
Submitted:
21 July 2023
Posted:
27 July 2023
You are already at the latest version
Abstract
Background: This review aims to explore the regulatory mechanism of microglial exosomes, particularly focusing on the role of miR-124-3p, in modulating neuroinflammation and promoting neuronal repair following traumatic brain injury (TBI). Methods: Multiple studies investigating the impact of microglial exosomal miRNAs, specifically miR-124-3p, on injured neurons and brain microvascular endothelial cells (BMVECs) in the context of TBI were reviewed. Results: Animal models of TBI, in vitro cell culture experiments, RNA sequencing analysis, and functional assays were employed to elucidate the mechanisms underlying the effects of miR-124-3p-loaded exosomes on neuroinflammation and neuronal repair. The increased expression of miR-124-3p in microglial exosomes after TBI inhibited neuronal inflammation and promoted neurite outgrowth. This effect was achieved by promoting the anti-inflammatory M2 polarization of microglia, suppressing mTOR signaling through downregulation of PDE4B, and inducing autophagy in BMVECs. These actions collectively contributed to neuroprotection, reduced blood-brain barrier leakage, and improved neurologic outcomes in animal models of TBI. Conclusion: Microglial exosomes, particularly those carrying miR-124-3p, have emerged as promising candidates for therapeutic interventions in TBI. These exosomes exhibit neuroprotective effects, attenuate neuroinflammation, and promote neuronal repair and plasticity. Targeting miR-124-3p and utilizing microglial exosomes as therapeutic agents hold great potential for future clinical translation in TBI management. However, further research is required to fully elucidate the underlying mechanisms and optimize their delivery strategies for effective treatment in human TBI cases.
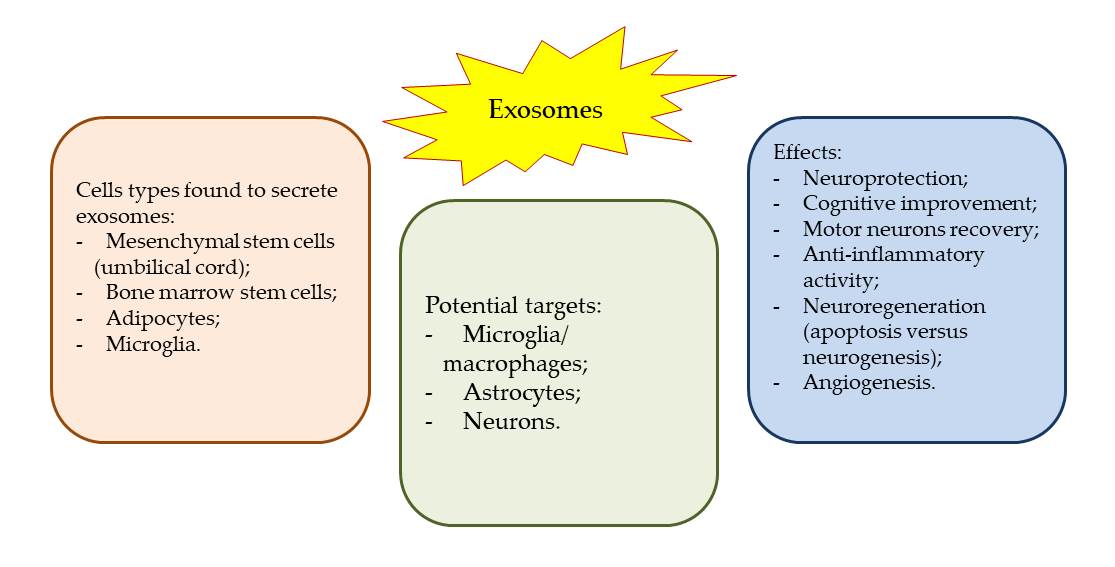
Keywords:
traumatic brain injury
; mesenchymal stem cells derived exosomes
; microglia
; mir-124-3p
1. Introduction
Lately, there is growing interest for the therapeutic potential of exosomes. Recent evidence showed that these small extracellular vesicles play a vital role in intercellular communication by transporting a wide range of bioactive molecules, including proteins, nucleic acids, and lipids [1-3]. The process through which they are generated is based on a complex cellular pathway involving the double invagination of the plasma membrane and the formation of intracellular multivesicular bodies (MVBs) that consist in intraluminal vesicles (ILVs) [4-9]. Initially, the plasma membrane invaginates to form a cup-shaped structure that includes cell surface and soluble proteins associated with the extracellular space. Afterwards the formation of an early-sorting endosome (ESE) is performed by direct merging with a preexisting ESE in some cases while interacting with trans-Golgi network and endoplasmic reticulum. ESEs can mature into late-sorting endosomes, which eventually generate MVBs, also known as multivesicular endosomes. The inward invagination of the endosomal limiting membrane results in the formation of MVBs containing several ILVs that are ultimately secreted as exosomes with a size range of approximately 40 to 160 nm in diameter through MVB fusion to the plasma membrane and exocytosis [5]. As Kalluri et al [4] described, the MVBs degradation can occur via two pathways: lysosomes or autophagosomes-assisted degradation, or plasma membrane resorption with exosomes formation (ILVs release).
Exosomes exhibit heterogeneity that could be attributed to their size, content, functional impact on recipient cells, and cellular origin [7]. Additionally, the microenvironment and biology of cells may affect the content of exosomes and their biological markers. Exosomes can carry membrane proteins, cytosolic and nuclear proteins, extracellular matrix proteins, metabolites, and nucleic acids, such as mRNA, noncoding RNA species, and DNA [10-13]. Although exosomal cargo analyses require large pools of purified exosomes, the abundance of a given cargo in exosomes can differ, as shown by miRNA exosomal cargo [14].
Also, the functional heterogeneity that the exosomes show is associated with the recipient cells due to their variable expression of cell surface receptors, resulting in different effects, such as cell survival, apoptosis, and immunomodulation, among others, in different target cell types. The source organ or tissue of exosomes, including whether they originate from cancer cells [15], could also affect their properties, such as tropism to certain organs and uptake by specific cell types. Therefore, the complexity and heterogeneity of exosomes could be a result of a combination of these features.
Traumatic brain injury (TBI) is a major cause of death and disability worldwide, with no clinically successful treatment available to date [16,17]. Mesenchymal stem cell (MSC)-based therapy has been identified as a promising approach for TBI treatment, but the precise mechanisms underlying their therapeutic effects are still not fully understood [18]. Due to their ability to transfer proteins, lipids, mRNA, miRNA between cells, and promoting cell-to-cell communication, exosomes are regarded as potential therapeutic agents in TBI. Moreover, as exosomes can interact with brain parenchymal cells and the neurogenic niche, leading to neurogenesis and brain remodeling, the implication of exosomes in TBI treatment can be beneficial for cognitive repair. This, we aimed to review the most recent studies investigating the role of exosomes in TBI treatment and highlight the advantages of this approach. We also aimed to discuss the potential limitations of exosome therapy and identify areas for further research. Understanding the mechanisms underlying exosome-mediated TBI repair could provide new insights into the development of effective therapeutic strategies for TBI.
2. Materials and Methods
2.1. Literature search
A comprehensive literature search was conducted using electronic databases such as PubMed, Scopus, and Web of Science. The keywords used were "exosomes", "microvesicles", "extracellular vesicles", "traumatic brain injury", "concussion", and "post-concussion syndrome". The search was limited to articles published in English between 2000 and 2023.
2.2. Inclusion and Exclusion Criteria
Studies were included if they were related to the role of exosomes in the treatment of traumatic brain injury, concussion, or post-concussion syndrome (PCS). Only articles written in English and published in peer-reviewed journals were included in this review. Studies that focused on exosomes in other diseases or injuries were excluded.
2.3. Data Extraction and Analysis
Two reviewers independently extracted data from the selected studies. The extracted data included the author, year of publication, study design, sample size, type of exosomes used, administration route, outcomes measured, and conclusions. The data were analyzed qualitatively.
2.4. Data Synthesis
The extracted data were synthesized and presented in a narrative format. The findings were discussed in the context of their potential clinical applications and future research directions.
3. Results
According to the latest research of the molecular mechanisms in which exosomes are implicated after TBI, they appear to promote functional recovery and reduce neuronal apoptosis by modulating microglia and astrocytes activation during brain injury. The recent studies that were considered presented a wide range of TBI animal models on which the effects of the exosomes administration was tested. Besides in vivo experiments, in vitro studies had the purpose of describing some of the molecular pathways in which the beneficial effects of exosomes are obtained (Figure 1).
Xian et al. investigated the therapeutic effects of mesenchymal stem cell-derived exosomes (MSC-Exo) on inflammation-induced alterations in astrocytes, which play a key role in maintaining brain homeostasis and responding to injury [19]. It was shown that reactive astrogliosis and inflammatory responses can be attenuated when MSC-Exo were incorporated into hippocampal astrocytes both in vitro and in vivo. In this way, the amelioration of ameliorated learning and memory impairments in mice with inflammation-induced astrocytic activation was obtained. They suggested that the Nrf2-NF-κB signaling pathway is involved in regulating astrocyte activation in mice, and that MSC-Exo may be a promising nanotherapeutic agent for the treatment of neurological diseases with hippocampal astrocyte alterations.
In a weight-drop-induced TBI rat model, Chen et al. obtained functional recovery after injecting human adipose mesenchymal stem cell-derived exosomes intracerebroventricularly [20]. Moreover, the study showed that the beneficial effects of these exosomes were correlated with suppressed neuroinflammation, reduced neuronal apoptosis, and increased neurogenesis [20] and proposed that the mechanisms through which the exosomes act are related to entering microglia/macrophages and suppressing their activation during brain injury.
Liu et al. also explored the potential of mesenchymal stem cells-derived exosomes in TBI treatment [21]. They showed that 3D-printed collagen/silk fibroin/hypoxia-pretreated human umbilical cord mesenchymal stem cells-derived exosomes scaffolds exhibit favorable physical properties that suggest increased biocompatibility and biodegradability in a TBI beagle dog model. Their effects include promoting neuroregeneration and angiogenesis, while inhibiting nerve cell apoptosis and proinflammatory factor expression. Furthermore, they showed that hypoxia-induced exosomes could offer superior biocompatibility and neuroregeneration ability suggesting that hypoxia-driven stress could promote reparatory processes. Similarly, Li et al [22] showed that the recovery of neurological function could be achieved by intracerebroventricular administration of umbilical cord MSCs-derived exosomes and suggested that their mechanism of action is correlated with the suppression of microglia and astrocytes response to brain injury.
Also, Zhang et al showed that exosome treatments improved sensorimotor and cognitive function, reduced hippocampal neuronal cell loss, promoted angiogenesis and neurogenesis, and reduced neuroinflammation in a rat model of TBI [23] and set the scene for a preliminary timeline for the efficiency of exosomes treatment following TBI by suggesting that the administration of exosomal products at 1 day post-TBI was more efficient in offering functional and histological outcomes, as compared to other 2 delayed treatments.
Ghosh et al. discussed the potential of exosome therapies for neural injuries, and highlighted their advantages [24] by stating that human stem cells and MSC-Exo could be a promising approach for neuronal injury healing.
Furthermore, exosome treatment attenuated the severity of neurologic injury and allowed for faster neurologic recovery in a clinically realistic large animal model of TBI and hemorrhagic shock. MSC-Exo were shown to retain some of the characteristics of their parent MSCs, such as immune system modulation, regulation of neurite outgrowth, promotion of angiogenesis, and the ability to repair damaged tissue. In a brain transcriptome study, Williams et al. [25] showed that a single dose of MSC-Exo induces transcriptomic changes that are suggestive for neuroprotection.
Han et al. investigated the effect of administration of multipotent MSC-Exo on functional recovery, neurovascular remodeling, and neurogenesis in a rat model of intracerebral hemorrhage and suggested that MSC-Exo effectively improve functional recovery, possibly by promoting endogenous angiogenesis and neurogenesis [26].
Liu et al. investigated the potential of incorporating bone marrow MSC-Exo into hyaluronan-collagen hydrogel to achieve both mimicking of brain matrix and steady release of exosomes, and thus realizing TBI repair [27]. Their results demonstrated that the combination of exosomes and hydrogel effectively induced angiogenesis and neurogenesis, promoting axonal regeneration, remyelination, synapse formation, and even brain structural remodeling, leading to neurological functional recovery of TBI.
Lu et al. investigated the effect of systemic administration of bone mesenchymal stem cells-derived extracellular vesicles on the loss of motor function after spinal cord injury and examined the potential mechanisms underlying their effects [28]. Their results showed that these extracellular vesicles could reduce brain cell death, enhanced neuronal survival and regeneration, and improved motor function when administered at 30 minutes after the spinal cord injury in the tail vein of the rats. This study was the one that suggested that extracellular vesicles could reduce pericyte migration via downregulation of NF-κB p65 signaling and to further modulate the permeability of the blood-spinal cord barrier.
Zhang et al. investigated the protective effects of human umbilical cord MSC-derived exosomes in both in vivo and in vitro TBI models [29]. Their results revealed that exosome treatment improved neurological function, decreased cerebral edema, attenuated brain lesion after TBI, and suppressed TBI-induced cell death, apoptosis, pyroptosis, and ferroptosis. Additionally, the exosomes activated PINK1/Parkin pathway-mediated mitophagy after TBI.
Cui et al isolated exosomes derived from umbilical cord mesenchymal stem cells and intraventricularly injected them into a rat model of TBI. The results demonstrated that the studied exosomes promoted functional recovery and reduced neuronal apoptosis in TBI rats [30]. Furthermore, the study found that umbilical cord mesenchymal stem cells-derived exosomes inhibited the activation of microglia and astrocytes during brain injury, which may contribute to functional recovery. However, the study failed to demonstrate the modulatory effects of this treatment onto the inflammatory factors that were evaluated from the rats’ plasma. Overall, the findings suggest that umbilical cord mesenchymal stem cells-derived exosomes may have therapeutic potential for TBI by inhibiting microglia and astrocyte activation and promoting functional recovery. These results provide a basis for the development of new therapeutic strategies for central nervous system diseases.
The administration of MSC-Exo within one hour after TBI and hemorrhagic shock has been shown to provide neuroprotection in animal models [31]. However, the underlying mechanisms responsible for the neuroprotective effects are not completely understood. To further investigate this treatment strategy, this study aimed to analyze the transcriptomic changes in the brain associated with the administration of MSC-Exo. Yorkshire swine weighing 40-45 kg were subjected to a severe TBI and hemorrhagic shock. After one hour of shock, animals were randomly assigned to receive either lactated Ringer's or exosomes suspended in lactated Ringer's. Brain swelling and lesion size were assessed after 6 hours of observation, and peri-injured brain tissue was processed for RNA sequencing. Results from high-throughput RNA sequencing data analysis were compared between control and experimental groups. In this experimental context it was demonstrated that a single dose of exosomes could particularly modulate the expression of some genes associated with blood-brain barrier stability, neurogenesis, neuronal development, synaptogenesis, and neuroplasticity and with genes associated with stroke, neuroinflammation, neuroepithelial cell proliferation, and non-neuronal cell proliferation (reactive gliosis). However, further studies are needed to determine the potential of exosomes as a treatment for TBI in humans and the traceability of effects on the transcriptome correlated with neuroprotection, as seen in the animal models.
Regarding the efficiency of exosomes treatment, Williams et al showed that even a single-dose administration could attenuate the effects of neurological injuries by reducing the swelling of the brain lesion and modulating inflammation and apoptosis in a 7-day survival model using a swine animal model of TBI [32]. Also, it was found that this single dose of exosomes could promote neural plasticity that could last as long as 7 days. Specifically, the exosome-treated animals had significantly lower neurologic severity scores, faster neurologic recovery, smaller brain lesion sizes, lower levels of inflammatory markers, and lower levels of apoptotic markers. They also had higher levels of the mediator of neural plasticity, brain-derived neurotrophic factor.
Ge et al. investigated the role of miRNAs in microglial exosomes in regulating post-traumatic neurodegeneration [33]. The study demonstrated that the level of miR-124-3p in microglial exosomes from the injured brain was significantly altered in the acute, sub-acute, and chronic phases after repeated mild TBI. In this way, in a reoccurring scratch-injury model, they showed that miR-124-3p upregulated microglial exosomes could significantly decrease the activity of the pathways associated with neurodegeneration. They proposed that this effect was obtained due to the potential of microglial exosomes to target Rela, an inhibitory transcription factor that is implicated in β-amyloid proteolytic breakdown, thereby a potential inhibitor of β-amyloid pathway abnormalities. Also, in this mice model it was observed that the intravenous administration of microglial exosomes could have significant effects within the brain, meaning that they can pass through the blood-brain barrier – this being an important feature of treatments that target the central nervous system. Additionally, miR-124-3p within the exosomes was transferred into hippocampal neurons and alleviated neurodegeneration by targeting the Rela/ApoE signaling pathway suggesting that these exosomes could also have cognitive effects following repeated mild TBIs.
Huang et al. also used animal models of rodents and repetitive TBI-like injuries to study the neuronal inflammation modulation using microglial exosomes and miRNA activity against injured neurons [34]. In this way, the injured brains of the mice were collected during the acute and the chronic phase of TBI and an extract was used on in vitro cultured microglia. The analysis of the microglial exosomes-derived miRNA showed a significant increase of miR-124-3p levels that lead to neuroinflammation inhibition possibly by promoting the anti-inflamed M2 polarization in microglia and by inhibiting mTOR signaling PDE4B gene expression. Moreover, exosomal miR-124-3p contributed to neurite outgrowth characterized by an increase in the number of neurite branches and total neurite length and a decrease in the expression of neurodegenerative proteins (Aβ-peptide and p-Tau). Furthermore, neuroprotective and anti-inflammatory properties were shown in repetitive TBI design. These results suggest that miR-124-3p containing exosomes could exhibit significant neuroregenerative and anti-inflammatory properties that could provide innovative therapeutic approach for brain injury and other neurological disorders.
In their study, Zhao et al investigated the effects of miR-124-3p on BMVEC function to determine the mechanistic basis of its activity [35]. The bEnd.3 cell scratch wound model was used to simulate TBI-associated endothelial cell injury, and Lipofec-tamine3000 was employed to overexpress miR-124-3p in endothelial cells. The upregulation of miR-124-3p was found to suppress bEnd.3 cell apoptotic death following in vitro scratch injury while promoting the upregulation of the tight junction proteins ZO-1 and occludin in these cells, thereby reducing the degree of BBB leakage. Thus, due to the ability to modulate mTOR signaling and autophagy, it is thought that miR-124-3p exert protective effects against apoptosis. Once again it was shown that miR-124-3p could modulate mTOR signaling and autophagy and thus contributing to their protective properties (Table 1).
4. Discussion
As TBI and its effects, such as PCS and chronic traumatic encephalopathy (CTE), are major public health concerns worldwide, the lack of effective treatments increase their burden on the patients and caregivers [4,5]. The potential therapeutic effects of MSC-derived and microglial exosomes have been widely studied in recent years, with promising results [36-40]. The exosomes potential to be used as therapeutic agents is given by their ability to carry proteins, lipids, mRNA, and miRNA, and to be implicated in the communication between cells [41,42]. Studies have shown that MSC-derived exosomes can contribute to the therapeutic effects of MSCs in TBI and hemorrhagic shock models, promoting neurogenesis and brain remodeling [25,31,32]. Administration of a single dose of exosomes induces transcriptomic changes suggestive of neuroprotection, which can reduce genes associated with stroke, neuroinflammation, and non-neuronal cell proliferation contributing to reactive gliosis. Moreover, MSC-derived exosomes effectively improve functional recovery in a rat model of intracerebral hemorrhage (ICH) by promoting endogenous angiogenesis and neurogenesis. Another study demonstrated that incorporation of MSC-derived exosomes into hyaluronan-collagen hydrogel (DHC-BME) can achieve both mimicking of brain matrix and steady release of exosomes, resulting in TBI repair [26,27,29]. Similarly, microglial exosomes have been found to have a potential therapeutic role in TBI [33,34]. They can help modulate inflammation, reduce oxidative stress, and promote neurogenesis. Microglia is resident immune cells in the central nervous system that play an important role in the pathophysiology of TBI. They can release exosomes that contain cytokines, chemokines, and microRNAs, which can interact with other central nervous system cells to promote neuroprotection and repair. Studies have shown that microglial exosomes can promote neuronal survival and neurite outgrowth, reduce microglial activation and inflammatory responses, and improve cognitive and motor functions after TBI.
Overall, the studies suggest that exosomes derived from MSCs or other stem cells hold promise as a potential therapeutic approach for TBI and SCI, and that exosome-based therapies may have neuroprotective effects, promote endogenous angiogenesis and neurogenesis, and suppress cell death, apoptosis, pyroptosis, and ferroptosis. However, further studies are needed to fully understand the mechanisms involved and to determine the efficacy and safety of these therapies in human clinical trials.
However, it is important to note that the heterogeneity of exosomes can be a significant challenge in their therapeutic applications. Exosomes can have different sizes, contents, and functional impacts on recipient cells, which can result in different therapeutic effects. In addition, the isolation and purification methods of exosomes can affect their quality and quantity, and the potential for contamination with other extracellular vesicles is always present. Therefore, more research is needed to optimize the isolation and characterization methods of exosomes to ensure their therapeutic efficacy and safety in clinical applications.
5. Conclusions
The therapeutic potential of mesenchymal and microglial exosomes in TBI and PCS is promising. They can promote neuroprotection, repair, and regeneration by modulating inflammation, reducing oxidative stress, and promoting neurogenesis. Further research is needed to optimize their isolation and characterization methods and to elucidate their mechanisms of action, but the potential of exosomes as a novel therapeutic strategy for TBI and PCS is significant.
Funding
The authors are thankful to the Romanian Ministry of Research, Innovation and Digitization, within Program 1—Development of the national RD system, Subprogram 1.2—Institutional Performance—RDI excellence funding projects, Contract no. 11PFE/30.12.2021, for financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: biogenesis, biologic function and clinical potential. Cell Biosci. 2018, 9, 19. [Google Scholar] [CrossRef]
- Edgar, J.R. Q&A: What are exosomes, exactly? BMC Biol. 2016, 14, 46. [Google Scholar] [CrossRef]
- Chen, H.; Wang, L.; Zeng, X.; Schwarz, H.; Nanda, H.S.; Peng, X.; Zhou, Y. Exosomes, a New Star for Targeted Delivery. Front. Cell Dev Biol, 2021, 9, 751079. [Google Scholar] [CrossRef]
- Kalluri, R. The biology and function of exosomes in cancer. J. Clin. Invest. 2016, 126, 1208–1215. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol 2018, 19, 213–228. [Google Scholar] [CrossRef]
- McAndrews, K.M.; Kalluri, R. Mechanisms associated with biogenesis of exosomes in cancer. Mol. Cancer, 2019, 18, 52. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol, 2019, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Willms, E.; Cabañas, C.; Mäger, I.; Wood, M.J.A.; Vader, P. Extracellular vesicle heterogeneity: subpopulations, isolation techniques, and diverse functions in cancer progression. Front. Immunol 2018, 9, 738. [Google Scholar] [CrossRef]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef]
- Moore, M.; Sandsmark, D.K. Clinical Updates in Mild Traumatic Brain Injury (Concussion). Neuroimaging Clin N Am. 2023, 33(2), 271–278. [Google Scholar] [CrossRef]
- Mavroudis, I.; Kazis, D.; Chowdhury, R.; Petridis, F.; Costa, V.; Balmus, I.M.; Ciobica, A.; Luca, A.C.; Radu, I.; Dobrin, R.P.; Baloyannis, S. Post-Concussion Syndrome and Chronic Traumatic Encephalopathy: Narrative Review on the Neuropathology, Neuroimaging and Fluid Biomarkers. Diagnostics (Basel). 2022, 12(3), 740. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, S.; Ge, X.; Yin, Z.; Li, M.; Guo, M.; Hu, T.; Han, Z.; Kong, X.; Li, D.; Zhao, J.; Wang, L.; Liu, Q.; Chen, F.; Lei, P. Mesenchymal stromal cell treatment attenuates repetitive mild traumatic brain injury-induced persistent cognitive deficits via suppressing ferroptosis. J Neuroinflammation. 2022, 19(1), 185. [Google Scholar] [CrossRef] [PubMed]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; Gangoda, L.; Mathivanan, S. ExoCarta: A web-based compendium of exosomal cargo. J. Mol. Biol, 2016; 428, 688–692. [Google Scholar] [CrossRef]
- Pathan, M.; Fonseka, P.; Chitti, S.V.; Kang, T.; Sanwlani, R.; Van Deun, J.; Hendrix, A.; Mathivanan, S. Vesiclepedia 2019: A compendium of RNA, proteins, lipids and metabolites in extracellular vesicles. Nucleic Acids Res 2019, 47, D516–D519. [Google Scholar] [CrossRef] [PubMed]
- van Balkom, B.W.; Eisele, A.S.; Pegtel, D.M.; Bervoets, S.; Verhaar, M.C. Quantitative and qualitative analysis of small RNAs in human endothelial cells and exosomes provides insights into localized RNA processing, degradation and sorting. J. Extracell. Vesicles 2015, 4, 26760. [Google Scholar] [CrossRef] [PubMed]
- Lasda, E.; Parker, R. Circular RNAs co-precipitate with extracellular vesicles: A possible mechanism for circRNA clearance. PLOS ONE 2016, 11, e0148407. [Google Scholar] [CrossRef]
- Chevillet, J.R.; Kang, Q.; Ruf, I.K.; Briggs, H.A.; Vojtech, L.N.; Hughes, S.M.; Cheng, H.H.; Arroyo, J.D.; Meredith, E.K.; Gallichotte, E.N.; Pogosova-Agadjanyan, E.L.; Morrissey, C.; Stirewalt, D.L.; Hladik, F.; Yu, E.Y.; Higano, C.S.; Tewari, M. Quantitative and stoichiometric analysis of the microRNA content of exosomes. Proc. Natl. Acad. Sci. 2014, 111, 14888–14893. [Google Scholar] [CrossRef]
- Wen, S.W.; Lima, L.G.; Lobb, R.J.; Norris, E.L.; Hastie, M.L.; Krumeich, S.; Möller, A. Breast cancer-derived exosomes reflect the cell-of-origin phenotype. Proteomics 2019, 19, e1800180. [Google Scholar] [CrossRef]
- Xian, P.; Hei, Y.; Wang, R.; Wang, T.; Yang, J.; Li, J.; Di, Z.; Liu, Z.; Baskys, A.; Liu, W.; Wu, S.; Long, Q. Mesenchymal stem cell-derived exosomes as a nanotherapeutic agent for amelioration of inflammation-induced astrocyte alterations in mice. Theranostics. 2019, 9(20), 5956–5975. [Google Scholar] [CrossRef]
- Chen, Y.; Li, J.; Ma, B.; Li, N.; Wang, S.; Sun, Z.; Xue, C.; Han, Q.; Wei, J.; Zhao, RC. MSC-derived exosomes promote recovery from traumatic brain injury via microglia/macrophages in rat. Aging (Albany NY). 2020, 12(18), 18274–18296. [Google Scholar] [CrossRef]
- Liu, X.; Wang, J.; Wang, P.; Zhong, L.; Wang, S.; Feng, Q.; Wei, X.; Zhou, L. Hypoxia-pretreated mesenchymal stem cell-derived exosomes-loaded low-temperature extrusion 3D-printed implants for neural regeneration after traumatic brain injury in canines. Front Bioeng Biotechnol. 2022, 10, 1025138. [Google Scholar] [CrossRef]
- Li, D.; Huang, S.; Yin, Z.; Zhu, J.; Ge, X.; Han, Z.; Tan, J.; Zhang, S.; Zhao, J.; Chen, F.; Wang, H.; Lei, P. Increases in miR-124-3p in Microglial Exosomes Confer Neuroprotective Effects by Targeting FIP200-Mediated Neuronal Autophagy Following Traumatic Brain Injury. Neurochem Res. 2019, 44(8), 1903–1923. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Chopp, M.; Zhang, ZG.; Mahmood, A.; Xiong, Y. Mesenchymal Stem Cell-Derived Exosomes Improve Functional Recovery in Rats After Traumatic Brain Injury: A Dose-Response and Therapeutic Window Study. Neurorehabil Neural Repair. 2020, 34(7), 616–626. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Garg, S.; Ghosh, S. Cell-Derived Exosome Therapy: A Novel Approach to Treat Post-traumatic Brain Injury Mediated Neural Injury. ACS Chem Neurosci. 2020, 11(14), 2045–2047. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.M.; Dennahy, I.S.; Bhatti, U.F.; Halaweish, I.; Xiong, Y.; Chang, P.; Nikolian, V.C.; Chtraklin, K.; Brown, J.; Zhang, Y.; Zhang, Z.G.; Chopp, M.; Buller, B.; Alam, H.B. Mesenchymal Stem Cell-Derived Exosomes Provide Neuroprotection and Improve Long-Term Neurologic Outcomes in a Swine Model of Traumatic Brain Injury and Hemorrhagic Shock. J Neurotrauma. 2019, 36(1), 54–60. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Seyfried, D.; Meng, Y.; Yang, D.; Schultz, L.; Chopp, M.; Seyfried, D. Multipotent mesenchymal stromal cell-derived exosomes improve functional recovery after experimental intracerebral hemorrhage in the rat. J Neurosurg. 2018, 131(1), 290–300. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, C.; Zhang, Y.; Chen, S.; Ding, J.; Chen, Z.; Wu, K.; Wu, X.; Zhou, T.; Zeng, M.; Wei, D.; Sun, J.; Fan, H.; Zhou, L. Hyaluronan-based hydrogel integrating exosomes for traumatic brain injury repair by promoting angiogenesis and neurogenesis. Carbohydr Polym. 2023, 306, 120578. [Google Scholar] [CrossRef]
- Lu, Y.; Zhou, Y.; Zhang, R.; Wen, L.; Wu, K.; Li, Y.; Yao, Y.; Duan, R.; Jia, Y. Bone Mesenchymal Stem Cell-Derived Extracellular Vesicles Promote Recovery Following Spinal Cord Injury via Improvement of the Integrity of the Blood-Spinal Cord Barrier. Front Neurosci. 2019, 13, 209. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, Y.; Bai, W.; Sun, L.; Tian, M. Human umbilical cord mesenchymal stem cell-derived exosome suppresses programmed cell death in traumatic brain injury via PINK1/Parkin-mediated mitophagy. CNS Neurosci Ther. 2023. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Luo, W.; Jiang, W.; Li, H.; Xu, J.; Liu, X.; Wang, B.; Wang, J.; Chen, G. Human umbilical cord mesenchymal stem cell-derived exosomes promote neurological function recovery in rat after traumatic brain injury by inhibiting the activation of microglia and astrocyte. Regen Ther. 2022, 21, 282–287. [Google Scholar] [CrossRef]
- Williams, A.M.; Higgins, G.A.; Bhatti, U.F.; Biesterveld, B.E.; Dekker, S.E.; Kathawate, R.G.; Tian, Y.; Wu, Z.; Kemp, M.T.; Wakam, G.K.; Liu, B.; Li, Y.; Buller, B.; Alam, H.B. Early treatment with exosomes following traumatic brain injury and hemorrhagic shock in a swine model promotes transcriptional changes associated with neuroprotection. J Trauma Acute Care Surg. 2020, 89(3), 536–543. [Google Scholar] [CrossRef]
- Williams, A.M.; Wu, Z.; Bhatti, U.F.; Biesterveld, B.E.; Kemp, M.T.; Wakam, G.K.; Vercruysse, C.A.; Chtraklin, K.; Siddiqui, A.Z.; Pickell, Z.; Dekker, S.E.; Tian, Y.; Liu, B.; Li, Y.; Buller, B.; Alam, H.B. Early single-dose exosome treatment improves neurologic outcomes in a 7-day swine model of traumatic brain injury and hemorrhagic shock. J Trauma Acute Care Surg. 2020, 89(2), 388–396. [Google Scholar] [CrossRef]
- Ge, X.; Guo, M.; Hu, T.; Li, W.; Huang, S.; Yin, Z.; Li, Y.; Chen, F.; Zhu, L.; Kang, C.; Jiang, R.; Lei, P.; Zhang, J. Increased Microglial Exosomal miR-124-3p Alleviates Neurodegeneration and Improves Cognitive Outcome after rmTBI. Mol Ther. 2020, 28(2), 503–522. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Ge, X.; Yu, J.; Han, Z.; Yin, Z.; Li, Y.; Chen, F.; Wang, H.; Zhang, J.; Lei, P. Increased miR-124-3p in microglial exosomes following traumatic brain injury inhibits neuronal inflammation and contributes to neurite outgrowth via their transfer into neurons. FASEB J. 2018, 32(1), 512–528. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, Y.; Wang, D.; Yan, W.; Zhang, S.; Li, D.; Han, Z.; Chen, F.; Lei, P. MiR-124-3p attenuates brain microvascular endothelial cell injury in vitro by promoting autophagy. Histol Histopathol. 2022, 37(2), 159–168. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y. G.; Wang, J.L.; Zhang, Y.X.; Li, L.; Reza, A.M.M.T.; Gurunathan, S. Biogenesis, Composition and Potential Therapeutic Applications of Mesenchymal Stem Cells Derived Exosomes in Various Diseases. Int J Nanomed. 2023, 18, 3177–3210. [Google Scholar] [CrossRef] [PubMed]
- Yari, H.; Mikhailova, M.V.; Mardasi, M.; Jafarzadehgharehziaaddin, M.; Shahrokh, S.; Thangavelu, L.; Ahmadi, H.; Shomali, N.; Yaghoubi, Y.; Zamani, M.; Akbari, M.; Alesaeidi, S. Emerging role of mesenchymal stromal cells (MSCs)-derived exosome in neurodegeneration-associated conditions: a groundbreaking cell-free approach. Stem Cell Res Ther. 2022, 13(1), 423. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Wang, S.; Zhao, R.C. Exosomes from mesenchymal stem/stromal cells: a new therapeutic paradigm. Biomarker Res. 2019, 7, 8. [Google Scholar] [CrossRef]
- Reza-Zaldivar, E.E. , Hernández-Sapiéns, M.A., Minjarez, B., Gutiérrez-Mercado, Y.K., Márquez-Aguirre, A.L., Canales-Aguirre, A.A. Potential Effects of MSC-Derived Exosomes in Neuroplasticity in Alzheimer's Disease. Front Cel Neurosci. 2018, 12, 317. [Google Scholar] [CrossRef]
- Fayazi, N.; Sheykhhasan, M.; Soleimani Asl, S.; Najafi, R. Stem Cell-Derived Exosomes: a New Strategy of Neurodegenerative Disease Treatment. Mol. Neurobiol. 2021, 58(7), 3494–3514. [Google Scholar] [CrossRef]
- Hade, M.D.; Suire, C.N.; Suo, Z. Mesenchymal Stem Cell-Derived Exosomes: Applications in Regenerative Medicine. Cells 2021, 10(8), 1959. [Google Scholar] [CrossRef]
- Rarinca, V.; Nicoara, M.; Ciobica, A.; Mavroudis, I. A short editorial view on the relevance of exosomes in some neuropsychiatric manifestations - model studies, Academy of Romanian Scientists Annals, Series on Biological Sciences. 2022, Academy of Romanian Scientists Volume 11, No. 2, 124-126. [CrossRef]
Figure 1.
Sources of exosomes and models in which they proved their efficiency.
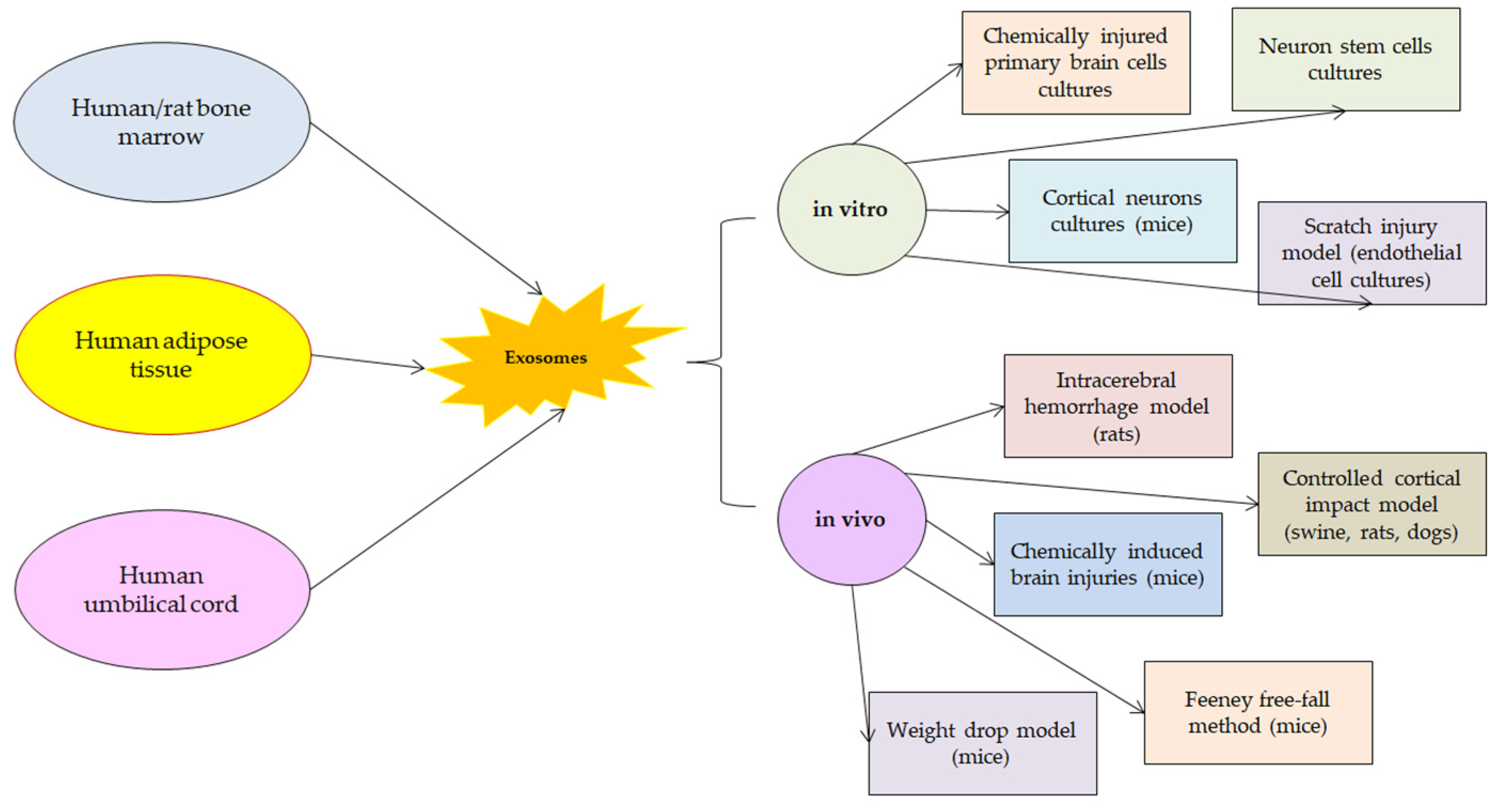

Table 1.
Summarized presentation of the studies regarding the effects of MSC-Exo in TBI models.
| Study | Objective | Models and administration | Key Findings |
|---|---|---|---|
| [19] | The therapeutic effects of MSC-Exo on astrocytic alterations induced by inflammation | In vitro: in cell cultures, hippocampal astrocytic inflammation was obtained by administering lipopolysaccharide to primary cultures;In vivo: in mice, pilocarpine administration produced status epilepticus.Treatment with MSC-Exo was performed in both cases. | The studied exosomes were incorporated into hippocampal astrocytes and attenuated reactive astrogliosis and inflammatory responses. They also ameliorated learning and memory impairments in mice with inflammation-induced astrocytic activation. |
| [20] | The effects of MSC-Exo on functional recovery, neuroinflammation, neuronal apoptosis, and neurogenesis | In vivo: weight-drop TBI rat model Human adipose MSC-Exo were administered intracerebroventricularly by microinjection | The studied exosomes promoted functional recovery, suppressed neuroinflammation, reduced neuronal apoptosis, and increased neurogenesis. They specifically entered microglia/macrophages and suppressed their activation during brain injury. |
| [21] | The potential of MSC-derived exosomes to regenerate the brain after TBI | In vivo: TBI canine model (Beagle dogs receiving small shocks using an electronic cortical contusion impactor through a hole in the skull).Hypoxia-treated human umbilical cord mesenchymal stem cells-derived exosomes were used for a 3D-printed collagen/silk fibroin/exosomes scaffold. | Exosomal treatment significantly promoted neuroregeneration and angiogenesis and inhibited nerve cell apoptosis and proinflammatory factor expression. Neuroregenerative properties and good biocompatibility were observed for hypoxia-induced human umbilical cord-derived MSC exosomes. |
| [23] | The effects of MSC-Exo in regarding the doses and times of administration | In vivo: unilateral moderate cortical concussion rat models.Single intravenous administration of exosomes at different time points after brain injury. | Exosome treatment improved sensorimotor and cognitive function, reduced hippocampal neuronal cell loss, promoted angiogenesis and neurogenesis, and reduced neuroinflammation.Earlier treatment administration (1-day post-TBI) provided a significantly greater improvement in functional and histological outcomes, as compared to delayed treatments. |
| [25] | The effects of MSC-Exo from bone marrow when administered in large animal models of TBI and hemorrhagic shock | In vivo: swine models that received TBI and hemorrhagic shock by a computer-controlled cortical impact device. MSC-Exo were separated from a single human donor of bone marrow and administered intravenously at 9h, 1, 5, 9, 13 days post-injury. | A single dose of MSC-Exo could attenuate the effects of the brain injury and provide better functional neurological recovery.They speculate that in large animal models the effects of MSC-Exo could be the result of central nervous system and peripheral immune response modulation. |
| [26] | The effects of multipotent MSC-Exo on functional recovery, neurovascular remodeling, and neurogenesis | In vivo: Wistar rat model of intracerebral hemorrhageMSC-Exo were administered in the tail at 24h after the injury | MSC-Exo promoted significant improvement of functional neurological recovery (spatial learning and motor functions). It was speculated that MSC-Exo could modulate endogenous angiogenesis and neurogenesis. |
| [27] | The efficiency of hyaluronan-collagen hydrogel incorporated bone marrow MSC-Exo in TBI treatment | In vitro: neuronal stem cells cultures | MSC-Exo incorporated in hydrogel could induce angiogenesis and neurogenesis. Axonal regeneration, remyelination, and synapse formation were observed. Brain structural remodeling could also be obtained by MSC-Exo treatment suggesting that neurological functional recovery after TBI could be achieved. |
| [28] | The effects of bone marrow MSC-Exo treatment in spinal cord injury. The potential mechanisms through which MSC-Exo protect the blood spinal cord barrier. | In vivo: rat model of spinal cord injury carried out by contusive injury with a spinal cord impactorMSC-Exo were isolated from rat bone marrow and administered intravenous 1-day post-injury. | It was showed that MSC-Exo could promote regeneration and neuronal survival reduced brain cell death, enhanced neuronal survival and regeneration, and improved motor functions.NF-κB p65 signaling in pericytes was identified as a possible action pathway of MSC-Exo. |
| [29] | The protective effects of human umbilical cord MSC-Exo in both in vivo and in vitro TBI models | In vitro: mice cortical neurons cultures exposed to mechanical stretch injury.In vivo: mice models of TBI (controlled cortical impact).Human umbilical cord MSC-Exo were obtained from expanded cultures and administered in 5 different doses in animal models, and in 3 doses in cultured neurons. | Improvement of neurological function, and decrease of cerebral edema were found to contribute to brain regeneration after TBI lesion. MSC-Exo suppressed apoptosis, pyroptosis, and ferroptosis and modulated PINK1/Parkin pathway-mediated mitophagy. |
| [30] | The mechanism through which umbilical cord MSC-Exo promote repair after TBI injury | In vivo: rat model of Feeney free-fall method for brain injuryMSC-Exo were isolated from 3rd-5th generation of umbilical cord stem cells cultures | MSC-Exo treatment promoted functional recovery and reduced neuronal apoptosis after TBI injury. Also, microglia and astrocytes were inhibited. |
| [31] | The molecular mechanisms that are contributing to brain repair after TBI injury and treatment with MSC-Exo | In vivo: swine model of severe TBI and hemorrhagic shock Bone marrow derived MSC-Exo were administered intravenously 1 hour after the injury. | Genes expression associated with neurogenesis, neuronal development, synaptogenesis, and neuroplasticity were significantly increased. Genes expression associated with stroke, neuroinflammation, neuroepithelial cell proliferation, and non-neuronal cell proliferation were significantly reduced. |
| [32] | The impact of early single-dose exosome treatment in a 7-day survival model | In vivo: swine model of severe TBI and hemorrhagic shock Bone marrow derived MSC-Exo were administered intravenously 1 hour after the injury. | It was showed that one dose of exosomes extract could improve survival of a large TBI model by decreasing brain lesion size, inhibiting inflammation and apoptosis, and promoting neural plasticity, when administered early. |
| [33] | The role of miRNAs in regulating post-traumatic neurodegeneration | In vivo: mice TBI models (close impact injury) | miR-124-3p modulated neurodegeneration and inhibited β-amyloid abnormalities. miR-124-3p could pass through the blood-brain barrier. miR-124-3p improved the cognitive outcome after repetitive mild TBI. |
| [34] | To explore the regulatory mechanism of microglial exosomes on neuronal inflammation in TBI by investigating the impact of microglial exosomal miRNAs on injured neurons | In vivo: mice model of controlled cortical impact In vitro: scratch injury model of cultured neurons | miR-124-3p inhibited neuronal inflammation and contributed to neurite outgrowth. |
| [35] | The effects of miR-124-3p on brain microvascular endothelial cells function and their molecular mechanisms | In vitro: specific cell scratch wound model for endothelial cell injury.Overexpression of miR-124-3p in endothelial cells was performed by treatment with Lipofectamine3000. | miR-124-3p overexpression prevented apoptosis and reduced blool-brain barrier leakage.miR-124-3p could be implicated in mTOR signaling and autophagy modulation in endothelial cells. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



