
Submitted:
27 July 2023
Posted:
28 July 2023
You are already at the latest version
Abstract
Trypanosomes are the extracellular protozoan parasites that cause human African trypanosomiasis disease in humans and nagana disease in animals. The disease is endemic in African countries where tsetse flies which act as a vector for the transmission of disease are present. The animals infected by these parasites become useless or workless and disease can become fatal if not treated. Old drugs used for treatment have many side effects and some are responsible for death in 5% of the patients. The parasite possesses a major surface protein known as variant surface glycoprotein. The immune system of the host develops antibodies against this antigen but due to antigenic variation, parasites evade the immune response. Currently, no vaccine is available that provides complete protection. Only partial protection using certain antigens in murine models was investigated. The tools of molecular biology and immunology have enabled a novel approach to the development of vaccines against trypanosomes. Immunization is the sole method for the control of disease because the eradication of the vector from endemic areas is an impossible task. Genetic vaccines can carry multiple genes encoding different antigens of the same parasite or different parasites. DNA immunization induces the activation of both cellular immune response and humoral immune response along with the generation of memory. This review highlights the importance of DNA vaccines and advances in the development of DNA vaccines against T. brucei.
Keywords:
DNA vaccine
; delivery system
; variant surface glycoprotein
; nanoparticle
; antigenic variation
; endemic
1. Introduction
Trypanosomes are the causative agent of human African trypanosomiasis and nagana disease in animals [1-2]. Trypanosoma brucei sporadically causes disease in humans but it is the etiologic agent of trypanosomiasis in animals [3]. This is a vector-borne disease transmitted by the bite of a tsetse fly and often becomes fatal if not treated [4]. The diagnosis of the disease depends on the microscopic examination of a blood smear to find the extracellular flagellate protozoa in the blood of infected animals [5]. Old drugs are used for the therapy that also has side effects such as hypotension, anaphylactic shock, hypoglycemia, neurotoxic symptoms, adverse cutaneous reactions, and renal failure. These drugs include suramin, pentamidine, melarsoprol, eflornithine, and nifurtimox [6].
It is a parasitic disease and clinically characterized by two stages such as the early stage known as haemolymphatic infection and late stage meningoencephalatic infection [7]. It is endemic in Sub-Saharan Africa and its control primarily depends on the prevention of tsetse fly interaction with animals and humans, vaccination of the susceptible hosts, and treatment of the infected individuals [5]. The advancement in the field of molecular biology and immunology has revealed many proteins encoded by the genome of parasite and opened opportunities for the identification of target antigens so that a new drug or vaccine against this parasite become realistic. Trypanosoma brucei has over 800 genes involved in the synthesis of proteins and meshed in such a way as to evade the immune responses of the host [8]. Trypanosomatids have an outer coating made up of variant surface glycoproteins (VSGs) having antigenic variation. It is known that pseudogenes and hybrid genes are involved in the synthesis of VSGs, where the domains of C- and N-terminals are formed by separate genes [9].
Trypanosoma brucei can evade the humoral immune response of the host by utilizing variant surface glycoproteins. The surface membrane of the parasite creates a physical barrier between the host immunity and the pathogen and protects them from the mechanism of the immune system [7]. Many researchers are looking to find invariable membranous proteins which can act as candidates for the development of vaccines or drugs. Principally the VSG is an ideal candidate for the development of an effective vaccine if its antigenic variation is neglected because it is highly immunogenic, bloodstream stage-specific, and is the surface protein of the parasite [9]. Variant surface glycoprotein is an immunodominant antigen that elicits both cellular immune response and humoral immune response. Trypanosoma brucei has extensive diversity in its genome and displays an adaptation and specificities of the host with huge diversity. Therefore, the development of efficacious vaccines against trypanosomiasis is an urgent need worldwide [10].
Trypanosomes can activate macrophages through their coating of lipopolysaccharide and Glycosylated phosphatidyl inositol (GPI) anchor. Interferon-γ facilitates the activation of these macrophages [11]. IFN-γ provides a signal for the growth of the parasites which are induced by T cells using trypanin (a microtubule-binding protein). The immune response elicited by the activation of T cells during infection is of helper T cell 1 or helper T cell 2 types depending on the cytokines produced at the inflammatory site [10]. The activation of B cells and T cells aids in controlling the acute infection. Tolerance of the disease occurs by the immune system during the balance between pro-inflammatory and anti-inflammatory production of cytokines which control the extent of inflammation in the host to enhance its longevity [12-13]. From the above-mentioned results, it is concluded that DNA immunization can stimulate both cell-mediated immunity and humoral immunity against parasites causing disease in animals as well as humans (Figure 1).
2. Is it rational to design DNA vaccines for T. brucei?
The third generation of vaccines is the DNA or nucleic acid vaccines being evaluated against various parasitic diseases for their efficacy, safety, and cross-protection in animals. The development of third-generation vaccines offers opportunities for the prevention and control of protozoal parasitic diseases like trypanosomiasis [14]. This novel approach emerged a decade ago in the immunological prophylaxis of pathogenic diseases and trials have been conducted in laboratory animals and non-human primates to evaluate both immune responses namely humoral and cellular immune responses in them [15]. In 1990, Wolff et al. demonstrated that the direct inoculation of DNA or RNA encoding antigens without any delivery system could result in the expression of proteins in the myocytes of the murine, and this activity remained in the skeletal muscle for about 60 days post-inoculation [16]. Later in 1992 Tang in another study demonstrated that inducing an immune response in the host against foreign proteins requires the purification of proteins which is a time-consuming and labor-intensive process. The direct injection of genetic material or plasmid encoding human growth hormone elicited the production of antibodies in mice [17]. Therefore, the concept of DNA vaccine development for the control of trypanosomiasis would revolutionize the prevention of tropical diseases.
2.1. Advantages
DNA vaccination has many beneficial features that make it attractive for the control of various pathogenic diseases and mass vaccination of animals. There is a chance for the reversal of virulence in the case of killed or live-attenuated vaccines, however, the DNA vaccines are subunit vaccines that diminish the reversal of virulence because the whole pathogens are not involved in the synthesis of vaccines [13]. The manufacturing of subunit DNA vaccines is easy, time and labor-saving when compared with the subunit recombinant protein vaccines. Recombinant protein subunit vaccines are not like each other, but all DNA vaccines have similarities with each other. The immunization of plasmid or nucleic acid-based vaccines induces both protective responses such as cellular immune response and humoral immune response [14]. The developing countries could not afford the cost associated with the transportation and maintenance of the cold chain, therefore DNA vaccines serve as the best method for this purpose [13]. Multiple antigens of the same pathogen or different pathogens can be incorporated into a single DNA vaccine [18]. These facts suggest the potential of DNA vaccine development.
2.2. Disadvantages
The health and safety of the animals are very important. The incorporation of the vaccinating DNA into the genome of the host can result in activation of the oncogenes and anti-DNA antibodies can destroy the foreign nucleic acid. These are the limitations of DNA vaccines but are rarely recognized in laboratory animals during experiments. The immunogenicity of the DNA vaccine is poor compared to other methods of immunization. The appropriate level of adjuvants is required to overcome this disadvantage [14]. For the administration of the vaccine, two routes are employed namely intramuscular injection of naked plasmid in murine and gene gun-mediated inoculation. The first approach is very effective in eliciting antibody production in laboratory animals, however, the results are not sufficient in humans and non-human primates [18]. The administration of inoculum through a gene gun showed a better immune response as compared to intramuscular injection of plasmid [14]. If these limitations could be removed, then DNA-based vaccines become more effective.
3. How does the DNA vaccine work?
The immune response shown by the organism to foreign antigens includes humoral immune response (production of antibodies), cytotoxic T lymphocytic response, and T helper responses (Th1 and Th2) mediated by cytokines. Besides these, the immune response is determined by the antigen-presenting cells (APCs) and B lymphocytes which act as detectors, B lymphocytes, and CD8+ toxic T cells which act as effectors, and CD4+ T cells which act as facilitators [18]. When DNA vaccine is introduced into the cell, polypeptides are encoded from the nucleic acid activating the local immune response of the host. It is different from gene therapy in which the introduction of genes into the host genome is desired but in DNA immunization expression of the protein is required [14]. As DNA vaccine is administered by either Intramuscular injection or gene gun into the skin, the expression of plasmid occurs in myoblast and somatic cells (fibroblast and keratinocytes) respectively. The expression of major histocompatibility complex class I molecules can occur at the site of inoculation of plasmid which activates T cells [18]. The proteins expressed by the DNA in host cells are involved in the activation of T cells (CD8+ and CD4+) and B cells [19]. At present, the presentation of antigens by plasmid immunization involves three mechanisms such as transfection and presentation of expressed proteins by dendritic cells, priming by keratinocytes and myocytes, and transfection of somatic cells and antigen-presenting cells with the resultant protein presented to T cells by other APCs [20]. The cytotoxic function is performed by CD8+ T cells which act as an effector of a cell-mediated immune response. On the other hand, CD4+ T cells are involved in the activation and differentiation of B cells into plasma cells which ultimately secrete antibodies and memory cells namely B cells [21]. Therefore, to induce complete immune protection, activation of professional APCs and accurate delivery of plasmid in tissue and proper cell type are necessary (Figure 2).
4. Nanoparticulate and microparticulate for the delivery of DNA vaccines
The immunogenicity of the naked DNA vaccines is poor due to cellular nuclease enzymes which degrade plasmids before expression. Therefore, to enhance its immunogenicity an adjuvant or proper delivery system is required. These microparticulate and nanoparticulate have their role in improving and generating immune responses [19]. The size of the particles also has a role in the transfection of the genes and the preferred range of size is 1 to 10 µm so that the plasmid can be easily taken up by the APCs over other cell types. The nanoparticle delivery system of DNA vaccine has certain characteristics over the microparticle delivery system such as their direct approach to lymph nodes, multiple routes of uptake, and more efficiency of transfection [21]. Various strategies are adopted to boost the potency of plasmids such as the use of genetic adjuvants, optimization of the codon, utilization of highly efficient promotors, intradermal delivery, and electroporation [22]. Such plasmids can be constructed through the same principles followed by recombinant DNA technology. The resultant plasmid when inoculated into animals or humans elicit an immune response [23]. The introduction of foreign DNA into the host mediated by nanoparticle delivery system has certain benefits over the traditional delivery methods such as simple and cost-effective procedure, high solubility, and stability, protection from degradation by cellular DNases, and promotion of endocytosis for cellular uptake of the genes. A nanoparticle delivery system is a controlled administration of antigens in terms of time at a specific site where antigens are recognized by professional APCs due to diffusion, dissolution of nano- or microparticles, and degradation of polymers [24]. The nanoparticle structure has a resemblance to certain viruses such as adenovirus. Therefore, with some modifications, these viruses can serve as an ideal delivery vehicle for DNA vaccines [23]. The non-viral gene transfer agents include cationic polymers and cationic lipids. When plasmid is incorporated with lipids or polymers the resultant product becomes positively charged and small enough to pass the cell membrane with the interventions of receptor-mediated endocytosis. Chitosan is a biodegradable polymer that acts as an ideal agent for the delivery of DNA [24]. The types of nanoparticulate are polymers, metals, ceramics, lipid-based nanoparticles, carbon-based nanoparticles, semiconductors, biological molecules, and immune stimulating complexes (ISCOMs) with many shapes such as round, branched, or carapace-like.
4.1. Polymeric nanoparticulate and microparticulate
It is proved that the efficacy of DNA vaccines can be enhanced when encapsulated in micro- or nanoparticulate that maintains the function of plasmid and taken orally to induce both mucosal and systematic immune responses. Polymers are macromolecules that can encapsulate or dissolve nucleic acid to provide better protection from nucleases, control the release of DNA, and facilitate reaching a specific target cell [25]. The polymeric particle delivery system was also evaluated for gene therapy and tissue engineering. An example of such polymer nano- and microparticulate is polylactide-co-glycolide (PLGA). It is used to encapsulate plasmids against parasitic infections as well as certain bacterial and viral infections [21]. This delivery system elicits a systemic humoral immune response. Cationic PLGA microparticulate loaded with DNA is involved in long-lasting immunity along with higher titer of antibodies and proliferation of lymphocytes [26]. Besides the benefits of polymer microparticles, it also has some demerits such as their large size as compared to nanoparticle upon degradation making the environment of the cell acidic which decrease the immunogenicity of plasmid and vaccine efficacy. Therefore, PLGA nanoparticles are an alternative to these PLGA microparticles [21]. The cationic glycol chitosan shell developed by Lee et al. (2010) can enhance the nucleic acid loading capacity of the delivery system along with the pH-dependent release of plasmid inside the cell. These nanoparticles can transfect the Langerhans cells in the dermis of the host and professional APCs residing in the dermis of the host. The expression of the foreign gene is increased in the lymph node after the migration of Langerhans cells demonstrating the capacity of the delivery system to activate APCs and their movement towards lymph nodes required for the stimulation of B and T naïve cells [27]. Chitosan is a natural polymer obtained from the chitin of insects and crustaceans. It is a partially deacetylated compound having a positive charge that can form electrostatic interaction with a negatively charged DNA phosphate group. This interaction modifies the complex into the nanoscale. Moreover, chitosan has an amine group that makes it suitable for a variety of chemical alterations required for the enhancement of the delivery of the foreign gene, its intracellular dissociation, and the target of the specific cell [28]. Besides the usefulness of chitosan, it also has low immunogenicity and transfection efficacy. Due to its high positive surface charge, it has limited stability in the presence of negatively charged serum proteins [21].
4.2. Lipid nano- and micro particulates
Lipid-based nanoparticles provide a promising delivery system for DNA immunization. They provide improved delivery of plasmid to professional APCs, cellular uptake of DNA, and protection from cellular nuclease enzymes. An example of such a particle used for the delivery of DNA in humans is cationic solid lipid nanoparticulate. Along with protection, it also provides control release of DNA inside the cell. The manufacturing of this nanoparticle is easy, and it offers limited or no toxicity [29]. Cationic lipids are used for the delivery of non-viral genes such as parasitic genes. This delivery platform can control the size of liposomes and is involved in the activation of protein recognition receptors; and innate immune receptors [30].
4.3. Inorganic nanoparticulate
The scientist also investigated the inorganic material that can be used as a delivery platform for genes like DNA or RNA immunization, gene therapy, drug delivery, vaccine adjuvants, and tissue engineering. As organic particles have many advantages, similarly inorganic particles also have certain advantages such as the chances of microbial infection for these inorganic particles are minimal, cheap to prepare, manufacturing is easy, and have good storage capacity [23]. They can be manufactured in a variety of small-sized, and offer a larger surface area to incorporate DNA or plasmid. Currently, examples of such inorganic particles that are used for the delivery of antigens include silver, gold, and magnetic nanoparticles. Gold has also proved to have anti-cancer properties [31]. The lamina propria of the intestine contains many APCs, macrophages, and dendritic cells. Therefore, the administration of DNA vaccine through an oral route encapsulated with metal nanoparticles provides good local and systemic immunity. These APCs activate both B-cells and T-cells [21].
5. Advancement in the development of anti-Trypanosoma brucei DNA-based vaccines
Researchers have demonstrated the importance of DNA vaccines for the prevention of protozoal diseases like trypanosomiasis (Table 1). The complete mechanism of the DNA vaccine still needs to be investigated. However, how DNA vaccines provoke an immune response is discussed above. Based on the strategy that DNA vaccine induces protection. Lanca et al. (2011) conducted research in which they studied the immunization provided by the genes encoding trans-sialidase. It could induce the production of anti-T. brucei antibodies and provide partial protection against this parasitic infection. They also studied the immunogenic role of invariant surface glycoprotein (ISG). Plasmid DNA (ISGpVAX1) encodes ISG and can be a promising candidate for the development of vaccines due to its high immunogenicity. Th1 like IgG2a antibodies produced by the expression of these genes gives partial protection in mice [7]. Silva et al. (2009) studied the humoral immune response induced by DNA immunization in murine. T. brucei trans-sialidase (TbTSA) is expressed only on the surface of the parasite’s procyclic form. Plasmid (nTSApVAX1) encodes amino acids and catalytic domains of TbTSA when inoculated resulting in the production of IgG antibodies against T. brucei. Thus, it showed that a humoral immune response was induced by this antigen, and it provided partial protection of up to 60% when compared with the control group [32].
Shoda et al. (2001) studied the activation of macrophages by DNA encoding a specific antigen of T. brucei. Interestingly, DNA was involved in the stimulation of B-lymphocyte proliferation and activation of macrophages resulting in the production of interleukin-12, tumor necrosis factor-alpha, and nitric oxide. The stimulation of the immune response is dependent on the unmethylated cytosine-guanine dinucleotide (CpG). This unmethylated CpG-dependent mechanism of stimulation of B-lymphocytes and macrophages by parasitic DNA provides the mechanism of innate immune response to control the parasitic infection through the development of a vaccine based on this mechanism [33]. The other genetic-based vaccine includes the genes encoding thiol-specific antioxidants and provides partial protection against T. brucei infection [34].
6. Conclusion and future perspectives
For the control of pathogenic diseases, vaccines are the most effective tool. DNA-based vaccines are a novel approach for the control of infection caused by T. brucei and they provide long-lasting immunity as well as elicit both cell-mediated immune response and humoral immune response that distinguish them from the conventional method of immunization [10,35]. Epitopes derived from the protein of parasites could serve as the best candidate for the development of the vaccine. The attachment of these epitopes with the major histocompatibility complex (MHC) protein is necessary for the stimulation of the immune response. Antigens derived from the VSGs are no longer effective for inducing a protective immune response due to the ability of the parasite to make an antigenic variation. But invariant surface glycoprotein derived from the bloodstream stage of the parasite was evaluated to provide partial protection against the lethal dose of the parasite [2,7]. During infection, the extracellular protozoa are continuously eliminated from the bloodstream of the host by the immune system through IgG anti-T. brucei antibodies and other immune cells. These antibodies are produced against VSG which is not constant and change over time. This mechanism of the parasite to evade the immune system of the host is the major obstacle to the development of a vaccine that also provides memory. The solution for the prevention of T. brucie infection is only the development of a new DNA-based vaccine effective against all different VSGs variants of the parasite. The plasmid encoding ISG (the second most abundant protein on the surface of parasites), and tubulin could be potential candidates for the development of novel vaccines against trypanosomes [34-35]
The expression of DNA in cells of the host is limited due to the presence of cellular nucleases that degrade the DNA and decrease the expression of antigens. Therefore, the use of a delivery system not only provides protection from these enzymes but also enhances the immunogenicity of the vaccine and targets professional APCs [36]. The advancement in nanotechnology has provided such particles that are effectively used for the delivery of antigens via various routes. From the above discussion, we have concluded that only T. brucie is very sharp in changing its antigens so the identification of a new target that is equally effective against all variants of this parasite is necessary. The third-generation vaccine (DNA vaccine) can carry multiple antigens against the same pathogen and can be promising in the control of parasitic infection. The nanoparticles and microparticles have a role in generating the immune response.
Author Contributions
Writing—original draft: A.A., F.M., and M.T.A., Writing—review &editing: Z.D.S., M.T.A., F.M., M.M.A. and A.A. Conceptualization: M.T.A. and A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This article did not receive any research funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available within the article.
Acknowledgments
Thanks to all authors who reviewed and revised this article.
Conflicts of Interest
The authors declare that there is no conflict of interest regarding the publication of this article
References
- Brun, R.; Blum, J.; Chappuis, F.; Burri, C. Human African trypanosomiasis. Lancet 2010, 375, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Casas-Sanchez, A.; Ramaswamy, R.; Perally, S.; Haines, L.R.; Rose, C.; Aguilera-Flores, M.; Portillo, S.; Verbeelen, M.; Hussain, S.; Smithson, L.; et al. The Trypanosoma brucei MISP family of invariant proteins is co-expressed with BARP as triple helical bundle structures on the surface of salivary gland forms, but is dispensable for parasite development within the tsetse vector. PLOS Pathog. 2023, 19, e1011269. [Google Scholar] [CrossRef] [PubMed]
- Büscher, P.; Cecchi, G.; Jamonneau, V.; Priotto, G. Human African trypanosomiasis. Lancet 2017, 390, 2397–2409. [Google Scholar] [CrossRef] [PubMed]
- Steverding, D. The history of African trypanosomiasis. Parasites Vectors 2008, 1, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Simarro, P.P.; Franco, J.R.; Diarra, A.; Jannin, J.G. Epidemiology of human African trypanosomiasis. Clin. Epidemiology 2014, 6, 257–275. [Google Scholar] [CrossRef] [PubMed]
- Legros, D.; Ollivier, G.; Gastellu-Etchegorry, M.; Paquet, C.; Burri, C.; Jannin, J.; Büscher, P. Treatment of human African trypanosomiasis—present situation and needs for research and development. Lancet Infect. Dis. 2002, 2, 437–440. [Google Scholar] [CrossRef]
- Lança, A.S.C.; de Sousa, K.P.; Atouguia, J.; Prazeres, D.M.F.; Monteiro, G.A.; Silva, M.S. Trypanosoma brucei: Immunisation with plasmid DNA encoding invariant surface glycoprotein gene is able to induce partial protection in experimental African trypanosomiasis. Exp. Parasitol. 2011, 127, 18–24. [Google Scholar] [CrossRef]
- Baral, T.N. Immunobiology of African Trypanosomes: Need of Alternative Interventions. J. Biomed. Biotechnol. 2010, 2010, 389153. [Google Scholar] [CrossRef]
- Berriman, M.; Ghedin, E.; Hertz-Fowler, C.; Blandin, G.; Renauld, H.; Bartholomeu, D.C.; Lennard, N.J.; Caler, E.; Hamlin, N.E.; Haas, B.; et al. The Genome of the African Trypanosome Trypanosoma brucei. Science 2005, 309, 416–422. [Google Scholar] [CrossRef]
- Akhoon, B.A.; Slathia, P.S.; Sharma, P.; Gupta, S.K.; Verma, V. In silico identification of novel protective VSG antigens expressed by Trypanosoma brucei and an effort for designing a highly immunogenic DNA vaccine using IL-12 as adjuvant. Microb. Pathog. 2011, 51, 77–87. [Google Scholar] [CrossRef]
- Rhind, S.G.; Shek, P.N. Cytokines in the pathogenesis of human African trypanosomiasis: antagonistic roles of TNF-α and IL-10. Progress in human African trypanosomiasis, sleeping sickness. Springer-Verlag. 1999, 119–135. [CrossRef]
- Kurup, S.P.; Tewari, A.K. Induction of protective immune response in mice by a DNA vaccine encoding Trypanosoma evansi beta tubulin gene. Veter- Parasitol. 2012, 187, 9–16. [Google Scholar] [CrossRef]
- Wilson, P.C.; Andrews, S.F. Tools to therapeutically harness the human antibody response. Nat. Rev. Immunol. 2012, 12, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Smooker, P.M.; Rainczuk, A.; Kennedy, N.; Spithill, T.W. DNA vaccines and their application against parasites – promise, limitations and potential solutions. Biotechnol. Annu. Rev. 2004, 10, 189–236. [Google Scholar] [CrossRef] [PubMed]
- Hasson, S.S.A.A.; Al-Busaidi, J.K.Z.; Sallam, T.A. The past, current and future trends in DNA vaccine immunisations. Asian Pac. J. Trop. Biomed. 2015, 5, 344–353. [Google Scholar] [CrossRef]
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct gene transfer into mouse muscle in vivo. Science 1990, 247, 1465–1468. [Google Scholar] [CrossRef]
- Tang, D.-C.; DeVit, M.; Johnston, S.A. Genetic immunization is a simple method for eliciting an immune response. Nature 1992, 356, 152–154. [Google Scholar] [CrossRef]
- Cui, Z. DNA Vaccine. Adv. Genet. 2005, 54, 257–289. [Google Scholar] [CrossRef]
- Mohajeri, P.; Soltani, S.; Farahani, A.; Dastranj, M.; Momenifar, N.; Emamie, A.D. DNA vaccine: Methods and mechanisms. Adv. Hum. Biol. 2018, 8, 132. [Google Scholar] [CrossRef]
- Gurunathan, S.; Klinman, D.M.; Seder, R.A. DNA Vaccines: Immunology, Application, and Optimization. Annu. Rev. Immunol. 2000, 18, 927–974. [Google Scholar] [CrossRef]
- Farris, E.; Brown, D.M.; E Ramer-Tait, A.; Pannier, A.K. Micro- and nanoparticulates for DNA vaccine delivery. Exp. Biol. Med. 2016, 241, 919–929. [Google Scholar] [CrossRef]
- Saade, F.; Petrovsky, N. Technologies for enhanced efficacy of DNA vaccines. Expert Rev. Vaccines 2012, 11, 189–209. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.A.A.; He, N.; Li, Z.; Ali, Z.; Zhang, L. Nanoparticles for DNA vaccine delivery. J. Biomed. Nanotechnol. 2014, 10, 2332–2349. [Google Scholar] [CrossRef] [PubMed]
- Kersten, G.; Hirschberg, H. Antigen delivery systems. Expert Rev. Vaccines 2004, 3, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Rodríguez, S.A.; Puel, F.; Briançon, S.; Allémann, E.; Doelker, E.; Fessi, H. Comparative scale-up of three methods for producing ibuprofen-loaded nanoparticles. Eur. J. Pharm. Sci. 2005, 25, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Briones, M.; Ott, G.; O'Hagan, D. Cationic microparticles: A potent delivery system for DNA vaccines. Proc. Natl. Acad. Sci. USA 2000, 97, 811–816. [Google Scholar] [CrossRef]
- Lee, P.-W.; Hsu, S.-H.; Tsai, J.-S.; Chen, F.-R.; Huang, P.-J.; Ke, C.-J.; Liao, Z.-X.; Hsiao, C.-W.; Lin, H.-J.; Sung, H.-W. Multifunctional core-shell polymeric nanoparticles for transdermal DNA delivery and epidermal Langerhans cells tracking. Biomaterials 2010, 31, 2425–2434. [Google Scholar] [CrossRef]
- Liu, W.G.; Zhang, X.; Sun, S.J.; Sun, G.J.; De Yao, K.; Liang, D.C.; Guo, G.; Zhang, J.Y. N-Alkylated Chitosan as a Potential Nonviral Vector for Gene Transfection. Bioconjugate Chem. 2003, 14, 782–789. [Google Scholar] [CrossRef]
- Wilson, K.D.; de Jong, S.D.; Tam, Y.K. Lipid-based delivery of CpG oligonucleotides enhances immunotherapeutic efficacy. Adv. Drug Deliv. Rev. 2009, 61, 233–242. [Google Scholar] [CrossRef]
- Lonez, C.; Bessodes, M.; Scherman, D.; Vandenbranden, M.; Escriou, V.; Ruysschaert, J.-M. Cationic lipid nanocarriers activate Toll-like receptor 2 and NLRP3 inflammasome pathways. Nanomedicine: Nanotechnology, Biol. Med. 2014, 10, 775–782. [Google Scholar] [CrossRef]
- Mukherjee, P.; Bhattacharya, R.; Wang, P.; Wang, L.; Basu, S.; Nagy, J.A.; Atala, A.; Mukhopadhyay, D.; Soker, S. Antiangiogenic Properties of Gold Nanoparticles. Clin. Cancer Res. 2005, 11, 3530–3534. [Google Scholar] [CrossRef]
- Silva, M.S.; Prazeres, D.M.F.; Lança, A.; Atouguia, J.; Monteiro, G.A. Trans-sialidase from Trypanosoma brucei as a potential target for DNA vaccine development against African trypanosomiasis. Parasitol. Res. 2009, 105, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Shoda, L.K.M.; Kegerreis, K.A.; Suarez, C.E.; Roditi, I.; Corral, R.S.; Bertot, G.M.; Norimine, J.; Brown, W.C. DNA from Protozoan ParasitesBabesia bovis, Trypanosoma cruzi, andT. bruceiIs Mitogenic for B Lymphocytes and Stimulates Macrophage Expression of Interleukin-12, Tumor Necrosis Factor Alpha, and Nitric Oxide. Infect. Immun. 2001, 69, 2162–71. [Google Scholar] [CrossRef] [PubMed]
- A Carvalho, J.; Rodgers, J.; Atouguia, J.; Prazeres, D.M.; A Monteiro, G. DNA vaccines: a rational design against parasitic diseases. Expert Rev. Vaccines 2010, 9, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Danazumi, A.U.; Gital, S.I.; Idris, S.; Dibba, L.B.; Balogun, E.O.; Górna, M.W. Immunoinformatic design of a putative multi-epitope vaccine candidate against Trypanosoma brucei gambiense. Comput. Struct. Biotechnol. J. 2022, 20, 5574–5585. [Google Scholar] [CrossRef]
- Li, L.; Saade, F.; Petrovsky, N. The future of human DNA vaccines. J. Biotechnol. 2012, 162, 171–182. [Google Scholar] [CrossRef]
Figure 1.
Both types of immune responses induced by DNA vaccine.
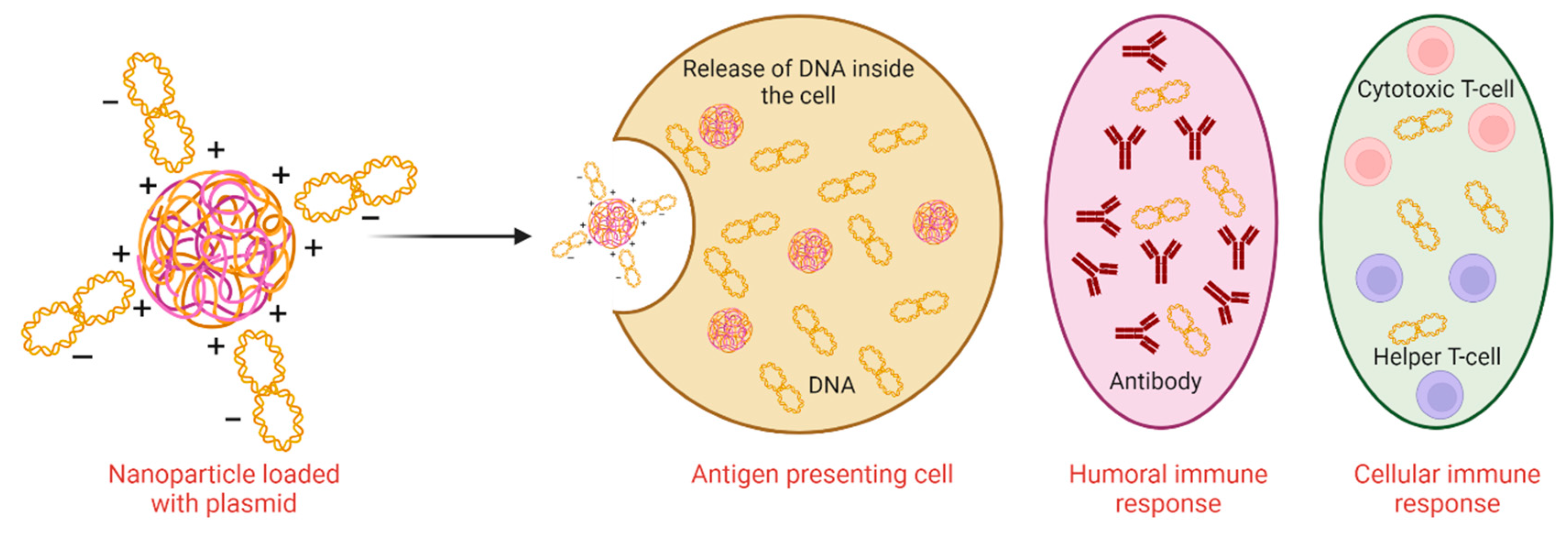
Figure 2.
Mechanism of DNA immunization in the host.
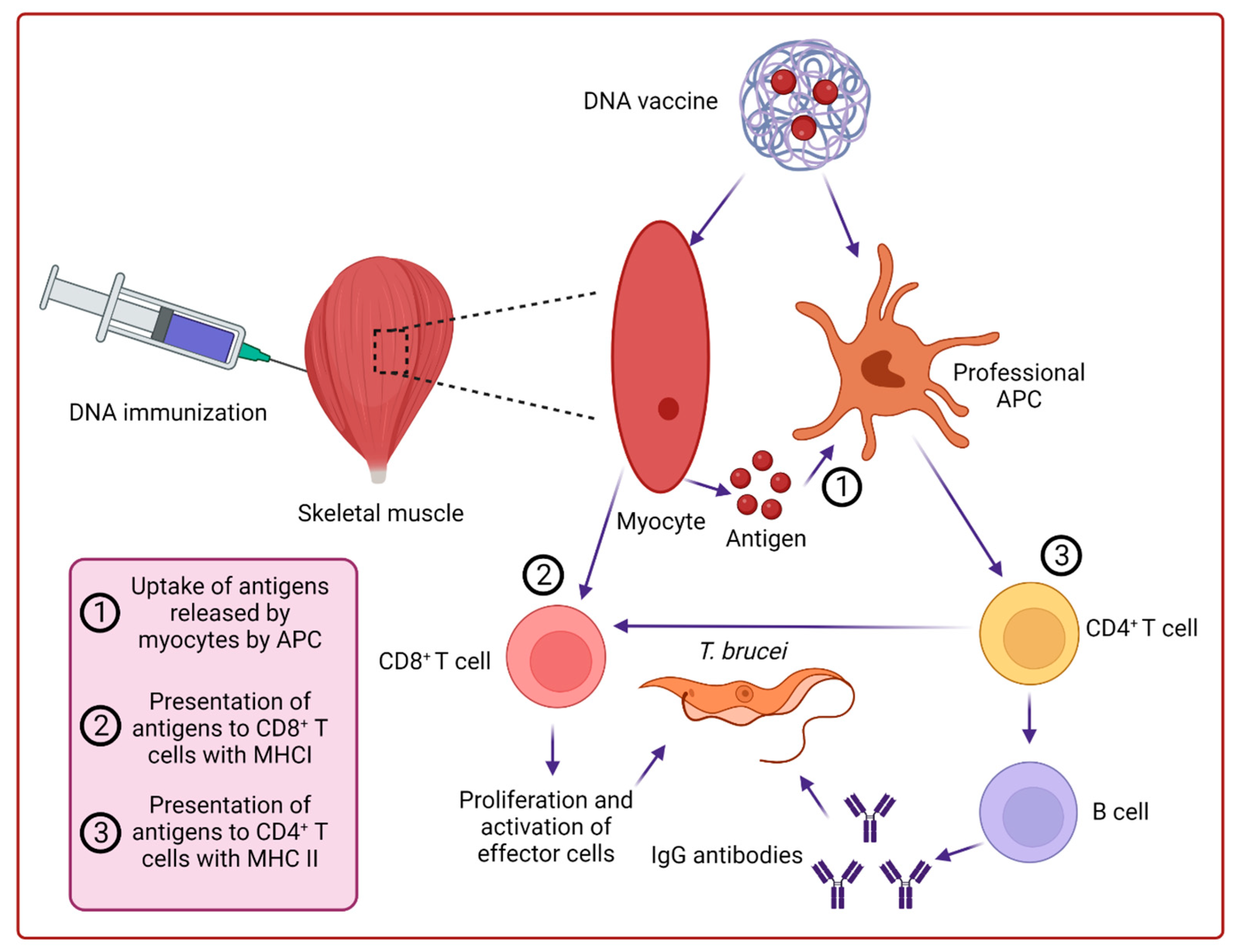

Table 1.
Studies conducted for the evaluation of DNA-based vaccines against T. brucei.
| Candidate antigen | Strain | Extraction site | Host | Effect | Reference |
|---|---|---|---|---|---|
| Metacyclic invariant surface proteins | Trypanosoma brucei brucei strain AnTat 1.1 90:13 | T. brucei-infected tsetse saliva | BALB/c 6–8 weeks old female mice | May act as a transmission blocking vaccine target | [2] |
| DNA encoding trans-sialidase 5′-terminal region | T. brucei brucei GVR 35/1.5 | Genome | BALB-c mice | IgG antibodies were produced and provided 60% protection in challenged mice | [32] |
| DNA encoding invariant surface glycoprotein specific for bloodstream stage | T. b. brucei GVR 35/1.5 | Genome | Balb/C mice | Anti-trypanosoma IgG antibodies were produced suggesting T helper cell type 1 response (Th1) | [7] |
| T. brucei DNA | - | Trypomastigotes were isolated from rats | Cattle macrophages | Promotes B-lymphocyte proliferation | [33] |
| Variant surface glycoprotein | In silico study | Immunogenic | [10] | ||
| Proteins harboring transmembrane helices | May stimulate cellular and humoral immune responses | [35] | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



