
Submitted:
26 July 2023
Posted:
28 July 2023
You are already at the latest version
Abstract
bZIP transcription factors, basic leucine zipper, play significant roles in plants’ growth and development processes, as well as in response to biological and abiotic stress. Hypericum perforatum is one of the world's top three best-selling herbal medicines, mainly used to treat depression. However, there is no systematic identification or functional analysis of the bZIP gene family in H. perforatum. In this study, 79 HpbZIP genes were identified. Based on phylogenetic analysis, the HpbZIP gene family was divided into ten groups, designated A-I, and S. The physicochemical properties, gene structures, protein conserved motifs, and Gene Ontology enrichments of all HpbZIPs were systematically analyzed. The expression patterns of all genes in different tissues of H. perforatum (root, stem, leaf, and flower) were analyzed by qRT-PCR, revealing the different expression patterns of HpbZIP under abiotic stresses. HpbZIP69 protein is localized in the nucleus. According to the results of the yeast one-hybrid (Y1H) assays, HpbZIP69 can bind to the HpASMT2 (N-Acetylserotonin O-methyltransferase) gene promoter ( G-box cis-element ) to activate its activity. Overexpressing HpbZIP69 transgenic Arabidopsis lines enhanced tolerance to drought. The content of MDA and H2O2 content was significantly decreased, and the activity of superoxide dismutase (SOD) was considerably increased under the drought stress. These results may aid in additional functional studies of HpbZIP transcription factors and cultivating drought-resistant medicinal plants.
Keywords:
bZIP transcription factor
; Hypericum perforatum
; Drought stress
1. Introduction
bZIP TFs, also known as basic leucine zipper, are a widely distributed and highly conserved multigene family in eukaryotes [1]. It plays an essential role in regulating plant growth and development process and promoting the synthesis of secondary metabolites bZIP protein is famous for its highly conserved bZIP domains consisting of 40~60 amino acids [2]. This domain consists of two structural features: one is the primary binding region (N-X7-R/K-X9), and the other is the leucine zipper dimer domain [3]. The basic binding region consists of about 18 amino acid residues and binds to specific DNA sequences with the help of a fixed N-X7-R/K structure [4]. The leucine zipper dimer domain is composed of several heptavalent repeats of leucine or other hydrophobic amino acids, which bind tightly to the essential region. The leucine zipper forms an amphiphilic α helix, which can regulate the homologous or heterodimerization of bZIP protein before binding to DNA [5]. bZIP proteins in plants have binding specificity for DNA sequences containing ACGT elements, which promotes its preferential binding to G-box (CACGTG), C-box (GACGTC), and A-box (TACGTA) [6]. On DNA binding, one-half of the n-terminal basic binding region inserts into the large groove of double-stranded DNA. The remaining half of the C-terminal leucine zipper regulates dimerization, thus forming a superimposed helical coil structure [7]. Such a structure also determines that bZIP transcription factors can regulate the expression of multiple downstream genes by interacting with cis-acting elements in the promoter region, thus participating in the transcriptional regulation process.
At present, a variety of bZIP transcription factors have been shown to regulate the transcriptional expression of related genes by interacting with genes in response to biotic and abiotic stresses [8]. Under salt stress conditions, AtbZIP17 in Arabidopsis directly or indirectly regulates some salt stress-related response genes, thereby participating in the salt signaling cascade [9]. AtbZIP53 can interact with AtbZIP10 to form dimers, further regulate proline metabolism and participate in abiotic stress response [10]. AtbZIP53 can also bind to AtbZIP1, participate in carbon and nitrogen metabolism pathways, and affect the decomposition and metabolism of sugars and amino acids [11]. In the study of Brachypodium distachyon and Oryza sativa, it was found that the expression levels of BdbZIP30 and BdbZIP41 and their homologous genes OsbZIP63 and OsbZIP05 in O. sativa increased under high salt conditions[12, 13]. Overexpression of wheat (Triticum aestivum) gene TabZIP6 in Arabidopsis reduced its frost resistance [14]. Overexpression of Camellia sinensis bZIP6 can enhance the ability to resist low temperature stress in Arabidopsis [15]. Furthermore, bZIP transcription factors can specifically bind to ABRE (abscisic acid response element) in response to drought stresses [16, 17]. For example, AtbZIP36 can be combined with the cis-acting element ABRE in the promoter region to promote and inhibit the expression of stress regulatory proteins and participate in the stress response pathway, thus improving the drought tolerance of Arabidopsis [18]. Moreover, ramie BnbZIP2 was overexpressed in Arabidopsis, which made transgenic Arabidopsis show stronger drought resistance compared with the wild type [19]. Exogenous hormone ABA significantly affects bZIP transcription factor. In the study of TabZIP14-B in wheat, it was found that TabZIP14-B transgenic plants were more sensitive to ABA than wild types, which severely inhibited plant root growth [20]. OsbZIP62, a stress-responsive bZIP transcription factor, was found that improved drought and oxidative tolerance in rice [21]. On the basis of expression pattern analyses, JcbZIP49 and JcbZIP50 are likely involved in responses to drought stress in Jatropha curcas [22]. The overexpression of TabZIP8-7A conferred greater drought resistance and ABA sensitivity in Arabidopsis [23]. Transcript accumulation of AtbZIP62 and AtPYD1 showed that both were highly up-regulated by drought stress in wild type (WT) plants [24]. The TGA subfamily of bZIP transcription factors has been studied in depth, which proves that this family is relevant to plant disease resistance [4, 25, 26]. For instance, TGA2, TGA5, and TGA6 are essential for plant disease resistance and have redundant functions [27].
The planting area of traditional Chinese medicine is constantly increasing in China, and the demand for land is also increasing. Promoting the cultivation of H. perforatum in arid and semi-arid areas has become one of the important means to alleviate its resource shortage. Drought is a highly destructive and frequent global natural disaster that restricts the yield and quality of traditional Chinese medicinal materials. Therefore, studying of stress resistance genes and improving the ability of H. perforatum to resist drought stress is of great significance for improving its yield and quality [28]. Numerous studies have shown that the bZIP transcription factors have many important biological functions, particularly in enhancing plant drought resistance [29, 30]. To reveal the detail and to facilitate future research on bZIP TF family in H.perforatum, the gene figure, classification, and stress-produced expression modes of bZIP TF family members in H. perforatum were systematically analyzed based on whole genome data. The phenotype and RNA-seq information of HpbZIP69 transgenic Arabidopsis were analyzed to investigate the characteristics and molecular functions of HpbZIP69 under drought stress. The outcome will lay a foundation for molecular biology of drought resistance in H. perforatum.
2. Result
2.1. Identification and sequence feature
The Arabidopsis thaliana bZIPs are taken as the comparison objects. Through screening out of the H. perforatum genome database (No. PRJNA588586), 79 members of the H. perforatum bZIPs were finally obtained and named as HpbZIP1 to HpbZIP79. By bioinformatics software analysis, the physicochemical properties of the bZIPs of H. perforatum are shown in Supplemental Table 2. The lengths of the proteins encoded by the H. perforatum bZIPs range from 119 aa (HpbZIP46) to 575 aa (HpbZIP79); the iso-electric points range from 4.65 (HpbZIP21) to 10.02 (HpbZIP53); the molecular weights range from 14.27 KDa (HpbZIP46) to 73.8 KDa (HpbZIP32). The subcellular localization prediction of all genes is carried out, and the results are shown in Supplemental Table 2.
2.2. Phylogenetic analysis of HpbZIPs
To seek the evolution relationship of bZIP TFs, we generated a phylogenetic tree, including 75 Arabidopsis and 79 H. perforatum bZIPs based on the alignment of the amino acid sequences (Figure 1). On account of the topology of phylogenetic tree and category of Arabidopsis bZIPs, ten groups (named as A-I, and S) were categorized. According to the distribution characteristics of conserved domains and other structural components in its protein structure, the H. perforatum bZIP gene family can be divided into ten groups correspondingly. The structure diagram of each subfamily represents the protein is shown in Supplemental Figure 1. Moreover, HpbZIP5 and HpbZIP36 did not classify into any group, which indicates that the sequence have varied in the progress of evolution, and the functions may have changed considerably. Based on the phylogenetic tree, the largest cluster is S group, containing 20 AtbZIPs and 19 HpbZIPs. The second largest cluster is Group A, which contains 12 AtbZIPs and 14 HpbZIPs. According to the gene functions of different groups, Group A is mainly involved in ABA and stress-mediated signal transduction. In contrast, Group S is mainly involved in biological processes in response to stress, such as cold, drought and injury. Members in Group S and Group A are almost involved in drought and ABA stress response, further demonstrating that Group S and Group A play an essential role in the stress regulation of plants. Different from Arabidopsis, Group S, and Group I can be subdivided into four subgroups (designated as Sa, Sb, Ia, and Ib) due to minor differences in gene clustering.
2.3. Gene structure and conserved motif analyses
To further research the phylogenetic connections among HpbZIP TFs, an unrooted phylogenetic tree only containing HpbZIPs was made, and the exon-intron structure of and allocations of conserved domains in HpbZIPs were analyzed (Figure 2). The HpbZIPs were classified and named as before. The results of multiple sequence alignment forecasted that every HpbZIP had a conserved bZIP domain, and its N-terminal region was an alkaline region containing N-X7-R/K motif, as well as its C-terminal leucine zipper region contained leucine every seven hydrophobic amino acids. Exon-intron structures of HpbZIPs between members were shown in Figure 2. Genetic structure suggested a significant specificity between groups. Because the same subtribe in introns and exons is relatively similar, the distribution and quantity of splice sites were fairly conservative and which same subtribe gene on the evolutionary relationships was more closely. Some HpbZIP genes (28%) were intronless. Six genes only had CDS sequences, which called HpbZIP18, HpbZIP13, HpbZIP53, HpbZIP63, HpbZIP15, and HpbZIP68, respectively. The number of exons was 12 at most and one at least. The number of introns was 11 at most and one at least. Compared with other genes in the bZIP gene family, the length of HpbZIP46 was the largest, about 5-6 times the average length of other genes, with highly long introns and very short CDS sequences. There were seven groups (HpbZIP6/HpbZIP7, HpbZIP31/HpbZIP41, HpbZIP17/HpbZIP67, HpbZIP50/HpbZIP79, HpbZIP38/HpbZIP74, HpbZIP19/HpbZIP51 and HpbZIP57/HpbZIP62) that contained the same number of exons and similar length of exons, suggesting that there may be tandem repeats in the HpbZIP gene family.
We identified 15 conserved motifs in 79 HpbZIP proteins (Supplemental Figure 2) based on the phylogenetic tree and MEME software. Different bZIP genes have different types of conserved motifs, and the number of motifs is different, suggesting different potential functional sites and different biological functions that may be involved. Although different bZIP genes have different types of conserved motifs, the type and number of conserved motifs in the same subfamily are still relatively similar. The length of all motifs was between 6 to 50 amino acids. Some the bZIP members had six motifs; other bZIP members owned two at least. Motif 1 existed in almost every HpbZIP. Some motifs existed in various genes. For example, motif 14 was identified in 50 HpbZIPs. A few motifs, like motif 13, only existed in 4 HpbZIP, while most conserved motifs occurred in particular groups. For instance, motif 9 and motif 3 were found in group B, motif 8 and motif 2 were identified in group D, and motif 5 and motif 12 existed in group C (Supplemental Table 3). The similar composition and distribution patterns of exon–intron structure and conserved domain facilitated the phylogenetic connection and category of the HpbZIP TFs.
2.4. Transcript abundance profiling
The upstream 1,5 kb of ATG was used to predict cis-elements. As shown in Supplemental Figure 3 and Supplemental Table 4, plenty of cis-elements connected with growth and development, hormonal regulations and stresses were identified in 79 HpbZIP gene promoters. There were 64 genes were responding to drought, including DRE and MBS and 176 involved in photoresponse regulation, such as GT1-motif, MNF1, Box I, 3-AF1 binding site, ATCC -motif, 4cl-CMA2b, and 45 contributing to abscisic acid response (ABRE). Analysis of cis-acting elements of HpbZIP gene family members, especially the functional annotation contribute to the subsequent available study of bZIP genes. For the GO annotation analysis, 74 HpbZIP proteins were summarized into 25 functional subcategories (Supplemental Figure 4) of the three main ontologies according to amino acid similarity. There were only four proteins enriched in membrane, membrane part, organelle part, and protein-containing complex. The number of proteins enriched in a cell, cell par, and organelle was 74. Among the molecular functions, the proteins enriched in the GO annotation on the nutrient storage activity were the minimum, and the proteins enriched in the binding and transcription regulator activity were the most intensive. In addition, some proteins were increased in response to stimulus and biological regulation processes, accounting for about 50% of the total protein number. It is speculated that members of this protein family can bind cis-acting elements to activate the expression of critical genes in the transcriptional regulation process.
2.5. Expression patterns analysis
Our laboratory has established the transcriptome database of H. perforatum in different tissues (flower, leaf, root, and stem; SRR8438983-SRR8438986). The formula log10(FPKM) from the RNA-seq data were applied to hierarchical clustering. The HpbZIP genes displayed different expression patterns in four organs showed in the heatmap (Figure 3A). For example, the expressions of HpbZIP59, HpbZIP70, and HpbZIP77 were similar in all tissues. However, some genes were highly expressed in specific sites and low in others, such as HpbZIP11, HpbZIP15, and HpbZIP27, implying that gene expression might be development-specific.
In addition, in order to investigate the response of members of the HpbZIP gene family to abiotic stresses, we selected 1-2 representative genes from 10 subfamilies respectively according to the results of evolutionary groups, and analyzed the expression patterns of 12 genes under hormone treatment (ABA; Figure 4A) and abiotic stresses (NaCl, PEG; Figure 4B and 4C). RT-qPCR was used to detect the expression levels at 0, 1, 3, 6, and 12 h. The expression levels of different genes were increased under ABA treatment. With the increase in treatment time, HpbZIP14, HpbZIP69, and HpbZIP37 showed a significant up-regulation trend, while HpbZIP31 and HpbZIP40 showed a down-regulated trend. The expression of HpbZIP31 increased sharply at a specific time point. Under drought and high salt treatment, gene expression levels were generally higher, and the expression levels showed a more apparent temporal difference (Figure 4B, C). Among them, the relative expression levels of HpbZIP69 were significantly increased under drought treatment compared with the control (0 h). The plenty of expression patterns revealed the different roles of HpbZIP genes in abiotic stress-response pathways.
2.6. Characterization of Transcription Activity of HpbZIP69
According to the evolutionary relationship, HpbZIP69 is homologous to ABF3 (AtbZIP37) and ABF4/AREB2 (AtbZIP38) [18, 31]which is involved in the regulation of abiotic stress such as drought. It is speculated that HpbZIP69 also has the similar function. Meanwhile, combined with the analysis of gene expression pattern under drought treatment, the expression level of HpbZIP69 was also significantly increased. The full length of HpbZIP69 cDNA is 1,125 bp, encoding 374 amino acids, with a molecular weight of 40.28 KDa and the IP of 9.4. Bioinformatics software analysis showed that HpbZIP69 was mainly localized in the nucleus (Supplemental Figure 5), which was consistent with the general characteristics of transcription factors. To verify the localization of HpbZIP69, the HpbZIP69 was fused with GFP driven by 35S promoter and transiently expressed in onion epidermal cells. The HpbZIP69-GFP was detected in the nucleus (Figure 5A), pointing that HpbZIP69 was a nucleus-localized protein. As shown in Figure 5B, the N-terminal region (1-298 aa) without bZIP domain had the minimal demand for its trans activity as a matter of facts that bZIP transcription factors bind the G-box motif (CACGTG). The yeast-one-hybrid assay indeed showed that HpbZIP69 binds G-box with a conserved sequence (Figure 5C). The results above-mentioned showed that HpbZIP69 was a characteristic bZIP transcription factor.
2.7. Overexpression of HpbZIP69 in Arabidopsis
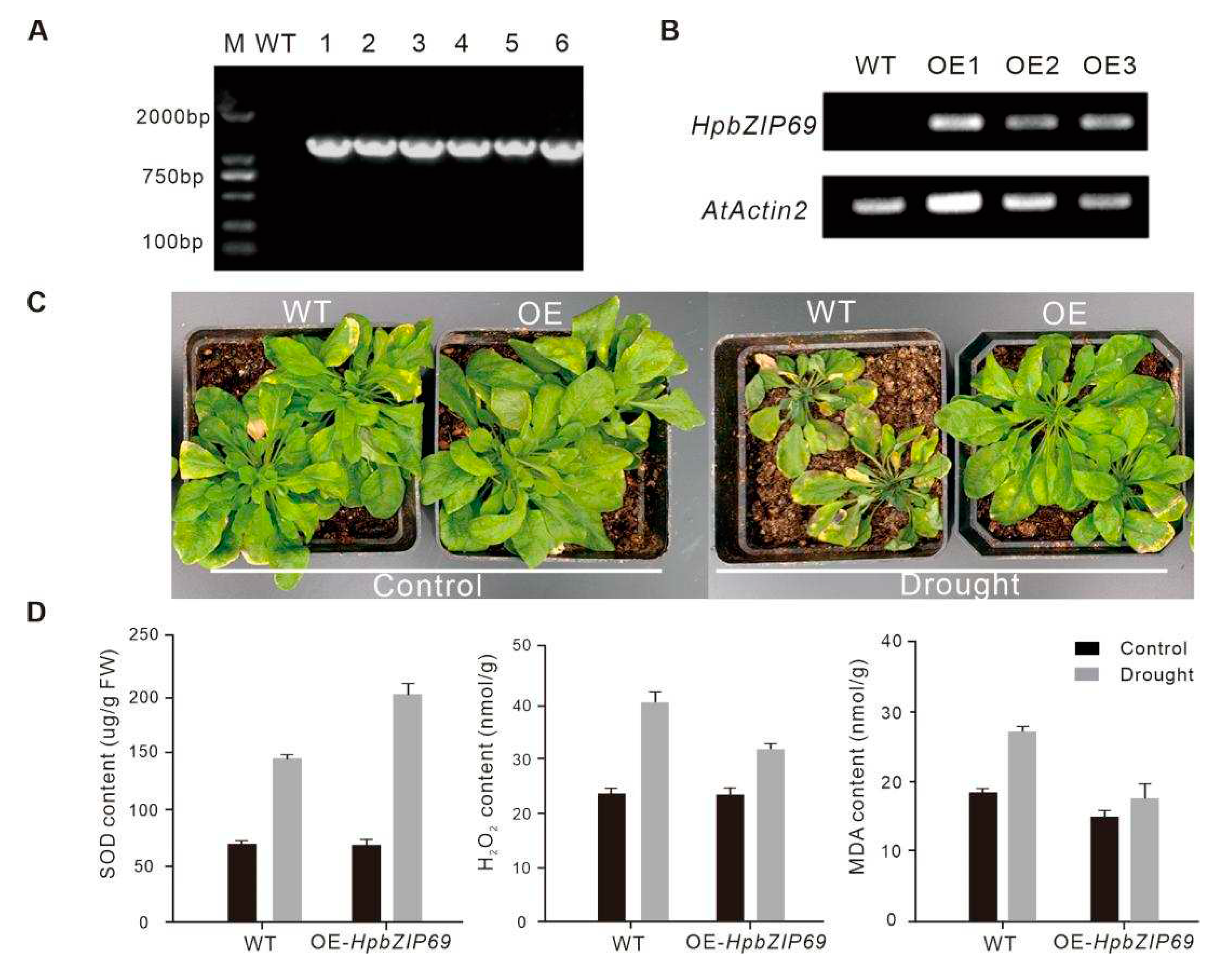
To investigate the function of HpbZIP69, pEarleyGate202-HpbZIP69 was transferred into Arabidopsis (Col-0). The target gene was amplified and identified to verify whether the transgenic plants were positive. HpbZIP69 were cloned from six candidate transgenic lines, but no bands were found at 1000-2000 bp in WT (Figure 6A). Three independent stable homozygous lines were acquired. They were confirmed using semi-qRT-PCR (Figure 6B). According to the result of semi-qPCR, the expression of HpbZIP69 is higher than OE-2 and OE-3, OE-1 line was selected for subsequent functional assays. To see whether HpbZIP69 can enhance the drought of plants, one-month-old field Arabidopsis seedlings were used for drought-tolerance analysis. The growth development of OE lines was obviously better than that of WT without water for 15 days (Figure 6C). The content of SOD in OE lines was higher than those in WT under drought treatment, and the MDA and H2O2 contents in OE lines were notably lower than in WT (Figure 6D). The consequences above-mentioned revealed that the ectopic expression of HpbZIP69 enhanced the drought tolerances in Arabidopsis.
3. Discussion
The H. perforatum bZIP gene family is multigene widely existing and highly conserved in eukaryotes, which plays a vital role in regulating plant growth and development, participating in stress response, and promoting the synthesis of secondary metabolites [32]. In this study, based on H. perforatum genome-wide database, we screened 79 HpbZIP genes, and the relationships between the physical and chemical properties, system development evolution, gene structures, conservative motifs, gene function enrichment regions, cis-elements, as well as under different tissues and different hormone gene expression patterns and made a preliminary analysis. Phylogenetic analysis showed that the bZIP gene family of H. perforatum had the same evolutionary group as that of A. thaliana, which was also divided into ten groups, among which group A was mainly responsible for transcriptional regulation and various stress responses [33].
According to the clustering results of the evolutionary tree, HpbZIP69 was closely clustered with ABF3 (AtbZIP37) and ABF4/AREB2 (AtbZIP38) which is involved in the regulation of abiotic stress such as drought. Therefore, we speculate that the two may have similar functions. In this study, the tissue culture seedlings of H. perforatum at two months of seedling age were treated with drought. qRT-PCR results showed that the expression levels of HpbZIP69 were significantly increased under drought treatment, suggesting that it may be involved in regulating drought stress. HpbZIP69 has cis-acting elements that respond to drought, abscisic acid, and excessive salt stimulation, which can promote transcriptional activation of genes related to the stress response. Therefore, the expression patterns of HpbZIP69 under drought, abscisic acid, and high salt treatment were analyzed simultaneously. The results showed that: The gene expression levels of HpbZIP69 increased after these treatments at different times, but the gene expression levels were different in time and space. We also constructed pEarleyGate103 (CD3-685) subcellular localization vector. We found that HpbZIP69 was only expressed in the nucleus through transient expression of the target gene in onion with the help of gene gun transformation technology. According to the results of transcriptional self-activation and yeast single hybridization experiments, we found that HpbZIP69 has self-activation activity and can interact with the G-box motif.
In this investigation, HpbZIP69 were found to be resistant to drought stress to a certain extent, but the mechanism through which they respond to drought stress remains to be further studied. It has been reported that group A members in A. thaliana, such as ABF3 (AtbZIP37) and ABF4/AREB2 (AtbZIP38) have the strongly transcriptional regulated ability. Meanwhile, it can control the transcriptional activity of essential enzyme genes in the drought response pathway by binding cis-acting elements on promotors, participating in stress response process. Analysis of cis-acting element in this study showed that the cis-acting element AREB was closely related to stress regulation. Subsequent experiments could explore whether the HpbZIP69 transcription factors bind to promoters and are pertaining to ABA-mediated transcriptional activation. In addition, ABA-mediated transcriptional activity is related to flavonoid synthesis [34], so whether HpbZIP69 plays a role in the flavonoid metabolism pathway can be explored, and the drought response mechanism of HpbZIP69 can be further elucidated.
Previous studies have reported that ABF3 (AtbZIP37) and ABF4/AREB2 (AtbZIP38) can regulate the transcriptional activation of essential enzyme genes in the stress response pathway in A. thaliana [18, 31], thereby responding to the stress response. Based on the phylogenetic analysis of H. perforatum and A. thaliana, it was found that HpbZIP69 was closely clustered with AtbZIP37/38, suggesting that it may have a similar function in response to drought. In this study, wild-type and overexpressing transgenic Arabidopsis lines were subjected to both control conditions and drought treatments. Phenotypic analysis showed that the overall growth of OE-HpbZIP69 lines was better than that of the wild type. We also determined the physiological indexes related to drought resistance. The results showed that the MDA content [35], H2O2 content [36], and reactive oxygen species content of OE-HpbZIP69 lines were significantly lower than those of wild type, and the activity of superoxide dismutase (SOD) [37]was considerably higher than that of the wild type under the drought stress. Based on the above results, we believe that HpbZIP69 can enhance the drought resistance of plants, suggesting that it plays an influential role in regulating plant drought resistance.
4. Conclusions
In conclusion, the bZIP TF is of vital for the growth and development of H. perforatum. Based on the phylogenetic, gene structure analyses, stress and hormone-related cis-acting elements, and expression patterns in different tissue and under abiotic stresses of H. perforatum bZIP TFs were analyzed by bioinformatics and qRT-PCR. Most H. perforatum bZIP TFs may be involved in plenty of abiotic stress responses. Overexpression of HpbZIP69 enhanced drought tolerance in Arabidopsis. However, the function of HpbZIP69 in H. perforatum are supposed to be proved experimentally in further research.
5. Materials and Methods
5.1. Identification and sequence analysis
The typical protein sequences of Arabidopsis and rice downloaded from TAIR (http://www.Arabidopsis.org/) [38], and RGAP (http://rice.plantbiology.msu.edu/) were used as a query to probe in H. perforatum genome assembly of our lab, using hmmer3.1 [39, 40] and Pfam ( http://pfam.sanger.ac.uk/ ) [41] . The E value was set to be one during comparison. The initial sequences were searched in Conserved Domains Search (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) [42]. After deleting repeated and incomplete gene sequences, the remaining ones containing bZIP domains were considered H. perforatum bZIP family members. The arrangements were named in order they be screened out from the library. Every HpbZIP protein sequence was uploaded to ExPASy ( https://web.expasy.org/protparam/) [43] to figure the amino acid quantity, molecular weight, and theoretical iso-electric point. The conserved motifs in bZIP proteins were analyzed utilizing the MEME (https://meme-suite.org/meme/tools/meme) [44]. The e-values of motifs less than 1e-10 were kept for subsequent analysis.
5.2. Phylogenetic analysis, cis-acting element predictions, and Gene Ontology annotations
Protein sequences of bZIPs from H. perforatum and Arabidopsis were used for phylogenetic analysis, which were aligned by Clustal X2.0.8. The unrooted phylogenetic trees were generated using MEGA 7.0 with the neighbor-joining method. In parameter settings, the gaps/missing data treatment was set to pairwise deletion, and the model developed to Poisson Model. The bootstrap test was installed as 1000 times, and checked other parameters to default values. To predict and classify the promoter cis-acting element composition, the 1.5kb upstream sequence of each HpbZIP from ATG was uploaded to Bioedit and PlantCARE (http://bioinformatics.psd.ugent.be/webtools/plantcare/html/) [45]. The protein sequences were matched to the NCBI non-redundant protein database with Blast2GO [46] to acquire a Gene Ontology (GO) annotation for each one of HpbZIPs. Furthermore, WEGO software [47] was applied to generate GO functional classifications and the allocation of gene functions at the macro level.
5.3. Plant materials, stress treatments, and expression analysis
Seeds of H. perforatum purchased from Gansu Province. The seeding methods and cultivation conditions of two-month and two-year seedlings are mentioned in the previous description [48]. The 2-month-old seedlings were used for stress and induction related expression profile analysis. For drought and high salinity treatment, 20% PEG6000 and 200 mM NaCl solution was used to treat seedlings. For hormone induction, the seedlings were sprayed with ABA with concentration of 100 mM. The above samples were obtained at 0h, 1h, 3h, 6h, and 12h after each stress treatment, then stored at -80 ℃ until used. Tissue samples (stem, root, leaf, and flower) were collected from 2-year-old plants were used for RNA-sequencing. The resulting data were normalized using TBtools v 0.58 [49] to generate a heatmap. Other samples were used for real-time quantitative PCR (RT-qPCR) analysis.
The seeds of Arabidopsis were surface-sterilized and sown in 1/2 MS medium containing 2% sucrose. Plants were grown in growth chambers with a long-day condition (16-h light and 8-h darkness) at 22 °C. After ten days, different lines were transferred to the soil. In the experimental group, 4-week-old seedlings were treated for 15 days without watering. The control group was watered once every 3 d. After 15 days of treatment, the phenotypes of these three Arabidopsis were observed. Three replicates were conducted, and each replicate contained at least 9 seedlings.
Total RNA was extracted by HiPure HP Plant RNA Mini Kit (Magen, Guangzhou, China). cDNA was synthesized by Prime Script RT Master Mix (Takara, Dalian, China.). The acquired cDNA was diluted 10-fold. RT-qPCR was carried out on the Roche Light Cycler 96 system (Roche Diagnostics GmbH) using the SYBR® qPCR Master Mix (Takara, Dalian, China) [50]. Technical and biological replicates were repeated three times. All the primers checked by Primer-BLAST (Supplemental Table 1) were designed using Genscript. The relative expression levels of HpbZIPs were figured in the 2−ΔΔCt method [51]. HpActin2 (MK054303) was screened out as the internal reference [52].
5.4. Subcellular localization of the HpbZIP69
The whole-length coding nucleotide sequences of HpbZIP69 were cloned into the vector pDONR207 using a BP reaction of the Gateway (Invitrogen, Carlsbad, US) [53]. The pDONR207-HpbZIP69 were recombined into the destination vector pEarleyGate103 (CD3-685) to obtain subcellular localization expression. All primers used in the experiment are shown in Supplementary Table 1. Meanwhile, pEarleyGate103 was used as a positive control. The vectors were bombarded with the gene gun as the method we described before [54]. The images were shot in 475 nm from living onion cells were recorded by Leica DM6000B microscope (Leica, Germany) after incubation on solid MS medium at 28 °C for 24 h in the dark.
5.5. Transcriptional Activation and Y1H Assay
To experiment the transcriptional activity of HpbZIP69, the sequence with full-length coding region (374 amino acids) and fragmentary sequences containing 1-298 amino acids (N-terminal), 1-319 amino acids, 229-374 amino acids, and 320-374 amino acids (C-terminal) were inserted into the pGBKT7, separately. The recombinant vectors and the empty pGBKT7 (negative control) were transferred into the AH109 yeast-competent cells (Weidi, Shanghai, China). The transformants were grew on SD/-Trp and SD/-Trp/-Ade/-His/X-α-gal deficiency media. Then the transcriptional activities were tested according to their growth status at 29 ℃ for three days in darkness. Since the promoter sequence of HpASMT2 contains four G-box elements, it was constructed on the pHis2 vector as a reporter vector. The pGADT7-HpbZIP69 was constructed as an effector vector. All recombinant plasmids were transformed into yeast competent cells Y187 (Weidi, Shanghai, China) and screened on SD/-Trp /-Leu medium at 29 °C for 48-72 h. Then transferred the surviving colonies to SD/-Trp/-Leu/-His mediums with and without 60 mM 3-AT (3-amino-1,2,4-triazole, Coolaber, Beijing, China). The variety of p53His and pGADT7-p53 vectors served as positive controls, while the combination of p53His and pGADT7 served as negative controls. The interaction between HpbZIP69 and G-box motif was valued by the transformants’ growth status. The primers used in the experiment are listed in Supplementary Table 1.
5.6. Overexpressing HpbZIP69 in Arabidopsis
The coding sequence of HpbZIP69 containing attB and attP sites was inserted into pDONR207 by BP reaction, according to the principle of Gateway (Invitrogen, Carlsbad, United States). Then, pEarleyGate202-HpbZIP69 was constructed by LR reaction. The constructs, 35S:: HpbZIP69 and empty pEarleyGate202, were transferred into Agrobacterium tumefaciens strain GV3101 (Weidi, Shanghai, China). Transgenic Arabidopsis lines were obtained by Agrobacterium-mediated transformation method [55] and screening of corresponding antibiotics. They were detected and identified at DNA level and RNA level, respectively. Homozygous T3 transgenic lines were detected by semi-quantitative PCR as previously [54], and the seedlings used for subsequent analysis. The primer sequences can be found in Supplementary Table 1.
5.7. Physiological index measurement
One-month-old WT and OE field plants were analyzed for drought resistance. Malondialdehyde (MDA) and Hydrogen Peroxide (H2O2) concentration was measured using the MDA Assay Kit and H2O2 Assay Kit (Solarbio, Beijing, China). The Superoxide dismutase (SOD) level was measured using a SOD Assay kit (Solarbio, Beijing, China), and then examined for values by Multiskan FC microplate photometer (Thermo Fisher, US), with three biological and three technical replicates. ANOVA analysis was used for statistical analysis, and the probability value p< 0.05 was considered statistically significant.
Supplementary Materials
Figure S1: Structure of representative HpbZIP proteins from each subgroup. Figure S2: Seqlogo of conserved motifs in HpbZIP proteins. Figure S3: Cis-Acting elements analysis of HpbZIP gene promoters. The cis-acting elements were identified by the online PlantCARE program using 1.5 kb upstream of transcription initiation site of HpbZIP genes. The graph was generated based on the presence of cis-acting elements in response to specific processes /elicitors/conditions (x-axis) in HpbZIP family members (y-axis). Figure S4: Gene Ontology annotation results for HpbZIP proteins. The y-axis on the left side indicates the percentage of a specific category of genes in the main category, and the right side indicates the number of genes in a category. Figure S5: Predicted subcellular localization of HpbZIP69 using WoLFPSOR (https://wolfpsort.hgc.jp/). Table S1: Primers were used in this study. Table S2: Analysis of the physicochemical properties of the bZIP gene family of Hypericum perforatum. Table S3:Typical expression sequences of 15 motifs identified in HpbZIP genes.Table S4: Putative cis-acting elements were identified in the promoter regions of 79 HpbZIP genes.
Author Contributions
Conceptualization, R.X. and S.Y.; methodology, Y.S. and R.X.; validation, W.Z., D.W. and Z.W.; formal analysis, Y.S.; investigation, R.X. and S.Y.; writing—original draft preparation, W.Z.; writing—review and editing, Z.W. and W.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (No. 32100308 and 32170378), the foundation of the Shaanxi Academy of Sciences (No. 2022K-15).
Conflicts of Interest
The authors declare no conflict of interest..
References
- Hurst, H.C. Transcription factors 1: bZIP proteins. Protein Profile 1995, 2, 101–168. [Google Scholar]
- Singh, K.; Foley, R.C.; Oñate-Sánchez, L. Transcription factors in plant defense and stress responses. Curr Opin Plant Biol 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Nijhawan, A.; Jain, M.; Tyagi, A.K.; Khurana, J.P. Genomic survey and gene expression analysis of the basic leucine zipper transcription factor family in rice. Plant physiology 2008, 146, 333–350. [Google Scholar] [CrossRef]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends in plant science 2002, 7, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Landschulz, W.H.; Johnson, P.F.; McKnight, S.L. The leucine zipper: a hypothetical structure common to a new class of DNA binding proteins. Science 1988, 240, 1759–1764. [Google Scholar] [CrossRef] [PubMed]
- Izawa, T.; Foster, R.; Chua, N.H. Plant bZIP protein DNA binding specificity. Journal of molecular biology 1993, 230, 1131–1144. [Google Scholar] [CrossRef] [PubMed]
- Ellenberger, T.E.; Brandl, C.J.; Struhl, K.; Harrison, S.C. The GCN4 basic region leucine zipper binds DNA as a dimer of uninterrupted alpha helices: crystal structure of the protein-DNA complex. Cell 1992, 71, 1223–1237. [Google Scholar] [CrossRef]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription Factors Associated with Abiotic and Biotic Stress Tolerance and Their Potential for Crops Improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.X.; Srivastava, R.; Howell, S.H. Stress-induced expression of an activated form of AtbZIP17 provides protection from salt stress in Arabidopsis. Plant Cell Environ 2008, 31, 1735–1743. [Google Scholar] [CrossRef]
- Weltmeier, F.; Ehlert, A.; Mayer, C.S.; Dietrich, K.; Wang, X.; Schütze, K.; Alonso, R.; Harter, K.; Vicente-Carbajosa, J.; Dröge-Laser, W. Combinatorial control of Arabidopsis proline dehydrogenase transcription by specific heterodimerisation of bZIP transcription factors. Embo j 2006, 25, 3133–3143. [Google Scholar] [CrossRef]
- Hartmann, L.; Pedrotti, L.; Weiste, C.; Fekete, A.; Schierstaedt, J.; Göttler, J.; Kempa, S.; Krischke, M.; Dietrich, K.; Mueller, M.J.; Vicente-Carbajosa, J.; Hanson, J.; Dröge-Laser, W. Crosstalk between Two bZIP Signaling Pathways Orchestrates Salt-Induced Metabolic Reprogramming in Arabidopsis Roots. The Plant cell 2015, 27, 2244–2260. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chu, Z. Genome-wide evolutionary characterization and analysis of bZIP transcription factors and their expression profiles in response to multiple abiotic stresses in Brachypodium distachyon. BMC genomics 2015, 16, 227. [Google Scholar] [CrossRef]
- Liu, C.; Mao, B.; Ou, S.; Wang, W.; Liu, L.; Wu, Y.; Chu, C.; Wang, X. OsbZIP71, a bZIP transcription factor, confers salinity and drought tolerance in rice. Plant molecular biology 2014, 84, 19–36. [Google Scholar] [CrossRef]
- Cai, W.; Yang, Y.; Wang, W.; Guo, G.; Liu, W.; Bi, C. Overexpression of a wheat (Triticum aestivum L.) bZIP transcription factor gene, TabZIP6, decreased the freezing tolerance of transgenic Arabidopsis seedlings by down-regulating the expression of CBFs. Plant Physiol Biochem 2018, 124, 100–111. [Google Scholar] [CrossRef]
- Wang, L.; Cao, H.; Qian, W.; Yao, L.; Hao, X.; Li, N.; Yang, Y.; Wang, X. Identification of a novel bZIP transcription factor in Camellia sinensis as a negative regulator of freezing tolerance in transgenic arabidopsis. Annals of botany 2017, 119, 1195–1209. [Google Scholar] [CrossRef]
- Yao, T.; Zhang, J.; Xie, M.; Yuan, G.; Tschaplinski, T.J.; Muchero, W.; Chen, J.G. Transcriptional Regulation of Drought Response in Arabidopsis and Woody Plants. Front Plant Sci 2020, 11, 572137. [Google Scholar] [CrossRef]
- Hossain, M.A.; Cho, J.I.; Han, M.; Ahn, C.H.; Jeon, J.S.; An, G.; Park, P.B. The ABRE-binding bZIP transcription factor OsABF2 is a positive regulator of abiotic stress and ABA signaling in rice. J Plant Physiol 2010, 167, 1512–1520. [Google Scholar] [CrossRef]
- Fujita, Y.; Fujita, M.; Satoh, R.; Maruyama, K.; Parvez, M.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in Arabidopsis. The Plant cell 2005, 17, 3470–3488. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zhou, J.; Jie, Y.; Xing, H.; Zhong, Y.; Yu, W.; She, W.; Ma, Y.; Liu, Z.; Zhang, Y. A Ramie bZIP Transcription Factor BnbZIP2 Is Involved in Drought, Salt, and Heavy Metal Stress Response. DNA and cell biology 2016, 35, 776–786. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, L.; Xia, C.; Gao, L.; Hao, C.; Zhao, G.; Jia, J.; Kong, X. A Novel Wheat C-bZIP Gene, TabZIP14-B, Participates in Salt and Freezing Tolerance in Transgenic Plants. Front Plant Sci 2017, 8, 710. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xu, K.; Chen, S.; Li, T.; Xia, H.; Chen, L.; Liu, H.; Luo, L. A stress-responsive bZIP transcription factor OsbZIP62 improves drought and oxidative tolerance in rice. BMC Plant Biol 2019, 19, 260. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhu, J.; Yuan, W.; Wang, Y.; Hu, P.; Jiao, C.; Xia, H.; Wang, D.; Cai, Q.; Li, J.; Wang, C.; Zhang, X.; Chen, Y.; Wang, Z.; Ou, Z.; Xu, Z.; Shi, J.; Chen, J. Genome-wide characterization of bZIP transcription factors and their expression patterns in response to drought and salinity stress in Jatropha curcas. Int J Biol Macromol 2021, 181, 1207–1223. [Google Scholar] [CrossRef]
- Wang, B.; Li, L.; Peng, D.; Liu, M.; Wei, A.; Li, X. TaFDL2-1A interacts with TabZIP8-7A protein to cope with drought stress via the abscisic acid signaling pathway. Plant science : an international journal of experimental plant biology 2021, 311, 111022. [Google Scholar] [CrossRef]
- Rolly, N.K.; Imran, Q.M.; Shahid, M.; Imran, M.; Khan, M.; Lee, S.U.; Hussain, A.; Lee, I.J.; Yun, B.W. Drought-induced AtbZIP62 transcription factor regulates drought stress response in Arabidopsis. Plant Physiol Biochem 2020, 156, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Zavaliev, R.; Wu, Q.; Zhou, Y.; Cheng, J.; Dillard, L.; Powers, J.; Withers, J.; Zhao, J.; Guan, Z.; Borgnia, M.J.; Bartesaghi, A.; Dong, X.; Zhou, P. Structural basis of NPR1 in activating plant immunity. Nature 2022, 605, 561–566. [Google Scholar] [CrossRef]
- Fan, W.; Dong, X. In vivo interaction between NPR1 and transcription factor TGA2 leads to salicylic acid-mediated gene activation in Arabidopsis. The Plant cell 2002, 14, 1377–1389. [Google Scholar] [CrossRef]
- Zhang, Y.; Tessaro, M.J.; Lassner, M.; Li, X. Knockout analysis of Arabidopsis transcription factors TGA2, TGA5, and TGA6 reveals their redundant and essential roles in systemic acquired resistance. The Plant cell 2003, 15, 2647–2653. [Google Scholar] [CrossRef]
- Zhang, R.; Ji, Y.; Zhang, X.; Kennelly, E.J.; Long, C. Ethnopharmacology of Hypericum species in China: A comprehensive review on ethnobotany, phytochemistry and pharmacology. J Ethnopharmacol 2020, 254, 112686. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Lakra, N.; Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. A unique bZIP transcription factor imparting multiple stress tolerance in Rice. Rice (New York, N.Y.) 2019, 12, 58. [Google Scholar] [CrossRef]
- Dröge-Laser, W.; Snoek, B.L.; Snel, B.; Weiste, C. The Arabidopsis bZIP transcription factor family-an update. Curr Opin Plant Biol 2018, 45, 36–49. [Google Scholar] [CrossRef]
- Singh, D.; Laxmi, A. Transcriptional regulation of drought response: a tortuous network of transcriptional factors. Front Plant Sci 2015, 6, 895. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, Z.; Ren, W.; Yan, S.; Xing, N.; Zhang, Z.; Li, H.; Ma, W. Identification of the bZIP gene family and regulation of metabolites under salt stress in isatis indigotica. Front Plant Sci 2022, 13, 1011616. [Google Scholar] [CrossRef]
- Que, F.; Wang, G.L.; Huang, Y.; Xu, Z.S.; Wang, F.; Xiong, A.S. Genomic identification of group A bZIP transcription factors and their responses to abiotic stress in carrot. Genetics and molecular research : GMR 2015, 14, 13274–13288. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, Y.; Xu, C.; Wang, X.; Wang, P.; Huang, S. ABA collaborates with lignin and flavonoid to improve pre-silking drought tolerance by tuning stem elongation and ear development in maize (Zea mays L.). The Plant journal : for cell and molecular biology 2023.
- Zhou, Q.; Li, Y.; Wang, X.; Yan, C.; Ma, C.; Liu, J.; Dong, S. Effects of Different Drought Degrees on Physiological Characteristics and Endogenous Hormones of Soybean. Plants (Basel) 2022, 11, 2282. [Google Scholar] [CrossRef]
- Li, L.; Yi, H. Enhancement of drought tolerance in Arabidopsis plants induced by sulfur dioxide. Ecotoxicology (London, England) 2022, 31, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Saed-Moucheshi, A.; Sohrabi, F.; Fasihfar, E.; Baniasadi, F.; Riasat, M.; Mozafari, A.A. Superoxide dismutase (SOD) as a selection criterion for triticale grain yield under drought stress: a comprehensive study on genomics and expression profiling, bioinformatics, heritability, and phenotypic variability. BMC Plant Biol 2021, 21, 148. [Google Scholar] [CrossRef] [PubMed]
- Reiser, L.; Subramaniam, S.; Zhang, P.; Berardini, T. Using the Arabidopsis Information Resource (TAIR) to Find Information About Arabidopsis Genes. Current protocols 2022, 2, e574. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic acids research 2018, 46, W200–w204. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, Y.; Li, B.; Petijová, L.; Hu, S.; Zhang, Q.; Niu, J.; Wang, D.; Wang, S.; Dong, Y.J.J. o. P. R. Whole-genome sequence data of Hypericum perforatum and functional characterization of melatonin biosynthesis by N-acetylserotonin O-methyltransferase. 2021, 70, e12709.
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; Finn, R.D.; Bateman, A. Pfam: The protein families database in 2021. Nucleic acids research 2021, 49, D412–d419. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; Thanki, N.; Yamashita, R.A.; Yang, M.; Zhang, D.; Zheng, C.; Lanczycki, C.J.; Marchler-Bauer, A. CDD/SPARCLE: the conserved domain database in 2020. Nucleic acids research 2020, 48, D265–d268. [Google Scholar] [CrossRef]
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, the Swiss Bioinformatics Resource Portal, as designed by its users. Nucleic acids research 2021, 49, W216–w227. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic acids research 2015, 43, W39–49. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic acids research 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S. Blast2GO: A comprehensive suite for functional analysis in plant genomics. International journal of plant genomics 2008, 2008, 619832. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zhang, Y.; Cui, H.; Liu, J.; Wu, Y.; Cheng, Y.; Xu, H.; Huang, X.; Li, S.; Zhou, A.; Zhang, X.; Bolund, L.; Chen, Q.; Wang, J.; Yang, H.; Fang, L.; Shi, C. WEGO 2.0: a web tool for analyzing and plotting GO annotations, 2018 update. Nucleic acids research 2018, 46, W71–W75. [Google Scholar] [CrossRef]
- Zhou, W.; Zhang, Q.; Sun, Y.; Yang, L.; Wang, Z. Genome-wide identification and characterization of R2R3-MYB family in Hypericum perforatum under diverse abiotic stresses. Int J Biol Macromol 2020, 145, 341–354. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Zhou, W.; Yang, S.; Yang, L.; Xiao, R.; Chen, S.; Wang, D.; Wang, S.; Wang, Z.J.I.J. o. M. S. Genome-Wide Identification of the Hypericum perforatum WRKY Gene Family Implicates HpWRKY85 in Drought Resistance. 2023, 24, 352.
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2ˆ(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostatistics, bioinformatics and biomathematics 2013, 3, 71–85. [Google Scholar]
- Zhou, W.; Wang, S.; Yang, L.; Sun, Y.; Zhang, Q.; Li, B.; Wang, B.; Li, L.; Wang, D.; Wang, Z.J.P. Reference genes for qRT-PCR normalisation in different tissues, developmental stages, and stress conditions of Hypericum perforatum. 2019, 7, e7133.
- Reece-Hoyes, J.S.; Walhout, A.J.M. Gateway Recombinational Cloning. Cold Spring Harbor protocols 2018, 2018, pdb–top094912. [Google Scholar] [CrossRef]
- Zhou, W.; Yang, S.; Zhang, Q.; Xiao, R.; Li, B.; Wang, D.; Niu, J.; Wang, S.; Wang, Z. Functional Characterization of Serotonin N-Acetyltransferase Genes (SNAT1/2) in Melatonin Biosynthesis of Hypericum perforatum. Front Plant Sci 2021, 12, 781717. [Google Scholar] [CrossRef]
- Zhang, X.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nature protocols 2006, 1, 641–646. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic analysis and subgroup classification of HpbZIP proteins. The phylogenetic tree of bZIP proteins was performed based on the homologous proteins in Arabidopsis. The 79 HpbZIP proteins and 75 AtbZIP proteins were categorized into ten groups (A through I, and S). The neighbor-joining tree was performed in MEGA 7.0 software.
Figure 1.
Phylogenetic analysis and subgroup classification of HpbZIP proteins. The phylogenetic tree of bZIP proteins was performed based on the homologous proteins in Arabidopsis. The 79 HpbZIP proteins and 75 AtbZIP proteins were categorized into ten groups (A through I, and S). The neighbor-joining tree was performed in MEGA 7.0 software.
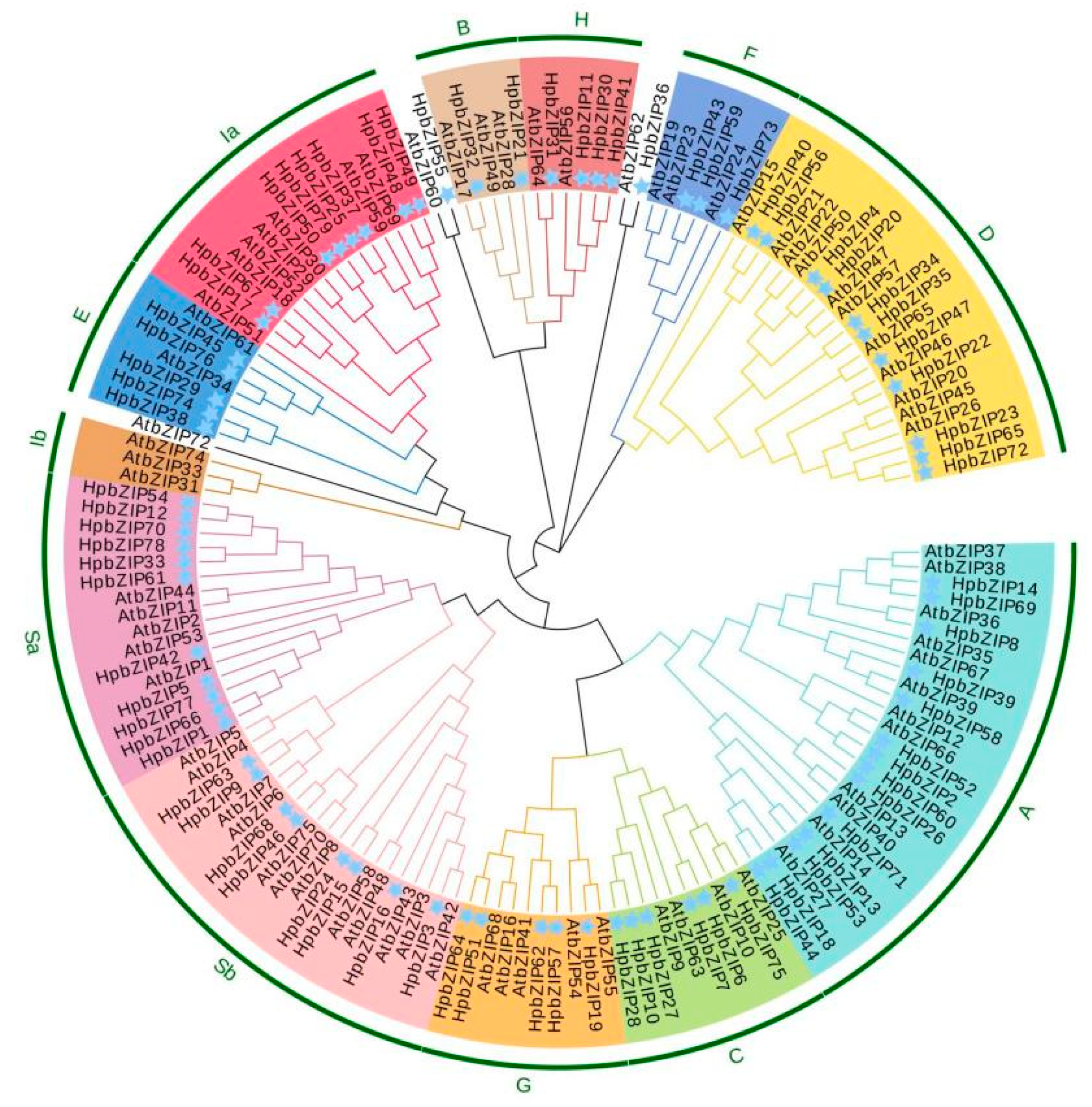
Figure 2.
Phylogenetic trees, conserved motifs, and gene structures of HpbZIP genes. In the left part, Different subgroups were indicated with various background colors. In the middle part, boxes represent motif distributions. In the right position, green boxes, grey lines, and yellow boxes represent exons, introns, and UTRs, respectively. The results were displayed using TB tools.
Figure 2.
Phylogenetic trees, conserved motifs, and gene structures of HpbZIP genes. In the left part, Different subgroups were indicated with various background colors. In the middle part, boxes represent motif distributions. In the right position, green boxes, grey lines, and yellow boxes represent exons, introns, and UTRs, respectively. The results were displayed using TB tools.
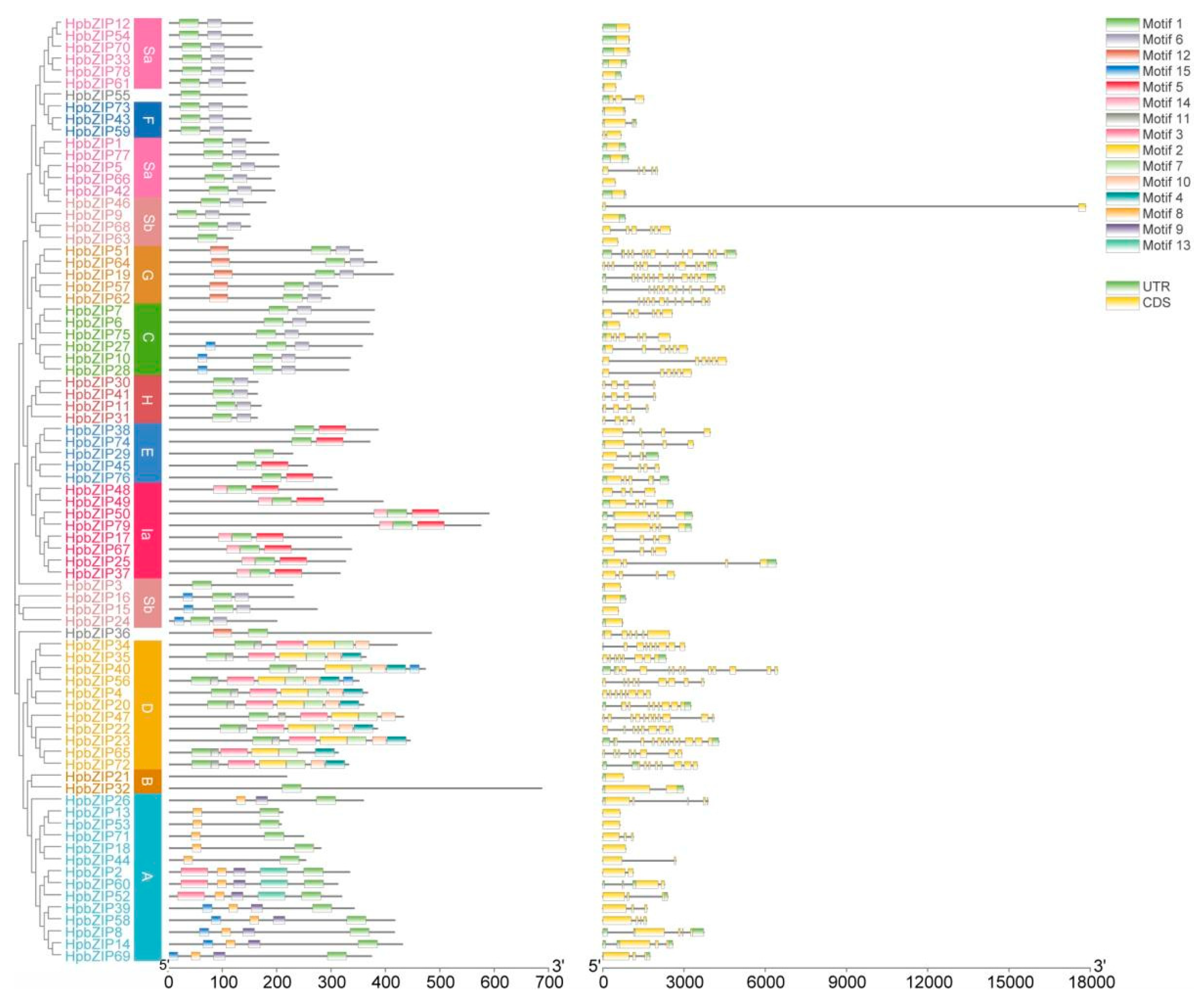
Figure 3.
Expression profiles of HpbZIP genes. Hierarchical clustering of expression profiles of 79 HpbZIP genes in different tissues (root, stem, leaf, and flower).
Figure 3.
Expression profiles of HpbZIP genes. Hierarchical clustering of expression profiles of 79 HpbZIP genes in different tissues (root, stem, leaf, and flower).
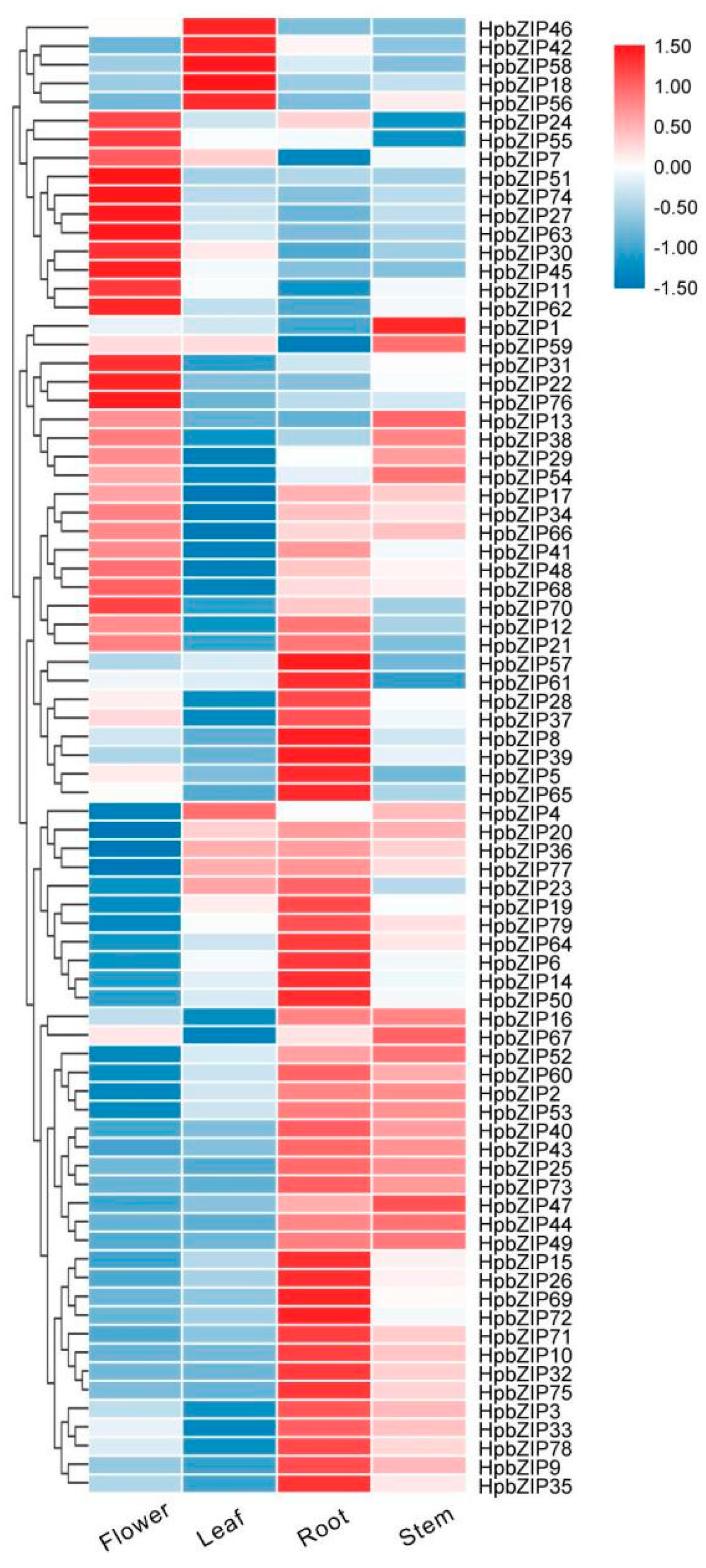
Figure 4.
Expression analysis of 12 representative HpbZIP genes under ABA induction (A), NaCl (B), and PEG (C)treatments. Data were normalized to HpActin2, and vertical bars indicate standard deviation. Each treatment contains three biological replicates. Asterisks indicate significant differences compared with the untreated control. Data are presented as Mean ± SD (n = 3) (*: p<0.05; **: p<0.01; ***: p<0.001, One-way ANOVA test).
Figure 4.
Expression analysis of 12 representative HpbZIP genes under ABA induction (A), NaCl (B), and PEG (C)treatments. Data were normalized to HpActin2, and vertical bars indicate standard deviation. Each treatment contains three biological replicates. Asterisks indicate significant differences compared with the untreated control. Data are presented as Mean ± SD (n = 3) (*: p<0.05; **: p<0.01; ***: p<0.001, One-way ANOVA test).
Figure 5.
HpbZIP69 is a representative bZIP transcription factor. (A) The subcellular location of HpbZIP69. Scale bars: 30 µm. (B) Transactivation activity test in the yeast GAL4 system and the protein structure of HpbZIP69. (C) Yeast-one-hybrid assay reveals that HpbZIP69 can bind to the G-box motif.
Figure 5.
HpbZIP69 is a representative bZIP transcription factor. (A) The subcellular location of HpbZIP69. Scale bars: 30 µm. (B) Transactivation activity test in the yeast GAL4 system and the protein structure of HpbZIP69. (C) Yeast-one-hybrid assay reveals that HpbZIP69 can bind to the G-box motif.
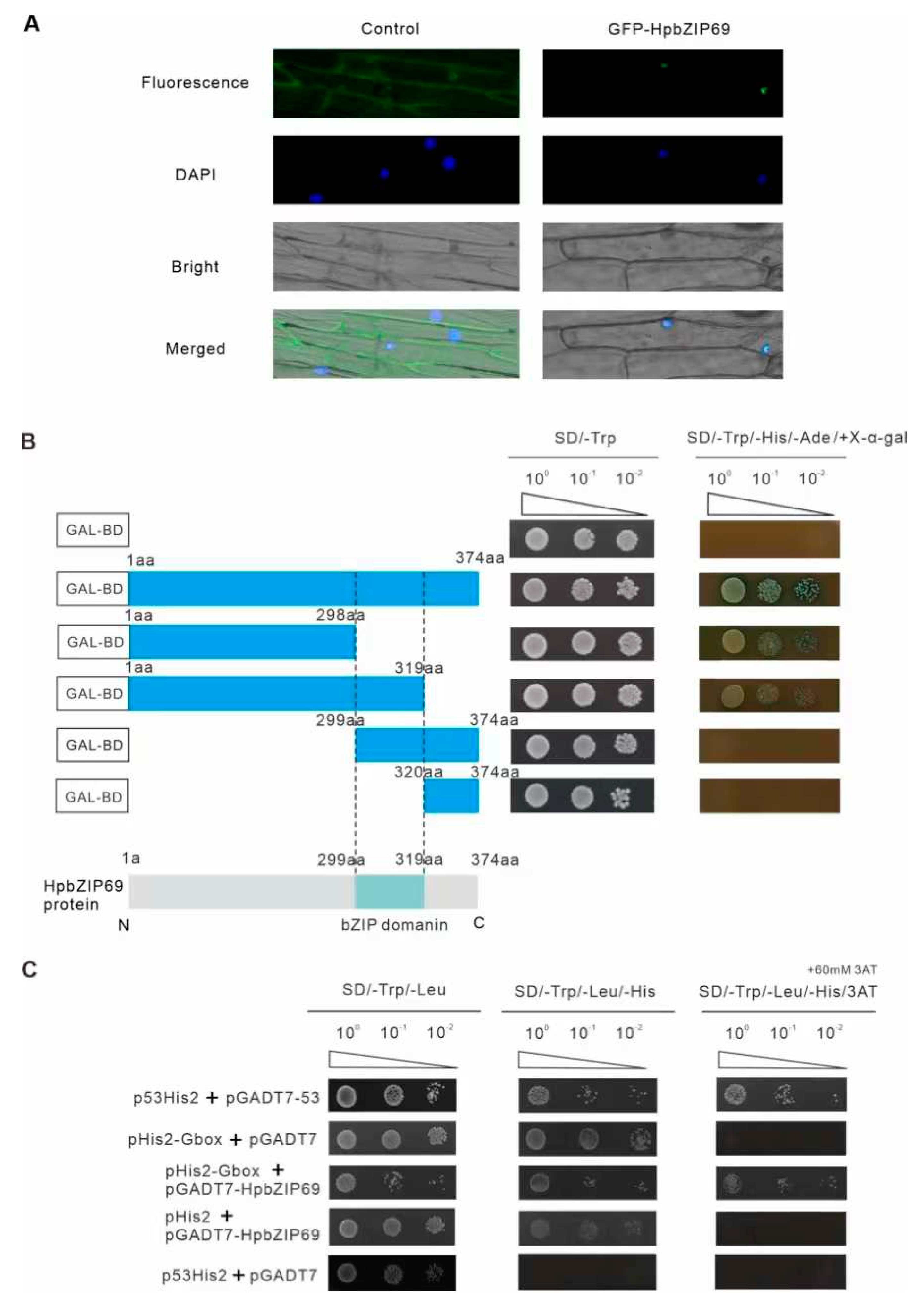
Figure 6.
Function analysis of HpbZIP69 under drought treatment in transgenic Arabidopsis. (A) PCR results of HpbZIP69 genes. HpbZIP69 cannot be cloned from WT lines, but can be cloned from OE transgenic lines. (B) RT-PCR identification of positive transgenic A. thaliana lines. The AtActin2 gene is used as an inner reference. WT line is used as the negative control. (C) Observations of WT and OE seedlings under drought test. (D) The analysis of SOD, H2O2, and MDA content in WT and OE lines in control and drought conditions. Data are the Mean ± SD of three independent biological replicates. (*: p<0.05; One-way ANOVA test).
Figure 6.
Function analysis of HpbZIP69 under drought treatment in transgenic Arabidopsis. (A) PCR results of HpbZIP69 genes. HpbZIP69 cannot be cloned from WT lines, but can be cloned from OE transgenic lines. (B) RT-PCR identification of positive transgenic A. thaliana lines. The AtActin2 gene is used as an inner reference. WT line is used as the negative control. (C) Observations of WT and OE seedlings under drought test. (D) The analysis of SOD, H2O2, and MDA content in WT and OE lines in control and drought conditions. Data are the Mean ± SD of three independent biological replicates. (*: p<0.05; One-way ANOVA test).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



