
Submitted:
24 July 2023
Posted:
26 July 2023
You are already at the latest version
Abstract
This study investigated the presence and characteristics of Juglans neotropica in three ecosystems in Southern Ecuador: Montane Evergreen Forest, Evergreen Seasonal Lower Montane Forest, and Semideciduous Foot Montane Forest. The main focus was on the species' multipurpose nature as both a fruit and timber source. Six study sites, totaling at least 0.5 hectares each, were established, with four in Loja province and two in Zamora Chinchipe province.The results showed significant differences in dendrometric variables across the sites, with the most favorable growth recorded in The Tundo, where trees exhibited an average diameter at breast height (DBH) of 45.16 cm, basal area (G) of 1.41 m2, total height (TH) of 19.22 m, canopy height (CH) of 13 m, cubic volume (CV) of 3.55 m3, and total volume (TV) of 5.22 m3. The species displayed a clumped distribution pattern, as indicated by a Morisita index greater than 1. Regarding abundance, the highest density of 297 trees per hectare was found in Argelia, while Victoria had the lowest density of 46 trees per hectare. This research highlights the importance of Juglans neotropica as a multipurpose species, providing both valuable fruits and wood resources in the studied ecosystems of Southern Ecuador.
Keywords:
Fruit
; forest
; ecosystem
; wood
; native
; Ecuador
1. Introduction
Understanding the distribution of a species is of utmost importance as it represents the geographic area where the species occurs and interacts with the ecosystem over time, be it short, medium, or long term [25]. While some species have a wide natural distribution, others have a limited range. A broader distribution increases the likelihood of encountering genetic variation, as different populations adapt to diverse local environments.
The Juglans genus belongs to the Juglandaceae family, which originated on the Asian continent approximately 56 million years ago. This genus includes species such as ailantifolia, mandshurica, and regia, with the latter being the most extensively cultivated and widely distributed across both the old and the New World. This is primarily due to its highly nutritious fruits and valuable wood [8- 3].
Around 23 million years ago, other American walnut species emerged, retaining physical characteristics inherited from their Asian counterparts. These species migrated from North America to Central America, specifically Mexico, with J. olanchana as the epicenter of speciation for other Central and South American species. Among these species, J. boliviana is found in Bolivia, J. australis in Argentina, and J. neotropica in Colombia. The latter species also extends its presence to Peru and was discovered in Ecuador around the 15th century [14].
Understanding the historical distribution patterns and speciation events of Juglans species is essential for various reasons. Firstly, it provides insights into the evolutionary processes that have shaped the genetic diversity and adaptations observed in different populations. Secondly, it aids in conservation efforts by identifying regions of high species richness and potential genetic reservoirs. Lastly, it enables the development of informed strategies for the management and sustainable use of these valuable resources. (2)
By examining the distribution of Juglans species across time and space, researchers can unravel the intricate relationships between the organisms and their environment. This knowledge can contribute to the conservation and sustainable utilization of these species, ensuring their survival in the face of environmental changes and human activities. Moreover, it offers opportunities for further research on the ecological interactions, evolutionary history, and potential applications of these remarkable trees.
In this study, we aim to expand our understanding of the distribution patterns and evolutionary dynamics within the Juglans genus. By examining historical records, genetic data, and ecological factors, we seek to shed light on the factors influencing the distribution and diversification of Juglans species. Through this research, we hope to contribute to the broader knowledge of plant evolution, conservation, and sustainable land management practices [1]
Juglans neotropica, commonly known as "Nogal" or "Tocte," is an arboreal plant native to the South American Andes, specifically Ecuador, Colombia, Peru, and Bolivia. This species is of particular interest due to its wide distribution and cultural significance in Ecuador. It has been reported in various provinces of Ecuador, including Bolívar, Loja, Azuay, Tungurahua, Chimborazo, Pichincha, Napo, and Galapagos [23].
In Ecuador, J. neotropica is predominantly found in the inter-Andean region, encompassing the valleys and foothills of the Andes mountain range. The species thrives in diverse ecosystems characterized by different altitudinal ranges. Within the Southern Montane Evergreen Forest of the Eastern Cordillera of the Andes, J. neotropica is distributed between 2200 and 3000 meters above sea level (masl). Specifically, it can be found in towns such as The Argelia, The Zañe, The Tibio, and The Merced. Furthermore, in the Catamayo-Alamor Semideciduous Foot Montane Forest ecosystem, which ranges from 400 to 1600 masl, J. neotropica is present in the provenances of The Tundo and The Victoria. Notably, in The Tundo provenance, walnut trees were also observed in the Evergreen Seasonal Montane Forest ecosystem, situated under the Catamayo-Alamor, at altitudes ranging from 1600 to 2000 masl.
The distribution of J. neotropica extends beyond South America, with a surprising occurrence in New Zealand, specifically in the city of Auckland. However, its presence in South America, particularly in Ecuador, holds historical and cultural significance. There is evidence of its cultivation in the Equatorial Andes since pre-Columbian times, highlighting its long-standing importance to local communities [4].
Understanding the distribution patterns and ecological preferences of J. neotropica is crucial for its conservation and sustainable management. Moreover, investigating the factors that contribute to its successful adaptation to different altitudes and ecosystems can provide valuable insights into the species' ecological resilience and potential for future cultivation. This article aims to further explore the distribution, habitat preferences, and cultural significance of J. neotropica in Ecuador, shedding light on its ecological role and potential conservation strategies. By analyzing existing literature and conducting field surveys, we aim to contribute to the broader understanding of this valuable species and promote its sustainable use and preservation in Ecuador and beyond.
Regarding the regulatory and protection entities responsible for endangered species in Ecuador, particularly J. neotropica, there is currently a lack of comprehensive information regarding its occurrence, distribution, abundance, and overall conservation status. As a result, the conservation efforts for this species have been hindered.
J. neotropica is currently classified as an endangered species, facing severe threats that have greatly hindered its regeneration in the early stages of growth. Anthropogenic activities, such as the recruitment of natural regeneration and expansion of agricultural land into forested areas, have played a significant role in this decline. This species is known to grow in a diverse range of soil types, including Ultisols, Entisols, Inceptisols, Alfisols, and Mollisols.
In terms of its ecological guild, J. neotropica exhibits a semi-heliophytic nature, requiring shade during its initial stages of growth but becoming heliophytic as an adult, dominating the upper canopy of the forest. Additionally, as a deciduous species, the leaf litter of J. neotropica plays a vital role in maintaining ecosystem balance by providing a source of energy, contributing nutrients, and forming humic substances in the soil. The most significant impact on this species is observed in South America's dry and montane forests, which serve as its natural habitat [10,11,12,13,14,15,16,17,18,19,20,21,22,23,24].
2. Materials and Methods
2.1. Study Area
The study area of this research is situated in southern Ecuador or Zone 7 of Ecuador, encompassing the geographic coordinates 3°30' and 5°0' south latitude, and 78°20' and 80°30' west longitude. It shares borders with zones 5 and 6 to the north, Peru to the south and east, and Peru and the Pacific Ocean to the west [21]. This region holds great ecological significance due to its distinct topographic, altitudinal, climatic, and soil characteristics.
To conduct the present study, we conducted fieldwork in the three provinces within Zone 7 that had documented occurrences of the species J. neotropica. These provinces are Loja (11065.6 km2), El Oro (5866.6 km2), and Zamora Chinchipe (10559.7 km2), collectively covering an area of 27491.9 km2, which represents approximately 11% of the total land area of Ecuador [21].
In order to provide a comprehensive analysis of J. neotropica and its associated factors, we selected these provinces as they exhibit diverse characteristics that are vital to understanding the species' distribution and ecology. By focusing on these specific regions, we aim to contribute valuable insights into the habitat preferences, population dynamics, and conservation implications of J. neotropica within Zone 7.
Through our investigation, we seek to shed light on the ecological importance of this area and provide valuable information that can guide future conservation efforts. The findings of this study will help to improve our understanding of J. neotropica's habitat requirements, enabling more effective conservation strategies to protect this species and its associated ecosystems in Southern Ecuador.
2.2. Occurrence
To determine the occurrence of the species in Southern Ecuador, we consulted databases from nationally and internationally recognized entities such as Geo-Cat, Tropics Database, and the Ecuadorian Biodiversity System (BNDB). These platforms contain a comprehensive collection of regional and international botanical data sourced from various institutions, collectors, and research projects, both public and private.
Additionally, we gathered information from the decentralized Autonomous Governments (GAD´s), public institutions like MAATE, and private landowners. We also accessed data from the National University of Loja's Reinaldo Espinosa Herbarium and the Private Technical University of Loja, which house collections of books containing valuable information on the flora of the Andean forest.
To validate the occurrence of the species, we carefully verified the geographic details of all the collected information. Furthermore, we utilized internet resources to obtain reports on the species' occurrence. Notably, the most reliable and accurate information came from residents and owners of private land where the species was suspected to exist.
All the collected information was meticulously entered into the ARCGIS 10.8 geographic information system to generate precise geographic locations for the occurrence of the species.
By following this comprehensive methodology, we ensured that our research on the occurrence of the species in Southern Ecuador was based on reliable data from reputable sources and validated through geographic verification and resident reports. The utilization of the ARCGIS 10.8 system further enhanced the accuracy of our findings.
2.3. Structure
Following the identification of species occurrence, a series of field trips were conducted to select suitable sites where J. neotropica was found. These field trips spanned a duration of 12 months. Each site was visited a minimum of three and a maximum of five times, depending on the forest size.
To assess the parameters and structural indices, a comprehensive forest inventory was conducted, employing two distinct methods:
1. Statistical Method: A 100% inventory was performed for areas ≤ one hectare. For areas larger than one hectare, a simple random sampling approach was implemented.
2. Objective-based Method: An inventory was carried out for the management of natural forests, with the intensity ranging from 1% to 5% of the total area [6].
To determine the reliability and representativeness of the sampled area, the sampling intensity formula (I) was applied.
By applying these methods, we ensured a rigorous and comprehensive assessment of the forest structure, allowing for accurate data collection and analysis. The statistical method provided a systematic approach, while the objective-based method facilitated the management of natural forests. The combination of these approaches ensured a robust evaluation of the species' habitat and supported the reliability of our findings.
The utilization of multiple visits to each site and the implementation of various inventory methods enabled us to capture a comprehensive picture of the forest's structure and characteristics. This approach contributes to the overall validity and reliability of our study, ensuring that our conclusions are well-founded and representative of the larger population of interest.
Where:
I = Sampling intensity
Ss = Surface of the sample
Pa = Population area
100 = Constant
2.3.1. Structural Parameters
To assess the structural characteristics of the study area, individuals with a diameter at breast height (DBA1.30m) of ≥ 10 cm were georeferenced. These individuals, commonly referred to as stems in forestry, were located using a Garmin Montana 650 GPS device.
Once the inventory and georeferencing of J. neotropica individuals were completed, various dasometric parameters were calculated, including DBH, TH, CH, G(m2), and V(m3). The DBH data of the inventoried individuals were then organized into diameter classes of 10 cm width, following the established protocol [11].

Table 1.
Dasometric parameters.
| Denomination | Formula |
|---|---|
| Diameter at breast height, above ground level | DBH1.30 m |
| Total height | TH(m) |
| Commercial height | CH(m) |
| Basal area of an individual | (2) |
| Basal area of the population | (3) |
| Volume of the stem in cubic meters | (4) |
2.3.2. Structural Indices
Subsequently, a structural valuation index was calculated:
Morisita index:
Where:
Iδ= Spatial distribution index
q= number of frames
ni= Number of individuals in í-th square
N= Total number of individuals in all q squares
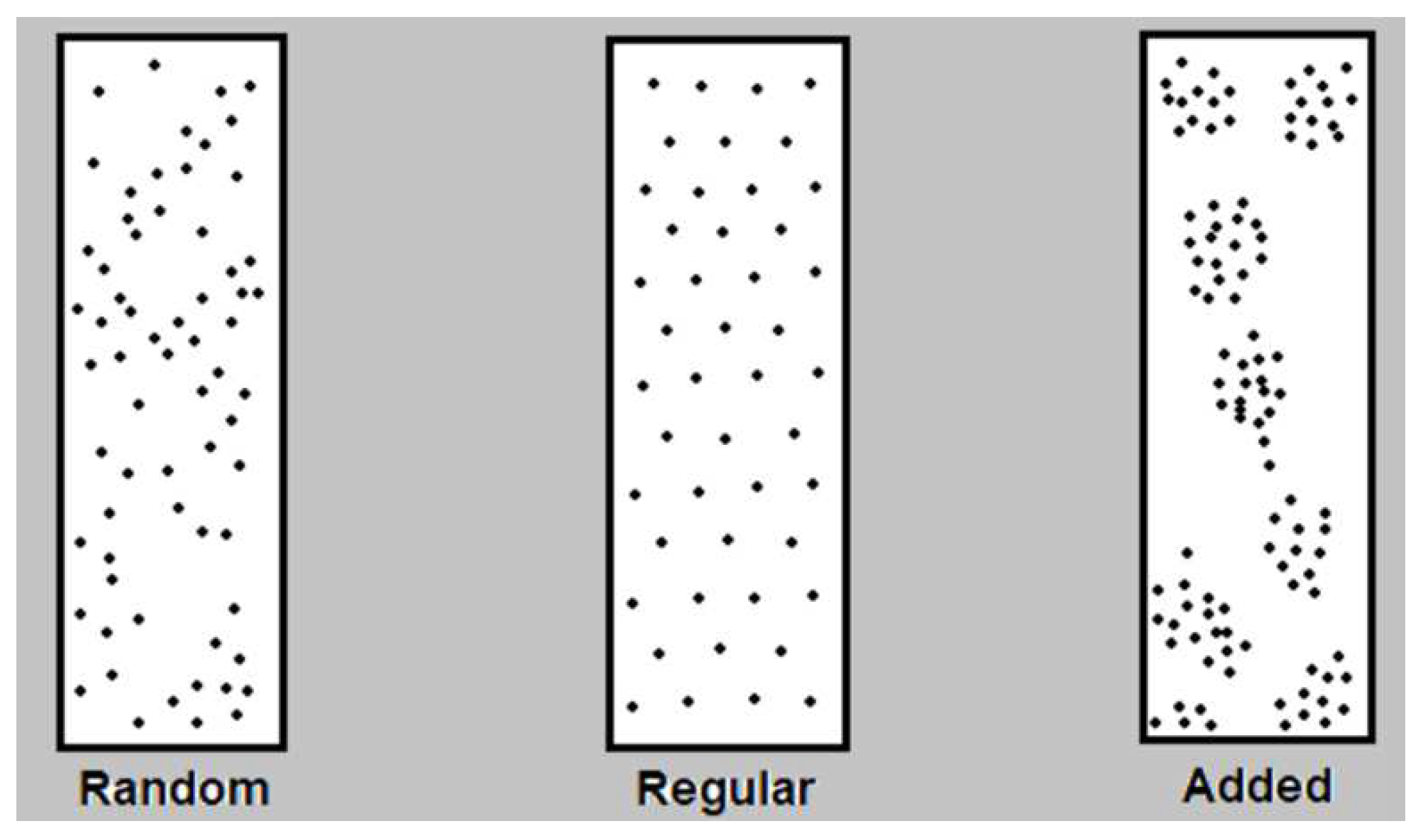
Values of < 1 regular or uniform distribution, equal to one random distribution and greater than one indicate an aggregated distribution, respectively (Morisita 1959).
2.4. Abundance
To assess the abundance of J. neotropica, we followed the guidelines set forth in [12], as outlined in ministerial agreement 125, article 11. According to these guidelines, a species is considered to have low abundance when the density is less than one tree per three hectares (0.33 trees/ha−1). Hence, we determined the abundance of J. neotropica by identifying the reference tree in each site and applying the specified criteria. This approach allows for an accurate evaluation of the species' abundance and facilitates comparisons across different sites.
Finally, the following formula was applied:
Where:
Ai = Absolute abundance
= Number of individuals of a species present in an area
3. Results
3.1. Ocurrence the J. neotropica
The natural and artificial occurrence of J. neotropica species was observed in six localities within southern Ecuador, encompassing the provinces of Loja, El Oro, and Zamora Chinchipe. These occurrences were identified in forested areas measuring ≥ 0.5 hectares. Specifically, four instances were recorded in the province of Loja, while the remaining two were documented in the province of Zamora Chinchipe (Table 2).
In this study, the provenances were located in terms of political and geographical location, as can be seen in (Figure 2)
3.2. Structural Parameters
In terms of structural parameters, we successfully determined the dasometric variables for all the localities with areas exceeding 0.5 hectares. The data obtained represents the number of individuals recorded within the sampled areas, appropriately adjusted to hectares to facilitate interpretation (Figure 3).
3.2.1. Horizontal Structure
The analysis of the horizontal structure across all provenances revealed significant variations in diameter classes and the number of individuals within each class. The Bosque Protector el Tundo exhibited the highest diversity with 12 distinct diameter classes. Additionally, this provenance recorded the largest individuals in terms of diameter (DBH1.30), with an impressive measurement of 134 centimeters. These findings emphasize the importance of considering provenance when assessing the horizontal structure of forest stands, as it can significantly influence the distribution and size of trees within a given area. These results contribute to our understanding of forest dynamics and have implications for sustainable forest management practices.
Among the different provenances studied, The Argelia naturalized forest displayed the highest density of individuals per hectare, with a remarkable count of 399 ind/ha−1. This finding highlights the exceptional abundance and richness of this particular forest ecosystem. The results underscore the significance of The Argelia as a biodiverse habitat, showcasing its potential for conservation efforts and sustainable management. The high density of individuals within this forest emphasizes its ecological value and warrants further investigation into the factors contributing to its remarkable success. These findings provide valuable insights for forest management strategies and conservation initiatives in similar ecosystems.
- Morisita Index
The analysis of the Morisita Index (Iδ) revealed a distribution pattern greater than 1 in all the studied sampling units of the various provenances. This finding indicates an aggregated distribution pattern.
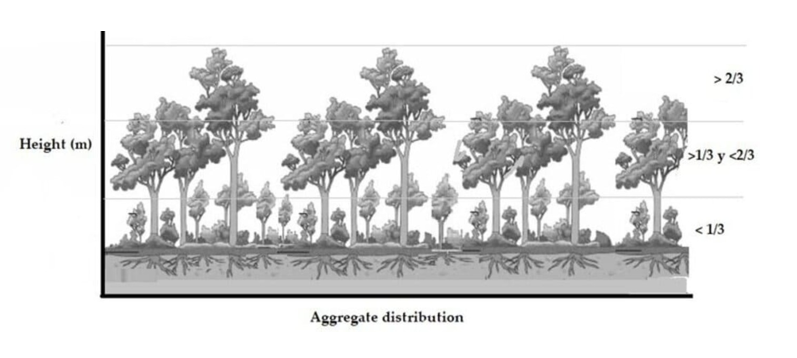
3.2.2. Vertical Structure
In relation to the vertical structure observed in all the identified provenances, our findings demonstrate that the arboreal component of J. neotropica attains impressive heights, ranging from 24 to 36 meters, depending on the provenance. Notably, the tallest individuals were found within the The Tibio Protected Forest and the Tundo provenances. This study sheds light on the remarkable vertical growth potential of J. neotropica and highlights the significant role played by specific provenances, such as The Tibio Protected Forest and the Tundo, in nurturing exceptionally tall individuals of this species. These results provide valuable insights for the management and conservation of J. neotropica populations (Figure 5).
3.2.3. Growth Variability
Regarding the growth behavior of dasometric variables among provenances in southern Ecuador, our study successfully demonstrated significant differences between the J. neotropica forests' dasometric variables using ANOVA. These findings highlight the variability in growth patterns across different provenances. The results contribute valuable insights into the understanding of tree growth in this region and have important implications for forest management and conservation strategies. Our study underscores the need for considering provenance-specific factors when developing sustainable forestry practices. These findings provide a solid foundation for future research and decision-making in the context of forest ecosystems in Southern Ecuador (Figure 6).
3.3. Abundance
The number of registered individuals of the J. neotropica species varied across different localities per hectare (Figure 7). The total count for each locality is as follows:
4. Discussion
4.1. Occurrence
The occurrence of the J. neotropica species was investigated in southern Ecuador, specifically in the provinces of Loja and Zamora Chinchipe, excluding the province of Oro. Six localities with areas greater than 5000 m2 were identified as potential habitats for the species. Previous studies support the findings of this research on the occurrence of the species in the identified localities.
One study conducted by [7] reported the presence of the Tundo walnut forest in the Sozoranga Canton of the province of Loja. This forest comprises 96 hectares of native forest, located at altitudes ranging from 800 to 2645 meters above sea level (masl). The area has a strongly inclined relief, which aligns with the findings of our study regarding the occurrence of the J. neotropica species.
Additionally, [18]documented the occurrence of the species in the Loja canton of the Loja province, specifically in the Argelia sector. In this location, a planted forest approximately 70 years old has provided a suitable habitat where the J. neotropica species has naturalized effectively. The forest covers an area of 0.7 hectares and spans an altitudinal range of 2170 to 2250 masl, corroborating the findings of our study.
However, our research has revealed new occurrences of the J. neotropica species in previously unreported locations. The first location is Cerro el Zañe in the Loja canton, characterized by an altitudinal range of 2100 to 2800 masl and a strongly inclined relief. The second location, the Victoria, corresponds to the Macará canton and spans an altitudinal range of 1400 to 1600 meters above sea level in the province of Loja. Furthermore, the species has been observed in the Merced, which spans an altitudinal range of 200 to 2200 masl in the Zamora canton, as well as in The Tibio, with an altitudinal range of 2400 to 2600 masl, also located in the Zamora canton of the Zamora Chinchipe province. The precise locations of these occurrences have been reported for the first time in this study.
According to [9], the Juglans species exhibits a relative gregariousness in mature forest conditions, resulting in its scarcity within natural ecosystems. This pattern aligns with the findings of our research, further supporting the similarity between our study and previous observations of Juglans species.
4.2. Structure
Regarding the structural parameters of the forest in the discovered provenances of the J. neotropica species, all the dendrometric variables were correctly identified. However, the limited availability of research related to this species makes it challenging to compare our findings with other studies. Nevertheless, a study by [17] notes that the planted forest of J. neotropica in Algeria exhibits very similar dendrometric variables to those observed in our study. Additionally, [5] mentions that young walnut trees have well-formed stems, reaching heights of 25 to 30 meters with a diameter at breast height (DBH) of 90 cm. Our observations align with these findings at the study sites.
In terms of Morisita's structural index, the species demonstrates an aggregated distribution behavior (Iδ > 1.0), indicating a preference for dominating specific sectors within the ecosystem. As described by [15], this type of distribution is not random but rather irregular, occurring in response to local habitat differences (microhabitats). The authors also highlight that aggregate distribution is the most frequent pattern observed in nature and is a result of the inherent tendency of individuals to aggregate. Therefore, plants tend to disperse their seeds in close proximity or in the same location where they reside.
Furthermore, the distribution of the species indicates a decreasing number of individuals over time, suggesting a gradual decline. However, it can still be found, albeit with difficulty, between 2200-3000 meters above sea level (masl) in natural forest ecosystems covering areas greater than 5000 m2.
4.3. Abundance
The abundance data obtained reveals notable differences in the number of individuals per hectare for the species J. neotropica across different provenances. The highest abundance was observed in the town of Argelia, followed by Zañe and Tibio. According to a previous study [17], the Algerian forest, being a forest plantation, exhibited the highest number of individuals per unit area in this investigation. This particular provenance, with an approximate age of 70 years, has adapted remarkably well to the prevailing climatic conditions at the site.
The Tundo Protected Forest and the Tibio provenance, which are native forests, also reported a relatively higher number of individuals per unit area according to another study [13]. Despite being located in different provinces, these forests share similar climate characteristics, which contribute to their higher abundance.
However, when considering the area extension, it is worth noting that the Tundo provenance boasts the largest forest area, covering approximately 130 hectares, whereas Argelia has the smallest extent, with only 0.7 hectares.
In terms of J. neotropica abundance, variations were observed in the number of individuals per hectare across the different provenances. These differences can be attributed to the severe threat posed to these ecosystems by various anthropic activities, such as livestock rearing and agriculture [16]. As a consequence, the species has been categorized as endangered by [10].
To better understand the abundance patterns and conservation status of J. neotropica, it is crucial to consider the interplay between provenance, climate conditions, and anthropic activities. The Algerian provenance, originating from a forest plantation, exhibited the highest abundance due to its successful adaptation to the local climatic conditions over several decades. Meanwhile, the Tundo Protected Forest and the Tibio provenance, being native forests, demonstrated a relatively higher abundance owing to their favorable climate characteristics despite their geographical separation. However, the overall abundance of J. neotropica is alarmingly affected by human activities, with livestock rearing and agriculture exerting significant pressure on the species' survival.
5. Conclusions
In southern Ecuador, there are not many relicts or fragments of native forests of J.neotropica, the few fragments that exist presented a vertical structure in the form of an inverted j, which means that there is good growth dynamics, insofar as to the vertical structure, the individuals presented a dominance of the site with respect to other individuals that share their habitat. However, the number of individuals per hectare is not abundant in all locations.
The data obtained from the analysis of variance between the dasometric variables in relation to the different provenances of J. neotropica indicate that there are significant statistical differences, finding better growth in the fragments of forests with ecosystems of Semideciduous Forest Foot Montane of Catamayo-Alamor, which has a altitudinal variation of 400-1600 masl, as well as ecosystems of the Lower Montane Seasonal Evergreen Forest of Catamayo-Alamor, which has an altitudinal variation of 1600-2000 masl.
The number of trees found in the Argelia forest provenance is greater in relation to the other provenances because this is a naturalized plantation with more than 70 years of age. However, its forest area is very small with 0.7 hectares and with lower dasometric growth than the rest of the provenances studied.
Wild fruit trees, especially the J. neotropica species, play a crucial role in preserving biodiversity and providing natural resources in various ecosystems. It is essential to recognize the significance of these species in conserving flora and fauna, particularly in places like the Tundo walnut forest in the canton of Sozoranga, where favorable conditions exist for their development. The protection and promotion of these wild trees are fundamental to maintaining environmental harmony and balance, ensuring a sustainable future for future generations.
Author Contributions
Conceptualization, methodology, writing, B.P.H, Review, S.P.L and D.P.C. All authors have read and agree with the published version of the manuscript.
Funding
This research did not receive external funding
Data Availability Statement
The data presented in this study are available upon request from the corresponding Author.
Acknowledgments
Private forest owners as well as government entities. Volunteers from the private research laboratory NEOSILVICULTURA TROPICAL who helped in the acquisition of field data over several years.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aradhya, M. K., Potter, D. A. N. I. E. L., & Simon, C. J. (2004, November). Origin, evolution, and biogeography of Juglans: a phylogenetic perspective. In V International Walnut Symposium 705 (pp. 85-94). [CrossRef]
- Frei, J. (2021). A Brief History of Juglandaceae. Arnoldia, 78(3), 10-17.
- Guo, W., Chen, J., Li, J., Huang, J., Wang, Z., & Lim, K. J. (2020). Portal of Juglandaceae: A comprehensive platform for Juglandaceae study. Horticulture research, 7. [CrossRef]
- Bachman, S., Moat, J., Hill, A. V., De la Torre, J., & Scott, B. (2011). Supporting Red List threat assessments With. e-Infrastructures for data publishing in biodiversity science, 150, 117.
- Barreto, G., & Herrera, J. (1990). INDERNED. Publisher Guadalupe LTDA.
- CATIE. (2002). Forest inventories for broadleaf forests in Central America (Orozco Lorena & Brumér Cecelia, Eds.). CATIE. (Google Scholar) (Cross Ref).
- Dueñas, D., Guevara, O., & Santacruz, S. (2022). Valoración económica de los bienes y servicios ecosistémicos del bosque protector Jatumpamba-Jorupe. Revista Geoespacial, 19(1), 12-32.
- Fjellstrom, RG, & Parfitt, DE (1995). Phylogenetic analysis and evolution of the genus Juglans (Juglandaceae) as determined from nuclear genome RFLPs. Plant Systematics and Evolution, 197(1-4), 19-32. [CrossRef]
- Garcia Rodriguez, Calixto Jose. 2002. Ecological, silvicultural and utilization study of walnut (Juglans olanchana Standl. & LO Williams) in broadleaf forests of Honduras. Special project as a requirement to qualify for the title of agricultural engineer. Zamorano, Honduras. 38p.
- IUCN Red List of Threatened Species: Juglans neotropica. IUCN Red List of Threatened Species. (IUCN.
- Lamprecht, H. (1990). Silvicultura en los trópicos: los ecosistemas forestales en los bosques tropicales y sus especies arbóreas; posibilidades y métodos para un aprovechamiento sostenido.
- MAE. (2015). Las Normas para el Manejo Forestal Sostenible de los Bosques Húmedo. Acuerdo, N. 0125.
- MAE. 2013. Ecuador, M. D. (2013). Sistema de clasificación de los Ecosistemas del Ecuador Continental. Subsecretaría de Patrimonio Natural. Quito.
- Manning, W. E. (1960). The Genus Juglans in South America and the West Indies. Brittonia, 12(1), 1. [CrossRef]
- Morláns, C., & María, B. (2014). Introducción a la ecología de poblaciones.
- Nieto, V. M., & Rodríguez, J. (2002). Juglans neotropica Diels. Tropical seed manual. United States Department of Agriculture Forest Service. USA.
- Palacios, B., Aguirre, Z., Pucha, D., Mocha, J., Feijoo, C., Jumbo, J., & Rojas, A. (2017). Form Factor and Productivity of a Plantation of Juglans neotropica Diels. Established in the premises of the National University of Loja.
- Palacios, B., Mendoza, ZA, Lozano, D., & Yaguana, C. (2016). WEALTH, STRUCTURE AND TREE DIVERSITY OF THE LOWER MOUNTAIN FOREST, ZAMORA CHINCHIPE-ECUADOR. Bosques Latitud Cero, 6(2), Art. 2. https://revistas.unl.edu.ec/index.php/bosques/article/view/233.
- Ramos Veintimilla, RA, Murillo Gamboa, O., & Gallo, LA (2020). Potential for genetic improvement in Juglans neotropica Diels, at 10 months of age in Tunshi, Chimborazo/Potential of Genetic Improvement in Juglans neotropica Diels, at 10 months of age in Tunshi, Chimborazo. KnE Engineering. [CrossRef]
- Ronnie de Camino, V. (1999). Sustainable forest management in Latin America: relevant actors and policies.
- SEMPLADES 2017. General Undersecretary of Planning for Good Living. Quito, Ecuador.
- Townsend, J. P., Cavalieri, D., & Hartl, D. L. (2003). Population genetic variation in genome-wide gene expression. Molecular Biology and Evolution, 20(6), 955-963.
- Tropics. (1982). Juglans neotropica. https://www.tropicos.org/name/16700014 (Tropicos.org).
- Toro Vanegas, E., & Roldán Rojas, I. C. (2018). Estado del arte, propagación y conservación de Juglans neotropica Diels., en zonas andinas. Madera y bosques, 24(1).
- Zunino, M., & Palestrini, C. (1991). El concepto de especie y la biogeografía. Anales de biología, vol. 17, 1991.
Figure 2.
Geographic location of occurrences of the specie J. neotropica in areas greater than 0.5 hectares of forest in southern Ecuador.
Figure 2.
Geographic location of occurrences of the specie J. neotropica in areas greater than 0.5 hectares of forest in southern Ecuador.
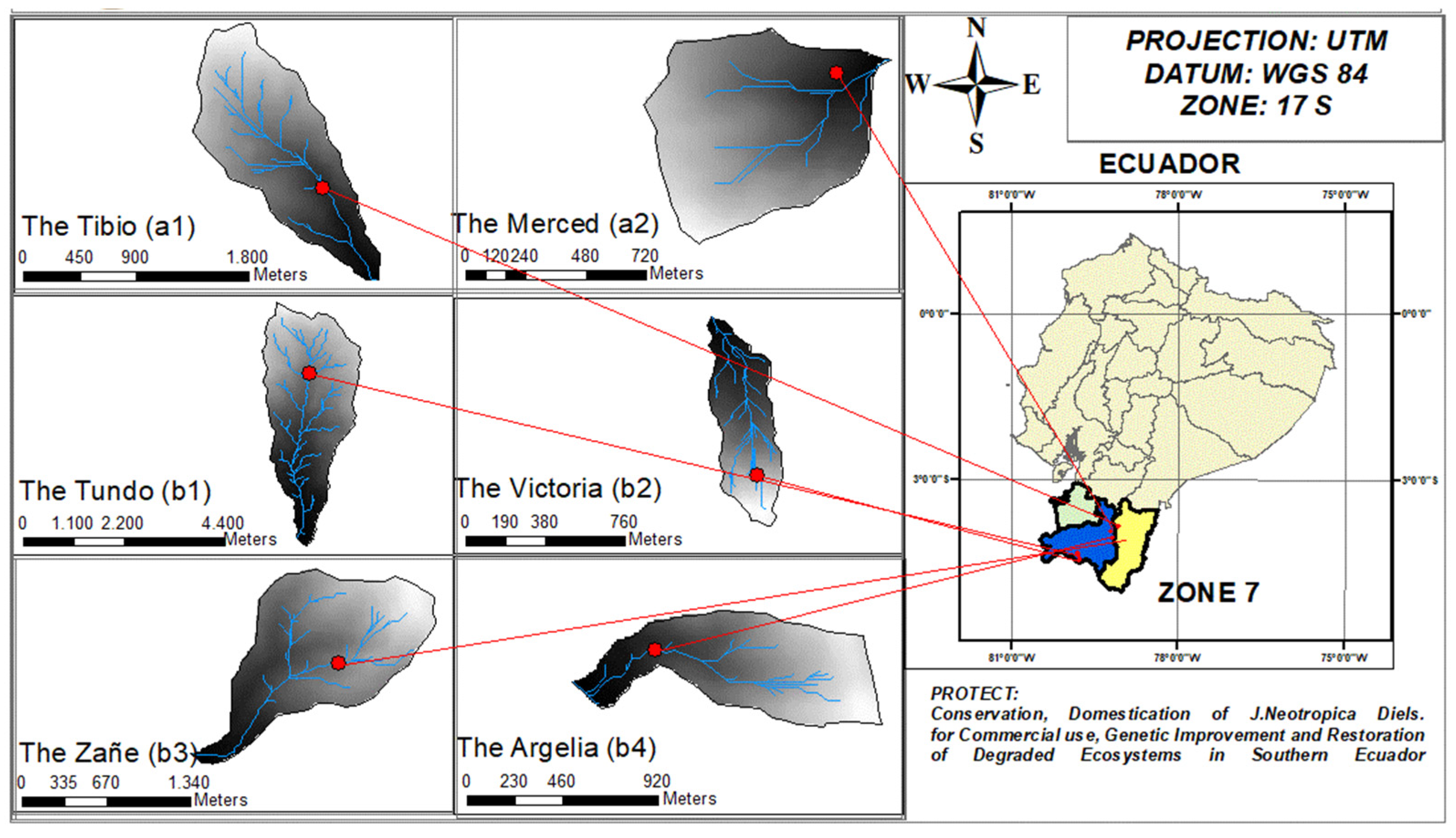
Figure 3.
Measurement of structural parameters in the different localities of southern Ecuador, The Tibio (a1), The Merced (a2); The Tundo (b1) The Victoria (b2), The Zañe (b3), The Argelia (b4).
Figure 3.
Measurement of structural parameters in the different localities of southern Ecuador, The Tibio (a1), The Merced (a2); The Tundo (b1) The Victoria (b2), The Zañe (b3), The Argelia (b4).


Figure 4.
Number of individuals per class diameter classes in different provenances of the study area at DBH(1,30cm). The Tibio (a1), The Merced (a2), The Tundo (b1), The Victoria (b2), The Zañe (b3), The Argelia (b4).
Figure 4.
Number of individuals per class diameter classes in different provenances of the study area at DBH(1,30cm). The Tibio (a1), The Merced (a2), The Tundo (b1), The Victoria (b2), The Zañe (b3), The Argelia (b4).
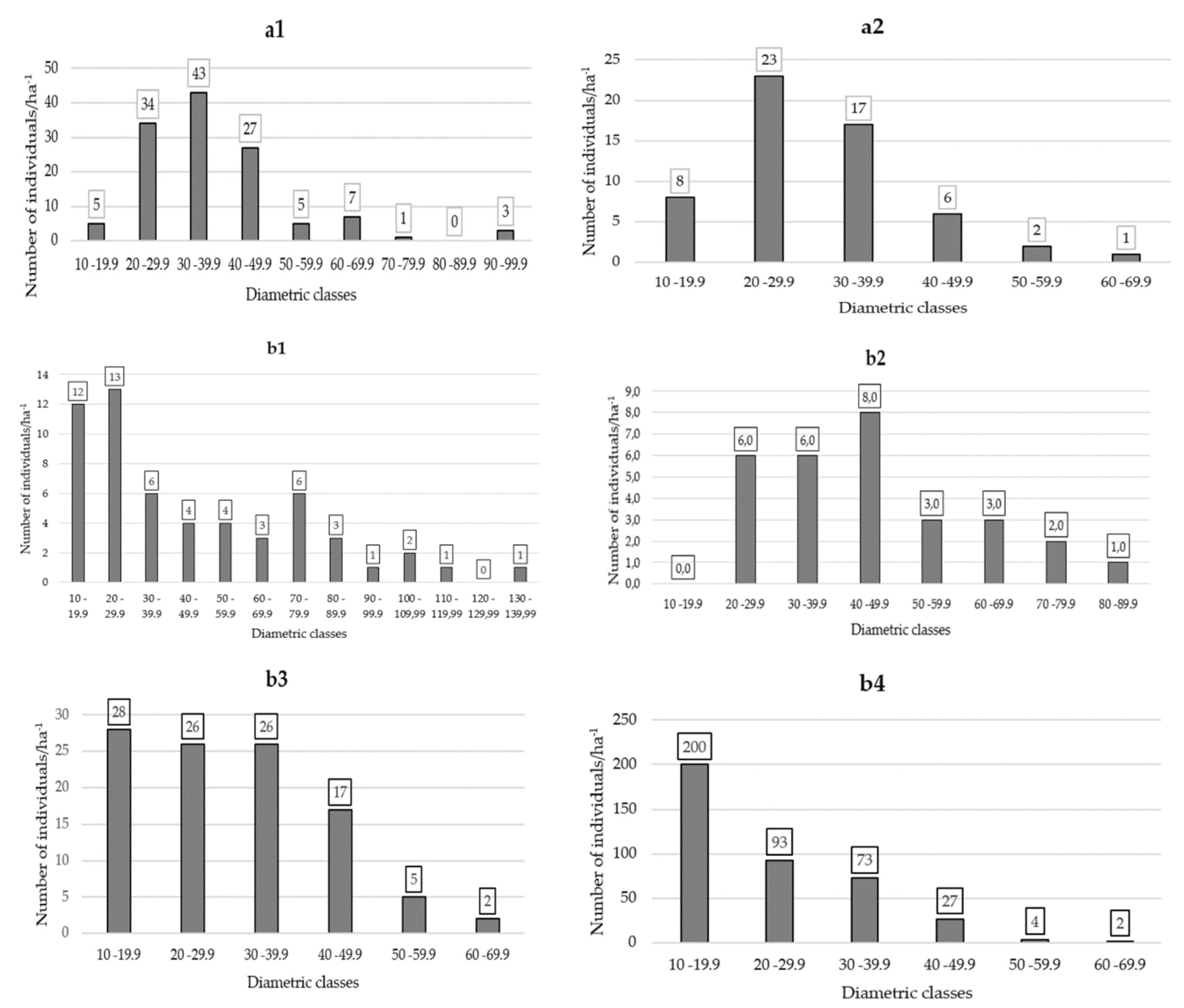
Figure 5.
Vertical structure in the different provenances of the study area The Tibio (a1), The Merced (a2), The Tundo (b1), The Victoria (b2), The Zañe (b3), The Argelia (b4).
Figure 5.
Vertical structure in the different provenances of the study area The Tibio (a1), The Merced (a2), The Tundo (b1), The Victoria (b2), The Zañe (b3), The Argelia (b4).
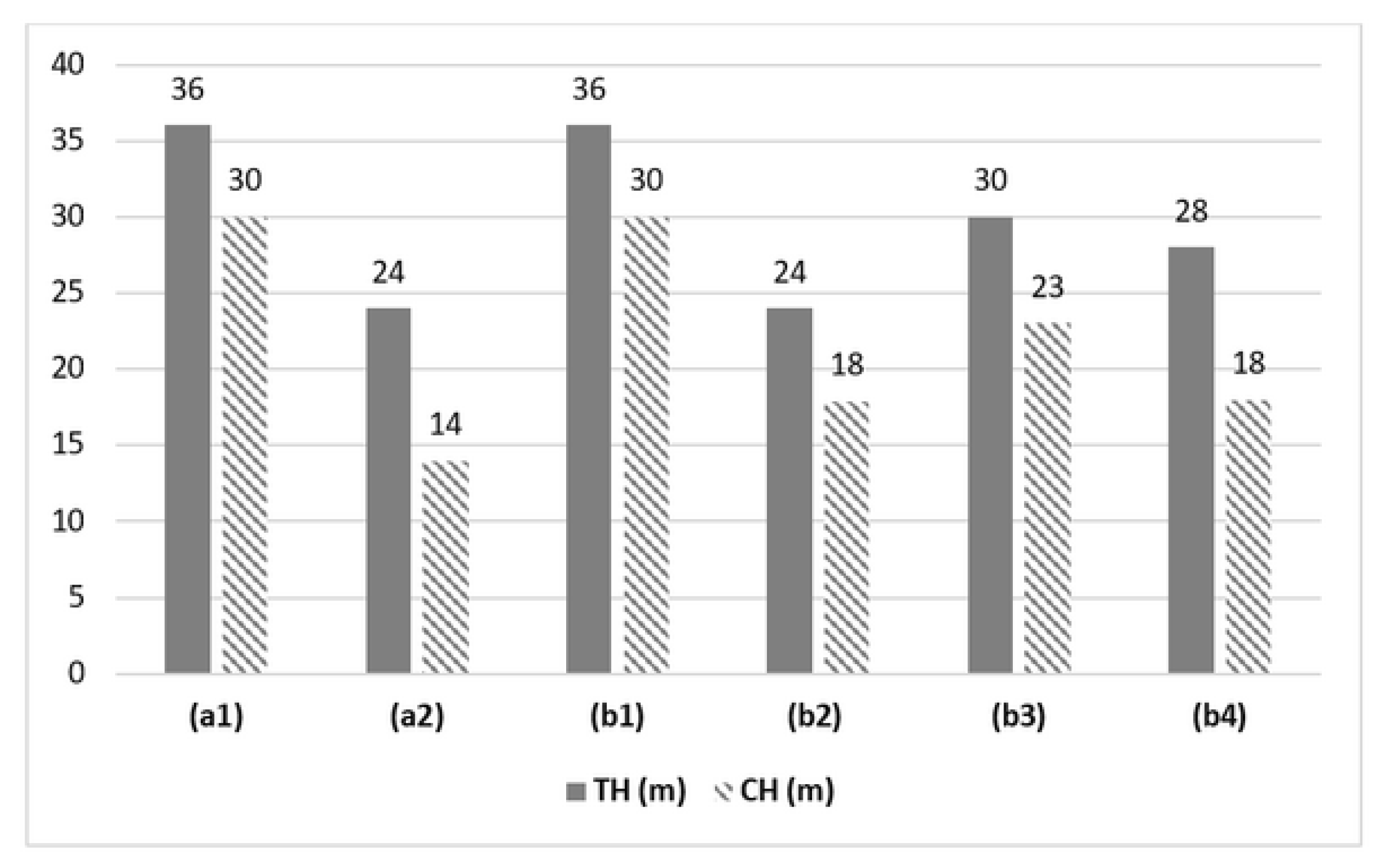
Figure 6.
Analysis of variance of dasometric variables in the different provenances of the study area The Tibio (a1), The Merced (a2), The Tundo (b1), The Victoria (b2), The Zañe (b3), The Argelia (b4).
Figure 6.
Analysis of variance of dasometric variables in the different provenances of the study area The Tibio (a1), The Merced (a2), The Tundo (b1), The Victoria (b2), The Zañe (b3), The Argelia (b4).
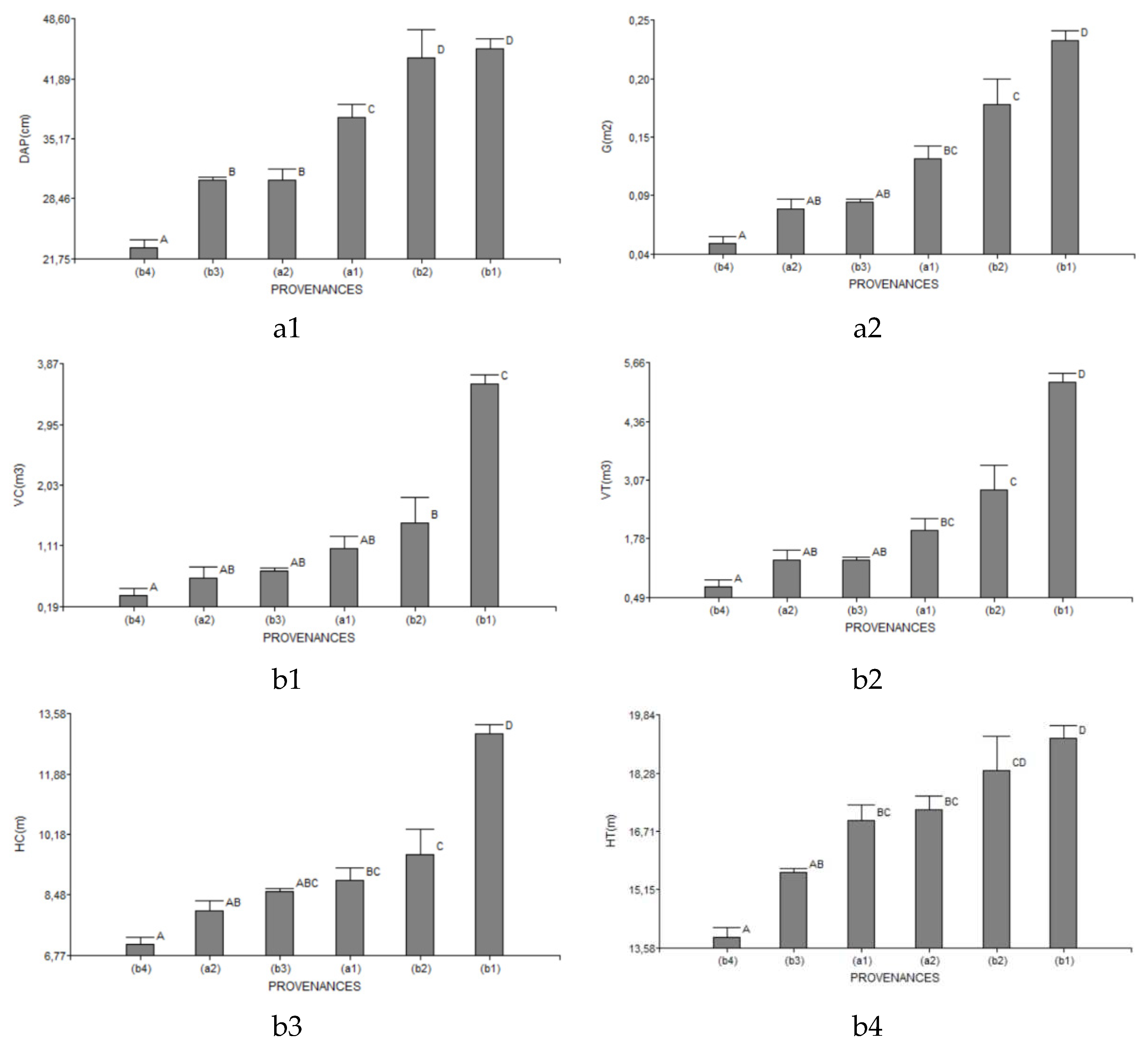
Figure 7.
Number of individuals of the species recorded per hectare in the different provenances of the study area The Tibio (a1), The Merced (a2), The Tundo (b1), The Victoria (b2), The Zañe (b3), The Argelia (b4).
Figure 7.
Number of individuals of the species recorded per hectare in the different provenances of the study area The Tibio (a1), The Merced (a2), The Tundo (b1), The Victoria (b2), The Zañe (b3), The Argelia (b4).
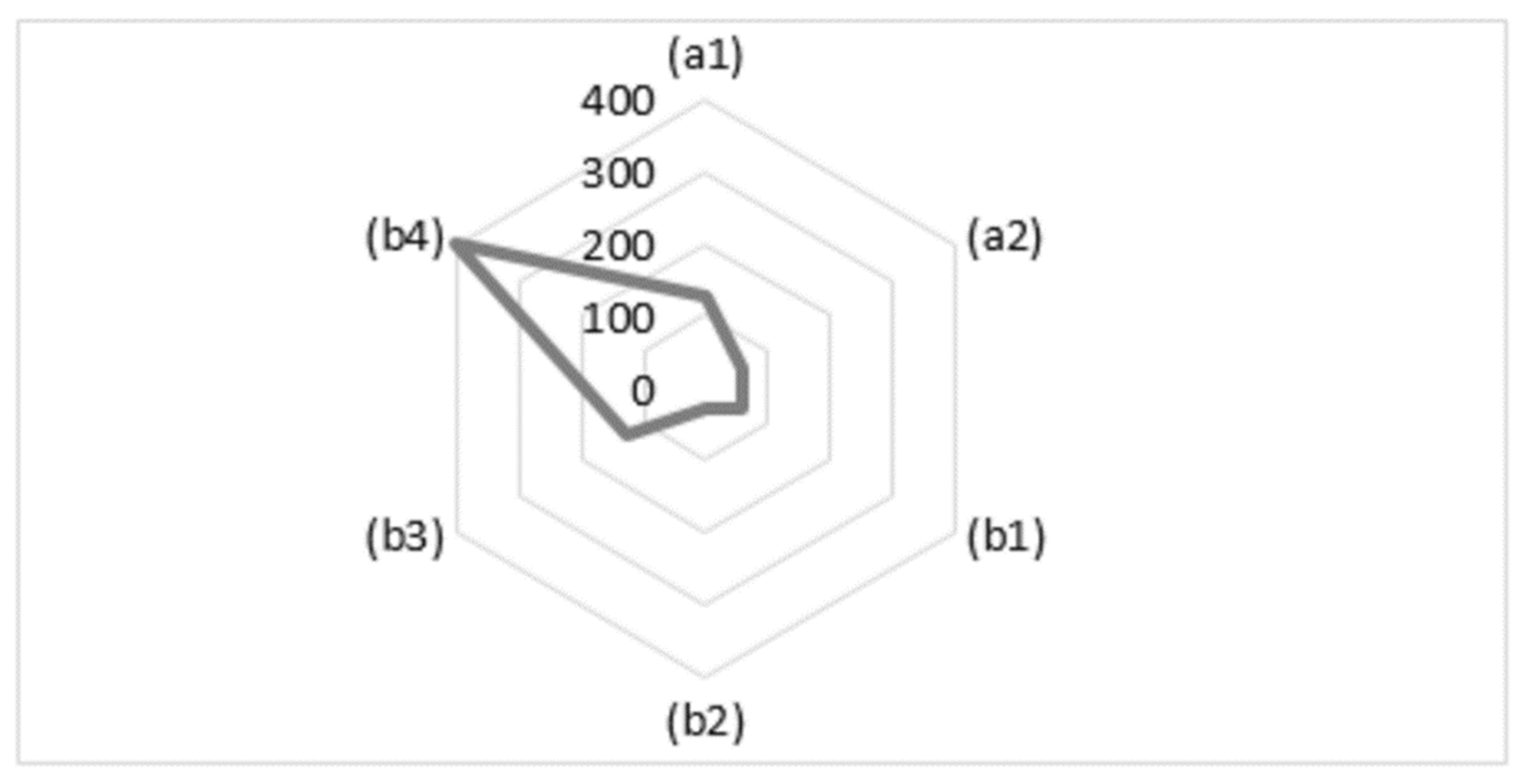
Table 2.
Occurrence of J. neotropica Diels growth sites in Southern Ecuador.
| PROVINCE | CANTON | FOREST SITE | AREA (ha) |
|---|---|---|---|
| Zamora chinchipe | Zamora | "El Tibio" (a1) | 0.8 |
| "La Merced" (a2) | 3 | ||
| Loja | Sozoranga | "El Tundo" (b1) | 120 |
| Macará | "La Victoria" (b2) | 0.9 | |
| Loja | "El Zañe" (b3) | 92 | |
| "La Argelia" (b4) | 0.7 | ||
| El Oro | No records | No records | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



