Submitted:
24 July 2023
Posted:
25 July 2023
You are already at the latest version
Abstract
Interspecific competition between herbivorous insects is a major selection pressure affecting the distribution, abundance, and structure of their populations. Facilitator-mediated interactions, such as mutualism, can influence competition. Furthermore, the temporal dynamics of competitive relationships affect the interaction’s outcome. Here, we re-evaluated the data on the competition for space between two herbivorous insects commonly known as scales (Toumeyella martinezae and Opuntiaspis philococcus) in either the presence or absence of Liometopum apiculatum (a mutualistic species of T. martinezae) and its variations over time. We selected 27 Myrtillocactus geometrizans plants on which the studied insects were present; the plants were classified into one of five different conditions: either of the scale species were present on the plant, without its competitor; T. martinezae with neither its mutualistic species nor the competitor; and both scale species competing in either presence or absence of the mutualistic species. We kept a photographic record of each condition, measured the size of (as an indicator of the development stage) and area occupied by the individual scales, estimated the total coverage of each scale species, and assessed their relative occupation of space and their competitive intensity. We found temporal variations in competitive intensity. T. martinezae occupied more space during the first months, whereas O. philococcus did so towards the end of the study period. The population structure changed over time and between species, affecting the competitive interactions. In conclusion, the dynamics of competition change over time, and the mutualistic species has a positive effect on T. martinezae when the scales are in competition. However, temporal variations resulting from changes in the life cycle of the scales allow the two competitors to coexist in the same place at the same time.
Keywords:
competition
; scale insects
; mutualism
; temporal dynamics
; xerophytic shrubland
1. Introduction
Competition between herbivorous insects is one of the best documented interactions and is regarded as a major selection pressure affecting the distribution, abundance, and population structure of phytophagous insects, especially sessile, sap-feeding species, such as members of the Homoptera and Hemiptera [1,2,3,4]. Interference competition is the most common mechanism by which these organisms compete for space, with outcomes ranging from competitive exclusion to coexistence [5,6]. Major factors affecting the outcome of this interaction include the host plant [7,8,9], the intensity of the intraspecific competition [1], natural enemies [2,3,4], and environmental factors such as light exposure, humidity, and others [2].
These factors and interactions shape insect communities and are particularly important when the species utilizing the same resource also share similar ecological and phylogenetic characteristics [2,10]. Competition is predicted to be intense in these cases, commonly leading to the exclusion of one of the competitors. However, coexistence has been shown to also be a possible outcome as species face multiple trade-offs between survival and reproduction, which promote coexistence [11].
When competition fluctuates over time, species dominance can also change over time. In addition, the mobility of organisms can reduce the intensity of competition, even if competition only occurs during a short period of time in the lifecycle [5,6].
Homopterans provide good systems for studying competition. They exhibit life history traits that are conducive to intensely competitive interactions, including rapid population growth [12,13], which allows for populations with large numbers of individuals [13]; a sessile lifestyle during most of their lifecycle, which is a product of their diet [14,15,16,17,18]; and considerable plasticity in the number of generations per year [19]. Univoltine life cycles have been documented to be more common in low-temperature regions, whereas bivoltine or multivoltine cycles are found in warmer places [20,21]. Competition between species with different voltinism can lead to exclusion, because populations of multivoltine insects grow faster [22,23] and are therefore able to appropriate resources more efficiently, thus having a competitive advantage over bivoltine and univoltine species [24].
Our team has been studying the interactions between insects associated with Myrtillocactus geometrizans (Mart.), a cactus species endemic to Mexico [25,27]. Callejas-Chavero et al. [27] described the arthropod community associated with this cactus in various populations in Mexico, and recorded the highest species richness in the locality of Huichapan (State of Hidalgo). Two sessile herbivores often co-occurred on the same plants at this locality: Opuntiaspis philococcus (Cockerell), which is also known as the armored scale due to the waxy cover that protects its body [28], and Toumeyella martinezae [29], known as the soft scale [30]. The two species have similar lifecycles lasting 12–24 weeks and encompassing three developmental stages (egg, nymph, and adult), with four distinct nymphal stages [31]. Both species feed on sap and compete for space [26]. O. philococcus is a multivoltine species that undergoes four reproductive events per year, whereas T. martinezae is bivoltine [32,33].
T. martinezae maintains a mutualistic interaction with the escamolera ant Liometopum apiculatum (Mayr), which attends, cares for, and protects the scale from natural enemies [34]. This mutualistic relationship is particularly intense during nymphal Stages two, three, and four, when scales produce the greatest amount of honeydew, a substance that ants feed on [26,30,32]. Thereafter, when the females become adults and grow into their final size, the ants only groom them.
A previous study evaluated the effect of these herbivores on the plant’s fitness and examined the relationship between the scales’ competition and the intensity of herbivory on garambullo plants [26]. We found that the armored scale is more affected by the presence of its competitor than the soft scale. The average abundance of O. philococcus decreased when it coexisted with the soft scale, while the abundance of the soft scale decreased to a lesser extent. The competitive advantage of the soft scales depends on the presence of the mutualistic ant. When ants were excluded, the abundance of the soft scale decreased, while the abundance of the armored scale increased. However, despite this asymmetry, the armored scale is not excluded from the garambullo plants. In light of this, we decided to re-evaluate the results of the study, incorporating two measurements of the space occupied by each species, rather than a record of the abundance of each species. Although there is a close relationship between abundance and space occupied, many small individuals may not take up enough space to exclude other competing species. In the same way, the temporal dynamics caused by the ant and by the phenology of the scales can affect the intensity of the competition and, therefore, the results of the interaction. Therefore, the effects of competition on the population structure of both species were re-evaluated.
This study aimed to evaluate the space occupied by and the changes in the population structure of the competing scales Toumeyella martinezae and Opuntiaspis philococcus in either the presence or absence of the mutualistic ant Liometopum apiculatum, and their variations over time. We pursued the following objectives: (1) to examine the competition over time, (2) to evaluate the effect of competition on the scales’ population structure (using body size as a proxy for development stage), (3) to determine the temporal dynamics of competition between the scales in the presence or absence of the mutualistic ant, and 4) to determine if the coexistence of the two scales is possible and what would determine such an outcome.
2. Materials and Methods
2.1. Study site and organisms
The study was carried out in a xerophytic shrubland near the town of Zequetejé in the municipality of Huichapan, State of Hidalgo, Mexico (20°22’47.2”N, 99°36’23.9”W). The elevation in the area is 2100 m above sea level, and the climate is dry with a rainy summer season (June through September).
Myrtillocactus geometrizans (Mart. ex Pfeiff.) Console 1897 (Cactaceae) is a cactus species endemic to Mexico that is very abundant in this area. It inhabits arid and semi-arid regions, especially those with xerophytic vegetation [25]. It has ascending stems that can reach up to 5 m in height [35,36,37].
Toumeyella martinezae Kondo and González 2014, the soft scale, is a sessile insect (Hemiptera: Coccidae) with a convex, oval body measuring 4.2–7.2 mm in length and 3.1–5.7 mm in width [29]. This species has only been found associated with M. geometrizans in the Huichapan locality (it is probably also found on the cactus Pachycereus marginatus, according to field observations) and in trophobiosis with the ant Liometopum apiculatum Mayr 1870. The ant attends to the scales day and night, feeding on the honeydew they produce; in exchange, it protects the scales from its natural enemies, grooms them, and transports them to sites suitable for their establishment and development [29,31,38,39].
Opuntiaspis philococcus Cockerell 1893, the armored scale, is another sessile Hemipteran (Diaspididae) with an elongated body measuring 3.5–4 mm in length and 1.5–2 mm in width, with white-to-grey armor exhibiting a slight longitudinal crest and a flattened edge [40,41]. This scale species is polyphagous and has been recorded on different hosts in the genera Agave [42] and Opuntia [41]. O. philococcus does not produce honeydew and does not establish trophobiosis with ants or any other mutualistic relationships [33,41].
2.2. Field conditions
We haphazardly selected 27 adult Myrtillocactus geometrizans plants in a ~1.5 ha area covered by xerophytic shrubland containing a total of approximately 250 garambullo plants. This design is the same used by Callejas-Chavero et al., [26] with the same plants. The average height of the selected plants was 2.6 0.4 m, and each bore 387 184 branches on average. The plants were classified into one of the five conditions shown in Figure 1: either scale species was present on the plant, without the competitor; T. martinezae was present with neither its mutualistic species nor the competitor; and both scale species competing in either the presence or absence of the mutualistic species. Six branches were randomly chosen on each of five plants in each condition, and a permanent 72 cm2 quadrat was delimited on each of them (to be used as a sampling unit with a constant area; Figure S1B–E in the Supplementary Information). The quadrats were located so that the conditions described were readily evident. Ants were excluded (in Conditions 3 and 5, see Figure 1) by placing a ring of solid, unscented petroleum jelly at the base of each branch to prevent them from moving onto the branch where the permanent quadrat was located. Ants that were already present prior to placing the ring were removed with a brush and tweezers. The condition of O. philococcus without competition plus L. apiculatum was not included, as this interaction does not occur in the study site under natural conditions (Figure 1, first condition). The scales’ mobile stage was not recorded in situ due to the high mobility and small size of these organisms.
In total, 162 permanent quadrats were established. We considered each quadrat as an independent sampling unit and only regarded the competitive interaction between the herbivores. Each quadrat was photographed every 6 weeks from October 2017 to April 2018 for a total of five censuses of 162 quadrats (N = 810). All the photographs were taken 15 cm above the quadrat’s center and included a 5 mm reference scale. The size of each individual scale and the abundance of each scale species in each photograph were recorded using the program MOTIC IMAGES 2000 Version 1.3 Ink for Windows [43].
Given the different body shapes of the two scale species, competition between them was examined in terms of the area occupied by the individuals rather than the number of individuals. A sample of 138 individuals of Opuntiaspis philococcus and 168 of Toumeyella martinezae was taken. The anteroposterior length and area covered by each scale individual were measured using the software MOTIC IMAGES 2000 Version 1.3 Ink for Windows [43]. Third-order polynomial models relating the scales’ anteroposterior length and area covered were constructed separately for each species. These models are shown in the Supplementary Material. The model for the soft scale had an adjusted R2 of 0.9701 (F3,164= 1810, P < 0.0000); the model for the armored scale had an adjusted R2 of 0.9568 (F3,134= 1012, P < 0.00001). Analyses were performed using the base R v4.1.1 (2021) software. The models were then used to estimate the area occupied by each individual in the sample (n = 42512 in total; 25393 soft scales and 17119 armored scales).
2.3. Measures of competition and statistical analyses
As we have already mentioned, in a previous study, we evaluated the competition between these two species of scale using abundance (the total number of individuals in the quadrat) [26]. However, we consider that abundance may be a biased measure, since the number of individuals does not necessarily measure the space they occupy (many small individuals can take up less space than one large one). In this sense, the direct estimation of the size of each individual defines the space it occupies more clearly. For this reason, we decided to analyze the data and considered the space used by each species as a new measurement of competition.
Since scale insects are sessile for most of their lifespan and usually do not interfere with each other through chemical substances or aggressive contact [13], we evaluated their competition for space using two different approaches. The first approach examined the total area occupied (OA) by each species in each quadrat (the sum of the area covered by each individual). Because these species have different shapes, as already mentioned, with this estimate, we could know how much area was covered by each species. A zero-inflated generalized linear mixed model with a gamma distribution and a logarithmic link was fitted separately for each species. The response variable was the species’ OA; the predictive variables were the total area occupied by the competing species, the presence/absence of the ant, and the month of the year when the measurement was made. As the quadrats’ data are nested within individual cacti, each cactus’s identity was entered into the model as a random effects factor. These models were fitted using the R package TMB. Secondly, we evaluated the Space Occupation Index (SOI) as a measure of space appropriation [44,45,46]. SOI values range from 0 to 100%, with those close to 100 denoting a greater occupation of space by the competitor and values close to 50% for both competing species indicating an equal appropriation of space [47], the space occupation index was calculated as follows
where:
Acc = ;
Bcc = .
2.4. Population structure
Population structure was determined on the basis of the criteria proposed by Martínez-Hernández et al. [31] for T. martinezae, and by Islas-Estrada [33] for O. philococcus. Those authors pointed out that, in these species, body size (i.e., anteroposterior length) indicates the development stage. Thus, the life cycle of Toumeyella martinezae comprises three distinct development stages with the following size categories (lengths): egg, 179 μm; first-stage nymph, 0.6–0.7 mm; second-stage nymph, 0.71–1.21 mm; third-stage nymph, 1.22–3.0 mm; fourth-stage nymph, 3.1–5.0 mm; and adult, 5.1–7.82 mm [31]. The lifecycle of Opuntiaspis philococcus comprises three development stages (egg, nymph, and adult) with the following size categories [41]: first-stage nymph, 0.1–0.7 mm long; second-stage nymph, 0.71–1.1 mm; third-stage nymph, 1.11–1.9 mm; fourth-stage nymph, 1.91–2.9 mm; and adult, 2.91–5.3 mm. All the scale individuals were measured and then assigned to the corresponding size categories or development stages.
To examine the effect of competition on the population structure, the changes over time in the number of individuals in each size category were analyzed by means of generalized linear models (GLM) fitted separately for each scale species. The number of live individuals of a given size recorded on a given census date (the response variable) was modeled as a function of age (size) category, the census date (five dates), and condition (with or without the ant, in competition, or without competition), plus the second- and third-order interactions between these variables. The models were built assuming a Poisson probability distribution and using a logarithm link function with the software SPSS v25 [49].
3. Results
3.1. Competition for space
The space occupied by the soft scale at the beginning of the experiment (in October) was larger than that occupied by the armored scale, either separately or in competition, and in the presence or absence of the mutualistic ant (Figure 2A,B). However, the space occupied by each of the two species changed drastically at the end of the experiment (in April). The area covered by the soft scale (without competition) experienced reductions of 97% and 92% (in the presence or absence of the ant, respectively), as they occupied only 3% and 8% of the space they had held at the beginning. The space occupied by the armored scale (without competition) was reduced by 75% compared with the space initially occupied (Figure 2A). When in competition in the presence of the mutualistic ant, the soft scale kept about 61% of the area it initially covered, while the armored scale retained only 52%. By contrast, in absence of the ant, the soft scale kept only 2% of the area it initially covered, while the armored scale retained 62% (Figure 2B).
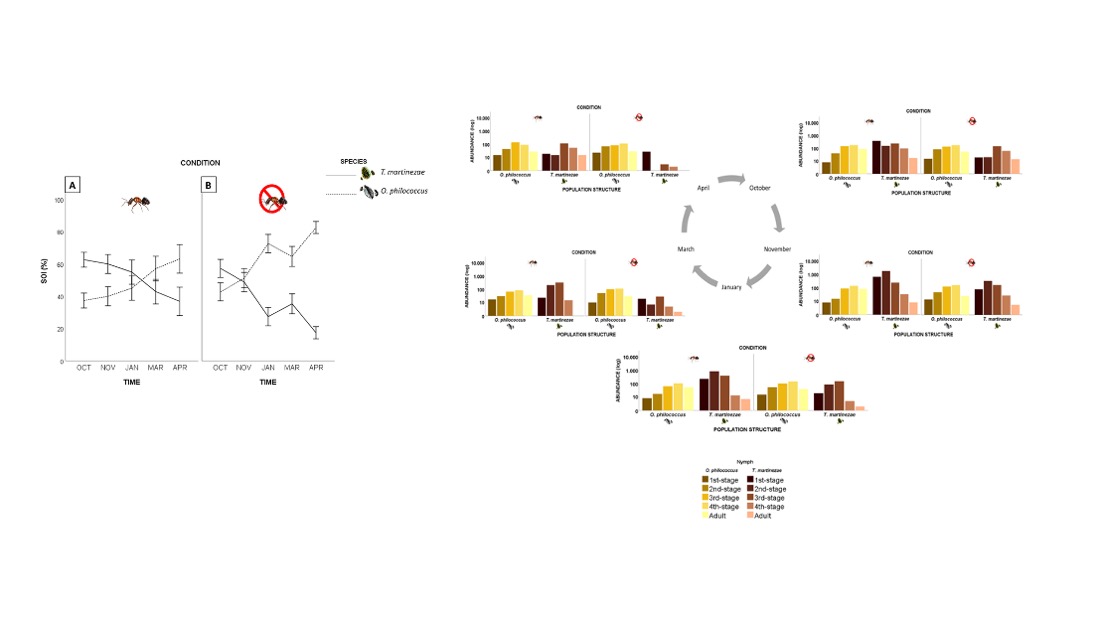
The values of the SOI showed that the space occupied by the soft scale in competition in the presence of the ant was noticeably reduced after January (Figure 3A). In the absence of the ant, the space occupied decreased from November to April (Figure 3B). The armored scale showed the opposite pattern, as its coverage increased towards the end of the sampling period, either in the absence or the presence of the ant (Figure 3A,B).
In summary, the area occupied by the armored scale (GLM fitted R2=0.297, AIC= 2267.383, BIC= 2326.101) showed that (a) the presence of the soft scale caused a significant reduction in the coverage of the armored scale, (b) the presence of the ant had no significant effect on the armored scale (Table 1), and (c) the coverage of the armored scale decreased over time, except in November (Table 1). The soft scale’s model showed that (a) the presence of the armored scale had no significant effect on the area occupied by the soft scale, (b) the presence of the ant had a significant effect only in April, and (c) the area occupied by the soft scale increased a little in January, March, and April (Table 1) (GLM R2=0.464, AIC=2166.887, BIC=2236.615).
3.2. Population structure
The first graphs in Figure 4 (A,B) show the data of the population structure under the control condition, that is, the natural conditions found in the field. The results for O. philococcus showed that there were similar amounts of individuals in all categories at all times, but for T. martinezae, a decrease in the amount of individuals was clearly seen, especially for the last categories towards the end of the study period. However, in the presence of both its competitor and the mutualistic ant, the population structure of O. philococcus remained stable, while its abundance decreased to fewer than 200 individuals in each size category. Size categories 1, 2, and 3 were the ones that decreased the most. The same effect was observed when O. philococcus was in competition with T. martinezae in the absence of L. apiculatum, with the decreases being greater for Category 4 and adults (Figure 4 A,B).
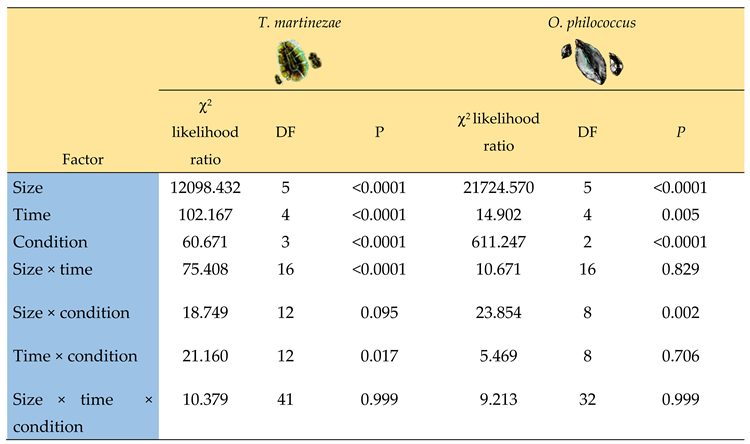
Our results showed the significant effects of competition, the census date, and the presence of the mutualistic ant on the population structure of the two species. The number of live T. martinezae individuals was significantly affected by all these factors (Table 2). The abundance of all the size categories of the soft scale showed a marked reduction in March and April under most conditions, except when in competition without the ant (Table 2 and Figure 4B). This change was due to the time taken for an individual to pass from one stage to another, combined with scarce recruitment at that time of the year [31].
The number of live O. philococcus individuals was significantly affected by competition and the presence/absence of the ant, and varied over time, but there was no interaction between these factors (Table 2). That is, its population structure did not change significantly between census dates. When O. philococcus occurred in the absence of its competitor, the population comprised scales in all size categories, with high and similar abundances, although the categories best represented in all census dates were for Stages 2, 3, and 4 (Figure 4A).
4. Discussion
As already found in a previous study, we observed that when the two phytophagous species shared the same space at the same time, the soft scale was the better competitor due to its mutualistic relationship with the ant; this advantage disappeared when the ant was excluded and the competition between T. martinezae and O. philococcus became symmetric. Although T. martinezae is the superior competitor, it is not always the dominant species, being dominant only when its abundance increases in the presence of the ant, particularly at the start of the competitive interaction. The competitive advantage of species maintaining mutualistic relationships has been widely documented [48,50,51,52], and it usually leads to competitive exclusion of the non-mutualistic species [51]. In this case, however, T. martinezae did not exclude O. philococcus, as there was no truly dominant species in this system, with the abundance of both species changing over time.
The two species differed in both the area they occupied (OA) and their space occupation index (SOI), and these changed over time. At the beginning of the study, when both species were in their early developmental stages, they occupied similar amounts of space. As time passed and they entered adulthood, the space occupied by the soft scale decreased gradually, and this pattern continued until the end of the study period. By the end of the life cycle of both species, however, the armored scale occupied more space, likely because the next generation of scales had begun to become established; this was confirmed by the results of the population structure analysis. The population structure of T. martinezae varied over time, depending on whether its mutualist was present or not, whereas the population structure of O. philococcus remained relatively constant. In these circumstances, O. philococcus would be expected to be the dominant species as it does not depend on the ant and is able to maintain its population structure. However, its growth rate is slower and, due to its smaller size, it occupies less space. In contrast, by being tended by its mutualistic ant, the population of T. martinezae can recover quickly, allowing it to appropriate more space. Another interesting finding was that the space occupied by the soft scale decreased over time even without competition. This reduction might be due to the fact that the scale’s lifecycle ends and recruitment becomes scarce and intermittent, leading to a lower abundance of all the developmental stages (unpublished data [32]) and less intense competition.
The presence of the mutualistic ant conferred a competitive advantage on T. martinezae by increasing its capacity to appropriate space. On the other hand, although O. philococcus was less able to occupy space than the soft scale (due to the lack of a mutualistic interaction), this did not affect its population structure or its permanence in the system. This has seldom been recorded in phytophagous species [53], although a similar result was found by Martínez [54]. An explanation based on the number of reproductive events that each species has during the year might be relevant for this case, since the recruitment rate of multivoltine species is constant, while it is intermittent in univoltine species [22,23].
Humidity and temperature have been reported to affect the number of reproductive events per year in some insect species [55]. When the weather is warmer and drier, insects tend to have several reproductive events (bivoltine or multivoltine) [19,56,57]. This rule seems to hold for the two competing species in our study system, as the armored scale has four events per year and is regarded as multivoltine, while the soft scale is bivoltine, based on field observations. This factor can cause variations in the space occupied and the competitive intensity over time [19,54,58,59,60]. As T. martinezae is a bivoltine species, its population size will decrease in some months of the year, thus allowing O. philococcus to recover from the adverse effects of its competitor. Having four reproductive events per year allows O. philococcus to maintain a stable population size over time and not be competitively excluded from the system. In addition to differences in spatial occupancy (OA) caused by the different number of reproduction events, the two scale species tended to gradually occupy different areas, as T. martinezae became more concentrated in the lower parts of the branch and O. philococcus in the upper parts, which also facilitated their coexistence [54]. Another important point to highlight is that the mobile stage of both species was not included in this study. The main reason for this is that the mobile stage is very difficult to observe in the field due to the insects’ mobility and small size [32,54]. Although the effect of mobile stages is unknown, they have been observed to usually establish themselves in areas where there is no other scale, which would help to reduce the effects of competition [8,9]. Further studies aimed at assessing the role of the mobile stage in competitive interactions would be useful.
When the number of generations per year and mutualism of the soft scale are jointly examined, the trade-off [61] between supporting the ant and reproducing becomes evident. Supporting the mutualistic ant involves a cost that reduces the number of reproductive events the soft scale can afford per year [53,61,62]. On the other hand, the multivoltine lifecycle of the armored scale might be possible because this species does not invest energy in maintaining a relationship with a mutualistic species and is therefore able to allocate all its energy to reproduction [63,64,65]. If the two species can thrive in different time windows, then they could coexist at the same site at the same time [66,67]. Having a specific measure that can be tracked over time would be useful for monitoring changes in the population structure and the intensity of competition between the species involved. This has been reported for other systems, where changes in the species’ life histories have been shown to modify interspecific interactions over time, progressing from facilitation to competition, to a reduction in the competitive intensity because of environmental changes, and finally ending with the coexistence of both competitors [53,68,69]. One of the necessary conditions for coexistence is that the species whose abundance has declined must possess the ability to re-enter the space where it was replaced [70]; this is what seems to happen here with the armored scale [54].
5. Conclusions
In this century, ecology faces new challenges and new questions. Among these, it is considered fundamental to know how many species can coexist in the same space and time, and the mechanisms for this coexistence [71]. In this context, interspecific competition is an important factor for understanding the changes in diversity in communities, since the coexistence in time and space of species that use the same resources is intense [2]. In particular, insects can be a good model for analyzing coexistence in competition, given the fact that it is common to find several species in the same place exploiting the same resources [3].
In this sense, we can say that there is an ecological timeframe during which coexistence is feasible, but this does not necessarily match the evolutionary time required to maintain coexistence. When the possibilities for spatial segregation are limited, coexistence can be favored for an indefinite period of time [72,73]. Interspecific competition is an important factor, since the coexistence of species in time and space entails the utilization of the same resources [2,3,71].
Thus, in summary, the occupation of space by T. martinezae and O. philococcus changes over time, with competition affecting the population structure of scales. Even though the presence of Liometoum apiculatum makes T. martinezae a better competitor when the soft scale is not abundant, O. philococus can re-enter the system, thus maintaining coexistence.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure 1. A) Distribution of garambullos in the field, each color represents a condition; Data and models that related total length with surface occupied by each species of scale insects.
Author Contributions
ACCH, AJMO, and CFVM designed the study and executed the fieldwork; ACCH provided information on the biology and ecology of the species; SSC, AJMO, and CFVM carried out the statistical analyses; AJMO, SSC, ACCH, and CFVM wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by grants SIP-20181233 and SIP-20195598 from Instituto Politécnico Nacional
Data Availability Statement
The data used for this study are available upon request from the corresponding authors.
Acknowledgments
We thank the authorities of Ejido Zequetejé for allowing us to conduct fieldwork on their land. Víctor Hugo López-González drew Figure 1, and Elena Flores-Callejas drew the graphic abstract. We thank Arturo Flores for his critical reading of the manuscript and valuable suggestions. María Elena Sánchez-Salazar and Salvador Sánchez-Colón translated the manuscript into English.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Connell, J. H. On the prevalence and relative importance of interspecific competition: evidence from field experiments. Am. Nat 1983, 122, 661–696. [Google Scholar] [CrossRef]
- Denno, R. F.; McClure, M. S.; Ott, J. R. Interpecific interactions in phytophagous insects: competition reexamined and resurrected. Ann. Rev. Entomol. 1995, 40, 297–331. [Google Scholar] [CrossRef]
- Bell, G. The co-distribution of species in relation to the neutral theory of community ecology. Ecology 2005, 86, 1757–1770. [Google Scholar] [CrossRef]
- Kaplan, I.; Denno, R. F. Interspecific interactions in phytophagous insects revisited: a quantitative assessment of competition theory. Ecol. Lett. 2007, 10, 977–994. [Google Scholar] [CrossRef]
- Mooney, K.; Jones, P.; Agrawal, A. Coexisting congeners: demography, competition, and interactions with cardenolides for two milkweed-feeding aphids. Oikos 2008, 117, 450–458. [Google Scholar] [CrossRef]
- Smith, R. A.; Mooney, K. A.; Agrawal, A. Coexistence of three specialist aphids on common milkweed, Asclepia syriaca. Ecology 2008, 89, 2187–2196. [Google Scholar] [CrossRef] [PubMed]
- Inbar, M.; Doostdar, H.; Leibee, G. L.; Mayer, R. T. The role of plant rapidly induced responses in asymmetric interspecific interactions among insect herbivores. J. Chem. Ecol. 1999, 25, 1961–1979. [Google Scholar] [CrossRef]
- Denno, R. F.; Peterson, M. A.; Gratton, C.; Cheng, J.; Langellotto, G. A.; Huberty, A. F.; Finke, D. L. Feeding-induced changes in plant quality mediate interspecific competition between sap-feeding herbivores. Ecology 2000, 81, 1814–1827. [Google Scholar] [CrossRef]
- Denno, R.F.; Kaplan, I. Plant-mediated interactions in herbivorous insects: mechanisms, symmetry, and challenging the paradigms of competition past. In Ecological Communities: Plant Mediation in Indirect Interaction Webs; Ohgushi, T.; Craig, T.P.; Price, P.W. Cambridge University Press, Cambridge, UK, 2007; pp. 19–50.
- Yamamichi, M.; Gibbs, T.; Levine, J. M. Integrating eco-evolutionary dynamics and modern coexistence theory. Ecol. Lett. 2022, 25, 2091–2106. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A. A. A scale-dependent framework for trade-offs, syndromes, and specialization in organismal biology. Ecology 2020, 101, e02924. Available online: https://www.cabdirect.org/cabdirect/abstract/19610501730. [CrossRef] [PubMed]
- Ito, Y. Ecological studies on population increase and habitat segregation among barley aphids. Bull. Natn. Inst. Agric. Sci. 1960, C11, 45–130. [Google Scholar]
- Denno, R. F.; Roderick, G. K. Density-Related dispersal in planthoppers : Effects of interspecific crowding. Ecology 1992, 73, 1323–1334. [Google Scholar] [CrossRef]
- McClure, M. S.; Price, P. W. Competition among sympatric Erythroneura leafhoppers (Homoptera: Cicadellidae) on America sycamore. Ecology 1975, 56, 1388–1397. [Google Scholar] [CrossRef]
- DeBach, P.; Hendrickson, R. M.; Rose, M. Competitive displacement: extinction of the yellow scale, Aonidiella citrina (Coq.) (Homoptera: Diaspididae), by its ecological homologue, the California red scale, Aonidiella aurantii (Mask.) in Southern California. Hilgardia 1978, 46, 1–35. [Google Scholar] [CrossRef]
- McClure, M. S. Competition between exotic species: scale insects on Hemlock. Ecology 1980, 61, 1391–1401. [Google Scholar] [CrossRef]
- McClure, M. S. Biology, population trends, and damage of Pineus boerneri and P. coloradensis (Homoptera: Adelgidae) on Red Pine. Environ. Entomol. 1989, 18, 1066–1073. [Google Scholar] [CrossRef]
- Gullan, P. J.; Martin, J. H. Sternorrhyncha: (jumping plant-lice, whiteflies, aphids, and scale insects). In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé R., T., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2009; pp. 957–967. [Google Scholar] [CrossRef]
- McClure, M. S. Effects of voltinism, interspecific competition and parasitism on the population dynamics of the hemlock scales, Fiorinia externa and Tsugaspidiotus tsugae (Homoptera: Diaspididae). Ecol. Entomol. 1981, 6, 47–54. [Google Scholar] [CrossRef]
- Gullan, P. J.; Kosztarab, M. Adaptations in scale insects. Annu. Rev. Entomol. 1997, 42, 23–50. [Google Scholar] [CrossRef]
- Forrest, J. R. Complex responses of insect phenology to climate change. Curr. Opin. Insect Sci. 2016, 17, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Zeuss, D.; Brunzel, S.; Brandl, R. Environmental drivers of voltinism and body size in insect assemblages across Europe. Glob. Ecol. Biogeogr. 2017, 26, 154–165. [Google Scholar] [CrossRef]
- Just, M. G.; Frank, S. D. Thermal tolerance of gloomy scale (Hemiptera: Diaspididae) in the Eastern United States. Environ. Entomol. 2020, 49, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, Y.; Ezoe, H.; Yamauchi, A. Evolutionarily stable seasonal timing of univoltine and bivoltine insects. In Insect life-cycle polymorphism, Series Entomologica, Danks, H. V., Ed.; Springer: New York, NY, USA, 1994, 43, pp. 69–89. [Google Scholar] [CrossRef]
- Arias, S.; Gama, S.; Guzman, L. Flora del Valle de Tehiacán-Cuicatlán. Fascículo 14. Cactaceae A.L. Juss. Universidad Nacional Autónoma de México. DF Ciudad de México, México, 1997; pp. 146. Available online: http://www.ibiologia.unam.mx/BIBLIO68/fulltext/fasiculosfloras/fas14.pdf.
- Callejas-Chavero, A.; Vargas-Mendoza, C. F.; Flores-Martínez, A.; Martínez-Hernández, D.; Moncada-Orellana, A.; Dias-Quiñones, J. Y. Herbivory in cacti: Fitness effects of two herbivories, one tending-ant on Myrtillocactus geometrizans (Cactaceae). In Evolutionary Ecology of Plant-Herbivore Interaction; Nuñez-Farfán, J.; Valverde, P., Eds; Springer: New York, NY, USA; 2020 pp. 109–134. [CrossRef]
- Callejas-Chavero, A.; Martínez, D. G.; Nicolás, D. E. Diversidad de artrópodos asociados a Myrtillocactus geometrizans (MART) en tres poblaciones del Valle de México. In Memorias del XLVIII Congreso Nacional de Entomología; Equihua, M.A., Estrada, V.E, Acuña, S.J., Chaires, G.P., Eds.; Sociedad Mexicana de Entomología: Ciudad de México, México; 2013; pp. 685–689. Available online: http://www.entomologia.socmexent.org/revista/2013/EC/685-689.pdf.
- Beardsley Jr, J. W.; Gonzalez, R. H. The biology and ecology of armored scales. Annu. Rev. Entomol. 1975, 20, 47–73. Available online: https://www.antwiki.net/wiki/images/4/40/Diaspididae_armored_scales_annurev1975.pdf. [CrossRef] [PubMed]
- Kondo, T.; González, H. A new species of Toumeyella Cockerell (Hemiptera: Coccidae) on Myrtillocactus geometrizans (Cactaceae) from Mexico with a checklist of known species of Toumeyella in the world. Insecta Mundi 2014, 0396, 1–10. Available online: https://digitalcommons.unl.edu/cgi/viewcontent.cgi?article=1899&context=insectamundi.
- Martínez, D.G. Efecto de Liometopum apiculatum (Hymenoptera: Formicidae) sobre la tasa de parasitoidismo de Toumeyella martinezae (Hemiptera: Coccidae) asociados a Myrtillocactus geometrizans (Cactaceae) en un Matorral Xerófilo de Huichapan, Hidalgo. Bachelor’s thesis, Instituto Politécnico Nacional. Ciudad de México, México, 2015.
- Martínez-Hernández, D. G.; Arriola-Padilla, V. J.; Callejas-Chavero, A.; González-Hernández, H.; Gijón-Hernández, A. Biología de la escama blanda Toumeyella martinezae Kondo y Gonzalez (Hemiptera: Coccidae) del Garambullo (Myrtillocactus geometrizans) (Mart. ex Pfeiff) Console) en Huichapan, Hidalgo. Entomol. Mex. 2015, 2, 172–179. Available online: http://www.entomologia.socmexent.org/revista/2015/BHN/PAG172-179.pdf.
- González-Villa, H. Efecto del parasitoidismo y mutualismo sobre la demografía de Toumeyella martinezae (Hemiptera: Coccidae) asociada a Myrtillocactus geometrizans (Cactaceae) en un matorral xerófilo de Huichapan, Hidalgo. Master Degree, Instituto Politécnico Nacional. Ciudad de México, México, 2021.
- Islas-Estrada, S. A. Ciclo de vida de Opuntiaspis philococcus (Cockerell, 1893) y sus enemigos naturales, asociados a Myrtillocactus geometrizans en Hidalgo, México. Bachelor’s thesis, Instituto Politécnico Nacional. Ciudad de México, México, 2020.
- Velasco-Corona, C.; Corona-Vargas, M. del C.; Peña-Martínez, R. Liometopum apiculatum (Formicidae: Dolichoderinae) y su relación trofobiótica con Hemiptera Sternorrhyncha en Tlaxco, Tlaxcala, México. Acta Zool. Mex. 2007, 23, 31–42. Available online: http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S0065-17372007000200003.
- Bravo-Hollis, H. Las cactáceas de México (Vol. 1) Universidad Nacional Autónoma de México: Ciudad de México, México,. 1978, 1, pp. 743.
- Bustamante, E.; Búrquez, A. Fenología y biología reproductiva de las cactáceas columnares. Cact. Suc. Mex. 2005, 50, 68–80. Available online: http://web.ecologia.unam.mx/cactsucmex/CACTACEAS2005_3.pdf.
- Hernández, M.; Terrazas, T.; Delgado, A.; Luna, M. Los estomas de Myrtillocactus geometrizans (Mart. ex. Pfeiff.) Console (Cactaceae): Variación en su área de distribución. Rev. Fitotec. Mex. 2007, 30, 235–240. Available online: https://www.redalyc.org/pdf/610/61003004.pdf. [CrossRef]
- Del Toro, I.; Pacheco, J. A.; Mackay, W. P. Revision of the ant genus Liometopum (Hymenoptera: Formicidae). Sociobiology 2009, 53, 299–369. Available online: https://www.researchgate.net/publication/260866074_Revision_of_the_Ant_Genus_Liometopum_Hymenoptera_Formicidae.
- Cruz-Labana, J. D.; Tarango-Arámbula, L. A.; Alcántara-Carbajal, J. L.; Pimentel-López, J.; Ugalde-Lezama, S.; Ramírez-Valverde, G.; Méndez-Gallegos, S. J. Habitat use by the escamolera ant (Liometopum apiculatum Mayr) in Central Mexico. Agrociencia 2014, 48, 569–582. Available online: http://www.colpos.mx/agrocien/Bimestral/2014/ago-sep/art-1.pdf.
- González, R.H. Estrategias de control de la Escama de San José. Rev. Frutíc. 1980, 1, 11–14. [Google Scholar]
- Hamon, A. B. Opuntiaspis philococcus (Cockerell) (Homoptera: Coccoidea: Diaspididae). Entomology Circular 1980, 214, 1–2. [Google Scholar]
- Rodríguez, W. D.; Navarrete-Heredia, J. L.; Vásquez-Bolaños, M.; Rodríguez-Macías, R.; Briceño-Félix, G. A.; Coronado Blanco, J. M.; Ruíz-Cancino, E. Insects associated with the genus Agave spp. (Asparagaceae) in Mexico. Zootaxa 2019, 4612, 451–493. [Google Scholar] [CrossRef] [PubMed]
- Motic Images 2000, version 1.3; Motic China Group Co LTD) and statistical data analyses were processed using Microcal Origin 8.5 Pro package software. Available online: https://imaris.oxinst.com/packages?gad=1&gclid=Cj0KCQjwho-lBhC_ARIsAMpgMoePWZaj3hwhbqh1qml-wRthy575_wd8lWYBEXiv_bMBupO_1An5sU4aArSiEALw_wcB (accessed on 6 Agust 2018).
- Weigelt, A.; Jolliffe, P. Indices of plant competition. J. Ecol. 2003, 91, 707–720. [Google Scholar] [CrossRef]
- Ghosh, P. K.; Manna, M. C.; Bandyopadhyay, K. K.; Ajay, A. K.; Tripathi, R. H.; Wanjari, R. H.; Hati, K. M.; Misra, A. K.; Acharya, C. L.; Subba Rao, A. Interspecific interaction and nutrient use in soybean/sorghum intercropping system. Agron. J. 2006, 98, 1097–1108. [Google Scholar] [CrossRef]
- Grace, J. B. On the measurement of plant competition intensity. Ecology 1995, 76, 305–308. [Google Scholar] [CrossRef]
- Yates, F.; Dutton, S. J. A simple procedure for the analysis of experiments on mixtures of varieties or species of agronomic interest. J. Agric. Sci. 1988, 110, 13–24. [Google Scholar] [CrossRef]
- Pekas, A.; Aguilar, A.; Tena, A.; Garcia-Marí, F. Influence of host size on parasitism by Aphytis chrysomphali and A. melinus (Hymenoptera: Aphelinidae) in Mediterranean populations of California red scale Aonidiella aurantii (Hemiptera: Diaspididae). Biol. Control. 2010, 55, 132–140. [Google Scholar] [CrossRef]
- IBM Corp. Released 2017. IBM SPSS Statistics for Windows, Version 25.0. Armonk, NY: IBM Corp.
- Stadler, B.; Dixon, A. F. G. Ecology and evolution of aphid-ant interactions. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 345–372. [Google Scholar] [CrossRef]
- Mehrparvar, M.; Zytynska, S. E.; Balog, A.; Weisser, W. W. Coexistence through mutualist-dependent reversal of competitive hierarchies. Ecol. Evol. 2018, 8, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J. A.; López-Gallego, E.; La-Spina, M. The impact of ant mutualistic and antagonistic interactions on the population dynamics of sap-sucking hemipterans in pear orchards. Pest Manag. Sci. 2020, 76, 1422–1434. [Google Scholar] [CrossRef] [PubMed]
- Hill, M. G.; Mauchline, N. A.; Hall, A. J.; Stannard, K. A. Life table parameters of two armoured scale insect (Hemiptera: Diaspididae) species on resistant and susceptible kiwifruit (Actinidia spp.) germplasm. N. Z. J. Crop Hortic. 2009, 37, 335–343. [Google Scholar] [CrossRef]
- Martínez, D. G. Efecto de las interacciones que establecen los herbívoros Toumeyella martinezae y Opuntiaspis philococcus sobre la calidad y reproducción de Myrtillocactus geometrizans Master Degree, Instituto Politécnico Nacional. Ciudad de México, México, 20217. Available online: http://tesis.ipn.mx/handle/123456789/28371.
- Bale, J. S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; Good, J.E.G.; Harrington, R.; Hartley, S.; Jones, T.H.; Lindroth, R.L.; Press, M.C.; Symrnioudis, I.; Watt, A.D.; Whittaker, J.B. Herbivory in global climate change research: direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Schmitt, R. J. , & Holbrook, S. J. Seasonally fluctuating resources and temporal variability of interspecific competition. Oecologia 1986, 69, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Backe, K. M.; Frank, S. D. Chronology of gloomy scale (Hemiptera: Diaspididae) infestations on urban trees. Environ. Entomol. 2019, 48, 1113–1120. [Google Scholar] [CrossRef]
- Nishikawa, K. C. Competition and the evolution of aggressive behavior in two species of terrestrial salamanders. Evolution 1985, 39, 1282–1294. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B. D.; Grime, J. P. An experimental test of plant strategy theory. Ecology 1992, 73, 15–29. [Google Scholar] [CrossRef]
- Aerts, R. Interspecific competition in natural plant communities: mechanisms, trade-offs and plant-soil feedbacks. J. Exp. Biol. 1999, 50, 29–37. [Google Scholar] [CrossRef]
- Bronstein, J. L. The costs of mutualism. Am. Zool. 2001, 41, 825–839. [Google Scholar] [CrossRef]
- Lieutier, F.; Paine, T. D. Responses of mediterranean forest phytophagous insects to climate change. In Insects and diseases of mediterranean forest systems; Paine T.D.; Lieutier F. Eds.; Springer: New York, NY, USA, 2016; pp. 801–858. [Google Scholar] [CrossRef]
- Stadler, B.; Dixon, A. F. G. Costs of ant attendance for aphids. J. Anim. Ecol. 1998, 67, 454–459. [Google Scholar] [CrossRef]
- Jones, E. I.; Bronstein, J. L.; Ferrière, R. The fundamental role of competition in the ecology and evolution of mutualisms. Ann. N. Y. Acad. Sci. 2012, 1256, 66–88. [Google Scholar] [CrossRef] [PubMed]
- Ntiri, E. S.; Calatayud, P.-A.; Van den Berg, J.; Le Ru, B. P. Density dependence and temporal plasticity of competitive interactions during utilisation of resources by a community of lepidopteran stemborer species. Entomol. Exp. Appl. 2017, 162, 272–283. [Google Scholar] [CrossRef]
- Moll, J. D.; Brown, J. S. Competition and coexistence with multiple life-history stages. Am. Nat. 2008, 171, 839–843. [Google Scholar] [CrossRef]
- Leisnham, P. T.; Juliano, S. A. Spatial and temporal patterns of coexistence between competing Aedes mosquitoes in urban Florida. Oecologia 2009, 160, 343–352. [Google Scholar] [CrossRef]
- Chamberlain, S. A.; Bronstein, J. L.; Rudgers, J. A. How context dependent are species interactions? Ecol. Lett. 2014, 17, 881–890. [Google Scholar] [CrossRef]
- Soliveres, S.; Lehmann, A.; Boch, S.; Altermatt, F.; Carrara, F.; Crowther, T. W.; Delgado-Baquerizo, M.; Kempel, A.; Maynard, D. S.; Rillig, M. C.; Singh, B. K.; Trivedi, P.; Allan, E. Intransitive competition is common across five major taxonomic groups and is driven by productivity, competitive rank and functional traits. J. Ecol. 2018, 106, 852–864. [Google Scholar] [CrossRef]
- Siepielski, A. M.; McPeek, M. A. On the evidence for species coexistence: a critique of the coexistence program. Ecology 2010, 91, 3153–3164. [Google Scholar] [CrossRef]
- Sutherland, W. J.; Freckleton, R. P.; Godfray, H. C. J.; Beissinger, S. R.; Benton, T.; Cameron, D. D. . Wiegand, T. Identification of 100 fundamental ecological questions. J. Ecol. 2013, 101, 58–67. [Google Scholar] [CrossRef]
- Ghilarov, A. M. The paradox of the plankton reconsidered; or, why do species coexist? Oikos 1984, 43, 46–52. [Google Scholar] [CrossRef]
- Abrams, P. A. Ecological vs evolutionary consequences of competition. Oikos 1990, 57, 147–151. [Google Scholar] [CrossRef]
Figure 1.
Field conditions. First: armored scale without competition. Second: soft scale plus its mutualistic ant, without competition. Third: soft scale without the mutualistic ant and without competition. Fourth: both scale species in competition, along with the mutualistic ant. Fifth: both scale species in competition without the mutualistic ant.
Figure 1.
Field conditions. First: armored scale without competition. Second: soft scale plus its mutualistic ant, without competition. Third: soft scale without the mutualistic ant and without competition. Fourth: both scale species in competition, along with the mutualistic ant. Fifth: both scale species in competition without the mutualistic ant.
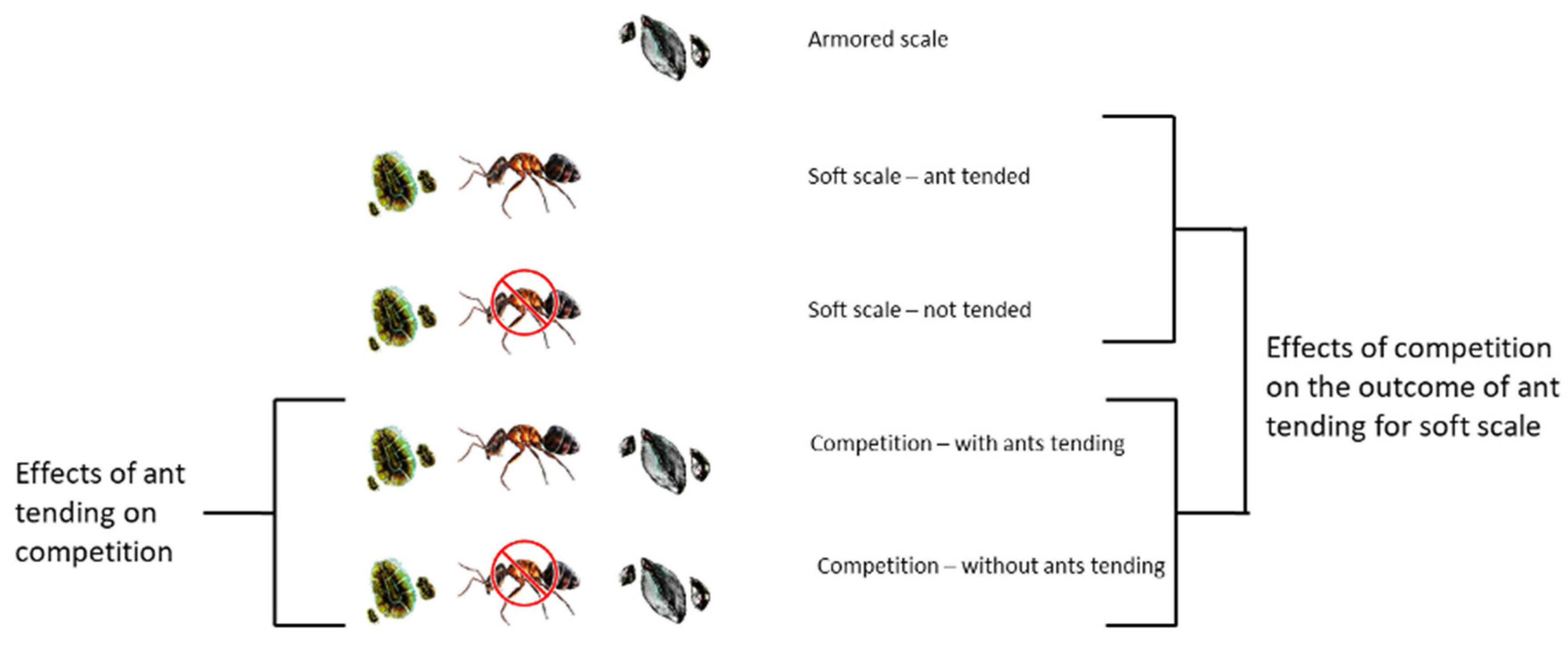
Figure 2.
Area occupied by each species without (A) or with (B) competition at the beginning (October) and end (April) of the study period, and in the presence or absence of the mutualistic ant.
Figure 2.
Area occupied by each species without (A) or with (B) competition at the beginning (October) and end (April) of the study period, and in the presence or absence of the mutualistic ant.
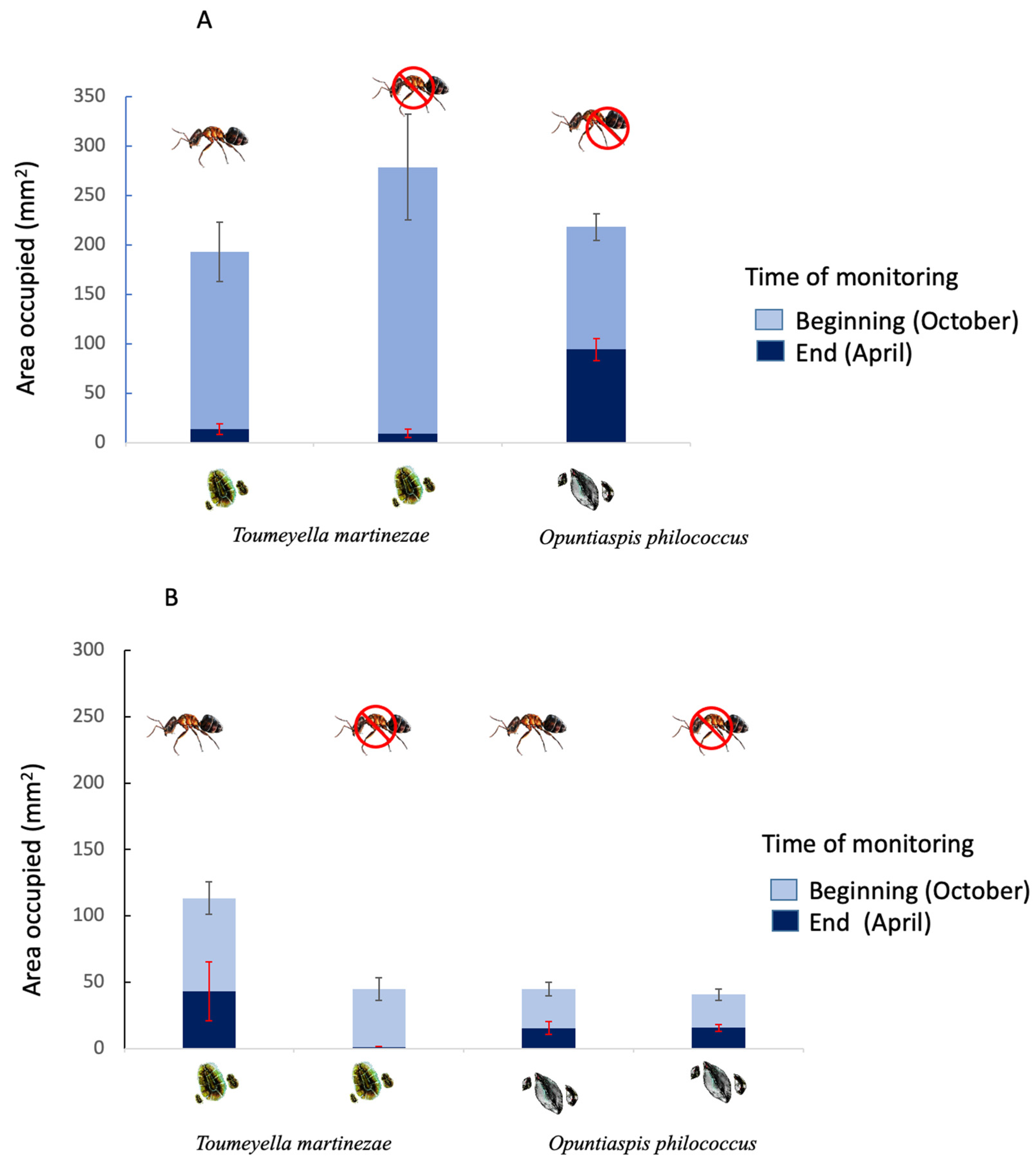
Figure 3.
Space occupation index (SOI) for T. martinezae (solid line) and O. philococcus (dashed line) coexisting either in the presence (A) or absence (B) of L. apiculatum. The vertical lines are the standard errors.
Figure 3.
Space occupation index (SOI) for T. martinezae (solid line) and O. philococcus (dashed line) coexisting either in the presence (A) or absence (B) of L. apiculatum. The vertical lines are the standard errors.
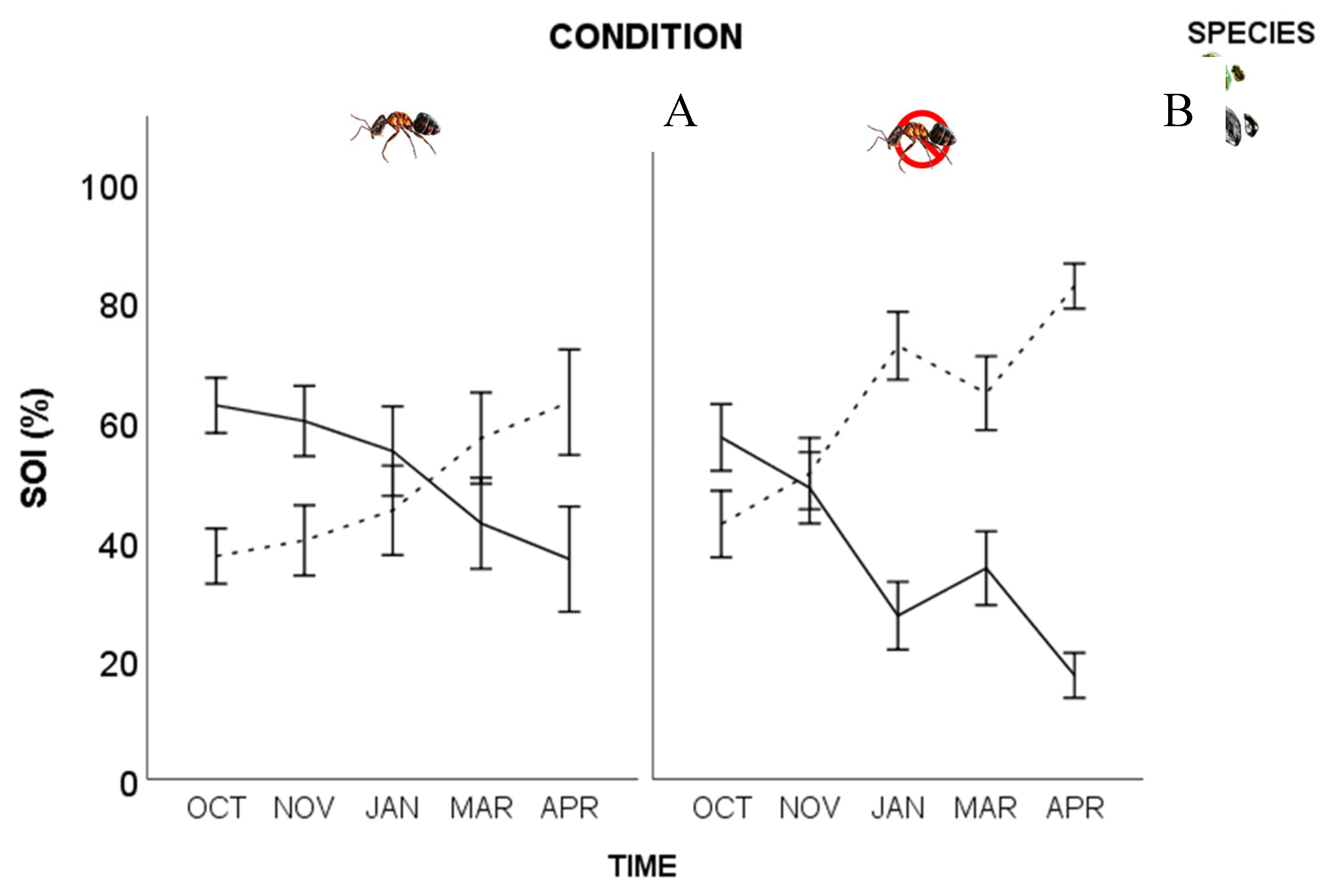
Figure 4.
Population structure of (A) O. philococcus and (B) T. martinezae under different conditions over time, shown as the base-10 logarithm (log) of the number of individuals. The absence of the ant is indicated by the crossed-out red circle. The colored bars represent different stages of the lifecycle. Competition is indicated by images of both species.
Figure 4.
Population structure of (A) O. philococcus and (B) T. martinezae under different conditions over time, shown as the base-10 logarithm (log) of the number of individuals. The absence of the ant is indicated by the crossed-out red circle. The colored bars represent different stages of the lifecycle. Competition is indicated by images of both species.
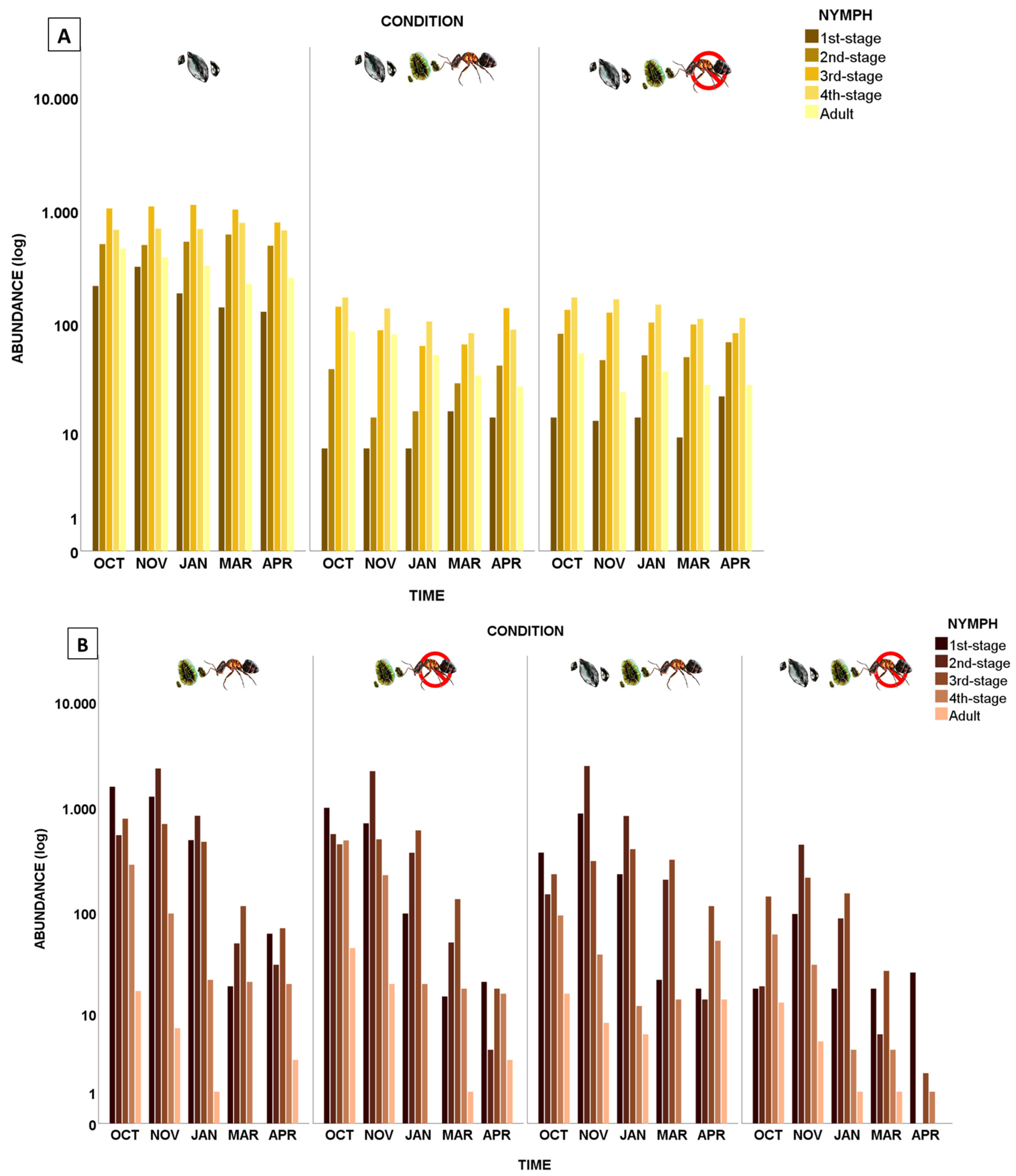

Table 1.
Generalized linear mixed model. The response variables are the coverage of each competing species; the predictive variables are the coverage of the other species, the presence/absence of the ant Liometopum apiculatum, and the month of the year in which the measurement was made. The values of the estimated coefficients are expressed relative to October. Individual data are nested (five data points were taken from the same plant, with a zero-inflation model). Standard errors (SE), Z, and P-values are given.
Table 1.
Generalized linear mixed model. The response variables are the coverage of each competing species; the predictive variables are the coverage of the other species, the presence/absence of the ant Liometopum apiculatum, and the month of the year in which the measurement was made. The values of the estimated coefficients are expressed relative to October. Individual data are nested (five data points were taken from the same plant, with a zero-inflation model). Standard errors (SE), Z, and P-values are given.
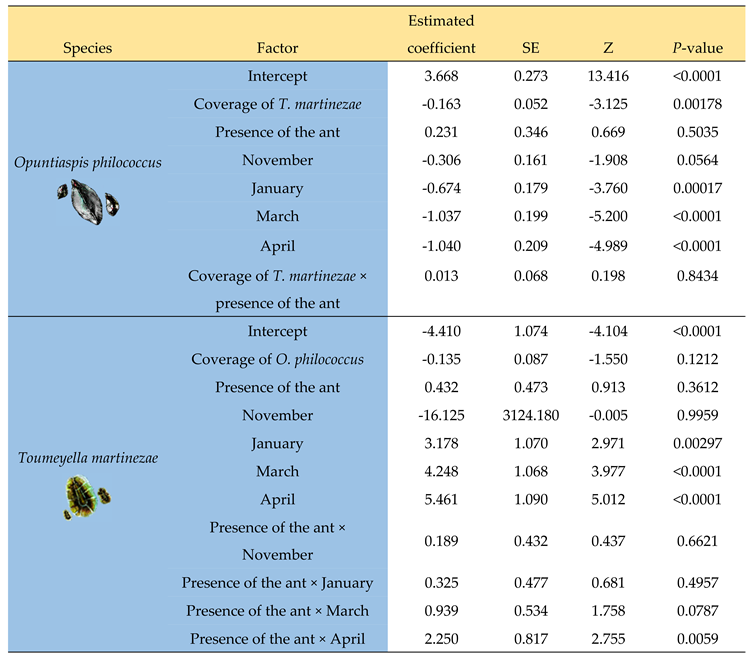 |
Table 2.
Results of the generalized linear models (GLM) used to examine the effect of competition on the population structure. The models were fitted separately to each scale species. The number of live individuals recorded on a given census date (as a response variable) was modeled as a function of the age (size) category, the census date (five dates), and condition (with or without the ant; in competition or without competition), plus the second- and third-order interactions of these variables. The likelihood ratio, degrees of freedom (DF), and P-values are given.
Table 2.
Results of the generalized linear models (GLM) used to examine the effect of competition on the population structure. The models were fitted separately to each scale species. The number of live individuals recorded on a given census date (as a response variable) was modeled as a function of the age (size) category, the census date (five dates), and condition (with or without the ant; in competition or without competition), plus the second- and third-order interactions of these variables. The likelihood ratio, degrees of freedom (DF), and P-values are given.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



