
Submitted:
28 July 2023
Posted:
31 July 2023
You are already at the latest version
Abstract
The bean weevil (Acanthoscelides obtectus (Say.)) is one of the main pests of stored beans in the world. Heterogeneous Inbreed Families (HIF) Population of Near Isogenic Lines (NILs) could be an exceptional strategy to study the inheritance of the resistance against A. obtectus. We developed a HIF population of NILs of R-bufa-80-12 (Phaseolus vulgaris) to study the inheritance related to bean weevil resistance and to search NILs with high resistance to the pest within the HIF population. The objective of this research was to study the inheritance in a HIF population and to identify the best lines to provide weevil resistance in bean breeding for commercial genotypes that could reduce pesticide applications. To achieve this objective, we developed a set of 148 NILs which were near-homozygous in the genome, except in the region under study. The HIF were evaluated in two environments under natural infestation with A. obtectus in free choice bioassays in a warehouse highly infested. The population showed wide variability for the traits studied, from 17.2 to 29.2 g for 100 seed weight, 0.0-0.8 for consumption in grams, 2.0-30.0 for consumption in percent and 1.2-9.6 for preference (number of adults in the bottle to the end of the experiments. The heritabilities for resistance traits were from very low to low 0.0 to 0.19. The heritability for 100 seed weight was the highest, with a value of 0.90. All traits studied in this research have a normal distribution, so they are considered to be quantitatively inherited. However, a great variability was found for the traits studied, which allows selecting lines with a high level of resistance within the R-bufa-80-12 HIF population. All traits studied in present research are of quantitative inheritance. The best lines of the HIF population for resistance to A. obtectus were the Line-45, Line-129, Line-124, Line-108 and Line-47.
Keywords:
bean weevil
; common bean
; HIF population
; inheritance
; weevil resistance
1. Introduction
Climate change is a current and future cause of hunger and the main constraint to guarantee food security, mainly in third world countries where the capability of response to extreme climate events is poor and insufficient [1]. Actual human population expansion and global climate change may have an impact on food stockpiles and plant seed availability in the near future. Food security under climate change is the major challenge of humanity; the goal is to produce food to feed more than 9 billion humans by 2050 with the same amount of land and resources [2]. Similarly, world grain production has increased each year, highlighting the need to invest in components of the postharvest system. In addition, to allow marketing after harvest, it is necessary to store the grains and preserve their quality during this period [3]. Nevertheless, storage temperatures and relative humidity associated with bean moisture content, prolonged storage period, and the presence of microorganisms constitute unsuitable storage conditions for bean storage [4].
Dry bean (Phaseolus vulgaris L.) is a staple food in many developing regions manly in Latin America and Africa. This legume does not require any industrial processing to be consumed. Its grain is used as auto consumption and the spare grain to interchange for other products. The area planted with bean in 2013, globally, exceed 29.5 million of hectares, with a total production of 22.8 million of tones [5]. In Mexico, common beans (Phaseolus vulgaris L.) is the second most important crop by planted area with more than 1,615,875 hectares with a production of 1,157,643 tons in 2021 and an average yield of 0.716 kg per hectare [6]. This yield is low mainly due to the effect of the drought since 90% of the surface is harvested in rainfed conditions. And of this 90, 70% is found in the highlands of Mexico where precisely the rainfall is scarce and poorly distributed [6]. In addition to this, the low densities of plants harvested by the farmers are the second problem in bean production since they harvest an average of 50,000 plants·ha-1 when they should harvest between 100,000 and 150,000 plants·ha-1 [7] [8]. There are pests in Mexico that cause severe damage in the field such as the mexican bean beetle, Epilachna varivestis Mulsant (Coleoptera: Coccinellidae), the bean weevil, Apion godmani Wagner (Coleoptera: Curculionidae) [9], and the bean grashopper, Brachystola magna Girard (Orthoptera: Acrididae) [10]. Likewise, the two main pests in storage are the mexican bean weevil, Zabrotes subfasciatus Boheman (Coleoptera: Bruchidae) and the bean weevil, Acanthoscelides obtectus Say (Coleoptera: Bruchidae). A. obtectus is the pest that attacks the highlands, the largest area of bean production in Mexico [11,12].
Nutritionists characterize beans as an exceptional food resource because of its high protein content and its combination of carbohydrates, dietary fiber, and minerals (particularly iron and zinc) [13]. This legume also contains thiamine and some neuroprotective and anticancerines properties [14]. However, to take care of the nutritional and nutraceutical properties of the bean, excellent storage care is required. On the other side, bean farmers obtain low yields due to many factors as lack of technology, use of low quality seeds, drought, adverse climate, pest and diseases between others [15,16,17]. Furthermore, problems do not over in the field, storage plague also damage the grains reducing in quality and quantity the already low production. Pest-caused post-harvest loss may surpass 20% in underdeveloped and tropical nations due to insufficient management measures and environmental factors that allow pests to reproduce quickly, particularly in emerging countries [18]. Acanthoscelides obtectus (Coleoptera: Bruchidae) (Say) is one of the most important post-harvest pests of dry bean Phaseolus vulgaris L [19]. Also, it is one of the most serious insect pests of stored beans worldwide in the post-harvest period [20]. Some studies have reported losses around the 7–40% [21]. This equates to a loss of 1.59 to 9.12 million of tones [19]. Other authors have been reported losses in the range of 20–100% in stored beans [17,22]. The main storage pest reported for P. vulgaris are the weevils: Acanthoscelides obtectus (Coleoptera: Bruchidae) (Say) and Zabrotes subfasciatus Boh [19,23,24].
Coleoptera is the largest order of insects and are the most common and damaging stored product pests [25]. Damage to beans caused by weevils can be great, depending on storage facilities and conditions [26]. These insects are highly adaptive, distributed in rural and urban areas, in a wide range of climates (temperate to tropical) and as a consequence are a serious problem to storage grain legumes. The percentage of damage range from 30% (in the field) to 74% (in storage facilities) with losses up to 9 millions of tones yearly [19,27]. The bean weevil (Acanthoscelides obtectus) is a neotropical, multivoltine pest distributed in the five continents manly in Latin America and Africa, its place of origin was in Central America [28]. Many species of the Acanthoscelides genus are found around the world. Since the middle of the 1980s A. obtectus has been increased its distribution in cooler countries such as Russia [29]. The bean weevil is highly adaptive and its infection starts in the field and continues during storage, where it causes the biggest damage. Its preference host is P. vulgaris but also infects P. lunatus and other grain legumes.
There are many ways to control or reduce the damage of the bean weevil since physical methods like freezing, use of chemicals products, farming practices or natural enemies such as the predatory mites Blattisocius tarsalis, Amblyseius swirskii and the larval parasitoids Anisopteromalus calandrae and Lariophagus distinguendus, and one of the most effective the use of resistant genotypes. Some of these practices have a negative impact on the quality of the seeds and the environment [27,30]. The alternative, the development of resistant lines for insect resistance has several economic and environmental advantages.
On the other hand, some reports mention that wild bean Mexican genotypes have been identified as highly resistant to this plague, such as the accession, G12952, the resistance was found to be inherited as two recessive complementary genes with segregation of 15S:1R, in this research the resistant genotypes were relatively stable as expected with recessively inherited traits [31]; recently Jiménez et al. [32], found a major gene with segregation 3:1 for resistance to A. obtectus in two F2 biparental populations of Phaseolus spp., in addition found pipecolic acid as responsible of the resistance. It has been described that the chemical compounds in the plant will define the host as resistant, tolerant or susceptible; for example, specific proteins in the seed make some genotypes of common bean resistant to Callosobruchus chinensis and Zabrotes subfasciatus, and some wild beans germoplasm are resistant to bean weevil due to the arcelin found in the genotypes [33].
Several researches have been carried out with Rosa Bufa (R-bufa) genotype for drought tolerance with PEG-6000 [34]; drought tolerance and precocity in field experiments [35] and the effect of moisture restriction on morphological and physiological characteristics of the grain [36]; the humidity restriction and high night temperature significantly and synchronously accelerated leaves and pods senescence in cv. R-bufa. In contrast, in leaves of cv. OTI, the loss of green color began several days earlier than in pods [37].
To date the research on weevil resistance in commercial genotypes dry bean is scarce. Only two QTL mapping reports [27,38], and two inheritance studies have been reported in biparental crosses, a segregation 15S: 1R by Kornegay and Cardona. [31] and 3R: 1S by Jiménez-Galindo et al. [32]. Inheritance of the resistance to A. obtectus with HIF populations of bean has not been reported to date. The advantage of HIF populations is that they are formed from a segregating plant for the trait [39] and it is possible to reach the genetic sequence of resistance, in this case to A. obtectus. The objective of this research was to study the inheritance in a HIF population and to identify the best lines of the HIF population generated to provide weevil resistance in bean breeding for commercial genotypes that could reduce pesticide applications to control A. obtectus in stored and field beans, being a very cheap and safe way for controlling this pest.
2. Materials and methods
2.1. Plant materials
The HIF method was used to develop the HIF population under study [40]. An offspring analysis was performed on individually harvested plants of the R-bufa genotype to develop the HIF population of 148 lines (Table 1). In 2016, 21 seeds of R-bufa were sown and the seed of each plant was harvested individually. An offspring study was realized in the plants R-bufa-80-2, R-bufa-80-4, R-bufa-80-12 and R-bufa-60 for resistance to A. obtectus from Spain. The pest of the offspring analysis was recollected and reproduced in Pontevedra, Spain. It was found that plant number 12 (R-bufa-80-12) showed segregation for the resistance to this pest. Subsequently, all the seeds of plant 12 were sown to increase the seed of each one and for the 148 lines of the HIF population in Bachiniva, Chihuahua. A scheme of the development process of the HIF population is shown in Figure 1.
2.2. Experimental design
The seeds used from the lines and the parents harvested and the adults of A. obtectus used in this experiment were recollected in 2019 in Bachiniva, Chihuahua. Two experiments were carried out in lattice with two repetitions for the analysis of the resistance of the 148 lines of the HIF population. Both experiments started on April 12, 2021 and ended on February 11, 2022. The material used from the lines and the parents were harvested in 2019 in Bachiniva, Chihuahua. One hundred forty-eight lines of the HIF population were evaluated in plastic bottles of 5 cm in diameter and 5 cm in height. With 10 seeds per repetition and the genotype that gave rise to the HIF population: R-bufa-80-12, as well as T-amarillo as resistant control. The bottles were left open inside a warehouse highly infested with A. obtectus. Four aleatory samples of 1 m2 size were taken to determine the level of infestation, on average 367 adults were found per square meter. The adults of A. obtectus were kept in contact with the seeds of the pure lines for 10 months and were evaluated. The following variables were measured: 1) Weight of 100 seeds, obtained with the formula: Weight of 100 seeds = 100 × weight of 10 seeds ÷ 10; 2) Consumption in grams, obtained by subtracting the final seed weight from the initial weight; 3) Consumption in percent, obtained with the formula: Consumption % = (initial weight − final weight) × 100 ÷ initial weight; 4) number of adults in bottle (preference), determined by accounting the number of adults per repetition at the end of the experiments.
2.3. Statistical analysis
The ANOVA was performed using the SAS General Linear Model (PROC GLM) [42]. The distribution of all resistance characteristics and the weight of 100 seeds were analyzed using the univariate procedure (PROC UNIVARIATE) of SAS [42]. All factors: replicates, blocks within replicates, and NILs were considered random effects. Heritabilities (h2) across environments were estimated for each trait on a family mean basis as described by Holland et al. [43]. Genetic and phenotypic correlations between traits were calculated following Holland. [44]. All previous analyzes were done in SAS software [42].
3. Results
3.1. Analysis of variance and heritability of parents and controls
Significant differences were found for almost all the variables studied between the control T-amarillo and the line that gave rise to the HIF population: R-bufa-80, except the variable of preference, number of adults in the bottle at the end of the experiments. The population showed wide variability for the traits studied, from 17.2 to 29.2 g for 100 seed weight, 0.0–0.8 for consumption in grams, 2.0–30.0 for consumption in percent and 1.2–9.6 for preference, number of adults in the bottle at the end of the experiments. The heritabilities for resistance traits were low and very low from 0.0 to 0.19. The heritability for 100 seed weight was the highest, with a value of 0.90 (Table 2).
3.2. Genetic and phenotypic correlations
Positive and significant genetic correlations were found between consumption in grams and percentage of consumption (0.94). Positive and significant phenotypic correlations were also found between 100 seed weight and consumption (g) (0.15), between preference and consumption (g) (0.20) and between preference and consumption (%) (0.20). And finally, positive correlations between preference and consumption (%) (0.20) and a very obvious, high, positive correlation between consumption (g) and consumption (%) (0.99) (Table 3).
3.3. Normality analysis
The pure lines (NILs) of the HIF population showed a great variability for all the traits analyzed. The 100 seed weight varied between 17.2–29.2 g while the preference of adults had a variation from 1.2 to 9.6 adults per repetition at the end of experiments. Consumption in grams ranged from 0.0 to 0.8 g. The traits of consumption in percent had variability from 2.0 to 30.0% (Figure 2). All traits studied in this research have a normal distribution, so they are considered to be quantitatively inherited. However, a great variability was found for the traits studied, which allows selecting lines with a high level of resistance within the R-bufa-80-12 HIF population.
3.4. Analysis of variance for 5 best and 3 most inferior lines for resistance to A. obtectus
The best NILs within the HIF population for resistance to A. obtectus were Line-45, Line-129, Line-124, Line-108 and Line-47. Previous lines presented 0.0 of consumption in grams and 0.0% of consumption in the seed. These lines were compared in an analysis of variance together with the lines that had the worst response for resistance to A. obtectus: Line-3, Line-62 and Line-63 with consumption in grams of 0.6 to 0.7 and a range of consumption in percent from 25.0 to 30.0 (Figure 3).
4. Discussion
Tepary beans from Mexico (Phaseolus acutifolius) are great source of resistance against A. obtectus to improve dry bean [32]. Also, some ancient beans from Mexico could be excellent sources of resistance to A. obtectus, as is the case of the R-bufa genotype.
In the case of the inheritance of resistance to A.obtectus, two biparental crosses have been reported by Kornegay and Cardona. [31] and in two biparental populations by Jiménez- Galindo et al. [32]. In the first case a two recessive complementary and independent genes were suggested to be responsible for the number of days to adult emergence of A. obtectus in two F2: A36 × G12952 and Pijao × G12952 with the segregation 15 [susceptible (short period to adult emergence)]:1 [resistant (long period)]; the resistance was found in cotyledons. In the second case a Mendelian gene was suggested to be responsible for the number of adults of the first generation in two F2: P-saltillo × T-amarillo and T-amarillo × T-cafe with the segregation 3 [resistant lines (with cero adults of the first generation)]:1[susceptible line (with one or more first generation adults)] (R:S). At the contrary of research reported by Kornegay and Cardona. [31] in this last case the resistance was found in the testa of resistant lines. Our present research is according with Li et al. [27] and Minney. [45] that found a quantitative inheritance of the weevil resistance trait controlled by recessive polygenes.
There are no reports of HIF populations to study the inheritance of resistance to A. obtectus. In this case, the traits studied in the present HIF population are quantitatively inherited. The advantage of these populations is that are near-homozygous in the genome, except in the region under study [39]. It is also important to mention that there would probably be more lines resistant to A. obtectus from Spain since we believe that the pest is adapted to different populations around the world. We are according with Rendon-Huerta et al. [46] who considers that A. obtectus have a high genetic variability and a more geographic dispersal as compared to Z. subfasciatus.
The HIF population for resistance to A. obtectus was obtained with weevils from Spain. The present study was carried out with A. obtectus collected in Mexico. It is highly probably that the weevils from Spain and from Mexico are genetically different and adapted to the bean genotypes that are produced and consumed in each region. There are maybe also bean populations with resistance to various populations of A. obtectus insects. Jiménez-Galindo et al. [32] found a high level of resistance to A. obtectus from Spain in the F2 population of the cross P-saltillo × T-amarillo. Besides Hernández et al. [47], in the same F3 population found a high level of resistance to A. obtectus from Mexico. We also mention that this HIF population from R-bufa-80-12 presents variability to study resistance to A. obtectus from Spain and Mexico, even though the pest is genetically different. Our results are according with Thakur. [48] and Duan et al. [49] who found in 1992, the first record of A. obtectus described from India, the research found that many characters of the weevils from India were similar to the American native population but different from the A. obtectus found in Galapagos.
The degree of variation and heritability of traits have influence in the selection in breeding programs [50]. In the present study, the HIF population of NILs showed a heritability of 0.90 for the variable of weight of 100 seeds. Our results are in agreement with White et al. [51] who found a heritability of 0.57 to 0.80 for the same trait. In the present research the heritabilities for the resistance traits to A. obtectus were very low and low from 0.0 to 0.19. This is in agreement with Galwey and Evans. [52] and Kornegay and Temple. [53] who found that leafhopper tolerance and antixenosis are quantitatively inherited with low heritability in crosses of small black beans tolerant to the “Tacaragua” leafhoppers and pinto beans Sierra with medium susceptibility to leafhoppers. Gonzales et al. [54] also reported low heritability for antixenosis in beans to leafhoppers.
Kornegay et al. [55] found that seed size is negatively correlated with adult weight but not with days to adult emergence. The correlation coefficients, between seed size and number of eggs, number of adults, adult weight, and life cycle were 0.64, 0.69, 0.63, and −0.42, respectively [56]. In the present study, positive and significant phenotypic correlations were found between the weight of 100 seeds and consumption in grams (0.15). This means that probably in some lines with larger seeds the adults consume a greater amount of the grain due to the accessibility of food, and a greater number of adults emerge. Also, between preference and consumption in grams (0.20), and percentage of consumption (0.20). The seed consumption by A. obtectus is highly correlated with the number of first generation adults [19]; [32], could also be attributed to beneficial thermal conditions in a range of 20–28 °C [57]. So in free choice trials as in the present investigation the consumption traits are very interesting. In this type of experiments many resistance variables cannot be measured and it is not possible to elucidate whether it is high preference of the susceptible lines or high antixenosis and antibiosis of the resistant ones.
All the characters studied in the present study have a normal distribution, considered to be of quantitative inheritance. However, a great variability was found for the studied characters that allow selecting lines with a high level of resistance within the HIF population of R-bufa-80-12. Other studies have also found normal distributions for resistance traits to A. obtectus. In fact, the F2 reported by Kornegay and Cardona. [31] showed a continuous, but skewed, distribution of the variable days to adult emergence from low to high. The F3 generation showed an overall decrease in levels of resistance compared to the original evaluations.
Kornegay and Cardona. [31] found only a few F2 individuals with the level of resistance of the G12952 genotype. In the present study many inbred lines of the HIF population with resistance were found.
5. Conclusions
All traits studied in the present research are of quantitative inheritance. The best lines of the HIF population for resistance to A. obtectus are the Line-45, Line-129, Line-124, Line-108 and Line-47. The HIF population presents high variability for resistance to A. obtectus from Mexico. This HIF population is suitable for genomics, proteomics and metabolomics studies to discover new compounds, proteins and resistance genes to A. obtectus. Further investigation of sequencing analysis could identify candidate gene and genetic sequences for A. obtectus resistance.
Author Contributions
A.Y.A.-R., G.C.-P., and J.C.J.-G. conceived the study and discussed the results. J.C.J.-G. developed HIF population. A.Y.A.-R., G.C.-P. and J.C.J.-G. carried out field experiments, and performed statistical analysis of the data. A.Y.A.-R., G.C.-P., J.C.J.-G., N.R.-C., O.R.-V., J.J.F.-G., and R.A.M. drafted the initial manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by National Institute of Forestry, Agriculture and Livestock Research of México. Grant number: 1-1.6-9385034743-A-M.2-2 “Regiones genómicas asociadas a la resistencia a plagas de almacén en una población MAGIC de maíz”.
Data Availability Statement
The data sets generated and analyzed during the current study and material are available from José Cruz Jiménez-Galindo upon reasonable request.
Acknowledgments
Adriana Ayala-Ruiz is grateful to the National Institute of Forestry, Agriculture and Livestock Research (INIFAP) in Mexico for the funding that supported his Bachelor Research.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Mbow, C. , et al., Food security. 2019.
- Roser, M. and L. Rodés-Guirao, Future population growth. Our world in data, 2013.
- Ziegler, V., R. T. Paraginski, and C.D. Ferreira, Grain storage systems and effects of moisture, temperature and time on grain quality-A review. Journal of Stored Products Research, 2021. 91: p. 101770.
- Ferreira, C.D.; Ziegler, V.; Lindemann, I.d.S.; Hoffmann, J.F.; Vanier, N.L.; de Oliveira, M. Quality of black beans as a function of long-term storage and moldy development: Chemical and functional properties of flour and isolated protein. Food Chem. 2018, 246, 473–480. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Statistical Database. In Food and Agriculture Organization of the United Nations; FAO: Rome, Italy, 2022. [Google Scholar]
- SIAP-SADER. 2023. Sistema de Información Agro Pecuaria de la Secretaria de Desarrollo Rural. Available online: https://nube.siap.gob.mx/cierreagricola/ (accessed on 20 May 2023).
- Jiménez Galindo, J.C. and J.A. Acosta Gallegos, Efecto y correlación de fechas de siembra, fertilización y densidad en el rendimiento de frijol Pinto Saltillo de temporal en Chihuahua. Revista mexicana de ciencias agrícolas, 2013. 4(1): p. 115-127.
- Galindo, J.C.J.; Gallegos, J.A.A. Efecto de la densidad a simple y doble hilera en el rendimiento de frijol de temporal en Chihuahua, Mexico. Revista mexicana de ciencias agrícolas 2018, 4, 393–407. [Google Scholar] [CrossRef]
- Acevedo-Peralta, I. , et al., Plagas insectiles y su efecto en el rendimiento en tres variedades de frijol en Texcoco, Mexico. AGROProductividad, 2010. 3(2): p. 22-26.
- Bustillos-Rodríguez, J.C. , et al., Laboratory Assessment of Metarhizium spp. and Beauveria spp. Isolates to Control Brachystola magna1 in Northern México. Southwestern Entomologist, 2016. 41(3): p. 643-656.
- Jiménez-Galindo, J.C. , et al., Inheritance and metabolomics of the resistance of two F 2 populations of Phaseolus spp. to Acanthoscelides obtectus. Arthropod-Plant Interactions, 2020. 14(5): p. 641-651.
- Jiménez, J.C.; de la Fuente, M.; Ordás, B.; Domínguez, L.E.G.; Malvar, R.A. Resistance categories to Acanthoscelides obtectus (Coleoptera: Bruchidae) in tepary bean (Phaseolus acutifolius), new sources of resistance for dry bean (Phaseolus vulgaris) breeding. Crop. Prot. 2017, 98, 255–266. [Google Scholar] [CrossRef]
- Uebersax, M.A. , et al., Dry beans (Phaseolus vulgaris L.) as a vital component of sustainable agriculture and food security—A review. Legume Science, 2023. 5(1): p. e155.
- Broughton, W.J. , et al., Beans (Phaseolus spp.)–model food legumes. Plant and soil, 2003. 252: p. 55-128.
- Ayvaz, A.; Sagdic, O.; Karaborklu, S.; Ozturk, I. Insecticidal Activity of the Essential Oils from Different Plants Against Three Stored-Product Insects. J. Insect Sci. 2010, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Baldin, E.L.; Lara, F.M. Resistance of stored bean varieties toAcanthoscelides obtectus(Coleoptera: Bruchidae). Insect Sci. 2008, 15, 317–326. [Google Scholar] [CrossRef]
- Ogendo, J.O.; Deng, A.L.; Birech, R.J.; Bett, P.K. Plant-Based Products as Control Agents of Stored-Product Insect Pests in the Tropics. 2012. [Google Scholar] [CrossRef]
- Aidoo, K. Post-harvest storage and preservation of tropical crops. Int. Biodeterior. Biodegradation 1993, 32, 161–173. [Google Scholar] [CrossRef]
- Jiménez, J.C.; de la Fuente, M.; Ordás, B.; Domínguez, L.E.G.; Malvar, R.A. Resistance categories to Acanthoscelides obtectus (Coleoptera: Bruchidae) in tepary bean (Phaseolus acutifolius), new sources of resistance for dry bean (Phaseolus vulgaris) breeding. Crop. Prot. 2017, 98, 255–266. [Google Scholar] [CrossRef]
- Baldin, E.L.L.; Lara, F.M.; Camargo, R.S.; Pannuti, L.E.R. Characterization of resistance to the bean weevil Acanthoscelides obtectus Say, 1831 (Coleoptera: Bruchidae) in common bean genotypes. Arthropod-Plant Interactions 2017, 11, 861–870. [Google Scholar] [CrossRef]
- Mbogo, K., J. Davis, and J. Myers, Transfer of the arcelin-phytohaemagglutinin-α amylase inhibitor seed protein locus from tepary bean (Phaseolus acutifolius A. Gray) to common bean (P. vulgaris L.). Biotechnology, 2009. 8(3): p. 285-295.
- Ebinu, J.; Nsabiyera, V.; Otim, M.; Nkalubo, S.; Ugen, M.; Agona, A.; Talwana, H. Susceptibility to bruchids among common beans in Uganda. Afr. Crop. Sci. J. 2016, 24, 289. [Google Scholar] [CrossRef]
- Kusolwa, P.M. , Breeding for bruchid resistance in common bean (Phaseolus vulgaris L.): interspecific introgression of lectin-like seed proteins from tepary bean (P. acutifolius A. Gray), genetic control and bruchid resistance characterization. 2007.
- Mallqui, K.S.V.; Oliveira, E.E.; Guedes, R.N.C. Competition between the bean weevils Acanthoscelides obtectus and Zabrotes subfasciatus in common beans. J. Stored Prod. Res. 2013, 55, 32–35. [Google Scholar] [CrossRef]
- Hernandez-Lambraño, R.; Pajaro-Castro, N.; Caballero-Gallardo, K.; Stashenko, E.; Olivero-Verbel, J. Essential oils from plants of the genus Cymbopogon as natural insecticides to control stored product pests. J. Stored Prod. Res. 2015, 62, 81–83. [Google Scholar] [CrossRef]
- Askey, J. , et al., Oxygen demand of bean bruchids (Acanthoscelides obtectus Say). Journal of Stored Products and Postharvest Research, 2020. 11(2): p. 8-14.
- Li, X. , et al., QTL mapping and identification of genes associated with the resistance to Acanthoscelides obtectus (Say) in cultivated common bean using a high-density genetic linkage map. 2022.
- Oliveira, M.R.C. , et al., Mesoamerican origin and pre-and post-Columbian expansions of the ranges of Acanthoscelides obtectus Say, a cosmopolitan insect pest of the common bean. PloS one, 2013. 8(7): p. e70039.
- Kaplin, V.G. Distribution and Biology of Invasive Species of Bean Bruchid Acanthoscelides obtectus (Insecta, Coleoptera, Bruchidae). Russ. J. Biol. Invasions 2022, 13, 41–57. [Google Scholar] [CrossRef]
- Iturralde-García, R.D.; Castañé, C.; Wong-Corral, F.J.; Riudavets, J. Biological control of Acanthoscelides obtectus and Zabrotes subfasciatus in stored dried beans. BioControl 2020, 65, 693–701. [Google Scholar] [CrossRef]
- Kornegay, J.L.; Cardona, C. Inheritance of resistance to Acanthoscelides obtectus in a wild common bean accession crossed to commercial bean cultivars. Euphytica 1991, 52, 103–111. [Google Scholar] [CrossRef]
- Jiménez-Galindo, J.C. , et al., Inheritance and metabolomics of the resistance of two F 2 populations of Phaseolus spp. to Acanthoscelides obtectus. Arthropod-Plant Interactions, 2020. 14(5): p. 641-651.
- Ishimoto, M.; Sato, T.; Chrispeels, M.J.; Kitamura, K. Bruchid resistance of transgenic azuki bean expressing seed α-amylase inhibitor of common bean. Entomol. Exp. et Appl. 1996, 79, 309–315. [Google Scholar] [CrossRef]
- Jimenez-Galindo, J.C.; Alvarez-Iglesias, L.; Revilla-Temino, P.; Jacinto-Soto, R.; Garcia-Dominguez, L.E.; de La Fuente-Martinez, M.; Malvar-Pintos, R.A.; Ordas-Lopez, B.; Wal, A.J.V.; Osorno, J.M. Screening for Drought Tolerance in Tepary and Common Bean Based on Osmotic Potential Assays. Plant 2018, 6, 24. [Google Scholar] [CrossRef]
- Jiménez Galindo, J.C. and J.A. Acosta Gallegos, Caracterización de genotipos criollos de frijol Tepari (Phaseolus acutifolius A. Gray) y común (Phaseolus vulgaris L.) bajo temporal. Revista mexicana de ciencias agrícolas, 2012. 3(8): p. 1565-1577.
- Cilia García, M. , et al., Effects of water restriction on carbohydrates concentration, starch granules size and amylolytic activity in seeds of Phaseolus vulgaris L. and P. acutifolius A. Gray. Botanical Sciences, 2021. 99(2): p. 364-376.
- Cayetano-Marcial, M.; Peña-Valdivia, C.; Esteva, A.G.; Galindo, J.J.; Escobedo, I.G.; Chacón, D.P. Humidity Restriction, High Night Temperature and their Combination, during Post Flowering on Common Bean (Phaseolus vulgaris L.) Canopy and Pod Senescence. Legum. Res.-Int. J. 2021; 44, 634–640. [Google Scholar] [CrossRef]
- Kamfwa, K.; Beaver, J.S.; Cichy, K.A.; Kelly, J.D. QTL Mapping of Resistance to Bean Weevil in Common Bean. Crop. Sci. 2018, 58, 2370–2378. [Google Scholar] [CrossRef]
- Jiménez-Galindo, J.C.; Malvar, R.A.; Butrón, A.; Caicedo, M.; Ordás, B. Fine analysis of a genomic region involved in resistance to Mediterranean corn borer. BMC Plant Biol. 2018, 18, 169. [Google Scholar] [CrossRef]
- Tuinstra, M.R.; Ejeta, G.; Goldsbrough, P.B. Heterogeneous inbred family (HIF) analysis: a method for developing near-isogenic lines that differ at quantitative trait loci. Theor. Appl. Genet. 1997, 95, 1005–1011. [Google Scholar] [CrossRef]
- Singh, S.P. , A key for identification of different growth habits of Phaseolus vulgaris L. 1981.
- SAS Institute. Base SAS 9.4 Procedures Guide: Statistical Procedures. Version 9.4; SAS Institute: Cary, NC, USA, 2016. [Google Scholar]
- Holland, J.B.; Nyquist, W.E.; Cervantes-Martínez, C.T. Estimating and Interpreting Heritability for Plant Breeding: An Update. Plant Breed. Rev. 2002, 22, 9–112. [Google Scholar] [CrossRef]
- Holland, J.B. Estimating Genotypic Correlations and Their Standard Errors Using Multivariate Restricted Maximum Likelihood Estimation with SAS Proc MIXED. Crop. Sci. 2006, 46, 642–654. [Google Scholar] [CrossRef]
- Minney, B.H. , Breeding phaseolus vulgaris (common bean) for resistance to the major pest bruchids Zabrotes subfasciatus and Acanthoscelides obtectus. Biochemical bases for seed resistance in wild lines. 1990, Durham University.
- Rendón-Huerta, J.A. , et al., Insecticide effect of wild plant powders on bean weevil (Zabrotes subfasciatus Boheman; Coleoptera: Bruchidae) in vitro. African Journal of Agricultural Research, 2013; Volume 8, pp. 971–977. [Google Scholar]
- Hernández C., A. , Mayra C. S. C., María A. F. C., Juan L. J. C., Rafael A. P. Q., and José Cruz J. G. 2021. Resistencia a Acanthoscelides obtectus en una población F2:3 de frijol P-saltillo × T-amarillo. Tesis de Licenciatura. Universidad Autónoma de Chihuahua, México.
- Thakur, D. , Taxonomy, distribution and pest status of Indian biotypes of Acanthoscelides obtectus (Coleoptera: Chrysomelidae: Bruchinae)-A new record. Pakistan Journal of Zoology, 2012. 44(1).
- Duan, C. , et al., Genetic diversity and differentiation of A canthoscelides obtectus S ay (C oleoptera: B ruchidae) populations in C hina. Agricultural and Forest Entomology, 2017. 19(2): p. 113-121.
- Mhlaba, Z.B.; A Shimelis, H.; Amelework, B.; Modi, A.T.; Mashilo, J. Variance components and heritability of yield and yield-related traits in tepary bean (Phaseolus acutifolius). South Afr. J. Plant Soil 2019, 36, 117–128. [Google Scholar] [CrossRef]
- White, J.W. , et al., Inheritance of seed yield, maturity and seed weight of common bean (Phaseolus vulgaris) under semi-arid rainfed conditions. The Journal of Agricultural Science, 1994. 122(2): p. 265-273.
- Galwey, N. and A.M. Evans, The inheritance of resistance to Empoasca kraemeri R oss & M oore in the common bean, Phaseolus vulgaris L. Euphytica, 1982. 31(3): p. 933-952.
- Kornegay, J.L.; Temple, S.R. Inheritance and Combining Ability of Leafhopper Defense Mechanisms in Common Bean 1. Crop. Sci. 1986, 26, 1153–1158. [Google Scholar] [CrossRef]
- Gonzales, J.W., D. P. Coyne, and D.T. Lindgren, Inheritance and heritability of leafhopper resistance in common beans (Phaseolus vulgaris). ANNUAL REPORT-BEAN IMPROVEMENT COOPERATIVE, 2002. 45: p. 80-81.
- Kornegay, J. , et al., Breeding for insect resistance in beans. A. Schoonhoven & O. van and Voysest (Eds.), Common Beans: Research for Crop Improvement, 1991: p. 619-648.
- Schoonhoven, A.V.; Cardona, C.; Valor, J. Resistance to the Bean Weevil and the Mexican Bean Weevil (Coleoptera: Bruchidae) in Noncultivated Common Bean Accessions. J. Econ. Entomol. 1983, 76, 1255–1259. [Google Scholar] [CrossRef]
- Soares, M.A.; Quintela, E.D.; Mascarin, G.M.; Arthurs, S.P. Effect of temperature on the development and feeding behavior of Acanthoscelides obtectus (Chrysomelidae: Bruchinae) on dry bean (Phaseolus vulgaris L.). J. Stored Prod. Res. 2015, 61, 90–96. [Google Scholar] [CrossRef]
Figure 1.
Scheme of obtaining 148 NILs from plant 12 of the offspring analysis of R-bufa-80-12 genotype.
Figure 1.
Scheme of obtaining 148 NILs from plant 12 of the offspring analysis of R-bufa-80-12 genotype.
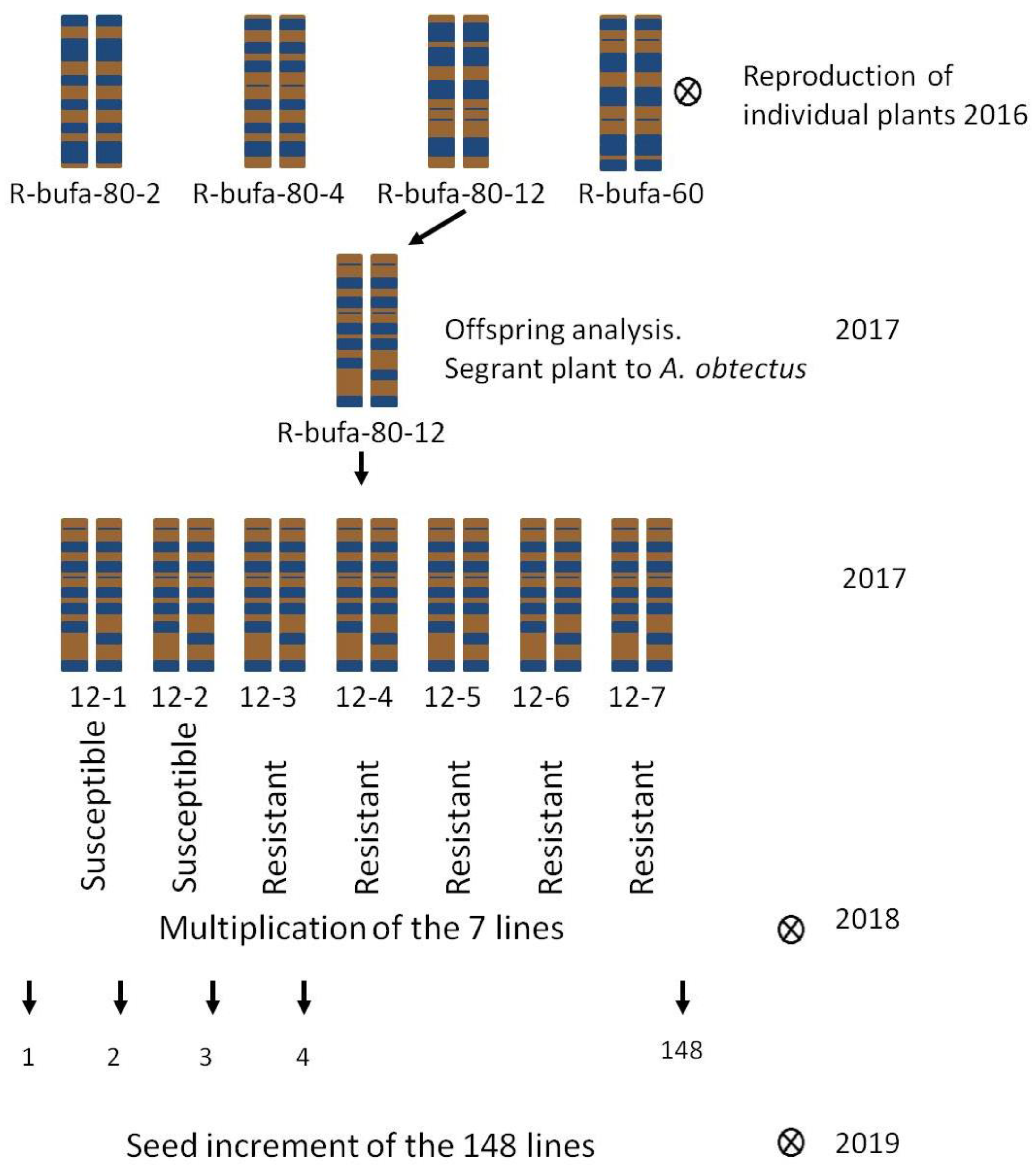
Figure 2.
A). Normality analysis for 100 seed weight. B). Preference (n adults per repetition at the end of experiments). C). Consumption in grams and D). Consumption in percent; of 148 NILs of the HIF population of the R-bufa-80-12.
Figure 2.
A). Normality analysis for 100 seed weight. B). Preference (n adults per repetition at the end of experiments). C). Consumption in grams and D). Consumption in percent; of 148 NILs of the HIF population of the R-bufa-80-12.
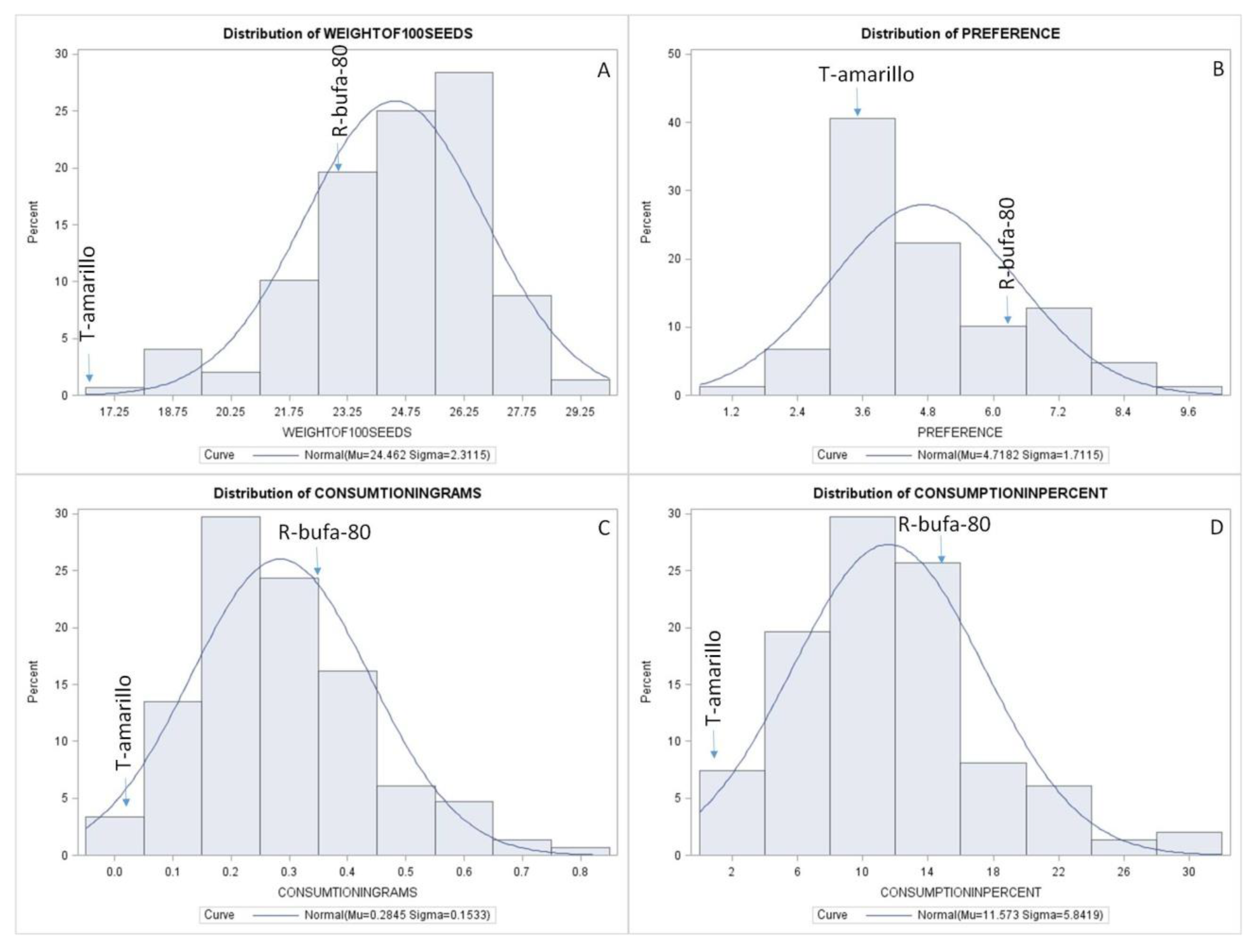
Figure 3.
Means and standard error for: A). 100 seed weight, B). Preference (n adults), C). consumption in grams and C). Consumption in percent of 8 contrasting NILs; of the HIF population of R-bufa-80-12 genotype.
Figure 3.
Means and standard error for: A). 100 seed weight, B). Preference (n adults), C). consumption in grams and C). Consumption in percent of 8 contrasting NILs; of the HIF population of R-bufa-80-12 genotype.
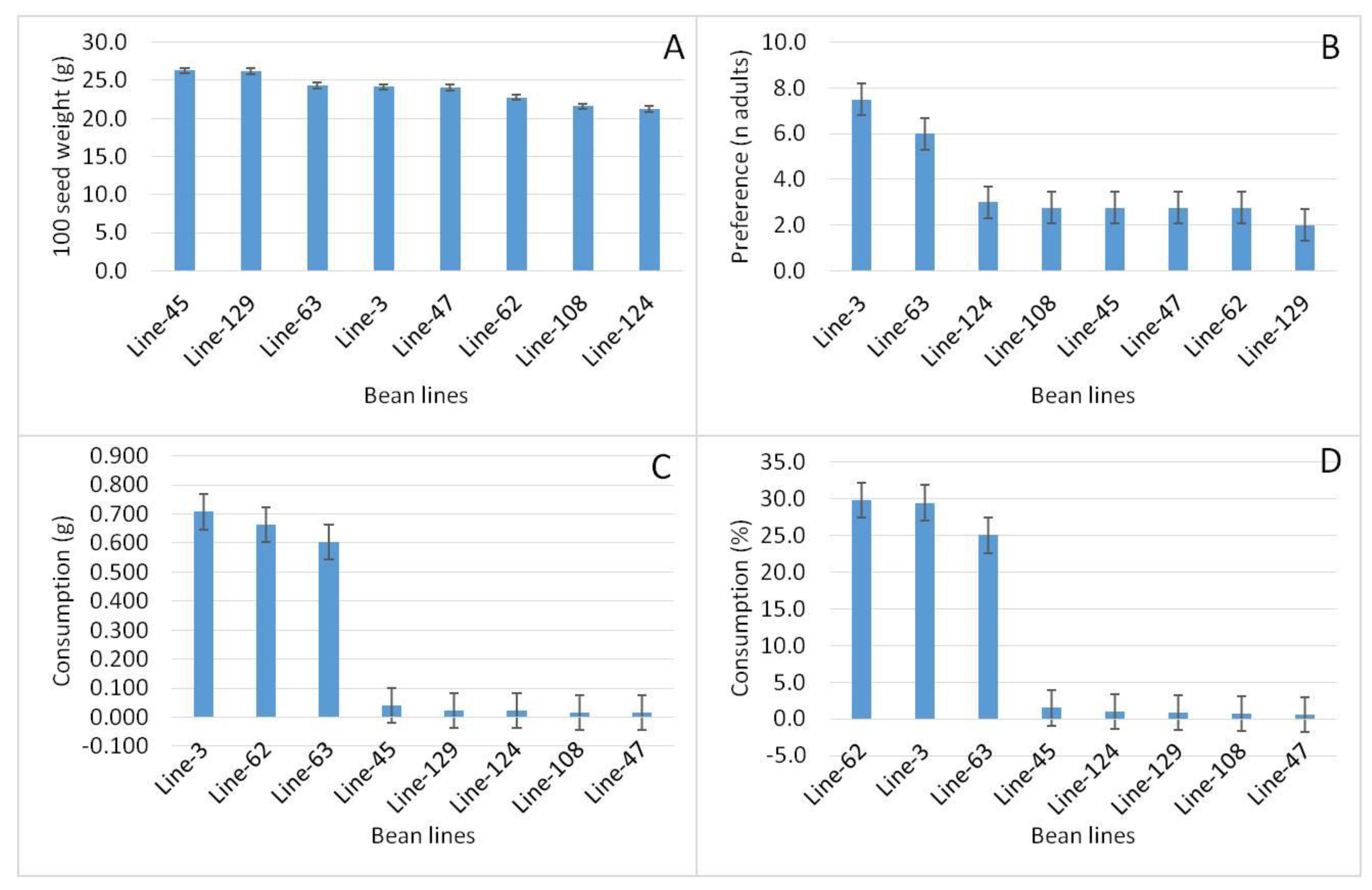

Table 1.
Characteristics of two bean genotypes used to create the HIF population analyzed for resistance to A. obtectus and the resistant control T-amarillo genotype.
Table 1.
Characteristics of two bean genotypes used to create the HIF population analyzed for resistance to A. obtectus and the resistant control T-amarillo genotype.
| Genotype | Species | Growth habit | Resistance level1 |
|---|---|---|---|
| R-bufa-80 | P. vulgaris | II | Susceptible |
| R-bufa-80-12 | P. vulgaris | II | Segregant |
| T-amarillo | P. acutifolius | III | Resistant |
Table 2.
Means and their standard errors, rank and heritabilities (h2) of the HIF population derived from the segregating plant R-bufa-80-12 for resistance to A. obtectus for 100 seed weight and resistance traits evaluated in two trials under infestation with A. obtectus. Mean comparisons, LSD and ±SE of the population, parent and control are also shown.
Table 2.
Means and their standard errors, rank and heritabilities (h2) of the HIF population derived from the segregating plant R-bufa-80-12 for resistance to A. obtectus for 100 seed weight and resistance traits evaluated in two trials under infestation with A. obtectus. Mean comparisons, LSD and ±SE of the population, parent and control are also shown.
| Seed trait | Resistance traits | ||||
| 100 seed weight | Consumption (g) | Consumption (%) | Adult preference (n) | ||
| 148 NILs | |||||
| Media | 24.5 | 0.284 | 11.5 | 4.5 | |
| ± SE | 0.11 | 0.01 | 0.46 | 0.21 | |
| Rank | 17.2–29.2 | 0.0–0.8 | 2–30 | 1.2–9.6 | |
| h2 | 0.90 | 0.19 | 0.18 | 0.08 | |
| Parent and control | |||||
| R-bufa-80 | 22.05 a | 0.356 a | 15.0 a | 6.37 a | |
| T-amarillo | 13.98 b | 0.021 b | 1.47 b | 3.50 a | |
| LSD | 1.8 | 0.273 | 11.3 | 4.3 | |
| ± SE | 1.23 | 0.09 | 3.16 | 1.27 | |
The heritabilities (h2) for each trait were estimated according to Holland et al. [43].
Table 3.
Genetic (below) and phenotypic (above) correlation coefficients between agronomic traits and resistance to A. obtectus registered of the HIF population derived from the genotype R-bufa-80-12 evaluated in two free choice experiments with A. obtectus.
Table 3.
Genetic (below) and phenotypic (above) correlation coefficients between agronomic traits and resistance to A. obtectus registered of the HIF population derived from the genotype R-bufa-80-12 evaluated in two free choice experiments with A. obtectus.
| 100 seed weight (g) | Preference | Consumption (g) | Consumption (%) | |
|---|---|---|---|---|
| 100 seed weight (g) | 0.06 | 0.15* | 0.06 | |
| Preference | 0.30 | 0.20* | 0.20* | |
| Consumption (g) | 0.45 | 2.9 | 0.99* | |
| Consumption (%) | 0.12 | 2.7 | 0.94* |
*Correlation coefficients that exceed twice their standard error.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



