
Submitted:
25 July 2023
Posted:
27 July 2023
You are already at the latest version
Abstract
Persistent Organic Pollutants (POP) such as dichlorodimethyltrichloroethane (DDT) are present and ubiquitous in the environment due to their resilient nature. DDT is a prevalent endocrine disruptor still found in detectable amounts in organisms and the environment even after its use was banned in the 1970s. Studies show that exposure to DDT can cause adverse health effects in humans and animals, impairing fertility and increasing the risk of developing cancer. The aim of this work is to provide a comprehensive overview of the available literature on the effect of DDT and its metabolites on female reproductive tract, leading to infertility and cancer. Medline and Google Scholar were systematically searched to detect all relevant animal and human studies published in the last 20 years (January 2003 to February 2023) in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-analyses (PRISMA) statement. In total, 38 studies were included for qualitative synthesis. This systematic search and review indicated that exposure to DDT is associated with female reproductive health decline and that DDT acts as a sufficient carcinogen instigating reproductive cancers.
Keywords:
Dichlorodimethyltrichloroethane
; Endocrine Disruptors
; Female
; Infertility
; Neoplasms
; Persistent Organic Pollutants
; DDTs
1. Introduction
Dichlorodiphenyltrichloroethane (DDT) is an organochlorine pesticide (OCP), classified as a Persistent Organic Pollutant (POP) [1]. POPs are a group of organic compounds resistant to degradation with long half-lives. Their ability to bioaccumulate through different trophic levels and the potential to be transported over large distances by air or water render them immobilized in the ecosystem for long periods of time [2]DDT was initially used to treat insect-borne diseases such as malaria, and typhus in humans [3] and later as an agricultural pesticide in the 1940s and 1950s [4]. Due to its adverse environmental and toxicological effects and its persistence in the environment attributed to its stable structure, the use of DDT in agriculture was banned in the US in the 1970s, in Asia in the 1980s and 1990s, and worldwide in 2001 by the Stockholm Convention on POP. DDT is still used to control vector-borne diseases in South Africa, Uganda, Ethiopia, and Swaziland [5]. However, even after its widespread use was discontinued, it is still detected in toxic amounts in humans and animals. Exposure to DDT occurs through different routes such as inhalation, ingestion, and contact with the skin.
The primary route of DDT entry is consumption of contaminated meat, fish, and crops with high-fat content. It t can accumulate in breast milk fat, passing to infants through breastfeeding [6]. DDT can also cross placental barrier [7]. In-utero exposure to DDT is linked with reduced birth weight and length, reduced head circumference, preterm birth, and fetal loss [8]. Lactational transfer is significantly higher than placental transfer. Effects of DDT include toxicity of the liver and central nervous system, reproductive cancers, and estrogenic, antiandrogenic, and epigenetic effects [9,10]. Exposure to DDT can also have transgenerational effects such as obesity, early onset of puberty, insulin resistance, testis, ovary, and kidney pathologies in offspring of exposed organisms [11,12]. Ancestral DDT exposure significantly increased the incidence of polycystic ovaries and the development of uterine infections in future generations in rats [13]. Fetal, neonatal, or pubertal exposure cause impaired reproductive function. DDT and its metabolites can cause a widespread reduction in population size by accumulating in the eggs of birds and reptiles affecting the eggshell thickness and consequently decreasing reproductive success [14]. It reduces fertility in mice causing a decline in sperm quality and quantity, rendering them sterile after exposure [15]. DDT and its metabolites are endocrine-disrupting chemicals (EDC) that mimic the action of estrogen disturbing the estrogen receptor (ER) pathways [16]. It also displays anti-androgenic effects in combination with the androgen receptor (AR). DDT induces a disrupter effect in target cells which involves competing with testosterone to bind to AR causing receptor signaling impairment. It also increases the estrogen synthesis, implicating hormone production and alterations that increase the probability of somatic and reproductive disorders in later life [16]. An imbalance in the production of testosterone and estrogen due to excessive estrogen production is actively associated with an increased risk of feminization. It can also trigger estrogen-related cancers and cardiovascular disorders [17,18].
2. Methodology
Medline (Ovid Medline Epub Ahead of Print, In-Process & Other Non-Indexed Citations, Ovid MEDLINE(R) Daily, and Ovid MEDLINE(R)) and Google Scholar were systematically searched to detect all relevant animal and human studies published during the last 20 years (January, 2003 to February, 2023) in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-analyses (PRISMA) statement [19] using the following search terms “DDT”, “DDT and Female infertility” and “DDT and female reproductive tract cancers”. We primarily focused on research articles containing epidemiological information on human and animal data, including mechanistic and molecular information. Reference lists of selected studies were screened for other potentially eligible studies. After excluding duplicates, citations in abstract form, and non-English citations, the titles/abstracts of full papers were screened for relevance, defined as original research focusing on the topic “Effects of DDT AND Female infertility AND Female reproductive tract cancers”. Two review authors (S.S. and S.Q.) independently scanned the title and the abstract content, or both, of every record retrieved to determine which studies should be assessed further evaluated and extracted all data. Disagreements were resolved through consensus or by consultation with a third author (S.A.). A final draft of the manuscript was prepared after several revisions and approved by all authors.
3. Results and Discussion
In this systematic search and review, a total of 38 studies were finally included for qualitative synthesis (20-57). Figure 1 provides a visualization of the review process.
3.1. Role of DDT in female infertility
3.1.1. In-vitro studies
Experimental studies investigating the role of DDT and its metabolites on female infertility include in-vitro assessments on follicular [20], granulosa, endometrial [21] , chorion [22], and placental cell cultures [23]. Six of the 12 in-vitro assays included in this systematic search and review, have been conducted on cows [21,22,23,24,25,26], two on porcine [20,27] , one on hamsters [28], two on Sprague-Dawley (SD) rats [29,30], and one on Danio rerio [31]. DDT and its metabolites impair fertility via hormone interference, cell cytotoxicity, and inhibition of enzymatic activity disrupting the ion balance, membrane permeability, uterine contractility, and ovarian steroidogenesis. This ultimately increases the risks of spontaneous abortions, miscarriages, delayed puberty, and reduced fecundability. Traces of DDT and its metabolites particularly p,p΄-DDE has been detected in the ovarian follicular cells of cattle, porcine, sheep, and pigs with the highest concentrations found in cows. The follicular cell membrane is permeable to low and high-molecular-weight compounds. DDT and its metabolites tend to alter the structure and function of the follicular cell membrane and eventually the oocyte membrane, which can have detrimental impacts on oocyte maturation, development of granulosa cells, and hormone secretion which leads to impaired fertility over time.
The mechanisms via which DDT and its metabolites cause fluctuations in the normal processes and maintain the balance of the reproductive system and associated hormones are complex and vary among different studies. Estradiol (E2) is produced in the ovaries and is responsible for fertility in non-pregnant women of childbearing age. DDT has both estrogenic and anti-estrogenic properties in a dose-and-exposure-dependent manner. At lower doses ο,p΄-DDT, ο,p΄-DDE, and ο,p΄-DDD decrease E2 secretion indicating their anti-estrogenic nature while at higher doses, they exhibit estrogenic characteristics. On the other hand, p,p΄-DDT and p,p΄-DDE are estrogenic independent of dosages but their repeated exposure decreases E2 levels [20]. To confirm the disruptive action of DDT, authors later studied its impact on ovarian steroidogenesis. They established DDT as an Estrogen Receptor Beta (Erβ) agonist and antagonist corroborating their earlier findings that DDT is both estrogenic and anti-estrogenic [31]. However, no sole mechanism can be considered a standard pathway via which DDT decreases E2 secretion. DDT has anti-androgenic properties particularly p,p΄-DDE [31] thus, it reduces testosterone levels, the prime substrate for Cytochrome P450 Enzymes. These P450 enzymes are responsible for the oxidation of xenobiotics such as DDT and converting androgens into estrogens. Thus, DDT either reduces E2 synthesis by antagonistic action the on Erβ receptor, reducing the testosterone levels which leads to less conversion of androgens into estrogens and inhibits the P450 activity via lack of substrate [27] which can be visualized in Figure 2.
Lyche et al. [31] reported delayed puberty, skewed sex ratio, and feminization during pubertal development as possible implications of androgen inhibition by DDT and its metabolites. DDT and DDE increase Oxytocin (OT) secretion from granulosa and luteal cells [21]. This finding is further supported by Mlynarczuk et al. [24] through transcriptome analysis which displayed increased mRNA expression for Neurophysin-1 (NP-I)/Oxytocin (OT) in pregnant cows particularly for 9-12 weeks. During the same pregnancy duration, the mRNA expression for Prostaglandin A (PGA) is also observed to decrease in luteal cells thus, indicating the impact of DDT and its metabolites on prostaglandin synthesis. In mammals, the Prostaglandin F2 alpha (PGF2α) and Prostaglandin E2 (PGE2) are important for the establishment of early pregnancy and for the course of the estrous cycle. Any imbalances in their ratios can have serious implications, particularly on fertilization and embryo implantation. Wrobel et al. [21] indicated that DDT and DDE influence the endometrial cells to secrete more PGF2a and less PGE2. This PGF2a: PGE2 ratio can increase myometrial contractions and accelerate luteal regression, which may alter progesterone (P4) levels responsible for pregnancy maintenance. A positive feedback loop exists between PGF2a and OT secretion during luteal regression. Thus, the increase in PGF2a is responsible for accelerated luteal regression and OT secretion. While performing a study on chorion explants obtained from cows Mlynarczuk et al. [22] reported similar results by indicating that DDT impairs the secretion of PGE2, PGF2α OT, and P4 from the smooth chorion. A study conducted on pregnant cows indicated that only DDE increases P4 levels. However, the ratio of P4 to OT decreases in response to DDT and DDE. In the case of pregnant cows, this alteration in secretion patterns can induce strong myometrial contractions of the uterine strips [24]. This disturbs the regulation of processes in the cattle placenta and increases the risk of abortions and preterm/premature births [22] which has also been proven by multiple epidemiologic studies on human female subjects. Kwekwl et al. [29] also reported uterine motility dysregulation in mice and rats by ο,p΄-DDT via specie-specific uterine hypertrophy. Myometrial hypertrophy was more evident in mice which increased their Uterine Wet Weight (UWW) while stromal hypertrophy was observed in rats. Stromal Edema was, however, common in both species. In addition, Luminal Epithelial Height (LEH), a characteristic market for estrogen exposure in the uterus is also increased under the influence of ο,p΄-DDT, indicating that the chemical is somewhat estrogenic in nature. A comprehensive depiction of these interlinked processes is shown in Figure 3 and Figure 4.
On the contrary, Salleh et al. [30] showed that exposure to DDT reduces uterine contractions in rats. He proposed two mechanisms via which DDT impacts uterine motility and contractions; through inhibition of uterotonic pathways such as PGF2a and OT secretion which has previously been mentioned as well or through alteration in the Ca2+ influx and intracellular release as shown in Figure 5 that ultimately reduces the uterine contractions. These compounds may also stimulate P4 secretion independently of E2 secretion from the chorion, which increases the ratio of P4 to E2 [25]. The levels of E2 and P4 maintain the placental barrier and maternal-fetal connections by regulation of Connexin (Cx) genes; Cx26 and Cx43, the expressions of which are observed to decrease and increase under the influence of DDT, respectively. DDE exposure increased Cx32 and Cx43 expression in the placental site [23]. These chemical and hormonal changes are speculated to impair the placental barrier function, disrupt trophoblast invasion, and impact the secretory activity of the placenta in cows [25]. Uterine Contractions are important in transporting sperm, ovum, and placental and fetal expulsion at birth. Any dysregulation might lead to various adverse effects on fertility and reproduction.
p,p΄-DDT decreases the activity of human chorionic gonadotropin/Luteinizing Hormone Receptor (hCG/LHR) in a dose-dependent manner via negative allosteric modulation of the beta-arrestin 2 (B-arrestin 2) and cyclic 3΄,5΄-monophosphate (cAMP) pathways which alter the levels of Human Chorionic Gonadotropin (hCG), which is required to support pregnancy and Luteinizing Hormone (LH) which is crucial to uterus preparation for pregnancy leading to an increased risk of spontaneous abortions and miscarriages [28] (Figure 6).
The viability of the cells was unaffected by the doses of DDT and DDE used by Wrobel et al. [21] in their study. Thus, these alterations in the secretions of PGF2a, PGE2, and OT by endometrial and granulosa cells cannot be attributed to their cytotoxic effects. Instead, it could be that DDT and DDE impair prostaglandin synthesis directly by impacting the synthesis of a common precursor or specific synthases, or both or indirectly by stimulating the Leukemia Inhibitor Factor (LIF) synthesis in the myometrium. LIF regulates decidualization, and embryo-endometrial interaction, and makes the uterus receptive to implantation [26]. The dysregulation of hormone levels in response to DDT and its metabolites is summarized in Table 1 [20,21,22,24,25,27,28,31]
3.1.2. Epidemiological studies
The epidemiological studies included in the systematic search and review consist of six case-control studies [32,33,34,35,36,37], seven cohort studies [38,39,40,41,42,43,44], two cross-sectional studies [45,46], and three pilot studies [47,48,49] which have been conducted on women from different geographical areas with different ethnic backgrounds. The ethnicities of the women involved in the studies include American, Mexican, Indian, Chinese, Bolivian, Latina, German, and Laotian. The results of this systematic search and review reveal that out of 18 epidemiological studies included, 13 (72.2%) showed a negative association between DDT and female infertility, and 1 (5.5%) showed a positive association. In contrast, 4 (22.2%) of them showed no association. This suggests that many of the studies included show consistent results. DDT and its metabolites exert adverse effects on female fertility and reproductive health through various mechanisms of action, either by their activity as endocrine disruptors, anti-androgens, and xenoestrogens or through interference in the binding of progesterone to its receptor thereby disrupting the closure of sodium ion channels in membranes as well as causing oxidative stress by the formation of reactive oxygen species (ROS). These detrimental effects of exposure to DDT and its metabolites can cause infertility, reduced fecundability, preterm birth (PTB), fetal loss, shorter time-to-pregnancy (TTP), decreased length of gestation (LOG), and reduced menstrual cycle length. An overview of how DDT and its metabolites have impacted women belonging to different ethnicities is presented in Table 2 [32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49]
3.1.2.1. DDT and menstrual cycle length
It has been reported by several studies that DDT can reduce the length of the menstrual cycle in young women. Ouyang et al. [46] studied the effects of DDT exposure on age at first menstruation and menstrual cycle length. It was observed that a 10ng/g increase in serum concentration of DDT reduced the age at which the first menstruation was experienced. The cause of this effect can be attributed to the activity of p,p΄-DDE as an endocrine disrupter which plays a role in increasing the amount of estradiol in the body by converting androgens to estrogens which in turn accelerates the maturation of oocytes causing early menarche. p,p΄-DDE can increase the activity of human granulosa cells in combination with FSH. Therefore, it can increase the amount of estradiol in the body due to the action of granulosa cells influenced by FSH that can aromatize androgens to estrogens. This can also lead to an upset in the hormonal balance implicating changes in the menstrual cycle. These findings can also be further confirmed by another study by Windham et al. [49] who researched the effects of exposure to organochlorides and their ramifications on female ovarian function. Women with a higher serum concentration of DDE had a reduction in their menstrual cycle length as well as their luteal phase length and their progesterone metabolite levels. DDT has been reported to interact with the progesterone-response pathway in various ways including binding to the progesterone receptor, inhibiting progesterone-induced enzymes, and decreasing the frequency of implantation of eggs thereby causing sterility and decreased serum progesterone levels as observed in rats exposed to high levels of DDT. In this way it can also alter the luteal phase length, making it shorter due to decreased progesterone production. Although these studies prove the negative impact DDT has on menstrual cycles Chen et al. [47] found that neither p,p΄-DDT nor o,p΄-DDT had any connection with menstrual cycle length, duration of menses or heaviness of menstrual flow in a pilot study they conducted on 60 Chinese women. This confirms that more investigation is required to thoroughly understand the mechanism behind the influence DDT has on the menstrual cycle.
3.1.2.2. DDT and infertility/fecundability
Studies in the scientific literature have provided evidence that DDT at low levels of exposure can cause adverse reproductive outcomes in humans and animals. However, many of the studies analyzing the relationship between DDT and infertility focus on males, while female reports are meager. According to the presently available studies on DDT and its impact on female reproductive health, DDT has an inverse relationship with female fertility. Cohn et al. [34] assessed the fecundability ratios for daughters exposed to maternal DDT and its metabolites in utero to monitor any induced transgenerational effects. According to the results p,p΄-DDT decreased the daughters’ conception probability by 32% for every 10μg/L p,p΄-DDT present in maternal serum. Surprisingly, the opposite effect was observed with p,p΄-DDE as it increased the conception probability of daughters by 16% for every 10μg/L p,p΄-DDE in the maternal serum. The antiandrogenic effects of p,p΄-DDT induce a detrimental impact on the ovaries during gestation and early life. However, the mechanism behind the positive effects of p,p΄-DDE on conception is not well understood. In a similar study Weiss et al. [48] found that German women with high concentrations of total serum DDT suffered from a lower pregnancy rate attributed to DDT’s xenoestrogenic behavior. Perry et al. [43] studied serum DDT levels and progesterone and estrogen levels across the menstrual cycle in women of reproductive age. The findings stated that DDT could cause a reduction in estrogen levels during ovulation and progesterone levels when it is required as an indicator of corpus luteal function essential for early pregnancy maintenance. Furthermore, DDT-PdG and E1C associations can harm female reproductive health, such as impaired fertility and early pregnancy loss. DDT affects progesterone and estrogen metabolite profiles by multiple mechanisms such as p,p΄-DDE binding to the progesterone receptor or o,p΄-DDT, and o,p΄-DDE binding to the estrogen receptor, inhibiting the binding of endogenous estradiol. Kezios et al. [40] concluded that p,p΄-DDE was negatively associated with the gestation length. It was also observed that o,p΄-DDT and p,p΄-DDE were linked with decreased birth weight while it was the opposite for p,p΄-DDT. This could be because they operate through different biological pathways leading to contrasting effects. While p,p΄-DDE acts as an anti-androgen, o,p΄-DDT is weakly estrogenic causing the influence change. Arrebola et al. [38] targeted the relationship between maternal o,p΄-DDT and p,p΄-DDE serum levels with birth outcomes. It was observed that p,p΄-DDE was positively associated with birth weight while it was the opposite for o,p΄-DDT. Ouyang et al. [42] reported that the women who were deficient in vitamin B2 but had high DDT serum concentrations were more susceptible to reduced clinical pregnancy (CP) and early pregnancy loss (EPL). In cases where Vit B was sufficient, DDT was not linked with CP, suggesting Vit B may help protect women against the adverse reproductive effects of DDT exposure. Although most studies found a negative association between DDT exposure and female fertility Harley et al. [45] determined that there was no association between p,p΄-DDT, o,p΄-DDT, or p,p΄-DDE and TTP in a cross-sectional study performed on 402 Latina women. This indicates insufficient research regarding women’s reproductive health and that this subject needs to be studied further.
3.1.2.3. DDT and preterm birth
DDT has been associated with preterm birth (PTB) and reduced LOG and TTP. Torres-Arreola et al. [36] conducted a case-control study to find the association between maternal serum p,p΄-DDE levels, and preterm birth. It was observed that p,p΄-DDE increased the risk of preterm birth by twofold at low levels (OR=1.87, 95% CI=0.95-3.68 for 111.6-228.8 ng/g and OR=1.67, 95% CI=0.84-3.31 for >228.8 ng/g). It was observed that p,p΄-DDE exhibited a dose-response effect on preterm birth at levels exceeding 21μg/L. The exact process explaining the correlation between p,p΄-DDE, and preterm birth is not yet determined. However, the antiandrogenic behavior of p,p΄-DDE and its interference in the binding of progesterone to its receptor could be attributed to its effect on pregnancy duration consequently causing preterm birth. Tyagi et al. [37] studied the accumulation of POP in pregnant women and the placenta to examine their exposure’s possible effects on PTB and LOG. Women with substantial levels of DDT made up 8.8% of PTB cases contrasting 2.7% for mothers with no detectable levels of DDT in their blood. A significant amount of DDT was also found in the placental tissues of PTB cases. A negative correlation was also found between LOG and DDE levels in maternal blood and high levels of DDT in breast milk suggesting it has a role in early birth. Anand et al. [32] analyzed the residues of OCP in the placenta of females in preterm and full-term deliveries to evaluate the levels of different oxidative stress markers and to correlate them with OCP levels. The investigation revealed that oxidative stress caused by OCP residues plays a significant role in preterm deliveries. It was hypothesized that oxidative stress in trophoblastic placental tissue caused by the generation of reactive oxygen species (ROS) resulted in adverse reproductive outcomes. However, it could also be plausible to assume that the xenoestrogenic nature attributed to DDT alters the natural hormonal balance between estrogen and progesterone responsible for maintaining pregnancy, causing harmful pregnancy outcomes. Anand et al. [33] conducted another study on 90 Indian women and observed that women exposed to OCP were 1.7 times more likely to deliver a preterm babies than pregnant women who were not exposed to them. Around 100% of pregnant women had detectable levels of p,p΄-DDE in their placenta and umbilical cord in PTB cases, confirming that OCP compounds are responsible for reproductive toxicity. Farhang et al. [39] examined 20,754 women in a longitudinal cohort study to study the effects of maternal serum DDT and DDE concentrations and their influence on male infants’ PTB, SGA, and birth weight. Results showed that no statistically significant relationship existed between the serum measurements of DDT or DDE and birth weight, length of gestation, and SGA. However, these results could not be generalized to female infants.
3.1.2.4. DDT and fetal loss
The endocrine disruptive behavior of DDT allows it to inhibit the deactivation and inactivation of the sodium channels in the membrane of tissues such as the placenta causing prolonged currents that keep the channel from closing. Longnecker et al. [41] verified this in their study aimed to confirm the relationship between maternal serum DDE levels and fetal loss in previous pregnancies. The results revealed that increasing maternal serum DDE levels were positively associated with fetal loss while DDT showed no interconnection. The odds ratio was 1.4 (95% CI 1.1-1.6) for a 60μg/L increase in serum level. The mechanism of action was postulated to be acute DDT toxicity induced by obstruction of closure of sodium channels in the placenta. The antiandrogenic behavior of DDE, blocking progesterone from binding to its receptor, was also considered a valid mechanism of action in causing fetal loss. Venners et al. [44] carried out a similar study purposed of finding how preconception serum total DDT had an impact on pregnancy loss. Among the DDT metabolites, the most abundant in concentration was p,p΄-DDE which made up around 92% while p,p΄-DDT accounted for 6% of the total mass of DDT metabolites. It was estimated that every 10ng/g (OR=1.19, 95% CI 1.04, 1.36) increase in serum total DDT was linked to the relative odds of total and early pregnancy losses. Results confirmed the positive linear correlation of serum total DDT concentration and early pregnancy losses. Although the causal pathway could not be identified, it was hypothesized that the disruption in the closure of sodium channels caused by DDT and its tendency to prevent progesterone from binding to its receptor could be relevant to the cause of early fetal loss. Mahalingaiah et al. [35] demonstrated the link between DDT and DDE serum levels with implantation failure, chemical pregnancy, and spontaneous abortion in women undergoing in vitro fertilization procedures. The study concluded that no statistically significant associations existed between serum and follicular concentrations of DDT/DDE and oocyte number, quality, fertilization, or pregnancy rates. It was determined that the likelihood of implantation failure was increased due to the suppression of luteal progesterone leading to the incomplete maturation of the endometrial lining or of early pregnancy loss from significantly low levels of progesterone production. The effects of exposure to DDT on female infertility and its mode of action are not entirely understood with the current information available. Further investigation and studies are required to determine the full extent of the potentially harmful impacts of exposure to DDT on females’ reproductive health.
3.2. Role of DDT in female reproductive cancers
Cervical, ovarian, and uterine cancers are prevalent cancers among women from all regions and DDT has been found in their samples. However, the past 20 years, not much has been studied about the impact of DDT on female reproductive tract cancers. The eight studies regarding the role of DDT in female reproductive cancers included in this review mainly highlight the presence of DDT and its metabolites in female reproductive cancer patients [50,51,52,53,54,55,56,57]. Most of the data has been reported from India where DDT is still used in agriculture and thus crops such as wheat have higher pesticide content. However, cases with higher OCP levels have been reported from Mexico, and North America which renders the above statement a mere speculation. Earlier studies focused more on understanding the potential mechanisms through which DDT directly or indirectly plays a part in the progression of relevant cancers. Later on, the focus of research shifted towards epidemiology.
Ndebele et al. [54] conducted a study on Human Cervical Cancer Cells (HeLa) to understand the mechanism through which xenoestrogens such as DDT modulate cancer biology. An increase in Cyclin A and D Proteins levels was observed to variable degrees, indicating that DDT induces changes in B-cell lymphoma 2 (BCL-2), the protein that regulates cell death. It also suppressed HeLa cell proliferation in a concentration-dependent manner and thus, has cytotoxic effects on cells. Earlier findings also reported the influences of DDT on the development and growth of leiomyoma by affecting angiogenesis and apoptosis [50]. According to Mathur et al. [53], though women aged 41-50 had a higher risk of developing reproductive tract cancers, higher DDT residues were more prevalent in women aged 21-30 in Jaipur, India. Gibson and Saunders [51] worked on 524 Ishikawa cell lines established from endometrial adenocarcinoma cells of a 39-year old woman 525 and observed that specifically o,p΄-DDT had 100-fold more estrogenicity in reproductive tissues 526 than p,p΄-DDT. A case-control study conducted in North India also reported significant results regarding the presence of DDT and DDE at higher levels in the blood among epithelial ovarian cancer patients [57]. Kalinina et al. [52] treated normal human endometrial cells with DDT and observed upregulation of miRNA 190b. A similar pattern was reported in the uterus of rats indicating an increase in the expression of oncogene Tumor protein p53-inducible nuclear protein 1 (Tp53inp1) which increases the chances of uterine cancers. The blood and cervical tissue samples from cervical cancer patients were studied to detect the levels of OCP among women of East Delhi. DDT was significantly higher in the blood samples of cases while no correlation was observed between levels of DDT in cervical tissue samples and blood samples. The higher pesticide levels in the blood were also attributed to the region’s higher DDT content in wheat [55]. A similar study conducted in Mexico also showed higher DDE and DDD levels in blood samples. Women from Tizimin, Progresso, and Kanasin had high DDD, DDT, DDE, and DDD levels, respectively [56].
4. Conclusion
This systematic search and review indicated that exposure to DDT is associated with female reproductive health decline and that DDT acts as a sufficient carcinogen instigating reproductive cancers.
Author Contributions
Conceptualization, S.A.; investigation S.S., S.Q., N.K., and S.; analysis and writing- original draft preparation, S.S. and S.Q.; methodology, S.S., S.Q., and M.E., visualization S.S., supervision, S.A. and S.T.A.S.; review and editing, C.M., H.A., H.E., A.T., S.S., and S.Q. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
No conflicts of interest.
Abbreviations
| AR | Androgen Receptor |
| cAMP | Cyclic Adenosine 3΄, 5΄-monophosphate |
| CP | Clinical Pregnancy |
| Cx | Connexin |
| DDD | Dichlorodiphenyldichloroethane |
| DDE | Dichlorodiphenyldichloroethylene |
| DDT | Dichlorodiphenyltrichloroethane |
| E2 | Estradiol |
| EDC | Endocrine-disrupting Chemicals |
| EPL | Early Pregnancy Loss |
| ER | Estrogen Receptor |
| Erβ | Estrogen Receptor Beta |
| FSH | Follicle Stimulating Hormone |
| hCG | Human Chorionic Gonadotropin |
| hCG/LHR | Human Chorionic Gonadotropin/Luteinizing Hormone Receptor |
| LEH | Luminal Epithelial Height |
| LH | Luteinizing Hormone |
| LIF | Leukemia Inhibitor Factor |
| NP-I | Neurophysin-1 |
| ο,p΄-DDD | ortho, para΄-Dichlorodiphenyldichloroethane |
| ο,p΄-DDE | ortho, para΄-Dichlorodiphenyldichloroethylene |
| ο,p΄-DDT | ortho, para΄-Dichlorodiphenyltrichloroethane |
| OCP | Organochlorine Pesticide |
| OCP | Organochlorine Pesticide |
| OT | Oxytocin |
| p,p΄-DDE | para, para΄-Dichlorodiphenyldichloroethylene |
| p,p΄- DDT | para, para΄-Dichlorodiphenyltrichloroethane |
| p,p΄-DDD | para, para΄-Dichlorodiphenyldichloroethane |
| P4 | Progesterone |
| PGA | Prostaglandin A |
| PGE2 | Prostaglandin E2 |
| PGF2α | Prostaglandin F2 alpha |
| LOG | Length of gestation |
| POP | Persistent Organic Pollutant |
| PRISMA | Preferred Reporting Items for Systematic Reviews and Meta-Analysis |
| PTB | Preterm Birth |
| ROS | Reactive Oxygen Species |
| SGA | Small for Gestational Age |
| TTP | Time to Pregnancy |
| UWW | Uterine Wet Weight |
References
- Burgos-Aceves, M.A.; Migliaccio, V.; Di Gregorio, I.; Paolella, G.; Lepretti, M.; Faggio, C.; Lionetti, L.J.E.T.; Pharmacology. 1, 1, 1-trichloro-2, 2-bis (p-chlorophenyl)-ethane (DDT) and 1, 1-Dichloro-2, 2-bis (p, p’-chlorophenyl) ethylene (DDE) as endocrine disruptors in human and wildlife: A possible implication of mitochondria. Environmental Toxicology and Pharmacology 2021, 87, 103684. [Google Scholar] [CrossRef]
- Nadal, M.; Marquès, M.; Mari, M.; Domingo, J.L. Climate change and environmental concentrations of POPs: A review. Environ. Res. 2015, 143, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Mariano, J. .; Baltazar-Reyes, M.C.; Salazar-Martínez, E.; Cupul-Uicab, L.A. Exposure to the pesticide DDT and risk of diabetes and hypertension: Systematic review and meta-analysis of prospective studies. Int. J. Hyg. Environ. Heal. 2021, 239, 113865. [Google Scholar] [CrossRef] [PubMed]
- Russo, F.; Ceci, A.; Pinzari, F.; Siciliano, A.; Guida, M.; Malusà, E.; Tartanus, M.; Miszczak, A.; Maggi, O.; Persiani, A.M. Bioremediation of Dichlorodiphenyltrichloroethane (DDT)-Contaminated Agricultural Soils: Potential of Two Autochthonous Saprotrophic Fungal Strains. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef]
- Binelli, A.; Provini, A. DDT is still a problem in developed countries: the heavy pollution of Lake Maggiore. Chemosphere 2003, 52, 717–723. [Google Scholar] [CrossRef]
- Bouwman, H.; Kylin, H.; Sereda, B.; Bornman, R. High levels of DDT in breast milk: Intake, risk, lactation duration, and involvement of gender. Environ. Pollut. 2012, 170, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Nicolella, H.D.; de Assis, S.J.I.J.o.M.S. Epigenetic inheritance: Intergenerational effects of pesticides and other endocrine disruptors on cancer development. International Journal of Molecular Sciences 2022, 23, 4671. [Google Scholar] [CrossRef]
- Fucic, A.; Duca, R.C.; Galea, K.S.; Maric, T.; Garcia, K.; Bloom, M.S.; Andersen, H.R.; Vena, J.E. Reproductive Health Risks Associated with Occupational and Environmental Exposure to Pesticides. Int. J. Environ. Res. Public Heal. 2021, 18, 6576. [Google Scholar] [CrossRef]
- Qi, S.-Y.; Xu, X.-L.; Ma, W.-Z.; Deng, S.-L.; Lian, Z.-X.; Yu, K. Effects of Organochlorine Pesticide Residues in Maternal Body on Infants. Front. Endocrinol. 2022, 13, 890307. [Google Scholar] [CrossRef]
- Sifakis, S.; Androutsopoulos, V.P.; Tsatsakis, A.M.; Spandidos, D.A. Human exposure to endocrine disrupting chemicals: effects on the male and female reproductive systems. Environ. Toxicol. Pharmacol. 2017, 51, 56–70. [Google Scholar] [CrossRef]
- E Nilsson, E.; Ben Maamar, M.; Skinner, M.K. Role of epigenetic transgenerational inheritance in generational toxicology. Environ. Epigenetics 2022, 8, dvac001. [Google Scholar] [CrossRef]
- Petrakis, D.; Vassilopoulou, L.; Mamoulakis, C.; Psycharakis, C.; Anifantaki, A.; Sifakis, S.; Docea, A.O.; Tsiaoussis, J.; Makrigiannakis, A.; Tsatsakis, A.M. Endocrine Disruptors Leading to Obesity and Related Diseases. Int. J. Environ. Res. Public Heal. 2017, 14, 1282. [Google Scholar] [CrossRef]
- Brehm, E.; Flaws, J.A.J.E. Transgenerational effects of endocrine-disrupting chemicals on male and female reproduction. Endocrinology 2019, 160, 1421–1435. [Google Scholar] [CrossRef] [PubMed]
- de Solla, S.R.; King, L.E.; Gilroy. A. Environmental exposure to non-steroidal anti-inflammatory drugs and potential contribution to eggshell thinning in birds. Environ. Int. 2023, 171, 107638. [Google Scholar] [CrossRef]
- de Jager, C.; Patrick, S.; Aneck-Hahn, N.; Bornman, M. Environmental Toxicants and Sperm Production in Men and Animals. Proceedings of XIIIth International Symposium on Spermatology; pp. 47–59.
- Amir, S.; Shah, S.T.A.; Mamoulakis, C.; Docea, A.O.; Kalantzi, O.-I.; Zachariou, A.; Calina, D.; Carvalho, F.; Sofikitis, N.; Makrigiannakis, A.; et al. Endocrine Disruptors Acting on Estrogen and Androgen Pathways Cause Reproductive Disorders through Multiple Mechanisms: A Review. Int. J. Environ. Res. Public Heal. 2021, 18, 1464. [Google Scholar] [CrossRef]
- Interdonato, L.; Siracusa, R.; Fusco, R.; Cuzzocrea, S.; Di Paola, R. Endocrine Disruptor Compounds in Environment: Focus on Women’s Reproductive Health and Endometriosis. Int. J. Mol. Sci. 2023, 24, 5682. [Google Scholar] [CrossRef] [PubMed]
- Mrema, E.J.; Rubino, F.M.; Brambilla, G.; Moretto, A.; Tsatsakis, A.M.; Colosio, C. Persistent organochlorinated pesticides and mechanisms of their toxicity. Toxicology 2012, 307, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Int. J. Surg. 2021, 88, 105906. [Google Scholar] [CrossRef] [PubMed]
- Wójtowicz, A.K.; Gregoraszczuk, E.L.; Ptak, A.; Falandysz, J. Effect of single and repeated in vitro exposure of ovarian follicles to o,p'-DDT and p,p'-DDT and their metabolites. Pol. J. Pharmacol. 2004, 56. [Google Scholar]
- Wrobel, M.; Mlynarczuk, J.; Kotwica, J. The adverse effect of dichlorodiphenyltrichloroethane (DDT) and its metabolite (DDE) on the secretion of prostaglandins and oxytocin in bovine cultured ovarian and endometrial cells. Reprod. Toxicol. 2009, 27, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Mlynarczuk, J.; Górska, M.; Wrobel, M. Effects of DDT, DDE, aldrin and dieldrin on prostaglandin, oxytocin and steroid hormone release from smooth chorion explants of cattle. Anim. Reprod. Sci. 2020, 223, 106623. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowska, A.; Młynarczuk, J.; Kotwica, J.J.P.J.o.V.S. The protein expression disorders of connexins (Cx26, Cx32 and Cx43) and keratin 8 in bovine placenta under the influence of DDT, DDE and PCBs. Polish Journal of Veterinary Sciences 2018, 21, 721–729. [Google Scholar]
- Mlynarczuk, J.; Wrobel, M.H.; Kotwica, J. Effect of environmental pollutants on oxytocin synthesis and secretion from corpus luteum and on contractions of uterus from pregnant cows. Toxicol. Appl. Pharmacol. 2010, 247, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowska, A.; Mlynarczuk, J.; Kotwica, J. Changes in the mRNA expression of structural proteins, hormone synthesis and secretion from bovine placentome sections after DDT and DDE treatment. Toxicology 2017, 375, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wrobel, M.H.; Bedziechowski, P.; Mlynarczuk, J.; Kotwica, J. Impairment of uterine smooth muscle contractions and prostaglandin secretion from cattle myometrium and corpus luteum in vitro is influenced by DDT, DDE and HCH. Environ. Res. 2014, 132, 54–61. [Google Scholar] [CrossRef]
- Wójtowicz, A.K.; Kajta, M.; Gregoraszczuk, E. . DDT- and DDE-induced disruption of ovarian steroidogenesis in prepubertal porcine ovarian follicles: a possible interaction with the main steroidogenic enzymes and estrogen receptor beta. J. Physiol. Pharmacol. : Off. J. Pol. Physiol. Soc. 2007, 58. [Google Scholar]
- Munier, M.; Ayoub, M.; Suteau, V.; Gourdin, L.; Henrion, D.; Reiter, E.; Rodien, P.J.A.o.T. In vitro effects of the endocrine disruptor p, p′ DDT on human choriogonadotropin/luteinizing hormone receptor signalling. Archives of Toxicology 2021, 95, 1671–1681. [Google Scholar] [CrossRef]
- Kwekel, J.C.; Forgacs, A.L.; Williams, K.J.; Zacharewski, T.R.J.T.; pharmacology, a. op′-DDT-mediated uterotrophy and gene expression in immature C57BL/6 mice and Sprague–Dawley rats. Toxicology and applied pharmacology 2013, 273, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Salleh, N.; Giribabu, N.; Feng, A.O.M.; Myint, K.J.I.j.o.m.s. Bisphenol A, dichlorodiphenyltrichloroethane (DDT) and vinclozolin affect ex-vivo uterine contraction in rats via uterotonin (prostaglandin F2α, acetylcholine and oxytocin) related pathways. International journal of medical sciences 2015, 12, 914. [Google Scholar] [CrossRef]
- Lyche, J.L.; Nourizadeh-Lillabadi, R.; Almaas, C.; Stavik, B.; Berg, V.; Skåre, J.U.; Alestrøm, P.; Ropstad, E. Natural Mixtures of Persistent Organic Pollutants (POP) Increase Weight Gain, Advance Puberty, and Induce Changes in Gene Expression Associated with Steroid Hormones and Obesity in Female Zebrafish. J. Toxicol. Environ. Heal. Part A 2010, 73, 1032–1057. [Google Scholar] [CrossRef]
- Anand, M.; Agarwal, P.; Singh, L.; Taneja, A. Persistent organochlorine pesticides and oxidant/antioxidant status in the placental tissue of the women with full-term and pre-term deliveries. Toxicol. Res. 2015, 4, 326–332. [Google Scholar] [CrossRef]
- Anand, M.; Singh, L.; Agarwal, P.; Saroj, R.; Taneja, A. Pesticides exposure through environment and risk of pre-term birth: a study from Agra city. Drug Chem. Toxicol. 2017, 42, 471–477. [Google Scholar] [CrossRef]
- A Cohn, B.; Cirillo, P.M.; Wolff, M.S.; Schwingl, P.J.; Cohen, R.D.; I Sholtz, R.; Ferrara, A.; E Christianson, R.; Berg, B.J.v.D.; Siiteri, P.K. DDT and DDE exposure in mothers and time to pregnancy in daughters. Lancet 2003, 361, 2205–2206. [Google Scholar] [CrossRef]
- Mahalingaiah, S.; Missmer, S.A.; Maity, A.; Williams, P.L.; Meeker, J.D.; Berry, K.; Ehrlich, S.; Perry, M.J.; Cramer, D.W.; Hauser, R.J.E.h.p. Association of hexachlorobenzene (HCB), dichlorodiphenyltrichloroethane (DDT), and dichlorodiphenyldichloroethylene (DDE) with in vitro fertilization (IVF) outcomes. Environmental health perspectives 2012, 120, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Torres-Arreola, L.; Berkowitz, G.; Torres-Sánchez, L.; López-Cervantes, M.; E Cebrián, M.; Uribe, M.; López-Carrillo, L. Preterm Birth in Relation to Maternal Organochlorine Serum Levels. Ann. Epidemiology 2003, 13, 158–162. [Google Scholar] [CrossRef]
- Tyagi, V.; Garg, N.; Mustafa, M.D.; Banerjee, B.D.; Guleria, K. Organochlorine pesticide levels in maternal blood and placental tissue with reference to preterm birth: a recent trend in North Indian population. Environ. Monit. Assess. 2015, 187, 471. [Google Scholar] [CrossRef]
- Arrebola, J.P.; Cuellar, M.; Bonde, J.P.; González-Alzaga, B.; Mercado, L.A.J.E.R. Associations of maternal o, p′-DDT and p, p′-DDE levels with birth outcomes in a Bolivian cohort. Environmental Research 2016, 151, 469–477. [Google Scholar] [CrossRef]
- Farhang, L.; Weintraub, J.M.; Petreas, M.; Eskenazi, B.; Bhatia, R. Association of DDT and DDE with Birth Weight and Length of Gestation in the Child Health and Development Studies, 1959–1967. Am. J. Epidemiology 2005, 162, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Kezios, K.L.; Liu, X.; Cirillo, P.M.; Cohn, B.A.; Kalantzi, O.I.; Wang, Y.; Petreas, M.X.; Park, J.-S.; Factor-Litvak, P. Dichlorodiphenyltrichloroethane (DDT), DDT metabolites and pregnancy outcomes. Reprod. Toxicol. 2012, 35, 156–164. [Google Scholar] [CrossRef]
- Longnecker, M.P.; Klebanoff, M.A.; Dunson, D.B.; Guo, X.; Chen, Z.; Zhou, H.; Brock, J.W. Maternal serum level of the DDT metabolite DDE in relation to fetal loss in previous pregnancies. Environ. Res. 2005, 97, 127–133. [Google Scholar] [CrossRef]
- Ouyang, F.; Longnecker, M.P.; Venners, S.A.; Johnson, S.; Korrick, S.; Zhang, J.; Xu, X.; Christian, P.; Wang, M.-C.; Wang, X.J.T.A.J.o.C.N. Preconception serum 1, 1, 1-trichloro-2, 2, bis (p-chlorophenyl) ethane and B-vitamin status: independent and joint effects on women’s reproductive outcomes. The American Journal of Clinical Nutrition 2014, 100, 1470–1478. [Google Scholar] [CrossRef] [PubMed]
- Perry, M.J.; Ouyang, F.; Korrick, S.A.; Venners, S.A.; Chen, C.; Xu, X.; Lasley, B.L.; Wang, X. A Prospective Study of Serum DDT and Progesterone and Estrogen Levels across the Menstrual Cycle in Nulliparous Women of Reproductive Age. Am. J. Epidemiology 2006, 164, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Venners, S.A.; Korrick, S.; Xu, X.; Chen, C.; Guang, W.; Huang, A.; Altshul, L.; Perry, M.; Fu, L.; Wang, X. Preconception Serum DDT and Pregnancy Loss: A Prospective Study Using a Biomarker of Pregnancy. Am. J. Epidemiology 2005, 162, 709–716. [Google Scholar] [CrossRef]
- Harley, K.G.; Marks, A.R.; Bradman, A.; Barr, D.B.; Eskenazi, B.J.J.o.o.; Occupational, e.m.A.C.o.; Medicine, E. DDT exposure, work in agriculture, and time to pregnancy among farmworkers in California. Journal of occupational and environmental medicine/American College of Occupational and Environmental Medicine 2008, 50, 1335. [Google Scholar] [CrossRef]
- Ouyang, F.; Perry, M.J.; A Venners, S.; Chen, C.; Wang, B.; Yang, F.; Fang, Z.; Zang, T.; Wang, L.; Xu, X.; et al. Serum DDT, age at menarche, and abnormal menstrual cycle length. Occup. Environ. Med. 2005, 62, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Zhang, J.; Zhou, L.; Gao, E.-S.; Chen, L.; Rogan, W.J.; Wolff, M.S. DDT serum concentration and menstruation among young Chinese women. Environ. Res. 2005, 99, 397–402. [Google Scholar] [CrossRef]
- Weiss, J.M.; Bauer, O.; Blüthgen, A.; Ludwig, A.K.; Vollersen, E.; Kaisi, M.; Al-Hasani, S.; Diedrich, K.; Ludwig, M. Distribution of persistent organochlorine contaminants in infertile patients from Tanzania and Germany. J. Assist. Reprod. Genet. 2006, 23, 393–399. [Google Scholar] [CrossRef]
- Windham, G.C.; Lee, D.; Mitchell, P.; Anderson, M.; Petreas, M.; Lasley, B. Exposure to Organochlorine Compounds and Effects on Ovarian Function. Epidemiology 2005, 16, 182–190. [Google Scholar] [CrossRef]
- Bredhult, C.; Bäcklin, B.-M.; Bignert, A.; Olovsson, M. Study of the relation between the incidence of uterine leiomyomas and the concentrations of PCB and DDT in Baltic gray seals. Reprod. Toxicol. 2008, 25, 247–255. [Google Scholar] [CrossRef]
- A Gibson, D.; Saunders, P.T.K. Endocrine disruption of oestrogen action and female reproductive tract cancers. Endocrine-Related Cancer 2013, 21, T13–T31. [Google Scholar] [CrossRef]
- Kalinina, T.S.; Kononchuk, V.V.; Ovchinnikov, V.Y.; Chanyshev, M.D.; Gulyaeva, L.F. Expression of the miR-190 family is increased under DDT exposure in vivo and in vitro. Mol. Biol. Rep. 2018, 45, 1937–1945. [Google Scholar] [CrossRef]
- Mathur, V.; John, P.J.; Soni, I.; Bhatnagar, P. Bhatnagar, P. Blood Levels of Organochlorine Pesticide Residues and Risk of Reproductive Tract Cancer Among Women from Jaipur, India. 2008, 617, 387–394. 617. [CrossRef]
- Ndebele, K.; Graham, B.; Tchounwou, P.B.J.I.J.o.E.R.; Health, P. Estrogenic activity of coumestrol, DDT, and TCDD in human cervical cancer cells. International Journal of Environmental Research and Public Health 2010, 7, 2045–2056. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, G.; Priyadarshini, V.; Singh, A. Association of Serum and Cervical Tissue Levels of Organochlorine Pesticides with Cervical Cancer in Women of East Delhi: A Case Control Pilot Study. J. South Asian Fed. Obstet. Gynaecol. 2019, 11, 190–193. [Google Scholar] [CrossRef]
- Rodríguez. G.P.; López, M.I.R.; Casillas, T..D.; León, J.A.A.; Mahjoub, O.; Prusty, A.K. Monitoring of organochlorine pesticides in blood of women with uterine cervix cancer. Environ. Pollut. 2017, 220, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.; Banerjee, B.D.; Mazumdar, D.; Tyagi, V.; Thakur, G.; Guleria, K.; Ahmed, R.S.; Tripathi, A.K. Association of organochlorine pesticides and risk of epithelial ovarian cancer: A case control study. J. Reprod. Heal. Med. 2015, 1, 76–82. [Google Scholar] [CrossRef]
Figure 1.
PRISMA Flowchart.
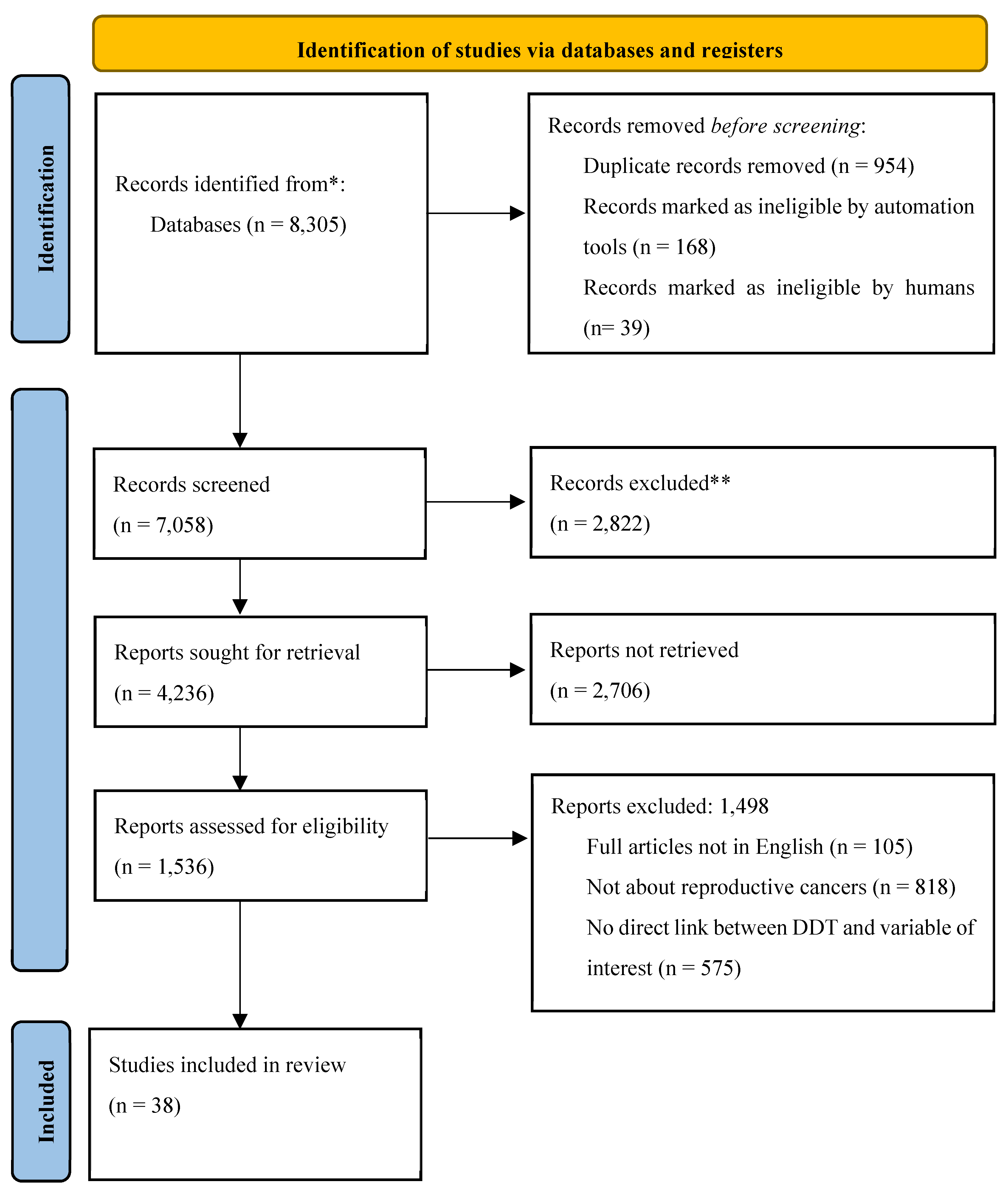
Figure 2.
Reduction of Estrogen in Follicular Cells. DDT acts as an antagonistic ligand to Er𝛽 receptor of E2 and inhibits the enzymatic activity of Cytochrome P450, both of which result in reduced estrogen secretions.
Figure 2.
Reduction of Estrogen in Follicular Cells. DDT acts as an antagonistic ligand to Er𝛽 receptor of E2 and inhibits the enzymatic activity of Cytochrome P450, both of which result in reduced estrogen secretions.
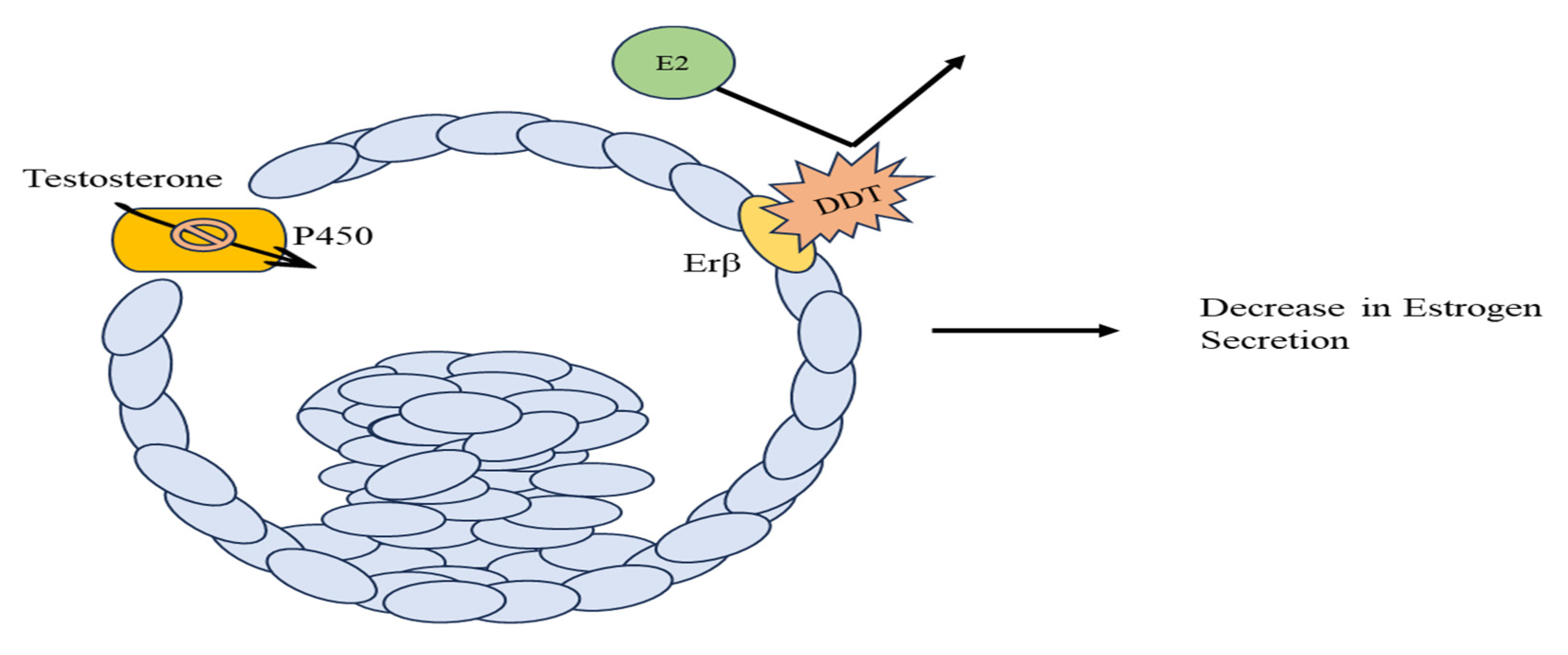
Figure 3.
Effect of DDT on Myometrial, Endometrial, Granulosa, and Luteal Cells. DDT causes Myometrial Hypertrophy which increases uterine contractions and Increases the synthesis of Leukemia Inhibitory Factor (LIF) and alters the synthesis of Prostaglandins in myometrial and endometrial cells which results in reduced PGE2 levels and elevated PGF2∝ levels. The increase in PGF2∝ is in a positive feedback mechanism with the secretion of Oxytocin by Luteal Cells. Along with Luteal cells, Granulosa Cells also secrete more Oxytocin under the impact of DDT which enhances the Luteal regression thereby, disrupting the P4: OT.
Figure 3.
Effect of DDT on Myometrial, Endometrial, Granulosa, and Luteal Cells. DDT causes Myometrial Hypertrophy which increases uterine contractions and Increases the synthesis of Leukemia Inhibitory Factor (LIF) and alters the synthesis of Prostaglandins in myometrial and endometrial cells which results in reduced PGE2 levels and elevated PGF2∝ levels. The increase in PGF2∝ is in a positive feedback mechanism with the secretion of Oxytocin by Luteal Cells. Along with Luteal cells, Granulosa Cells also secrete more Oxytocin under the impact of DDT which enhances the Luteal regression thereby, disrupting the P4: OT.
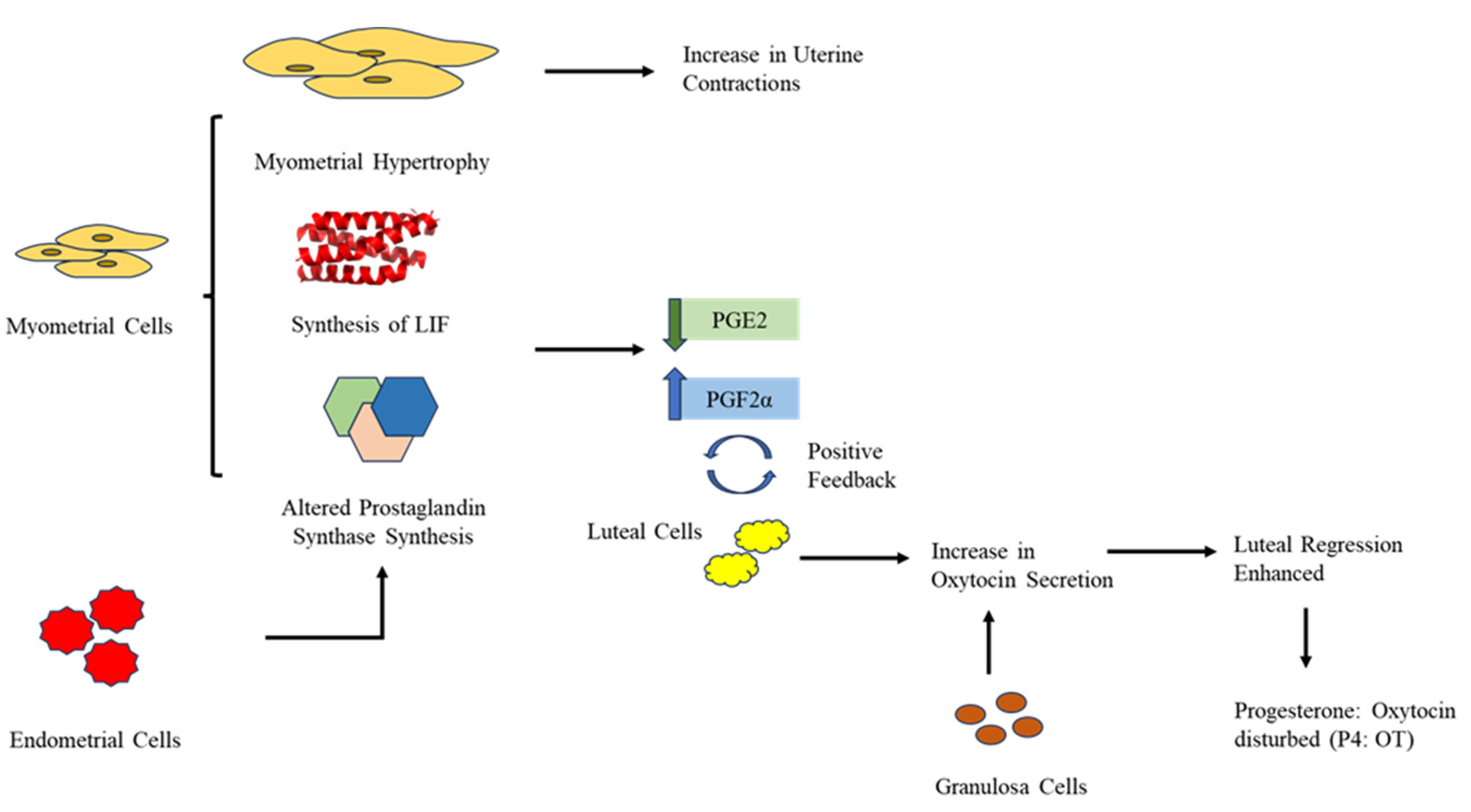
Figure 4.
DDT exposed stromal cells. DDT causes Stromal Hypertrophy and Edema which results in increased Uterine Wet Weight (UWW), both of which increase nuclear contractions.
Figure 4.
DDT exposed stromal cells. DDT causes Stromal Hypertrophy and Edema which results in increased Uterine Wet Weight (UWW), both of which increase nuclear contractions.
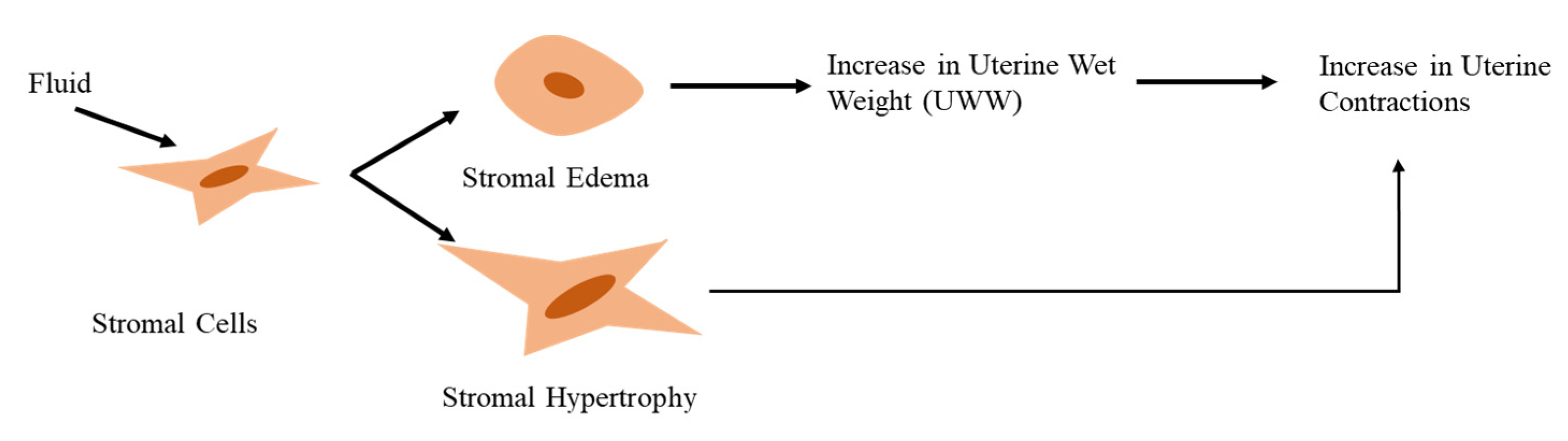
Figure 5.
Ion Imbalance in Uterine Cell Membrane. DDT causes reduced influx and intracellular release of Calcium Ions through the Uterine Cell Membrane which results in decreased uterine contractions.
Figure 5.
Ion Imbalance in Uterine Cell Membrane. DDT causes reduced influx and intracellular release of Calcium Ions through the Uterine Cell Membrane which results in decreased uterine contractions.
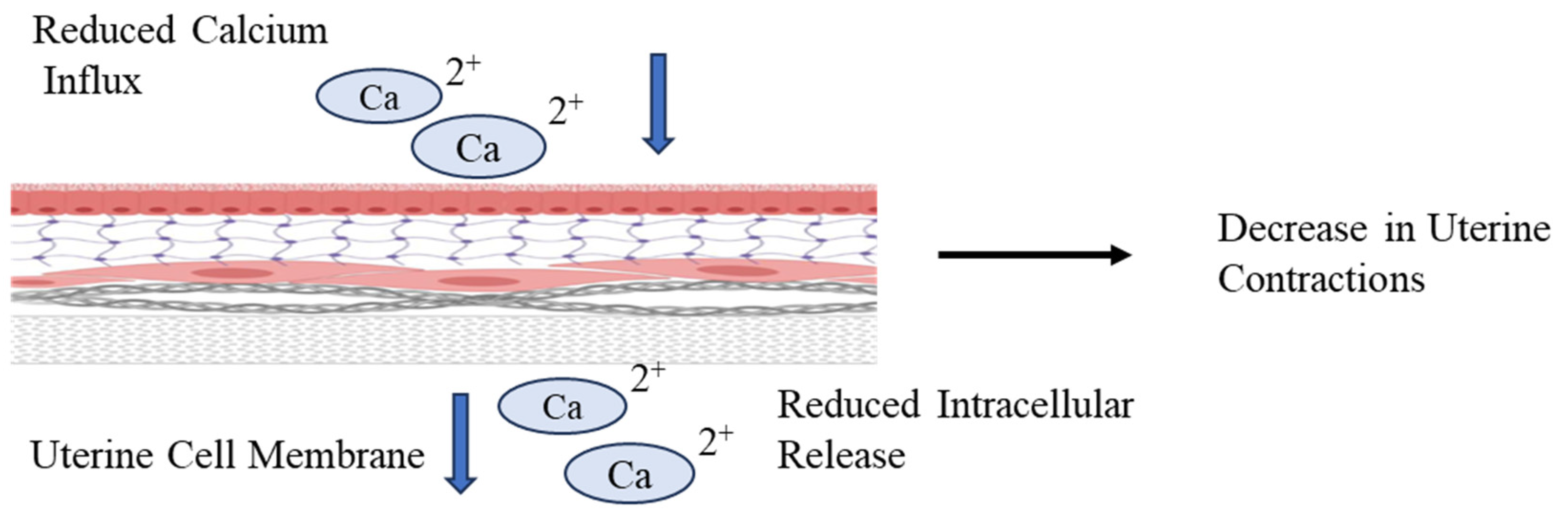
Figure 6.
Pathway Inhibition by DDT. DDT alters the levels of Human Chorionic Gonadotropin (hCG) and Luteinizing Hormone (LH) by inhibition of cAMP and 𝛽- Arrestin Pathways.
Figure 6.
Pathway Inhibition by DDT. DDT alters the levels of Human Chorionic Gonadotropin (hCG) and Luteinizing Hormone (LH) by inhibition of cAMP and 𝛽- Arrestin Pathways.
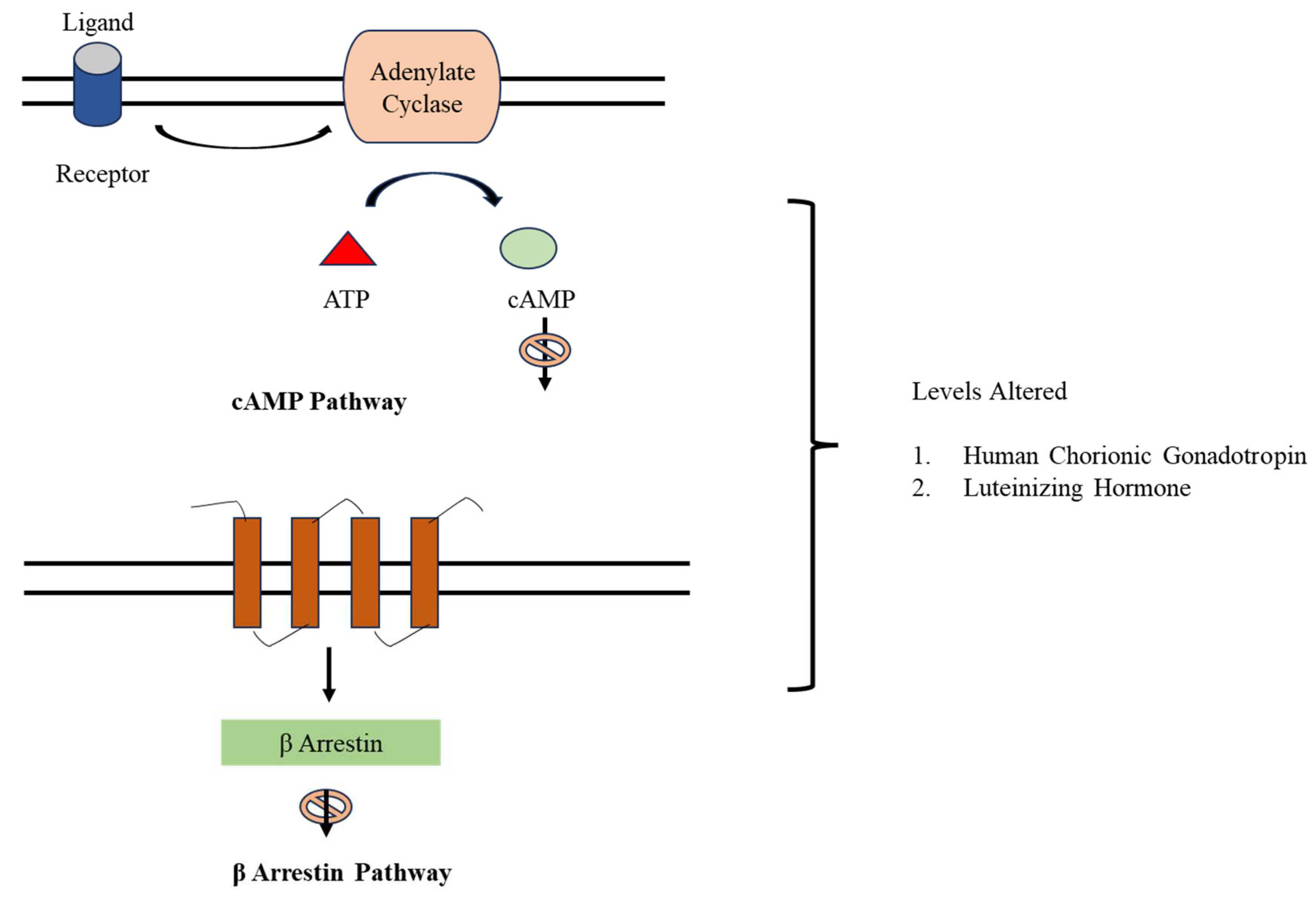

Table 1.
Impact of DDT and its metabolites on hormone levels. DDT and its metabolites alter hormone secretion levels by increasing or decreasing their production through various mechanisms.
Table 1.
Impact of DDT and its metabolites on hormone levels. DDT and its metabolites alter hormone secretion levels by increasing or decreasing their production through various mechanisms.
| Chemical/Hormone | DDT Isomer | Levels of Chemical/Hormone |
| Estradiol | ο,p΄-DDT | Increase or Decrease |
| ο,p΄-DDE | Increase or Decrease | |
| ο,p΄-DDD | Increase or Decrease | |
| p,p΄-DDT | Increase | |
| p ,p΄-DDE | Increase | |
| Oxytocin | DDT | Increase |
| DDE | Increase | |
| Prostaglandin A | DDT | Decrease |
| DDE | Decrease | |
| Prostaglandin F2a | DDT | Increase |
| DDE | Increase | |
| Prostaglandin E | DDT | Decrease |
| DDE | Decrease | |
| Progesterone | DDE | Increase |
| hCG/LHR | p, p’- DDT | Decrease |
Table 2.
Impact of DDT and its metabolites on reproductive disorders based on ethnicities. DDT and its metabolites were associated with the incidence of various parameters of reproductive disorders in women belonging to different ethnic backgrounds.
Table 2.
Impact of DDT and its metabolites on reproductive disorders based on ethnicities. DDT and its metabolites were associated with the incidence of various parameters of reproductive disorders in women belonging to different ethnic backgrounds.
| Sr no. | Study Type | Ethnicity | Population | Metabolite | Parameter | Effect |
| 1 | Case-control | American | 289 | p,p΄-DDT p,p΄-DDE | TTP | Decreased |
| 2 | Case-control | Mexican | 233 | p,p΄-DDE | PTB | Increased |
| 3 | Nested Case- Control | American | 720 | DDT/DDE | Infertility | No statistically significant association |
| 4 | Case-control | North Indian | 100 | DDT/DDE | PTB | Increased |
| 5 | Case-control | Indian | 90 | p,p΄-DDE | PTB | Increased |
| 6 | Case-control | Indian | 90 | p,p΄-DDT p,p΄-DDE | PTB | Increase |
| 7 | Cohort | American | 2613 | DDT/DDE | Fetal loss | DDE Increased, no relation with DDT |
| 8 | Cohort | American | 20,754 | DDT/DDE | PTB | No statistically significant association |
| 9 | Cohort | Chinese | 287 | o,p΄-DDT o,p΄-DDE | Menstrual cycle length | Increased |
| 10 | Cohort | American | 1752 | p,p΄-DDT, o,p’-DDT, p,p΄-DDE | POG | Decreased |
| 11 | Prospective | Chinese | 291 | DDT | Clinical pregnancy | Decreased |
| 12 | Cohort | Bolivian | 200 | p,p΄-DDE o,p΄-DDT | POG | Decreased |
| 13 | Prospective | Chinese | 388 | DDT | Fetal loss | Increased |
| 14 | Cross-sectional | Chinese | 466 | p,p΄-DDE/DDT | Menstrual cycle length | Reduced age at menarche |
| 15 | Cross-sectional | Latina | 402 | p,p΄-DDT, o,p'-DDT p,p΄-DDE | TTP | No statistically significant association |
| 16 | Pilot | Chinese | 60 | p,p΄-DDT o,p΄-DDT | Menstrual cycle length | No statistically significant association |
| 17 | Pilot | German | 89 | DDT | Infertility | Increased |
| 18 | Pilot | Laotian | 50 | DDT/DDE | Menstrual cycle length | Decreased |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



