
Submitted:
26 July 2023
Posted:
27 July 2023
You are already at the latest version
Abstract
Acid stress poses a common challenge for bacteria in diverse environments by the presence of inorganic (e.g., mammals’ stomach) or organic acids (e.g., feed additives; acid-based disinfectants). Limited knowledge exists regarding acid-tolerant strains of specific serotypes, clonal lineages, or sources in human/animal pathogens, namely non-typhoidal Salmonella enterica (NTS) and Enterococcus faecium (Efm). This study evaluated acidic pH and peracetic acid (PAA) susceptibility of Efm (n=72) and NTS (n=60) from diverse epidemiological/genetic backgrounds, and with multiple antibiotic resistance profiles. Efm minimum growth/survival-pH was 4,5-5/3-4 and for NTS 4,0-4,5/3,5-4,0. Among Efm, only those of clade-non-A1 (non-hospital associated) or food chain demonstrated greater tolerance to acidic pH compared to clade-A1 (hospital-associated clones) or clinical isolates (P<0.05). MDR (multidrug-resistant) NTS survived better to acidic pH (P<0.05). The MICPAA/MBCPAA was 70-120/80-150mg/L for Efm and 50-70/60-100mg/L for NTS. Efm-clade-A1 or MDR strains exhibited higher PAA tolerance than clade-non-A1 or non-MDR ones (P<0.05). Higher tolerance was found in non-MDR and clinical NTS than in food chain isolates (P<0.05), but not between different serogroups. This unique study identifies specific NTS or Efm populations more tolerant to acidic pH or PAA, emphasizing the need for further research to tailor control measures of public health and food safety within a One Health framework.
Keywords:
Biocides
; One Health
; Food chain
; Antimicrobial resistance
1. Introduction
Acid stress poses a widespread challenge for bacteria in various natural and transient environments, where exposure to organic or inorganic acids is commonplace. It occurs through natural geochemical or microbial metabolic processes, in mammal and bird stomachs (pH 1.5-3.7), in specific cells during infection (e.g., macrophages: pH 5.0-5.5), in acid-rich foods (e.g., citrus fruits), in acidified feed (pH 3.5-4.6), or through the exposure to acid-based disinfectants (pH<5) [1,2,3]. Inorganic acids (e.g., HCl) occur in animal stomachs, while organic acids, such as formic, propionic or citric, are often used in animal feed as preservatives or to promote animal health and growth by modulating gut microbiota [4,5,6]. On the other hand, the peracetic acid (PAA) is an organic acid-based broad-spectrum biocide used for multiple purposes (applications or concentration allowed differ according to geographic regions) as hand disinfection (150-2000 mg/L) [7], disinfection of fresh produce (<80 mg/L, USA) [8], poultry carcass (220 mg/L, USA) [9], or animal drinking water (25mg/L), food processing equipment and food contact/surfaces (20-3000mg/L), animal feet/animal houses (100-5000 mg/L), clinical settings surfaces (125-1500 mg/L) or reduction of fecal bacteria counts in waste water/sewage before leaving treatment plants (1.5 mg/L in the effluent) [7]. PAA stands out among other biocides due to its notable advantages, including its rapid action and short contact time, effectiveness even in the presence of high organic loads, rapid biodegradability and minimal environmental impact (PAA breaks down into harmless byproducts—acetic acid, oxygen, and water) [3,7,10]. The effectiveness of organic acids relies on their ability to penetrate cell membranes as protonated acids, allowing the undissociated forms to freely diffuse through the cell membrane into the cytoplasm at low pH [1,11]. Inside the cell, acid dissociation occurs as a result of the elevated pH, leading to the release of charged anions and protons that accumulate in the cytoplasm, disrupting crucial enzymatic activity and exerting detrimental effects on protein and DNA/RNA synthesis, as well as the proton motive force [12]. Consequently, the cell’s ability to restore its cytoplasmic alkalinity is compromised, profoundly impacting vital processes such as cell growth, metabolism, nutrient absorption, substrate degradation, and the synthesis of proteins and nucleic acids [12,13,14,15]. Besides the acid action of PAA, this biocide also damages cellular components and genetic material through the generation of reactive oxygen species [16,17,18]. On another hand, inorganic acids (e.g., HCl in the stomach) primarily act by reducing the cytoplasmatic pH of bacteria [12].
Bacteria have developed multiple strategies to respond to acid stress, including the production of neutralizing products (e.g., NH3), ATP consumption for proton elimination (e.g., decarboxylation of amino acids), efflux of anions through membrane pumps (e.g., F1-F0-ATPase proton pump), or membrane modifications (e.g., fluidity, lipid composition) [1,12,19], while the tolerance mechanisms to PAA are not fully understood [3]. While acidic environments are prevalent and bacteria have well-documented adaptive strategies to cope with them [1,20], the precise impact of these factors on the selection of acid-tolerant and/or antibiotic-resistant strains, particularly those relevant to human health, remains poorly understood.
Limited knowledge exists regarding the occurrence of acid-tolerant strains in the food chain where acid stress is common, namely those associated with human and/or animal infections or used as hygiene indicator of drinking water, food and food contact surfaces, as the case of non-typhoidal Salmonella enterica (NTS) and/or Enterococcus faecium (Efm). NTS or Enterococcus spp. have adaptive responses to acid tolerance [21,22,23], enabling their survival in diverse acidic environments across the food chain and beyond [24,25] but studies concerning PAA susceptibility are scarce and for NTS have been showing variable levels of tolerance [26,27,28]. Moreover, most of the studies on NTS and Efm often overlook the inclusion of isolates from diverse epidemiological and genetic backgrounds impairing understanding if particular strains (e.g., serogroups/serotypes, clonal lineages, source-related or antibiotic resistance ones), are better selected in specific acidic environments [26,29,30,31,32,33]. The aim of this study was to assess the susceptibility to acidic pH and PAA of a comprehensive collection of antibiotic-resistant NTS and Efm strains from diverse epidemiological and genetic backgrounds.
2. Materials and Methods
2.1. Epidemiological Background of Bacterial Isolates
A collection of 132 isolates including Efm (n=72) and NTS (n=60) obtained in previous surveillance studies [24,25,34,35] and representative of different geographical regions, sources, time spans and genomic backgrounds was analysed. Efm were isolated between 1996 and 2018 in Portugal, Spain, Tunisia and Angola from human (n=29; 23 clinical isolates, 6 from fecal colonization of healthy human or long-term facility care patients), food chain (n=42; 28 from poultry skin, 8 from piggeries, 3 from supermarket trout, 2 from bovine meat; 1 from ready to eat salad) and environmental (n=1) sources. In previous studies these Efm were identified as clade A1 (n=21; mostly associated in literature with human infections and hospital outbreaks) or non-clade A1 (n=37, non-hospital associated, mostly associated with animal colonization) [36], while some were not typed (n=14). Sixty-one (85%) were multidrug-resistant (MDR; resistant to three or more antibiotics from different families) [37], with 25 showing resistance to vancomycin and 44 to ampicillin. Only two isolates were susceptible to all antibiotics tested in previous studies. Efm data concerning acid pH and PAA susceptibility from poultry were previously published [25] but included here for source comparison.
NTS were recovered between 2002 and 2018 in Portugal from human (n=20 clinical isolates), food chain (n=37; 12 from pork meat/pork products; 15 poultry meat/skin and byproducts; 5 from pig and piggeries; 4 from trout aquaculture; 1 chicken manure, animal feed, surface/drain and foodstuff each) sources. They belong to 17 NTS serotypes (23 S. Typhimurium and its variant 1,4,[5],12:i:-, 4 S. Heidelberg, 4 S. Rissen, 4 S. Infantis, 3 S. Derby, 3 S. Enteritidis, 3 S. Mbandaka, 3 S. Virchow, 3 S. Stanley, 3 S. Hadar, 2 S. Kentucky, and 1 S. Bovismorbificans, S. Abony, S. Guerin, S. Linguere, S. Newport each). Most were from serogroups B (n=33) or C (n=23). Forty-four were MDR, with 9 being resistant to ciprofloxacin or pefloxacin, 6 to colistin, 5 to cefotaxime or to cefotaxime. Eleven isolates did not show any resistance to the antibiotics tested in previous studies. NST data concerning acidic pH and PAA susceptibility from 5 poultry isolates were previously published [24] but included here for source comparison.
2.2. Susceptability to Acidic pH
Susceptibility to acidic pH was performed using an adaptation of the microdilution standard method (ISO 20776-1:2019) [24,25,38]. To determine the minimum growth pH and survival pH of the bacteria, Mueller-Hinton II broth (BD BBL™, Franklin Lakes, NJ, USA) culture media was used and adjusted to a pH range of 2.0 to 6.5 (in 0.5 intervals) using hydrochloric acid-HCl (Merck, Darmstadt, Germany). A freshly prepared 96-well microtiter plate was used for each assay. Bacterial suspensions in log-phase growth were adjusted and inoculated in each well with the corresponding pH to reach a final inoculum of 5 × 105 CFU/ml. To confirm the inoculum for each isolate tested, colony counts were performed in the surface of Mueller-Hinton 2 agar plates (bioMérieux, Marcy-l’Étoile, France). The microdilution and the Mueller-Hinton 2 agar plates were incubated at 37°C for 20±2 hours. The minimum growth pH was determined by identifying the lowest pH at which visible bacterial growth was observed. To determine the minimum survival pH, 10 μL of the wells without visible growth were inoculated on BHI agar (Liofilchem, Roseto degli Abruzzi, Italy) and incubated at 37°C for 24-48 hours. The minimum survival pH was determined as the lowest pH showing at least one colony growth in BHI agar. Control strains Enterococcus faecalis ATCC 29212 (minimum growth pH = 4.5; minimum survival pH = 4.0), Escherichia coli ATCC 25922 (minimum growth pH = 4.5; minimum survival pH = 3.5) and Salmonella Typhimurium LT2 (minimum growth pH = 4.0; minimum survival pH = 4.0) were included in all assays. All acidic pH susceptibility assays were performed 2 to 5 times for each bacteria. The mean of the replicas was calculated as the final result for each isolate.
2.3. Susceptability to PAA
Susceptibility to PAA was performed using an adaptation of the microdilution standard method (ISO 20776-1:2019) [38]. The Minimum Inhibitory Concentration (MICPAA) was determined by identifying the first concentration of PAA without visible growth in Mueller-Hinton II broth supplemented with PAA (15% stock solution, CAS No. 79-21-0; PanReac AppliChem, Darmstadt, Germany) at concentrations ranging from 50-90 mg/L for NTS and 60-160 mg/L to Efm, both with a 10 mg/L interval, and distributed in a freshly prepared 96-well microtiter plate for each assay. Bacterial suspensions in log-phase growth were adjusted and inoculated in each well with the corresponding PAA concentration to reach a final inoculum of 5 × 105 CFU/ml, confirmed for each isolate by colony counts in Mueller-Hinton 2 agar, followed by incubation at 37°C for 20±2 hours. The Minimum Bactericidal Concentration (MBCPAA) was established as the lowest PAA concentration for which the number of colonies was equal to or less than the rejection value defined by CLSI:1999 (former NCCLS:1999) guidelines [39], based on the final bacterial inoculum of each well after incubation confirmed by actual count. To determine the MBCPAA, 10 μL of the wells without visible growth were plated on BHI agar (37°C for 24-48 h). The pH of PAA concentrations tested was also determined for each assay, ranging between 5.5 and 7, in which non-dissociated PAA was present at 99.8–98% (PAA pKa = 8.2 at 20°C) [40]. Control strains E. faecalis ATCC 29212 (MICPAA = 100 ppm; MBCPAA = 120 ppm), E. coli ATCC 25922 (MICPAA = 60 ppm; MBCPAA = 60 ppm) and Salmonella Typhimurium LT2 (MICPAA = 50 ppm; MBCPAA = 70 ppm) were included in all assays. All PAA susceptibility assays were performed 2 to 5 times for each bacteria. The mean of the replicas was calculated as the final result of each isolate.
2.3. Statistical Analysis
Differences in distribution of acid pH and PAA susceptibility values among Efm or NTS isolates considering sources, clonal lineages, serogroups or susceptibility to antibiotics were analyzed by the Mann-Whitney test (α = 0.05), and comparison between proportions by the Fisher exact test (α = 0.05), using Prism software, version 8.1.1 (GraphPad).
3. Results and Discussion
3.1. Susceptability to Acidic pH
3.1.1. Enterococcus Faecium
Efm exhibited a minimum growth pH and minimum survival pH ranging between 4.5-5 and 3-4, respectively (Figure 1). In terms of minimum survival pH, the distribution of isolates from clade non-A1 (n=37) suggests greater tolerance compared to those from clade A1 (n=21) (P<0.05) [Figure 2b]. Also, the food chain isolates (n=42) distribution suggests higher tolerance compared to the clinical isolates (n=23) (P<0.05) [Figure 2d]. The comparison of Efm from environmental sources or human colonization was not considered due to the limited number of isolates available. In contrast, statistical analysis revealed no significant differences (P>0.05) in the minimum survival pH of Efm among MDR and non-MDR isolates [Figure 2f]. Regarding the minimum growth pH of isolates, identical results were observed comparing different clades, sources, or MDR/non-MDR profiles (P>0.05) [Figure 2a,c,e]. The 45 isolates with the lowest minimum growth at pH=4.5 were from diverse sources (clinical, n=13; human colonization, n=5; food chain, n=26; environment, n=1) or clades (A1, n=14; non-A1, n=20; non-identified, n=11), with most being MDR (n=36). The 9 isolates with the lowest value of minimum survival at pH=3 were recovered from poultry meat (n=7) raised in farms using organic acids in feed and recovered in a slaughterhouse using PAA as a sanitizer as well as from supermarket trout (n=2), most belonging to clade non-A1 (n=7) and being MDR (n=7). Efm resistant to clinically relevant antibiotics vancomycin or ampicillin showed similar minimum growth pH or minimum survival pH values to other isolates susceptible to these antibiotics.
The pH values at which Efm grew and survived in this study were consistent with previous findings for this species [30,32,41]. Isolates from diverse sources and antibiotic resistance profiles tolerate acidic pH values, supporting Efm widespread in diverse environments and hosts. However, the observation of greater tolerance in Efm isolates recovered from food chain or belonging to clade non-A1 (usually identified in animal colonization), compared to clinical isolates or those from clade A1 (usually identified in human infection and hospital outbreaks), suggests distinct adaptive needs for acid stress in subpopulations that have adapted to different niches. The increased tolerance of isolates from food-animal sources may be attributed to their exposure to acidic environments during food production, processing and preservation [5,42,43]. In fact, the most recent poultry isolates included in the study were recovered from chicken meat which had been sourced from chickens fed with diets supplemented with organic acids. Furthermore, the absence of significant differences in minimum growth pH and minimum survival pH between MDR and non-MDR isolates suggests that acidic environments may not be a significant driver selecting such strains. However, it is worth noting that even at such low pHs (3-3.5), the survival of antibiotic-resistant Efm strains, such as those resistant to clinically relevant vancomycin and ampicillin, is not impaired, as for example during passage through the stomach and gut of both humans and animals.
3.1.2. Non-Typhoidal Salmonella
Minimum growth pH was for most isolates 4.0, with just one S. Typhimurium from a pig carcass showing 4.5. The minimum survival pH for NTS ranged between 3.5-4.0, (Figure 3). MDR isolates distribution suggest they survive better to lower pH values than non-MDR ones [Figure 4f]. No statistical differences were observed among minimum growth pH and minimum survival pH when comparing isolates from different sources, with/without MDR profile (except for minimum growth pH) or serogroups (B and C) (Figure 4). The 32 isolates with the lowest value of minimum survival at pH=3.5 were recovered from different samples of food chain (n=23) and patients (n=9), with most being from serogroup B (n=20) and MDR (n=26). NTS isolates resistant to the clinically relevant antibiotics ciprofloxacin or pefloxacin, 3rd generation cephalosporins or colistin showed minimum growth pH or minimum survival pH values similar to isolates susceptible to these antibiotics.
The pH tolerance values observed for NTS were consistent with those reported in the literature for the serotypes frequently associated with human infections, specifically S. Typhimurium (serogroup B) [44,45,46]. The lack of variation in acid tolerance from serogroups B and C or sources may be explained by the fact that NTS is a zoonotic pathogen, with most isolates of the serogroups studied being similarly adapted to the diverse food chain environment acid challenges. In contrast to Efm, there was an association between acid tolerance and MDR NTS, suggesting that acid stress may play a role in selecting such populations. It is also important to note that NTS ability to tolerate acidic conditions facilitates its survival and passage through the gastrointestinal tracts of humans and animals, including of strains showing resistance to clinically relevant antibiotics such as quinolones, 3rd generation cephalosporins, or colistin, especially of the emergent serotypes belonging to serogroups B or C often causing human infections.
3.2. Susceptability to PAA
3.2.1. Enterococcus Faecium
Efm MICPAA and MBCPAA varied between 70-120 mg/L and 80-150 mg/L, respectively (Figure 5). Efm from clade A1 (n=21) or with a MDR profile were more tolerant to PAA considering MBC distribution values than those of clade non-A1 or non-MDR, respectively (P<0.05) [Figure 6b,f]. Most of clade A1 isolates (52%, n=11/21) had the highest values of MBCPAA =130-150 mg/L comparing to clade non-A1 (24%, n=9/37) (P<0.05). The 20 isolates showing the higher values of MBCPAA (130, 140 or 150 mg/L) were associated with hospitalized of long-care facility patients (n=10), with poultry meat collected in a slaughterhouse using PAA as a disinfectant (n=6), and the remaining 4 from piggeries environment and healthy human faeces. No statistical differences were observed when comparing MBCPAA values of Efm from different sources [Figure 6d]. Regarding MICPAA, similar results were observed comparing clades, sources or antibiotic resistance profiles [Figure 6a,c,e]. The 5 isolates with the highest MICPAA=100-120mg/L were all recovered from poultry meat from a slaughterhouse using PAA as sanitizer and organic acids in feed, belonged to clade non-A1 (n=2) or were non-identified (n=2), and were MDR. Vancomycin or ampicillin resistant Efm showed MICPAA=80-90mg/L or 70-90mg/L, respectively, and MBCPAA=90-150mg/L, each. Our analysis of Efm from all sources revealed no significant association between MICPAA and minimum growth pH or between MBCPAA and minimum survival pH (P>0.05) (Figure 7).
As far as we know there are limited studies showing the susceptibility of Efm populations to PAA and they use diverse methodological strategies [33,47,48], which difficult comparison of our data with isolates from other collections. However, this study shows that Efm can survive above the minimum concentration of PAA used in food chain (20mg/L-sanitation of automatic spraying in closed systems; 125 mg/L-hand disinfection) and in the clinical settings (125 mg/L-hand disinfection in hospitals, health and animal care areas) [7]. Although similar proportions of clinical Efm presented MBC above or below the 125mg/L (n=9/23 isolates and n=14/23, respectively; P>0.05), most clade A1 isolates were above this value suggesting they can at least survive hand disinfection. This concern extends to lower concentrations of PAA used in food industry surfaces or those expected to occur in sewage effluents (1.5 mg/L), as they may not completely eliminate all Efm strains. This limitation in efficacy against such hygiene or fecal indicators raises concerns about the containment of antibiotic resistant isolates in the environment. Thus, strains exhibiting higher MBCs warrant careful surveillance across different settings.
While a recent study indicated that exposure of Efm to low doses of PAA did not lead to changes in the relative abundance of the highly prevalent transferable erm(B) gene (macrolide, lincosamide, and streptogramin B resistance) despite bacterial adaptation to PAA stress [49], further data is needed to evaluate whether PAA is an effective choice as a sanitizer that does not promote the selection of antibiotic resistant Efm. This evaluation becomes particularly important in the clinical setting where this species is a major pathogen.
3.2.2. Non-Typhoidal Salmonella
Salmonella MICPAA and MBCPAA varied among 50-70 mg/L and 60-100 mg/L respectively (Figure 8). Isolates belonging to serogroup B demonstrated a greater ability to grow under PAA compared to those from serogroup C, as indicated by the MICPAA distribution (P<0.05) [Figure 9a] but were not able to survive better, as similar values of MBCs were found (P>0.05) [Figure 9b]. Moreover, both human associated infections and non-MDR isolates exhibited higher tolerance to PAA, as reflected by the MBC [Figure 9d,f]. Of note, 75% of the isolates causing human infection (n=15/20) or that were non-MDR (n=12/16) showed the highest values observed (MBCPAA=90-100 mg/L; P<0.05), contrasting with the those of food chain (40%, n=16/40) or MDR (43%, n=19/44), respectively. No statistical differences were observed when comparing MICPAA of NTS of different sources or with MDR or non-MDR profiles [Figure 9c,e]. The 13 isolates with the highest MICPAA=70 mg/L were mostly from the food chain (n=9) followed from patients (n=4), belong to serogoups B (n=11), C or D (one each), and 11 were MDR. The 5 isolates with the highest value of MBCPAA=100mg/L were recovered from the food chain (n=3) and patients (n=2), belong to serogroups B (n=3) or C (n=2) and 3 were MDR. Most of ciprofloxacin or pefloxacin resistant isolates had a MICPAA=60-70 mg/L and MBCPAA=70-100 mg/L, those resistant to colistin MICPAA=50-70mg/L and MBCPAA=90-100 mg/L, and those resistant to 3rd generation cephalosporins a MICPAA=60-70 mg/L and MBCPAA=70 mg/L. No association between MICPAA and minimum growth pH or between MBCPAA and minimum survival pH was detected (Figure 10) in NTS from all sources analyzed (P>0.05), as isolates with high or low pH growth or survival presented diverse MICPAA or MBCPAA.
Our findings emphasize that under certain conditions, the currently recommended concentrations of PAA used in disinfection products applied in the food and feed industry (20-3000mg/L for Product-Type PT 4) may be ineffective against NTS strains, including those recovered from cases of human infection. This is due to the fact that the MIC/MBC values to PAA for these strains fell within the range of the suggested concentrations to be used in food and feed industry. The MBCPAA was also found to be higher in the non-MDR subpopulation, which comprised 44% of isolates associated with human infections, thus justifying the overlap of data between sources and antibiotic resistance profiles.
Significant variations in MICs and MBCs of PAA have been reported among different Salmonella strains in various studies [50,51,52,53]. However, these variations, ranging from 7-80 mg/L to 500-1760 mg/L for MICs and from 20-80 mg/L to 200-1000 mg/L for MBCs, can be attributed to the diversity of methodological approaches that have been employed. The observed variations can be attributed to factors such as differences in culture medium, incubation temperature, contact time with the compound, as well as the limited number of isolates, serotypes, or clones tested [51,53,54,55], highlighting the need for standardized methods to accurately assess bacteria susceptibility to disinfectants.
4. Conclusions
Our study stands out as a unique and comprehensive analysis of the susceptibility of specific populations of NTS and Efm to acidic conditions and PAA, uncovering notable differences among them. Due to its rapid degradation in hydrogen peroxide (which is also unstable) and acetic acid, lack of surface residues, and unspecific mode of action, PAA has been proposed has having a low likehood of bacterial resistance development [7]. However, while PAA seems to be effective in most disinfection practices, the heightened tolerance observed in NTS associated with human infections and clade A1 or MDR Efm raises concerns and emphasizes the need for ongoing surveillance to monitor the evolution of bacterial tolerance to this environmentally-friend biocide. While acid pH may play a role in the MDR profile of NTS, it does not seem to have the same impact on Efm, but in both cases isolates resistant to clinically relevant antibiotics tolerate low pHs values, facilitating animal and human gut colonization. Further research is crucial to investigate the acidic pH and PAA susceptibility of NTS and Efm populations representative of various selective pressure scenarios worldwide, as well as to explore their specific genetic and physiological factors contributing to tolerance to these stresses. By addressing such research gaps, we can advance our knowledge regarding the survival and persistence of pathogens in food-related and clinical environments, facilitating the implementation of customized control measures and selection of suitable biocides that are tailored to the local microbiota. Ultimately, these efforts will contribute to more effective strategies in mitigating the impact of antibiotic resistant pathogens on public health and food safety within a One Health context.
Author Contributions
Conceptualization, Carla Novais and Patrícia Antunes; methodology, Carla Novais, Patrícia Antunes and Andreia Rebelo; software, Carla Novais and Andreia Rebelo; validation, Carla Novais, Patrícia Antunes; formal analysis, Andreia Rebelo, Carla Novais and Patrícia Antunes; investigation, Andreia Rebelo, Bárbara Duarte, Carla Novais and Patrícia Antunes; resources, Carla Novais, Patrícia Antunes and Luísa Peixe; data curation, Andreia Rebelo, Carla Novais and Patrícia Antunes; writing—original draft preparation, Andreia Rebelo, Carla Novais and Patrícia Antunes; writing—review and editing, Andreia Rebelo, Bárbara Duarte, Ana R. Freitas, Luísa Peixe Carla Novais and Patrícia Antunes; visualization, Andreia Rebelo, Carla Novais, Patrícia Antunes; supervision, Carla Novais and Patrícia Antunes; project administration, Carla Novais and Patrícia Antunes; funding acquisition, Carla Novais, Patrícia Antunes and Luísa Peixe. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financed by national funds from FCT - Fundação para a Ciência e a Tecnologia, I.P., in the scope of the project UIDP/04378/2020 and UIDB/04378/2020 of the Research Unit on Applied Molecular Biosciences - UCIBIO, the project LA/P/0140/2020 of the Associate Laboratory Institute for Health and Bioeconomy - i4HB, by the AgriFood XXI I&D&I project (NORTE-01-0145-FEDER-000041) cofinanced by European Regional Development Fund (ERDF) and through the NORTE 2020 (Programa Operacional Regional do Norte 2014/2020), and by the exploratory project EXPL/SAU-INF/0261/2021. Andreia Rebelo was supported by a PhD fellowship from FCT (SFRH/BD/137100/2018), co-financed by European Social Fund through Norte Portugal Regional Operational Programme (NORTE 2020). ARF acknowledges the Junior Research Position (CEECIND/02268/2017) granted by FCT - Fundação para a Ciência e a Tecnologia. The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Data Availability Statement
Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Lund, P.A.; De Biase, D.; Liran, O.; Scheler, O.; Mira, N.P.; Cetecioglu, Z.; Fernández, E.N.; Bover-Cid, S.; Hall, R.; Sauer, M.; et al. Understanding How Microorganisms Respond to Acid pH Is Central to Their Control and Successful Exploitation. Front. Microbiol. 2020, 11, 556140. [Google Scholar] [CrossRef]
- Ricke, S. Perspectives on the Use of Organic Acids and Short Chain Fatty Acids as Antimicrobials. Poult. Sci. 2003, 82, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Wessels, S.; Ingmer, H. Modes of Action of Three Disinfectant Active Substances: A Review. Regul. Toxicol. Pharmacol. 2013, 67, 456–467. [Google Scholar] [CrossRef] [PubMed]
- European Union Regulation (EC) No 1831/2003 of the European Parliament and of the council of 22 September 2003 on additives for use in animal nutrition, Off J Eur Union. 2003;L 268 Oct 18.
- Saleem, K.; Saima; Rahman, A. ; Pasha, T.N.; Mahmud, A.; Hayat, Z. Effects of Dietary Organic Acids on Performance, Cecal Microbiota, and Gut Morphology in Broilers. Trop. Anim. Health Prod. 2020, 52, 3589–3596. [Google Scholar] [CrossRef]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to Antibiotics for Maximizing Growth Performance and Feed Efficiency in Poultry: A Review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef] [PubMed]
- European Chemicals Agency (ECHA); Biocidal Products Committee (BPC) Regulation (EU) No 528/2012 Concerning the Making Available on the Market and Use of Biocidal Products – Evaluation of Active Substances, Assessment Report of Peracetic Acid (Product Type 1-6). 2015.
- U.S. Food and Drug Administration Code of Federal Regulation. Title 21—Food and Drugs—Chemicals Used in Washing or to Assist in the Peeling of Fruits and Vegetables 2012.
- U.S. Department of Agriculture, Food Safety and Inspection Service Safe and Suitable Ingredients Used in the Production of Meat, Poultry, and Egg Products 2021.
- Petri, E.; Virto, R.; Mottura, M.; Parra, J. Comparison of Peracetic Acid and Chlorine Effectiveness during Fresh-Cut Vegetable Processing at Industrial Scale. J. Food Prot. 2021, 84, 1592–1602. [Google Scholar] [CrossRef] [PubMed]
- Huyghebaert, G.; Ducatelle, R.; Immerseel, F.V. An Update on Alternatives to Antimicrobial Growth Promoters for Broilers. Vet. J. 2011, 187, 182–188. [Google Scholar] [CrossRef]
- Guan, N.; Liu, L. Microbial Response to Acid Stress: Mechanisms and Applications. Appl. Microbiol. Biotechnol. 2020, 104, 51–65. [Google Scholar] [CrossRef]
- Cherrington, C.A.; Hinton, M.; Chopra, I. Effect of Short-Chain Organic Acids on Macromolecular Synthesis in Escherichia coli. J. Appl. Bacteriol. 1990, 68, 69–74. [Google Scholar] [CrossRef]
- Trček, J.; Mira, N.P.; Jarboe, L.R. Adaptation and Tolerance of Bacteria against Acetic Acid. Appl. Microbiol. Biotechnol. 2015, 99, 6215–6229. [Google Scholar] [CrossRef]
- Mani-López, E.; García, H.S.; López-Malo, A. Organic Acids as Antimicrobials to Control Salmonella in Meat and Poultry Products. Food Res. Int. 2012, 45, 713–721. [Google Scholar] [CrossRef]
- Shi, C.; Li, C.; Wang, Y.; Guo, J.; Barry, S.; Zhang, Y.; Marmier, N. Review of Advanced Oxidation Processes Based on Peracetic Acid for Organic Pollutants. Water 2022, 14, 2309. [Google Scholar] [CrossRef]
- Kitis, M. Disinfection of Wastewater with Peracetic Acid: A Review. Environ. Int. 2004, 30, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, W.P.; Carlos, T.D.; Cavallini, G.S.; Pereira, D.H. Peracetic Acid: Structural Elucidation for Applications in Wastewater Treatment. Water Res. 2020, 168, 115143. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, H.; Lin, Z.; Xu, P. Mechanisms of Acid Tolerance in Bacteria and Prospects in Biotechnology and Bioremediation. Biotechnol. Adv. 2015, 33, 1484–1492. [Google Scholar] [CrossRef]
- Lund, P.; Tramonti, A.; De Biase, D. Coping with Low pH: Molecular Strategies in Neutralophilic Bacteria. FEMS Microbiol. Rev. 2014, 38, 1091–1125. [Google Scholar] [CrossRef]
- Gaca, A.O.; Lemos, J.A. Adaptation to Adversity: The Intermingling of Stress Tolerance and Pathogenesis in Enterococci. Microbiol. Mol. Biol. Rev. 2019, 83, e00008–19. [Google Scholar] [CrossRef]
- Álvarez-Ordóñez, A.; Prieto, M.; Bernardo, A.; Hill, C.; López, M. The Acid Tolerance Response of Salmonella spp.: An Adaptive Strategy to Survive in Stressful Environments Prevailing in Foods and the Host. Food Res. Int. 2012, 45, 482–492. [Google Scholar] [CrossRef]
- Spector, M.P.; Kenyon, W.J. Resistance and Survival Strategies of Salmonella Enterica to Environmental Stresses. Food Res. Int. 2012, 45, 455–481. [Google Scholar] [CrossRef]
- Mourão, J.; Rebelo, A.; Ribeiro, S.; Peixe, L.; Novais, C.; Antunes, P. Atypical Non-H2S-Producing Monophasic Salmonella Typhimurium ST3478 Strains from Chicken Meat at Processing Stage Are Adapted to Diverse Stresses. Pathogens 2020, 9, 701. [Google Scholar] [CrossRef]
- Rebelo, A.; Duarte, B.; Ferreira, C.; Mourão, J.; Ribeiro, S.; Freitas, A.R.; Coque, T.M.; Willems, R.; Corander, J.; Peixe, L.; et al. Enterococcus spp. from Chicken Meat Collected 20 Years Apart Overcome Multiple Stresses Occurring in the Poultry Production Chain: Antibiotics, Copper and Acids. Int. J. Food Microbiol. 2023, 384, 109981. [Google Scholar] [CrossRef]
- Alonso-Hernando, A.; Capita, R.; Prieto, M.; Alonso-Calleja, C. Adaptation and Cross-Adaptation of Listeria monocytogenes and Salmonella enterica to Poultry Decontaminants. J. Microbiol. 2009, 47, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Hernando, A.; Capita, R.; Prieto, M.; Alonso-Calleja, C. Comparison of Antibiotic Resistance Patterns in Listeria monocytogenes and Salmonella enterica Strains Pre-Exposed and Exposed to Poultry Decontaminants. Food Control 2009, 20, 1108–1111. [Google Scholar] [CrossRef]
- Gantzhorn, M.R.; Pedersen, K.; Olsen, J.E.; Thomsen, L.E. Biocide and Antibiotic Susceptibility of Salmonella Isolates Obtained before and after Cleaning at Six Danish Pig Slaughterhouses. Int. J. Food Microbiol. 2014, 181, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Morandi, S.; Brasca, M.; Alfieri, P.; Lodi, R.; Tamburini, A. Influence of pH and Temperature on the Growth of Enterococcus faecium and Enterococcus faecalis. Le Lait 2005, 85, 181–192. [Google Scholar] [CrossRef]
- Shi, Y.; Zhai, M.; Li, J.; Li, B. Evaluation of Safety and Probiotic Properties of a Strain of Enterococcus faecium Isolated from Chicken Bile. J. Food Sci. Technol. 2020, 57, 578–587. [Google Scholar] [CrossRef]
- Fernández, A.; Cebrián, G.; Álvarez-Ordóñez, A.; Prieto, M.; Bernardo, A.; López, M. Influence of Acid and Low-Temperature Adaptation on Pulsed Electric Fields Resistance of Enterococcus faecium in Media of Different pH. Innov. Food Sci. Emerg. Technol. 2018, 45, 382–389. [Google Scholar] [CrossRef]
- Vassos, D.; Bezirtzoglou, E.; Voidarou, C.; Alexopoulos, A.; Maipa, V. Biochemical and Antimicrobial Profile of Enterococcus faecium and E. faecalis Isolated from Traditional Dairy Products and Infant Intestinal Microbiota. Microb. Ecol. Health Dis. 2009, 21, 241–250. [Google Scholar] [CrossRef]
- Suchomel, M.; Lenhardt, A.; Kampf, G.; Grisold, A. Enterococcus Hirae, Enterococcus faecium and Enterococcus faecalis Show Different Sensitivities to Typical Biocidal Agents Used for Disinfection. J. Hosp. Infect. 2019, 103, 435–440. [Google Scholar] [CrossRef]
- Rebelo, A.; Mourão, J.; Freitas, A.R.; Duarte, B.; Silveira, E.; Sanchez-Valenzuela, A.; Almeida, A.; Baquero, F.; Coque, T.M.; Peixe, L.; et al. Diversity of Metal and Antibiotic Resistance Genes in Enterococcus Spp. from the Last Century Reflects Multiple Pollution and Genetic Exchange among Phyla from Overlapping Ecosystems. Sci. Total Environ. 2021, 787, 147548. [Google Scholar] [CrossRef]
- Mourão, J.; Rebelo, A.; Ribeiro, S.; Peixe, L.; Novais, C.; Antunes, P. Tolerance to Arsenic Contaminant among Multidrug-resistant and Copper-tolerant Salmonella Successful Clones Is Associated with Diverse ars Operons and Genetic Contexts. Environ. Microbiol. 2020, 22, 2829–2842. [Google Scholar] [CrossRef]
- Arredondo-Alonso, S.; Top, J.; McNally, A.; Puranen, S.; Pesonen, M.; Pensar, J.; Marttinen, P.; Braat, J.C.; Rogers, M.R.C.; van Schaik, W.; et al. Plasmids Shaped the Recent Emergence of the Major Nosocomial Pathogen Enterococcus faecium. mBio 2020, 11, e03284–19. [Google Scholar] [CrossRef]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- International Organization for Standardization (ISO). ISO 20776–1:2019: Susceptibility Testing of Infectious Agents and Evaluation of Performance of Antimicrobial Susceptibility Test Devices—Part 1: Reference Method for Testing the in Vitro Activity of Antimicrobial Agents against Rapidly Growing Aerobic Bacteria Involved in Infectious Diseases; International Organization for Standardization (ISO): Geneva, Switzerland, 2019. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Determining Bactericidal Activity of Antimicrobial Agents; Approved Guideline; CLSI Document M26-A; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 1999. [Google Scholar]
- Dean, J.A. , Lange, N.A. Lange’s Handbook of Chemistry; 13. ed.; McGraw-Hill: New York, 1985; ISBN 978-0-07-016192-4. [Google Scholar]
- Mohammed, S.; Çon, A.H. Isolation and Characterization of Potential Probiotic Lactic Acid Bacteria from Traditional Cheese. LWT 2021, 152, 112319. [Google Scholar] [CrossRef]
- Park, S.-H.; Choi, M.-R.; Park, J.-W.; Park, K.-H.; Chung, M.-S.; Ryu, S.; Kang, D.-H. Use of Organic Acids to Inactivate Escherichia coli O157:H7, Salmonella Typhimurium, and Listeria monocytogenes on Organic Fresh Apples and Lettuce. J. Food Sci. 2011, 76, M293–M298. [Google Scholar] [CrossRef]
- Thomas, C.; Schönknecht, A.; Püning, C.; Alter, T.; Martin, A.; Bandick, N. Effect of Peracetic Acid Solutions and Lactic Acid on Microorganisms in On-Line Reprocessing Systems for Chicken Slaughter Plants. J. Food Prot. 2020, 83, 615–620. [Google Scholar] [CrossRef]
- Lin, J.; Lee, I.S.; Frey, J.; Slonczewski, J.L.; Foster, J.W. Comparative Analysis of Extreme Acid Survival in Salmonella Typhimurium, Shigella flexneri, and Escherichia coli. J. Bacteriol. 1995, 177, 4097–4104. [Google Scholar] [CrossRef]
- Koutsoumanis, K.P.; Kendall, P.A.; Sofos, J.N. Modeling the Boundaries of Growth of Salmonella Typhimurium in Broth as a Function of Temperature, Water Activity, and pH. J. Food Prot. 2004, 67, 53–59. [Google Scholar] [CrossRef]
- Álvarez-Ordóñez, A.; Fernández, A.; Bernardo, A.; López, M. Acid Tolerance in Salmonella Typhimurium Induced by Culturing in the Presence of Organic Acids at Different Growth Temperatures. Food Microbiol. 2010, 27, 44–49. [Google Scholar] [CrossRef]
- Li, K.; Chiu, Y.-C.; Jiang, W.; Jones, L.; Etienne, X.; Shen, C. Comparing the Efficacy of Two Triple-Wash Procedures With Sodium Hypochlorite, a Lactic–Citric Acid Blend, and a Mix of Peroxyacetic Acid and Hydrogen Peroxide to Inactivate Salmonella, Listeria monocytogenes, and Surrogate Enterococcus faecium on Cucumbers and Tomatoes. Front. Sustain. Food Syst. 2020, 4, 19. [Google Scholar] [CrossRef]
- Zhu, M.-J.; Shen, X.; Sheng, L.; Mendoza, M.; Hanrahan, I. Validation of Enterococcus faecium NRRL B-2354 as a Surrogate for Listeria monocytogenes on Fresh Apples during Pilot Spray-Bar Peroxyacetic Acid Intervention. Food Control 2021, 119, 107472. [Google Scholar] [CrossRef]
- Turolla, A.; Sabatino, R.; Fontaneto, D.; Eckert, E.M.; Colinas, N.; Corno, G.; Citterio, B.; Biavasco, F.; Antonelli, M.; Mauro, A.; et al. Defence Strategies and Antibiotic Resistance Gene Abundance in Enterococci under Stress by Exposure to Low Doses of Peracetic Acid. Chemosphere 2017, 185, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Humayoun, S.B.; Hiott, L.M.; Gupta, S.K.; Barrett, J.B.; Woodley, T.A.; Johnston, J.J.; Jackson, C.R.; Frye, J.G. An Assay for Determining the Susceptibility of Salmonella Isolates to Commercial and Household Biocides. PloS One 2018, 13, e0209072. [Google Scholar] [CrossRef]
- Micciche, A.C.; Feye, K.M.; Rubinelli, P.M.; Lee, J.A.; Knueven, C.J.; Ricke, S.C. Comparison of Acid Sanitizers on Salmonella Typhimurium Inoculated Commercial Poultry Processing Reuse Water. Front. Sustain. Food Syst. 2019, 2, 90. [Google Scholar] [CrossRef]
- Alonso-Hernando, A.; Alonso-Calleja, C.; Capita, R. Effects of Exposure to Poultry Chemical Decontaminants on the Membrane Fluidity of Listeria monocytogenes and Salmonella enterica Strains. Int. J. Food Microbiol. 2010, 137, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Jolivet-Gougeon, A.; Sauvager, F.; Bonnaure-Mallet, M.; Colwell, R.R.; Cormier, M. Virulence of Viable but Nonculturable S. Typhimurium LT2 after Peracetic Acid Treatment. Int. J. Food Microbiol. 2006, 112, 147–152. [Google Scholar] [CrossRef]
- Bauermeister, L.J.; Bowers, J.W.J.; Townsend, J.C.; McKee, S.R. The Microbial and Quality Properties of Poultry Carcasses Treated with Peracetic Acid as an Antimicrobial Treatment. Poult. Sci. 2008, 87, 2390–2398. [Google Scholar] [CrossRef] [PubMed]
- Mathew, E.N.; Muyyarikkandy, M.S.; Bedell, C.; Amalaradjou, M.A. Efficacy of Chlorine, Chlorine Dioxide, and Peroxyacetic Acid in Reducing Salmonella Contamination in Wash Water and on Mangoes Under Simulated Mango Packinghouse Washing Operations. Front. Sustain. Food Syst. 2018, 2, 18. [Google Scholar] [CrossRef]
Figure 1.
Distribution of Enterococcus faecium (n=72) by acidic pH values.
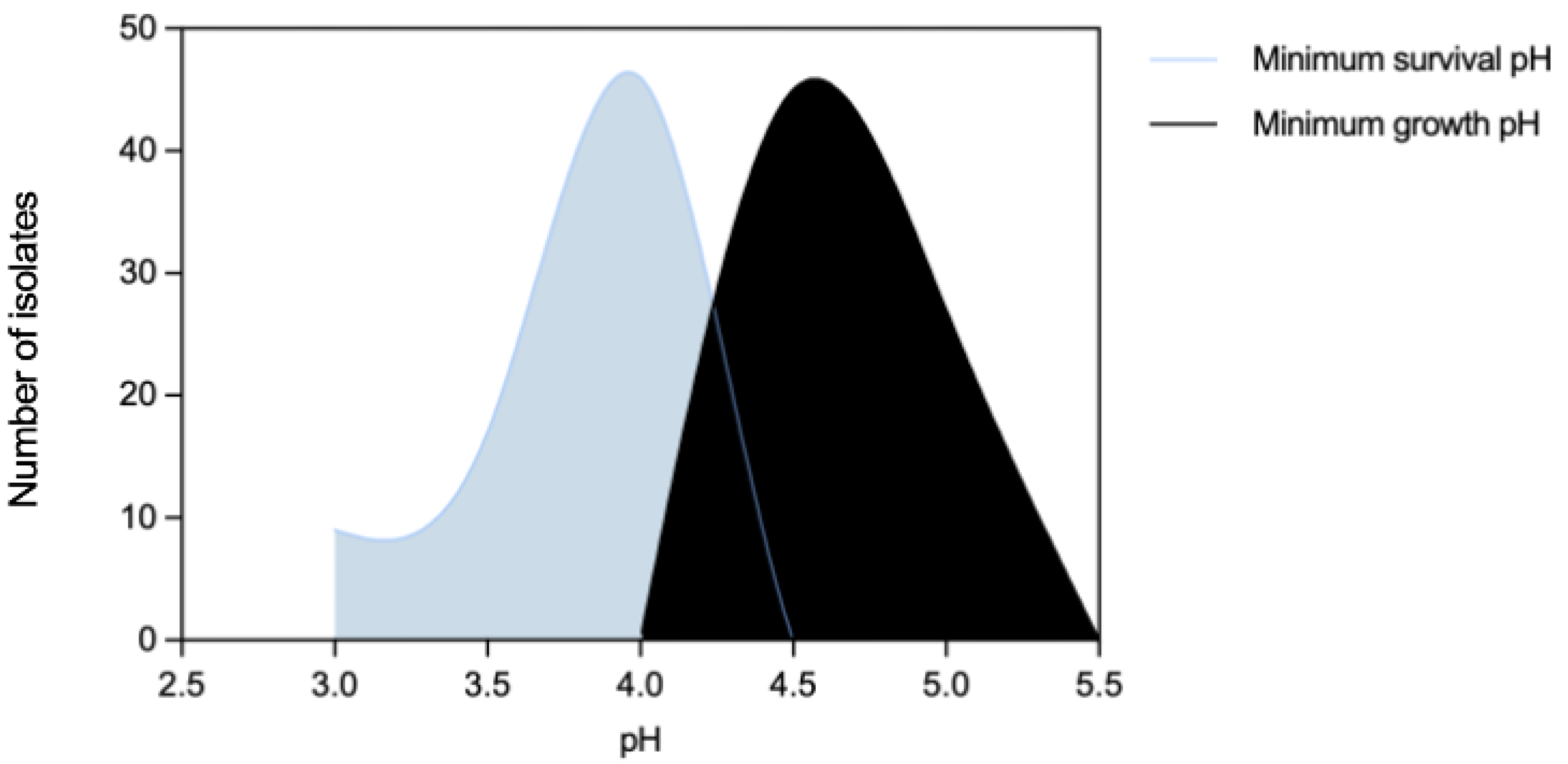
Figure 2.
Distribution of Enterococcus faecium (n=72) by Minimum Growth pH and Minimum Survival pH values according to clade type [(a), (b)], source [(c), (d)] and MDR profile [(e), (f)].
Figure 2.
Distribution of Enterococcus faecium (n=72) by Minimum Growth pH and Minimum Survival pH values according to clade type [(a), (b)], source [(c), (d)] and MDR profile [(e), (f)].
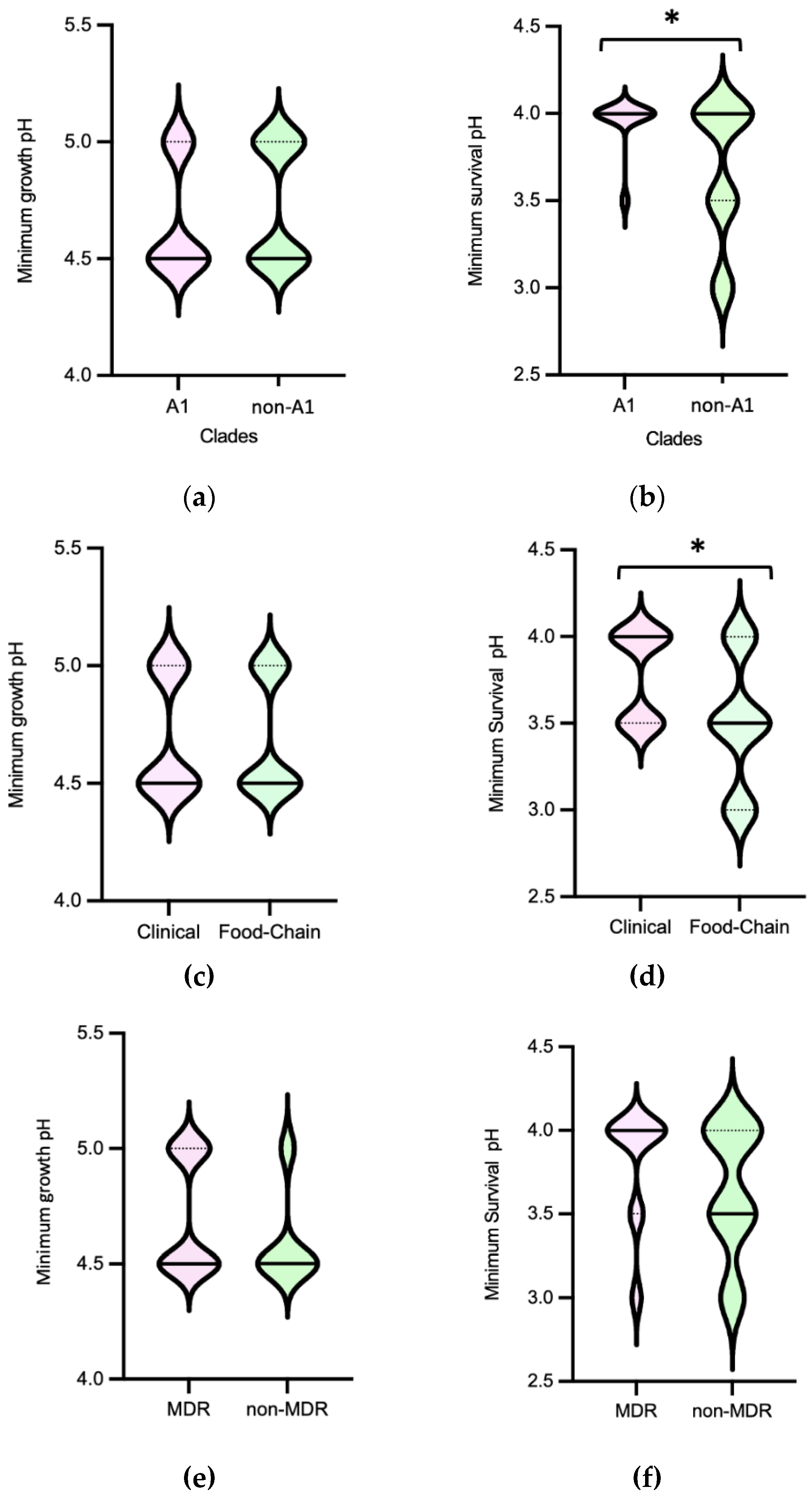
Figure 3.
Distribution of non-typhoidal Salmonella (n=60) by acidic pH values.
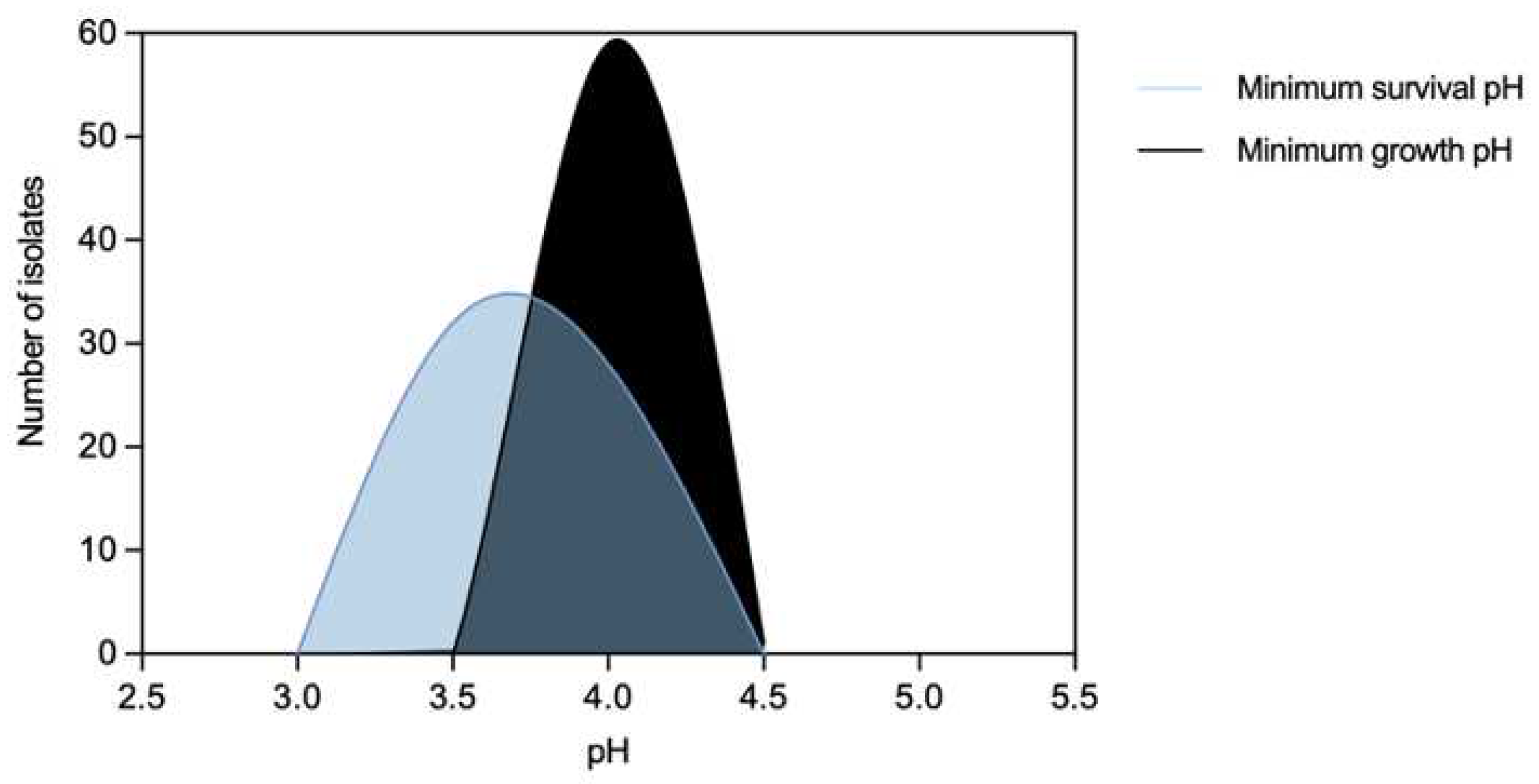
Figure 4.
Distribution of non-typhoidal Salmonella (n=60) by Minimum Growth pH and Minimum Survival pH values according to serogroup [(a), (b)], source [(c), (d)] and MDR profile [(e), (f)].
Figure 4.
Distribution of non-typhoidal Salmonella (n=60) by Minimum Growth pH and Minimum Survival pH values according to serogroup [(a), (b)], source [(c), (d)] and MDR profile [(e), (f)].
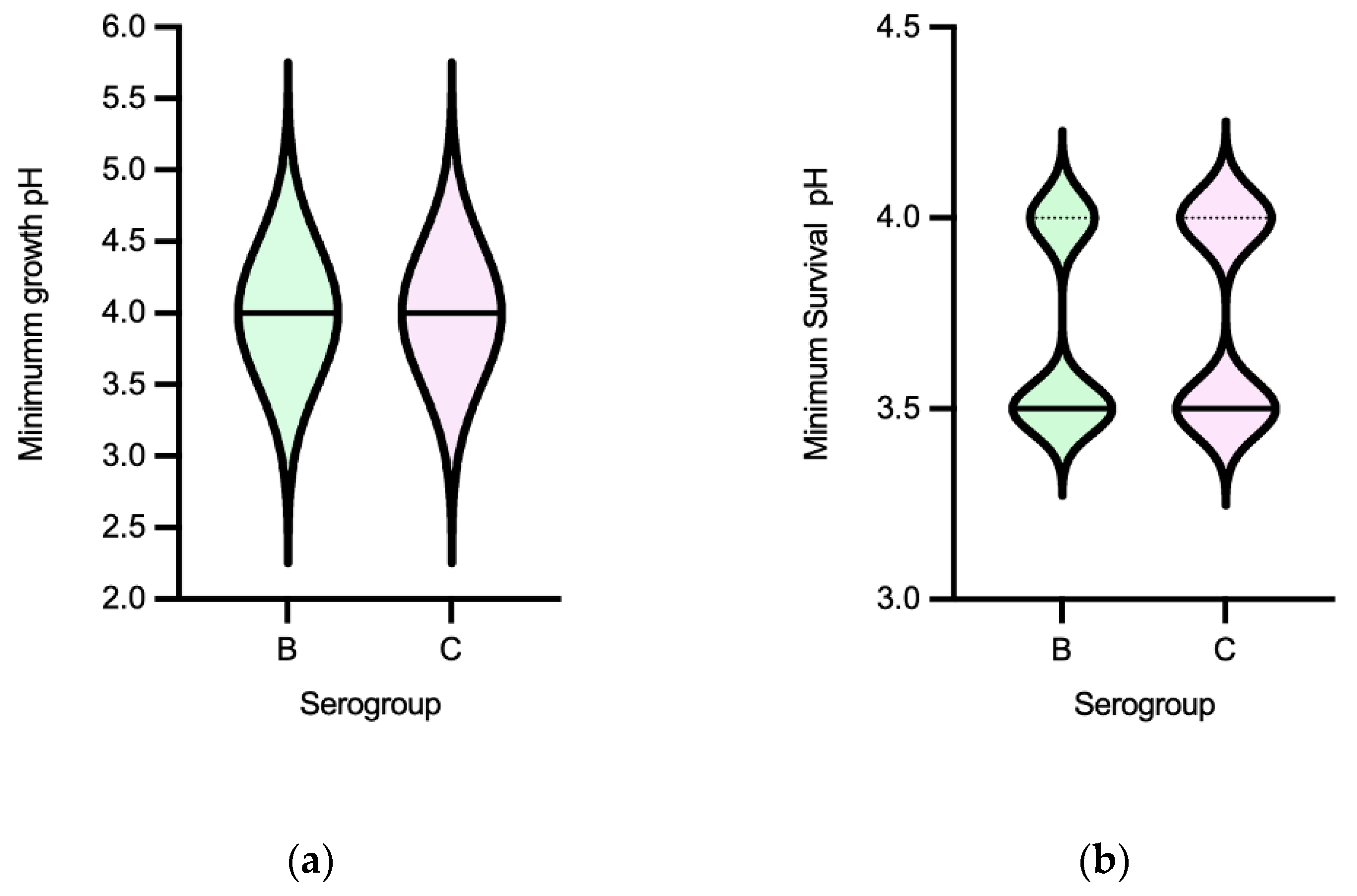
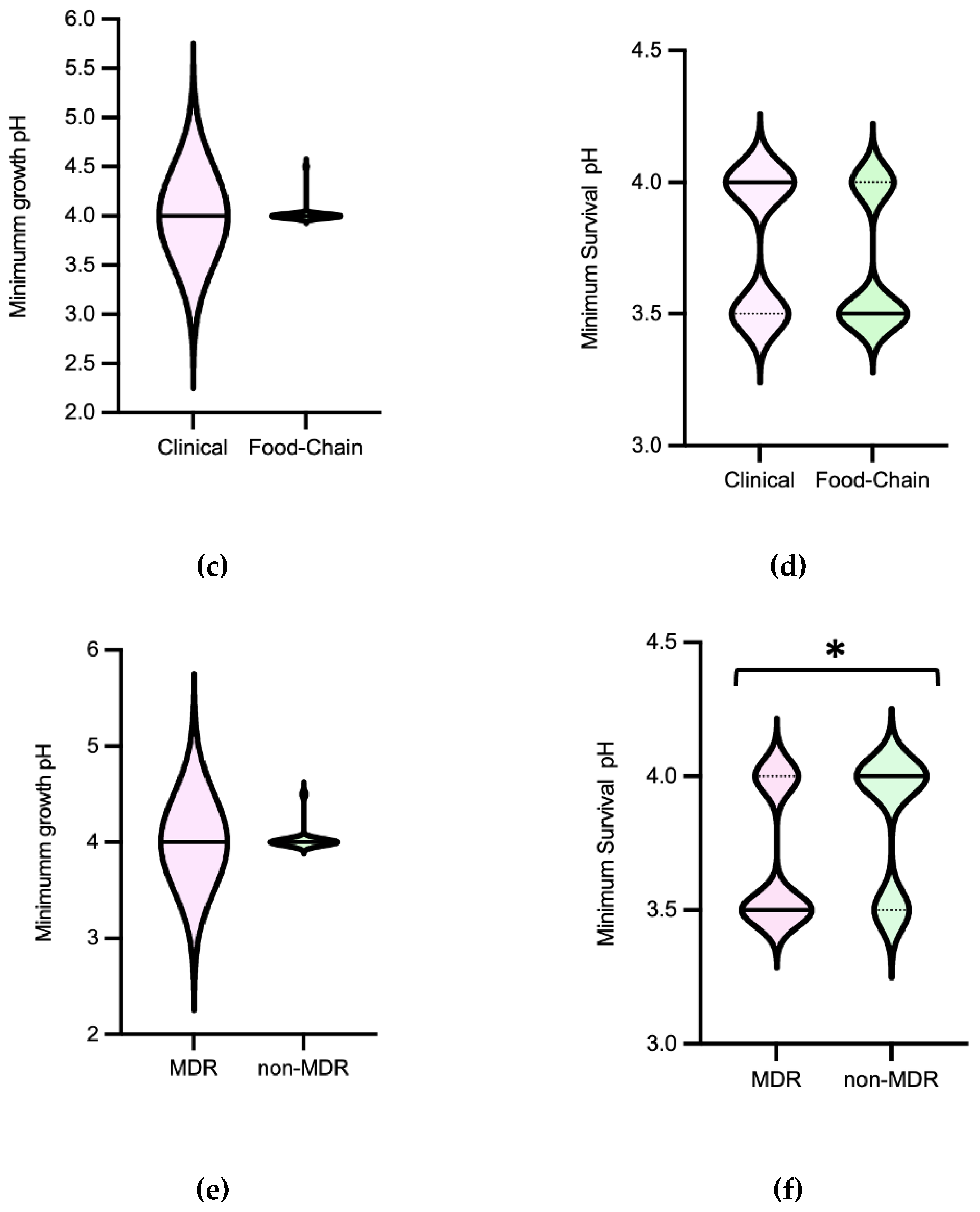
Figure 5.
Distribution of Enterococcus faecium (n=72) by Peracetic acid Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC).
Figure 5.
Distribution of Enterococcus faecium (n=72) by Peracetic acid Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC).
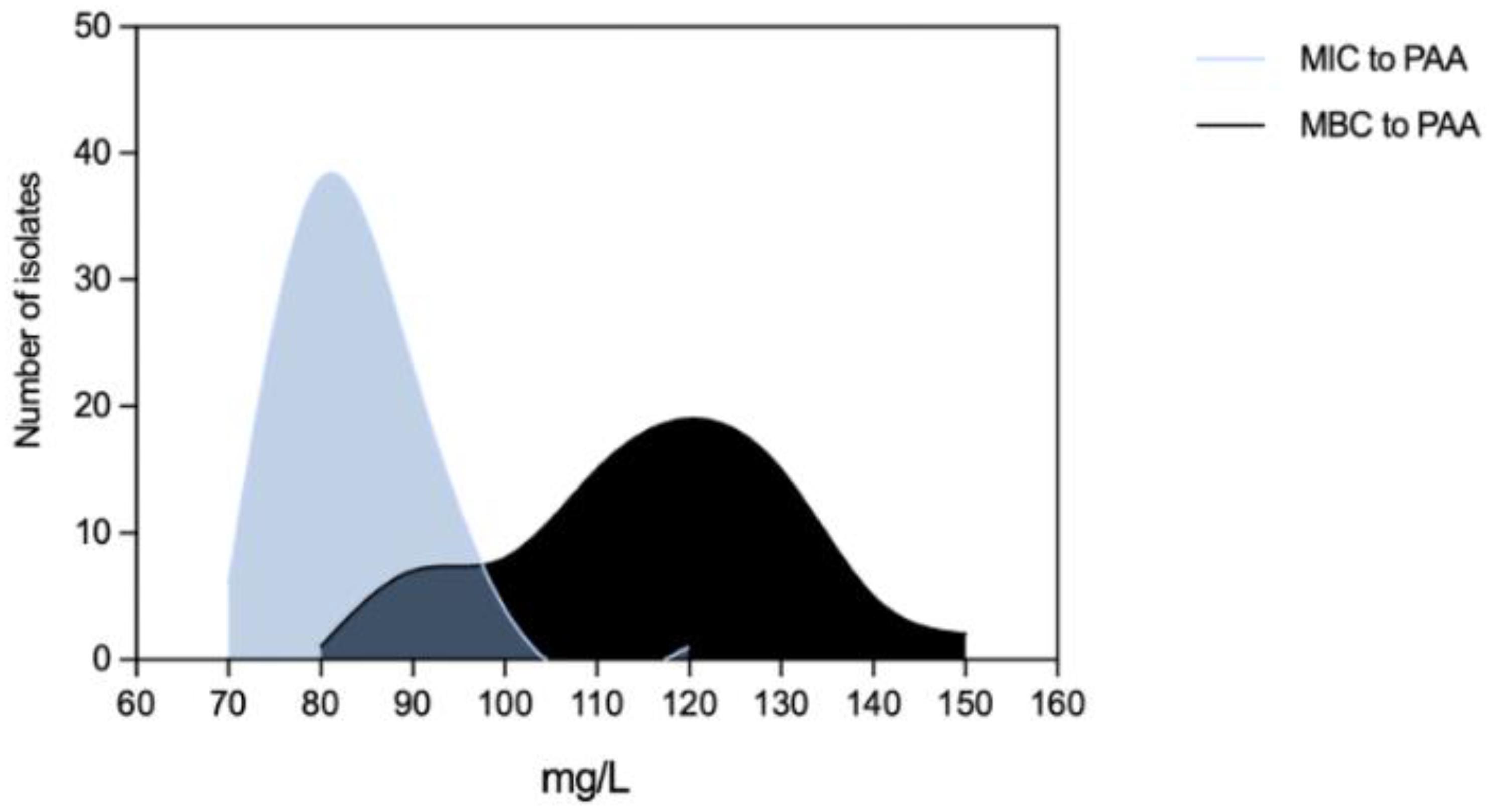
Figure 6.
Distribution of Enterococcus faecium (n=72) by Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC) to Peracetic acid according to clade type [(a), (b)], source [(c), (d)] and MDR profile [(e), (f)].
Figure 6.
Distribution of Enterococcus faecium (n=72) by Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC) to Peracetic acid according to clade type [(a), (b)], source [(c), (d)] and MDR profile [(e), (f)].
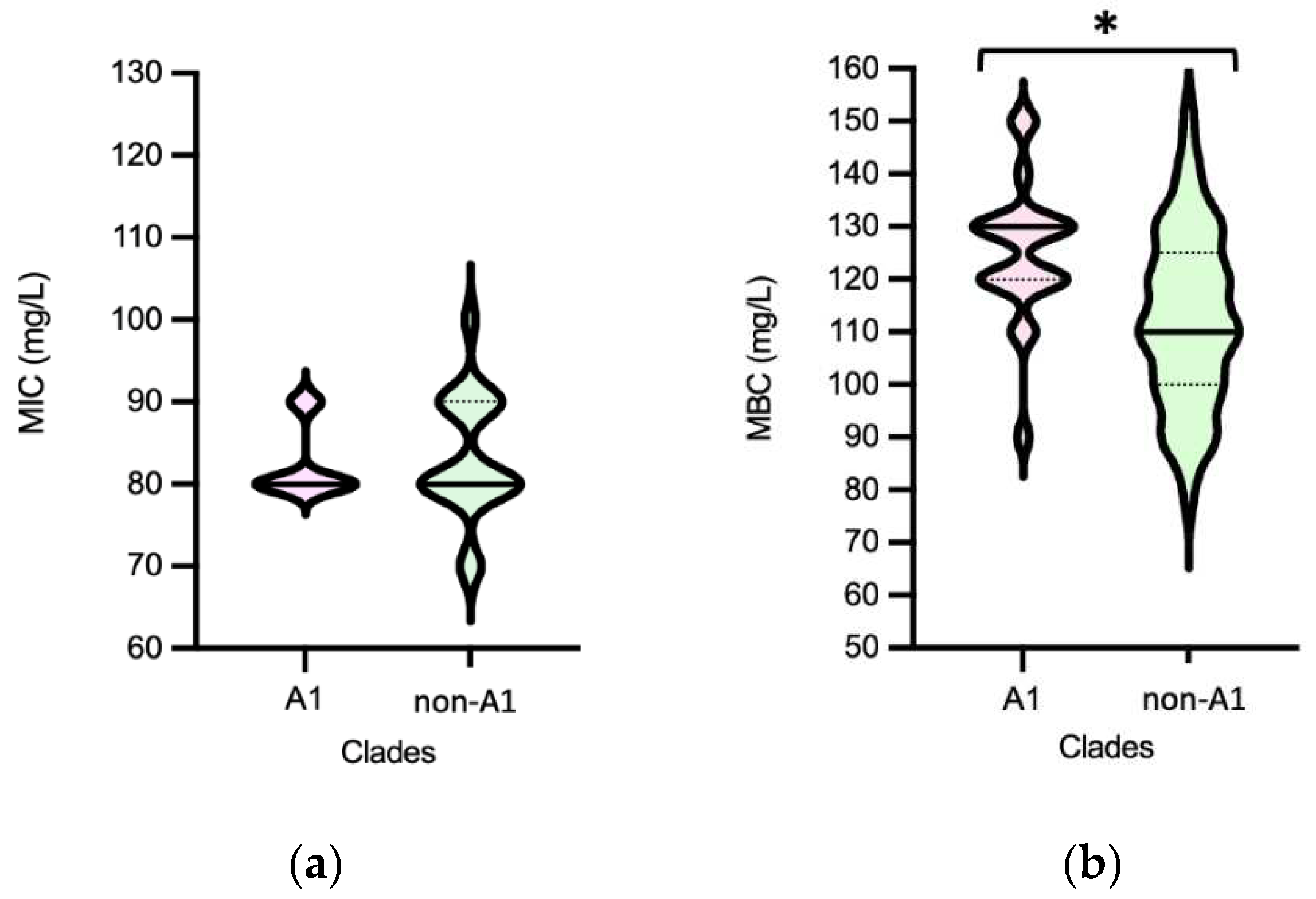
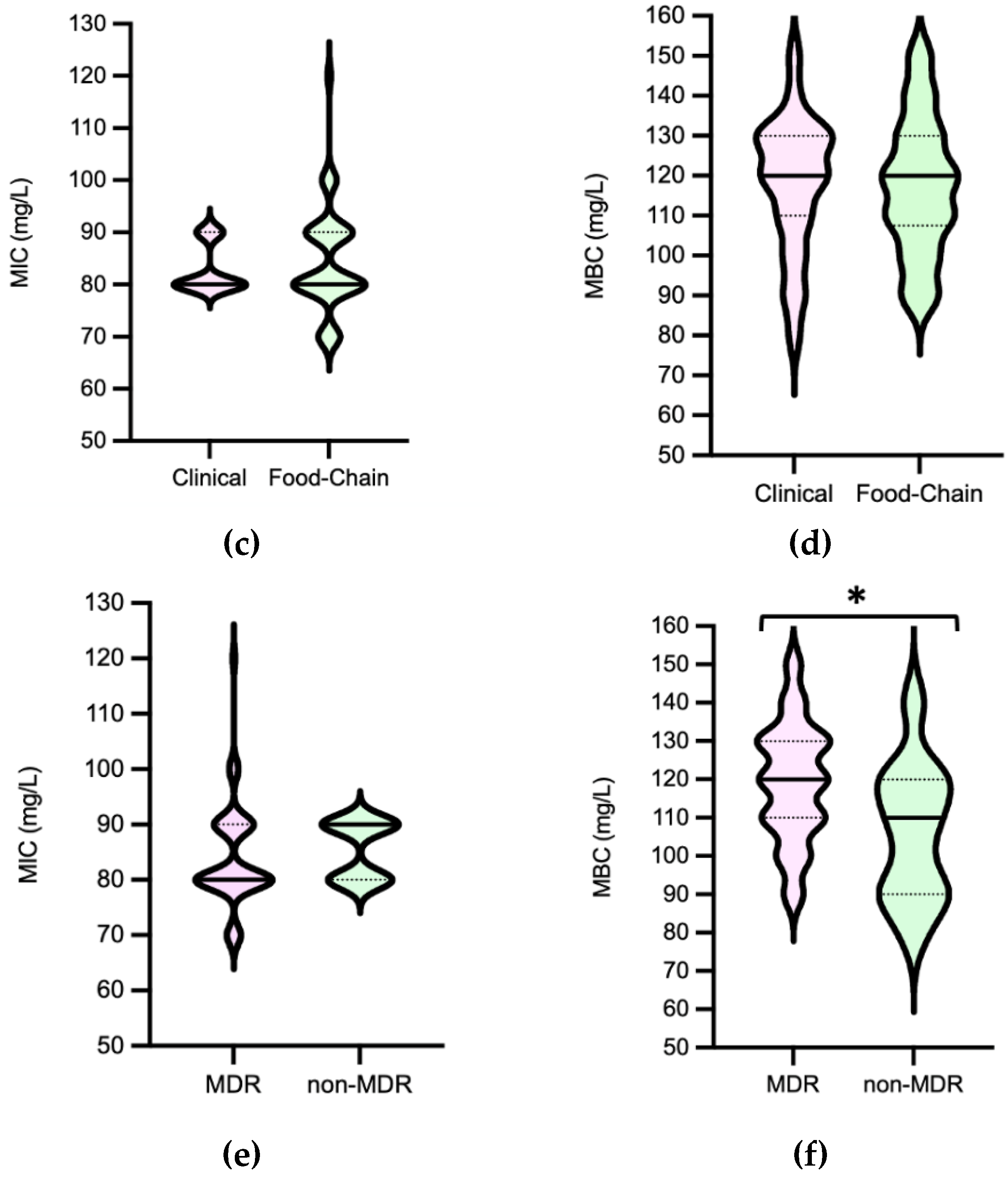
Figure 7.
Association of Peracetic Acid Minimum Inhibitory Concentrations (MIC) with Minimum Growth pH values [(a)] and Peracetic Acid Minimum Bactericidal Concentrations (MIC) with Minimum survival pH values [(b)] of Enterococcus faecium (n=72). The grey color means there is an overlapping of pink (clinical) and green (food-chain) isolates.
Figure 7.
Association of Peracetic Acid Minimum Inhibitory Concentrations (MIC) with Minimum Growth pH values [(a)] and Peracetic Acid Minimum Bactericidal Concentrations (MIC) with Minimum survival pH values [(b)] of Enterococcus faecium (n=72). The grey color means there is an overlapping of pink (clinical) and green (food-chain) isolates.
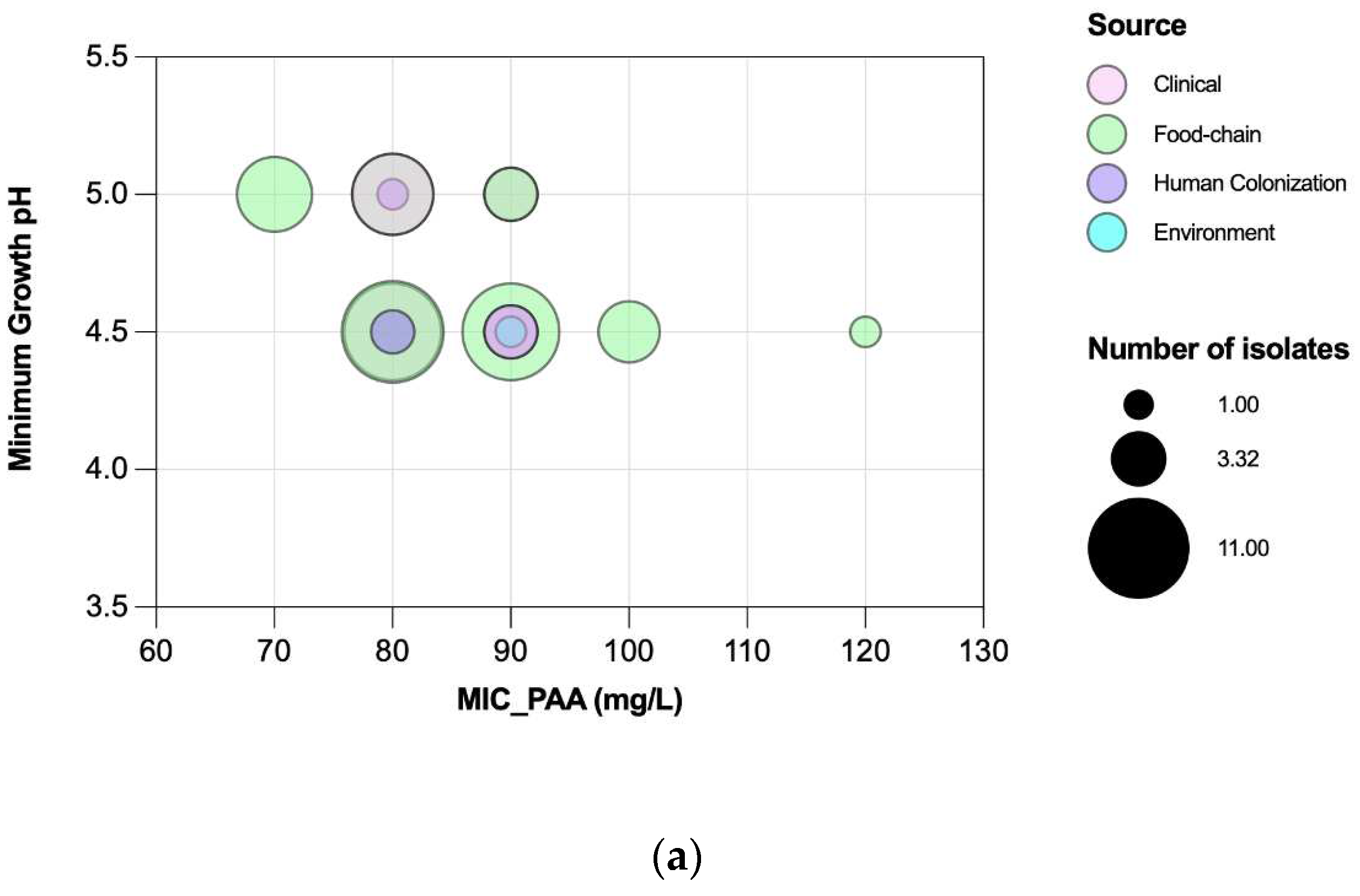
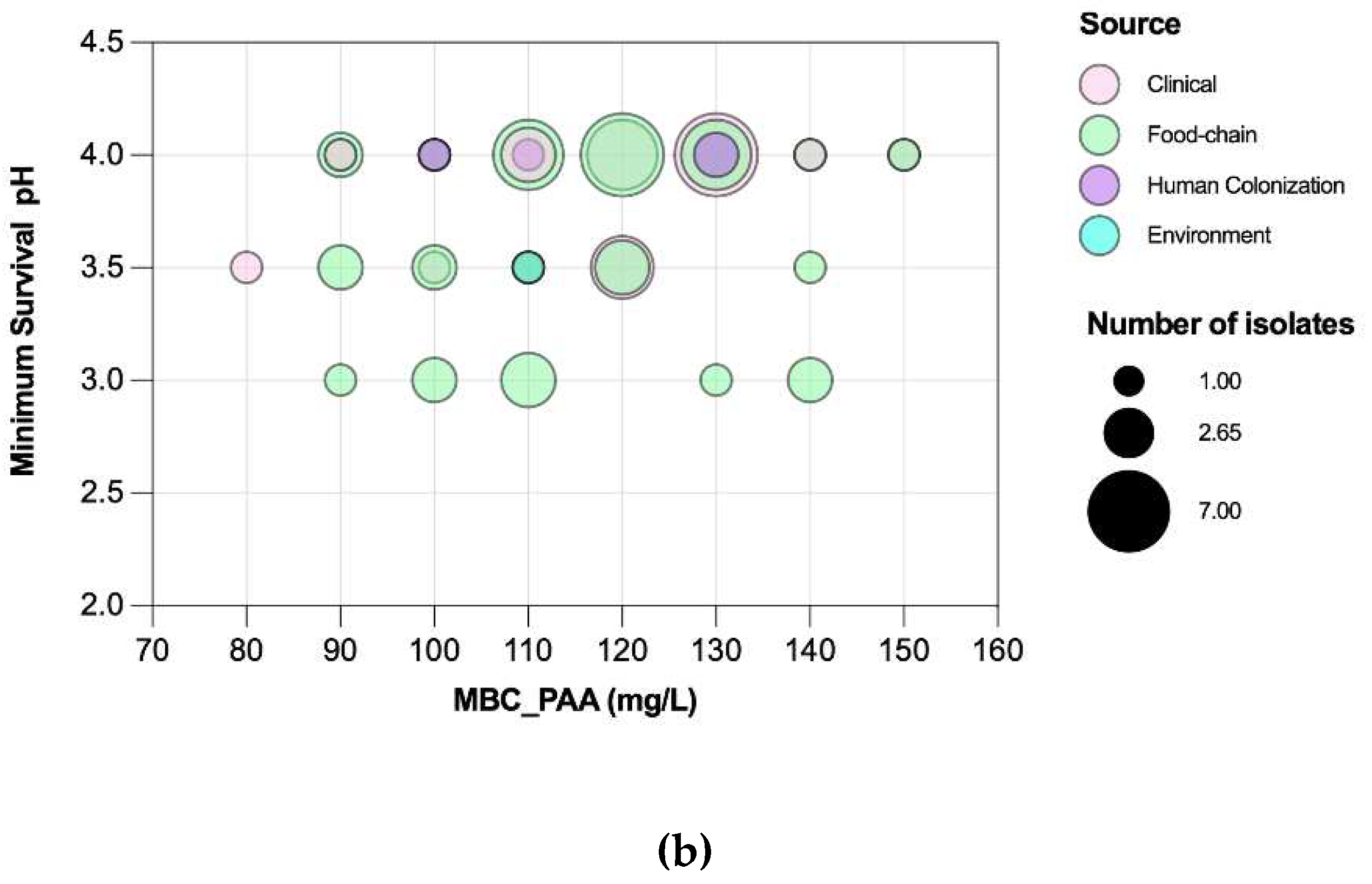
Figure 8.
Distribution of non-typhoidal Salmonella (n=60) by Peracetic acid Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC).
Figure 8.
Distribution of non-typhoidal Salmonella (n=60) by Peracetic acid Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC).
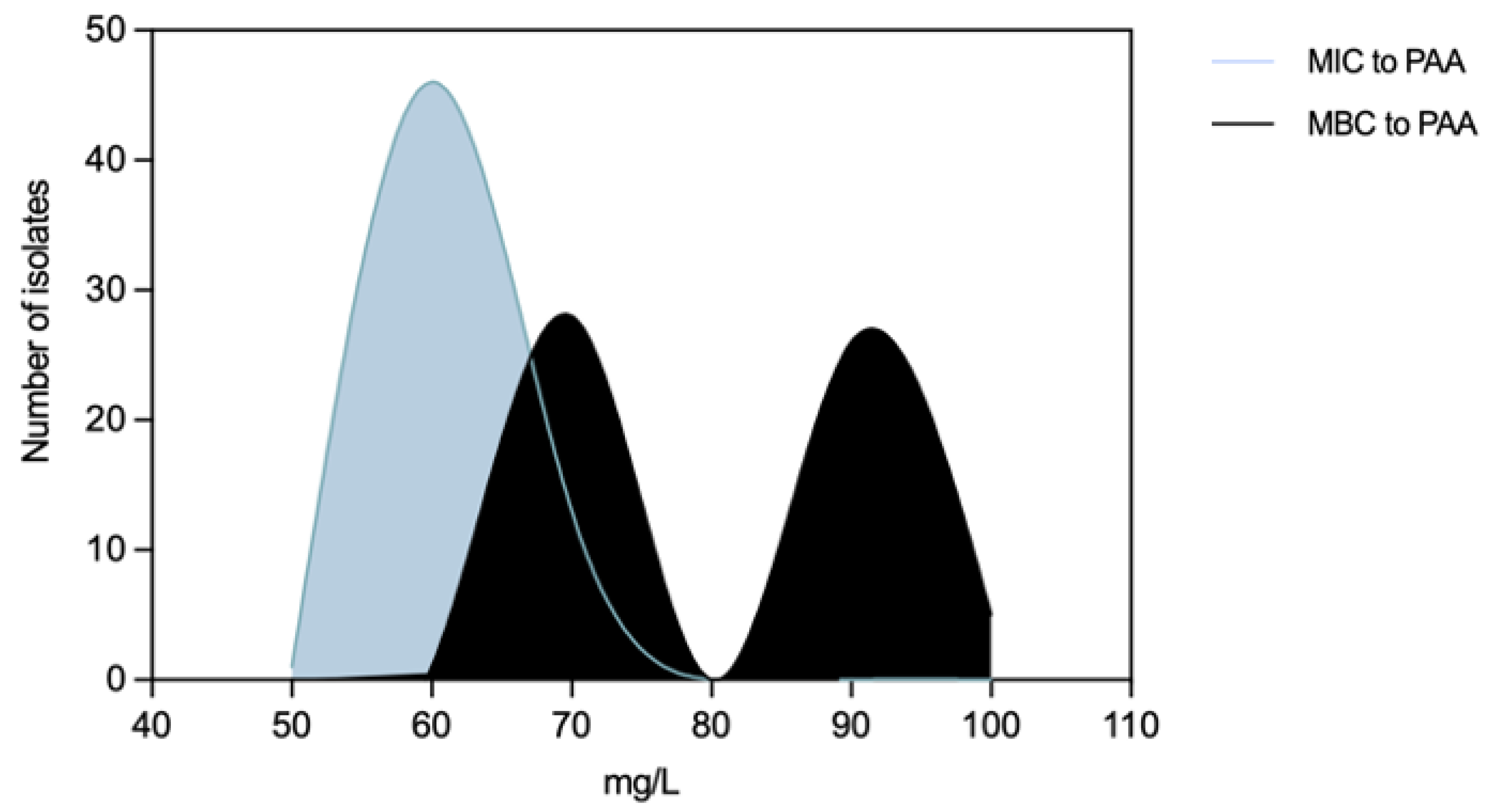
Figure 9.
Distribution of non-typhoidal Salmonella (n=60) by Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC) to Peracetic acid according to serogroups [(a), (b)], source [(c), (d)] and MDR profile [(e), (f)].
Figure 9.
Distribution of non-typhoidal Salmonella (n=60) by Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC) to Peracetic acid according to serogroups [(a), (b)], source [(c), (d)] and MDR profile [(e), (f)].
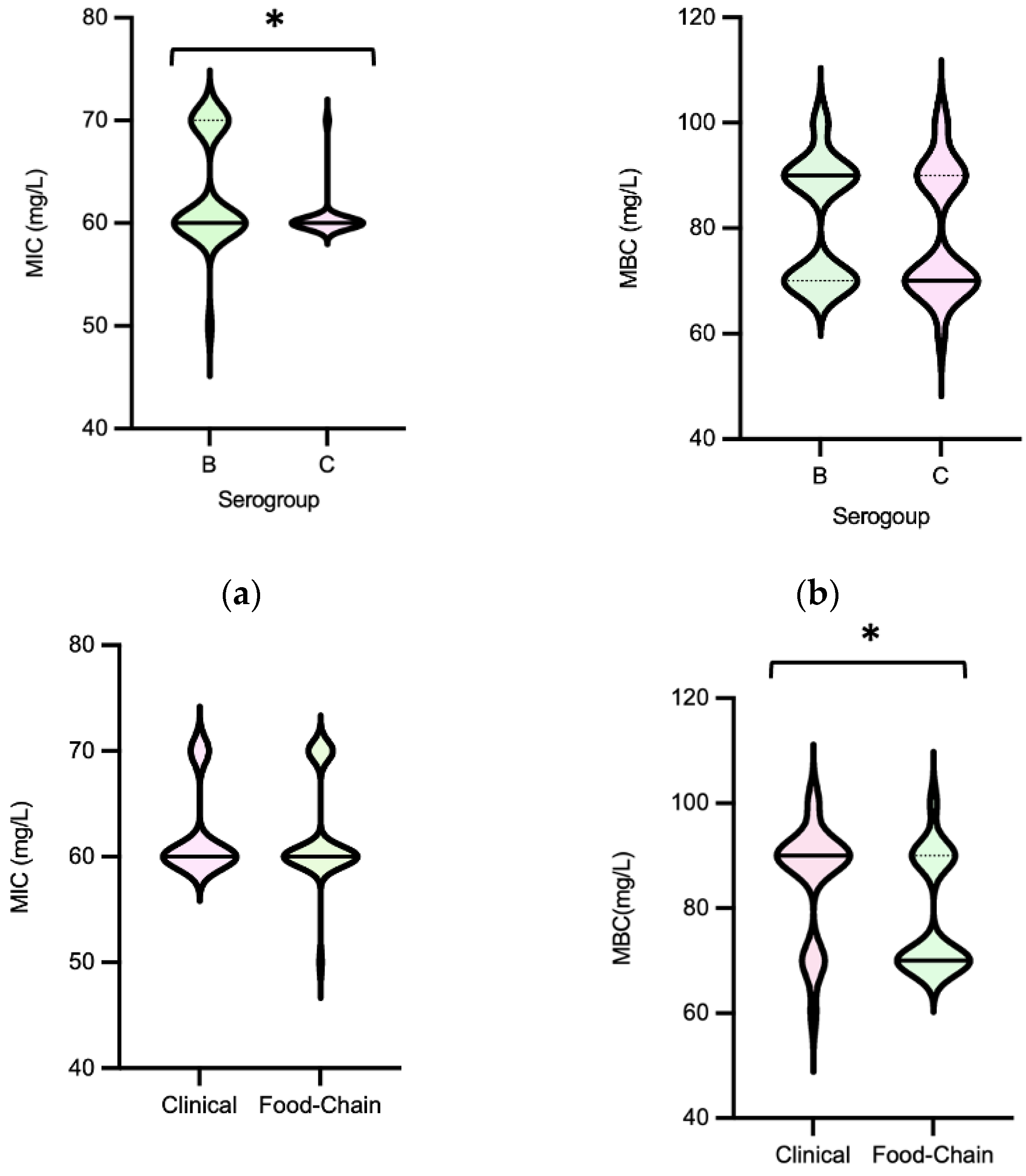
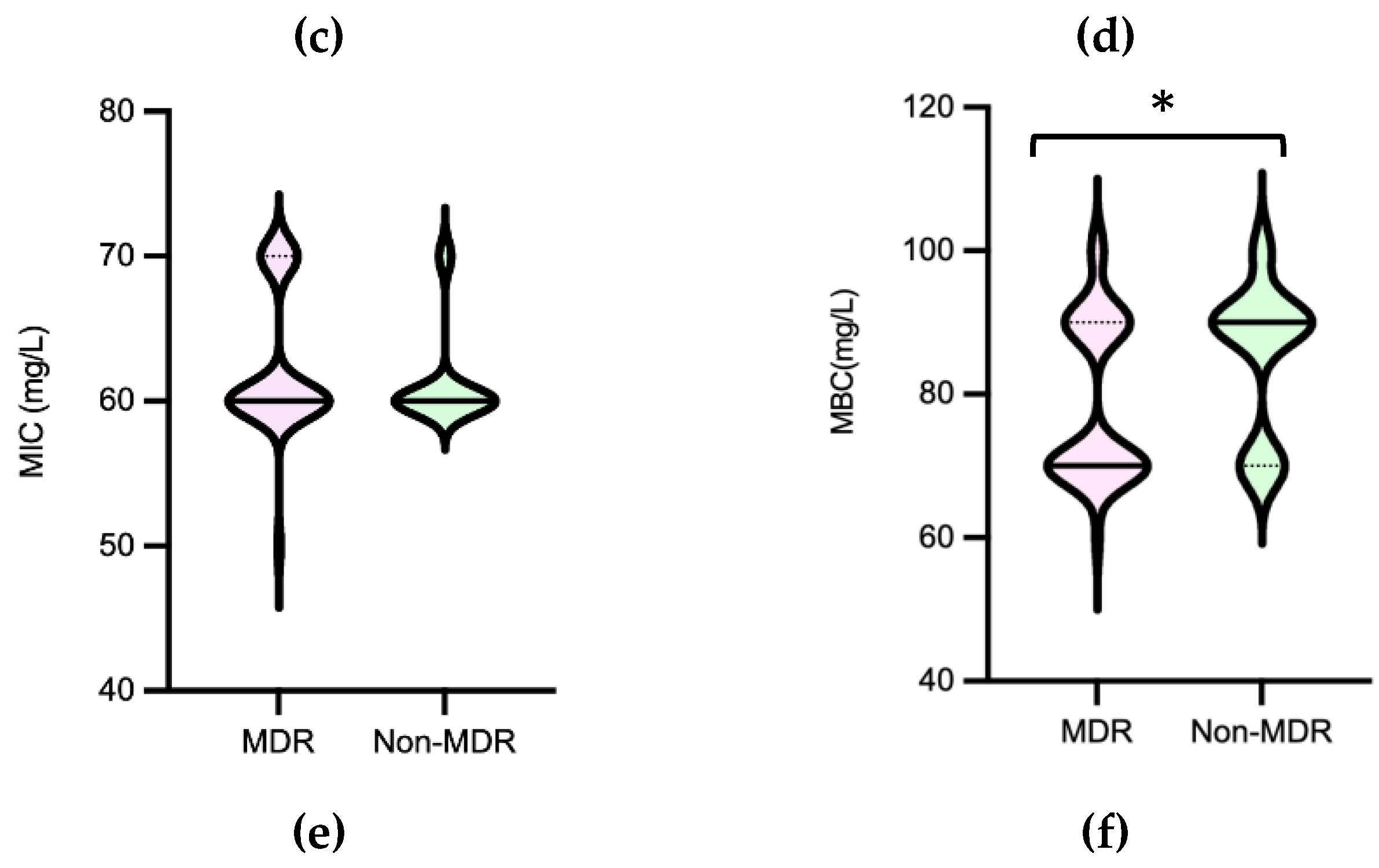
Figure 10.
Association of Peracetic Acid Minimum Inhibitory Concentrations (MIC) with Minimum Growth pH values [(a)] and Peracetic Acid Minimum Bactericidal Concentrations (MIC) with Minimum survival pH values [(b)] of non-typhoidal Salmonella (n=60). The grey color means there is an overlapping of pink (clinical) and green (food-chain) isolates.
Figure 10.
Association of Peracetic Acid Minimum Inhibitory Concentrations (MIC) with Minimum Growth pH values [(a)] and Peracetic Acid Minimum Bactericidal Concentrations (MIC) with Minimum survival pH values [(b)] of non-typhoidal Salmonella (n=60). The grey color means there is an overlapping of pink (clinical) and green (food-chain) isolates.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



