Submitted:
21 July 2023
Posted:
24 July 2023
You are already at the latest version
Abstract
Round bodies in spirochete cultures have been a controversial subject since their description seven decades ago. We report the existence of round bodies (spherical cells) in cultures of Mucispirillum schaedleri, a spiral bacterium phylogenetically distant from spirochetes. Furthermore, when grown in biofilms, M. schaedleri demonstrates a unique morphology known as cording, which has been previously described only in mycobacteria. Thus, M. schaedleri has two distinct features, each previously thought to be unique to two different phylogenetically distant groups of bacteria.
Keywords:
Mucispirillum schaedleri
; pleomorphism
; biofilm
; round body
; spirochete
; cording
; mycobacteria
1. Introduction
The Mucisprillum schaedleri (Deferribacteraceae) is a spiral-shaped, flagellated, Gram negative, obligately anaerobic bacterium that was first described in 1987 as “a spirochete”[1]. Its original designation, Altered Schaedler Flora strain 457 (ASF 457) relates to its inclusion in commercially supplied consortia used to establish putatively standardized bacterial communities in gnotobiotic mice [2,3]. A decade later, unclassified murine gut isolates similar to ASF 457 (but not then recognized as such) were obtained during a thesis project [4]; 5 had novel 16S rRNA gene sequences most closely related to Geovibrio and Deferribacter. Shortly thereafter, a molecular phylogenetic study on the Altered Schaedler Flora found the 16S rRNA gene sequence of ASF 457 to be closely related to those of the unclassified murine isolates [5]. Six years on, 7 additional murine isolates were obtained that had 16S rRNA gene sequences differing by maximally 4 nucleotides, and 4 had 100% similarity to ASF 457. Based on the close phylogenetic relationships and certain phenotypic features, the designation Mucispirillum schaedleri was proposed [6]. The type strain (HRI |17) was deposited in the Australian Collection of Microorganisms (ACM 5223) and in the American Type Culture Collection (ATCC BAA-1009), but it is no longer available. Two variants of ASF 457 referred to as AYGZ [7] and MCS [8] have been reported, and a variant named SH1 (likely to be MCS) is available from the German Collection of Microorganisms and Cell Cultures (DMSZ). The strains have few genetic differences [8] and phenotypic differences have yet to be reported. Published microscopy is limited to an electron microscopy micrograph [6] and fluorescent in situ hybridization (FISH) documentation in artificially colonized mice [8,9].
Murine intestinal samples are enriched in M. schaedleri in the context of infection [10], as well as in genetic [11,12,13] and chemical [14] colitis models. Intestinal enrichment has also been described in other murine models including stress [15], high-fat diet [16] and rheumatoid arthritis [17]. Importantly, molecular data suggest that M. schaedleri colonizes the nasal, vaginal and GI mucosae of humans, albeit at a lower prevalence than in mice or other sources[18]. M. schaedleri abundance was found to be increased in a human study of Parkinson’s disease [19]. Enrichment may relate to the ability of M. schaedleri to reduce nitrate [8], which is abundant in inflammatory conditions [18]. It harbors only a few genes encoding glycan-degrading enzymes; instead it utilizes monosaccharides, amino acids and short-chain fatty acids [8]. M. schaedleri also contains numerous genes associated with nitrate reduction, as well as genes for superoxide reductase, catalase and cytochrome c oxidase genes [8]: features that may facilitate colonization of mucosal surfaces in the presence of epithelium-derived reactive oxygen and nitrogen species [20,21].
Spirochetes, a group of Gram-negative anaerobic motile spiral bacteria unrelated to M. schaedleri, can show pleomorphic growth in which spherical bodies are intermixed with the typical spiral morphologies. Multiple terms appear in the literature (cysts, round bodies, spheroids, gemmae), some of which have been proven inapplicable (L-forms, cell-wall deficient forms); the term round body will be used here. Best studied in the Lyme disease pathogen Borrelia burgdorferi and in Treponema spp., round bodies contain rod-like cells appressed to the inner membrane of the sphere in a folded or looping manner. Their formation is thought to take place through the development of a bleb in the outer membrane of the typical spiral cell (frequently on the end) into which the cell moves by rolling up to fit within the diameter of the expanding bleb [22]. Electron micrographs show a large portion of the sphere’s diameter can be occupied by cellular cross-sections and such images can give the impression that multiple cells are sometimes present. Spheres can also contain amorphous material. Formation is reported to be induced by stressors (antibiotics, hypotonic conditions, changes in serum or complement in the medium and changes in oxygen tension) [23,24,25] which favor membrane blebbing and development of gaps between the outer membrane and cell wall. Round bodies are typically observed in greater numbers in older cultures [23,26]. They have been described as reproductive or resistant forms from which a single cell or multiple cells emerge [27,28] and they have been observed in vivo in a murine infection model [29]. Reviews of the Lyme disease literature have not shown a clear connection between round bodies and disease, but at the same time, they have noted a potential role for pleomorphy in infection and persistence [30,31].
The present study demonstrates that M. schaedleri produces round bodies identical to those observed in spirochetes and that M. schaedleri exhibits a striking in vitro biofilm morphology which mirrors cording in mycobacteria.
2. Materials and Methods
2.1. Bacterial strains and growth conditions
M. schaedleri ASF 457 (AYGZ variant) was kindly provided by Dr. Michael Wannemuehler (Iowa State University) and cultured on Akkermansia minimal medium (AMM) broth [8] containing 1% KNO3 in a Whitley A35 anaerobic chamber (Microbiology International; N2/CO2/H2 80/10/10, 37o C. Treponema denticola ATCC 35405 was grown in NOS broth [32] in the anaerobic chamber. GFP-expressing Mycobacterium bovis BCG [33] was grown in Middlebrook 7H9 base supplemented with 0.2% (v/v) glycerol at 37oC in a 3% CO2 incubator.
2.2. Initial microscopic characterization
All manipulations except centrifugation and microscopy were carried out in the anaerobic chamber, and all liquids were equilibrated in the chamber overnight. For initial observation of cells from culture, washed cells were resuspended in BacLight Live/Dead viability stain (Invitrogen) prepared in anaerobic phosphate-buffered saline (PBS) and examined by confocal microscopy (Leica SP8, Leica Microsystems). Excitation wavelength was 488 nm, and emission was collected simultaneously at 505-530 nm (Syto9 – “live”) and 600-630 nm (propidium iodide – “dead”) using a 100x (1.4 NA) oil-immersion lens. Optical section thickness and z-step settings were those recommended by the system software. Transmitted white-light micrographs of wet-mount preparations were obtained using a 100x (1.4 NA) oil-immersion lens on a Leica DM LB2 microscope with a Hamamatsu camera.
2.3. Transmission electron microscopy
Cell pellets from broth culture or loopfuls of material from AMM plates were fixed for 48 hrs at 4o C in 0.1M cacodylate buffer (pH 7.4) containing glutaraldehyde/paraformaldehyde (2.5%/1%), then washed 3 times with cacodylate buffer. The pellets were dehydrated in ethanol and propylene oxide, then embedded in EMBed 812 resins (Electron Microscopy Science). Thin sections (80 nm) were cut using an Ultracut-UCT ultramicrotome (Leica), placed onto 300-mesh copper grids, then stained with saturated uranyl acetate in 50% methanol followed by lead citrate. Grids were viewed with a JEM-1200EXII electron microscope (JEOL Ltd) at 80kV; images were recorded on a mid-mounted CCD camera (XR611M, 10.5 Mpixel; Advanced Microscopy Techniques Corp).
2.4. Biofilm experiments
For M. schaedleri biofilms, a 4-day-old broth-culture cell pellet was resuspended in AMM/NO3 to 0.05 OD600. Three microliters of the standardized cell suspension were added to wells of duplicate chamber slides (Ibidi LLC; µ-Slide 8 Well, untreated glass), each containing 300 µL of AMM/NO3. After 18 hrs, one set of slides was processed by removal of spent culture followed by 2 washes with 250 µL PBS. Two hundred microliters of BacLight Live/Dead stain were added to the biofilms and the slides were incubated for 15 min prior to confocal microscopy as described above. The second set of slides was processed the following day (ca. 48 hr incubation). In addition to high resolution images, six randomly selected fields-of-view around the center of the chamber were recorded using a 20x, 0.75 NA dry lens. For flowcell-grown biofilms, 2 mLs of the standardized suspension were injected into two µ-Slide I 0.8 glass-bottom chambers (Ibidi LLC) connected in series. After 60 min of attachment time, flow of AMM/NO3 was begun at 10 mls/hr (Ismatec Regalo ICC pump, Cole-Parmer). After 18 hrs, one flowcell was removed and slowly injected with 1 mL viability stain; the second flowcell was processed after 48 hrs incubation. High-resolution and low-magnification images were collected as for chamber biofilms. Imaris V9.7 (Bitplane AG) was used to compute biovolume, to produce maximum intensity projections, and to generate movies. For Treponema denticola biofilms, an overnight culture was diluted 10x into 300 µL fresh NOS in an Ibidi chamber slide. After 18 hrs of growth in the anaerobic chamber, the biofilm was examined using Live/Dead stain as above. For Mycobacterium bovis biofilms, an overnight culture was diluted 10x into 300 µL fresh medium in an Ibidi chamber slide. After 18 hrs of growth in the CO2 incubator, the pellicle biofilm at the air/liquid interface was lifted off using an inoculating loop, placed on a wetted microscope slide, and examined as above but using intracellular GFP fluorescence rather than LiveDead stain.
2.5. Enumeration of round bodies by microscopy
For experiments on temporal aspects of round body production, cell pellets from 3- or 6-day-old ASF 457 broth cultures were resuspended in AMM and incubated for 3 hrs at 37°C inside an anaerobic chamber. After incubation, cells were pelleted and resuspended in anaerobic PBS with BacLight Live/Dead stain. Culture ages investigated here correspond approximately to mid-log (3 days) and plateau (6 days) phases on a growth curve measured by OD8. Wet-mounted samples were visualized by confocal fluorescence microscopy using an LSM710 (Zeiss) with a 63x (1.4 NA) oil-immersion objective. The excitation/emission wavelengths for “live” (Syto9) and “dead” (propidium iodide; PI) stains were 488/531 nm and 534/616 nm, respectively. For each experimental condition, 9 scanned images (3048 x 3048 pixels, 269,0 µm x 269,0 µm) were taken and 4 independent experiments were performed (n=4). Raw image data were imported to Matlab in tiff format. Syto9 and PI tiff images were segmented using 3 labels from the Pixel Classification algorithm from ilastik (version 1.3.3Post3: https://www.ilastik.org/). Two labels were identified as the Syto9 and PI channels, and the background was set as the third channel. Objects found by segmentation were processed using MatLab (version 2020b: https://www.mathworks.com/). A first criteria of size was used to remove all objects having a size smaller than 0.4 µm2 (corresponding to 50 pixels). Objects were then selected based on their Eccentricity and Solidity parameters. Threshold values of Eccentricity lower than 0.9 and Solidity greater than 0.7 were used to identify round bodies, whereas objects not corresponding to these criteria were identified as rods. Finally, a mean intensity threshold value of 25 was used to reject faint round objects from the Syto9 channel (Supplementary Figure S1).
2.6. Statistical analyses
For 2-group comparisons, Mann-Whitney U was performed using Prism 9 (version 9.5.1) and significance was established at p<0.05.
3. Results
3.1. M. schaedleri ASF457 produces round bodies
M. schaedleri ASF 457 presented 3 prominent morphological features in static broth culture. First, rods were frequently found appressed to one another in an arrangement similar to that observed in, e.g., Capnocytophaga [34] (Figure 1 A-C). Second, round bodies were often attached to the end of rods and also distributed as single entities, in which case they were often larger than when attached to a rod. The round bodies typically contained cells appressed to the inner wall of the sphere, presenting either a c-shaped or spiral appearance (Figure 1 A, D, E). Electron microscopy confirmed that the cell inside the round body can be wound along the sphere’s wall, thereby giving the appearance of multiple cells, and that many round bodies had amorphous content (Figure 1 F, G).
For comparison, confocal micrographs of round bodies produced by Treponema denticola biofilms grown as part of the current study (Figure 1 H, I), and electron micrographs from a different study [35] on Borrellia burgdorferi (Figure 1 J, K) are shown. Third, image quantification analysis of broth-grown cultures demonstrated that the ratio of “live” round bodies to “live” rods increased as cultures aged (Figure 2 and Supplementary Table S1). In 3-day-old cultures, round bodies made up 20.3% of all cells counted and 15.3% of the round bodies were “live”. In 6-day-old cultures, 48% of counted cells were identified as round bodies, of which 36.5% were “live”. Regardless of culture age, more than 95% of rods were scored as “live”. Notably, the proportion of “live” rods was significantly decreased at 6 days (49.4%) compared to 3 days (76.8%) (p = 0.03), whereas the proportion of “live” round bodies was significantly increased at 6 days (36.5%) compared to 3 days (15.3%) (p = 0.03). These data are consistent with previously-described age-associated formation of round bodies in spirochete cultures [36,37].
3.2. M. schaedleri ASF 457 biofilms are composed of cords and contain round bodies.
M. schaedleri ASF 457 presented a striking biofilm morphology. When grown in AMM/NO3 medium, it displayed large rope-like structures resembling masses of the closely appressed cells seen in planktonic broth culture; these structures will be termed cords. After 48 hrs of growth, cords were much larger than after 18 hrs of growth (Figure 3 A-D), and surface coverage was much higher in the continuous culture system of flowcells than under static conditions in chamber-slides. The biofilm morphology overall is the same as cording documented in mycobacterial biofilms, here shown in M. bovis biofilms grown as part of the present study (Figure 3 E, F), as well as in M. marinum biofilms from different studies [38,39] (Figure 3 G). Image analysis (Figure 4) demonstrated that average biovolume per field-of-view in chambers increased 3-fold between 18 and 48 hrs (reaching 4.0x104 µm3) and maximum biofilm height (measured as the height at which cells were no longer visible in the microscope) increased 1.8-fold (reaching 28.2 µm). In contrast, biovolume in flowcell-grown biofilms increased 40-fold (reaching 4.3x105 µm3), and maximum height increased 2.4-fold (reaching 64.9 µm). At 18 hrs of growth, high standard deviation was apparent in average biovolume measurements for flowcell-grown biofilms, whereas the range of initial biovolumes in chamber-grown biofilms was comparatively narrow. However, by 48 hrs, flowcell-grown biofilms had reached confluence whereas chamber biofilms still had uncolonized areas (Figure 3).
A feature common to ASF 457 biofilms regardless of growth condition was the presence of round bodies identical to those in broth cultures. Round bodies were more prominent and more likely to be stained PI-positive in chamber biofilms than in flowcell biofilms; round bodies in flowcell-grown biofilms often appeared completely green or red rather than containing a green or red cell along the inner wall (Figure 5 and Supplementary Material – Videos S1-S4).
4. Discussion
The present study demonstrates three key findings: 1) M. schaedleri displays pleomorphic forms distinct from those reported for other gut bacteria, e.g., Helicobacter [40,41], but which are identical to round bodies described for spirochetes – these round bodies are seen in biofilms in vitro; 2) round bodies increase in M. schaedleri cultures in an age-dependent manner and many round bodies are considered as “live” in a live/dead viability assay; 3) M. schaedleri biofilms in vitro display the distinct mycobacterial morphology known as cording.
In T. denticola, round bodies occur not only in broth culture, but also in biofilms in vitro [42] (an observation that has been documented here as well; Figure 1). They likely also occur in biofilms of B. burgdorferi [35] and may be a common feature among all spirochetes. The present study reports an age-dependent increase in round bodies in M. schaedleri, which is also a characteristic of treponeme round bodies.
M. schaedleri biofilms displayed a corded morphology strikingly similar to that described in M. bovis [43](documented here as well; Figure 3) and in M. marinum [38,39]. In mycobacteria, genes known as cording factors are involved in mycolic acid synthesis/modification and in secretion (Type VII) [39,44]: both are important factors in the production of extracellular polymeric substances (i.e., biofilm matrix). Cording in vitro is typically studied using mycobacterial biofilms grown as pellicles at the air-liquid interface; an exception is cording described in rotary cell culture [43]. In contrast to pellicle biofilms, submerged mycobacterial biofilms have a non-corded, lichen-like morphology [39]. Cording occurs intracellularly (i.e., within the host) and is thought to impair host immune surveillance [44]. Round bodies have not been described in mycobacteria. In contrast to mycobacteria, M. schaedleri is an anaerobic commensal, does not form biofilms at the gas-liquid interface, and its genome does not contain cording factors8. Although the present study on M. schaedleri did not address the mechanism of cording nor its potential function in vivo, no cording or round bodies have been observed in sections of murine gut from gnotobiotic animals which have received rectal and oral gavages of M. schaedleri [8,18].
Viability staining demonstrated that in M. schaedleri biofilm cultures (particularly those formed in chamber slides), round bodies had reduced membrane integrity and/or transmembrane potential compared to rods, especially once the presence of an enclosed cell was difficult to verify. However, quantification of PI-stained round bodies in broth cultures demonstrated that many round bodies were viable, including at 6 days.
Biofilm culture in chamber slides is a batch culture procedure. In contrast, culture in flowcells is analogous to a fixed-bed reactor in which fresh medium is constantly supplied; nutrient depletion can occur only if biomass and cellular activity become so high that nutrients are exhausted during transit through the flowcell. The present experiments document that sufficient nutrients were available to support growth of M. schaedleri biofilms in chambers over the period of 18-48 hrs, albeit with a small (3-fold) increase in biovolume. During this time, PI-positive round bodies became more prominent. However, in flowcells, growth over the same period resulted in a 40-fold increase in biovolume, and PI-positive round bodies were less apparent; instead, round bodies lacking a c-shaped cell but staining positive with Syto 9 were observable. The homogenous staining of round bodies suggests that these may contain the amorphous material seen in electron microscopy but, in the case of Syto-positive cells, a membrane potential across the round body membrane remains. Such cells may also become PI positive if nutrients are no longer plentiful. Results from biofilm experiments suggest that, once formed, round bodies may be more susceptible to senescence than are rods during extended periods of nutrient depletion.
Scant evidence exists for the presence of round bodies in spirochete infections. Likewise, cording is not easily identifiable in mycobacterial infections. A function for these forms has yet to be identified in vitro. The presence of both forms in M. schaedleri presents an opportunity to examine the relevance of round bodies and of cording in a tractable model system such as in gnotobiotic mouse models.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: Identification of round bodies and rods based on Eccentricity and Solidity; Table S1: Frequencies and viability of rods and round bodies in 3- and 6-day-old cultures; Video S1: Chamber 20X; Video S2: Chamber 100X; Video S3: Flowcell 20X; Video S4: Flowcell 100X.
Author Contributions
Conceptualization, R.J.P. and E.L.F.; methodology, A.D., R.J.P., P.S., D.F..; software, D.F..; formal analysis, A.D., R.J.P..; investigation, A.D., R.J.P.; resources, R.J.P., E.L.F.; writing—original draft preparation, A.D., R.J.P., E.L.F.; writing—review and editing, A.D., D.F., R.J.P., E.L.F; visualization, A.D., D.F., R.J.P.; supervision, E.L.F.; funding acquisition, E.L.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a Canada Research Chair (Tier 2) in Role of the Microbiome in Primary Immune Deficiencies, Canadian Institutes of Health Research (CIHR), the Fonds de Recherche du Québec-Santé (FRQ-S) Clinician Scholar award, the J-Louis Lévesque Foundation Research Chair, and by the Intramural Research Program of the National Institute of Dental and Craniofacial Research (NIDCR) and the NIDCR Imaging Core ZIC DE000750-01.
Data Availability Statement
The method for identifying round bodies and rods based on Eccentricity and Solidity is shown in Supplementary Figure S1. Any additional microscopy images or extended biofilm movies will be made available upon request. MatLab script used to identify, and enumerate round bodies and rods by microscopy will be provided upon request.
Acknowledgments
We thank Dr. Michael Wannemuehler (Iowa State University) for providing Mucispirillum schaedleri ASF 457 variant AYGZ.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Orcutt, R.P.; Gianni, F.J.; Judge, R.J. Development Of An “Altered Schaedler Flora” For Nci Gnotobiotic Rodents. Microecology And Therapy 1987, 17, 59. [Google Scholar]
- Sarma-Rupavtarm, R.B.; Ge, Z.M.; Schauer, D.B.; Fox, J.G.; Polz, M.F. Spatial Distribution And Stability Of The Eight Microbial Species Of The Altered Schaedler Flora In The Mouse Gastrointestinal Tract. Applied And Environmental Microbiology 2004, 70, 2791–2800. [Google Scholar] [CrossRef] [PubMed]
- Wymore Brand, M.; Wannemuehler, M.J.; Phillips, G.J.; Proctor, A.; Overstreet, A.-M.; Jergens, A.E.; Orcutt, R.P.; Fox, J.G. The Altered Schaedler Flora: Continued Applications Of A Defined Murine Microbial Community. Ilar Journal 2015, 56, 169–178. [Google Scholar] [CrossRef]
- Robertson, B.R. The Molecular Phylogeny And Ecology Of Spiral Bacteria From The Mouse Gastrointestinal Tract. University Of New South Wales, Sydney, Australia, 1998. [Google Scholar]
- Dewhirst, F.E.; Chien, C.-C.; Paster, B.J.; Ericson, R.L.; Orcutt, R.P.; Schauer, D.B.; Fox, J.G. Phylogeny Of The Defined Murine Microbiota: Altered Schaedler Flora. Applied And Environmental Microbiology 1999, 65, 3287–3292. [Google Scholar] [CrossRef] [PubMed]
- Robertson, B.R.; O'rourke, J.L.; Neilan, B.A.; Vandamme, P.; On, S.L.W.; Fox, J.G.; Lee, A. Mucispirillum Schaedleri Gen. Nov., Sp Nov., A Spiral-Shaped Bacterium Colonizing The Mucus Layer Of The Gastrointestinal Tract Of Laboratory Rodents. International Journal Of Systematic And Evolutionary Microbiology 2005, 55, 1199–1204. [Google Scholar] [CrossRef]
- Wannemuehler, M.J.; Overstreet, A.-M.; Ward, D.V.; Phillips, G.J. Draft Genome Sequences Of The Altered Schaedler Flora, A Defined Bacterial Community From Gnotobiotic Mice. Genome Announcements 2014, 2, E00287-00214. [Google Scholar] [CrossRef]
- Loy, A.; Pfann, C.; Steinberger, M.; Hanson, B.; Herp, S.; Brugiroux, S.; Neto, J.C.G.; Boekschoten, M.V.; Schwab, C.; Urich, T.; et al. Lifestyle And Horizontal Gene Transfer-Mediated Evolution Of Mucispirillum Schaedleri, A Core Member Of The Murine Gut Microbiota. Msystems 2017, 2. [Google Scholar] [CrossRef]
- Herp, S.; Brugiroux, S.; Garzetti, D.; Ring, D.; Jochum, L.M.; Beutler, M.; Eberl, C.; Hussain, S.; Walter, S.; Gerlach, R.G.; et al. Mucispirillum Schaedleri Antagonizes Salmonella Virulence To Protect Mice Against Colitis. Cell Host & Microbe 2019, 25, 681–694.E688. [Google Scholar] [CrossRef]
- Belzer, C.; Gerber, G.K.; Roeselers, G.; Delaney, M.; Dubois, A.; Liu, Q.; Belavusava, V.; Yeliseyev, V.; Houseman, A.; Onderdonk, A.; et al. Dynamics Of The Microbiota In Response To Host Infection. Plos One 2014, 9, E95534. [Google Scholar] [CrossRef]
- Rooks, M.G.; Veiga, P.; Wardwell-Scott, L.H.; Tickle, T.; Segata, N.; Michaud, M.; Gallini, C.A.; Beal, C.; Van Hylckama-Vlieg, J.E.; Ballal, S.A.; et al. Gut Microbiome Composition And Function In Experimental Colitis During Active Disease And Treatment-Induced Remission. Isme J 2014, 8, 1403–1417. [Google Scholar] [CrossRef]
- Vereecke, L.; Vieira-Silva, S.; Billiet, T.; Van Es, J.H.; Mc Guire, C.; Slowicka, K.; Sze, M.; Van Den Born, M.; De Hertogh, G.; Clevers, H.; et al. A20 Controls Intestinal Homeostasis Through Cell-Specific Activities. Nat Commun 2014, 5, 5103. [Google Scholar] [CrossRef] [PubMed]
- Selvanantham, T.; Lin, Q.; Guo, C.X.; Surendra, A.; Fieve, S.; Escalante, N.K.; Guttman, D.S.; Streutker, C.J.; Robertson, S.J.; Philpott, D.J.; et al. Nkt Cell-Deficient Mice Harbor An Altered Microbiota That Fuels Intestinal Inflammation During Chemically Induced Colitis. J Immunol 2016, 197, 4464–4472. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.; Kuzyk, O.; Rauch, I.; Heider, S.; Schwab, C.; Hainzl, E.; Decker, T.; Muller, M.; Strobl, B.; Schleper, C.; et al. Intestinal Microbiota Signatures Associated With Inflammation History In Mice Experiencing Recurring Colitis. Front Microbiol 2015, 6, 1408. [Google Scholar] [CrossRef] [PubMed]
- Jasarevic, E.; Howard, C.D.; Misic, A.M.; Beiting, D.P.; Bale, T.L. Stress During Pregnancy Alters Temporal And Spatial Dynamics Of The Maternal And Offspring Microbiome In A Sex-Specific Manner. Sci Rep 2017, 7, 44182. [Google Scholar] [CrossRef]
- Ussar, S.; Griffin, N.W.; Bezy, O.; Fujisaka, S.; Vienberg, S.; Softic, S.; Deng, L.; Bry, L.; Gordon, J.I.; Kahn, C.R. Interactions Between Gut Microbiota, Host Genetics And Diet Modulate The Predisposition To Obesity And Metabolic Syndrome. Cell Metab 2015, 22, 516–530. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Fu, X.; Ni, Y.; Chen, J.; Jian, S.; Wang, L.; Li, L.; Du, G. Protective Effects Of Paederia Scandens Extract On Rheumatoid Arthritis Mouse Model By Modulating Gut Microbiota. J Ethnopharmacol 2018, 226, 97–104. [Google Scholar] [CrossRef]
- Herp, S.; Durai Raj, A.C.; Salvado Silva, M.; Woelfel, S.; Stecher, B. The Human Symbiont Mucispirillum Schaedleri: Causality In Health And Disease. Med Microbiol Immunol 2021, 210, 173–179. [Google Scholar] [CrossRef]
- Lin, C.H.; Chen, C.C.; Chiang, H.L.; Liou, J.M.; Chang, C.M.; Lu, T.P.; Chuang, E.Y.; Tai, Y.C.; Cheng, C.; Lin, H.Y.; et al. Altered Gut Microbiota And Inflammatory Cytokine Responses In Patients With Parkinson's Disease. J Neuroinflammation 2019, 16, 129. [Google Scholar] [CrossRef]
- Mu, K.; Yu, S.; Kitts, D.D. The Role Of Nitric Oxide In Regulating Intestinal Redox Status And Intestinal Epithelial Cell Functionality. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Burgueno, J.F.; Fritsch, J.; Santander, A.M.; Brito, N.; Fernandez, I.; Pignac-Kobinger, J.; Conner, G.E.; Abreu, M.T. Intestinal Epithelial Cells Respond To Chronic Inflammation And Dysbiosis By Synthesizing H(2)O(2). Front Physiol 2019, 10, 1484. [Google Scholar] [CrossRef]
- Merilainen, L.; Herranen, A.; Schwarzbach, A.; Gilbert, L. Morphological And Biochemical Features Of Borrelia Burgdorferi Pleomorphic Forms. Microbiology-Sgm 2015, 161, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Alban, P.S.; Johnson, P.W.; Nelson, D.R. Serum-Starvation-Induced Changes In Protein Synthesis And Morphology Of Borrelia Burgdorferi. Microbiology-Uk 2000, 146, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Brorson, Ø.; Brorson, S.H. In Vitro Conversion Of Borrelia Burgdorferi To Cystic Forms In Spinal Fluid, And Transformation To Mobile Spirochetes By Incubation In Bsk-H Medium. Infection 1998, 26, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Murgia, R.; Cinco, M. Induction Of Cystic Forms By Different Stress Conditions In Borrelia Burgdorferi. Apmis 2004, 112, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Mursic, V.P.; Wanner, G.; Reinhardt, S.; Wilske, B.; Busch, U.; Marget, W. Formation And Cultivation Of Borrelia Burgdorferi Spheroplast-L-Form Variants. Infection 1996, 24, 218–226. [Google Scholar] [CrossRef]
- Brorson, O.; Brorson, S.H. Transformation Of Cystic Forms Of Borrelia Burgdorferi To Normal, Mobile Spirochetes. Infection 1997, 25, 240–246. [Google Scholar] [CrossRef]
- Gruntar, I.; Malovrh, T.; Murgia, R.; Cinco, M. Conversion Of Borrelia Garinii Cystic Forms To Motile Spirochetes In Vivo. Apmis 2001, 109, 383–388. [Google Scholar] [CrossRef]
- Dunham-Ems, S.M.; Caimano, M.J.; Pal, U.; Wolgemuth, C.W.; Eggers, C.H.; Balic, A.; Radolf, J.D. Live Imaging Reveals A Biphasic Mode Of Dissemination Of Borrelia Burgdorferi Within Ticks. Journal Of Clinical Investigation 2009, 119, 3652–3665. [Google Scholar] [CrossRef]
- Bamm, V.V.; Ko, J.T.; Mainprize, I.L.; Sanderson, V.P.; Wills, M.K.B. Lyme Disease Frontiers: Reconciling Borrelia Biology And Clinical Conundrums. Pathogens 2019, 8. [Google Scholar] [CrossRef]
- Lantos, P.M.; Auwaerter, P.G.; Wormser, G.P. A Systematic Review Of Borrelia Burgdorferi Morphologic Variants Does Not Support A Role In Chronic Lyme Disease. Clinical Infectious Diseases 2013, 58, 663–671. [Google Scholar] [CrossRef]
- Fenno, J.C. Laboratory Maintenance Of Treponema Denticola. Current Protocols In Microbiology 2006, 00, 12b.11.11–12b.11.21. [Google Scholar] [CrossRef] [PubMed]
- Dhandayuthapani, S.; Via, L.E.; Thomas, C.A.; Horowitz, P.M.; Deretic, D.; Deretic, V. Green Fluorescent Protein As A Marker For Gene-Expression And Cell Biology Of Mycobacterial Interactions With Macrophages. Molecular Microbiology 1995, 17, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Poirier, T.P.; Tonelli, S.J.; Holt, S.C. Ultrastructure Of Gliding Bacteria - Scanning Electron Microscopy Of Capnocytophaga Sputigena, Capnocytophaga Gingivalis, And Capnotcytophaga Ochracea. Infection And Immunity 1979, 26, 1146–1158. [Google Scholar] [CrossRef] [PubMed]
- Sapi, E.; Bastian, S.L.; Mpoy, C.M.; Scott, S.; Rattelle, A.; Pabbati, N.; Poruri, A.; Burugu, D.; Theophilus, P.A.S.; Pham, T.V.; et al. Characterization Of Biofilm Formation By Borrelia Burgdorferi In Vitro. Plos One 2012, 7, E48277. [Google Scholar] [CrossRef] [PubMed]
- Delamater, E.D.; Haanes, M.; Wiggall, R.H.; Pillsbury, D.M. Studies On The Life Cycle Of Spirochetes: Viii. Summary And Comparison Of Observations On Various Organisms. Journal Of Investigative Dermatology 1951, 16, 231–256. [Google Scholar] [CrossRef] [PubMed]
- Pillot, J.; Dupouey, P.; Ryter, A. La Signification Des Formes Atypiques Et La Notion De Cycle Evolutif Chez Les Spirochtes. Annales De L’institut Pasteur 1964, 107, 484–502, 663–677. [Google Scholar]
- Hall-Stoodley, L.; Brun, O.S.; Polshyna, G.; Barker, L.P. Mycobacterium Marinum Biofilm Formation Reveals Cording Morphology. Fems Microbiology Letters 2006, 257, 43–49. [Google Scholar] [CrossRef]
- Savijoki, K.; Myllymäki, H.; Luukinen, H.; Paulamäki, L.; Vanha-Aho, L.-M.; Svorjova, A.; Miettinen, I.; Fallarero, A.; Ihalainen Teemu, O.; Yli-Kauhaluoma, J.; et al. Surface-Shaving Proteomics Of Mycobacterium Marinum Identifies Biofilm Subtype-Specific Changes Affecting Virulence, Tolerance, And Persistence. Msystems 2021, 6, E00500–00521. [Google Scholar] [CrossRef]
- Silva, H.; Oliveira, R.; Almeida, C.; Azevedo, N.F.; Fernandes, R.M.; Silva, H.; Azevedo, N.F.; Vieira, M.J. Morphological Transition Of Helicobacter Pylori Adapted To Water. Future Microbiology. 2017, 12, 1167–1179. [Google Scholar] [CrossRef]
- Zerebinski, J.; Ottemann, K.M.; Hathroubi, S.; Ottemann, K.M.; Freitag, N.E. Helicobacter Pylori Biofilm Involves A Multigene Stress-Biased Response, Including A Structural Role For Flagella. Mbio. 2018, 9. [Google Scholar] [CrossRef]
- Mitchell, H.L.; Dashper, S.G.; Catmull, D.V.; Paolini, R.A.; Cleal, S.M.; Slakeski, N.; Tan, K.H.; Reynolds, E.C. Treponema Denticola Biofilm-Induced Expression Of A Bacteriophage, Toxin-Antitoxin Systems And Transposases. Microbiology-Sgm 2010, 156, 774–788. [Google Scholar] [CrossRef] [PubMed]
- Cantillon, D.; Wroblewska, J.; Cooper, I.; Newport, M.J.; Waddell, S.J. Three-Dimensional Low Shear Culture Of Mycobacterium Bovis Bcg Induces Biofilm Formation And Antimicrobial Drug Tolerance. Npj Biofilms And Microbiomes 2021, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Lerner, T.R.; Queval, C.J.; Lai, R.P.; Russell, M.R.G.; Fearns, A.; Greenwood, D.J.; Collinson, L.; Wilkinson, R.J.; Gutierrez, M.G. Mycobacterium Tuberculosis Cords Within Lymphatic Endothelial Cells To Evade Host Immunity. Jci Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Light- and electron microscopy of round bodies from Mucispirillum schaedleri, Treponema denticola, and Borrellia burgdorferi. (A) M. schaedleri ASF 457 from plate culture. Appressed cells (stars) and round bodies attached to tips of rods (arrows). Note crescent-shaped regions on the walls of round bodies. Transmitted light micrograph, bar = 5 µm. (B) Confocal micrograph of viability-stained cells from 3-day-old liquid culture. (C) Electron micrograph of appressed cells from liquid culture. Bar = 500 nm. (D) Confocal micrograph of viability-stained cells from 6-day-old broth culture showing crescent-shaped cells (round bodies); fully active cells (green) together with those having reduced activity (red, orange). (E) Confocal micrograph of viability-stained round body from 6-day-old broth culture showing spiral morphology. ¬(F, G) Electron micrographs of round bodies from broth culture. Crescent- and helical-shaped cellular cross-sections are visible within the round bodies (bar = 500 µm). Multiple cross-sections are visible in single round bodies, and round bodies also have amorphous contents. (H) Treponema denticola biofilm showing round bodies (from current study). (I) High-magnification micrograph of round body from center of H. (J, K) Electron micrographs of Borrellia burgdorferi from a different study [35].
Figure 1.
Light- and electron microscopy of round bodies from Mucispirillum schaedleri, Treponema denticola, and Borrellia burgdorferi. (A) M. schaedleri ASF 457 from plate culture. Appressed cells (stars) and round bodies attached to tips of rods (arrows). Note crescent-shaped regions on the walls of round bodies. Transmitted light micrograph, bar = 5 µm. (B) Confocal micrograph of viability-stained cells from 3-day-old liquid culture. (C) Electron micrograph of appressed cells from liquid culture. Bar = 500 nm. (D) Confocal micrograph of viability-stained cells from 6-day-old broth culture showing crescent-shaped cells (round bodies); fully active cells (green) together with those having reduced activity (red, orange). (E) Confocal micrograph of viability-stained round body from 6-day-old broth culture showing spiral morphology. ¬(F, G) Electron micrographs of round bodies from broth culture. Crescent- and helical-shaped cellular cross-sections are visible within the round bodies (bar = 500 µm). Multiple cross-sections are visible in single round bodies, and round bodies also have amorphous contents. (H) Treponema denticola biofilm showing round bodies (from current study). (I) High-magnification micrograph of round body from center of H. (J, K) Electron micrographs of Borrellia burgdorferi from a different study [35].
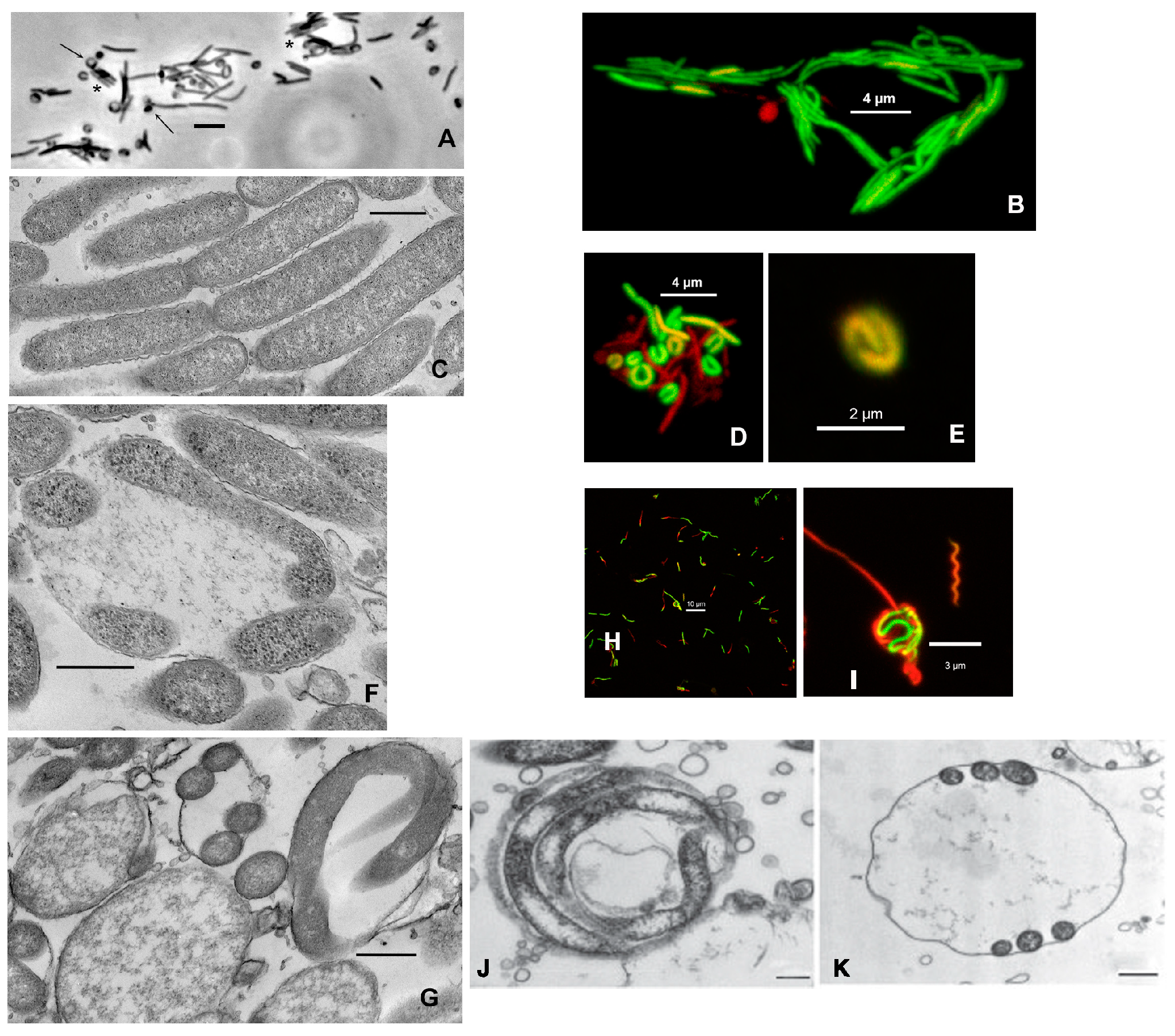
Figure 2.
Quantification of rods and round bodies in 3- and 6-day-old Mucispirillum schaedleri cultures. MS 3-day and 6-day cultures; n=4 independent experiments. All values are presented as the mean ± standard deviation (SD), p<0.05 was considered significant. MS, Mucispirillum schaedleri.
Figure 2.
Quantification of rods and round bodies in 3- and 6-day-old Mucispirillum schaedleri cultures. MS 3-day and 6-day cultures; n=4 independent experiments. All values are presented as the mean ± standard deviation (SD), p<0.05 was considered significant. MS, Mucispirillum schaedleri.
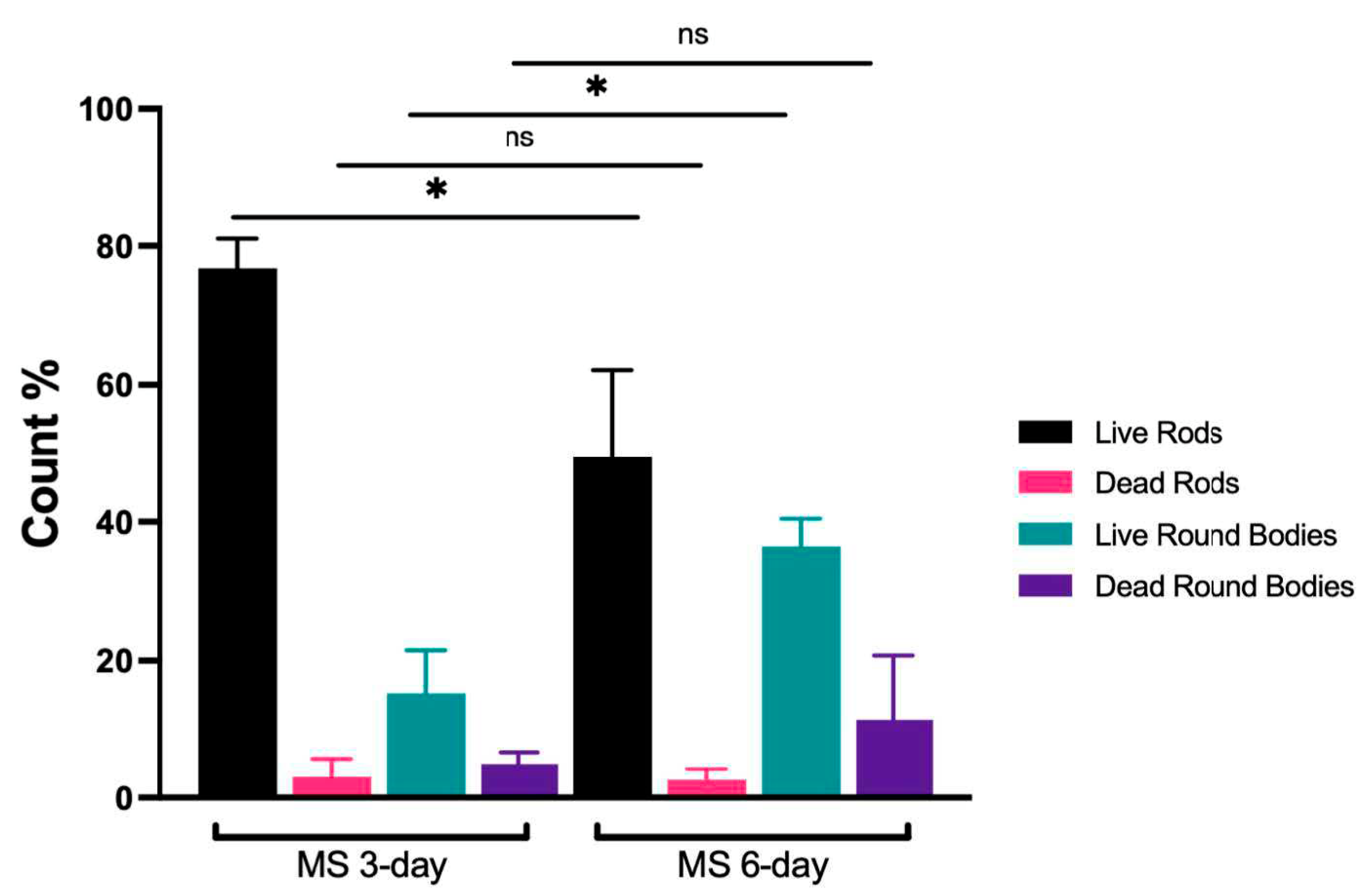
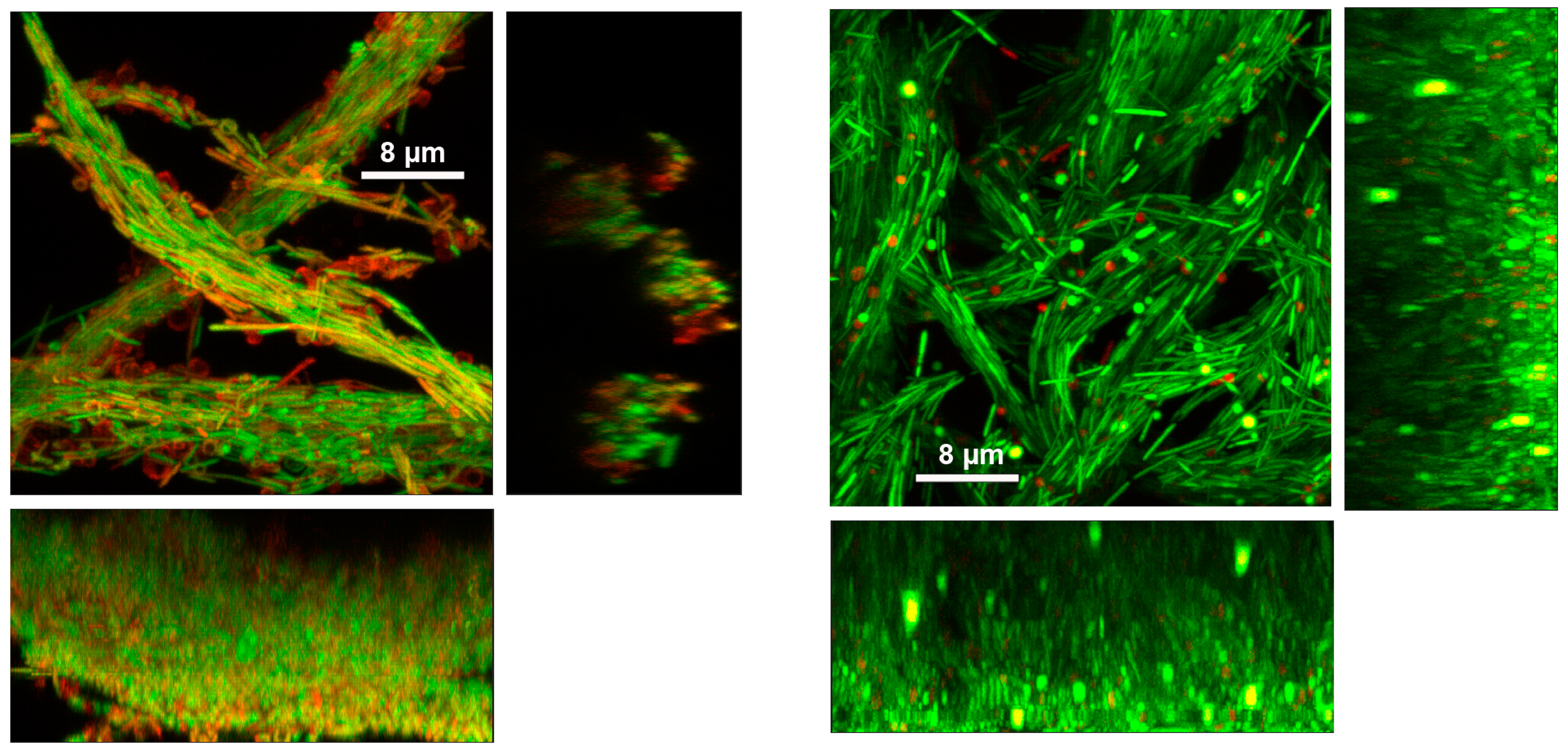
Figure 3.
Confocal micrographs document cording in biofilms of Mucispirillum schaedleri ASF 457 and of Mycobacterium spp. (A) Chamber-slide-grown M. schaedleri biofilm after 18 hours growth. Viability-stained. (B) Chamber-slide-grown biofilm after 48 hours growth. (C) Flowcell-grown M. schaedleri biofilm after 18 hrs growth. Viability-stained. (D) Flowcell-grown biofilm after 48 hrs growth. Bar = 50 µm in A, B, C, D. Rectangular panels beneath and to the right of the main (square) image are side-on views. (E, F) GFP-expressing Mycobacterium bovis pellicle biofilm from current study. (G) tdTomato-expressing M. marinum biofilm. Reprinted under license from reference [39].
Figure 3.
Confocal micrographs document cording in biofilms of Mucispirillum schaedleri ASF 457 and of Mycobacterium spp. (A) Chamber-slide-grown M. schaedleri biofilm after 18 hours growth. Viability-stained. (B) Chamber-slide-grown biofilm after 48 hours growth. (C) Flowcell-grown M. schaedleri biofilm after 18 hrs growth. Viability-stained. (D) Flowcell-grown biofilm after 48 hrs growth. Bar = 50 µm in A, B, C, D. Rectangular panels beneath and to the right of the main (square) image are side-on views. (E, F) GFP-expressing Mycobacterium bovis pellicle biofilm from current study. (G) tdTomato-expressing M. marinum biofilm. Reprinted under license from reference [39].
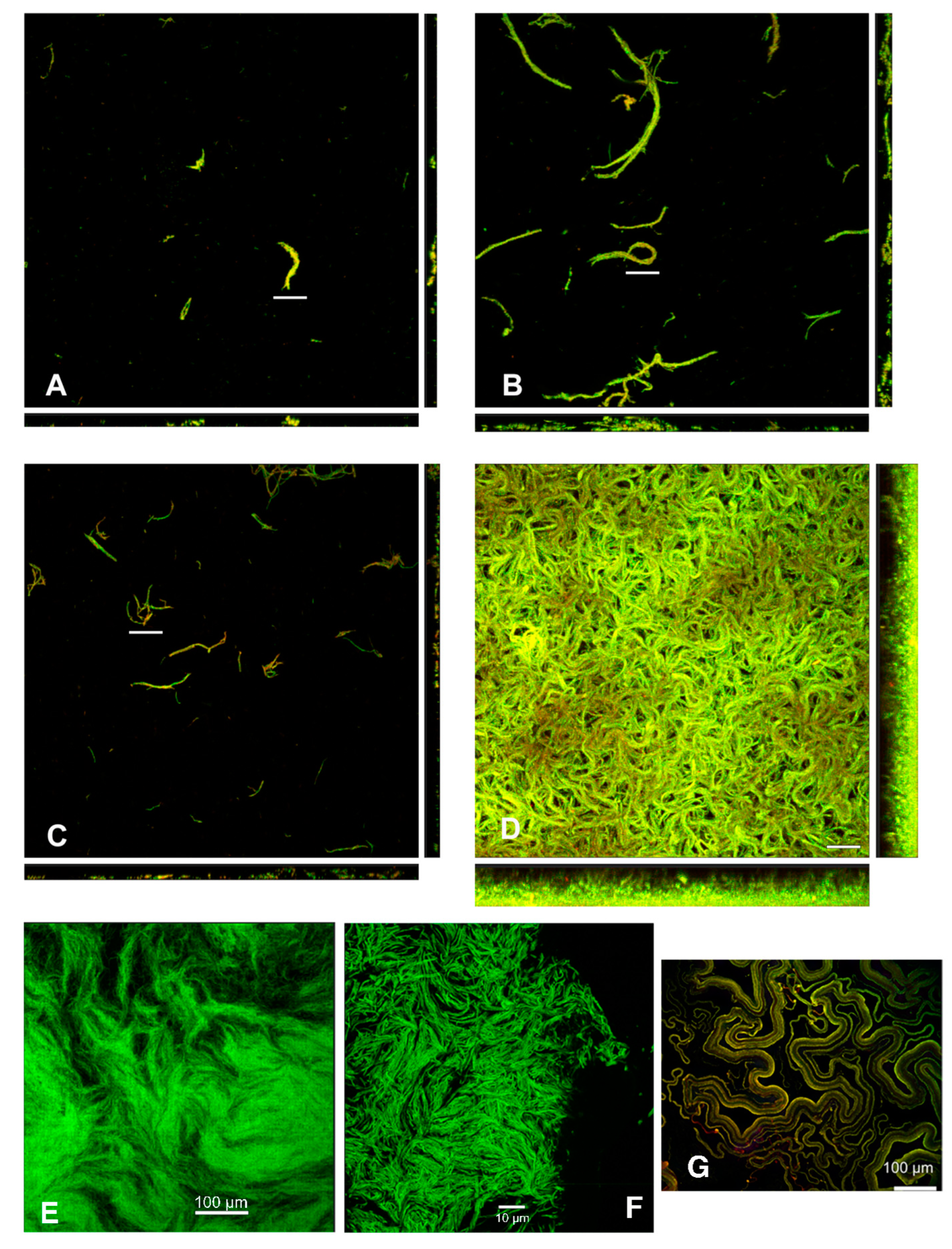
Figure 4.
Comparison of chamber- and flowcell-grown Mucispirillum schaedleri biofilms. Biovolume (left) and maximum height (right) of chamber-grown and flowcell-grown M. schaedleri biofilms after 18 hrs and 48 hrs. .
Figure 4.
Comparison of chamber- and flowcell-grown Mucispirillum schaedleri biofilms. Biovolume (left) and maximum height (right) of chamber-grown and flowcell-grown M. schaedleri biofilms after 18 hrs and 48 hrs. .
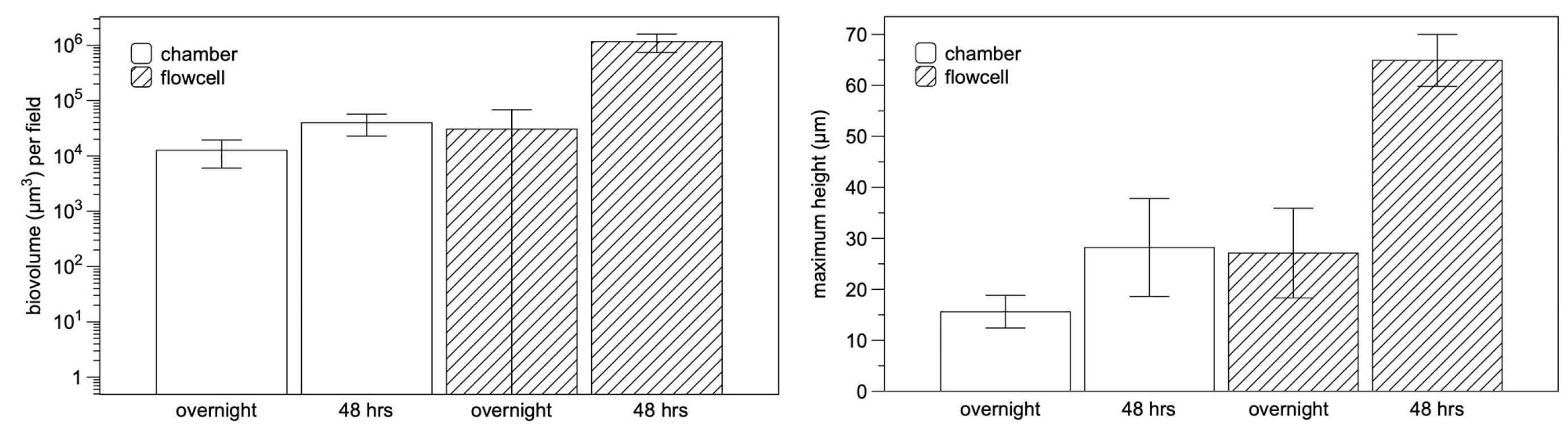
Figure 5.
High-magnification confocal micrographs of viability-stained 48-hr-old chamber-grown Mucispirllum schaedleri biofilm (left) and flowcell-grown biofilm (right). Rectangular panels beneath and to the right of the main (square) image are side-on views. Supplementary Videos S1-S4 show the series of z-planes through the samples.
Figure 5.
High-magnification confocal micrographs of viability-stained 48-hr-old chamber-grown Mucispirllum schaedleri biofilm (left) and flowcell-grown biofilm (right). Rectangular panels beneath and to the right of the main (square) image are side-on views. Supplementary Videos S1-S4 show the series of z-planes through the samples.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



