
Submitted:
21 July 2023
Posted:
25 July 2023
You are already at the latest version
Abstract
To identify soil microbial communities, it is essential to understand their phenotypic and genetic characteristics. Plant growth-promoting rhizobacteria (PGPR) are bacteria capable of promoting plant growth. In addition, rhizobacteria also serve as biological control agents. In addition to fixing atmospheric nitrogen, solubilising insoluble phosphate and secreting hormones such as indoloacetic acid, kinetics and gibberellins, plant growth-promoting rhizobacteria help plants grow directly. Furthermore, they facilitate plant growth indirectly by inducing systemic resistance, siderophores, antibiotics, lytic enzymes, hydrogen cyanide production and stress regulation. This review examines in detail the direct and indirect mechanisms of PGPR action and their interaction in plant growth and resistance.
Keywords:
Microbial biofertilizers
; microbial symbioses
; plant interactions
; crop resistance
1. Introduction
Rhizobacteria that supports plant growth are known as Plant Growth-Promoting Rhizobacteria (PGPR) [1,2]. The diversity of phenotypic and genotypic characteristics of soil microbiomes makes them complex and difficult to characterise [3]. However, as the rhizosphere has become increasingly important to the bio-sphere in recent years, several PGPRs have been identified that significantly have a great impact in plant growth, primarily because they act as an ecological unit [4]. The PGPRs affect plant growth by solubilising insoluble phosphates, fixing atmospheric nitrogen, and secreting hormones that control plant growth [5]. In addition, through induced systemic resistance (ISR), nutrient competition, antibiotics, parasitism, and suppression of rhizobacterial growth, PGPRs indirectly benefit plant growth [6]. These communities are very diverse, and their actions can take many forms, including antagonistic action against pathogens in the soil and inducing systemic resistance against pathogens throughout the plant [7]. Plants can be indirectly aided in growing by antagonistic rhizobacteria because they produce various substances that can control pathogens [8]. If the inducing bacteria and the challenging pathogen remain spatially separated, inducing systemic resistance (ISR) can be compared to pathogen-induced acquired systemic resistance (SAR). Different plant species have induced resistance that makes uninfected parts of the plant more resistant to pathogen attack [9]. Induction of resistance occurs by rhizobacteria either through salicylic acid-dependent SAR pathways or through the bacteria's perception of jasmonic acid and ethylene. Among the many characteristics of rhizobacteria are their antagonistic effects and ability to trigger inflammatory responses. Many studies have examined the use of PGPRs as a substitute for crop protection agents (fertilisers and pesticides) for plant growth promotion in recent years [10,11].The rhizobacteria can alter the soil structure, recycle essential elements, decompose organic matter, solubilise mineral nutrients, and serve as biocontrol agents for soil-borne and seed-borne pathogens [12,13,14]. A good understanding of rhizobacteria that promote plant growth and their interaction with bi-otic and abiotic factors is crucial to bioremediation techniques. This is also relevant for energy generation processes, and biotechnology industries, such as pharmaceutical, chemical and food industries [15]; rhizobacteria are also useful for reducing the use of chemical fertilisers. The main advantage of this approach is the increasing the productivity and sustainability of agricultural systems and soil fertility [16]. As a result, production costs are reduced, and the best soil and crop management practices are identified [17]. The aim of this review was to show the possible benefits of the application of rhizobacteria in plants, the direct and indirect mechanisms they influence, the possible applications of PGPR-based formulations in agriculture and the prospects of the use of rhizobacteria on crops.
2. Plant and soil effects of PGPRs
Rhizobacteria that promote plant growth are well known and essential, and this growth enhancement is due to rhizobacteria's characteristics [18]. PGPRs can enhance plant growth and development through various mechanisms [19]. In particular, rhizobacteria produce a variety of substances that alter the entire microbial community in the rhizosphere; and they are capable of supplying nutrients (nitrogen, phosphorus, potassium and essential minerals) or producing plant hormones [20]. For example, the inoculation of rhizobacteria in Astrophytum spp. grown in biochar-enriched substrates improves vegetative and root growth and plant flowering (Figure 1) [21]. By acting as biocontrol agents, environmental protectors, and root colonisers, PGPRs can also indirectly promote plant growth by reducing the effects of pathogens [22,23]. Sustainable agriculture and ecosystem stability are severely threatened by phytopathogenic microorganisms, which degrade soil fertility, alter the environment, and, ultimately, harm human health and contaminate groundwater [24]. By fixing nitrogen, mineralising organic compounds, solubilising mineral nutrients, and producing phytohormones, PGPRs also facilitate plant uptake of nutrients and increased the and increase resistance to biotic and abiotic stresses. Many species are able to survive particular environmental conditions, such as high temperatures and drought (Table 1) [25]. As an indirect means of achieving soil fertility and plant growth, PGPRs are crucial to a sustainable and ecological approach. This can be achieved through various mechanisms, including antibiotics, HCNs, siderophores, and hydrolytic enzymes, and as outlined before, PGPRs can be exploited to decrease the need for agrochemicals such as fertilisers and pesticides and increase soil fertility [26].
3. Mechanisms activated directly by Plant Growth Promoting Rhizobacteria
In terms of plant growth, phytohormones play a critical role. These are plant hormones that affect the plant's response to its environment. These hormones are produced at one point in the plant and then transferred to another part of the plant, where they are used to promote the growth [1]. Roots and leaves grow due to the physical responses caused by these hormones [27]. Some essential plant hormones are auxins, gibberellins, ethylene, cytokinins, and abscisic acid [28]. Rhizobacteria produce these phytohormones. In addition to auxins and gibberellins, ethylene, cytokinins and abscisic acid are important phytohormones [29]. In plant roots and shoots, cytokinins (CKs) play a role in cell division [30]. Among their benefits, there is the growth of cells, differentiation of cells, apical dominance, axillary bud development, and leaf senescence [31,32]. Plants synthesise this hormone, but yeast strains and PGPR strains can also prepare it. As well as phytopathogens, some phytopathogens can synthesise cytokinins. It has been reported that Azotobacter species, Pantoea agglomerans strains, Rhizobium species, Rhodospirillum rubrum strains, Bacillus subtilis strains, Pseudomonas fluorescens strains, and Paenibacillus polymyxa species all produce the cytokinin hormone [33,34]. Some rhizobacteria are able by their action to mitigate the effect of different types of stress such as water, salt and heat stress (Table 2) [35]. A class of important plant hormones, gibberellins (GA) control various developmental processes in plants. Their functions include stem elongation, dormancy, germination, flowering, and flower development. Several cytokinin-producing polymeric protein receptors synthesise gibberellin, a phytohormone involved in breaking dormancy and other aspects of germination. Gibberellin is the most crucial phytohormone synthesised by some PGPRs. Production and regulation of gibberellin and cytokinin are extremely important [36]. PGPRs and plants produce a variety of phytohormones, including indoloacetic acid. In addition to cell division, other proprieties like gene expression, organogenesis, pigmentation, root development, seed germination, stress resistance, tropical responses, and photosynthesis, play an essential role in plant cellular responses [37]. Plants and bacteria influence the amount of IAAs required to promote plant growth vigorously. The amount of IAA required to promote plant growth depends on the plant and bacterial species. PGPRs produce indole-3-acetic acid, which is responsible for root elongation and the formation of roots. Nearly all plants produce ethylene as a growth hormone, which is key in many physiological changes [38]. Plants respond to biotic and abiotic stresses negatively, affecting root growth and plant growth [39]. The PGPR enzyme 1-aminocyclopropane-1-carboxylate (ACC) deaminase can regulate ethylene production. Inoculation with PGPRs can maintain plant growth and development under stressful conditions, such as drought, salinity, cold, and soil pollution, plants synthesise abscisic acid [25]. This growth hormone activates stress-resistance genes. Abscissic acid producing strains, such as Bacillus licheniformis Rt4M10, Azospirillum brasilense sp. 245, and Pseudomonas fluorescens Rt6M10, increase the internal ABA content of plants. As a result, the plants become more resilient to drought. The unavailability of nitrogen can limit plant growth, but phosphorus is also essential for life [40]. There are large quantities of phosphate in soil, but they are in an insoluble form that plants cannot utilise for growth since they are insoluble. PGPRs can solubilise phosphate in soil via acidification, chelation or enzyme action [41]. For example, the PGPR Gluconacetobacter diazotrophicus, found in sugar cane, can solubilise phosphate by acidification [7].

Table 2.
Application of Rhizobacteria in mitigating heat stress in plants [35].
Table 2.
Application of Rhizobacteria in mitigating heat stress in plants [35].
| Microbes | Plant | Parameters | Stress |
|---|---|---|---|
|
Enterobacter SA187 |
Arabidopsis thaliana, Wheat plant | Increased Biomass, height, seed weight | Long term |
|
Septoglomus deserticola |
Solanum Lycopersicum |
Improved stomatal conductance, water content |
Heat Drought |
|
Pseudomonas fluorescens, Pantoea agglomerans |
Triticum aestivum |
Increased Antioxidant enzymes |
High Temp. |
| B. phytofirmans |
Solanum tuberosum |
Increased Proline and glycine betaine |
High Temp. |
| B. cereus | Soybean | Increased Chlorophyll and Carotenoid |
High Temp. |
4. Mechanisms indirectly activated by Plant Growth-Promoting Rhizobacteria
Microorganisms compete for nutrients and colonisation sites in their natural environment fiercely. Various mechanisms of PGPR species have evolved that allow them to reduce competition by releasing antibiotics, lytic enzymes, or weak organic acids into their environments (Figure 2) [21,42]. As a result, PGPRs are valuable tools that can be used against plant pathogens. However, there is a possibility of the development of resistant pathogens if antibiotic-producing bacteria are used more frequently. It was shown that PGPR enzymes secreted by these PGPRs could eliminate pathogens such as Botrytis cinerea, Fusarium oxysporum, Sclerotium rolfsii, Phytophthora spp, Pythium ultimum and Rhizoctonia solani [43,44]. They include cellulases, chitinases, lipases and proteases secreted by the plant. Plants respond to pathogens in two ways: acquired systemic resistance and induced systemic resistance. Induced systemic resistance is the same as Systemic Acquired Resistance (SAR) [45]. Acquired systemic resistance is triggered by the infection of a plant by a pathogen, whereas PGPR triggers acquired systemic resistance. The application of PGPR inoculum induces systemic resistance in the plant against many bacterial pathogens. Plants also need iron as a nutrient. Under aerobic conditions, iron is found as Fe3+, which is not soluble for use by microorganisms and plants [46]. Therefore, microbes produce and secrete iron chelators, known as siderophores. The chelators are highly iron-adjacent. They function as iron solubilising agents under limiting conditions. In the cell, Fe3+ is transformed into Fe2+, which is then released by the siderophores [47]. Rhizobacteria steal iron from other microorganisms by producing siderophores. In addition, PGPRs use siderophores to remove other heavy metals from soil and prevent the heavy metals from causing toxicity in plants. They can also be used for the bioremediation of heavy metal toxic soils [48]. As well as helping plants grow and ripen fruit, ethylene acts as a phytohormone in response to salt, drought or pathogenic bacteria. However, high amounts of ethylene can also cause plant harm. Many PGPR synthesises an ACC deaminase enzyme [50]. This enzyme destroys 1-aminocyclopropane-1-carboxylate, the precursor of ethylene. It relieves plant stress by reducing ethylene levels. Plant root surfaces can be colonised by harmful rhizobacteria that act as biocontrol agents for weeds. They produce toxic compounds known as cyanides, produced by many microorganisms, such as bacteria, algae, fungi and plants [51]. Biological weed control agents can be derived from host-specific rhizobacteria, which compete with their counterparts to survive. It has no negative impact on host plants when inoculated with cyanide-producing bacterial strains that produce cyanide [52]. In addition, weed biocontrol agents, such as hydro-gen cyanide, are produced, which inhibits the electron transport chain and energy supply to cells. Many harmful microbes compete with PGPRs for nutrients, but these nutrients are present only in trace amounts so that they can limit the disease's causative agent [53]. In fertile soils with abundant non-pathogenic microbes, they colonise plant surfaces quickly and utilise nutrients. These mechanisms can be challenging to study in the system because they inhibit pathogenic microbes from growing. One essential interaction that indirectly supports plant growth is the competition for nutrients between PGPR and pathogens [54].
5. Application of PGPR-based formulations
The support of crop management by beneficial microorganisms is an environmentally friendly alternative to the conventional techniques that are based on chemical inputs, with respect to the increasing consumer expectations of healthy products and current policies towards the implementation of environmentally friendly cropping systems [55]. Besides biotic stresses, useful microorganisms in agriculture have been shown to increase plant tolerance to abiotic stresses such as flooding, water shortages, and excess salinity [56]. Plant growth regulators of microbial origin are of great agrarian and ecological interest, since they offer significant opportunities for eco-friendly agronomic applications. As a result of selected strains, these regulators can also be used in the open field today, thus overcoming certain limitations. It is difficult to colonize the rhizosphere of an adult plant that is already well colonized by resident microorganisms due to high competition [57]. Soil type, temperature, introduced strains, inoculant density, and plant species can all influence the immediate response to PGPR soil administration. After inoculation, the introduced population typically drops rapidly, and it is possible that the number of PGPR colonizing roots is insufficient to achieve the desired results [58]. Other times, the introduced microorganisms cannot find a free ecological niche in the soil. As well as maintaining the desired character characteristics, the strains used must be capable of surviving the stresses associated with concentration and stabilisation processes during production. Agricultural crops can be inoculated in a variety of ways:
- covering the seed at the time of sowing;
- using confected seed, i.e. covered with matrices that have included beneficial microorganisms;
- distributing the product directly in the furrows at the time of sowing;
- performing covering treatments during plant growth.
Using seed inoculation allows farmers to sow and inoculate at the same time, thus saving time and money. Another option is to encapsulate microbial cells in polymers, particularly alginate, which protect them from environmental stress and allow them to be released into the soil slowly and in large quantities [59]. For example, alginate preparations have been proposed for Pseudomonas fluorescens as a biocontrol and biostimulating agent, and for Azospirillum brasilense as a biofertilizing and biostimulating agent [60].
6. Perspectives on the use of Plant Growth Promoting Rhizobacteria in agriculture
An essential and safe method to increase yield is PGPR. In terms of increasing productivity, it is a promising solution [61,62]. In addition, it protects plants from chemicals used to control pests, which can also have a negative impact on the environment. Plant diseases and pests can also be controlled with PGPRs, improving yields. In laboratory and greenhouse experiments, PGPR strains are advantageous [63,64]. The field of genetic engineering is emerging as a means to improve PGPR strains and explore their potential applications. Besides all these advances, some environmental barriers and adverse conditions greatly influence the activity of PGPRs [65]. Mixing of strains, use of improved inoculation techniques and transfer of the active gene source of antagonists to the host plant can improve the variable efficacy of PGPRs [66]. Furthermore, biocontrol agents need a specific ecological environment to grow and survive, so different conditions may influence their efficacy and use [58,67]. The efficacy of biocontrol agents can be modified by using compatible mixed inocula in different ecological niches. Besides these advantages, PGPRs face several challenges [68]. Due to natural variations, it is difficult to predict the behaviour of bacteria in the laboratory and on the farm. These variations can have a significant effect on the entire experiment. Plant type and season can also influence the propagation of PGPRs to recover their viability and biological activity [69,70]. According to the notion that these bacteria can be applied as biofertilisers in agriculture and forestry [71,72], monitoring their activity under stress conditions such as salinity, soil pollution and other environmental conditions that alter crop productivity and yield is essential to understand their applications in different sectors of agriculture. Soil moisture, electrical conductivity and N, P and K concentrations must be monitored under different climatic conditions and bacterial concentrations. This is important in order to develop real, concrete microbial application protocols suitable for different geographical locations.
7. Conclusions
A significant improvement in plant growth, health and yield has been observed with the use of PGPRs. Plant growth, health and yield can be improved directly or indirectly by PGPRs through different mechanisms of action. Plants can also grow faster by reducing phytopathogens that reduce yield and growth with PGPRs. Several factors can influence the inoculation of PGPRs, including the age of the plants and the chemical, physical and biological properties of the soil. To overcome these problems, field studies have to be carried out to identify the best application methods. This may one day ensure that PGPRs can replace chemical fertilisers in agriculture and ensure greater environmental sustainability.
Author Contributions
Conceptualization, D.P.; methodology, writing—original draft preparation D.P. and D.S.; software and investigation, R.F.; writing—review and editing, D.P.; funding acquisition, D.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the CREA Research Centre for Vegetable and Ornamental Crops.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data, tables, and figures in this manuscript are original.
Acknowledgments
The author would like to express his heartfelt gratitude to his colleagues at CREA Research Centre for Vegetable and Ornamental Crops in Pescia and to all other sources for their cooperation and guidance in writing this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Camaille, M.; Fabre, N.; Clément, C.; Ait Barka, E. Advances in Wheat Physiology in Response to Drought and the Role of Plant Growth Promoting Rhizobacteria to Trigger Drought Tolerance. Microorganisms 2021, 9, 687. [Google Scholar] [CrossRef]
- Di Salvo, L.P.; Cellucci, G.C.; Carlino, M.E.; De Salamone, I.E.G. Plant growth-promoting rhizobacteria inoculation and nitrogen fertilization increase maize (Zea mays L.) grain yield and modified rhizosphere microbial communities. Appl. Soil Ecol. 2018, 126, 113–120. [Google Scholar] [CrossRef]
- Ekinci, M.; Kocaman, A.; Argin, S.; Turan, M.; Dadaso-Glu, F. Rhizobacteria alleviate the adverse effects of salt stress on seedling growth of Capsicum annuum L. by modulating the antioxidant enzyme activity and mineral uptake. Taiwania 2021, 66, 287–297. [Google Scholar]
- El-Sawah, A.; El-Keblawy, A.; Ali, D.; Ibrahim, H.; El-Sheikh, M. Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Rhizobacteria Enhance Soil Key Enzymes, Plant Growth, Seed Yield, and Qualitative Attributes of Guar. Agriculture 2021, 11, 194. [Google Scholar] [CrossRef]
- Emmanuel, O.C.; Babalola, O.O. Productivity and quality of horticultural crops through coinoculation of arbuscular mycorrhizal fungi and plant growth promoting bacteria. Microbiol. Res. 2020, 239, 126569. [Google Scholar] [CrossRef] [PubMed]
- Enebe, M.C.; Babalola, O.O. The influence of plant growth-promoting rhizobacteria in plant tolerance to abiotic stress: A survival strategy. Appl Microbiol Biotechnol. 2018, 102, 7821–7835. [Google Scholar] [CrossRef]
- Forni, C.; Duca, D.; Glick, B.R. Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil. 2017, 410, 335–356. [Google Scholar] [CrossRef]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.S. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef]
- Grover, M.; Bodhankar, S.; Sharma, A.; Sharma, P.; Singh, J. PGPR mediated alterations in root traits: Way toward sustainable crop production. Front Sustain. Food Syst. 2021, 4, 39. [Google Scholar] [CrossRef]
- Kalsoom, M.; Rehman, F.; Shafique, T.; Junaid, S.; Khalid, N. Biological importance of microbes in Agriculture, Food and Pharmaceutical Industry: A Review. Innove. J. Life Sci. 2020, 8, 1–4. [Google Scholar] [CrossRef]
- Kenneth, O.C.; Nwadibe, E.C.; Kalu, A.U.; Unah, U.V. Plant growth promoting rhizobacteria (PGPR): A novel agent for sustainable food production. J. Agric. Biol. Sci. 2019, 14, 35–54. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Shahid, M.A.; Nasim, W.; Babar, M.A. Interaction between PGPR and PGR for water conserva-tion and plant growth attributes under drought condition. Biologia. 2018, 73, 1083–1098. [Google Scholar] [CrossRef]
- Kumari, B.; Mallick, M.A.; Solanki, M.K.; Solanki, A.C.; Hora, A. Plant Growth Promoting Rhizobacteria (PGPR): Modern Prospects for Sustainable Agriculture. In Plant Health Under Biotic Stress; Springer: Singapore, 2019; pp. 109–127. [Google Scholar]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S. Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: A methodical review. Sci. Total Environ. 2020, 743, 140682. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Gonzalez-Andres, F. Bacillus as a source of phytohormones for use in agriculture. Appl. Microbiol. Biotechnol. 2021, 105, 8629–8645. [Google Scholar] [CrossRef] [PubMed]
- Prasad, M.; Srinivasan, R.; Chaudhary, M.; Choudhary, M.; Jat, L.K. Plant growth promoting rhizobacteria (PGPR) for sustainable agriculture: Perspectives and challenges. In PGPR Amelioration in Sustainable Agriculture; Woodhead Publishing, 2019; pp. 129–157. [Google Scholar]
- Rehman, F.; Kalsoom, M.; Adnan, M.; Toor, M.D.; Zulfiqar, A. Plant Growth Promoting Rhizobacteria and their Mechanisms Involved in Agricultural Crop Production: A Review. SunText Rev Biotechnol. 2020, 1, 110. [Google Scholar]
- Rehman, F.; Kalsoom, M.; Nasir, T.A.; Adnan, M.; Anwar, S. Chemistry of Plant–Microbe Interactions in Rhizosphere and Rhizoplane. Ind. J. Pure App. Biosci. 2020, 8, 11–19. [Google Scholar] [CrossRef]
- Song, Q.; Song, X.S.; Deng, X.; Luo, J.Y.; Song, R.Q. Effects of plant growth promoting Rhizobacteria microbial on the growth, rhizosphere soil properties, and bacterial community of Pinus sylvestris var. mongolica seedlings. Scand J For Res. 2021, 36, 249–262. [Google Scholar] [CrossRef]
- Prisa, D. Biochar effects in the growing and control of biotic and abiotic stress in Astrophytum myriostigma and Astrophytum capricorne. GSC Biol. Pharm. Sci. 2021, 16, 186–194. [Google Scholar] [CrossRef]
- Tariq, M.; Noman, M.; Ahmed, T.; Hameed, A.; Manzoor, N. Antagonistic features displayed by plant growth promoting rhizobacteria (PGPR): A review. J. Plant Sci. Phytopathol. 2017, 1, 38–43. [Google Scholar]
- Ullah, N.; Ditta, A.; Imtiaz, M.; Li, X.; Jan, A.U. Appraisal for organic amendments and plant growth-promoting rhizobacteria to enhance crop productivity under drought stress: A review. J. Agron. Crop Sci. 2021, 207, 783–802. [Google Scholar] [CrossRef]
- Wang, H.; Liu, R.J.; You, M.P.; Barbetti, M.J.; Chen, Y.L. Pathogen biocontrol using plant growth-promoting bacteria (PGPR): Role of bacterial diversity. Microorganisms 2021, 9, 1988. [Google Scholar] [CrossRef] [PubMed]
- Abdelaal, K.; Alkahtani, M.; Attia, K.; Hafez, Y.; Király, L.; Künstler, A. The role of plant growth-promoting bacteria in alleviating the adverse effects of drought on plants. Biology. 2021, 10, 520. [Google Scholar] [CrossRef]
- Zeng, Q.; Ding, X.; Wang, J.; Han, X.; Iqbal, H.M.N. Insight into soil nitrogen and phosphorus availability and agricultural sustainability by plant growth-promoting rhizobacteria. Environ. Sci. Pollut. Res. 2022, 29, 45089–45106. [Google Scholar] [CrossRef]
- Dar, S.A.; Bhat, R.A.; Dervash, M.A.; Dar, Z.A. Azotobacter as biofertilizer for sustainable soil and plant health under saline environmental conditions. Microbiota and Biofertilizers. Springer 2021, 4, 231–254. [Google Scholar]
- Ghaffari, M.R.; Mirzaei, M.; Ghabooli, M.; Khatabi, B.; Wu, Y.; Zabet-Moghaddam, M.; Mohammadi-Nejad, G.; Haynes, P.A.; Hajirezaei, M.R.; Sepehri, M.; Salekdeh, G.H. Root endophytic fungus Piriformospora indica improves drought stress adaptation in barley by metabolic and proteomic reprogramming. Environ. Exp. Bot. 2019, 157, 197–210. [Google Scholar] [CrossRef]
- Liu, G.Y.; Chen, L.L.; Shi, X.R.; Yuan, Z.Y.; Yuan, L.Y.; Lock, T.R.; Kallenbach, R.L. Changes in rhizosphere bacterial and fungal community composition with vegetation restoration in planted forests. Land Degrad. Dev. 2019, 30, 1147–1157. [Google Scholar]
- Dubey, A.; Malla, M.A.; Khan, F.; Chowdhary, K.; Yadav, S.; Kumar, A.; Sharma, S.; Khare, P.K.; Khan, M.L. Soil microbiome: A key player for conservation of soil health under changing climate. Biodivers. Conserv. 2019, 28, 2405–2429. [Google Scholar] [CrossRef]
- Diagne, N.; Ndour, M.; Djighaly, P.I.; Ngom, D.; Ngom, M.C.N.; Ndong, G.; Svistoonoff, S.; Cherif-Silini, H. Effect of plant growth promoting rhizobacteria (PGPR) and arbuscular mycorrhizal fungi (AMF) on salt stress tolerance of Casuarina obesa (Miq. ). Front. Sustain. Food Syst. 2020, 4, 266. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Babalola, O.O.; Santoyo, G.; Perazzolli, M. The potential role of microbial biostimulants in the amelioration of climate change-associated abiotic stresses on crops. Front. Microbiol. 2022, 12, 829099. [Google Scholar] [CrossRef]
- Ramakrishna, W.; Yadav, R.; Li, K. Plant growth promoting bacteria in agriculture: Two sides of a coin. Appl. Soil Ecol. 2019, 138, 10–18. [Google Scholar] [CrossRef]
- Chiappero, J.; Del Rosario Cappellari, L.; Alderete, L.G.S.; Palermo, T.B.; Banchio, E. Plant growth promoting rhizobacteria improve the antioxidant status in Mentha piperita grown under drought stress leading to an enhancement of plant growth and total phenolic content, Ind. Crops Prod. 2019, 139, 111553. [Google Scholar] [CrossRef]
- Shaffique, S.; Khan, M.A.; Wani, S.H.; Pande, A.; Imran, M.; Kang, S.M.; Rahim, W.; Khan, S.A.; Bhatta, D.; Kwon, E.H. A Review on the Role of Endophytes and Plant Growth Promoting Rhizobacteria in Mitigating Heat Stress in Plants. Microorganisms 2022, 10, 1286. [Google Scholar] [CrossRef]
- Vorholt, J.A.; Vogel, C.; Carlström, C.I.; Müller, D.B. Establishing causality: Opportunities of synthetic communities for plant microbiome research. Cell Host Microbe. 2017, 22, 142–155. [Google Scholar] [CrossRef]
- Kaminsky, L.M.; Trexler, R.V.; Malik, R.J.; Hockett, K.L.; Bell, T.H. The inherent conflicts in developing soil microbial inoculants. Trends Biotechnol. 2019, 37, 140–151. [Google Scholar] [CrossRef]
- Naylor, D.; Degraaf, S.; Purdom, E.; Coleman-Derr, D. Drought and host selection influence bacterial community dynamics in the grass root microbiome. ISME J. 2017, 11, 2691–2704. [Google Scholar] [CrossRef]
- Ahluwalia, O.; Singh, P.C.; Bhatia, R. A review on drought stress in plants: Implications, mitigation and the role of plant growth promoting rhizobacteria. Resour. Environ. Sustain. 2021, 5, 100032. [Google Scholar] [CrossRef]
- Akhtar, N.; Ilyas, N.; Hayat, R.; Yasmin, H.; Noureldeen, A.; Ahmad, P. Synergistic effects of plant growth promoting rhizobacteria and silicon dioxide nanoparticles for amelioration of drought stress in wheat. Plant Physiol. Biochem. 2021, 166, 160–176. [Google Scholar] [CrossRef]
- Bhat, M.A.; Kumar, V.; Bhat, M.A.; Wani, I.A.; Dar, F.L.; Farooq, I.; Bhatti, F.; Koser, R.; Rahman, S.; Jan, A.T. Mechanistic insights of the interaction of plant growth-promoting rhizobacteria (PGPR) with plant roots toward enhancing plant productivity by alleviating salinity stress. Front. Microbiol. 2020, 11, 1952. [Google Scholar] [CrossRef]
- Saleem, M.; Law, A.D.; Sahib, M.R.; Pervaiz, Z.H.; Zhang, Q. Impact of root system architecture on rhizosphere and root microbiome. Rhizosphere 2018, 6, 47–51. [Google Scholar] [CrossRef]
- Niu, X.; Song, L.; Xiao, Y.; Ge, W. Drought-tolerant plant growth-promoting rhizobacteria associated with foxtail millet in a semi-arid agroecosystem and their potential in alleviating drought stress. Front. Microbiol. 2018, 8, 2580. [Google Scholar] [CrossRef] [PubMed]
- Narayanasamy, S.; Thangappan, S.; Uthandi, S. Plant growth-promoting Bacillus sp. cahoots moisture stress alleviation in rice genotypes by triggering antioxidant defense system. Microbiol. Res. 2020, 239, 126518. [Google Scholar]
- Rashid, U.; Yasmin, H.; Hassan, M.N.; Naz, R.; Nosheen, A.; Sajjad, M.; Ilyas, N.; Keyani, R.; Jabeen, Z.; Mumtaz, S. Drought tolerant Bacillus megaterium isolated from semi-arid conditions induces systemic tolerance of wheat under drought conditions. Plant Cell Rep. 2022, 41, 549–569. [Google Scholar] [CrossRef]
- Vandana, U.K.; Rajkumari, J.; Singha, L.P.; Satish, L.; Alavilli, H.; Sudheer, P.D.; Chauhan, S.; Ratnala, R. : Satturu, V.; Mazumder, P.B. The endophytic microbiome as a hotspot of synergistic interactions, with prospects of plant growth promotion. Biology 2021, 10, 101. [Google Scholar] [CrossRef]
- Carlson, R.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A.; Hassen, A.I.; Labuschagne, N. Rhizobacteria-induced systemic tolerance against drought stress in Sorghum bicolor (L. ) Moench. Microbiol. Res. 2020, 232, 126388. [Google Scholar] [CrossRef]
- Barnawal, D.; Bharti, N.; Pandey, S.S.; Pandey, A.; Chanotiya, C.S.; Kalra, A. Plant growth-promoting rhizobacteria enhance wheat salt and drought stress tolerance by altering endogenous phytohormone levels and TaCTR1/TaDREB2 expression. Physiol. Plant. 2017, 161, 502–514. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Ullah, I.; Ali, S.; Kang, S.-M.; Lee, I.-J. Alleviation of salt stress response in soy-bean plants with the endophytic bacterial isolate Curtobacterium sp. SAK1. Ann. Microbiol. 2019, 69, 797–800. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.; Flores, A.; Rojas-Sánchez, B.; Urtis-Flores, C.A.; Morales-Cedeño, L.R.; Valencia-Marin, M.F.; Chávez-Avila, S.; Rojas-Solis, D.; Santoyo, G. Plant growth-promoting bacteria as bioinoculants: Attributes and challenges for sustainable crop improvement. Agronomy 2021, 11, 1167. [Google Scholar] [CrossRef]
- Gadhave, K.R.; Devlin, P.F.; Ebertz, A.; Ross, A.; Gange, A.C. Soil inoculation with Bacillus spp. modifies root endophytic bacterial diversity, evenness, and community composition in a context-specific manner. Microb. Ecol. 2018, 76, 741–750. [Google Scholar] [CrossRef]
- Hafez, E.M.; Alsohim, A.S.; Farig, M.; Omara, A.E.D.; Rashwan, E.; Kamara, M.M. Synergistic effect of biochar and plant growth promoting rhizobacteria on alleviation of water deficit in rice plants under salt-affected soil. Agronomy 2019, 9, 847. [Google Scholar] [CrossRef]
- Azmat, A.; Yasmin, H.; Hassan, M.N.; Nosheen, A.; Naz, R.; Sajjad, M.; Ilyas, N.; Akhtar, M.N. Co-application of biofertilizer and salicylic acid improves growth, photosynthetic pigments and stress tolerance in wheat under drought stress. PeerJ. 2020, 8, e9960. [Google Scholar] [CrossRef] [PubMed]
- Lally, R.D.; Galbally, P.; Moreira, A.S.; Spink, J.; Ryan, D.; Germaine, K.J.; Dowling, D.N. Application of endophytic Pseudomonas fluorescens and a bacterial consortium to Brassica napus can increase plant height and biomass under greenhouse and field conditions. Front. Plant Sci. 2017, 8, 2193. [Google Scholar] [CrossRef] [PubMed]
- Głodowska, M.; Schwinghamer, T.; Husk, B.; Smith, D. Biochar based inoculants improve soybean growth and nodulation. Agric. Sci. 2017, 8, 1048–1064. [Google Scholar] [CrossRef]
- Finkel, O.M.; Salas-González, I.; Castrillo, G.; Con-Way, J.M.; Law, T.F.; Teixeira, P.J.P.L.; Wilson, E.D.; Fitz-Patrick, C.R.; Jones, C.D.; Dangl, J.L. A single bacterial genus maintains root growth in a complex microbiome. Nature 2020, 587, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Vurukonda, S.S.K.P.; Giovanardi, D.; Stefani, E. Plant growth promoting and biocontrol activity of Streptomyces spp. as endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, D.P.; Prabha, R.; Sahu, P.K.; Gupta, V.K. Abiotic stress responses and microbe-mediated mitigation in plants: The omics strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef]
- del Carmen Orozco-Mosqueda, M.; Fadiji, A.E.; Babalola, O.O.; Glick, B.R.; Santoyo, G. Rhizobiome engineering: Unveiling complex rhizosphere interactions to enhance plant growth and health. Microbiol. Res. 2022, 16, 127137. [Google Scholar] [CrossRef]
- Meena, M.; Swapnil, P.; Zehra, A.; Aamir, M.; Dubey, M.; Goutam, J.; Upadhyay, R. Beneficial Microbes for Disease Suppression and Plant Growth Promotion. In Plant-Microbe Interactions in Agro-Ecological Perspectives; Springer: Singapore, 2017; pp. 395–432. [Google Scholar]
- Tabassum, B.; Khan, A.; Tariq, M.; Ramzan, M.; Khan, M.S.I.; Shahid, N.; Aaliya, K. Bottlenecks in commercialisation and future prospects of PGPR. Appl. Soil Ecol. 2017, 121, 102–117. [Google Scholar] [CrossRef]
- Thakur, M.P.; Van Der Putten, W.H.; Apon, F.; Angelini, E.; Vreš, B.; Geisen, S. Resilience of rhizosphere microbial predators and their prey communities after an extreme heat event. Funct. Ecol. 2021, 35, 216–225. [Google Scholar] [CrossRef]
- Lugtenber, B.J.; Malfanova, N.; Kamilova, F.; Berg, G. Plant growth promotion by microbes. Mol. Microb. Ecol. Rhizosphere 2013, 2, 561–573. [Google Scholar]
- Naik, K.; Mishra, S.; Srichandan, H.; Singh, P.K.; Sarangi, P.K. Plant growth promoting microbes: Potential link to sustainable agriculture and environment. Biocatal. Agric. Biotechnol. 2019, 21, 101326. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Bera, T.; Chakrabarty, A.M. Microbial siderophore a boon to agricultural sciences. Biol. Control. 2020, 144, 104214. [Google Scholar] [CrossRef]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial siderophores and their potential applications: A review. Environ. Sci. Pollut. Res. 2016, 23, 3984–3999. [Google Scholar] [CrossRef]
- Trivedi, P.; Batista, B.D.; Bazany, K.E.; Singh, B.K. Plant–microbiome interactions under a changing world: Responses, consequences and perspectives. New Phytol. 2022, 234, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef]
- Berg, G. Plant–microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef]
- Anli, M.; Baslam, M.; Tahiri, A.; Raklami, A.; Symanczik, S.; Boutasknit, A.; Ait-El-Mokhtar, M.; Ben-Laouane, R.; Toubali, S.; Rahou, Y.A. Biofertilizers as strategies to improve photosynthetic apparatus, growth, and drought stress tolerance in the date palm. Front. Plant Sci. 2020, 11, 516818. [Google Scholar] [CrossRef]
- García-Fraile, P.; Menéndez, E.; Rivas, R.L. Role of bacterial biofertilizers in agriculture and forestry. AIMS Bioen-gineering 2015, 2, 183–205. [Google Scholar] [CrossRef]
- Basu, S.; Kumar, G.; Chhabra, S.; Prasad, R. Role of soil microbes in biogeochemical cycle for enhancing soil fertility, New and Future Developments in Microbial Biotechnology and Bioengineering. Elsevier 2021, 3, 149–157. [Google Scholar]
Figure 1.
Increased vegetative growth and flowering in Astrophytum capricorne (A) and Astrophytum myriostigma (B) in plants supplemented with rhizobacteria on biochar substrate [21].
Figure 1.
Increased vegetative growth and flowering in Astrophytum capricorne (A) and Astrophytum myriostigma (B) in plants supplemented with rhizobacteria on biochar substrate [21].

Figure 2.
PGPB promotes plant growth through the production of siderophores, increasing iron availability, and producing hormones such as auxins, gibberellins, and cytokinin that modulate the hormone balance of the host plant. The direct mechanism includes biological nitrogen fixation (BNF) by the activity of the nitrogenase enzyme complex; the solubilization of inorganic phosphate in the soil; and the production of siderophores. The indirect mechanisms are attributed to PGPB's occupation of niches and the production of substances that repel phytopathogens and nematodes [21].
Figure 2.
PGPB promotes plant growth through the production of siderophores, increasing iron availability, and producing hormones such as auxins, gibberellins, and cytokinin that modulate the hormone balance of the host plant. The direct mechanism includes biological nitrogen fixation (BNF) by the activity of the nitrogenase enzyme complex; the solubilization of inorganic phosphate in the soil; and the production of siderophores. The indirect mechanisms are attributed to PGPB's occupation of niches and the production of substances that repel phytopathogens and nematodes [21].
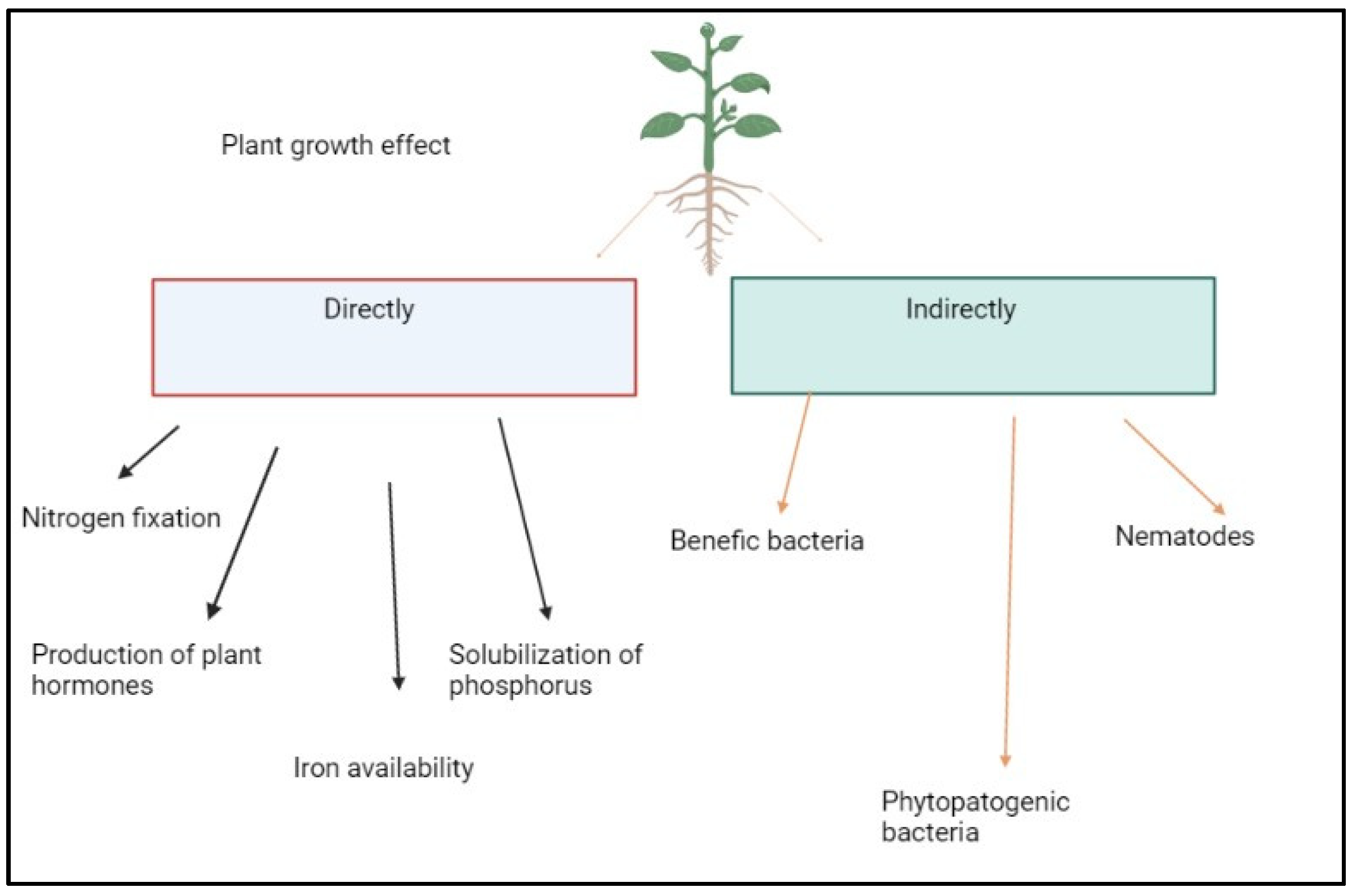
Table 1.
Bacterial genera and species that are drought resistant [25].
Table 1.
Bacterial genera and species that are drought resistant [25].
| Bacteria | Crop | Action Mechanism |
|---|---|---|
| Azospirillum sp. | Wheat | Highest amounts of N and auxin |
| Bacillus sp. | Grass | Responses of antioxidant system and early proline accumulation |
| Streptomyces sp. | Tomato | Increase the content of different sugars |
| Pseudomonas sp. | Arabidopsis | Higher ACC deaminase activity gibberellic acid, abscisic acid, indole acetic acid, and exopolysaccharide |
| Enterobacter sp. | Bean | Enhance proline, malondialdehyde, and antioxidant enzymes |
| Azospirillum brasilense | Wheat | Less accumulation of H2O2 with less enhanced production of proline and activities of catalase and peroxidase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



