. The southwestern mountainous region of China represents one of the few areas where leopards and wolves coexist. Notably, diverse habitats and abundant prey resources make it an excellent area for investigating the differences in habitat selection between these two species [13].
This study focuses on Xinlong County, situated within the southwestern mountain range. Using camera traps to investigate the disparities in suitable habitat distribution for leopards and wolves, as well as their responses to environmental factors. Our primary objective is to ascertain whether the habitat selection strategies employed by leopards and wolves in Xinlong County contribute to the differentiation of their spatial ecological niches. Thus, we propose the following hypotheses: (1) leopards and wolves exhibit significant differences in the distribution of suitable habitats within the same area; (2) leopards show a preference for forested areas, whereas wolves demonstrate a predilection for meadow areas; (3) anthropogenic disturbance and slope gradients are the primary environmental factors influencing the selection of suitable habitats for leopards and wolves, respectively, with both species favoring areas located far from settlements and featuring gentle slopes; (4) the area of suitable habitat patches is greater for wolves compared to leopards, but certain areas experience substantial human interference, resulting in fragmented suitable habitats.
2. Materials and Methods
2.1. study area
The Xinlong region, located in the Ganzi Tibetan Autonomous Prefecture of Sichuan Province, lies in the middle reaches of the Yalong River (99°37′~ 100°54′ E, 30°23′ ~ 31°32′ N). The topography of the region is characterized by higher elevations in the north gradually descending to lower elevations in the south. The Yalong River flows through the county from north to south, dividing it into eastern and western parts and creating a typical alpine valley terrain with an altitude range exceeding 3000 m. The region experiences a typical plateau monsoon climate, characterized by hot and rainy summers, and cold and dry winters, with significant vertical variations in climate. The interplay of terrain and climate has created diverse habitats for wildlife and sustained abundant wild flora and fauna resources. In addition to top-level carnivores such as leopards and wolves, the area is home to secondary predators like a red fox (Vulpes vulpes), leopard cat (Prionailurus bengalensis), and Pallas's cat (Otocolobus manul), as well as a rich array of prey species including blue sheep (Pseudois nayaur), tufted deer (Elaphodus cephalophus), wild boar (Sus scrofa), and Chinese serow (Capricornis milneedwardsii) [13]. The forests and meadows in the region are subjected to varying degrees of anthropogenic disturbance primarily caused by Tibetan herders, whose livelihoods predominantly rely on grazing practices. However, the cultural belief of most Tibetans that refrains from killing animals has contributed to the persistence of numerous endangered and rare species, thereby aiding in the preservation of biodiversity within the region. The combined influence of the natural environment and cultural customs have created highly favorable conditions for the coexistence of wild animals in the Xinlong region.
2.2. Data collection
2.2.1. Camera trapping surveys
Data on species distribution were obtained through infrared camera trapping. The monitoring was conducted during two distinct periods: September 2016 to May 2017, during which 149 cameras were utilized, and March to October 2022, during which 144 cameras were deployed. In the initial period, the infrared cameras were placed randomly, with a focus on areas known for frequent wildlife activities. In 2022, we established a systematic carnivore monitoring network in Xinlong County using a grid-based layout with 1 km ⨯ 1 km grid cells. Within each grid cell, we placed 1 camera, ensuring a minimum distance of 500m between cameras. The specific deployment criteria were as follows: 1) the presence of animal traces (such as hair, feces, and footprints) at the chosen sites; 2) ensuring that the monitoring target is at the center of the camera's field of view; 3) camera parameters were set as follows: 24-hour system operation, medium sensitivity, capturing three consecutive shots followed by a 20-second video, with a one-minute interval between each sequence.
Overall, the cameras recorded a total of 183 instances of leopard and wolf occurrences, comprising 83 leopard occurrences and 100 wolf occurrences.
2.2.2. Environment variables
Based on the objectives of this study and previous relevant research, we selected topography, vegetation, and disturbance as environmental factors for analyzing the distribution of suitable habitats for leopards and wolves (Table S2) [14]. Regarding topographic variables, seven factors were considered: elevation (ELE), slope (SLP), topographic relief (TRI), distance to water (DTW), distance to cliffs (DTC), distance to valleys (DTV), and distance to ridges (DTR). All these variables were extracted from the Digital Elevation Model (DEM) of Xinlong County using ArcGIS 10.8 for surface analysis and hydrological analysis. The DEM data were obtained from the geospatial data cloud platform (https://www.gscloud.cn/). Regarding vegetation variables, the Enhanced Vegetation Index (EVI) was employed. The EVI was calculated using Landsat 8 satellite imagery, which underwent atmospheric correction and radiometric calibration. The vegetation index tool was utilized to compute the average EVI values for May and August. The EVI data were corrected using ENVI 5.3. Disturbance variables encompassed the distance to roads (DTD) and the distance to settlements (DTS). Roads and settlements vector data, as well as surface cover raster data, were acquired from the National Catalogue Service for Geographic Information (https://www.webmap.cn/). The distance analysis tool in ArcGIS 10.8 was applied to generate raster data for DTD and DTS. Furthermore, the analysis included other variables, such as land cover (LC), which comprised cropland, forest, grassland, shrubland, and other categories.
2.3. Data analysis
2.3.1. Ensemble model
Species Distribution Models (SDMs) are valuable tools for integrating species distribution data with abiotic factors. However, with the availability of various model types, it is crucial to carefully select the appropriate model and adjust its parameters according to the specific research question [15]. Among the SDM approaches, the Maximum Entropy Model (MaxEnt) is an ecological niche model that utilizes only species occurrence points and environmental variables to predict the suitable habitat distribution of the target species in the study area. MaxEnt has gained popularity in recent years due to its ability to achieve relatively accurate predictions with limited data volume [16,17,18]. Nevertheless, one limitation of MaxEnt is the potential risk of overfitting. On the other hand, the Generalized Linear Model (GLM) is also a powerful method for species distribution prediction, but its accuracy heavily relies on the quality and quantity of available samples [19]. Ensemble models that combine MaxEnt and GLM integrated the outcomes of both models, offering a means to reduce individual algorithm prediction uncertainty to some extent.
To assess the quality of the models, the dataset was divided into two subsets: the training set and the test set. Random sampling was used to allocate 20% of the total data as the test set for model validation, while the remaining 80% was used as the training set for model training. The performance of the models was evaluated using the area under the receiver operating characteristic curve (AUC), which represents the accuracy of the model simulation. The AUC value ranges can be interpreted as follows: 0.5 to 0.6 indicates model failure, 0.6 to 0.7 indicates poor performance, 0.7 to 0.8 indicates moderate performance, 0.8 to 0.9 indicates good performance, and 0.9 to 1.0 indicates excellent performance [20]. Additionally, model weights were calculated based on the AUC values, and the predictions of the GLM model were combined with those of the MaxEnt model to obtain ensemble results.
The relative importance of environmental variables predicted by each model was calculated by conducting three separate runs of the model, and the average of these three runs was considered as the result. Then, the results were normalized individually. The ensemble model then determined the relative importance of environmental variables by taking a weighted average of the normalized results. Additionally, the response curves of environmental variables were predicted based on the outcomes of the ensemble model. All analyses were conducted using R version 4.2.1.
2.3.2. Assessment of habitat suitability
The habitat suitability maps of leopards and wolves were reclassified using ArcGIS 10.8 software with a threshold value of 0.3, resulting in habitat suitability level evaluation maps. The suitable habitat area was then calculated using the image element method.
2.3.3. Assessment of suitable habitat landscapes
In this study, the leopard and wolf habitats were classified as either suitable or unsuitable. The classification results were rasterized in ArcMap 10.8 software and then imported into Fragstats V4.2.0 software as a layer. Fragstats was used to extract landscape parameters at the class scale based on this layer. Seven indices were selected for the class scale, which can be categorized into four types of indicators: area indicator (Percentage of Landscape, PLAND), shape indicator (Landscape Shape Index, LSI, Edge Density, ED, Perimeter-Area Fractal Dimension, PAFRAC), connectivity index (Connectance index, CONNECT, Aggregation index, AI), and fragmentation index (Patch Density, PD). These indices were used to analyze the landscape characteristics of suitable and unsuitable habitats for leopards and wolves in the study area (Table S1).
3. Results
3.1. Ensemble model accuracy assessment
The AUC values for leopard and wolf habitat prediction were 0.81 and 0.82, respectively. According to the AUC evaluation criteria, these values suggest that the ensemble model results were more accurate in assessing the habitat suitability for leopards and wolves. Furthermore, the prediction accuracy of the GLM was higher than that of the MaxEnt as observed in Table 1.
3.2. Environmental variable importance analysis
The importance analysis of environmental variables using the ensemble model revealed that for suitable leopard habitats, the two most important variables were DTS and LC. Among these variables, DTS had the highest importance, contributing 84% to the suitability of the leopard habitat (Figure 2). On the other hand, for suitable wolf habitats, the two most important variables were SLP and DTC. Among these variables, SLP had the highest importance, contributing 92% to the suitability of wolf habitat (Figure 2).
By comparing the occurrence probabilities of leopards and wolves with three important environmental variables, SLP, DTC, and DTS (Figure 3), it becomes evident that both leopards and wolves exhibit preferences for areas with lower slopes and closer proximity to settlements. However, there are differences in their responses to these variables. Leopards show a stronger response to the distance from settlements, indicating a preference for areas that are farther away from settlements. On the other hand, wolves exhibit a stronger response to the slope, indicating a preference for areas with gentler slopes. Regarding the distance from the cliff, leopards and wolves demonstrate contrasting responses. Leopards show a preference for areas that are closer to cliffs, whereas wolves prefer areas that are farther away from cliffs.
3.3. Habitat suitability evaluation
The study area covered a total area of approximately 9127 km². The suitable habitat for leopards occupied an area of about 2111 km², accounting for 23% of the total study area. The suitable habitat was predominantly distributed along both banks of the Yalong River and extended to the surrounding areas (Figure 4A). Conversely, the unsuitable habitat covered an area of about 7016 km², representing 77% of the total study area. It was primarily located in the eastern and western parts of the study area (Figure 4A). For wolves, the suitable habitat accounted for 56% of the total study area, covering an area of approximately 5142 km². It was mainly distributed in the northwest, southwest, and east regions of the study area (Figure 4B). On the other hand, the unsuitable habitat occupied about 3985 km², making up 44% of the total study area. It was concentrated in the southeastern part of the study area (Figure 4B).
Overall, the suitable habitat for leopards was smaller in comparison to that for wolves within the study area, and the distribution of their major suitable habitats tended to be separate.
3.4. Habitat suitability landscape evaluation
The landscape pattern of the study area was assessed using Fragstats 4.2 software, which analyzed several landscape metrics including PLAND, PD, ED, LSI, AI, CONNECT, and PAFRAC. The findings are summarized as follows (Table 2): 1) Area Index: The PLAND of wolves (64.83%) was nearly four times larger than that of leopards (13.76%), indicating a greater extent of suitable habitat patches for wolves compared to leopards. 2) Fragmentation Index: The number of patches per unit area of suitable habitat was 0.35/km² for leopards and 1.17/km² for wolves, suggesting a higher degree of fragmentation in the suitable habitat of wolves than leopards. 3) Shape Index: Both the ED and LSI values were higher for wolves (ED: 41.35 m/ha, LSI: 126.82) than for leopards (ED: 5.78 m/ha, LSI: 41.17), indicating a more irregular and complex shape of suitable habitats for wolves. 4) Patch Aggregation Index: Both leopards and wolves exhibited high aggregation of suitable habitats, with values close to 100%. 5) Habitat Connectivity Index (threshold is 500m): The connectivity of suitable habitats for leopards was high (0.99), while it was significantly lower for wolves (0.06), suggesting limited connectivity between suitable habitat patches for wolves. 6) Disturbance Index: The landscape shape complexity of suitable habitats for both leopards and wolves was moderate, as indicated by the PAFRAC values (leopard: 1.35, wolf: 1.44).
4. Discussion
Large carnivores hold a crucial role as keystone species within terrestrial ecosystems, contributing significantly to the maintenance of ecosystem stability and biodiversity [1]. Interspecific competition among large carnivores of sympatric distribution can be more intense, with the result that vulnerable populations may be reduced or even extinct [21]. Large carnivores tend to spatially avoid one another to mitigate such outcomes [2]. Gaining a comprehensive understanding of the variations in suitable habitat selection among large carnivores is crucial for comprehending multi-species competition and coexistence. It also enhances our knowledge of their ecological behaviors, adaptations, and interactions with the environment. Such insights are vital for effective conservation and management strategies. We reveal the following insights: 1) The suitable habitat distribution for leopards and wolves in Xinlong County exhibits minimal overlap, indicating significant differences in their preferred habitats. 2) Both species exhibit a preference for areas characterized by gentle slopes and proximity to settlements. However, leopards are predominantly influenced by the distance from settlements, while wolves are primarily influenced by slope gradients. 3) Suitable habitats for both species demonstrate an overlap distribution pattern. Although the area of suitable habitat for wolves is greater than that of leopards, the fragmentation of suitable habitats and the complexity of patch shapes are more pronounced for wolves. These findings suggest that human activities have a more significant impact on wolves, despite their larger habitat area.
4.1. Divergent suitable habitats selection
The Leopards exhibit a preference for selecting habitats in the understory of forests, while wolves tend to favor grassland areas. This differential habitat selection could be driven by various factors, including interspecific competition, differences in prey distribution, and intrinsic differences. 1) Interspecific competition. Competition among large carnivores encompasses exploitative competition, which involves the utilization of shared resources, and interference competition, which entails direct interference or aggression. Prey resources in Xinlong County are abundant [13]. When prey resources are abundant, interference competition plays a stronger role in regulating population dynamics compared to exploitative competition [22]. In addition, the home range sizes of leopards and wolves are closely related to interspecific competition. Our results only indicate the distribution area of suitable habitat for two species in Xinlong County, which is approximately 2111 km2 and 5142 km2, respectively. Taking into account existing studies, the typical home range size for leopards ranges from 20-200 km2, while for wolves, it is generally between 500-8300 km2 [23,24,25,26,27]. We can predict that the suitable habitats in Xinlong County can support approximately 10-105 leopards and 1-10 wolves, respectively. However, based on infrared camera monitoring data, wolves have been observed multiple times at various camera sites, and residents have reported severe predation of domestic yaks by wolves. This suggests that the wolf population in Xinlong County is likely to be much larger than 10 individuals. There may be strong interspecies competition between leopards and wolves in Xinlong County, leading them to prefer different suitable habitats to reduce competition intensity. Several studies have found that adult leopards prefer understory areas with high vegetation cover and avoid scrub meadows [28,29,30], while wolves usually prefer areas with a high cover of tall grass or scrub vegetation and avoid understory areas [31,32,33]. Our results also further support the existing conclusions. 2) Differences in prey resource distribution. Studies have revealed significant dietary differences between leopards and wolves in Xinlong County and Shiqu County, where leopards primarily prey on the Chinese goral (Naemorhedus griseus) while wolves primarily target domestic yaks [34]. Chinese gorals tend to inhabit the understory of forests in Xinlong, while domestic yaks are mainly raised in alpine meadows. Therefore, the differential distribution of prey resources is likely a key factor driving the divergent habitat selection between wolves and leopards. 3) Intrinsic differences. The coat patterns of leopards are intricately connected to their natural environment. Research has revealed a correlation between the dappled patterns on a leopard's body and the patches of sunlight filtering through the vegetation, aiding their camouflage during forest hunting [35]. Similarly, the coat color of wolves can vary depending on the season and geographic location. In mountainous grassland regions, wolves often exhibit gray or brown fur, providing effective concealment [36]. Leopards possess exceptional visual acuity, enabling them to keenly detect potential prey. They are skilled climbers and jumpers, well-suited for ambushing prey in forested areas. In contrast, wolves possess a highly developed sense of smell and excel at tracking prey. They are adapted for endurance running and adept at navigating vast wilderness areas in search of prey [11,37]. 4) Differences in living habits. Leopards usually live alone while wolves often hunt in groups.
4.2. Divergence in main environmental factors
The main environmental factors driving habitat selection differ between wolves and leopards. The preference of wolves for areas closer to human settlements may be attributed to the fact that their primary prey, domestic yaks, are commonly found in proximity to human settlements. On the other hand, the leopards contradict the findings of studies on Amur leopards, which show a tendency to select habitats far away from roads and human habitation [38]. This could be attributed to the fact that the Tibetan residents in Xinlong County adhere to a belief system that prohibits killing animals. As a result, leopards in the vicinity of human settlements are almost unaffected by human hunting threats. Additionally, some prey species tend to appear near human settlements because they find abundant food sources in the crops cultivated by the residents, apart from being free from human hunting threats [39]. Both leopards and wolves prefer areas with lower slopes. Gentle slopes are generally more conducive for movement and provide a relatively open field of view, which facilitates prey detection. Compared to leopards, wolves are more affected by slope conditions (Figure 2), indicating a higher preference for areas with lower slopes. This preference aligns with the fact that wolves prefer grassland habitats and are well-adapted for running and hunting [31,32,33]。
4.3. Habitats fragmentation of wolf's near settlements
Habitat fragmentation can disrupt the balance of ecosystems by reducing biodiversity [8]. The suitable habitat for wolves exhibits higher fragmentation than that of leopards, with low connectivity and complex habitat shapes. This could be attributed to significant human influence. Referring to the evaluation map of habitat suitability levels (Figure 4), the areas with high habitat fragmentation for wolves primarily overlap with the suitable habitat distribution of leopards and coincide with the main residential areas. In Tibetan culture, leopards are generally associated with auspicious meanings, while wolves are more frequently involved in predation incidents on free-ranging livestock. Therefore, there may be differences in local tolerance towards leopards and wolves, leading to a greater aversion towards wolves. Additionally, since the implementation of natural forest protection projects, measures such as the prohibition of large-scale logging, reduced infrastructure development, and forest conservation have contributed to the recovery and protection of the forest habitat preferred by leopards. This may have facilitated population growth among leopards to some extent. Consequently, the wolves present in the region not only face interspecies suppression from leopards but also potentially encounter stronger human disturbances. These factors contribute to the higher level of habitat fragmentation observed in the suitable habitat for wolves compared to that of leopards.
4.4. Future work
Understanding the differences in habitat selection between leopards and wolves and their responses to environmental factors is of great significance for understanding large carnivore competition and coexistence and for developing scientific conservation management measures. The present study implies that large carnivores, leopards, and wolves, promote the differentiation of spatial ecological niches through differences in suitable habitat selection. However, this research has not yet considered the influence of prey resources on the habitat selection differences between the two species. Subsequent studies will further investigate the impact of prey resources in Xinlong County on the coexistence of carnivores, such as examining the differences in dietary preferences between the two species. This will provide valuable insights into the interplay between habitat selection and prey availability, shedding light on the mechanisms underlying their coexistence and facilitating more comprehensive conservation strategies.
Author Contributions
Conceptualization, T.T., X.C., and B.Y.; methodology, T.T., X.C, and H.P.; formal analysis, T.T., X.C, and H.P.; investigation, T.T., X.C., Y.J., X.Z., Y.X., and D.S.; writing—original draft preparation, T.T.; writing—review and editing, X.C., B.Y., and L.Z.; visualization, T.T., X.C, and H.P.; resources, X.C., B.Y, and L.Z.; data curation, T.T., and X.C.; funding acquisition, B.Y., and L.Z.; project administration, X.C., and Y.J.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was founded by the Ant Group and China Environmental Protection Foundation.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The work received financial support from the Ant Group and China Environmental Protection Foundation. We thank the support from the Sichuan Protected Areas Station and the Sichuan Wildlife Investigation and Protection Station.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix 1

Table S1.
Evaluation Indicators of Habitat Landscape Characteristics and Their Ecological Significance.
Table S1.
Evaluation Indicators of Habitat Landscape Characteristics and Their Ecological Significance.
| Index type | Exponential type | Meaning |
|---|---|---|
| Area index | Percentage of landscape (PLAND) | is the percentage (%) of the total area of a certain patch type in the entire landscape area. |
| Shape index | Landscape Shape Index(LSI) | The LSI value calculated the deviation degree of patch edge shape from a circle or square in the landscape. The closer the value was to 1, the closer the patch shape was to a circle or square. |
| Edge Density(ED) | Reflects the degree of landscape fragmentation, the larger the value, the greater the degree of fragmentation. | |
| Perimeter-Area Fractal Dimension(PAFRAC) | Reflecting the complexity of the landscape shape, 1≤PAFRAC≤2, the closer it tends to 1, the simpler the landscape shape is, and it may be less disturbed by humans. | |
| Connectivity index | Connectance index(CONNECT) | Reflecting the connectivity between patch types, the higher the value, the higher the landscape connectivity. |
| Aggregation index(AI) | Reflects the aggregation between patches of the same type, the smaller the value, the more discrete the landscape. | |
| Breakage index | Patch Density(PD) | Indicates the number of patches per unit area, reflecting the fragmentation of the landscape. |
Table S2.
Description of environmental variables in the Ensemble model.
| Variable | Code | Description | Variable types |
|---|---|---|---|
| Vegetation factor | EVI | Enhanced vegetation index | Continuous |
| Topographic factor | ELE | Elevation | Continuous |
| SLP | Slope | Continuous | |
| DTW | Distance to water | Continuous | |
| TRI | Terrain roughness index | Continuous | |
| DTC | Distance to cliffs | Continuous | |
| DTV | Distance to valleys | Continuous | |
| DTR | Distance to ridges | Continuous | |
| Disturbance factor | DTD | Distance to roads | Continuous |
| DTS | Distance to settlements | Continuous | |
| Other variables | LC | Land Cover | Categorical |
References
- Ripple, W.J.; Estes, J.A.; Beschta, R.L.; Wilmers, C.C.; Ritchie, E.G.; Hebblewhite, M.; Berger, J.; Elmhagen, B.; Letnic, M.; Nelson, M.P.; et al. Status and Ecological Effects of the World’s Largest Carnivores. Science 2014, 343, 1241484. [Google Scholar] [CrossRef]
- Li, Z.; Duo, L.; Li, S.; Wang, T. Competition and coexistence among terrestrial mammalian carnivores. Biodivers. Sci. 2021, 29, 81–97. [Google Scholar] [CrossRef]
- Hardin, G. The Competitive Exclusion Principle: An Idea That Took a Century to Be Born Has Implications in Ecology, Economics, and Genetics. Science 1960, 131, 1292–1297. [Google Scholar] [CrossRef] [PubMed]
- Odden, M.; Wegge, P.; Fredriksen, T. Do Tigers Displace Leopards? If so Why? Ecol Res. Ecol. Res. 2010, 25, 875–881. [Google Scholar] [CrossRef]
- Fennell, J.G.; Talas, L.; Baddeley, R.J.; Cuthill, I.C.; Scott-Samuel, N.E. Optimizing Colour for Camouflage and Visibility Using Deep Learning: The Effects of the Environment and the Observer’s Visual System. J. R. Soc. Interface. 2019, 16, 20190183. [Google Scholar] [CrossRef] [PubMed]
- Shahsavarzadeh, R.; Hemami, M.-R.; Farhadinia, M.S.; Fakheran, S.; Ahmadi, M. Spatially Heterogeneous Habitat Use across Distinct Biogeographic Regions in a Wide-Ranging Predator, the Persian Leopard. Biodivers. Conserv. 2023, 32, 2037–2053. [Google Scholar] [CrossRef]
- Ngoprasert, D.; Lynam, A.J.; Gale, G.A. Human Disturbance Affects Habitat Use and Behaviour of Asiatic Leopard Panthera Pardus in Kaeng Krachan National Park, Thailand. Oryx 2007, 41, 343–351. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat Fragmentation and Its Lasting Impact on Earth’s Ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef]
- De Lima Filho, J.A.; Vieira, R.J.A.G.; De Souza, C.A.M.; Ferreira, F.F.; De Oliveira, V.M. Effects of Habitat Fragmentation on Biodiversity Patterns of Ecosystems with Resource Competition. Physica. A. 2021, 564, 125497. [Google Scholar] [CrossRef]
- Ahmadi, M.; Balouchi, B.N.; Jowkar, H.; Hemami, M.-R.; Fadakar, D.; Malakouti-Khah, S.; Ostrowski, S. Combining Landscape Suitability and Habitat Connectivity to Conserve the Last Surviving Population of Cheetah in Asia. Divers. Distrib. 2017, 23, 592–603. [Google Scholar]
- Liu, X. Overlapping dietary and food ecological niches of three species of canids in eastern Inner Mongolia. Master thesis, Qufu Normal University, China, 2009. [Google Scholar]
- Zafar-ul Islam, M.; Volmer, R.; Al Boug, A.; Shehri, A.A.; Gavashelishvili, A. Modelling the Effect of Competition for Prey and Poaching on the Population of the Arabian Leopard, Panthera Pardus Nimr, in Saudi Arabia (Mammalia: Felidae). Zool. Middle East 2020, 66, 95–106. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, B.; Gu, X.; Song, D.; Yang, B. Camera Trapping Reveals Area of Conservation Significance for Large and Medium-Sized Mammals on the Eastern Tibetan Plateau. Oryx 2022, 56, 137–144. [Google Scholar] [CrossRef]
- Qiao, J. Distribution and behavior of leopard (Panthera pardus), snow leopard (Panthera unica) and gray wolf (Canis lupus) in Gongga Mountain, Sichuan Province. Master thesis, Sichuan Agricultural University, China, 2022. [Google Scholar]
- Kamino, L.H.Y.; Stehmann, J.R.; Amaral, S.; De Marco, P.; Rangel, T.F.; de Siqueira, M.F.; De Giovanni, R.; Hortal, J. Challenges and Perspectives for Species Distribution Modelling in the Neotropics. Biol. Letters 2011, 8, 324–326. [Google Scholar] [CrossRef]
- Liu, Z.; Gao, H.; Teng, L.; Su, Y.; Wang, X.; Kong, Y. Habitat suitability assessment of blue sheep in Helan Mountain based on MAXENT modeling. Acta Ecol. Sin. 2013, 33, 7243–7249. [Google Scholar] [CrossRef]
- Qi, Z.; Xu, W.; Xiong, X.; Ou, Y.; Zheng, H.; Gan, D. Assessment of potential habitat for Ursus thibetanus in the Qinling Mountains. Biodivers. Sci. 2011, 19, 343–352. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, Q.; Wang, B.; Gong, X.; Duan, C.; Zhang, Y. The habitat suitability evaluation of Salweenia bouffordiana based on MaxEnt model. Acta Ecol. Sin. 2020, 40, 6077–6085. [Google Scholar] [CrossRef]
- Brito, J.C.; Crespo, E.G.; Paulo, O.S. Modelling Wildlife Distributions: Logistic Multiple Regression vs Overlap Analysis. Ecography 1999, 22, 251–260. [Google Scholar]
- Swets, J. Measuring the Accuracy of Diagnostic Systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Interspecific Killing among Mammalian Carnivores. [CrossRef]
- Linnell, J.D.C.; Strand, O. Interference Interactions, Co-Existence and Conservation of Mammalian Carnivores. Divers. Distrib. 2000, 6, 169–176. [Google Scholar] [CrossRef]
- Snider, M.H.; Athreya, V.R.; Balme, G.A.; Bidner, L.R.; Farhadinia, M.S.; Fattebert, J.; Gompper, M.E.; Gubbi, S.; Hunter, L.T.B.; Isbell, L.A.; et al. Home Range Variation in Leopards Living across the Human Density Gradient. J. Mammal. 2021, 102, 1138–1148. [Google Scholar] [CrossRef]
- Gwynn, V.; Symeonakis, E. Rule-Based Habitat Suitability Modelling for the Reintroduction of the Grey Wolf (Canis lupus) in Scotland. PLoS ONE 2022, 17, e0265293. [Google Scholar] [CrossRef] [PubMed]
- Mondal, K.; Bhattacharjee, S.; Gupta, S.; Sankar, K.; Qureshi, Q. Home Range and Resource Selection of “problem” Leopards Trans-Located to Forested Habitat. Curr. Sci. India. 2013, 105, 338–345. [Google Scholar]
- Nams, V.O.; Parker, D.M.; Weise, F.J.; Patterson, B.D.; Buij, R.; Radloff, F.G.T.; Vanak, A.T.; Tumenta, P.N.; Hayward, M.W.; Swanepoel, L.H.; et al. Spatial Patterns of Large African Cats: A Large-scale Study on Density, Home Range Size, and Home Range Overlap of Lions Panthera Leo and Leopards Panthera Pardus. Mammal Rev. 2023, 53, 49–64. [Google Scholar] [CrossRef]
- Bryce, C.M.; Dunford, C.E.; Pagano, A.M.; Wang, Y.; Borg, B.L.; Arthur, S.M.; Williams, T.M. Environmental Correlates of Activity and Energetics in a Wide-Ranging Social Carnivore. Anim. Biotelem. 2022, 10, 1. [Google Scholar] [CrossRef]
- Zhou, Z. Periodic habitat utilization and its driving factors of North China Leopard. Master thesis, Northeast Forestry University, China, 2022. [Google Scholar]
- Lovari, S.; Ventimiglia, M.; Minder, I. Food Habits of Two Leopard Species, Competition, Climate Change and Upper Treeline: A Way to the Decrease of an Endangered Species? Ethol. Ecol. Evol. 2013, 25, 305–318. [Google Scholar] [CrossRef]
- Wei, W. The suitability evaluation of Amur Tiger ( Panthera tigris altaica) and Amur Leopard( Panthera pardus orientalis) habitat based on remote sensing and GIS. Master thesis, JinLin University, China, 2019. [Google Scholar]
- Han, L. Habitat selectivities and seasonal variations of wolves(Canis lupus) in Mount Kalamaili Ungulate Nature Reserve, Xinjiang,China. Master thesis, Xinjiang University, China, 2015. [Google Scholar]
- Wang, Y. Habitats selection during wintering of the steppe wolf (Canis lupus) in eastern Inner Mongolia. Master thesis, Qufu Normal University, China, 2007. [Google Scholar]
- Zhang, H.; Zhou, Y.; Dou, H.; Liu, X.; Zhang, C.; Liu, S. Prelimnary analysis on the habitat selection of wolf (Canis lupus) in Dalai Lake Natural Reserve in Inner Mongolia. Acta Ecol. Sin. 2010, 30, 3637–3644. [Google Scholar]
- Xinning, S.; Dazhao, S.; Qiaowen, H.; Sheng, L.; Meng, Y. Fast Surveys and Molecular Diet Analysis of Carnivores Based on Fecal DNA and Metabarcoding. Biodivers. Sci. 2019, 27, 543. [Google Scholar] [CrossRef]
- Allen, W.L.; Cuthill, I.C.; Scott-Samuel, N.E.; Baddeley, R. Why the Leopard Got Its Spots: Relating Pattern Development to Ecology in Felids. P. Roy. Soc. B-Biol. Sci. 2010, 278, 1373–1380. [Google Scholar] [CrossRef]
- Bannasch, D.L.; Kaelin, C.B.; Letko, A.; Loechel, R.; Hug, P.; Jagannathan, V.; Henkel, J.; Roosje, P.; Hytonen, M.K.; Lohi, H.; et al. Dog Colour Patterns Explained by Modular Promoters of Ancient Canid Origin. Nat. Ecol. Evol. 2021, 5, 1415. [Google Scholar] [CrossRef]
- Botts, R.T.; Eppert, A.A.; Wiegman, T.J.; Blankenship, S.R.; Rodriguez, A.; Wagner, A.P.; Ullrich, S.E.; Allen, G.R.; Garley, W.M.; Asselin, E.M.; et al. Does Moonlight Increase Predation Risk for Elusive Mammals in Costa Rica? Trop. Conserv. Sci. 2020, 13, 194008292095240. [Google Scholar] [CrossRef]
- Li, Z.; Wang, T.; Smith, J.L.D.; Feng, R.; Feng, L.; Mou, P.; Ge, J. Coexistence of Two Sympatric Flagship Carnivores in the Human-Dominated Forest Landscapes of Northeast Asia. Landscape Ecol. 2019, 34, 291–305. [Google Scholar] [CrossRef]
- Wang, J. Seasonal differences in prey abundance and food composition of the North China leopard (Panthera pardus fontanierii) in Tieqiaoshan Nature Reserve, Shanxi. Master thesis, Northeast Forestry University, China, 2020. [Google Scholar]
Figure 1.
a. Global distribution range of leopards and wolves; b. The location of Xinlong region in the Mountains of Southwest China; c. Distribution of camera-trapping in Xinlong County.
Figure 1.
a. Global distribution range of leopards and wolves; b. The location of Xinlong region in the Mountains of Southwest China; c. Distribution of camera-trapping in Xinlong County.
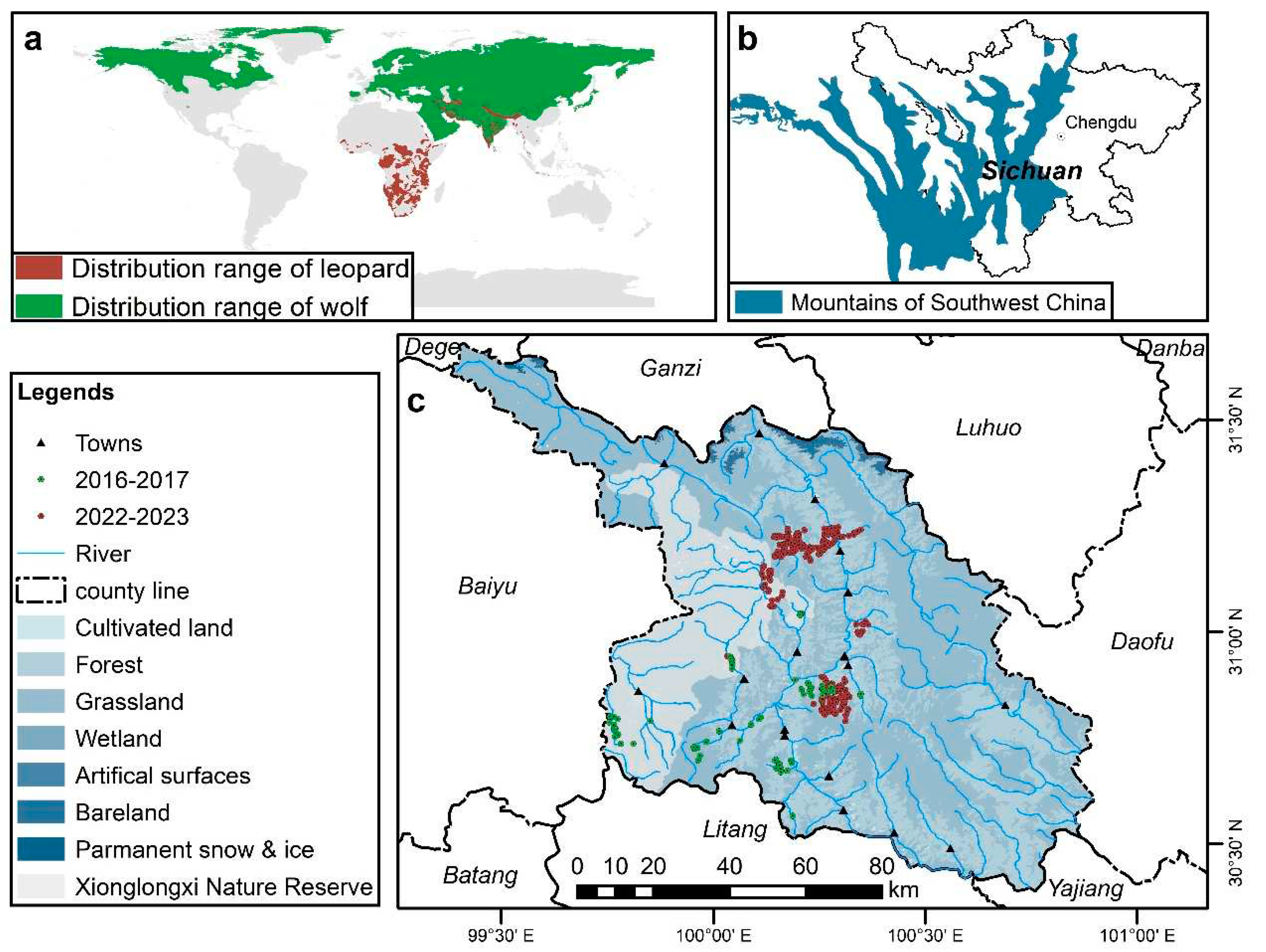
Figure 2.
Importance of environmental variables for suitable habitat. A pink solid circle represents leopards, while blue represents wolves. The size of the circle corresponds to the magnitude of the value, with larger circles indicating higher values.
Figure 2.
Importance of environmental variables for suitable habitat. A pink solid circle represents leopards, while blue represents wolves. The size of the circle corresponds to the magnitude of the value, with larger circles indicating higher values.
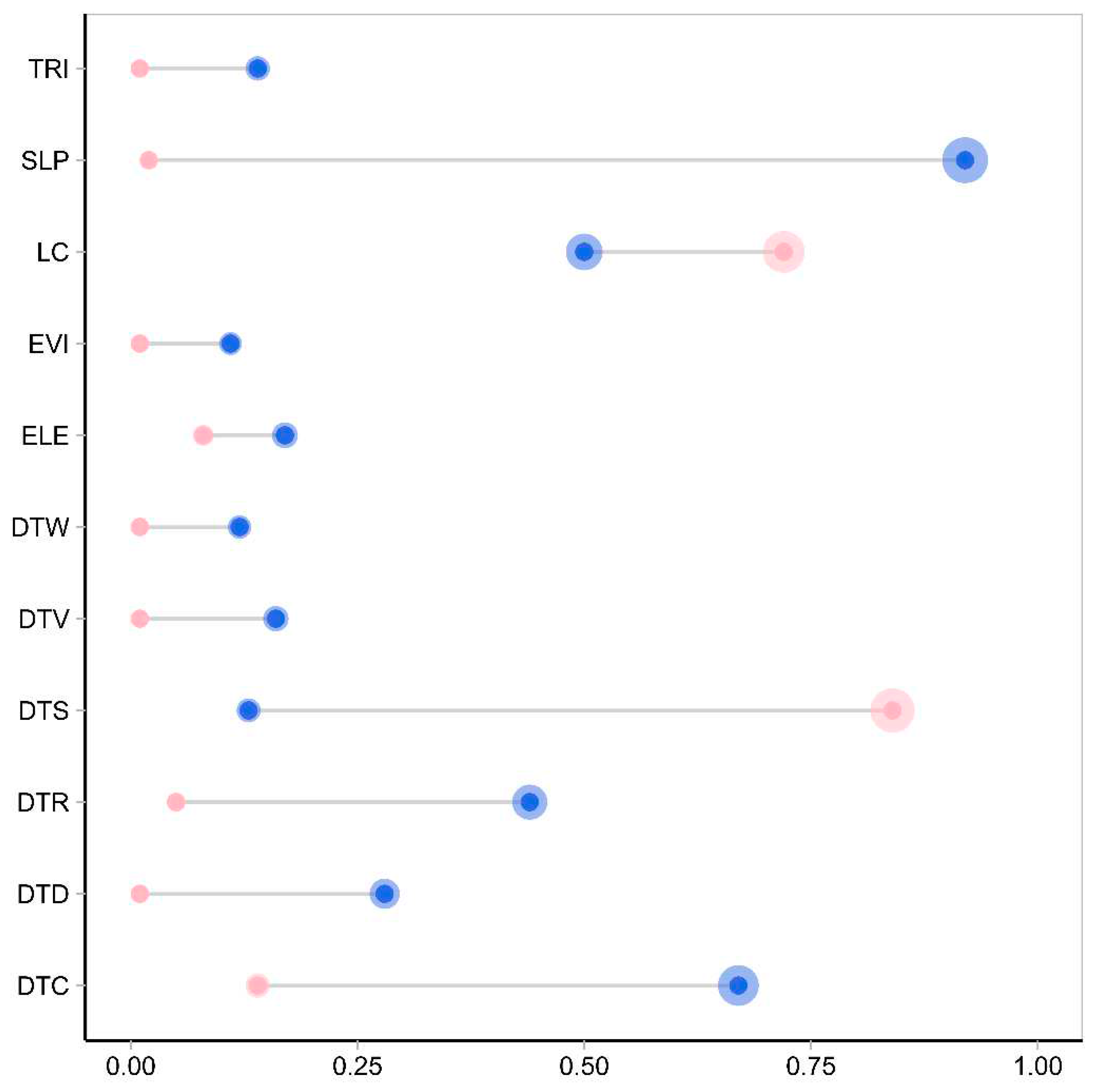
Figure 3.
Leopards (above) and wolfs (below) environmental response curves. From left to right: SLP, DTC, and DTS. The vertical axis represents the response values, while the horizontal axis represents the slope (left panel) and distance (middle and right panels). Gray-shaded areas represent 95% confidence intervals.
Figure 3.
Leopards (above) and wolfs (below) environmental response curves. From left to right: SLP, DTC, and DTS. The vertical axis represents the response values, while the horizontal axis represents the slope (left panel) and distance (middle and right panels). Gray-shaded areas represent 95% confidence intervals.
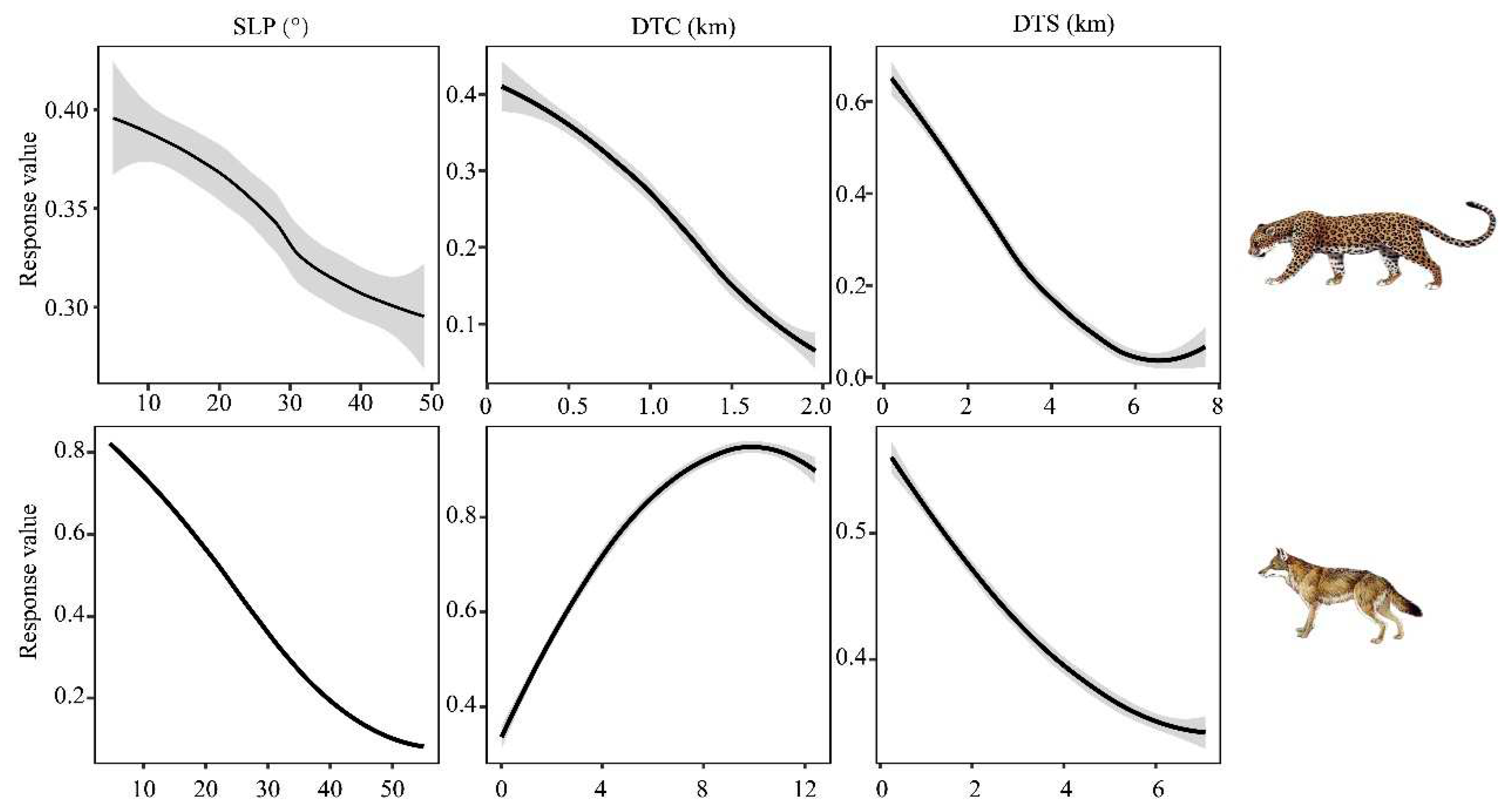
Figure 4.
Distribution of habitat suitability for leopards (A) and wolves (B) in Xinlong County. Gray-red for suitable habitat distribution and gray-blue for unsuitable habitat distribution.
Figure 4.
Distribution of habitat suitability for leopards (A) and wolves (B) in Xinlong County. Gray-red for suitable habitat distribution and gray-blue for unsuitable habitat distribution.
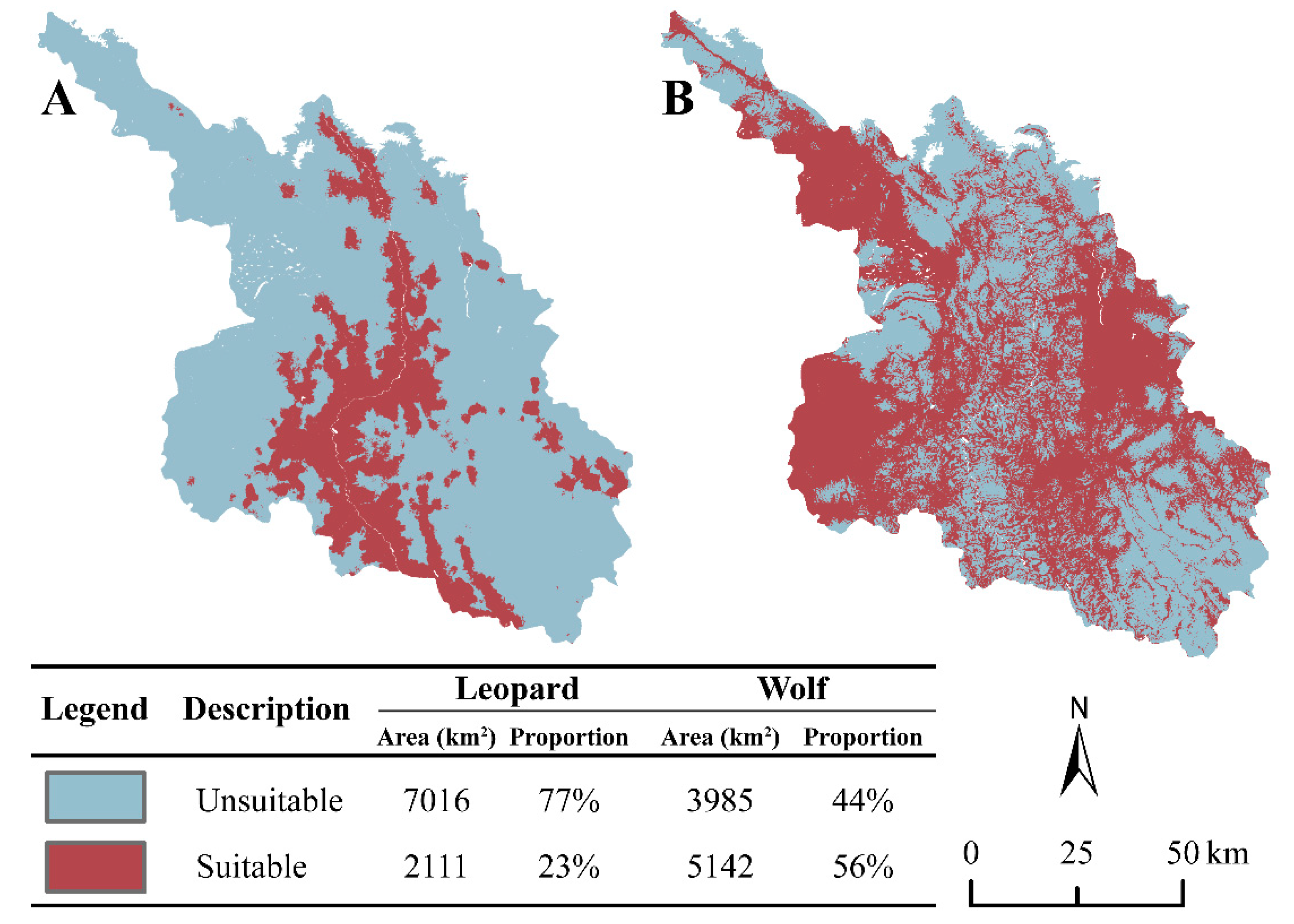
Table 1.
AUC Values of Different Species Distribution Models.
| Species | GLM | MaxEnt | Ensemble model |
|---|---|---|---|
| Leopard | 0.84 | 0.76 | 0.81 |
| Wolf | 0.86 | 0.73 | 0.82 |
Table 2.
Evaluation indices of landscape characteristics of suitable habitats for leopards and wolves.
Table 2.
Evaluation indices of landscape characteristics of suitable habitats for leopards and wolves.
| Species | PLAND | PD | ED | LSI | AI | CONNECT | PAFRAC |
|---|---|---|---|---|---|---|---|
| Leopard | 13.76 | 0.35 | 5.78 | 41.17 | 96.60 | 0.99 | 1.35 |
| Wolf | 64.83 | 1.17 | 41.35 | 126.82 | 95.09 | 0.06 | 1.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



